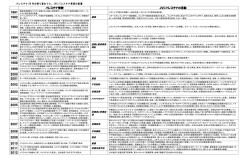
京府医大誌 122 (6),371~379,2013. 繊毛による左右決定機構 <特集「個体発生と細胞分化の医学」 > 繊毛は身体の「右と左」を決める 小林 大介*,横山 尚彦 京都府立医科大学大学院医学研究科生体機能形態科学 Ci l i aDet er mi neLef t r i ghtAs ymmet r yoft heBody Da i s uk eKo b a y a s hia ndTa ka hi k oY o k o y a ma De p a r t me nto fAna t o mya ndDe v e l o p me nt a lBi o l o g y , Ky o t oPr e f e c t ur a lUni v e r s i t yo fMe d i c i neGr a d ua t eSc h o o lo fMe d i c a lSc i e nc e 抄 録 身体の左右は,外見上は対称であるが,内部の臓器を見ると多くの左右非対称性を見いだすことが出 来る.この非対称性が逆転する変異体は,内蔵逆位と呼ばれる.近年,繊毛局在蛋白や繊毛の運動性に 関与する遺伝子が,内臓逆位を示すヒトやマウスの遺伝的変異体の原因遺伝子としてクローニングされ てきた. この左右非対称性決定に原始結節の細胞が持つ繊毛が重要な役割を果たしていることが分かってき た.繊毛は,細胞表面より突出した直径約 200nmの毛状の構造体で,運動性と非運動性繊毛がある. 左右非対称性決定には,繊毛運動に関与する遺伝子と関与しない遺伝子の両者が必要であることが分 かってきた.本総説では,左右非対称性決定に関わる繊毛の運動性と非運動性の原因遺伝子と,その作 用機序について紹介する. キーワード:繊毛,内臓逆位,カルタゲナー症候群,ダイニン,若年性ネフロン癆. Abs t r ac t Ourv e r t e b r a t eb o d yi se x t e r na l l ys y mme t r i c a l .Ho we v e r ,a l mo s ta l li nt e r na lo r g a ns ,s uc ha st he he a r ta nds t o ma c h,a r ea s y mme t r i c a l l ypo s i t i o ne d .Thec o nd i t i o ni nwhi c ht hi sa s y mme t r yi sr e v e r s e d i sc a l l e ds i t usi nv e r s us .Thel e f t r i g hta s y mme t r yi sg e ne t i c a l l yd e t e r mi ne d ,a ndanumb e ro fc a us a t i v e g e ne swho s ed e f e c tr e s ul t si ns i t usi nv e r s usha v eb e e nc l o ne d . Ana l y s i so ft he s eg e nepr o d uc t sha sr e v e a l e dt ha tc i l i aha v ea ni mpo r t a ntr o l et oe s t a b l i s ht hel e f t r i g hta s y mme t r y .Ci l i aa r eat i nyo r g a ne l l et ha tpr o t r ud e sf r o mt hec e l ls ur f a c e .So mec i l i as ho wmo t i l i t y . Bo t hmo t i l ea ndno nmo t i l ec i l i aa r eno wkno wnt ob ei mpo r t a ntt oe s t a b l i s ht hel e f t r i g hta s y mme t r y . Thi sr e v i e wi nt r o d uc e sc i l i a r yc o mpo ne nt st ha ta r ei mpo r t a ntf o rt hel e f t r i g hta s y mme t r ye s t a b l i s hme nt . KeyWor ds :Ci l i a ,Si t usi nv e r s us ,Ka r t a g e ne rs y nd r o me ,Dy ne i n,Ne phr o no pht hi s i s . 平成25年 5月 7日受付 *連絡先 小林大介 〒602 ‐8566京都市上京区河原町通広小路上ル梶井町465番地 kd a i s uk e @k o t o . kpum. a c . j p 371 小 372 は じ め 林 に 私たち脊椎動物の形態は,頭尾,背腹,左右 の 3つの軸によって決定されている.頭尾軸は 頭とお尻の方向を決定する.背腹軸は背中とお 腹の決定に関与する.左右軸は右と左の決定を する軸である.発生過程において左右軸は頭尾 軸,背腹軸が決定されてから決定される.たっ た一つの細胞から始まる私たちの身体がどのよ うに様々な形態や機能をもつ身体に形成されて くるかは,古くから多くの研究者を魅了してき ている.特に,体軸決定(頭尾軸,背腹軸,左 右軸決定)に関しては,対称性から非対称性の 形成という点から興味が持たれてきた. 