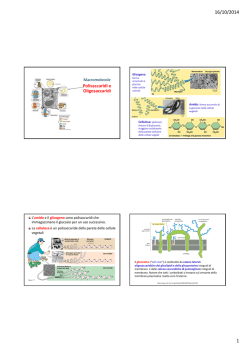
Enzimi amilolitici L’amido, uno dei polimeri più abbondanti sulla Terra, è un polimero di molecole di glucosio, legate mediante legami a-glicosidici: Amilosio glucano lineare con legami a-1,4 Amilopectina glucano lineare con legami a-1,4 e ramificato con legami a- 1,6 Le amilasi (a-amilasi, -amilasi, glucoamilasi) scindono le catene dei costituenti dell’amido, introducendo una molecola d’acqua, a livello dei legami glucosidici. ENDOAMILASI PM 15-50k Industria alimentare Attaccano all’interno delle catene del polimero a qualsiasi punto. Scindono il legame a-1,4 e portano alla formazione di DESTRINE di diversa lunghezza (eritrodestrine con PM elevato e le acrodestrine con PM più basso). Es. la ptialina salivare, l’amilasi pancreatica, la takadiastasi (a-amilasi da A. orizae) e diverse amilasi di microrganismi. ESOAMILASI (da vegetali o microrganismi) attaccano le estremità non riducenti delle catene di a-D-glucosio e liberano sia molecole di maltosio, idrolizzando il penultimo legame a-1->4, di glucosio, sia molecole di glucosio, idrolizzando l’ultimo legame a-1->4 o a-1->6 o a-1->63. Nel primo caso abbiamo le -amilasi, nel secondo caso le a-1->4 glucosidasi: glucoamilasi o -amilasi o amiloglucosidasi. Le a-1->6 glucosidasi sono l’isoamilasi (enzima deramificante), la pullulanasi (enzima R), e la destrinasi limite. Esse completano l’azione delle amilasi del tubo digerente. Utilizzate per la preparazione di sciroppi di glucosio. Endoamilasi salivari e pancreatiche: hanno una elevata attività, con pH ottimale: intorno a 7. L’amilasi pancreatica può idrolizzare 4 milioni di volte il suo peso in amido. Amilosio: lineare Amilopectina: ramificazioni circa 20-30 legami alfa 5 Maltosaccaridi lineari con grado di polimerizzazione (GP) 6-22 C Sebbene le diverse amilasi catalizzino la stessa reazione – idrolisi del legame aglicosidico sullo stesso substrato – sono differenti sia per struttura che per meccanismo di reazione. STRUTTURA Sia le a- che le -amilasi hanno una struttura a TIM-barrel [(/a)8-barrel fold]. (TIM= trioso-P-isomerasi) Il dominio catalitico consiste in una struttura a botte formata da 8 foglietti b paralleli, circondati da 8 a-eliche, collegate da regioni loop di varia lunghezza (le botti non sono identiche nei dettagli). Barrel= barile Colorazione dal blue (N-terminale) al rosso (Cterminale) La struttura delle glucoamilasi (-amilasi) consiste di 6 a-eliche parallele, circondate da altre 6 a-eliche, parallele tra loro ma antiparallele rispetto alle prime [(a/a)6-barrel fold], collegate anch’esse da regioni loop di varia lunghezza. Barrel= barile In base alle differenze e alle similarità delle strutture primarie, gli enzimi amilolitici sono stati classificati nella famiglia delle idrolasi glicosidiche (GH, EC 3.2.1.-): 1. a-amilasi – clan GH-H (famiglie GH13, GH70 e GH77) 2. -amilasi – famiglia GH14 3. glucoamilasi – famiglia GH15. Questa classificazione riflette anche le differenze nel meccanismo di reazione tra i tre tipi di amilasi. http://www.cazy.org/ Clan GH-H: famiglie GH13, GH70 e GH77 Comprende non solo idrolasi (EC 3), ma anche transferasi (EC 2) e isomerasi (EC 5): GH13, GH70 e GH77 rappresentano la famiglia delle a-amilasi. Le caratteristiche comuni ai circa 30 enzimi del clan GH-H sono: 1. Dominio catalitico TIM-barrel [(/a)8barrel fold], con un lungo loop che unisce il filamento 3 con l’elica a3, noto come dominio B; 2. Stesso meccanismo di reazione, in cui l’aspartato del filamento 4 funge da base (nucleofilo) e il glutammato 5 funge da donatore di protoni (catalisi acido-base), con l’aiuto di un terzo residuo di aspartato 7, essenziale per il legame con il substrato 3. Utilizzano un meccanismo che mantiene la conformazione anomerica del legame a-glicosidico. Presentano strutture primarie Molto diverse, sono conservati Dagli 8 ai 10 residui, essenziali Per l’attività enzimatica E H 3 e 7 D 4 e 7 Famiglia delle a-amilasi GH13 Comprende più di 30 enzimi e più di 2000 sequenze e rappresenta la più importante famiglia del clan GH-H. Famiglia delle -amilasi GH14 e delle glucoamilasi GH15 Entrambe utilizzano un meccanismo di