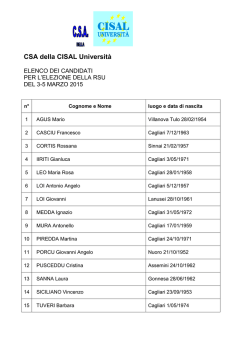
Cenni su…
Interazione luce-materia e
Spettroscopia atomica e molecolare
Corso di Biofisica, Università di Cagliari
1
Riferimenti
• D. McQuarrie, “Physical Chemistry: A molecular approach”,
chapters 13, 15.
• http://blogs.jccc.edu/astronomy/textbook/unit-two-conceptualand-observational-tools-of-astronomy/chapter-5-electromagneticradiation-and-matter/
• http://chemwiki.ucdavis.edu/Physical_Chemistry/Spectroscopy/
• Movies
• http://chemwiki.ucdavis.edu/Physical_Chemistry/Spectroscopy/
Electronic_Spectroscopy/Fluorescence/Jablonski_Diagram
%3a_Shockwave
• https://www.youtube.com/watch?v=O39avevqndU
• Exercise
• http://home.sou.edu/~chapman/ch445/Iodine.htm
Corso di Biofisica, Università di Cagliari
2
Cenni storici
• 1665 Newton disperde la luce mediante un prisma
• 1814 Fraunhofer: spettro a righe della radiazione solare
Corso di Biofisica, Università di Cagliari
3
Leggi di Kirchoff
• Prima legge: un solido, un liquido
o un gas sufficientemente denso
emettono luce a tutte le lunghezze
d’onda e producono uno spettro
continuo.
• Seconda legge: un gas caldo a
bassa densità emette luce il cui
spettro consiste di una serie di
linee di emissione che sono
caratteristiche della composizione
chimica del gas.
• Terza legge: un gas freddo
assorbe certe lunghezze d’onda
da uno spettro continuo, lasciando
linee di assorbimento scure
sovrapposte allo spettro continuo
(principio di reversibilità
Kirchoff-Bunsen).
Corso di Biofisica, Università di Cagliari
4
Radiazione elettromagnetica
• Può essere interpretata come fenomeno ondulatorio...
• …e come fenomeno quantistico (fotoni, legge di Planck)
E = hν = hc
λ
h = 6.62 ⋅10 −34 Js
Corso di Biofisica, Università di Cagliari
5
Spettro elettromagnetico
Insieme delle radiazioni elettromagnetiche è lo
spettro elettromagnetico: da onde radio a raggi γ (E crescenti).
Corso di Biofisica, Università di Cagliari
6
Interazione luce-materia
• La spettroscopia o spettrofotometria studia l’interazione
fra luce e materia (atomi, molecole, soluzioni, solidi).
• L’assorbimento di energia da parte della materia può
provocare delle variazioni chimiche o fisiche (proprietà
degli elettroni o del nucleo) della specie chimica.
• Il confronto con dati sperimentali noti o con simulazioni
teoriche permette di dedurre le proprietà della materia.
• Permette di caratterizzare vari parametri, tra cui le
strutture (elettroniche e atomiche) molecolari.
Corso di Biofisica, Università di Cagliari
7
Alcuni tipi di spettrometria
•
•
•
•
•
•
•
•
•
•
•
Assorbimento o Emissione Atomiche (AAS, AES)
Molecolare Ultravioletto-Visibile (UV-VIS)
Molecolare Infrarosso (IR)
Risonanza Magnetica Nucleare (NMR, EPR)
Spettrometria di massa
Rifrattometria
Polarimetria
Dispersione ottica rotatoria
Diffrazione dei raggi X
Nefelometria e Turbidimetria
Spettroscopia di Fluorescenza (FS, atomica e molecolare)
Corso di Biofisica, Università di Cagliari
8
Nel dettaglio…
λ (nm)
106
-
1010
103 - 105
3102
-
8102
ν (Hz)
107
-
1011
1011 - 1014
31014
-
7.81014
Regione
spettro
Interazioni
(spettroscopia)
Radio,
Spin nucleare (NMR)
microonde e elettronico (EPR)
IR
Visibile
(VIS)
Vibrazioni, rotazioni
2102 - 3102
1014 - 1016
UV
Transizioni
elettroniche
(spettroscopia ottica)
100 - 102
1017 - 1020
X
Gusci interni
Corso di Biofisica, Università di Cagliari
9
Cosa succede?