身体の左右非対称性が逆転する変異体は一般 的に内臓逆位と言われる.近年,内臓逆位を示 す人やマウスの遺伝的変異体の原因遺伝子ク ローニングや,左右非対称的に発現する遺伝子 の発見により,左右非対称性決定機構が大きく 解明されてきた. 左右非対称性形成は,1)左右相称に発生し てきた対称性の破れ,2)左右非対称な遺伝子 発現,3)左右特異的な器官形成,の 3ステッ プに分けられる.この中で繊毛が重要な役割を 大 介 ほか 果たしているのがステップ 1である.マウスに おいて受精後 7. 5日胚で一過的に形成される原 始結節(ノード)と呼ばれる器官が左右決定に 非常に重要な役割を果たしている.原始結節は 「へそ」のような構造で中心部がくぼみ(原始窩 pi t )周辺部がやや隆起(c r o wn )した構造をと る(図 1 ) .原始結節を形成する細胞は 1本の繊 毛を持つ.この繊毛は回転運動をし,その回転 運動により引きおこされる左向きの水流(ノー ド流)が左右の非対称性を生み出す(図 1 ) .こ の繊毛運動に障害が生じ,ノード流を作ること が出来ないマウスは左右軸の決定がランダマイ ズし,半数の個体で内臓逆位を生じる.また人 工的に右向きの水流を与えると,左右が逆転す ることも分かっている1).このノードに相当す る運動繊毛を持った器官は,哺乳類のみならず 鳥類,両生類,魚類でも保存されており2),ノー ド流に相当する水流や,運動繊毛の障害による 内臓逆位も報告されている.さらに,左右非対 称性決定における繊毛の機能はノード流を作り 出すことのみではないことがわかってきてい る.本稿では,運動性および非運動性繊毛の身 体の左右決定への関与について紹介する. 図 1 ノード(原始結節)の運動繊毛とノード流.ノードに存在する運動性繊毛が回転運動するこ とにより,ノード内に左向きの水流を生じる. 繊毛による左右決定機構 繊 毛 の 構 造 繊毛は,細胞表面より突出する直径約 200nm の毛状の構造体である.長さは,大体 1 μmから 10 μmまで様々である.内部は微小管が規則正 しく配列し,軸糸と呼ばれる構造を形成してい る(図 2A) .運動性と非運動性繊毛が知られてお り,軸糸構造が異なる.古典的に知られた運動繊 毛の軸糸は,A小管と B小管からなる 9個の周 辺微小管(Pe r i phe r a lMi c r o Tub ul e ,PMT)と 2 本の中心微小管(Ce nt r a lPa i r ,CP)を持ち,9+ 2構造と呼ばれている.PMTには外腕および内 腕ダイニン (Out e r / I nne rDy ne i nAr m, ODA/ I DA) と呼ばれるタンパク質複合体が付属している.ダ イニン腕(Dy ne i nAr m,DA)は ATP加水分解 反応のエネルギーにより繊毛を動かすモーター の役割を果たしている.またこれらの構造の他 に,隣接する PMTをつなぐネキシンリンク(ダ イニン制御複合体,d y ne i nr e g ul a t o r yc o mpl e x , DRCとも呼ばれる)や,CPから DAへシグナル を伝えるラディアルスポークなどの構造も鞭 毛・繊毛が動くための精密機械の一部としてそ の役割を果たしている.同じく運動性繊毛でも, ノード及びそれに相同な器官の繊毛は,CPを 欠いた 9+0構造を持ち,PMTにはDAが付属し ている.DAがモータータンパク質複合体であ ることからもわかるように,ノードの繊毛は運 動性を持つ3).一方多くの細胞がもつ一次繊毛 は,9+0構造であるが,DAをもたない.従っ 373 て,運動性を示さない.視細胞や嗅細胞では, この一次繊毛が特殊化した繊毛が,光や匂いの 受容体となっている. 運動繊毛障害症候群 運動繊毛は左右軸決定以外にも様々な生命現 象に関与することが知られている.例えば鼻 腔・気管上皮の運動繊毛は外界からの異物を体 外へ排出する働きを持ち,この機能が障害され ると,慢性の副鼻腔炎や気管支炎を引き起こ す.また,精子の鞭毛は,遊泳に必須であり, これが障害されると男性不妊となる.これらは 総称して運動繊毛障害症候群(Pr i ma r yCi l i a r y Dy s ki ne s i a ,PCD) と呼ばれ,そのうち内臓逆位を 併発するものをカルタゲナー症候群(Ka r t a g e ne r Sy nd r o me ,KS)と呼ぶ.興味深いのは,すべて の PCDにおいて内臓逆位が観察されるわけで はないことである.これはノード繊毛の構造が 他の部位の運動繊毛と異なることに起因すると 考えられるが,PCDの原因遺伝子の同定と表現 型の解析から,この予想が正しいことがわかっ てきた. PCD で観察される 繊毛構造異常と責任遺伝子並びに 内臓逆位の関係について PCD原因遺伝子はこれまでに 19遺伝子が同 4 ‐ 15) .観察される繊毛の構造 定されている(表 1 ) 異常に注目すると,内臓逆位を伴う PCDは DA 図 2 A:繊毛の構造(横断面) .B:クラミドモナス外腕ダイニンの構造(模式図)と対応するヒトのサブユニット. 