inversione della conformazione anomerica del legame a-glicosidico: i prodotti di reazione sono infatti anomeri. -amilasi GH14 I residui coinvolti nella catalisi sono Glu186 e Glu380, al C terminale dei filamenti 4 e 7, rispettivamente. Indispensabile anche il ruolo di Asp101 e Leu383. Glucoamilasi GH15 I residui di Glu179 e Glu400 sono i residui chiave per l’attività catalitica. Amilasi utilizzate nei processi industriali: a-amilasi L’enzima più utilizzato per la degradazione industriale dell’amido è la aamilasi, una a-1,4-glucanoidrolasi. L’enzima (EC 3.2.1.1) contiene una caratteristica tasca di legame per il substrato, che può accogliere da 4 a 10 unità di glucosio della molecola di substrato, sebbene ogni sito di legame ha affinità per una sola unità di glucosio della catena polisaccaridica di amido. Le differenze, strutturali e cinetiche, tra le varie a-amilasi sono determinate da: 1. numero di siti di legame per il substrato 2. posizione del dominio catalitico 3. lunghezza del frammento oligosaccaridico rilasciato in seguito all’idrolisi 4. natura del prodotto finale 5. Capacità di rompere un legame a-1,4 in prossimità di una ramificazione a1,6 6. temperatura ottimale di reazione (varia tra i 25 e i 95°C) 7. presenza di ioni Ca2+ Le a-amilasi catalizzano la rottura del legame a-1,4-glicosidico nella porzione interna della molecola di amido, causando una rapida diminuzione del peso molecolare. Possono essere divise in: 1. a-amilasi che provocano liquefazione substrato con più di 15 unità di glucosio 2. a-amilasi che provocano saccarificazione (processo industriale che consiste in una idrolisi amilasica dell’amido gelatinizzato*, che fornisce essenzialmente maltosio). L’idrolisi prolungata dell’amilosio produce maltosio, maltotriosio e catene oligosaccaridiche di varia lunghezza, cui generalmente segue un secondo stadio della reazione che produce glucosio dal maltotriosio. * Gelatinizzazione: perdita della struttura semicristallina del granulo di amido in seguito a trattamento idrotermico ( >50°C). Riguarda soprattutto l’amilosio di cui aumenta la suscettibilità alla idrolisi. Le a-amilasi vengono utilizzate in un gran numero di processi industriali, che avvengono sotto diverse condizioni fisiche e chimiche. Ogni enzima è specifico per ciascuna applicazione Generalmente, enzimi termostabili vengono preferiti, visto che le alte temperature: favoriscono la gelatinizzazione dell’amido abbassano mediamente la viscosità accelerano la velocità della reazione abbassano il rischio di contaminazioni batteriche inattivano gli enzimi dei prodotti alimentari, che potrebbero portare alla formazione di prodotti indesiderati durante la reazione. L’a-amilasi termostabile più utilizzata è quella prodotta dal Bacillus licheniformis rimane attiva per diverse ore alla temperatura di 90°C Pyrococcus woesei 40-130°C (optimum 100°C, pH 5.5) Pyrococcus furiosus 40-130°C (optimum 100°C, pH 6.5-7.5) Tuttavia, per i processi industriali, le a-amilasi dovrebbero rimanere attive ad un pH intorno a 4.0 Nessuna delle a-amilasi termostabili ha una stabilità a tale pH, quindi studi di ingegneria proteica stanno cercando di favorire questa proprietà. Amilasi utilizzate nei processi industriali: Enzimi deramificanti Ci sono due principali gruppi di enzimi endo-deramificanti, che tagliano il legame a -1,6-glicosidico di amilopectina, glicogeno, pullulani e relativi oligosaccaridi: Pullulanasi (EC 3.2.1.41) attaccano il legame a-1,6, liberando oligosaccaridi lineari di glucosio a-1,4. Neopullulanasi e amilopullulanasi (EC 3.2.1.135 e 3.2.1.1/41) attive verso entrambi i legami (a-1,4 e a-1,6) Le pullulanasi sono piuttosto termostabili: Klebsiella pneumoniae, Bacillus acidopullulyticus 50-60°C Thermus caldophilus 75°C - fino a 90°C, pH 5.5 La maggior parte delle amilopullulanasi viene estratta da batteri termofili: Bacillus subtilis Thermoanaerobium brockii Clostridium thermosulphuricum Thermus acquaticus Pyrococcus woeser 105°C, pH 6.0 Amilasi utilizzate nei processi industriali: Eso-idrolasi -amilasi (EC 3.2.1.2) glucoamilasi (EC 3.2.1.3) Entrambe agiscono alle estremità non riducenti di amilosio, amilopectina e