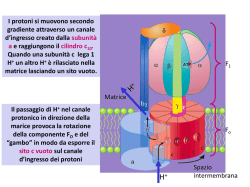
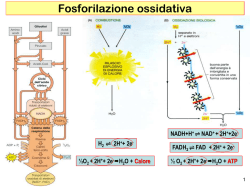
Il metabolismo cellulare 1 Il metabolismo cellulare CATABOLISMO ANABOLISMO NAD FAD NADP (per produrre energia) Nutriliti Biopolimeri (x es. proteine) ATP Intermedi di biosintesi (x es. aminoacidi) ADP Pool di precursori intracellulari energia Intermedi di sintesi Prodotti finali riduzione CALORE (sottrazione di e- come H) ossidazione e-+H+ Processo endoergonico (trasferimento di e- come H) Processo esoergonico Nutrienti dall’esterno (CO2, H2O, molecole organiche più ossidate di quelle di partenza) 2 RUOLO DELL’ ATP NEL METABOLISMO È un derivato dell’AMP (adenosin monofosfato) al quale sono legati altri 2 gruppi fosforici per mezzo di legami anidride ( ADP + Pi + energia = ATP ) fosfato inorganico I legami anidride sono legami ricchi di energia e quindi sono particolarmente reattivi - L’ATP è in grado di cedere gruppi fosforici a numerosi intermedi metabolici, convertendoli nella forma attivata con livelli di energia libera tali da consentire a questi intermedi fosforilati di partecipare alle reazioni biosintetiche, che sono termodinamicamente favorevoli (∆G°<0) - Una analoga reazione con un reagente in forma non fosforilata sarebbe termodinamicamente sfavorevole (∆G°>0) La generazione di ATP è quindi necessaria per il funzionamento dei processi biosintetici 3 Come ottengono ATP i microrganismi? Lettera O, non numero “zero”! RESPIRAZIONE: sintesi di ATP attraverso il sistema enzimatico della Fo-F1 ATP sintasi (o Fo-F1 ATPasi), al termine della catena di trasporto degli elettroni FOTOFOSFORILAZIONE: sintesi di ATP attraverso l’impiego di energia luminosa per creare una separazione di cariche FOSFORILAZIONE A LIVELLO DI SUBSTRATO: consiste nel diretto trasferimento di un fosfato ad alto contenuto energetico da un composto fosforilato ad ADP per formare ATP 4 REAZIONI di OSSIDO-RIDUZIONE - donatore di e- (agente riducente… che si ossida!) - accettore di e- (agente ossidante… che si riduce!) il valore del potenziale di RIDUZIONE è un indice di quanto la molecola in oggetto sia un buon donatore o un buon accettore di e2H+/H2 O2/H2O - 0.42 V (infatti H2 è un ottimo donatore di elettroni) + 0.82 V (infatti O2 è un ottimo accettore di elettroni) per OGNI composto che si OSSIDA in una semireazione DEVE esserci un composto che si RIDUCE in un’altra semireazione ad essa accoppiata 2H+ + 2 e- (semireazione 1) H2O (semireazione 1) H2 ½ O2 + 2H+ + 2 eREAZIONE NETTA H2 + ½ O2 H2O 5 REAZIONI di OSSIDO-RIDUZIONE donatore di e- (agente riducente… che si ossida!) - accettore di e (agente ossidante… che si riduce!) - il valore del potenziale di RIDUZIONE è un indice di quanto la molecola in oggetto sia un buon donatore o un buon accettore di e2H+/H2 O2/H2O - 0.42 V (infatti H2 è un ottimo donatore di elettroni) + 0.82 V (infatti O2 è un ottimo accettore di elettroni) per OGNI composto che si OSSIDA in una semireazione DEVE esserci un composto che si RIDUCE in un’altra semireazione ad essa accoppiata H2 ½ O2 + 2H+ + 2 e- 2H+ + 2 e- (semireazione 1) H2O (semireazione 1) REAZIONE NETTA H2 + ½ O2 H2O 6 Nei sistemi biologici le reazioni di ossido-riduzione UTILI ai fini energetici avvengono quando l’ e- e il protone (H+) che lo segue sono rimossi da un substrato (per