374 小 林 大 介 ほか 表 1 これまでに同定された PCDの責任遺伝子 略号:DNAI ;Dy ne i n,a x o ne ma l ,i l nt e r me d i a t ec ha i n:DNAH;Dy ne i n,a x o ne ma l ,he a v yc ha i n:CCDC;Co i l e d c o i ld o ma i n c o nt a i ni ng :DNAL;Dy ne i n, a x o ne ma l , l i g htc ha i n:TXNDC;t hi o r e d o x i nd o ma i nc o nt a i ni ng :NME;NME/ NM23f a mi l y me mb e r :LRRC;Le uc i ne r i c hr e pe a tc o nt a i ni ng :Dy ne i na x o ne ma la s s e mb l yf a c t o r :HEATR;HEAT r e pe a t c o nt a i ni ng :HYDI N;a x o ne ma lc e nt r a lpa i ra ppa r a t uspr o t e i n:RSPH4A;Ra d i a ls po k ehe a d4ho mo l o gA:RSPH9; Ra d i a ls po k ehe a d9ho mo l o g :PMT;Pe r i phe r a lMi c r o Tub ul e :CP;Ce nt r a lPa i r :RS;Ra d i a lSpo k e :DRC;Dy ne i n Re g ul a t o r yCo mpl e x .Mo r pha nt ;モルフォリノアンチセンスオリゴヌクレオチドによって一過的に遺伝子の機能を ノックダウンした胚.上記 Re f e r e nc eのうち参考文献リストにないものは, 4)Ko b a y a s hi&Ta k e d a ,Di f f e r e nt i a t i o n (2012 )を参照のこと. の異常を伴い,内臓逆位を伴わない PCDでは 他の構造の異常が観察される.後者のケースと して,HYDI N,RSPH4A,RSPH9に変異を持つ 患者の繊毛(主に 9 +2構造を持つ気管支や鼻 腔の繊毛が解析に利用されている)で観察され る構造異常は CP並びにそれに付属する構造の 異常である.これらはノードの繊毛には無い構 造であり,これらの因子がノードの繊毛には必 要ないことと一致している. 一方,内臓逆位を伴い DAに異常を持つタイ プの原因遺伝子はいくつかのタイプに分類で きる.最もよく観察される構造の異常は ODA の欠損である.この表現型を示す PCDの原因 遺 伝 子 は DNAH5 ,DNAI 1 ,DNAI 2 ,DNAL1 , TXNDC3/ NME8 ,CCDC114であり,CCDC114 を除き ODAのサブユニットをコードすること 繊毛による左右決定機構 が分かっている (図 2B) .DAが繊毛内に輸送さ れるためには,細胞質での複合体形成が必須で あり,DAサブユニットを欠いたクラミドモナ ス変異体ではこの複合体を形成できず,繊毛内 の DAの欠損が観察されることが知られてい る.これは ODAのサブユニット異常により ODAが欠損するヒトの症例とよく一致する. 例外的に DNAH11の症例では ODAが観察され るが,これは完全な機能喪失に至っていないた めと考えられている.また CCDC114は DAの PMTへの結合に必要な因子であることが分 かっている14)15). もう一つの例は ODA,I DA共に欠くものであ る.この中には細胞質内における DA複合体形 成に必須な細胞質因子に変異を持つ例が 3例 報告されている(DNAAF1/ LRRC50,DNAAF2/ 5) 16) 1 7) .この他の ODA,I DA KTU/ PF13,DNAAF3 ) を共に欠損するケースとして,HEATR2 ,LRRC6 , CCDC103が 報 告 さ れ て い る.CCDC103は, DAの軸糸へのアンカーに必須のタンパク質であ ることが分かっているが,HEATR2と LRRC6に 関して,正常な DAを形成するためのどのス テップに関連しているかの詳細な研究はまだな い8‐11). この他ネキシンリンクを欠く DRC1では内臓 逆位の報告はないが,マウスのノードにおける 発現は確認されており,左右軸の決定に関与す ,並び る可能性は十分にある13).更に CCDC39 に CCDC40では,繊毛の構造の広い範囲で異常 が観察されている18). 以上のように,内臓逆位をもたらす運動繊毛 の異常の遺伝的背景には様々な因子が存在す る. 繊毛運動に関係しない繊毛蛋白も 身体の左右決定に関与する 原始結節の繊毛の左右決定に関する役割は, その運動によってノード流を作り出すことのみ ではない.繊毛運動とは関係のない繊毛局在蛋 白も,左右決定に関与することが知られてい る. 繊毛運動に関与しなくても左右決定に関与す 375 る繊毛蛋白として常染色体優性多発性嚢胞腎 Aut o s o ma ld o mi na ntpo l y c y s t i cki d ne yd i s e a s e (ADPKD)と若年性ネフロン瘻(NPHP)があ る.面白いことに,これらの遺伝子は嚢胞性腎 疾患の原因遺伝子でもある. ADPKDはヒト遺伝性疾患の中でも頻度が最 も高い疾患であり,その原因遺伝子として, PKD1と PKD2がクローニングされている19)20). ヒト PKD1は 4302アミノ酸をコードしており, 大きな細胞外ドメインを有する非常に大きな蛋 白である.ヒト PKD2は 968アミノ酸をコード しており,非特異的カルシウムチャネルとして 機能すると考えられる.