glicogeno, producendo carboidrati di basso peso molecolare in conformazione -anomerica. Il principale prodotto finale dell’idrolisi catalizzata dalla -amilasi è il maltosio; la glucoamilasi produce glucosio. Strutturalmente, questi due enzimi fanno parte rispettivamente delle famiglie GH14 e GH15, ma, mentre le -amilasi presentano una struttura (/a)8-barrel fold, le glucoamilasi hanno una struttura (a/a)6-barrel fold. Tutte le -amilasi non sono in grado di tagliare i legami a-1,6 e i prodotti finali sono rappresentati da maltosio e destrine “-limit” ( polimeri prodotti dall’idrolisi dell’amilopectina con amilasi, che non può idrolizzare i punti di ramificazione a-1, 6) Quando quindi la degradazione dell’amilopectina è completa, solo il 50-60% risulta convertito in maltosio. Solo nel caso dell’amilosio, che non presenta o che presenta poche ramificazioni, la resa è del 75-90%. L’accumulo di destrine -limit non è desiderabile perché aumenta la viscosità degli sciroppi di maltosio. Le -amilasi sono presenti in molte piante superiori e in microrganismi quali Pseudomonas, Bacillus e Streptococcus, ma non resistono a temperature superiori ai 60°C. Le glucoamilasi tagliano preferenzialmente i legami a-1,4, ma possono tagliare anche quelli a-1,6 e a-1,3 le glucoamilasi sono capaci di degradare completamente l’amido in glucosio A concentrazioni medie di glucosio del 30-35% durante la reazione, le glucoamilasi possono anche catalizzare la reazione inversa e formare maltosio, isomaltosio e altri sottoprodotti, abbassando quindi la resa del processo. Fonti: - Piante - Animali - Microrganismi (Saccharomices, Endomycopsis, Aspergillus, Penicillium, Mucor, Clostridium) 45-60°C (come le -amilasi, sono rare tra i microrganismi termofili) pH 4.5-5.0 Applicazioni Idrolizzati di amido La reazione di degradazione dell’amido avviene in 2 step. 1. Liquefazione Il 30-40% dei granuli di amido è gelatinizzato, ad una temperatura di 90110°C. L’aggiunta di endoamilasi termostabili (EC 3.2.1.1) in questa fase del processo protegge dal rapido aumento della viscosità della soluzione di amido, causata dal rilascio di amilosio durante il rigonfiamento dei granuli. L’idrolisi enzimatica dell’amilosio da parte dell’a-amilasi procede fino a quando la lunghezza della catena del prodotto di reazione contiene circa 1020 unità di glucosio: a questo punto, infatti, il frammento di amido prodotto non riesce più a legare bene l’enzima. Si produce quindi una miscela di malto-oligosaccaridi che contengono legami a-1,6 che l’a-amilasi non è stata in grado di rompere. 2. Saccarificazione Questo processo avviene a temperature più basse e porta all’idrolisi degli oligosaccaridi con formazione di glucosio o maltosio in reazioni catalizzate dalle glucoamilasi (EC 3.2.1.3) o dalle -amilasi (3.2.1.2), rispettivamente. La resa della reazione può essere aumentata (fino al 94%) aggiungendo pullulanasi (EC 3.2.1.41) o altri enzimi deramificanti l’uso delle pullulanasi aumenta la resa fino al 94%. Preparazione di idrolizzati d’amido Ca2+ aumenta la stabilità dell’enzima DE= destrosio equivalente Destrosio= glucosio Per l’amido: DE=1 Per il maltosio: DE=ca.50 Per il glucosio puro: DE=100 Preparazione di idrolizzati d’amido a-amilasi termostabili (B. lichenifromis 90°C, pH 5.5-6.0) -amilasi e glucoamilasi non termostabili La necessità di aggiustare pH e temperatura, rendono il processo più costoso e rendono necessario un ulteriore passaggio cromatografico sul prodotto finale per eliminare l’NaCl formatosi Un importante sviluppo di questo processo potrebbe essere rappresentato dalla degradazione dell’amido in un unico step: 1. a-amilasi più stabile termicamente, che richieda un pH più basso e non richieda l’utilizzo di sali di calcio per la propria attività 2. Tutti gli enzimi della fase di saccarificazione attivi a condizioni compatibili con quelle dell’a-amilasi a-glucosidasi (EC 3.2.1.20) idrolisi del legame a-1,4 all’estremità non-riducente e a-1,6, con formazione di glucosio come prodotto finale. Isomerizzazione del glucosio L’isomerizzazione del glucosio derivato dall’amido in fruttosio conferisce una maggiore dolcezza agli sciroppi comunemente