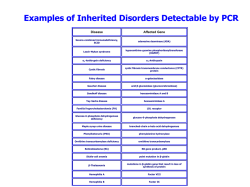
LA VIA DEL GUSTO UN PERCORSO TRA GENETICA E CULTURA DEL TERRITORIO PROGETTO DI DIDATTICA DI GENETICA MOLECOLARE BITTER RECEPTOR GENE LA VARIAZIONE GENETICA ASSOCIATA ALLA PERCEZIONE DEL GUSTO AMARO ANNO 2012-13 PROF.SSA BEATRICE ZANINI UNIVERSITA’ DEGLI STUDI DI GENOVA in collaborazione con COOP LIGURIA QUADRO GENERALE DELL’ATTIVITA’ MODULO DI GENETICA MOLECOLARE: MATERIALE: - Laboratorio di bioinformatica - Laboratorio di Genetica molecolare Copie del testo Testi in PP TECNICHE UTILIZZATE IN LABORATORIO ADATTA PER - Estrazione di DNA dalle cellule della mucosa boccale - PCR - Elettroforesi 4° E 5°anno Scuola Secondaria Superiore CONCETTI CHIAVE TEMPO DI REALIZZAZIONE - DNA/mRNA Gene/Locus/Allele Polimorfismo allelico Aplotipo Linkage Disequilibrium 2 moduli di 4 ore LABORATORIO DIDATTICO Università-IST-S.Martino PREREQUISITI 1 La struttura del DNA. 2.. Dal DNA al cromosoma 3. Aploidia/Diploidia 4 Gene/Locus/Allele. 5. Genotipo/Fenotipo 6. Duplicazione del DNA 7. Trascrizione del mRNA 8. Traduzione 9. Struttura dei geni 10. Regolazione dell’espressione genica 11. Il promotore 12. Splicing 13. Enzimi di restrizione 14. Polimorfismi 15. Polimorfismo allelico 16. Polimorfismi di sequenza ATTIVITA’ WEBSITE http://learn.genetics.utah.edu http:// www.dnalc.org http://www.ncbi.org 2 INDICE GUIDA PER IL DOCENTE A. B. C. D. pag.4 Obiettivi Background di conoscenze da fornire allo studente Argomenti correlati all’attività di Laboratorio: Strategia d’insegnamento Argomenti correlati all’attività di Laboratorio 1. Struttura del DNA 2. Replicazione del DNA 3. La trascrizione negli Eucarioti 4. Struttura ed espressione dei geni. Promotore. Enhancer. Esoni e introni. 5. Dal DNA ai cromosomi 6. Regolazione dell’espressione genica I livelli di regolazione genica 7. Enzimi di restrizione 8. Marcatori genetici e Polimorfismi 9. Polimorfismi a singolo nucleotide o SNP 10. Aplotipi 11. Analisi di Linkage 12. Tecniche utilizzate in laboratorio per l’ analisi molecolare del DNA: - Estrazione del DNA - PCR - Elettroforesi su gel di agarosio pag.5 pag.5 pag.6 pag.7 pag.9 pag.10 pag.12 pag.13 pag.11 pag.15 pag.16 pag.18 “ BITTER RECEPTOR GENE TAS2R38. LA VARIAZIONE GENETICA ASSOCIATA ALLA PERCEZIONE DEL GUSTO AMARO” Introduzione. Il gusto Genetica del gusto amaro I recettori dell’amaro. La famiglia TAS2R Polimorfismo rs 713598 Modulo di Bioinformatica Modulo di Laboratorio Bitter Taste Sensitivity Test Food Preference Questionnaire Bibliografia pag.21 pag.26 pag.28 pag 29 pag.31 pag.36 pag.39 pag. 40 pag.42 PAGINE PER LO STUDENTE pag.45 - Background di conoscenze - Glossario - Test per studenti Basic Knowledge Test Post Test pag 45 pag.46 3 Guida per il docente A. Obiettivi: In questa attività si studia la condizione nota come “Percezione del gusto amaro” correlata con il gene TAS2R38 che codifica per il recettore per l’amaro. Tre SNPs sono implicati nella espressione fenotipica che identifica soggetti “taster” e “non taster”; gli studenti potranno identificare uno dei tre SNPs, associati ai fenotipi AVI/AVI, PAV/AVI e PAV/AVI. B. Background di conoscenze da fornire allo studente 1. Leggi di Mendel e terminologia della Genetica mendeliana 2. Struttura chimica del DNA e livelli di organizzazione molecolare. 3. Duplicazione, trascrizione, traduzione 4 Meccanismi di regolazione C. Argomenti correlati all’attività di laboratorio 1. Meccanismi di regolazione 2. I polimorfismi di sequenza del DNA: i RFLP e gli SNP. Frequenza degli SNP e individuazione degli SNP 3. Aplotipi 4. Studi di Linkage 5. Tecniche di laboratorio D. Strategia d’insegnamento: 1. Attività di Bioinformatica: Abstract: Gli studenti navigano in Internet con accesso a Internet utilizzando il Modulo ”Percorso di Bioinformatica” per compiere una ricerca web e per alleli, SNP imparare l’approccio metodologico di un lavoro di ricerca Materiale: Computers Concetti chiave: Geni, alleli, SNP 2. Laboratorio di Genetica Molecolare Abstract: Il DNA genomico viene estratto dalle cellule della mucosa buccale. Il polimorfismo C/G è identificato mediante PCR e successiva digestione enzimatica. La corsa elettroforetica evidenzierà la lunghezza dei frammenti e permetterà di definire le varianti alleliche Durata: 2 mezze giornate Concetti chiave: Lo studio del carattere “Percezione del gusto amaro”, mediante l’analisi di uno dei tre SNPs, introduce al concetto di polimorfismo allelico Età: 17-18 anni 4 Argomenti correlati all’attività di laboratorio 1. STRUTTURA DEL DNA La molecola del DNA è un polimero, ossia è un insieme di tanti monomeri: i nucleotidi. Ogni nucleotide è costituito da tre componenti: un gruppo fosfato, uno zucchero (desossiribosio) e una base azotata. La molecola di DNA è formata da due catene polinucleotidiche avvolte l’una intorno all’altra con andamento destrorso. Le due catene sono antiparallele, cioè i due singoli filamenti sono orientati uno in direzione 5’->3’ e l’altro 3’->5’. Gli scheletri zucchero – fosfato si trovano all’esterno, le basi azotate all’interno. Le basi delle due catene sono unite tra loro mediante legami a idrogeno. Le basi sono complementari e il loro appaiamento è: A=T Adenina–Timina G =C Guanina-Citosina L’informazione genetica risiede nella sequenza di basi 2. REPLICAZIONE DEL DNA NEGLI EUCARIOTI La replicazione del DNA in tutte le cellule viventi, dai batteri all’uomo, è un processo complesso, che richiede l’intervento di più di una dozzina di enzimi diversi. La replicazione comincia in corrispondenza di siti detti origine di replicazione presenti ad intervalli in questi siti; alcune proteine srotolano la doppia elica di DNA, rompendo i legami a idrogeno tra le basi dei filamenti complementari. L’allineamento e unione tra loro dei nucleotidi complementari avviene per azione della DNA POLIMERASI che procede solo in direzione 5’-> 3’, cioè aggiunge nuove basi all’estremità 3’ del filamento esistente. La DNA polimerasi, per iniziare il processo, ha anche bisogno di un innesco (o primer), a cui attaccarsi e procedere con la polimerizzazione lungo i cromosomi. Il primer è costituito da una corta sequenza polinucleotidica di RNA. Poiché la DNA Polimerasi può sintetizzare il DNA in una sola direzione, ed i due filamenti di DNA procedono in direzioni opposte, le DNA Polimerasi sui due filamenti opposti devono sintetizzare il DNA in due direzioni opposte. Questo significa che un filamento può essere sintetizzato in modo continuo e veloce da un innesco iniziale (filamento principale), ma l’altro filamento (filamento in ritardo) deve essere sintetizzato in modo discontinuo con più eventi di innesco della replicazione e brevi tratti di DNA che vengono poi uniti tra loro ( frammenti di Okazaki) La replicazione è semiconservativa: ogni emi-elica (singolo filamento) della molecola madre serve da stampo per la sintesi di un nuovo filamento, per cui ogni doppia elica figlia sarà costituita da un filamento vecchio e da un filamento nuovo. Le molecole risultanti sono copie esatte dell’originale. Da una doppia elica madre derivano due doppie eliche figlie uguali tra loro e uguali alla molecola madre. 5 3. LA TRASCRIZIONE NEGLI EUCARIOTI L’informazione genetica contenuta nelle sequenze del DNA viene trasferita all’RNA e dall’RNA al polipeptide corrispondente. Durante la trascrizione, un complesso proteico, comprendente l’enzima RNA polimerasi, sintetizza le molecole di RNA sullo stampo delle sequenze di DNA che costituiscono le unità di trascrizione. La RNA polimerasi si lega al sito d’inizio della trascrizione insieme ad altre proteine, dette fattori di trascrizione. Questi fattori, mediante l’interazione con brevi sequenze di DNA presenti nella regione a monte dell’inizio della trascrizione (promotore), servono a posizionare la RNA polimerasi nel sito giusto e a separare i due filamenti di DNA per formare la bolla di trascrizione. Il processo continua fino a che la polimerasi incontra una sequenza di arresto. L’enzima usa come stampo uno dei due filamenti di DNA in direzione 5’->3’, catalizzando il legame fosfodiestere tra il gruppo ossidrilico legato al C3’ del ribonucleotide precedente e il fosfato del nuovo ribonucleotide. A questo punto si stacca e libera la catena di RNA, mentre la bolla di trascrizione si richiude e il DNA riassume la conformazione a doppia elica. L’RNA neosintetizzato ha la sequenza di basi identica a quella di uno dei due filamenti di DNA (il filamento senso), anche se la Timina è sostituita dall’Uracile. Da uno stesso gene possono essere trascritte consecutivamente numerose copie di RNA e il livello di trascrizione dipende da complessi meccanismi (vedi la regolazione della trascrizione). E’ importante ricordare che le cellule eucariotiche possiedono tre tipi di RNA polimerasi: - RNA polimerasi I trascrive i geni degli RNA ribosomiali - RNA polimerasi II trascrive i geni che codificano proteine sintetizzando i precursori degli RNA messaggeri e anche alcuni piccoli RNA - RNA polimerasi III trascrive i geni di tutti gli RNA transfer, un RNA ribosomiale e altri piccoli RNA. I precursori degli mRNA neosintetizzati (trascritti primari) devono subire una serie di modificazioni prima di essere trasferiti nel citoplasma per venire tradotti sui ribosomi. Questo processo di maturazione degli mRNA include le seguenti modificazioni: • Aggiunta all’estremità 5’ di un cappuccio (cap). Al primo nucleotide all’estremità 5’ della molecola di RNA nascente viene rimosso il fosfato terminale e viene aggiunta una molecola di Guanosina monofosfato (GMP) metilata in posizione 7’. Il capping serve per proteggere il trascritto dall’attacco delle esonucleasi che lo degraderebbero, e per facilitare il trasporto dal nucleo al citoplasma. • Rimozione di alcune sequenze che non vengono tradotte (processo di splicing). Quasi tutti i geni eucariotici sono divisi in sequenze codificanti, chiamate esoni, e sequenze non tradotte, dette introni. Questi ultimi vengono rimossi dai trascritti primari mediante il processo di splicing. Gli introni sono quindi sequenze di DNA, situate tra due esoni, le quali sono trascritte ma non tradotte. Salvo rare eccezioni, gli introni iniziano sempre con i nucleotidi GT e terminano con i nucleotidi AG (regola GT-AG). Nel processo di splicing si verifica prima la scissione all’inizio dell’introne (5’), poi l’estremità libera dell’introne si ripiega su se stessa formando una struttura simile ad un laccio e infine avviene il taglio a livello della giunzione 3’ dell’introne. Quindi i due esoni si uniscono mentre l’introne va perso. Una struttura macromolecolare (costituita da varie subunità di molecole di piccoli RNA nucleari, gli snRNP, e da una serie di proteine specifiche) promuove e controlla le reazioni dello splicing. • Aggiunta all’estremità 3’ di una coda poli-A. La maggior parte delle unità di trascrizione hanno una breve sequenza (AATAAA) che specifica il sito di termine della trascrizione. Circa 6 15-30 nucleotidi a valle di questo sito, l’RNA neosintetizzato viene scisso da un enzima, una endonucleasi, e alla molecola di RNA vengono aggiunti circa 200 residui di Adenosina monofosfato (AMP). Questa coda di poli-A ha lo scopo di stabilizzare le molecole degli mRNA maturi e di facilitare il loro trasporto dal nucleo al citoplasma. 4. STRUTTURA ED ESPRESSIONE DEI GENI Dal punto di vista della genetica molecolare per gene s’intende una sequenza di DNA potenzialmente trascrivibile in RNA funzionalmente attivo. Tale RNA può svolgere direttamente una funzione strutturale e/o catalitica (rRNA, tRNA) oppure trasportare l’informazione per la sintesi di una proteina (mRNA). Nel genoma umano si stima che siano presenti circa 23.000 geni codificanti proteine e 1000-2000 geni codificanti RNA strutturali. Da recenti studi emergerebbe però l’esistenza di diverse migliaia (o decine di migliaia) di trascritti non codificanti che potrebbero non avere alcuna funzione o, viceversa, svolgere un ruolo fondamentale nella regolazione della conformazione della cromatina e della trascrizione di geni codificanti proteine. IL PROMOTORE La regione a monte del sito d’inizio della trascrizione è detta promotore. In questa regione, di lunghezza variabile, si trova una serie di brevi sequenze che vengono riconosciute e legate da fattori di trascrizione. I fattori di trascrizione favoriscono il legame dell’RNA polimerasi al sito giusto per iniziare la sintesi di RNA. I geni che presentano elevati livelli di trascrizione, presentano nel promotore delle sequenze specifiche ( i TATA box a circa -25 bp dal sito d’inizio della trascrizione; il CAAT box, a -80 bp dal sito d’inizio della trascrizione, i GC box). Accanto a sequenze comuni a molti promotori vi sono elementi che sono riconosciuti da fattori di trascrizione tessuto-specifici. Anche i geni che mostrano un’espressione tessuto-specifica vengono spesso trascritti a livelli molto bassi in tutte le cellule. Vi sono altre sequenze che vengono riconosciute da fattori di trascrizione quali gli elementi di risposta, localizzati nel promotore o nella regione 5’ del gene, e gli elementi indicati come enhancer (intensificatori). GLI ENHANCERS Gli enhancers sono sequenze nucleotidiche cis-agenti che esplicano la loro funzione aumentando notevolmente (fino a 200 volte) la frequenza di trascrizione del gene che controllano. Dal punto di vista strutturale, un enhancer non differisce molto da un promotore. Gli enhancers non devono necessariamente essere vicini ai promotori: è possibile infatti trovare degli enhancer a parecchie centinaia di migliaia di paia di basi di distanza a valle o a monte del sito d'inizio della trascrizione. 