それぞれの遺伝子産物 は膜貫通蛋白であり,po l y c y s t i n1,po l y c y s t i n2 といわれる.PKD1,PKD2ともに腎臓嚢胞の原 因遺伝子と同定されたが,左右非対称性決定に おける PKDの関与は,p k d 2kno c k o utmo us eに おいて左右非対称性異常が出現する報告に始 k d 2変異を まった21).さらに,ヒトにおいても,p もち内蔵逆位が合併する症例が報告された22). Na ul i ら23)は,水流刺激により正常腎臓細胞か らカルシウムが流入するのに対して,ADPKD1 変異細胞や po l y c y s t i n2に対する抗体処理によ りカルシウムが流入しなくなることを報告して おり,Po l y c y s t i n (ポリシスチン)の機能として は,機械的刺激に対して反応してカルシウム流 入させると考えられている.Po l y c y s t i n1,2と もに繊毛に局在する.腎臓では,po l y c y s t i n1,2は複合体を形成することが知られている.原 始 結 節 に お い て は po l y c y s t i n1に 代 わ り po l y c y s t i n1L1 (PKD1L1原因遺伝子産物)が po l y c y s t i n2と複合体を形成して機能すると考 えられている.Pk d 1L1のマウス,メダカの変 異体で内臓逆位が生じる24)25). po l y c y s t i n2,po l y c y s t i n1L1は繊毛運動と無 関係だとすると,左右非対称性決定に関与して いる機能は何であろうか.仮説としては,原始 結節の繊毛の動きにより生じたノード流を po l y c y s t i n複合体が感じてカルシウムが流入す ることで左右非対称性が生じるということであ る.実際に,原始結節における細胞内カルシウ ムの発現に左右差があることが 2つ報告されて 376 小 林 いる.さらに,Y o s hi b aら26)は,カルシウム阻害 剤 Gd CL3, 2ABP , t ha ps i ng a r g i nで処理した胚で は,下流遺伝子の左右非対称的発現が消失する ことを報告した. 原始結節の繊毛では,中央のくぼんだ部位 (原始窩,pi t )に存在する細胞と原始窩周囲の土 手の部分(c r o wn )に存在する細胞の繊毛で挙 動が異なる.Pi tc e l l に存在する繊毛は可動性 であるが,c r o wnc e l l の繊毛は発生時期により 2 -/ -マウ 運動性が異なる.Y o s hi b aら26)は pkd スに c r o wnc e l l のみで pkd 2を発現させ,左右非 対称な遺伝子発現が回復することを示した.ま た,po l y c y s t i n2の繊毛局在シグナルを除去し た場合,po l y c y s t i n2の左右決定機能が消失す ることを示した. 以上から,原始結節における繊毛は,運動に よりノード流を形成するのみでなく,多分ノー ド流により形成される何かを感受し(機械的に もしくは化学的に) ,po l y c y s t i n2から流入した カルシウムが左右非対称的遺伝子発現を誘導し ていると考えられる. 内蔵逆位は,若年性ネフロン瘻(NPHP)に おいても観察される.NPHPの原因遺伝子は現 大 介 ほか 在まで 13遺伝子が報告されている.しかし,こ れら遺伝子の変異が見つかっているのは,全症 例の約 3割であり,残りの大多数の原因遺伝子 は同定されていない27). NPHPは内蔵逆位,嚢胞性腎疾患の原因とな る以外に,その異常は網膜色素変性症などの多 くの疾患を引き起こし,様々な症候群の原因と なることも分かっている.例えば,NPHP6は CEP290とも呼ばれているが,BBS(Ba r d e t Bi e d l 症候群) ,J BTS(J o ub e r t症候群) ,MKSの 原因ともなることが報告されている.PKDは, NPHP ,BBS,J BTS,MKSとの合併の報告はな い.従って,繊毛運動に関与せず左右決定に関 与する繊毛蛋白群は,PKDグループと NPHPグ ループの 2つに大きく分けることが出来ると考 えられる. Ne phr o c y s t i nは NPHPの遺伝子産物である が,Ne phr o c y s t i nsの繊毛における局在には特 28) .一言に繊毛といっても,繊 徴がある(図 3 ) 毛は単純な構造ではない.図 3に示すように, 繊毛の基部は基底小体と呼ばれるが,これは 母中心体と同じものである.細胞体から突出 する繊毛本体は,先端部(t i p) ,体部(s ha f t ), 図 3 ネフロシスチン(NPHP)の繊毛局在とネフロシスチン・モデュール 基底小体には nphp5,nphp6 ,移行帯に nphp1,nphp4,nphp8 ,I nvc o mpa r t me ntに nphp2/ I nv ,nphp3,nphp9が局在する. 繊毛による左右決定機構 移 行 帯(t r a ns i t i o na lz o ne )に 分 か れ て い る. Ne phr o c y s t i n1,4,8は移行帯,Ne phr o c y s t i n5,6は中心体,Ne phr o c y s t i n2,3,9は体部の I nvc o mpa r t me nt と呼ばれる基底小体に近い部 位に局在する.