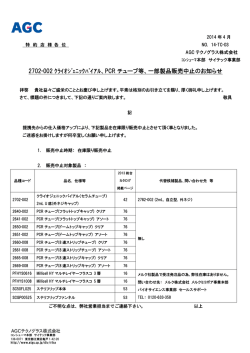
ダイズ共存細菌の群集構造解析における LNA-PCR クランプ技術 ダイズ共存細菌の群集構造解析における LNA-PCR クランプ技術 ○池永 誠・田淵雅和1)・日髙栄作・境 雅夫 1) 1) ○池永誠・田淵雅和 (鹿児島大農・ ・日髙栄作・境雅夫 鹿児島大院農) (鹿児島大農・1)鹿児島大院農) 【目的】 化した。ミトコンドリアに特異的な LNA オリゴととも 植物に共存する細菌が, 植物の生育に対して重要な 【目的】 植物に共存する細菌が, 植物の生育に対して に, 設計した LNA オリゴを加えて を行い, 効果的 LNA オリゴとともに, 設計した LNAPCR オリゴを加えて PCR 影響を及ぼしている事は古くから知られており, 植物 重要な影響を及ぼしている事は古くから知られており, な LNA オリゴの濃度を検討した。次に, DGGE 法で LNA を行い, 効果的な LNA オリゴの濃度を検討した。次に, 共存細菌の機能を微生物資材として農業利用するため 植物共存細菌の機能を微生物資材として農業利用する オリゴの有無およびダイズ各部位における群集構造の DGGE 法で LNA オリゴの有無およびダイズ各部位におけ には, 群集構造解析法の確立が不可欠である。 しかし, ためには, 群集構造解析法の確立が不可欠である。し 異同を解析した。 る群集構造の異同を解析した。 植物試料から抽出した DNA を用いて細菌用プライマー かし, 植物試料から抽出した DNA を用いて細菌用プラ 【結果および考察】 で PCR 増幅を行った場合, 植物オルガネラ(ミトコン イマーで PCR 増幅を行った場合, 植物オルガネラ(ミ オルガネラのSSUオルガネラのSSU rRNA遺伝子のPCR増幅を抑制するた 【結果および考察】 rRNA遺伝子のPCR増 ドリアおよびプラスチド) の SSU rRNA 遺伝子が過剰に トコンドリアおよびプラスチド) の SSU rRNA 遺伝子が めの効果的な濃度を検討したところ, 種子と葉につい 幅を抑制するための効果的な濃度を検討したところ, 増幅され, 共存細菌の群集構造が過小評価される重大 過剰に増幅され, 共存細菌の群集構造が過小評価され ては1.0µM以上, 茎については3.0µM以上, 根について 種子と葉については1.0µM以上, 茎については3.0µM以 な問題が存在する。以前の支部会では, 植物オルガネ る重大な問題が存在する。以前の支部会では, 植物オ は4.0µMで, オルガネラ遺伝子の増幅を抑制し, 共存細 上, 根については4.0µMで, オルガネラ遺伝子の増幅を ラのSSU rRNA遺伝子に特異的な配列を持つロックド核 ルガネラのSSU rRNA遺伝子に特異的な配列を持つロッ 菌の遺伝子を選択的にPCR増幅する事ができた。次に, 抑制し, 共存細菌の遺伝子を選択的にPCR増幅する事が 酸含有オリゴヌクレオチド(LNA オリゴ)をプライマ クド核酸含有オリゴヌクレオチド(LNA オリゴ)をプ DGGE法を用いてダイズ各部位に生息する共存細菌の群 できた。次に, DGGE法を用いてダイズ各部位に生息する ーと競合する位置に設計し, オルガネラ遺伝子の PCR ライマーと競合する位置に設計し, オルガネラ遺伝子 集構造解析を行った。その結果, LNAオリゴ使用しない 共存細菌の群集構造解析を行った。その結果, LNAオリ 増幅を抑制する事によって, 植物共存細菌の SSU rRNA の PCR 増幅を抑制する事によって, 植物共存細菌の 従来のDGGE法では検出されなかった新たなバンドが, ゴ使用しない従来のDGGE法では検出されなかった新た 遺伝子を選択的に増幅する PCR クランプ技術について SSU rRNA 遺伝子を選択的に増幅する PCR クランプ技術 LNAオリゴを使用する事で複数検出され, 新たに設計し なバンドが, LNAオリゴを使用する事で複数検出され, 報告した。しかしながら, 設計した設計した LNA オリゴにおい について報告した。しかしながら, LNA オリ たLNAオリゴの有効性が示された(図1)。また, 各部位に 新たに設計したLNAオリゴの有効性が示された(図1)。ま て, プラスチドに特異的な配列は 2 つに大別されるた ゴにおいて, プラスチドに特異的な配列は 2 つに大別 おける群集構造を比較したところ, 共通するバンドが た, 各部位における群集構造を比較したところ, 共通 め, ダイズなどの共存細菌の群集構造を解析する際は, されるため, ダイズなどの共存細菌の群集構造を解析 多かったものの, 各部位に特異的なバンドも認められ, するバンドが多かったものの, 各部位に特異的なバン 新たな LNA オリゴの設計が必要となる。今回は新たに する際は, 新たな LNA オリゴの設計が必要となる。今 これらのバンドに由来する共存細菌が, 植物の生長に ドも認められ,これらのバンドに由来する共存細菌が, プラスチドに特異的な LNA オリゴを設計したので, そ 回は新たにプラスチドに特異的な LNA オリゴを設計し 関与している可能性が示唆された。 植物の生長に関与している可能性が示唆された。 れを使用して共存細菌の遺伝子を選択的に PCR 増幅し, たので, それを使用して共存細菌の遺伝子を選択的に 群集構造した時の結果について報告する。 PCR 増幅し, 群集構造した時の結果について報告する。 【材料および方法】 供試試料にはダイズ(フクユタカ)を用いた。芽出 【材料および方法】 供試試料にはダイズ(フクユタカ) し後, 約 3 週間ポット栽培を野外で行い, 約 20 本の幼 を用いた。芽出し後, 約 3 週間ポット栽培を野外で行 個体から葉, 茎および根を採取した。ダイズ種子も供 い, 約 20 本の幼個体から葉, 茎および根を採取した。 試試料として用いた。種子, 葉, 茎および根の表面を ダイズ種子も供試試料として用いた。種子, 葉, 茎お 洗浄後, 磨り潰してから磨り潰してから DNA を抽出した。プラスチド よび根の表面を洗浄後, DNA を抽出し に特異的なLNAオリゴは, た。プラスチドに特異的なプラスチドおよび細菌のSSU LNA オリゴは, プラスチド rRNA 遺伝子を, および細菌の SSUプライマーとともにアライメントし, rRNA 遺伝子を, プライマーとともに プライマーのアニーリング部位と数塩基競合させる一 アライメントし、プライマーのアニーリング部位と数 方, 3′末端の伸長側にシフトした位置においてプラス 塩基競合させる一方, 3′末端の伸長側にシフトした位 チドに特異的な DNA を LNA に置換して設計した。また, 置においてプラスチドに特異的な DNA を LNA に置換し LNA オリゴから伸長が生じないように 3′末端をリン酸 て設計した。また, LNA オリゴから伸長が生じないよう 図 1.ダイズ各部位における共存細菌群の DGGE パター に 3′末端をリン酸化した。ミトコンドリアに特異的な ン. M;オルガネラマーカー, -;LNA 無, +;LNA 有 ─ 57 ─ p053-064cs6.indd 57 2014/07/31 8:58:28
© Copyright 2026 Paperzz