7 ESONI E INTRONI Il primo esone comincia al sito d’inizio della trascrizione, ma il primo tratto (fatto di 200300 bp) non è codificante; questo segmento pertanto è trascritto, ma non tradotto e viene indicato come regione 5’UTR ( UnTraslated Region). La regione 5’UTR è importante per l’efficienza della traduzione in quanto facilita il legame dell’mRNA ai ribosomi. La regione tradotta inizia generalmente con ATG, nel 1° esone. Il numero degli esoni presenti nei geni umani è altamente variabile. Vi sono geni piccoli costituiti da un singolo esone e altri che possiedono più di 100 esoni. Il numero medio è di 9-10 esoni per gene. I singoli esoni sono generalmente piuttosto piccoli con dimensioni medie di circa 200bp, ma esistono alcuni esoni eccezionalmente lunghi che possono superare anche le 5 kb. Al contrario degli esoni, la dimensione degli introni è molto variabile. Generalmente i geni piccoli hanno introni piccoli, mentre in quelli più grandi gli introni possono avere anche una lunghezza di 10-20 kb. Quasi tutti gli introni cominciano con GT (sito donatore di splicing) e terminano con AG (sito accettore di splicing). Questi dinucleotidi sono circondati da sequenze consenso, altamente conservate nel corso dell’evoluzione e molto simili tra loro. Il processo di splicing deve essere molto preciso dato che lo spostamento anche di un singolo nucleotide determinerebbe lo slittamento del modulo di lettura e quindi la sintesi di una proteina alterata. L’ultimo esone, così come il primo, contiene una sequenza trascritta, ma non tradotta, detta regione 3’UTR. Schema di un gene umano costituito da tre esoni. Sono indicati alcuni elementi del promotore e i siti importanti per la trascrizione e la maturazione dell’mRNA. La numerazione dei nucleotidi del promotore inizia da -1 che corrisponde al nucleotide che precede il sito d’inizio della trascrizione (indicato con +1). Nella regione del promotore - di lunghezza variabile – si trovano delle brevi sequenze che vengono riconosciute e legate da fattori di trascrizione. Questi – a loro volta - favoriscono il legame dell’RNA polimerasi al sito giusto per iniziare la sintesi di RNA dal +1. E’ rappresentato anche il trascritto primario ( precursore dell’mRNA) e l’mRNA maturo. 5’UTR (UnTraslated Region) è l’estremità 5’ del primo esone che viene trascritta ma non tradotta. 3’UTR (UnTraslated Region) è l’estremità 3’ dell’ultimo esone che viene trascritta ma non tradotta. - capping = all’estremità 5’ viene attaccato un nucleotide “rovesciato” (con un legame anidridico tra fosfati) - splicing = rimozione degli introni e giunzione degli esoni - poliadenilazione = rimozione della porzione 3’ del trascritto primario e nell’aggiunta di numerosi polinucleotidi adenilici (poliA) 8 5. DAL DNA AL CROMOSOMA Alla molecola di DNA sono associati gli istoni (proteine basiche), essenziali per permettere l’avvolgimento e il ripiegamento del DNA in strutture estremamente compatte, vale a dire i cromosomi, visibili solo durante la divisione cellulare. I cromosomi umani sono costituiti di cromatina. La cromatina è un complesso sovramolecolare di acidi nucleici (DNA e RNA) e proteine, una struttura altamente dinamica il cui grado di organizzazione o compattazione o “condensazione” varia a. da cellula a cellula, b. durante il ciclo vitale della cellula, c. tra zone diverse dei nuclei interfasici La cromatina può essere osservata in due diverse forme : 1. eucromatina - dispersa, debolmente colorata, attivamente espressa cioè trascrivibile. 2. eterocromatina - molto compattata, colorata marcatamente, ed inattiva cioè non trascrivibile Il cromosoma : Unità strutturale e colorabile che porta i geni disposti in modo lineare. E’ considerato anche come un insieme di geni - o gruppo di associazione organizzati secondo una successione lineare che tendono ad essere ereditati insieme Ogni cromosoma e’ costituito da una successione lineare di geni o loci. Il locus è la posizione occupata da un gene su un cromosoma I cromosomi sono presenti in coppie nelle cellule somatiche e i membri di una coppia sono i cromosomi omologhi. P' M' Ogni coppia di cromosomi omologhi contiene un cromosoma di origine paterna e un cromosoma di origine materna centromero' '''''''''''''Fase'S' REPLICAZIONE'DEL' DNA' Cromosomi'omologhi' Croma:di'fratelli' 9 Cromosomi omologhi (contengono forme alternative degli stessi geni) Gli alleli sono le forme diverse di uno stesso gene Replicazione+ del+DNA++ centromero cromatidi fratelli (sono identici) cromatidi fratelli (sono identici) Nell'immagine accanto, i due cromosomi omologhi sono rappresentati in nero. Il locus genetico è in turchese. Vediamo come le forme diverse dello stesso gene, gli alleli, sono raffigurati in diversi colori. Gli alleli possono essere uguali o diversi nei due cromosomi. Queste forme alternative si dicono alleli. Se gli alleli sono uguali sui due membri della coppia di cromosomi omologhi l'individuo è detto omozigote per quel particolare gene, se invece gli alleli sono diversi l'individuo è eterozigote. 6. LA REGOLAZIONE DELL’ESPRESSIONE GENICA Premessa. Esistono geni costantemente attivi (geni costitutivi), espressi ubiquitariamente, che codificano per proteine house-keeping che svolgono funzioni comuni a tutte le cellule (enzimi del metabolismo, proteine del citoscheletro, del Reticolo endoplasmatico e del Golgi) . Altri geni (geni regolatori) la cui espressione è regolata in base alle necessità della cellula o dell’organismo codificano per proteine specializzate. LIVELLI DI REGOLAZIONE GENICA NEGLI EUCARIOTI Il controllo dell’espressione genica si realizza a vari livelli: A. Lo stato di condensazione della cromatina Una conformazione della cromatina del gene “aperta”, ovvero i nucleosomi non compattati e, possibilmente, il DNA non associato agli istoni nel promotore. Generalmente la cromatina attiva – cioè decondensata - è caratterizzata da un alto livello di acetilazione degli istoni H3 e H4. Al contrario, gli istoni associati alla cromatina inattiva, non trascritta, sono tutti de-acetilati. Un’altra modifica degli istoni è la metilazione delle citosine. Elevata metilazione in regioni non codificanti al 5’ di un gene lo inattiva trascrizionalmente. Le citosine metilate vengono riconosciute da proteine determinando il reclutamento di altre proteine che modificano la cromatina e la compattano, facilitando l’inattivazione trascrizionale del gene stesso. Notevoli progressi sono stati fatti recentemente nell’identificazione di modificazioni della cromatina associate a una conformazione aperta (attiva) o compatta (inattiva). Tali modificazioni sono dette epigenetiche, perché non modificano la sequenza primaria del DNA, 10 B. La trascrizione Esistono due tipi principali di sequenze del DNA che regolano l’espressione genica: i promotori dei geni e le sequenze di regolazione genica (enhancer e silenziatori). Del promotore e dell’enhancer abbiamo già parlato nel paragrafo precedente. Specifici fattori di trascrizione interagiscono con brevi sequenze nel promotore del gene e con sequenze enhancer o silenziatori, e consentono l’assemblaggio del complesso di trascrizione. Il controllo dell’espressione genica mediante il legame di fattori trascrizionali proteici con le sequenze di regolazione è estremamente complesso e coinvolge numerosi fattori che possono essere grossolanamente distinti in fattori ubiquitari e tessuto-specifici. L’interazione di fattori specifici con gli elementi enhancer è importante per l’espressione genica tessuto-specifica. Elementi regolatori della trascrizione 1. fattori di trascrizione 2. elementi cis-acting 3. elementi di regolazione distanti anche 1 Mb 4. promotori alternativi/multipli 5. modificazioni in DNA e istoni (acetilazioni, metilazione)/accessibilità alla cromatina 6. piccoli RNAs di tanti tipi. Elementi cis-acting “Cis-acting” generalmente significa “che agisce sulla stessa molecola”. Nel contesto della regolazione della trascrizione, sono generalmente considerati “elementi cis-acting” delle sequenze di DNA, che attraverso i fattori di trascrizione, regolano l’espressione dei geni sullo stesso cromosoma. Un esempio di sequenza regolatoria cis-acting è l’”operatore” dell’operone lac. Questa sequenza di DNA è legata dal repressore lac che impedisce la trascrizione dei geni adiacenti sulla stessa molecola di DNA. Possiamo dire quindi che l’operatore lac “agisce in cis” nella regolazione dei geni vicini. L’operatore non codifica per alcuna proteina o RNA. C. Regolazione post-trascrizionale. Si esercita a livello di: - Stabilità dell’mRNA. L’emivita delle molecole di mRNA può essere molto variabile. - Trascritti alternativi e isoforme proteiche Un tempo si riteneva che ogni gene codificasse per un unico prodotto polipeptidico o molecola di RNA, mentre oggi si sa che la maggioranza di geni umani specifica due o più forme alternative di proteine (isoforme). I meccanismi con i quali vengono generate queste diverse isoforme sono: - l’uso di promotori alternativi - lo splicing alternativo 11 - la poliadenilazione alternativa Si conoscono diversi geni umani che hanno due o più promotori che sono attivi specificamente in determinati tessuti e dirigono la sintesi di isoforme tessuto-specifiche oppure vengono attivati durante un particolare stadio dello sviluppo. Il meccanismo più frequente con il quale si generano delle isoforme diverse è lo splicing alternativo, che consiste nell’assemblaggio differenziale di esoni durante la maturazione dell’RNA. Si stima che oltre il 60% dei geni umani produca due o più proteine mediante questo meccanismo. Molti esoni specificano domini proteici strutturali distinti che possono essere combinati in modo diverso nelle cellule dei diversi tessuti nei quali il gene è espresso. Pertanto a partire da un singolo gene possono venire generate proteine simili, ma non identiche che possono avere funzioni diverse nei vari tessuti. D. Regolazione traduzionale. Oltre all’emivita dell’mRNA, intervengono fattori d’inizio della traduzione e talvolta di proteine che si legano a regioni non tradotte dell’mRNA. E. Controllo post-traduzionale, Si basa su modificazioni della struttura/funzione, tagli proteolitici, degradazione delle proteine. Vi sono inoltre numerosi altri meccanismi da cui dipende il meccanismo dell’espressione genica, come l’attivazione dei fattori di trascrizione in seguito al legame di un ligando, per esempio un ormone, ad uno specifico recettore sulla superficie cellulare. Infatti l’espressione di molti geni è controllata da un ormone, da un fattore di crescita o da una molecola di segnale intracellulare (es. cAMP) i quali legandosi a recettori specifici determinano l’attivazione o inattivazione di determinati fattori di trascrizione (es. mediante la loro fosforilazione o defosforilazione). I fattori attivati interagiscono quindi con gli elementi di risposta presenti nel promotore e inducono l’espressione del gene corrispondente. 7. GLI ENZIMI DI RESTRIZIONE Un sito di restrizione o sequenza consenso viene definito come una particolare sequenza di DNA riconosciuta da un enzima di restrizione o endonucleasi come il punto in cui tagliare la molecola di DNA. Questi siti sono generalmente palindromici (cioè possono essere letti in entrambe le direzioni) la cui successione di basi è identica su entrambi i filamenti quando ciascuno di essi venga letto in direzione 5’ -> 3’. La sequenza riconosciuta non è unica e varia da enzima ad enzima, anche se con la stessa specificità di sequenza. Infatti, sebbene gli enzimi di restrizione isolati siano oltre 3500, le sequenze bersaglio che possono essere tagliate sono molto meno numerose (poco più di 200). I siti di restrizione sono delle normali sequenze di basi, lunghi dai 4 fino a diverse decine di paia di basi, per cui si possono trovare più o meno facilmente nel genoma. Il taglio prodotto dagli enzimi può generare frammenti di DNA con estremità piatte (blunt), o sporgenti ( 5’-protruding e 3’-protruding). Le estremità prodotte sono “appiccicose”, cioè possono formare ponti Idrogeno tra le due code a filamento singolo complementari. Le estremità coesive facilitano inoltre la reazione della DNA ligasi. L’enzima EcoRI produce un taglio sfalsato creando due estremità coesive a singolo filamento 12 al 5’ (5’-protruding): 5’...G/ AATTC.. 3’ 3’...CTTAA /G...5’ L’enzima HhaI opera un taglio sfalsato creando due estremità coesive a singolo filamento al 3’(3’-protruding ) : 5’… G C G /C… 3’ 3’… C/G C G… 5’ enzima Organismo di provenienza Sequenza d riconoscimento e posizione di taglio Pronuncia E.Coli RY13 G/A A T T C C T T A A/G Eco-ri-uno HindIII Haemophilus influenzae Rd A/A G C T T T T C G A /A Acca-ind-tre BamHI Bacillus amyloliquefaciens G/G A T C C C C T A G/G Bam-accauno EcoRI ENZIMI DI RESTRIZIONE. Sono prodotti dai batteri che li utilizzano per difendersi da un DNA estraneo, esempio un virus. Altri enzimi (le metilasi ) proteggono il DNA batterico grazie all’azione delle proprie endonucleasi di restrizione. Gli ER si indicano con un sistema di lettere e numeri che si riferisce al ceppo batterico da cui sono stati isolati. 