この c o mpa r t me ntは形態学的 には際立った特徴がない.また,Ne phr o c y s t i n 間の結合では,NPHP1,4,8と NPHP5,6およ びNPHP2, 3, 9が免疫沈降による結合が確かめら れている(図 3 ) .内臓逆位が観察されるのは,主 として I nvc o mpa r t me ntに局在する I nv / np h p 2, np h p 3,np h p 8/ ne k 8遺伝子に変異があった個体 である.したがって,NPHPの中でも,その局 在により機能が異なる可能性がある. これらの遺伝子産物は po l y c y s t i nと同様にカ ルシウム流入と関連があるのであろうか.我々 は,i nv / nphp2の変異マウスより分離した腎臓 尿細管細胞を用いてカルシウム流入を検討した が,野生型細胞との間に差は認められなかっ nv / nphp2遺伝子産物などの た29).したがって,i ne phr o c y s t i nは po l y c y s t i nとは異なった機構で 文 377 左右非対称性形成に関与していると考えられ る.Ne phr o c y s t i nの機能は腎臓で研究され様々 な提唱がなされているが,左右決定にどのよう な機構で関与しているかは現在のところ不明で ある. 終 わ り に 発生における繊毛機能の中で,左右決定にお ける役割を紹介した.しかし,繊毛の発生にお ける役割は,これ以外にも,脳,腎臓,手足, 骨,感覚器,精子形成など多岐にわたる.さら に,繊毛は中心体と深い関係があり,細胞周期 によりその存在は制御される.逆に,繊毛によ り細胞周期制御も考えられ,腫瘍形成との関与 も提唱されている. なぜ,繊毛がこのような多彩な機能をもつの か,また,持つようになったのか? 繊毛局在 因子の機能解明がさらに必要である. 開示すべき潜在的利益相反状態はない. 献 1)No na ka S,Shi r a t o r iH,Sa i j o hY ,Ha ma d a H. Lo g e sNT,Ra i d tJ ,e ta l .Re c e s s i v eHYDI NMut a t i o ns De t e r mi na t i o no fl e f t r i g htpa t t e r ni ngo ft hemo us e Ca us ePr i ma r yCi l i a r yDy s ki ne s i awi t ho utRa nd o mi z a - e mb r y ob ya r t i f i c i a lno d a lf l o w.Na t ur e2002;418:969. t i o no fLe f t Ri g htBo d yAs y mme t r y .TheAme r i c a n Pub Me dPMI D:d o i : 10. 1038/ na t ur e 00849. 2)Es s ne rJ J ,Vo g a nKJ ,Wa g ne rMK,Ta b i nCJ ,Y o s tHJ , J o ur na lo fHuma nGe ne t i c s2012. 7)Ca s t l e ma nVH,Ro mi oL,Cho d ha r iR,Hi r s tRA,d e Br ue c kne rM. Co ns e r v e df unc t i o nf o re mb r y o ni cno d a l Ca s t r oSCP ,Pa r k e rKA,e ta l .Mut a t i o nsi nRa d i a l c i l i a .Na t ur e2002;41 ( 86893 ) :37. Spo k e He a d Pr o t e i n Ge ne s RSPH9 a nd RSPH4A 3)Oka d aY ,Ta k e d aS,Ta na kaY ,Be l mo nt eJ CI ,x 00f a , Ca us e Pr i ma r y Ci l i a r y Dy s ki ne s i a wi t h Ce nt r a l - Hi r o ka waN.Me c ha ni s mo fNo d a lFl o w:ACo ns e r v e d Mi c r o t ub ul a r Pa i rAb no r ma l i t i e s .TheAme r i c a nJ o ur - Sy mme t r y Br e a ki ng Ev e nt i n Le f t Ri g ht Ax i s De t e r mi na t i o n.Ce l l2005;121:63344. na lo fHuma nGe ne t i c s2009;84:197209. 8)Pa ni z z iJ R,Be c k e r He c kA,Ca s t l e ma nVH,A Al - 4)Ko b a y a s hiD,Ta k e d a H.Ci l i a r y mo t i l i t y :The Mut a i r iD,Li uY ,Lo g e sNT,e ta l .CCDC103mut a t i o ns c o mpo ne nt sa nd c y t o pl a s mi c pr e a s s e mb l y me c ha - c a us e pr i ma r y c i l i a r y d y s ki ne s i ab y d i s r upt i ng ni s ms o ft he a x o ne ma ld y ne i ns . Di f f e r e nt i a t i o n; a s s e mb l yo fc i l i a r yd y ne i na r ms .Na t ur eg e ne t i c s2012; r e s e a r c hi nb i o l o g i c a ld i v e r s i t y20122;83:S23S9. 