Un pò di fenomenologia delle
soluzioni molecolari
Corso di Biofisica, Università di Cagliari
10
Interazione-luce materia
luce incidente
luce riflessa
luce rifratta
luce difratta
luce assorbita
Oggetto
(es. bicchiere d’acqua
con pigmento)
luce trasmessa
Corso di Biofisica, Università di Cagliari
Assorbimento (empirico, UV-VIS)
I0
campione
I
• Ogni sostanza può assorbire una diversa quantità di
radiazione.
• Tale quantità è diversa per le varie lunghezze d’onda
(colori).
Corso di Biofisica, Università di Cagliari
Legge di Lambert-Beer
• Si definisce trasmittanza T la frazione (intensità) di luce
trasmessa, e assorbanza A il log dell’inverso di T:
I sample
I
T= ≈
I0
I blank
I0
I blank
A = −logT = log ≈ log
I
I sample
• A direttamente proporzionale alla concentrazione del soluto
secondo la legge di Lambert-Beer:
I0
A = log = ε cl
I
•
•
•
ε coefficiente di estinzione molare
c concentrazione molare
l cammino ottico
• A di soluzioni è additiva:
Amixture = A1 + A2 + ...+ An
Corso di Biofisica, Università di Cagliari
Legge di Lambert-Beer
• In realtà A (ε) dipende dalla lunghezza d’onda:
Aλ = ε λ cl
• Inoltre, ε dipende anche dal pH e dalla natura del solvente,
mentre è indipendente dalla temperatura.
L-B
Real
Legge di Lamber-Beer vale
solo per soluzioni molto diluite
(trascura interazioni
molecolari).
Corso di Biofisica, Università di Cagliari
Coefficienti di estinzione di alcuni gruppi
Corso di Biofisica, Università di Cagliari
Esempio di assorbimento molecolare
Ripetendo l’esperimento a varie λ lo spettro di assorbimento permette
di risalire alla natura del campione (per confronto diretto con campioni
noti o tramite banche dati di spettri) o di controllarne il grado di
purezza.
Esempio di spettro UV-visibile di un’aldeide insatura. La
banda a 395 nm rende conto del fatto che il composto è
colorato in arancio, complementare al violetto che
corrisponde alla regione spettrale interessata (~ 400 nm)
Corso di Biofisica, Università di Cagliari
Relazioni colori assorbiti/osservati
Lunghezza d’onda
380-420
420-440
440-470
470-500
500-520
520-550
550-580
580-620
620-680
680-780
Assorbita
violetto
blu-viola
blu
blu-verde
verde
verde-giallo
giallo
arancione
rosso
viola
Osservata
verde-gialla
giallo
arancione
rosso
viola
violetto
blu
verde-blu
blu-verde
verde
Corso di Biofisica, Università di Cagliari
Altri esempi
• Assorbimento da parte di gruppi funzionali e molecole organiche.
• L’intensità dipende dal tipo di transizione elettronica (ππ* >> nπ*).
Corso di Biofisica, Università di Cagliari
Cenni di spettroscopia atomica
Corso di Biofisica, Università di Cagliari
19
Schema generale
Atomi ISOLATI
in fase gassosa
!
Atomizzazione
Campione
Interazione
radiazione-materia
Radiazione
Corso di Biofisica, Università di Cagliari
20
Spettroscopia Atomica
Tramite assorbimento (AAS), emissione (AES) o fluorescenza
(AFS) di radiazione UV-VIS.
Atomizzazione del campione con tecniche termiche/fisiche/chimiche.