8. MARCATORI GENETICI E POLIMORFISMI Il concetto di variabilita’ Il completamento del Progetto Genoma Umano, rendendo disponibile la sequenza del DNA umano, ha rilevato che ogni persona dimostra il 99.9% di identità genetica rispetto ad una qualsiasi altra. Questo risultato ha confermato che nel DNA è presente una piccola porzione, lo 01%, soggetta a variazioni all’interno di una popolazione. La somma di queste differenze costituisce la variabilità interindividuale caratterizzata principalmente da variazioni di sequenza definite polimorfismi. Il termine polimorfismo significa “esistenza di forme diverse”. In Genetica si definisce polimorfismo l’esistenza – ad un dato locus – di due o più alleli, ciascuno dei quali mostri una frequenza di almeno 1%. Le forme diverse, ossia le varianti genetiche possono riguardare un gene, vale a dire un tratto di DNA codificante una proteina (polimorfismo allelico), oppure un tratto di DNA non codificante (polimorfismo di sequenza). Dato che più del 98% del DNA umano è DNA non codificante, e che quindi la maggior parte di queste differenze è localizzata in sequenze non codificanti, il fenotipo di un polimorfismo di sequenza del DNA non è riconoscibile dall’esterno (ex. nei gruppi sanguigni). A causa dell’elevato numero di loci polimorfici, i polimorfismi di sequenza sono molto più frequenti dei polimorfismi allelici tradizionali (gruppi sanguigni, albinismo, colore degli occhi, ecc..) e conseguentemente più utili nella ricerca biologica e medica. Sulla base del tipo di variazione osservata e della distribuzione lungo il genoma, i polimorfismi possono essere classificati in: 1. Polimorfismi di Lunghezza dei Frammenti di Restrizione, o RFLP 2. i Polimorfismi del Singolo Nucleotide, o SNP 3. i Polimorfismi del numero delle ripetizioni in tandem VNTR (Variable Number of Tandem Repeats) che possono essere distinti in - minisatelliti - microsatelliti 13 9. POLIMORFISMO A SINGOLO NUCLEOTIDE o SNP Un polimorfismo a singolo nucleotide (o Single Nucleotide Polymorphism o SNP) è un polimorfismo (cioè una variazione a livello di una sequenza di acidi nucleici) che si presenta tra individui della stessa specie, caratterizzata da una differenza a carico di un unico nucleotide C T A A/G G T SNP A Gli SNPs sono sostituzioni di un singolo nucleotide di una base con un’altra. Naturalmente, possiamo avere 4 versioni per ogni SNP, una per ogni nucleotide, A, C, G, T e la distribuzione nella popolazione potrebbe risultare in una delle seguenti combinazioni. Sono stati individuati molti SNPs nella sequenza del DNA e la sfida per la ricerca è identificare gli SNPs correlati con un particolare effetto nel fenotipo. Gli SNP si verificano nella popolazione con una frequenza maggiore all’1%. Nel genoma umano si verificano SNPs all’incirca uno ogni 300 paia di basi. Questo significa che su 3 miliardi di nucleotidi presenti nel genoma umano avremo circa 10 milioni di SNPs. Gli SNPs costituiscono il 90% di tutte le variazioni genetiche umane. Un polimorfismo noto è quello dei gruppi sanguigni, ~20 loci. Non bisogna confondere una mutazione puntiforme con uno SNP ! Anche se si assomigliano, non sono la stessa cosa: - sono entrambe differenze di singoli nucleotidi, ma per parlare di SNP bisogna che questo sia presente in almeno l’1% della popolazione. - molte mutazioni correlate a malattie si trovano all’interno delle regioni codificanti del gene o in quelle regolatorie e interessano la proteina corrispondente a quel gene. Viceversa, gli SNPs non sono necessariamente localizzati nei geni, e non sempre alterano la funzione di una proteina. In base a quanto detto possiamo distinguere gli SNPs in due principali categorie: 1. Linked SNPs non si trovano all’interno dei geni e non alterano la funzione della proteina. 2. Causative SNPs alterano il funzionamento di una proteina, correlando con una malattia o con un tratto fenotipico. Si distinguono ancora 2 tipi di Causative SNPs: - Coding SNPs, localizzati nella regione codificante del gene, cambiano la sequenza di aminoacidi nella proteina. - Non Coding SNPs, situati nella regione regolatoria del gene, cambiano la quantità di proteina prodotta 14 I POLIMORFISMI DEL DNA sono utili come: - IDENTIFICATORI INDIVIDUALITA’ : • controllo relazioni parentali in famiglie con malattie mendeliane • Genetica di popolazione • Indagini di paternità • Indagini di medicina legale - MARCATORI GENETICI ANALISI DI LINKAGE • per identificare geni-malattia (diagnosi portatore) • mappaggio sia genetico (ordinamento dei geni sui cromosomi) che fisico (distanza fisica tra i geni) Come si individuano i Polimorfismi? Per analisi diretta della sequenza del DNA : - PCR + elettroforesi ( agarosio + et.br./acrilamide + et.br./acrilamide+fluoresc.) - microchip Un metodo utile per individuare gli SNPs è la valutazione dei cosiddetti polimorfismi di lunghezza dei frammenti di restrizione (Restriction fragment length polymorphisms) o RFLP. Se un allele contiene un sito di riconoscimento per un enzima di restrizione ed un altro no, la digestione dei due alleli genererà due frammenti di dimensione differente. In realtà oggi gli SNPs sono studiati principalmente attraverso i microarrays, che permettono l’analisi simultanea di centinaia di migliaia di diversi SNPs ed una veloce analisi elaborata da un computer. 10. APLOTIPI Al fine di trovare una associazione tra SNPs, viene considerata una serie di SNPs su un segmento più lungo di DNA. C T/C G A C T A A/G G A C C G/T A SNP SNP SNP Questi tre SNPs possono combinarsi in 23 ( ognuno dei tre SNPs con i due possibili nucleotidi), cioè otto differenti combinazioni C C C C C C C C T T T T C C C C G G G G G G G G A A A A A A A A C C C C C C C C T T T T T T T T A A A A A A A A AGTACCGA AGTACCTA GGTACCGA GGTACCTA AGTACCGA AGTACCTA GGTACCGA GGTACCTA POSSIBILI COMBINAZIONI DI SNPs Ogni combinazione di SNPs è chiamata APLOTIPO. Quindi, possiamo dire che in questa regione di DNA ci sono 8 possibili aplotipi. C C C C C C C C T T T T C C C C G G G G G G G G A A A A A A A A C C C C C C C C T T T T T T T T A A A A A A A A AGT AGT GGT GGT AGT AGT GGT GGT AC AC AC AC AC AC AC AC CGA CTA CGA CTA CGA CTA CGA CTA APLOTIPO 1 APLOTIPO 2 APLOTIPO 3 APLOTIPO 4 APLOTIPO 5 APLOTIPO 6 APLOTIPO 7 APLOTIPO 8 15 RICORDA ! Nel mondo della Genetica, ogni cosa è “in paia”: noi riceviamo un aplotipo dalla madre e uno dal padre. Questo significa che abbiamo due aplotipi ( o un paio di aplotipi). SNP PROFILE………….. che cos’è ? LA COPPIA DI APLOTIPI è lo “SNP PROFILE” 11. ANALISI DI LINKAGE (o di Associazione o di Concatenazione) Permette di determinare la posizione cromosomica del locus responsabile di un carattere genetico o di una determinata malattia rispetto a ”marcatori genetici” di cui è nota la posizione. Un marcatore genetico è un qualunque carattere che risponde alle seguenti caratteristiche: - è polimorfico - è facile da identificare e stabile nelle generazioni - segrega in modo mendeliano Quali sono ? RFLP,VNTR, STR, SNP. Ogni individuo possiede due copie di ciascun allele, uno ereditato dal padre e uno dalla madre. Durante la meiosi i cromosomi omologhi segregano separatamente. Quindi, se consideriamo due loci polimorfici situati su cromosomi diversi, un particolare allele del primo locus avrà il 50% di probabilità di segregare insieme ad un particolare allele del secondo locus. Se consideriamo invece due loci situati sullo stesso cromosoma, ci aspettiamo che i loro alleli vengano ereditati insieme ( cioè co-segregano) in base alla distanza che intercorre tra oro sul cromosoma. Più grande è la loro distanza, più è facile che un evento di crossing over separi i due alleli. L’evento di crossing over dà luogo alla formazione di gameti ricombinanti La probabilità che avvenga questa ricombinazione, espressa in percentuale, è nota come frequenza di ricombinazione. La frequenza di ricombinazione ci dice quindi la distanza genetica tra loci diversi. 16 L’analisi di concatenazione nell’uomo si basa sullo studio degli alberi genealogici e sulla ricerca delle meiosi informative (cioè quelle in cui si può identificare se un gamete è o non è ricombinante). Allo stesso modo si può mappare anche un numero consistente di loci, considerandone sempre due per volta. Un altro approccio è quello di studiare gli aplotipi, ovvero l’ordine degli alleli sui rispettivi cromosomi omologhi. E’ quindi necessaria una rielaborazione statistica dei dati riguardanti l’associazione tra i marcatori e la comparsa della malattia. Quali sono i problemi di uno studio di linkage ? 1. le famiglie da analizzare solitamente contano pochi individui 2. in famiglie poco numerose vi è un basso numero di ricombinanti 3. i risultati così ottenuti hanno una bassa significatività statistica Quali sono le possibili soluzioni ? Analizzare più famiglie insieme ! Quali popolazioni studiare ? 1. Popolazioni geneticamente isolate 2. Popolazioni giovani, con poche generazioni dal momento del fondatore, con pochi crossing over, con poche ricombinazioni 3. Popolazioni omogenee, cioè con poche famiglie fondatrici Esempi di popolazioni che presentano queste caratteristiche sono: - I Finlandesi: geneticamente e linguisticamente molto diversi dai loro vicini slavi. La popolazione è giovane in quanto fondata circa 2000 anni fa da poche famiglie fondatrici - I Sardi in Italia LINKAGE EQUILIBRIUM: indica una combinazione casuale di alleli a loci associati. Consideriamo per esempio il caso di due loci associati 1 e 2 con 2 possibili alleli ciascuno (A e a per il locus 1 e B e b per il locus 2) Gli aplotipi possibili in una determinata popolazione (AB Ab, aB, ab) si verificheranno con una frequenza che è il prodotto delle frequenze dei singoli alleli per ciascun aplotipo. Esempio: A = 0,2 B = 0,6 a = 0,8 b = 0,4 4 aplotipi possibili: AB Ab aB ab AB 0,2 x 0,6 = 0,12 Ab 0,2 x 0,4 = 0,08 aB 0,8 x 0,6 = 0,48 ab 0,8 x 0,4 = 0,32 } equilibrio di linkage LINKAGE DISEQUILIBRIUM: indica una combinazione non casuale di alleli a loci associati. Il linkage disequilibrium è spesso la conseguenza di un effetto "founder" (fondatore), cioè di una mutazione in un singolo individuo. Perchè l'effetto fondatore sia evidenziabile in una popolazione è necessario che i due loci siano vicini, in maniera tale che gli eventi di ricombinazione siano rari tra i due loci, e che non sia trascorso abbastanza tempo dalla comparsa del fondatore poichè la ricombinazione puo' ristabilire nel tempo l'equilibrio. Il linkage disequilibrium (LD) indica la presenza di associazione preferenziale tra specifici alleli relativi a due o più loci, presenti sullo stesso cromosoma, che costituiscono di solito un particolare aplotipo ancestrale, diffuso nella popolazione in cui è rilevato, perché trasmesso lungo la discendenza da un comune progenitore. Riprendiamo l’esempio dei 4 aplotipi AB, Ab, aB, ab. Se si trovassero questi valori AB = 0,04 – Ab = 0,16 + aB = 0,56 + ab = 0,24 – } linkage disequilibrium 17 12. TECNICHE UTILIZZATE PER L’ANALISI MOLECOLARE DEL DNA ESTRAZIONE DEL DNA. Il DNA può essere estratto da qualunque cellula nucleata. I tipi cellulari più utilizzati sono rappresentati dai leucociti di sangue periferico, colture cellulari (fibroblasti, amniociti, villi coriali). L’isolamento del DNA richiede l’utilizzo di enzimi capaci di distruggere le membrane cellulari e nucleari e di solventi organici in grado di separare le proteine dagli acidi nucleici. Nella procedura di estrazione del DNA genomico, le cellule vengono lisate e sottoposte a trattamento proteolitico con Proteinasi K. Il DNA purificato viene quindi raccolto con una soluzione di eluizione. La determinazione quantitativa della concentrazione del DNA estratto, calcolata in ng/µl, viene effettuata mediante lettura spettrofotometrica valutando l’assorbanza del campione a 260 nm. oppure mediante corsa su gel. LA TECNICA DELLA PCR L’introduzione della PCR, la tecnica che consente di amplificare selettivamente un tratto di DNA, ha rivoluzionato la genetica molecolare e le sue applicazioni sono praticamente infinite. Uno degli ambiti di utilizzo è la diagnosi di malattie genetiche mediante analisi di RFLP. L’utilizzo della PCR semplifica molte cose. Ad esempio, la PCR consente di analizzare uno specifico tratto di DNA, invece di dover lavorare su tutto il DNA nucleare di una cellula, ossia sul DNA genomico. La PCR (Polymerase Chain Reaction) è una tecnica di amplificazione in vitro di un frammento di DNA di cui si conosca la sequenza nucleotidica delle regioni terminali. Il principio è molto semplice. Data una sequenza di DNA genomico a doppio filamento e due corte sequenze oligonucleotidiche (primer), di cui una complementare ad un tratto di filamento a una estremità del DNA da amplificare (forward primer) e l’altra complementare ad un altro tratto posto all’altra estremità (reverse primer), in presenza di una DNA polimerasi termostabile e di una miscela di desossinucleotiditrifosfati (dNTPs), in appropriate condizioni di reazione, è possibile copiare numerosissime volte (30-40 volte) il tratto compreso tra i due primer, semplicemente facendo variare ciclicamente la temperatura di reazione. Infatti, raggiunta la temperatura di denaturazione (92-95°C), la doppia elica