5)Mi t c hi s o nHM,Sc hmi d t sM,Lo g e sNT,Fr e s ho urJ , a d v a nc eo nl i nepub l i c a t i o n. 9)Ho r a niA,Dr ul e y TE,Za r i wa l a MA,Pa t e lAC, Dr i t s o ul aA,Hi r s tRA,e ta l .Mut a t i o nsi na x o ne ma l Le v i ns o nBT,Va nAr e nd o nkLG,e ta l .Who l e Ex o me d y ne i na s s e mb l yf a c t o rDNAAF3c a us epr i ma r yc i l i a r y Ca pt ur ea ndSe q ue nc i ngI d e nt i f i e sHEATR2Mut a t i o n d y s ki ne s i a .Na t ur eg e ne t i c s 2012;a d v a nc eo nl i ne a s a Ca us eo fPr i ma r y Ci l i a r y Dy s ki ne s i a .The pub l i c a t i o n. 6)Ol b r i c hH,Sc hmi d t sM,We r ne rC,Ono uf r i a d i sA, Ame r i c a nJ o ur na l o fHuma nGe ne t i c s2012;91:68593. 10 )Ho r a niA,Fe r k o lTW,Sho s e y o vD,Wa s s e r ma nMG, 378 小 林 大 介 ほか Or e nYS,Ke r e m B,e ta l .LRRC6Mut a t i o nCa us e s Ve l d hui s e n B,Sa r i sJ J ,e ta l .PKD2,a g e ne f o r Pr i ma r yCi l i a r yDy s ki ne s i awi t hDy ne i nAr m De f e c t s . po l y c y s t i cki d ne yd i s e a s et ha te nc o d e sa ni nt e g r a l Pl o So ne201303/ 19;8:e 59436. me mb r a nepr o t e i n.Sc i e nc e1996Ma y31;27 ( 25266 ) : 11 )Ko t tE,Duq ue s no yP ,Co pi nB,Le g e nd r eM,Da s t o t - 133942.Pub Me dPMI D:8650545. LeMo a lF ,Mo nt a nt i n G,e ta l .Lo s s o f Func t i o n 20 )Thepo l y c y s t i cki d ne yd i s e a s e1g e nee nc o d e sa14 Mut a t i o nsi nLRRC6,aGe neEs s e nt i a lf o rPr o pe r kbt r a ns c r i pta ndl i e swi t hi nad upl i c a t e dr e g i o no n Ax o ne ma lAs s e mb l yo fI nne ra ndOut e rDy ne i nAr ms , c hr o mo s o me 16.The Eur o pe a n Po l y c y s t i cKi d ne y Ca us e Pr i ma r y Ci l i a r y Dy s ki ne s i a .The Ame r i c a n Di s e a s eCo ns o r t i um.Ce l l1994J un17;77:88194. J o ur na lo fHuma nGe ne t i c s2012;91:95864. 12 )Zit ki e wi c z E,Buk o wy Bi e r y o Z,Vo e l k e lK, Pub Me dPMI D:8004675. 21 )Pe nne ka mpP ,Ka r c he rC,Fi s c he rA,Sc hwe i c k e r tA, Kl i me kB, Dme s kaH, Po g o r z e l s ki A, e ta l . Mut a t i o ns Skr y a b i nB,Ho r s tJ ,e ta l .Thei o nc ha nne lpo l y c y s t i n- i nRa d i a lSpo k eHe a dGe ne sa ndUl t r a s t r uc t ur a lCi l i a 2i sr e q ui r e df o rl e f t r i g hta x i sd e t e r mi na t i o ni nmi c e . De f e c t si nEa s t Eur o pe a nCo ho r to fPr i ma r yCi l i a r y Cur r e ntb i o l o g y :CB2002J un4;12:93843.Pub Me d Dy s ki ne s i aPa t i e nt s .Pl o So ne201203/ 20;7:e 33667. 