azione di un enzima) per essere trasferiti ad un TRASPORTATORE che agirà come intermedio di trasporto tra un donatore e un accettore sempre associati a protoni!!! Gli e- non esistono allo stato libero nei sistemi biologici MA SI SPOSTANO DA DONATORI AD ACCETTORI ATTRAVERSO LA MEDIAZIONE DI TRASPORTATORI FISSI LIBERI Associati alla membrana NAD+ reazioni CATABOLICHE NADP+ reazioni ANABOLICHE cofattori di enzimi responsabili delle deidrogenazioni per esempio lattato deidrogenasi riduzione del NAD + NADH + H+ NAD+ + ossidazione del NAD Lattato Piruvato 7 NICOTINAMIDE ADENIN DINUCLEOTIDE FORMA OSSIDATA (NAD+) NICOTINAMIDE ADENIN DINUCLEOTIDE FORMA RIDOTTA (NADH + H+) NICOTINAMIDE NICOTINAMIDE ADENINA RIBOSIO 2 FOSFATI RIBOSIO ADENINA RIBOSIO 2 FOSFATI RIBOSIO Quando il NAD in forma ossidata (NAD+) acquista 2 elettroni, cioè si riduce, acquista anche un H+ e un H+ viene rilasciato nel citoplasma… per questo il NAD in forma ridotta deve essere indicato come NADH + H+ 8 Che bisogno c’è, da un punto di vista metabolico, di due piridinnucleotidi (NAD e NADP) così simili a livello funzionale? RISPOSTA: i piridinnucleotidi ossidati entrano come reagenti nei processi catabolici, mentre quelli in forma ridotta partecipano ai processi biosintetici Ciò significa che: - i piridinnucleotidi si devono trovare per lo più in forma ossidata perché i processi catabolici possano funzionare - per le reazioni biosintetiche si devono trovare in gran parte in forma ridotta SONO QUINDI NECESSARI DUE TIPI DI piridin-nucleotidi Il NAD è mantenuto largamente nello stato ossidato → interviene quindi nelle reazioni cataboliche (è importante ricordarlo per quando parleremo della fermentazione) Il NADP è mantenuto largamente nello stato ridotto → interviene quindi nelle reazioni anaboliche 9 I meccanismi di produzione di energia A - RESPIRAZIONE Completa demolizione del substrato (fonte di energia) in CO2 Permette il massimo recupero di energia dal substrato ridotto Respirazione AEROBIA: l’accettore finale di elettroni è l’O2 Respirazione ANAEROBIA: l’accettore finale di elettroni è un composto inorganico diverso dall’O2 come per esempio NO2-, CO3--, SO4-- B - FERMENTAZIONE Non comporta la completa demolizione del substrato (fonte si energia) Non prevede il coinvolgimento dell’O2 È veloce ma energeticamente meno efficiente: minore ATP prodotto rispetto alla respirazione L’accettore finale di elettroni è una molecola organica (per es. l’acetaldeide: CH3-COH) Ora ci occuperemo più nel dettaglio del metabolismo energetico di specifici batteri CHEMIO-ORGANOETEROTROFI che respirano o fermentano C - FOTOSINTESI D - LITOTROFIA: una molecola INORGANICA è la fonte di energia (di elettroni) Per es. METANOGENESI: un antico tipo di metabolismo di alcuni archeobatteri che utilizza H2 come fonte di energia e produce METANO 10 Il metabolismo energetico (catabolismo) degli eterotrofi Il destino del glucosio in una cellula batterica GLUCOSIO pathway dei pentoso-fosfati glicolisi pathway di Entner-Doudoroff 2 (via di Embden-Meyerhof-Parnas) 1 via della fruttosio-6fosfato fosfochetolasi (F6PPK) 4 3 PIRUVATO respirazione aerobica A respirazione anaerobica B fermentazione lattica, alcolica acido mista, eterolattica metabolismo dei bifidobatteri 1 2 Le tre vie attraverso cui i batteri