si apre (fase di denaturazione), rendendo disponibile lo stampo per la sintesi delle catene complementari. Se la temperatura si abbassa, in virtù delle loro minori dimensioni e della loro concentrazione, i primer si legheranno (fase di appaiamento o annealing) al DNA stampo prima che si rinaturi e in presenza di una DNA polimerasi con un optimum di temperatura elevato (circa 72°C), inizierà la sintesi di DNA a partire dai primer (fase di sintesi del DNA o extension), procedendo lungo i filamenti singoli. Al termine del primo ciclo di PCR da una doppia elica di DNA se ne ottengono due. Ripetendo il ciclo denaturazione – annealing – extension numerose volte (in genere da 30 a 40 volte), si ottiene una massiccia amplificazione specifica di un dato tratto di DNA, corrispondente a DNA in quantità tale da essere visualizzabile in un gel di agarosio mediante colorazione specifica. La PCR ha rivoluzionato la genetica molecolare. Le applicazioni della PCR sono praticamente infinite. I principali ambiti di utilizzo sono la diagnosi prenatale delle malattie genetiche e le indagini di medicina legale. I termociclatori Il successo della PCR è dovuto in gran parte alla possibilità di far avvenire l’intero processo in modo automatico all’interno di strumenti detti termociclatori (thermal cyclers) in grado di variare ciclicamente la temperatura tra le varie fasi di ogni ciclo di PCR. Un esempio di profilo di amplificazione standard impostato mediante un termociclatore è il seguente: 1. denaturazione del DNA: 30 sec. a 94°C 2. appaiamento (annealing) dei primer: 30 sec. a 50°-60°C 35 cicli 3. sintesi (extension) di DNA: 30 sec-5 min. a 72°C Il successo della PCR è stato possibile grazie anche all’uso di una DNA polimerasi termostabile estratta da batteri termofili (che vivono ad elevate temperature). Una DNA polimerasi utilizzata nelle reazioni della PCR è la Taq polimerasi, estratta dal batterio Thermus aquaticus. L’isolamento di DNA polimerasi termostabili ha sollevato gli operatori dall’ingrato compito di aggiungere enzima fresco ad ogni ciclo di reazione! 18 Scelta dei primer Per ogni PCR, è necessario usare due primer (forward e reverse). La scelta della coppia di primer è critica per una buona riuscita della PCR, ovvero per ottenere l’amplificazione di un tratto di DNA in modo specifico. I primer devono essere “disegnati” a monte e a valle dei siti di restrizione. Si tratta di oligonucleotidi, con dimensioni comprese tra le 15 e le 30 basi che ibridano su filamenti opposti in posizioni fiancheggianti la regione di interesse del DNA. Per minimizzare la formazione di artefatti è importante che le loro sequenze non contengano basi complementari (all’interno dello stesso primer o tra i due primer); inoltre la Temperatura di fusione dei due oligonucleotidi deve essere identica o almeno molto vicina. DIGESTIONE CON ENZIMI DI RESTRIZIONE (ER) Come abbiamo detto, gli ER sono endonucleasi che tagliano il legame fosfodiesterico nel DNA a doppia elica a livello di sequenze specifiche (siti di restrizione). La digestione con ER va condotta a 37°C in una soluzione tampone (fornita insieme all’enzima dal produttore) che garantisce le condizioni ottimali (di salinità e pH) per la digestione. Il tempo di incubazione varia a seconda se si digerisce DNA genomico o frammenti di DNA corti. Nel primo caso la digestione richiede almeno 8 ore (o tutta la notte). Nel secondo caso sono sufficienti da 1-4 ore. Gli ER sono reagenti costosi e delicati. Temono le contaminazioni (usare precauzioni nel prelevare l’enzima dalla soluzione stock) e l’inattivazione (si devono conservare a -20°C e, al momento dell’uso, mantenere sempre in un bagno di ghiaccio). ELETTROFORESI SU GEL DI AGAROSIO E’ una tecnica che consente di separare in base alle loro dimensioni (peso molecolare) molecole dotate di carica, facendole migrare su un gel in presenza di un campo elettrico. Il gel può essere immaginato come una rete tridimensionale attraverso le cui maglie migrano le molecole sotto l’azione di un campo elettrico. Il campo elettrico è generato da un apparecchio, detto alimentatore. Per separare molecole di DNA si usano gel di agarosio. Le molecole di DNA sono cariche negativamente per la presenza di gruppi fosfato e migrano dal polo negativo (catodo) verso il polo positivo (anodo). Per un certo intervallo di pesi molecolari, la velocità di migrazione è funzione del loro peso molecolare: tanto più grande è la molecola di DNA, tanto minore è la velocità di migrazione. E, viceversa, tanto più piccola è la molecola di DNA, tanto più velocemente migra. Le molecole di DNA di diversa lunghezza vengono pertanto separate in base alla diversa velocità di migrazione. Per poter determinare la lunghezza delle molecole di DNA in esame separate mediante elettroforesi, vengono “caricati” sul gel anche i cosiddetti marcatori di peso molecolare, ossia una miscela di frammenti di DNA di cui è noto il peso molecolare. Confrontando la posizione dei frammenti a peso molecolare noto con quella dei frammenti di DNA in esame, è possibile calcolarne il peso molecolare, ossia la lunghezza. Dato che il peso molecolare di un frammento di DNA è proporzionale al numero di coppie di nucleotidi (basi) che lo costituiscono, di solito esso viene espresso in paia di basi (bp). La separazione elettroforetica dura circa 45 minuti circa. Al termine, i vari frammenti di DNA, essendo incolori, possono essere visualizzati, con particolari sistemi di colorazione. Il DNA delle diverse classi di peso molecolare è visibile sotto forma di bande distinte: sono le cosiddette bande di DNA. Comunemente, per poter visualizzare il DNA, durante la preparazione del gel si aggiunge all’agarosio il bromuro di etidio, una sostanza che ha la proprietà di legarsi al DNA e di emettere fluorescenza se esposta a luce UV. Alla fine della 19 corsa, le bande si visualizzano esponendo il gel alla luce ultravioletta. Il bromuro di etidio va maneggiato con estrema cautela in quanto è un agente intercalante del DNA e, come tale, ha proprietà mutagene. Noi utilizzeremo un intercalante non tossico che non richiede precauzioni particolari. 20 BITTER RECEPTOR GENE TAS2R38. LA VARIAZIONE GENETICA ASSOCIATA ALLA PERCEZIONE DEL GUSTO AMARO INTRODUZIONE. IL GUSTO Il gusto è uno dei cinque sensi i cui recettori sono costituiti dai calici gustativi presenti nelle papille gustative della lingua, nel palato molle, nella faringe, nelle guance e nell' epiglottide. Il gusto dipende dalla percezione sinergica di cinque gusti fondamentali: amaro, aspro, dolce, salato e umami. Lo spazio gustativo è continuo. I profili di risposta dei diversi stimoli dolci possono essere estremamente diversi gli uni dagli altri, e lo stesso vale per gli stimoli amari. Anche gli stimoli amari sono molto diversi gli uni dagli altri. Oltre agli stimoli dolce e amaro, nello spazio gustativo sono situati altri stimoli di qualità non definita, in diverse direzioni dello spazio. Lo spazio gustativo è pluridimensionale. Nei mammiferi la percezione gustativa si realizza attraverso le gemme gustative, unità morfologiche e funzionali specializzate, che sono localizzate in tutto l’epitelio orale, sulla lingua, sul palato e sulla faringe. Le gemme gustative, a seconda delle specie, contengono aggregazioni di 50–150 cellule, includendo cellule precursori, cellule di supporto e cellule recettrici del gusto. Sulla lingua, le gemme gustative sono localizzate su specifiche protrusioni, le papille, che vengono generalmente classificate in tre tipi morfologici con proprietà funzionali distinte. Si riconoscono papille circumvallate, fungiformi e foliate. Le papille circumvallate si trovano prevalentemente nella zona posteriore della lingua, in un numero di circa 1000 nell’uomo e sono particolarmente sensibili alle sostanze amare. Le papille foliate sono localizzate nella porzione latero-posteriore della lingua, contengono da dozzine a centinaia di gemme gustative e sono sensibili al salato e all’amaro. Le papille fungiformi contengono una singola o poche gemme gustative, sono localizzate nella porzione anteriore della lingua sembrano mediare la percezione di composti dolci. Rappresentazione schematica dei tre tipi delle papille gustative e loro localizzazione sulla superficie dorsale della lingua. Nelle figura sono evidenziate le regioni specializzate per definiti stimoli gustativi (amaro, aspro, dolce e salato). Si nota che mentre differenti aree della lingua presentano forti preferenze per specifiche modalità gustative, esiste anche una significativa sovrapposizione tra le varie regioni. I tre tipi di papille gustative sono mostrate ingrandite e ne è mostrata la localizzazione nelle rispettive regioni . Le cellule deputate al riconoscimento di stimoli gustativi sono cellule epiteliali modificate connesse al cavo orale attraverso un sottile processo simile ad un dendrite. Le cellule recettrici del gusto presentano proprietà eccitatorie e sono attivate da stimoli gustativi. Un’apertura alla superficie dell’epitelio, il poro gustativo permette l’accesso degli stimoli chimici ai recettori localizzati sui microvilli apicali delle cellule gustative. 21 Il recettore, attivato in seguito alla stimolazione, genera una cascata trasduzionale che culmina nel rilascio di neurotrasmettitori al livello delle sinapsi con le fibre nervose (nervi facciale, vago e glossofaringeo) alla base delle gemme gustative. Le fibre nervose eccitate conducono lo stimolo, via talamo, ai centri corticali del gusto, dove l’informazione è integrata e processata. Schema di recettore gustativo. La superficie apicale contiene sia canali sia recettori accoppiati a proteine G che sono attivati da stimoli chimici. La superficie baso-laterale contiene canali al Na, K e Ca voltaggio dipendenti, come pure un apparato per la trasmissione sinaptica mediata dalla serotonina. I recettori presentano un corto dominio amino-terminale extracellulare e sequenze conservate in sette domini trans-membrana. La percezione di ognuno dei sapori fondamentali è associata ad una particolare via di trasduzione del segnale che schematicamente può essere riportata a due tipologie principali: - recettori legati a proteine G per l’amaro, il dolce e umami - recettori legati a canali ionici di membrana per aspro e salato 22 Dalla stimolazione della cellula si ottiene un potenziale di recettore che stimola l'ingresso di ioni Calcio nella cellula determinando la liberazione di neurotrasmettitori a livello basale e la genesi di un potenziale d'azione nelle fibre afferenti. Il gusto amaro viene trasdotto secondo almeno tre vie possibili. Nella prima, sostanze come la chinina determinano blocco dei canali apicali del potassio. Un secondo meccanismo sembra essere legato ad un particolare proteina G, la quale attiva una Fosfodiesterasi che fa diminuire le concentrazioni intracellulari di cAMP e cGMP. Il terzo consiste sempre nell'attivazione di una proteina G la quale attiva una fosfolipasi C che fa aumentare la concentrazione di IP3 il quale a sua volta determina liberazione di ioni calcio dai depositi intracellulari, depolarizzando la cellula. I meccanismi di trasduzione - come già detto - sono diversi a seconda del tipo di sostanza chimica. Ad esempio, la stimolazione gustativa prodotta dall’NaCl è caratterizzata dal passaggio dello stesso ione attraverso canali ionici amiloride sensibili. Il canale amiloride sensibile è permeabile anche agli ioni H+, quindi la trasduzione delle sostanze che vengono percepite come aspre è dovuta ad un ingresso di questi ioni attraverso canali Na+ sensibili all’amiloride. Acido e salato, in relazione alle loro concentrazioni nella saliva, interferiscono in parte tra loro a livello periferico. Le differenze che si osservano nella capacità di percepire il gusto dei diversi sali di Na+ potrebbero anche dipendere dalla diversa permeabilità dei rispettivi anioni attraverso le giunzioni strette e dalla conseguente diversa capacità di interessare i canali ionici localizzati a livello delle membrane basali laterali delle cellule gustative. La trasduzione dei segnali evocati dai composti dal sapore dolce comporta l’attivazione di recettori accoppiati a proteine G (GPCR) presenti sulla superficie apicale delle cellule gustative. Negli zuccheri, l’attivazione di questi recettori (GPCR) depolarizza le cellule gustative per attivazione dell’Adenilato Ciclasi, che a sua volta aumenta la concentrazione di cAMP che causa la chiusura diretta o indiretta dei canali del potassio presenti sulla membrana basale laterale. I dolcificanti sintetici come la saccarina attivano recettori accoppiati a proteine G (GPCR) differenti, che a loro volta attivano la Fosfolipasi C con formazione di Inositolotrifosfato (IP3) e Diacilglicerolo (DAG). Un aumento di IP3 fa aumentare la concentrazione intracellulare di Ca++, determinando il rilascio del trasmettitore. Un aumento del DAG attiva la proteina chinasi A (PKA), che a sua volta fosforila e causa la chiusura dei canali K+ della membrana basale laterale, amplificando in questo modo l’effetto. Entrambe queste vie di percezione delle sostanze dolci possono coesistere nella stessa cellula gustativa. I composti dal sapore amaro comprendono molte classi chimiche distinte. Alcune di queste sono alcaloidi come il chinino che, bloccando i canali al K+, determina