13 )Wi r s c he l lM,Ol b r i c hH,We r ne rC,Tr i t s c hl e rD, PMI D:12062060. 22 )Ba t a i l l eS,De mo ul i nN,De v uy s tO,Aud r e z e tMP , Bo we rR,Sa l eWS,e ta l .Thene x i nd y ne i nr e g ul a t o r y Da ha n K,Go d i n M,e ta l .As s o c i a t i o no fPKD2 c o mpl e xs ub uni tDRC1i se s s e nt i a lf o rmo t i l ec i l i a (po l y c y s t i n2 )mut a t i o ns wi t hl e f t r i g ht l a t e r a l i t y f unc t i o ni na l g a ea ndhuma ns .Na t ur eg e ne t i c s2013; d e f e c t s .Ame r i c a nj o ur na lo fki d ne yd i s e a s e s :t he 45:2628. o f f i c i a lj o ur na lo ft heNa t i o na lKi d ne yFo und a t i o n2011 14 )Ono uf r i a d i sA,Pa f fT,Ant o nyD,Sho e ma r kA,Mi c ha D,Kuy t B,e ta l .Spl i c e Si t e Mut a t i o ns i nt he Se p;58:45660.Pub Me dPMI D:21719175. 23 )Na ul i SM, Al e ng ha tFJ , LuoY , Wi l l i a msE, Va s s i l e vP , Ax o ne ma lOut e rDy ne i nAr m Do c ki ngCo mpl e xGe ne LiX,e ta l .Po l y c y s t i ns1a nd2me d i a t eme c ha no s e ns a - CCDC114 Ca us e Pr i ma r y Ci l i a r y Dy s ki ne s i a .The t i o ni nt hepr i ma r yc i l i um o fki d ne yc e l l s .Na t ur e Ame r i c a nJ o ur na lo fHuma nGe ne t i c s2013;92:8898. 15 )Kno wl e sMi c ha e lR,Le i g hMa r g a r e tW,Os t r o ws ki g e ne t i c s 2003 Fe b ;33:12937.Pub Me d PMI D: 12514735. La wr e nc eE,Hua ngL,Ca r s o nJ o hnnyL,Ha z uc ha 24 )Fi e l dS,Ri l e yKL,Gr i me sDT,Hi l t o nH,Si mo nM, Mi l a nJ ,e ta l .Ex o meSe q ue nc i ngI d e nt i f i e sMut a t i o ns Po wl e s Gl o v e rN,e ta l .Pkd 1l 1e s t a b l i s he sl e f t r i g ht i nCCDC114a saCa us eo fPr i ma r yCi l i a r yDy s ki ne s i a . a s y mme t r y a nd phy s i c a l l y i nt e r a c t s wi t h Pkd 2. TheAme r i c a nJ o ur na lo fHuma nGe ne t i c s20131/ 10/ ; De v e l o pme nt 2011 Ma r ; 138: 113142. Pub Me d 92:99106. 16 )Omr a nH,Ko b a y a s hiD,Ol b r i c hH,Ts uka ha r aT, PMI D:21307093.Pub me dCe nt r a lPMCI D:3042869. 