Corso di Biofisica, Università di Cagliari
21
Spettroscopia Atomica
• Emissione: atomi portati ad uno stato
eccitato tramite energia termica
emettono luce tornando allo stato
fondamentale. Si misura l’intensità
della radiazione emessa (sorgente =
campione).
• Assorbimento: atomi assorbono luce
da una sorgente a determinate
frequenze, e la luce non assorbita
raggiunge il rivelatore.
• Fluorescenza: atomi eccitati da una
sorgente di luce; decadendo allo
stato fondamentale emettono luce di
eguale o maggiore λ. Si chiama
chemiluminescenza se causata da
reazione chimica.
Corso di Biofisica, Università di Cagliari
AAS e AES
Spettri di righe (no componenti roto vibrazionali)
corrispondenti a transizioni elettroniche fra stati quantistici.
Corso di Biofisica, Università di Cagliari
23
AAS e AES
!
NO SOVRAPPOSIZIONI tra spettri atomici di elementi diversi
Corso di Biofisica, Università di Cagliari
24
Spettroscopia Atomica
• Permette determinazione qualitativa e quantitativa
degli analiti presenti nel campione.
• Selettiva perché righe di assorbimento molto strette e
energie di transizione elettroniche tipiche di ciascun
elemento.
Infatti…
• Ogni elemento assorbe (emette) solo a λ corrispondenti a
promuovere una transizione tra stati elettronici.
• Numero di transizioni caratteristico di ciascun elemento.
Corso di Biofisica, Università di Cagliari
25
Spettri atomici vs. molecolari
Spettro di assorbimento di atomi di
silicio nell-intervallo 250-253 nm.
Spettro di assorbimento del permanganato di
potassio (KMnO4) nell’intervallo 450-650 nm.
Corso di Biofisica, Università di Cagliari
26
Spettri atomici vs. molecolari
Spettro di assorbimento di atomi di
silicio nell-intervallo 250-253 nm.
Spettro di assorbimento del permanganato di
potassio (KMnO4) nell’intervallo 450-650 nm.
Corso di Biofisica, Università di Cagliari
27
Interazione luce-materia:
Interpretazione molecolare
Corso di Biofisica, Università di Cagliari
28
Livelli e transizioni molecolari
• Anche la più semplice molecola contiene almeno tre set di livelli
energetici:
– rotazionale (microonde, lontano IR)
– vibrazionale (IR)
– elettronico (UV-VIS)
• ΔErot << ΔEvib << ΔEel
• Un fascio di fotoni (radiazione) che colpisce la molecola può eccitare
uno di questi gradi di libertà, promuovendo il passaggio ad uno
stato a energia superiore, a patto che:
hν
E1
E0
molecola
E1
E0
hν = E1 − E0 = ΔE
molecola*
Corso di Biofisica, Università di Cagliari
29
Livelli e transizioni molecolari
Una volta assorbita, l’energia può essere rilasciata sotto forma di
radiazione o tramite processi non radiativi quando la molecola rilassa
nello stato inferiore:
M + hv → M
∗
∗
M → M + hv '+ heat
Emissione di radiazione:
(foto)luminescenza, es.
fluorescenza, fosforescenza Processi non radiativi:
conversioni interne, inter-sistema, trasferimento energia elettronica,
vibrazionale, rotazionale e traslazionale delle molecole circostanti Corso di Biofisica, Università di Cagliari
30
Livelli vibrazionali (mol. biatomica)
Descritti in prima approssimazione tramite oscillatore armonico.
Livelli energetici equispaziati
Gn = En
hc
= ( n + 12 ) ν
12
1 !k$
ν
ν =
=
c 2π c #" µ &%
n = 0,1, 2,...
Regole di selezione: Δn = ±1
Corso di Biofisica, Università di Cagliari
31
Livelli vibrazionali
Nella realtà l’energia potenziale è più simile a curva di Morse
Teoria perturbativa,
termine anarmonico.