la depolarizzazione delle membrana. Altri come la caffeina, gli L-amminoacidi, l’urea e anche sali come l’MgSO4 non utilizzano gli stessi recettori o le stesse vie di trasduzione del segnale. Il gusto umami, verrebbe trasdotto da un particolare tipo di recettore metabotropico del glutammato che viene anche espresso in alcune regioni cerebrali. Le grosse molecole lipidiche sono scisse dalla lipasi linguale in molecole di minor peso molecolare, capaci di stimolare i recettori gustativi. Inoltre, le membrane apicali dei bottoni gustativi contengono trasportatori degli acidi grassi, molecole che permettono l’entrata degli acidi grassi nelle cellule. Come abbiamo detto, la maggior parte dei meccanismi di trasduzione determina la depolarizzazione della membrana della cellula gustativa (potenziale del recettore), che a sua volta determina aumento della concentrazione di Ca++ per apertura di canali voltaggiodipendenti o per mobilizzazione da riserve intracellulari. L’aumento del Ca++ provoca l’esocitosi del mediatore chimico e la conseguente trasmissione del segnale alle fibre afferenti gustative primarie. Le fibre afferenti che convogliano le informazioni gustative alle strutture centrali appartengono a tre paia di nervi cranici (VII, IX, X) e possono avere funzioni diverse a seconda della loro provenienza dalle diverse parti della lingua. Ogni fibra può rispondere, in genere, a 2, 3 o tutte e 4 le qualità gustative, anche se con intensità che sono differenti. La porzione anteriore della lingua è innervata da un ramo del nervo facciale (corda del timpano VII). Infatti i bottoni gustativi delle papille fungiformi e delle papille fogliate, sono innervate da tale ramo. 23 Organizazione del sistema gustativo umano. (A) Il disegno a sinistra mostra il rapporto tra i recettori della cavità orale e della parte superiore del canale alimentare, e il nucleo del tratto solitario del bulbo. La sezione frontale sulla destra mostra il nucleo ventrale posteriore mediale del talamo e le sue connessioni con le regioni gustative della corteccia cerebrale. (B) Schema delle vie fondamentali deputate all’elaborazione delle informazioni gustative La sensibilità gustativa per un determinato stimolo varia grandemente tra gli individui e condiziona la scelta del cibo e quindi lo stato nutrizionale dell’individuo. Il sapore di un cibo o di una bevanda è infatti, fondamentale nel determinarne la commestibilità e l’appetibilità; il rifiuto di un cibo a causa del suo sapore sgradevole è importante per la sopravvivenza di molte specie animali. Di norma, le sostanze dolci, fonte di energia per l’organismo, provocano una sensazione piacevole e l’innesco di riflessi di salivazione, di deglutizione e di preparazione del tubo digerente alla digestione e all’assorbimento. Il sapore amaro provoca sovente il rifiuto del cibo o della bevanda e, se molto intenso, stimola il riflesso del vomito: questo è presumibilmente in relazione al fatto che la maggior parte delle sostanze tossiche presenti in natura ha sapore amaro. La sensibilità gustativa è normalmente alta per il gusto amaro, in accordo con quanto detto sulla funzione di monitoraggio della presenza di sostanze potenzialmente tossiche rispetto a quelle invece che possono essere utili per l’organismo. Il grado di piacevolezza di un sapore è tuttavia soggettivo e può essere influenzato dall’esperienza e dalle necessità nutrizionali. Sia l’animale che l’uomo tendono a rifiutare un cibo dal quale in passato siano stati intossicati. Lo stesso cibo può essere avvertito come molto gradevole e desiderabile all’inizio del pasto e sgradevole quando viene raggiunta la sazietà. In base alle informazioni gustative e olfattive si ha la capacità di operare una scelta tra diversi cibi e preferire quello che fornisce l’apporto nutritivo più consono alle esigenze dell’organismo. È interessante ricordare che il sapore può essere in relazione alla concentrazione. Ad esempio, il sale da cucina a bassa concentrazione viene percepito come dolce, ad alta concentrazione come amarognolo. La sensibilità gustativa può variare grandemente anche in funzione dell’età e del sesso dell’individuo. È noto che le donne mostrano una maggiore sensibilità rispetto agli uomini per gli stimoli dolci e salati, e una sensibilità minore per gli stimoli acidi. E’ noto che anche l’invecchiamento può determinare modificazioni della sensibilità gustativa che possono essere dovute anche a fattori diversi dalle sole modificazioni dell’invecchiamento delle strutture coinvolte. Non è stato infatti dimostrato che il numero dei calici gustativi e delle papille diminuisca con l’invecchiamento. In realtà, molti dei farmaci che sono assunti dai soggetti anziani alterano la percezione gustativa e influiscono negativamente sull’output salivare (peggiorando ulteriormente il senso del gusto). Anche la perdita delle capacità olfattive, legata all’età o a patologie, può giocare un ruolo importante nel diminuire la sensibilità gustativa. La capacità di percepire l’amaro ha avuto e sicuramente ha ancora delle notevoli implicazioni e conseguenze, sia positive che negative. Vediamone alcune: a) in passato ha comportato sicuramente un importante vantaggio selettivo permettendo di evitare l’ingestione di cibi amari che spesso in natura sono tossici o velenosi; 24 b) porta a eliminare dalla dieta cibi come le crucifere che, se introdotti in consistenti quantità, interagiscono con il metabolismo dello iodio producendo gozzo, una grave malattia della tiroide (a riprova di ciò va ricordato che i deficit tiroidei sono rari tra i taster); c) porta a preferire diete povere in frutta e vegetali (cosa che può tradursi in una ridotta prevenzione di tumori, specie quelli intestinali); d) si associa mediamente a una massa corporea inferiore valutata come indice di massa corporea o BMI (body mass index). I non-taster consumano una maggiore varietà di cibi e sono associati ad un maggior peso corporeo. E’ stata anche trovata una forte associazione tra lo stato di Non Taster e una maggiore adiposità nelle donne. e) comporta un numero percentualmente inferiore di carie. Per capire meglio l’influenza della percezione dell’amaro su malattie complesse, come l’obesità, sono state studiate popolazioni isolate geneticamente e geograficamente. La diversa capacità percettiva è un tipico carattere genetico ereditario ed è dovuta a un gene chiamato TAS2R38, responsabile della produzione di recettori gustativi per l’amaro. Uno studio pubblicato da un gruppo di ricercatori dell’Istituto di Biologia Evolutiva di Barcellona dimostra come anche l’uomo di Neanderthal, 48.000 anni fa, fosse in grado di percepire il gusto amaro dei cibi che ingeriva, proprio come l’uomo moderno. Lo studio genetico condotto dal gruppo del dottor Carles Lalueza-Fox dell’Istituto di Biologia Evolutiva di Barcellona ha analizzato il gene TAS2R38 in un campione di osso prelevato da un uomo di Neanderthal rinvenuto nel 2000 a El Sidron, nel nord della Spagna. I ricercatori hanno estratto il DNA dal fossile d’osso e hanno ottenuto la sequenza del gene specifico per il recettore delle sostanze amare e hanno scoperto che anche 48.000 anni fa, i nostri antenati erano in grado di percepire il gusto amaro dei cibi. Il dato rilevante è che nella loro popolazione, come oggi nella nostra, esistevano due gruppi di persone: quelle capaci di percepire l’amaro e quelli insensibili a questa sostanza. La ricerca, pubblicata su Biology Letters, dimostra la vicinanza genetica tra l’Homo Sapiens e l’Homo Neanderthalensis, che condividono un gene ereditato da un comune antenato, vissuto più di 300.000 anni fa. Hypotheses for the origin of PTC taster and nontaster alleles. (Left) Fisher et al. ‘‘Single Origin’’ hypothesis. Under this hypothesis, the taster (T) and nontaster (t) alleles diverged prior to the human–chimpanzee species divergence. Then both alleles were maintained separately in each species up to the present time. The maintenance of both alleles for such an extended period [Fisher (1939b) thought that it must be 1 million generations, or 20–30 million years] is unlikely if balancing selection has not been active because one allele or the other would be expected to go to fixation. (Right) Wooding et al.’s (2006) ‘‘Separate Origin’’ hypothesis.Under this hypothesis, nontaster alleles were derived from taster alleles twice— once in each species—after the human–chimpanzee species divergence. (Arrows indicate divergence events) L’abilità di percepire le sostanze amare rappresenta quindi una difesa contro le tossine vegetali e i veleni spesso contenuti in piante dal sapore amaro. I risultati di questo studio aprono però anche altre prospettive. La presenza di individui portatori del gene che li rende insensibili all’amaro, anche dopo 48.000 anni di evoluzione, suggerisce il fatto che questo tratto genetico si sia conservato per un vantaggio dei portatori. Questi individui potrebbero essere in grado di percepire la presenza di sostanze ancora sconosciute e inoltre la capacità di mangiare anche cibi amari sembra essere importante, viste le proprietà medicinali di alcuni cibi amari. Il TAS2R38 non è l’unico recettore per l’amaro. Il fatto che oggi circa il 30% della popolazione non percepisce l’amaro se stimolato dal PTC (Phenylthiocarbamide, una sostanza utilizzata per eseguire il test del gusto e discriminare tra individui capaci o incapaci di percepire l’amaro) significa 25 che evolutivamente c’è stato qualche svantaggio, altrimenti ci sarebbe un equilibrio al 50% tra taster e non taster. Questa situazione però potrebbe essere stata almeno in parte controbilanciata per far sì che i non taster siano ancora presenti nella popolazione. Tre diversi recettori sono stati sinora identificati per il gusto dolce, chiamato dolce-umami (T1Rs), e almeno 30 per l’amaro (T2Rs). Diversi altri recettori, molecole e vie di trasmissione del segnale che fornisce poi la percezione del gusto devono ancora essere identificati. Diversi composti presenti nei cibi come amminoacidi, peptidi, esteri e lattoni, fenoli e polifenoli, flavonoidi e terpeni, metilxantine (caffeina), sulfanilammidi (saccarina) ecc. stimolano la percezione dell’amaro. La sensibilità gustativa al PROP viene spesso usata come indice generale della percezione chemosensoriale dato che è stato dimostrato essere associata alla percezione di una vasta gamma di stimoli gustativi, come molti altri stimoli dal sapore amaro, sostanze dolci, sostanze irritanti e anche con la percezione delle sostanze grasse. Dato il valore nutrizionale che i lipidi hanno nella dieta la relazione tra sensibilità al PROP e accettabilità per cibi contenenti grassi risulta essere di notevole interesse. Diversi studi hanno riportato che i PROP “non tasters” rispetto ai “tasters” presentano una più bassa abilità nel riconoscere e percepire i grassi contenuti i cibi, mostrando una più alta accettabilità nei confronti di questi ultimi. Questa interpretazione è supportata da altri risultati che affermano, per l’appunto, una minore accettabilità degli individui PROP-sensibili nei confronti di cibi ad alto contenuto di grassi. Esistono evidenze a favore del fatto che i “PROP nontaster” hanno un più alto indice di massa corporea (BMI) rispetto ai “taster”, ma esistono anche dati che contraddicono questa affermazione. Nel comportamento alimentare i fattori cognitivi possono giocare un ruolo di fondamentale importanza, così come possono influire sullo stato del peso corporeo e della salute dell’uomo! Il controllo cognitivo dell’assunzione di cibo è associato ad un minor apporto energetico, una minor assunzione di grassi e un più frequente utilizzo di prodotti a basso contenuto lipidico. Queste osservazioni suggeriscono che nella scelta del cibo le persone che ne controllano volontariamente l’assunzione sono condizionate più fortemente dal loro peso corporeo che dallo stimolo gustativo Quindi, l’associazione tra sensibilità al PROP e BMI potrebbe essere mascherata nelle persone che bloccano volontariamente l’assunzione di cibo in modo da controllare il proprio peso corporeo, mentre sarebbe evidente un’associazione negativa solo nelle persone che mangiano liberamente. Tutti questi risultati portano all’ipotesi di una correlazione inversa tra il PROP status e l’indice di massa corporea (BMI). Altri lavori mostrano invece che non ci sia nessuna correlazione tra il PROP-taster status e questa variabile. Le differenze individuali della sensibilità al PROP potrebbero essere correlate con variazioni nella composizione chimicofisica della saliva. Fox per primo suggerì che la composizione salivare potrebbe essere uno dei fattori responsabili delle differenze individuali gustative, e ipotizzò che queste differenze possano dipendere dalla presenza o assenza nella saliva di alcuni costituenti organici come colloidi e/o proteine. Tra le proteine salivari di particolare interesse è la gustina, una anidrasi carbonica. La gustina è una zinco-metallo-proteina secreta dalle cellule acinose delle ghiandole salivari parotidi, sottomandibolari, ghiandole von Ebner nell’uomo. Diminuzioni della secrezione salivare di gustina sono state associate con ridotta o distorta funzione gustativa e olfattoria. GENETICA DEL GUSTO AMARO Gli studi sulla genetica del gusto iniziarono casualmente nel 1931 nel