25 )Ka mur aK,Ko b a y a s hiD,Ue ha r aY ,Ko s hi d aS,I i j i ma Lo g e sNT,Ha g i wa r aH,e ta l .Kt u/ PF13i sr e q ui r e df o r N,Kud oA,e ta l .Pkd 1l 1c o mpl e x e swi t hPkd 2o n c y t o pl a s mi c pr e a s s e mb l y o fa x o ne ma ld y ne i ns . mo t i l ec i l i aa ndf unc t i o nst oe s t a b l i s ht hel e f t r i g ht Na t ur e2008De c4;45 ( 67222 ) :6116.Pub Me dPMI D: a x i s .De v e l o pme nt2011Ma r ;138:11219.Pub Me d 19052621.e ng . PMI D:21307098. 17 )Lo g e sNT,Ol b r i c hH,Be c k e r He c kA,Ha f f ne rK, 26 )Y o s hi b a S,Shi r a t o r iH,Kuo I Y ,Ka wa s umiA, He e rA,Re i nha r d C,e ta l .De l e t i o ns a nd po i nt Shi no ha r aK,No na kaS,e ta l .Ci l i aa tt heno d eo f mut a t i o nso fLRRC50c a us epr i ma r yc i l i a r yd y s ki ne s i a mo us e e mb r y o ss e ns e f l ui d f l o w f o rl e f t r i g ht d uet od y ne i na r md e f e c t s . AmJHumGe ne t2009De c ; d e t e r mi na t i o nv i aPkd 2.Sc i e nc e2012Oc t12;338 85:8839.Pub Me dPMI D:19944400.e ng . (6104 ) :22631.Pub Me dPMI D:22983710. 18 )Me r v e i l l e AC, Da v i s EE, Be c k e r He c k A, 27 )Ha l b r i t t e rJ ,Po r a t hJ D,Di a zKA,Br a unDA,Ko hlS, Le g e nd r eM,Ami r a vI ,Ba t a i l l eG,e ta l .CCDC39i s Cha kiM,e ta l .I d e nt i f i c a t i o no f99no v e lmut a t i o nsi n r e q ui r e df o ra s s e mb l yo fi nne rd y ne i na r msa ndt he a wo r l d wi d e c o ho r to f 1, 056 pa t i e nt s wi t h a d y ne i nr e g ul a t o r yc o mpl e xa nd f o rno r ma lc i l i a r y ne phr o no pht hi s i s r e l a t e dc i l i o pa t hy .Huma ng e ne t i c s mo t i l i t yi nhuma nsa ndd o g s . Na t ur eg e ne t i c s2010;43: 728. 19 )Mo c hi z ukiT,WuG,Ha y a s hiT,Xe no pho nt o sSL, 2013Apr5.Pub Me dPMI D:23559409. 28 )Shi b aD,Y o k o y a maT.Thec i l i a r yt r a ns i t i o na lz o ne a ndne phr o c y s t i ns .Di f f e r e nt i a t i o n;r e s e a r c hi nb i o l o - 繊毛による左右決定機構 379 g i c a ld i v e r s i t y2012Fe b ;83:S916.Pub Me dPMI D: f l ui df l o w:b e nd i ngo fpr i ma r yc i l i aa ndCa 2 +i nf l ux . 22169048. Ce l ls t r uc t ur ea ndf unc t i o n2005;30:93100.Pub Me d 29 )Shi b aD,Ta ka ma t s uT,Y o k o y a maT.Pr i ma r yc i l i ao f PMI D:16474191. i nv / i nvmo us er e na le pi t he l i a lc e l l ss e ns ephy s i o l o g i c a l 著者プロフィール 小林 大介 Da i s uk eKo b a y a s hi 所属・職:京都府立医科大学大学院医学研究科生体機能形態科学・講師(学内) 略 歴:昭和4 2 年 新潟県生まれ 平成 3年 京都大学農学研究科農芸化学科卒業 平成 5年 京都大学大学院農学研究科農芸化学専攻修了 平成 8年 総合研究大学院大学生命科学研究科分子生物機構論専攻終了 学位(博士(理学) )取得 遺伝研,東大などを経て平成 19年 10月より現職 現在の興味:繊毛並びに左右軸形成 趣 味:狂言(大蔵流) ,写真
© Copyright 2026 Paperzz