utilizzati per i prodotti alimentari e le bevande (saccarosio 1; fruttosio 1,35-1,7; glucosio e galattosio 0,75; maltosio 0,4). Lo sciroppo di fruttosio viene generalmente prepaarto utilizzando un processo in continuo catalizzato dalla glucosio isomerasi (EC 5.3.1.5) immobilizzata, ad una temperatura di 55-60°C. A queste condizioni, solo il 40-42% di glucosio viene convertito in fruttosio. La resa del processo può essere aumentata alle alte temperature che spostano l’equilibrio della reazione verso un aumento della concentrazione del fruttosio. Questo però fa diminuire la vita media dell’enzima, estratto da un batterio mesofilo, e aumenta la quantità di sottoprodotti che si formano in seguito alla reazione di Maillard che interviene proprio ai valori di pH leggermente alcalini necessari per l’attività della glucosio isomerasi. Anche in questo caso è necessaria una gel filtrazione per allontanare il glucosio e altri grossi polissaccaridi presenti. Questo passaggio può essere evitato utilizzando una glucosio isomerasi maggiormente termostabile, che sia attiva ad un pH più acido, in modo da evitare indesiderate reazioni collaterali. Produzione di trealosio Gli sciroppi di amido o di maltosio possono essere trasformati in trealosio, in una reazione catalizzata da un enzima isolato da microrganismi termofili o mesofili. Il trealosio è un disaccaride stabile, non riducente, formato da un legame a-1,1 tra due molecole di glucosio. E’ coinvolto nella protezione delle strutture biologiche durante il congelamento, essiccamento o la cottura. I cristalli amorfi di trealosio : intrappolano le molecole biologiche, senza modificarne la struttura nativa e limitando di conseguenza i danni alle sostanze durante l’essiccamento sono permeabili all’acqua ma impermeabili agli esteri aromatici idrofobici: questo limita la perdita di composti aromatici responsabili de sapori e questo favorisce la produzione di cibi essiccati che mantengano lo stesso sapore del prodotto fresco. Il trealosio inoltre: è leggermente dolce è solubile è stabile a pH bassi riduce l’attività dell’acqua ha bassa igroscopicità abbassa la temperatura di congelamento non caramellizza non subisce la reazione di Maillard prezioso ingrediente alimentare Viene utilizzato per: bevande, cioccolata e caramelle, prodotti da forno, latticini, derivati della frutta…….. Gli enzimi utilizzati per la produzione di trealosio dall’amido sono: 1.maltooligosil-trealosio sintasi converte il legame a-1,4 terminale dell’estremità riducente del maltooligosaccaride nel legame a-1,1 del trealosio 2.maltooligosil-trealosio trealo-idrolasi rimuove il trealosio durante l’idrolisi del legame a-1,4 tra la seconda e la terza unità di glucosio, ripetendo questa operazione fino a quando la catena oligosaccaridica rimanente consiste di 2, massimo 3 unità di glucosio. Il maltooligosaccaride di partenza viene prodotto trattando l’amido liquefatto con enzimi deramificanti. Sintesi di ciclodestrine Gli enzimi amilolitici (batterici) vengono utilizzati anche per la produzione di ciclodestrine. 1. a-amilasi e pullulanasi oligosaccaridi non ramificati 2. Ciclomaltodestrina glucanotransferasi taglio degli oligosaccaridi lineari per ottenere oligosaccaridi di 8 unità. Questo enzima è estremamente termostabile (fino a 100°C) e possiede anche un’attività a-amilasica e questo consente la produzione di ciclodestrine senza l’aggiunta delle a-amilasi per la liquefazione preliminare. A causa della struttura elicoidale di questi oligosaccaridi, le due estremità di ciascuna molecola sono molto vicine e di conseguenza possono facilmente unirsi a formare la struttura caratteristica delle ciclodestrine. Il prodotto finale è una miscela di a-, - e -ciclodestrine, a 6, 7 o 8 residui di glucosio con legami a1,4. Le ciclodestrine hanno una struttura conica in cui i gruppi ossidrilici sono localizzati sulla superficie, mentre l’interno è apolare e può facilmente formare complessi con molecole idrofobiche . Es. 1. Rimozione dell’amaro dai succhi di agrumi 2. Protezione dei lipidi dall’ossidazione 3. Rimozione del colesterolo dalle uova Incapsulaggio di principi aromatici
© Copyright 2026 Paperzz