laboratorio del chimico britannico A. F. Fox. Un giorno, durante un esperimento per sintetizzare il composto PTC (feniltiocarbamide), una sostanza di sapore amaro appartenente alla famiglia delle tiouree, avvenne un’esplosione che disperse la sostanza nell’aria. Alcuni colleghi presenti percepirono un forte sapore amaro mentre Fox non percepì assolutamente nulla. A questa prima osservazione sono seguiti diversi studi che hanno dimostrato che l’incapacità di percepire il PTC (o un composto simile chiamato PROP) varia da popolazione a popolazione da un minimo del 3% nell’Africa occidentale a oltre il 40% in India. È interessante notare come circa il 25% della popolazione non percepisce assolutamente il sapore di questo costituente, anche alle alte concentrazioni. Queste persone hanno una 26 particolare insensibilità a questa sostanza veramente amara, diversamente dal restante 75%. La feniltiocarbamide è uno dei composti più amari, la cui soglia di identificazione è 6,5 · 10-8 mole/l (circa 10 mg/l), pertanto sono necessarie circa 1013 molecole/ 1 ml per identificarne il sapore amaro. L'intensità dell'amaro dipende dalla sostanza assaggiata, dalla sua concentrazione e dal tempo di reazione nella bocca. La percezione dell'amaro si sviluppa e raggiunge il suo massimo in pochi secondi, in contrasto con altri gusti che vengono percepiti quasi istantaneamente. Le soluzioni contenenti sostanze amare che vengono degustate a bassa temperatura tendono ad essere riconosciute più amare rispetto a quelle percepite a temperature più alte. Se presente nella soluzione, l'alcol intensifica la percezione dell'amaro, mentre i composti dolci la riducono. I composti amari più noti ed utilizzati nella ricerca come sostanze di riferimento nei test gustativi sono il chinino, la naringina e la caffeina. Tale caratteristica influenza le nostre abitudini alimentari e ci induce a preferire o eliminare dalla dieta determinati cibi. L'amaro è percepito come un avvertimento contro gli alimenti pericolosi, come indicazione della presenza di veleno o di cibi alterati. Viene anche associato ai preparati medicinali e alle erbe terapeutiche. D'altra parte, l'amaro costituisce in molti casi la necessaria componente del gusto in certi tipi di alimenti, consentendone un'estensione del flavor. Nella nostra popolazione il 30% circa è classificato come non taster, mentre il 70% è taster. I taster sono quelli che hanno una percezione molto elevata e che in genere allontanano subito la sostanza amara con repulsione (soglia di circa 1,0 x 10-4 mol al litro), i medium taster e i non taster sono invece quelli che percepiscono molto poco o per nulla l’amaro (soglia maggiore di 2,0 x 10-4 M). I taster, più sensibili all’amaro, non prediligono i cibi come le crucifere ricchi in tiouree (cavoli, broccoli, cavoletti di Bruxelles, rape ecc.), quelli contenenti caffeina, chinino, isoumuloni (amaro della birra), naringina (pompelmo); sono inoltre più sensibili alla percezione del piccante (irritante per effetto di sostanze quali la capsaicina del chili, la piperina del pepe nero, e lo zingerone presente nel ginger) e del grasso per una maggiore presenza di terminazioni del nervo trigemino sulla lingua e nel cavo orale. Ovviamente i non taster tendono a comportarsi in maniera completamente opposta. PROP sensitivity Food Perception Preference Selection Se ora ricordiamo la notevole diversa distribuzione di NON TASTER nelle popolazioni dell’Africa Occidentale (3%) rispetto a quelle dell’India (40%), sulla base di quanto sinora detto ci aspetteremo in India una cucina mediamente più piccante, grassa e con presenza di cibi amari rispetto a quella dell’Africa Occidentale. Distribuzione dei genotipi PAV e AVI in differenti aree geografiche/ gruppi etnici 27 I RECETTORI DELL’AMARO: LA FAMIGLIA TAS2R La famiglia dei geni codificanti chemiorecettori di composti amari, TAS2R, è stata inizialmente individuata mediante un approccio combinato bioinformatico e di analisi genetica. Alcuni dei geni, identificati mediante analisi di banche dati genomiche murine e umane, mappavano all’interno di loci associati con varie sensibilità a composti amari e suggerivano che i geni codificanti i recettori del gusto amaro, appartenessero alla famiglia TAS2R. John E. Hayes Allelic Variation in TAS2R Bitter Receptor Genes Associates with Variation in Sensations from and Ingestive Behaviors toward Common Bitter Beverages in Adults Chem. Senses 36: 2011 Abstract: The 25 human bitter receptors and their respective genes (TAS2Rs) contain unusually high levels of allelic variation, which may influence response to bitter compounds in the food supply. Phenotypes based on the perceived bitterness of single bitter compounds were first linked to food preference over 50 years ago. The most studied phenotype is propylthiouracil bitterness, which is mediated primarily by the TAS2R38 gene and possibly others. In a laboratory-based study, we tested for associations between TAS2R variants and sensations, liking, or intake of bitter beverages among healthy adults who were primarily of European ancestry . . . . . . . In summary, TAS2R polymorphisms appear to influence the sensations, liking, or intake of common and nutritionally significant beverages. Studying perceptual and behavioral differences in vivo using real foods and beverages may potentially identify polymorphisms related to dietary behavior even in the absence of known ligands. Allo stato attuale la famiglia genica TAS2R comprende 25 membri potenzialmente funzionali nell’uomo. Le proteine TAS2R all’interno della stessa specie mostrano dal 23% all’80% di identità. TAS2R: 25 geni che mappano in cluster su 3 cromosomi I 25 TAS2R sono codificati da cluster di geni localizzati sui cromosomi: - 15 geni sul cromosoma 12p - 9 sul cromosoma 7q - 1 sul cromosoma 5p 9 geni (TAS2R38,16,3,4,5) 15 geni (TAS2R19,43,44) TAS2R1 12 TAS2R espressi a livelli elevati nel 10% delle cellule recettoriali 6 espressi a livelli moderati nel <5% 7 espressi solo nell’ 1% Finora sono stati identificate circa 550 molecole in grado di legarsi a questi recettori. Tuttavia, questo numero rappresenta solo una piccola frazione delle migliaia di composti dal sapore amaro di origine vegetale che esistono in natura. Poiché il numero di composti supera notevolmente il numero di recettori, sembra probabile che i singoli recettori rispondono a più di un tipo di composto chimico dal sapore amaro. Il TAS2R38 è un esempio di recettore altamente selettivo per il quale sono noti solo 3 ligandi con strette analogie strutturali. Il recettore TAS2R38 che si lega al gruppo chimico N-C=S delle tiouree (responsabile del loro sapore amaro), come la feniltiocarbammide (PTC) e il 6-n-propiltiouracile (PROP). La genetica variabilità nella sensibilità a sentire il sapore amaro delle tiouree come il PROP e la PTC, rappresenta il più studiato fenotipo del gusto amaro nell’uomo. Le differenze individuali nella percezione gustativa dell’amaro sono ereditate geneticamente e riflettono la ricca diversità allelica dei recettori TAS2Rs. La capacità di legare uno o più agonisti per ogni recettore sembra essere dipendente non solo da un dato gene della famiglia TAS2R, ma anche dalla variazione allelica del gene stesso. Uno stesso recettore è attivato da composti simili. Esempio: - il recettore TAS2R16 risponde alla salicina e β- glucosidi correlati - il recettore TAS2R10 esibisce attivazione in risposta alla stricnina. - il recettore TAS2R19 è stato associato all’abilità a percepire il sapore amaro del chinino. - il recettore TAS2R38 è attivato da pheniltiocarbamide e da molti altri composti contenenti il gruppo –N=C=S, inclusa la propiltiourea. I TAS2R38 contengono un singolo esone codificante un recettore a serpentina della lunghezza media di 330 aminoacidi. Si trovano sul braccio lungo del cromosoma 7. 28 La sequenza del gene è qui sotto rappresentata. TAS2R38. GENE ORIGIN 1-1143 = EXON total length: 1143 1 cctttctgca ctgggtggca accaggtctt tagattagcc aactagagaa gagaagtaga 61 atagccaatt agagaagtga catcatgttg actctaactc gcatccgcac tgtgtcctat 121 gaagtcagga gtacatttct gttcatttca gtcctggagt ttgcagtggg gtttctgacc 181 aatgccttcg ttttcttggt gaatttttgg gatgtagtga agaggcaggc actgagcaac 241 agtgattgtg tgctgctgtg tctcagcatc agccggcttt tcctgcatgg actgctgttc 301 ctgagtgcta tccagcttac ccacttccag aagttgagtg aaccactgaa ccacagctac 361 caagccatca tcatgctatg gatgattgca aaccaagcca acctctggct tgctgcctgc 421 ctcagcctgc tttactgctc caagctcatc cgtttctctc acaccttcct gatctgcttg 481 gcaagctggg tctccaggaa gatctcccag atgctcctgg gtattattct ttgctcctgc 541 atctgcactg tcctctgtgt ttggtgcttt tttagcagac ctcacttcac agtcacaact 601 gtgctattca tgaataacaa tacaaggctc aactggcaga ttaaagatct caatttattt 661 tattcctttc tcttctgcta tctgtggtct gtgcctcctt tcctattgtt tctggtttct 721 tctgggatgc tgactgtctc cctgggaagg cacatgagga caatgaaggt ctataccaga 781 aactctcgtg accccagcct ggaggcccac attaaagccc tcaagtctct tgtctccttt 841 ttctgcttct ttgtgatatc atcctgtgtt gccttcatct ctgtgcccct actgattctg 901 tggcgcgaca aaataggggt gatggtttgt gttgggataa tggcagcttg tccctctggg 961 catgcagcca tcctgatctc aggcaatgcc aagttgagga gagctgtgat gaccattctg 1021 ctctgggctc agagcagcct gaaggtaaga gccgaccaca aggcagattc ccggacactg 1081 tgctgagaat ggacatgaaa tgagctcttc attaatacgc ctgtgagtct tcataaatat 1141 gcc L’abilità a sentire il sapore amaro delle tiouree è associata agli aplotipi del gene TAS2R38 che derivano da tre singoli polimorfismi che risultano in tre sostituzioni aminoacidiche. I tre SNPs responsabili di tre sostituzioni aminoacidiche nella proteina sono: rs713598 pos 229 G -> C A49P (Alanina/Prolina) ALA -> PRO rs1726866 pos. 869 T -> C A262V (Alanina/Valina) VAL -> ALA rs10246939 pos.970 A -> G I296V (Isoleucina/Valina) ILE -> VAL CDS ORIGIN – EXON - total length: 1,002 1 cctttctgca ctgggtggca accaggtctt tagattagcc aactagagaa gagaagtaga 61 atagccaatt agagaagtga catcatgttg actctaactc gcatccgcac tgtgtcctat 121 gaagtcagga gtacatttct gttcatttca gtcctggagt ttgcagtggg gtttctgacc 181 aatg ccttcg ttttcttggt gaatttttgg gatgtagtga agaggcaggc actgagcaac rs713598 241 agtgattgtg tgctgctgtg tctcagcatc agccggcttt tcctgcatgg actgctgttc C/G 301 ctgagtgcta tccagcttac ccacttccag aagttgagtg aaccactgaa ccacagctac 361 caagccatca tcatgctatg gatgattgca aaccaagcca acctc tggct tgctgcctgc 421 ctcagcctgc tttactgctc caagctcatc cgtttctctc acaccttcct gatctgcttg 481 gcaagctggg tctccaggaa gatctcccag atgctcctgg gtattattct ttgctcctgc 541 atctgcactg tcctctgtgt ttggtgcttt tttagcagac ctcacttcac agtcacaact 601 gtgctattca tgaataacaa tacaaggctc aactggcaga ttaaagatct caatttattt 661 tattcctttc tcttctgcta tctgtggtct gtgcctcctt tcctattgtt tctggtttct 721 tctgggatgc tgactgtctc cctgggaagg cacatgagga caatgaaggt ctataccaga 781 aactctcgtg accccagcct ggaggcccac attaaagccc tcaagtctct tgtctccttt 841 ttctgcttct ttgtgatatc atcctgtgct gccttcatct ctgtgcccct actgattctg rs1726866 901 tggcgcgaca aaataggggt gatggtttgt gttgggataa tggcagcttg tccctctggg C/T 961 catgcagcca tcctgatctc aggcaatgcc aagttgagga gagctgtgat gaccattctg rs10246939 1021ctctgggctc agagcagcct gaaggtaaga gccgaccaca aggcagattc ccggacactg A/G 1081tgctgagaat ggacatgaaa tgagctcttc attaatacgc ctgtgagtct tcataaatat 1141gcc 29 Protein product length: 333 translation "MLTLTRIRTVSYEVRSTFLFISVLEFAVGFLTNAFVFLVNFWDV VKRQALSNSDCVLLCLSISRLFLHGLLFLSAIQLTHFQKLSEPLN HSYQAIIMLWMIANQANLWLAACLSLLYCSKLIRFSHTFLICLAS WVSRKISQMLLGIILCSCICTVLCVWCFFSRPHFTVTTVLFMNNN TRLNWQIKDLNLFYSFLFCYLWSVPPFLLFLVSSGMLTVSLGRH MRTMKVYTRNSRDPSLEAHIKALKSLVSFFCFFVISSCVAFISV PLLILWRDKIGVMVCVGIMAACPSGHAAILISGNAKLRRA VM TILLWAQSSLKVRADHKADSRTLC Ecco il cambio di aminoacidi sulla proteina Il 2° e il 3° SNP sono in perfetto linkage disequilibrium Si conoscono due comuni aplotipi, la variante dominante PAV associata alla elevata sensibilità e la variante recessiva AVI associata a una bassa o nulla sensibilità alle tiouree. Queste varianti danno origine ai Taster (PAV) e ai Non Taster (AVI). Avremo quindi tre genotipi: - PAV/PAV super-taster con alta sensibilità per l’amaro - PAV/AVI medium taster con moderata sensibilità per l’amaro - AVI/AVI non taster non percepiscono l’amaro Gli individui omozigoti o eterozigoti per la variante PAV sono in grado di sentire il sapore amaro delle tiouree anche a basse concentrazioni, mentre gli individui che sono completamente incapaci di sentire le tiouree e quelli che le sentono solo ad alte concentrazioni sono omozigoti per la variante AVI I genotipi del TAS2R38 sono in grado di predire la maggior parte (55-85%), ma non la totalità delle variazioni nel fenotipo del PROP, implicando che altri fattori epigenetici possano essere coinvolti nell’espressione di questo carattere genetico. Non-genetic factors influencing PROP taste perception Approximately 50% of the observed variation in PROP detection thresholds is accounted for by genetic variation in the TAS2R38 gene. From replicate measurements we can estimate that the measurement error accounts for another 20% of the observed phenotype variability. This leaves approximately 30% of the phenotypic variation we observed to be explained by other factors. Consistent with reports in the literature [33–35] neither gender nor body mass index explain a significant portion of these remaining 30% of variation. Even age (Figure 3a), which is known to influence both overall and PROP-specific taste sensitivity explains only 3.8% of the overall variation in PROP detection thresholds. Nella popolazione bianca - la più studiata - la distribuzione dei tre fenotipi è di circa: • 30% Non-taster AVI/AVI • 45% Medium-taster AVI/PAV • 25% Supertaster PAV/PAV Alti livelli di variazioni alleliche sono state trovate all’interno dei loci TAS2R nelle popolazioni umane. Questa variabilità correla con l’enorme variabilità nella percezione di composti amari riscontrata nell’uomo. Le diversità nei geni TAS2R potrebbero essere dovuti alla selezione naturale, che può aver favorito alleli maggiormente responsivi a tossine naturali amare prodotte dalle piante. 