ottengono energia dalla degradazione di composti organici 11 Glicolisi e Via dei pentoso-fosfati 12 2 Via dei pentoso-fosfati (o degli esoso-monofosfati) La prima parte della via, definita fase ossidativa, vede una serie di reazioni che ossidano il glucosio-6fosfato a ribosio-5-fosfato, uno zucchero basilare per la sintesi dei nucleotidi. Nella seconda fase, definita non ossidativa, avviene l'epimerizzazione e l'isomerizzazione del ribosio-5-fosfato che produce, alla fine, fruttosio-6-fosfato e gliceraldeide-3-fosfato. Il fruttosio-6-fosfato è facilmente interconvertito in glucosio-6-fosfato attraverso l'enzima fosfofruttosio isomerasi e il ciclo può continuare FASE OSSIDATIVA ribosio-5-P xilulosio-5-P 6-P-gluconato-δ-lattone glucosio-6-P 6-P-gluconato ribulosio-5-P FASE NON OSSIDATIVA gliceraldeide-3P xilulosio-5-P eritrosio-4-P xilulosio-5-P sedoeptulosio-7-P ribosio-5-P gliceraldeide-3P fruttosio-6-P La via dei pentoso fosfati ha due scopi. Nella parte ossidativa serve a sintetizzare il ribosio-5fosfato, precursore dei nucleotidi, mentre nella parte non ossidativa avvengono delle trasformazioni che rimettono in ciclo il fruttosio-6-fosfato, convertito facilmente in glucosio-6fosfato. Questa seconda parte avviene perlopiù quando la cellula ha più necessità di potere riducente sotto forma di NADPH rispetto alla presenza di zuccheri o ribosio-6-fosfato 13 3 Via di Entner/Doudoroff glucosio Via alternativa alla glicolisi tipica dei batteri. Tipica di alcune specie appartenenti al genere Pseudomonas, che sono prive dell’enzima FOSFOFRUTTOCHINASI batteri Gram-negativi esochinasi È una via tipica anche di Zymomonas (yeast like bacterium), che produce ETANOLO a partire dall’acido piruvico glucosio-6-P della classe dei gamma-Proteobatteri (divisione Proteobatteri) glu-6-P deidrogenasi etanolo NAD+ gliceraldeide3-fosfato NADH+H+ 6-P-gluconato-δ-lattone acetaldeide piruvato lattonasi deidratasi 6-fosfogluconato chetodeossifosfogluconato aldolasi 2-cheto-3-deossi-6-fosfo gluconato piruvato 14 DISTRIBUZIONE DELLE DIVERSE VIE DI UTILIZZAZIONE DEGLI ZUCCHERI TRA ALCUNI BATTERI Microrganismo EmbdenMeyerhof Arthrobacter spp. Azotobacter chroococcum Alcaligenes eutrophus Bacillus spp. Escherichia coli e altri batteri enterici Pseudomonas spp. Rhizobium spp. Thiobacillus spp. Xanthomonas spp. + + + + - EntnerDoudoroff glicolisi + + + + + - La via di Entner-Doudoroff è di grandissima utilità per quei microrganismi che impiegano il gluconato o il mannonato come substrato - Alcuni microrganismi come Escherichia coli impiegano la glicolisi per il metabolismo del glucosio, ma quando il gluconato viene fornito come substrato, allora vengono indotti gli enzimi chiave della Entner-Doudoroff - A prescindere dal fatto che metabolizzi il glucosio attraverso la via Embden- Meyerhof o la via Entner-Doudoroff, un batterio possiede tutti o molti degli enzimi della via dei pentosofosfati (shunt dell’esoso monofosfato). Questa apparente ridondanza metabolica può trovare una spiegazione nelle funzioni vitali della via dei pentosofosfati, relative alla generazione di NADPH e di altri due precursori metabolici, il ribosio-5-fosfato e l’eritrosio-4-fosfato 15 A La respirazione glucosio piruvato Ciclo di Krebs ATP sintasi (FoF1 ATPasi) 16
© Copyright 2026 Paperzz