30 BIOINFORMATICA Bitter receptor gene TAS2R38 PREMESSA: Principali siti di riferimento www.ncbi.nlm.nih.gov L’NCBI (National Centre for Biotecnology Information) sorto nel 1988, crea database pubblici, conduce ricerche in bioinformatica, sviluppa software per analizzare dati genomici e divulga informazioni biomediche. L’obiettivo è una migliore comprensione dei processi molecolari riguardanti la salute umana e le malattie. Sul sito si trovano banche dati relative al genoma umano e di altri organismi, a sequenze nucleotidiche e aminoacidiche, a strutture molecolari, a pubblicazioni scientifiche (come PubMed, la principale banca dati bibliografica, pubblica e gratuita, del settore biomedico). In particolare NCBI –Entrez comprende un database di strutture biomolecolari 3D determinate sperimentalmente; forniscono informazioni sulla funzione biologica, la storia evolutiva e le relazioni tra le macromolecole. http://genome.ucsc.edu/ UCSC ( University of California Santa Cruz). Questo sito contiene le sequenze di riferimento e le schermate che mostrano un’ampia collezione di genomi. Fornisce inoltre un portale di accesso al Progetto Encode. www.ensembl.org ENSAMBL ( un gioco di parole tra ensamble -insieme- e EMBL, European Molecular Biology Laboratory) è un progetto sviluppato in collaborazione tra il Sanger Center di Cambridge e EMBL per sviluppare un software di annotazione automatica dei genomi animali. Ensambl aggiorna i dati almeno 10 volte l’anno. OBIETTIVI DELL’ATTIVITA’ : 1. Cos’è il recettore per il gusto amaro? 2. Come si fa a localizzare il gene d’interesse ? 3. Una volta che l’ho localizzato, come trovo tutte le informazioni necessarie ? 4. Come si caratterizza il gene ? 5. Come trovare il polimorfismo a singolo nucleotide rs 1766866 correlato con la percezione del gusto amaro? 6. Come disegnare la coppia di Primer F e R per amplificare il tratto di gene a monte e a valle dello Snp? 7. Come interpretare i dati statistici sulle frequenze alleliche nelle popolazioni 31 1°. OBIETTIVO: COS’E’ IL RECETTORE PER IL GUSTO AMARO ? Andiamo su GOOGLE . digitiamo NCBI. Si apre la Home Page. 2° OBIETTIVO: Come si fa a localizzare il gene TAS2R38 ? Search: Nucleotide. TAS2R38. Andare su GO. Si apre la pag seguente Cliccare su TAS2R38 (Homo Sapiens). Si aprirà la pagina contenente le informazioni sul gene 3° OBIETTIVO: Come si trovano tutte le informazioni necessarie ? Nella pagina aperta leggiamo: - Summary. Sotto: 32 Sotto ancora: La Bibliografia: 4° OBIETTIVO: Come si caratterizza il gene TAS2R38? Se torniamo sopra a Genomic regions….Puntiamo il mouse su NM176817.4 Leggiamo una seri di informazioni sul gene. Lunghezza: 1143 bp. Proteina 333 AA Clicchiamo sopra. Si ha una ulteriore grafica sul gene. Clicchiamo su GeneBank Scorriamo la pagina. Troviamo le informazioni su - gene 1….1143 - mRNA 1..1143 - CDS 85…1086 Ciccando sul rif NM 176817.4 dell’mRNA mi compare anche exon 1…1143 Sotto troviamo la sequenza del gene. Andiamo al Capitolo Genomic contest. Chr 7. Location 7q34 Vogliamo vedere il gene in un altro database: UCSC Qui possiamo vedere le sequenze conservate del gene Tas2R38 nelle specie Clicchiamo sul gene e andiamo ad una pagina molto ricca di informazioni 33 5. Individuazione del Polimorfismo SNP rs713598 Torniamo al NCBI nella pagina che avevamo lasciato aperta. Al Cap General gene information c’è un sotto cap Genotypes. Andare a TAS2R38 SNP e Ciccare su VarView Vediamo i tre SNPs interessanti per la variazione di percezione dell’amaro. A noi interessa il primo, rs713598. Clicchiamo sopra. Si apre la pagina con lo SNP C/G. E’ un missense alla posizione 229 della proteina. Scorrendo la pagina avremo in Fasta la sequenza intorno allo SNP. Qui isoliamo la regione intorno allo SNP Copiamo la sequenza a monte e a valle dello snp e cerchiamo i primer nel programma chiamato Primer3. Si apre la pagina 34 Nel riquadro incolliamo la sequenza copiata e clicchiamo su “submit primer” Comparirà il seguente pannello 6. Genetica delle popolazioni Quali frequenze alleliche di riferimento si riscontrano nella popolazione europea? 35 PROTOCOLLO DI LABORATORIO Individuazione del polimorfismo G/C rs713598 del gene TAS2R38 (Bitter receptor gene) STEP 1 Estrazione del DNA STRUMENTI E REAGENTI necessari § § § § § Bagno maria o termoblocco a 60°C Pipette con puntali Microcentrifuga provette da 1.5ml Vortex PRIMA DI INIZIARE 1. Preparare un bagnomaria a 60°C 2. Se si forma un precipitato nella soluzione di Lisi, scaldare a 60 °C per alcuni minuti 3. Prendere dal freezer la PK solution e scongelarla a temperatura ambiente. Parte A –Stabilizzazione del DNA 1. Aggiungere 500ul LS solution alla provetta contenente lo swab. 2. Aggiungere 20µl PK solution alla provetta. Vortexare. A questo punto il DNA è stabilizzato Si può continuare a isolare il DNA oppure conservare il swab stabilizzato in una provetta sigillata a temperatura ambiente anche per 3½ anni. Parte B – Isolamento del DNA 0 3. Mettere la provetta (contenente lo swab, la LS solution e la PK solution) in un bagnetto a 60 C per 1 ora. Vortexare brevemente. 4. Trasferire il liquido dalla provetta (approx. 400µl) da 5 ml in un’altra provetta da 1.5 ml usando un puntale pulito ( o una pasteur di plastica) . 5. Aggiungere 400µl CT solution nella provetta. Vortexare . 6. Centrifugare ad approx. 13K rpm per 7 minuti per pelletare il DNA. Tip: Mettere la provetta con la chiusura del tappo posizionato verso l’esterno così che il liquido possa essere rimosso sul lato opposto Nota - il pellet può non essere visibile 7. Rimuovere tutto il supernatante con attenzione usando una pipetta per non toccare il pellet di DNA. 8. Ri- centrifugare la provetta brevemente e rimuovere il liquido rimanente. Nota- è importante rimuovere tutto il liquido. 9. Aggiungere 100µl TE solution alla provetta. Il campione di DNA è ora pronto per la PCR 0 0 Conservare il campione di DNA a 4 C per un tempo breve o a -20 C per tempi più lunghi. La resa attesa è intorno a 10µg DNA (5 to 70 ng/µl). STEP 2. PREPARAZIONE DEL GEL DI AGAROSIO - Sciogliere l’Agarosio ( in quantità stabilita dalla %) nella beuta con la soluzione tamponeTAE. - Far bollire la soluzione nel microonde agitando di tanto in tanto fino a quando questa non diventa trasparente. - Quando la soluzione è tiepida, ma non ancora gelificata, aggiungere 3 µL.di intercalante del DNA - Preparare la vaschetta per elettroforesi con bordi di nastro adesivo e inserire il pettine. - Versare la soluzione di Agarosio nella vaschetta per elettroforesi e lasciare polimerizzare (solidificare). - Togliere il pettine quando il gel sarà solidificato. - Versare il tampone TAE nella camera di corsa STEP 3. CHECK DNA con gel di Agarosio allo 0.8% La presenza di DNA viene valutata con un check prelevando 10 µl di DNA. - Prelevare 2 µL colorante e disporlo su carta formando una goccia - Mescolare con la pipetta il DNA e il colorante - Si trasferisce il prelievo in un pozzetto del gel, facendo attenzione a non bucare il fondo 36 del pozzetto e a non far uscire il campione fuori dal pozzetto - Chiudere e attaccare gli elettrodi all’alimentatore. Far correre per 10- 20’. - Osservare al transilluminatore STEP 4. PCR MIX per 25 µL di PCR: Buffer 2.5 µL (10x) dnTP 0.5 µL (10mM) Foward 0.5 µL (10mM) Reverse 0.5 µL (10mM) MgCl2 0.75. µL (50mM) TAQ 0.10 µL H2 O 19.13 µL DNA 1 µl (100 ng totali) Lavorare in ghiaccio ! Ciclo PCR 95°C x 5’ (95°C 30”, 64°C 45”, 72°C 30”) x 35 72°C 10’ 4°C° store Durata: 2h.30’ circa L’amplimero che si ottiene con la PCR è di 221 bp. Per individuare lo SNP del gene TAS2R38, rs 713598 dobbiamo creare artificialmente un sito di restrizione. Per questo motivo disegniamo il primer Foward con un mismatch F 5’ CCTTCGTTTTCTTGGTGAATTTTTGGGATGTAGTGAAGAGGCAGG 3’ (101 – 144) in pos 143 porta un MISMATCH = ha una G mentre sul gene c’e’ A R 5’- AGGTTGGCTTGGTTTGCAATCAT – 3’ Il primer di attacco si lega al gene TAS2R38 a livello dei nucleotidi 101-144. Il mismatch A/G è fondamentale per l’ esperimento di PCR, perché il nucleotide A nella sequenza del gene TAS2R38, è rimpiazzato da una G in ognuno dei prodotti di amplificazione. Questo crea la prima G della sequenza di riconoscimento GGCC di HaeIII, permettendo il taglio dell’allele taster amplificato (GG). L’allele non taster amplificato legge invece GGGC e di conseguenza non subisce nessun taglio. STEP 5. CHECK PCR con gel di Agarosio al 2,5% 5 µL di PCR e 2 µL di colorante. In un pozzetto si mettono 2µL. di marker di 100 bp. Al termine della corsa (circa 20 min), si prende il gel e lo si osserva al transilluminatore STEP 6. DIGESTIONE ENZIMATICA 20 μL PCR 2,5 μL 10 x NE Buffer (10x) 2 μL H2O 0.5 μL di HaeIII La soluzione viene incubata a 37°C per 1 h STEP 7. Valutazioni sui risultati. GGCC allele Taster (digerito) 176 bp + 45 bp GGGC allele Non Taster (non digerito) 221 bp 37 tt - non taster - non digerito 221 bp digerito 221 bp TT taster - non digerito 221 bp digerito 176 +45 bp Tt taster - non digerito 221 bp digerito 221 bp 176 + 45 38 Bitter Taste Sensitivity Test L’intensità della sensazione dell’amaro viene stimata mediante la Scala LMS (LABELED MAGNITUDE SCALE) . Tale scala è una scala quasi-logaritmica, nella quale dei descrittori semantici corrispondono a determinate ampiezze della valutazione della sensazione evocata Questa scala minimizza l’effetto soglia e fornisce una misura continua, desiderabile quando si fa un’analisi quantitativa. La scala è ancorata verso il basso al descrittore “appena percettibile” e in alto al descrittore “il più forte immaginabile”. Quest’ultimo deve essere riferito alla sensazione orale più forte che il soggetto abbia mai avuto come esperienza nella sua vita (un cibo piccante, un dentifricio forte, una caramella ...). La posizione delle etichette verbali corrisponde, nella scala, ad una percentuale della lunghezza totale: appena percettibile 1.4, debole 6.1, moderato 17.2, forte 35.4, molto forte 53.3, il più forte immaginabile 100. Dopo aver effettuato la stimolazione, indicare il valore della scala che rappresenta meglio l’intensità della sensazione gustativa percepita. PROCEDURA 1. Gli studenti non devono mangiare, fumare e bere solo acqua da almeno un’ora prima del test. Posizionare la cartina PTC sulla parte anteriore e mediana della lingua 2. Sul foglio del punteggio si da un voto all’intensità percepita segnando un trattino con una matita nella Labeled Magnitude Scale (LMS) Labeleled Magnitude Scale 100 - Strongest imaginable 80 - 60 -- Very strong 40 - Strong 20 - Moderate 10 - Weak 50 Barely detectable SESSO ALTEZZA PESO PROVENIENZA REGIONALE E/O NAZIONALE DEI GENITORI 39 FOOF PREFERENCE QUESTIONNAIRE Prima di iniziare l’attività bisogna rispondere al seguente questionario tenendo presente che bisogna indicare non tanto quanto si mangia di questi cibi ma quanto si preferiscono (cioè quanto piace il sapore) indipendentemente dal fatto che si mangiano spesso o raramente. ID_______________ Sex______ AGE_________ ACCIUGHE 1 2 3 4 5 6 7 8 9 Never_tasted CARCIOFI 1 2 3 4 5 6 7 8 9 Never_tasted ASPARAGI 1 2 3 4 5 6 7 8 9 Never_tasted AVOCADO 1 2 3 4 5 6 7 8 9 Never_tasted PANCETTA 1 2 3 4 5 6 7 8 9 Never_tasted RADICCHIO 1 2 3 4 5 6 7 8 9 Never_tasted OLIVE_NERE 1 2 3 4 5 6 7 8 9 Never_tasted SUGO_ALLA_BOLOGNESE 1 2 3 4 5 6 7 8 9 Never_tasted FAVE 1 2 3 4 5 6 7 8 9 Never_tasted BROCCOLI 1 2 3 4 5 6 7 8 9 Never_tasted CAVOLO 1 2 3 4 5 6 7 8 9 Never_tasted PANETTONE 1 2 3 4 5 6 7 8 9 Never_tasted CAPPERI 1 2 3 4 5 6 7 8 9 Never_tasted CAVOLFIORE 1 2 3 4 5 6 7 8 9 Never_tasted CICORIA 1 2 3 4 5 6 7 8 9 Never_tasted PEPERONCINO 1 2 3 4 5 6 7 8 9 Never_tasted CETRIOLO 1 2 3 4 5 6 7 8 9 Never_tasted BIRRA_SCURA 1 2 3 4 5 6 7 8 9 Never_tasted CIOCCOLATO_FONDENTE 1 2 3 4 5 6 7 8 9 Never_tasted ZABAIONE 1 2 3 4 5 6 7 8 9 Never_tasted ACCIUGHE 1 2 3 4 5 6 7 8 9 Never_tasted CARCIOFI 1 2 3 4 5 6 7 8 9 Never_tasted 40 ASPARAGI 1 2 3 4 5 6 7 8 9 Never_tasted AVOCADO 1 2 3 4 5 6 7 8 9 Never_tasted ACCIUGHE 1 2 3 4 5 6 7 8 9 Never_tasted CARCIOFI 1 2 3 4 5 6 7 8 9 Never_tasted ASPARAGI 1 2 3 4 5 6 7 8 9 Never_tasted AVOCADO 1 2 3 4 5 6 7 8 9 Never_tasted PANCETTA 1 2 3 4 5 6 7 8 9 Never_tasted RADICCHIO 1 2 3 4 5 6 7 8 9 Never_tasted OLIVE_NERE 1 2 3 4 5 6 7 8 9 Never_tasted SUGO_ALLA_BOLOGNESE 1 2 3 4 5 6 7 8 9 Never_tasted FAVE 1 2 3 4 5 6 7 8 9 Never_tasted BROCCOLI 1 2 3 4 5 6 7 8 9 Never_tasted CAVOLO 1 2 3 4 5 6 7 8 9 Never_tasted PANETTONE 1 2 3 4 5 6 7 8 9 Never_tasted CAPPERI 1 2 3 4 5 6 7 8 9 Never_tasted CAVOLFIORE 1 2 3 4 5 6 7 8 9 Never_tasted CICORIA 1 2 3 4 5 6 7 8 9 Never_tasted ACCIUGHE 1 2 3 4 5 6 7 8 9 Never_tasted CARCIOFI 1 2 3 4 5 6 7 8 9 Never_tasted ASPARAGI 1 2 3 4 5 6 7 8 9 Never_tasted AVOCADO 1 2 3 4 5 6 7 8 9 Never_tasted PANCETTA 1 2 3 4 5 6 7 8 9 Never_tasted RADICCHIO 1 2 3 4 5 6 7 8 9 Never_tasted OLIVE_NERE 1 2 3 4 5 6 7 8 9 Never_tasted SUGO_ALLA_BOLOGNESE 1 2 3 4 5 6 7 8 9 Never_tasted 41 FAVE 1 2 3 4 5 6 7 8 9 Never_tasted BROCCOLI 1 2 3 4 5 6 7 8 9 Never_tasted CAVOLO 1 2 3 4 5 6 7 8 9 Never_tasted PANETTONE 1 2 3 4 5 6 7 8 9 Never_tasted CAPPERI 1 2 3 4 5 6 7 8 9 Never_tasted CAVOLFIORE 1 2 3 4 5 6 7 8 9 Never_tasted CICORIA 1 2 3 4 5 6 7 8 9 Never_tasted PEPERONCINO 1 2 3 4 5 6 7 8 9 Never_tasted CETRIOLO 1 2 3 4 5 6 7 8 9 Never_tasted BIRRA_SCURA 1 2 3 4 5 6 7 8 9 Never_tasted CIOCCOLATO_FONDENTE 1 2 3 4 5 6 7 8 9 Never_tasted ACCIUGHE 1 2 3 4 5 6 7 8 9 Never_tasted CARCIOFI 1 2 3 4 5 6 7 8 9 Never_tasted ASPARAGI 1 2 3 4 5 6 7 8 9 Never_tasted AVOCADO 1 2 3 4 5 6 7 8 9 Never_tasted PANCETTA 1 2 3 4 5 6 7 8 9 Never_tasted RADICCHIO 1 2 3 4 5 6 7 8 9 Never_tasted OLIVE_NERE 1 2 3 4 5 6 7 8 9 Never_tasted SUGO_ALLA_BOLOGNESE 1 2 3 4 5 6 7 8 9 Never_tasted FAVE 1 2 3 4 5 6 7 8 9 Never_tasted BROCCOLI 1 2 3 4 5 6 7 8 9 Never_tasted CAVOLO 1 2 3 4 5 6 7 8 9 Never_tasted PANETTONE 1 2 3 4 5 6 7 8 9 Never_tasted CAPPERI 1 2 3 4 5 6 7 8 9 Never_tasted CAVOLFIORE 1 2 3 4 5 6 7 8 9 Never_tasted 42 ACCIUGHE 1 2 3 4 5 6 7 8 9 Never_tasted CARCIOFI 1 2 3 4 5 6 7 8 9 Never_tasted ASPARAGI 1 2 3 4 5 6 7 8 9 Never_tasted AVOCADO 1 2 3 4 5 6 7 8 9 Never_tasted PANCETTA 1 2 3 4 5 6 7 8 9 Never_tasted RADICCHIO 1 2 3 4 5 6 7 8 9 Never_tasted OLIVE_NERE 1 2 3 4 5 6 7 8 9 Never_tasted MINESTRONE 43 BIBLIOGRAFIA 1. D.A. Prodi, D. Drayna, P. Forabosco, M.A. Palmas, G.B. Maestrale, D. Piras, M. Pirastu and A. Angius Bitter Taste Study in a Sardinian Genetic Isolate Supports the Association of Phenylthiocarbamide Sensitivity to the TAS2R38 Bitter Receptor Gene . 2. Julie A. Mennella, PhD, M. Yanina Pepino, PhD, and Danielle R. Reed, PhD ,Genetic and Environmental Determinants of Bitter Perception and Sweet Preferences 3. Jonathan L. Hansen, Danielle R. Reed, Margaret J. Wright, Nicholas G. Martin,and Paul A. S. Breslin Heritability and Genetic Covariation of Sensitivity to PROP,SOA, Quinine HCl, and Caffeine. 4.The Molecular Basis of Individual Differences in Phenylthiocarbamide and Propylthiouracil 5. Bernd Bufe, Paul A.S. Breslin, Christina Kuhn, Danielle R. Reed, Christopher D. Tharp, Jay P. Slack, Un-Kyung Kim, Dennis Drayna, and Wolfgang Meyerhof Bitterness Perception 6. Jian Yang, Beben Benyamin1, Brian P McEvoy, Scott Gordon, Anjali K Henders, Dale R Nyholt, Pamela A Madden, Andrew C Heath, Nicholas G Martin, Grant W Montgomery, Michael E Goddard & Peter M Visscher. Common SNPs explain a large proportion of the heritability for human height 6. Beverly J. Tepper, Yvonne Koelliker, Liqiang Zhao, Natalia V. Ullrich, Carmela Lanzara,Pio d'Adamo, Antonella Ferrara, Sheila Ulivi, Laura Esposito and Paolo Gasparini Variation in the Bitter-taste Receptor Gene TAS2R38, and Adiposity in a Genetically Isolated Population in Southern Italy 7. J. Tepper,a Elizabeth A. White,a Yvonne Koelliker, Carmela Lanzara,Pio d’Adamo, and Paolo Gasparinib Genetic Variation in Taste Sensitività to 6-n-Propylthiouracil and Its Relationship to Taste Perception and Food Selection Beverly 8. Stephen Wooding. Anecdotal, Historical and Critical Commentaries on Genetics. Phenylthiocarbamide: A 75-Year Adventure in Genetics and Natural Selection.Genetics,2009 9 Sensitivity of Genome-Wide-Association Signals to Phenotyping Strategy: The PROP TAS2R38 Taste Association as a Benchmark Ulrich K. Genick, Zolta´n Kutalik, Mirko Ledda, Maria C. Souza Destito, Milena M. Souza, Cintia A. Cirillo, Nicolas Godinot, Nathalie Martin, Edgard Morya, Koichi Sameshima, Sven Bergmann, Johannes le Coutre PLoS ONE ISSUE 2011 10 La percezione del gusto amaro nell’uomo: La sensibilità gustativa al 6-n-propiltiouracile (PROP) come marker della variabilità individuale con implicazioni fisiologico-nutrizionali Andrea Zonza 44 Guida per lo studente Background di Conoscenze su alcuni argomenti di Genetica Le malattie ereditarie monogeniche sono determinate da mutazioni in singoli geni che si trasmettono nelle famiglie secondo leggi ben definite scoperte da G. Mendel nella seconda metà dell’Ottocento. Per questo motivo le malattie monogeniche monofattoriali vengono anche chiamate“mendeliane”. In particolare bisogna ricordare che: - I geni possono avere alleli diversi (varianti alternative di uno stesso gene) ossia essere polimorfici; - Gli organismi diploidi hanno due copie di ogni gene, cioè due alleli. Se i due alleli sono uguali, l’individuo è omozigote (ad es, ha genotipo AA oppure aa). Se i due alleli sono diversi, l’individuo è eterozigote (ad es, il genotipo è Aa) - I legge di Mendel o legge dell’uniformità della prima generazione ibrida, afferma che l’incrocio tra individui della generazione parentale ciascuno omozigote per due alleli diversi di uno stesso gene (ad es, AA x aa) e che quindi differisce dall’altro genitore per una caratteristica (ad es, pelo nero o marrone), dà una progenie costituita da individui tutti identici tra loro (tutti eterozigoti; ad es, Aa); - Durante la meiosi i due alleli di un gene segregano e si distribuiscono ciascuno in un gamete apolide (II legge di Mendel, legge della segregazione). Un individuo omozigote produce un solo tipo di gamete (A oppure a) relativamente a un dato locus (la posizione occupata da un gene in un cromosoma). Un individuo eterozigote produce due tipi di gameti (A e a) in ugual quantità, cioè in rapporto 50% ciascuno. Una buona rappresentazione grafica della segregazione si ottiene costruendo il “quadrato di Punnett”. - Alleli appartenenti a geni diversi localizzati su cromosomi diversi segregano in modo indipendente (III legge di Mendel, legge della segregazione indipendente). - Allele dominante: allele che si manifesta allo stato eterozigote; - Allele recessivo: allele mascherato dall’allele dominante, che si manifesta solo allo stato omozigote; - Allele codominante: due alleli entrambi espressi allo stato eterozigote, che agiscono sul fenotipo in modi indipendenti e distinguibili. Un esempio classico si ha nel caso degli alleli IA e IB del gruppo sanguigno ABO. L’1% circa dei neonati è affetto da una malattia genetica monogenica. La mutazione genica è presente fin dal concepimento, quindi anche alla nascita, anche se non sempre si manifesta fenotipicamente alla nascita. Alcune di queste malattie comportano conseguenze già apprezzabili nel neonato. Altre malattie monogeniche si manifestano, invece, durante le età successive della vita. La corea di Huntington, ad esempio, è una malattia genetica a insorgenza tardiva che si manifesta solo in età adulta. - Si definisce malattia autosomica quella in cui l’allele malato è presente nelle coppie di cromosomi non coinvolti nella determinazione del sesso, gli autosomi; - si parla invece di malattia legata all’X quella in cui l’allele si trova nel cromosoma sessuale X. In questo caso la trasmissione segue modalità particolari in quanto la maggior parte dei geni rappresentati sul cromosoma X non sono rappresentati sul cromosoma Y. Il cromosoma Y, infatti, è molto più piccolo e contiene quasi esclusivamente i geni per la determinazione del sesso. Per schematizzare: - malattie autosomiche: se l’allele normale è dominante, la malattia si rende manifesta solo nella condizione omozigote recessiva, ossia negli individui in cui entrambi gli alleli del gene sono alterati (malattia autosomica recessiva); se invece è l’allele alterato a prevalere su quello sano, la malattia si rende sempre evidente, anche nella condizione eterozigote, ossia è sufficiente che uno solo dei due alleli sia alterato per avere la malattia (malattia autosomica dominante) - malattie legate all’X: se l’allele è recessivo, come nella maggior parte dei casi, i maschi ricevono l’X difettoso dalla madre, ma non possono bilanciarlo con un allele sano nell’Y in quanto questo cromosoma non contiene l’allele corrispondente; le femmine invece manifestano la malattia solo nelle condizione omozigote. Di conseguenza, la frequenza delle malattie recessive legate al cromosoma X è maggiore nei maschi che nelle femmine. Nel caso di malattie dominanti le femmine manifestano la malattia anche nella condizione eterozigote 45 GLOSSARIO Allele. è una delle possibili forme o varianti in cui si puo' presentare un gene, con conseguenze sulle caratteristiche espresse. Per ogni gene (ad un determinato locus sul cromosoma) esistono due alleli, ereditati dal padre e della madre. Quando i due alleli sono uguali, l'individuo è definito omozigote per quel specifico tratto; se gli alleli sono diversi, l'individuo è definito eterozigote. Annealing (appaiamento) formazione di molecole ibride di acidi nucleici basata sulla complementarietà delle basi. Elettroforesi tecnica che consente di separare molecole caricate elettricamente, mediante la migrazione in un campo elettrico. Enzima proteina che catalizza (accelera) una specifica reazione chimica di una via metabolica in un sistema vivente. Ha azione specifica, in quanto riconosce il substrato su cui agire. Enzimi di restrizione chiamati anche endonucleasi di restrizione o “forbici” molecolari : sono enzimi che tagliano il DNA a doppia elica (e non quello a singolo filamento!) in corrispondenza di specifiche sequenze nucleotidiche, dette siti di restrizione Fenotipo insieme delle caratteristiche visibili di un organismo, che risultano dall’interazione tra genotipo e ambiente. Frammenti di restrizione frammenti provenienti da una molecola di DNA più lunga per digestione con enzimi di restrizione Gene unità ereditaria funzionale corrispondente generalmente al segmento di DNA che codifica una proteina. Genoma patrimonio genetico di una cellula o di un organismo. Genotipo costituzione genetica di un organismo Linkage Equilibrium indica una combinazione casuale di alleli a loci associati Linkage disequilibrium indica una combinazione non casuale di alleli a loci associati Isoforma Un tempo si riteneva che ogni gene codificasse per un unico prodotto polipeptidico o molecola di RNA, mentre oggi si sa che la maggioranza di geni umani specifica 2 o più forme alternative di proteine Locus ( plurale loci) posizione fissa su un cromosoma occupata da un dato gene. Nel liguaggio comune il termine viene spesso usato come sinonimo di gene Marcatore genetico qualsiasi differenza fenotipica, controllata geneticamente, utilizzata nell’analisi genetica Oligonucleotide corta molecola di DNA (o RNA) a singolo filamento, costituita da poche decine di basi, ottenuta artificialmente e utilizzata in esperimenti di ibridazione molecolare e nella PCR PCR (polymerase chain reaction) tecnica di biologia molecolare utilizzata per amplificare in breve tempo segmenti specifici di DNA Polimorfismo esistenza di forme diverse di: alleli (polimorfismo allelico), proteine (polimorfismo proteico), DNA (polimorfismo del DNA). ciascuno dei quali con una frequenza di almeno 1%. Polimorfismo della lunghezza dei frammenti di restrizione (RFLP) Restriction Fragment Length Polymorphism (lunghezza variabile dei frammenti di restrizione). Polimorfismo del DNA, che si evidenzia trattando cromosomi omologhi con enzimi di restrizione. Determinato da una differenza di una base tra due DNA omologhi, differenza che causa la presenza o assenza di un sito di restrizione per uno specifico enzima di restrizione, e quindi frammenti di diversa lunghezza dopo trattamento con quell’enzima. Primer corta catena polinucleotidica, a DNA o RNA, alla quale vengono aggiunti nuovi nucleotidi nel corso della sintesi di DNA. Noto anche con il termine di innesco. Nella PCR si utilizza una coppia di primer Promotore regione del DNA a cui si lega la RNA-polimerasi per avviare la trascrizione del gene (cioè la sintesi dell'RNA-messaggero). Funziona come un "interruttore molecolare" che "accende" o "spegne" il gene. regioni di controllo sequenze del DNA che attivano e regolano l'espressione di un gene. La loro presenza è pertanto necessaria per la corretta e efficace espressione del gene. RNA-messaggero (mRNA) copia a singolo filamento della sequenza nucleotidica di un gene, 46 che trasporta ai ribosomi l'informazione per la sintesi (traduzione) della proteina corrispondente Splicing del RNA processo in cui le sequenze introniche sono eliminate dai trascritti primari di RNA nel nucleo durante la trascrizione, per dar luogo alla produzione di molecole di mRNA maturo. Splicing alternativo assemblaggio differenziale di esoni durante la maturazione dell’RNA. Si stima che più del 60% dei geni umani produca 2 o più proteine mediante questo meccanismo. Molti esoni specificano domini proteici strutturali distinti che possono essere combinati in modo diverso nelle cellule dei diversi tessuti nei quali il gene è espresso..si ottengono così isoforme tessuto-specifiche Sequenze palindromiche sequenza nucleotidica che è identica al suo filamento complementare quando entrambi vengono letti nella stessa direzione chimica (es 5'-->3'). Esempio GATC Siti di restrizione sequenza di basi ben determinata riconosciuta e tagliata dagli enzimi di restrizione in modo da lasciare corte sequenze a singola elica, dette estremità coesive poiché le loro basi sono complementari. PRE-TEST POST-TEST 47
© Copyright 2026 Paperzz