2
Gn = ( n + 12 ) ν e − x eν e ( n + 12 ) +...
x e costante anarmonicità e
equilibrio
Gap decresce all’aumentare di n
Valori tipici
ν e ≈ 10 2 −10 3 cm −1
x e ≈ 1−10 cm −1
Regole di selezione: Δn = ±1,2,3,…
Corso di Biofisica, Università di Cagliari
32
Livelli rotazionali (mol. biatomica)
Descritti in prima approssimazione tramite rotore rigido
FJ = EJ
B =
hc
= J ( J +1) B
h
2
, I
=
µ
R
e
8π 2 cI
J = 0,1, 2,...
gJ = 2J +1
Regole di selezione:
• µdipole permanente
• ΔJ = ±1
F0→1 = F1 − F0 = 2 B
F1→2 = F2 − F1 = 4 B
F2→3 = F3 − F2 = 6 B
...
FJ→J+1 = FJ+1 − FJ = 2(J +1)B
Linee spettrali appaiono
per multipli di 2B̃
Corso di Biofisica, Università di Cagliari
33
Livelli rotazionali vs. vibrazionali
B ≈ 10 −2 −10 2 cm −1
n=3 v ≈ 10 2 −10 3 cm −1
Ogni livello vibrazionale
contiene 100/1000
livelli rotazionali!
n=2 n=1 n=0 Corso di Biofisica, Università di Cagliari
34
Livelli rotazionali
Correzioni centrifuga, isotopica, e vibro-rotazionale
All’aumentare di J la forza centrifuga
distende leggermente il legame
2
FJ = J ( J +1) B − J 2 ( J +1) D
J = 0,1, 2,...
Costante di distorsione centrifuga
Corso di Biofisica, Università di Cagliari
35
Livelli rotazionali
Correzioni centrifuga, isotopica, e vibro-rotazionale
All’aumentare di J la forza centrifuga
distende leggermente il legame
2
FJ = J ( J +1) B − J 2 ( J +1) D
J = 0,1, 2,...
Costante di distorsione centrifuga
All’aumentare di µ aumenta il
momento di inerzia
1
B ∝
I
12
C 16 O
13
C 16 O
2 B = 3.842 cm −1
2 B = 3.673 cm −1
12
C 18 O
2 B = 3.659 cm −1
Corso di Biofisica, Università di Cagliari
36
Livelli rotazionali
Correzioni centrifuga, isotopica, e vibro-rotazionale
All’aumentare di J la forza centrifuga
distende leggermente il legame
2
FJ = J ( J +1) B − J 2 ( J +1) D
J = 0,1, 2,...
All’aumentare di n aumenta Re e
quindi diminuisce B̃ (~Re2)
B n = B e − α e ( n +1)
n = 0,1, 2,...
Costante di distorsione centrifuga
All’aumentare di µ aumenta il
momento di inerzia
13
C 16 O
2 B = 3.842 cm −1
2 B = 3.673 cm −1
12
C 18 O
2 B = 3.659 cm −1
12
1
B ∝
I
C 16 O
B e ≈ 10 −2 −10 2 cm −1
D ≈ 10 −8 −10 −2 cm −1
α e ≈ 10 −4 −10 cm −1
Corso di Biofisica, Università di Cagliari
37
Livelli rotazionali
Correzioni centrifuga, isotopica, e vibro-rotazionale
All’aumentare di J la forza centrifuga
distende leggermente il legame
All’aumentare di n aumenta Re e
quindi diminuisce B̃ (~Re2)
= B − α ( n +1)
B
e energetici
e
fra i nlivelli
2
FJ Tutti
= J ( Jgli
+1)effetti
B − J 2 ( Jriducono
+1) D
il gap
J = 0,1, 2,...all’aumentare di J, n e µ.n = 0,1, 2,...
Intensità
ordini
di grandezza inferiori rispetto a
Costante di alcuni
distorsione
centrifuga
termini principali.
−2
2
−1
All’aumentare di µ aumenta il
momento di inerzia
2 B = 3.842 cm −1
13
C 16 O
2 B = 3.673 cm −1
2B̃ 4B̃
6B̃
12
C 18 O
2 B = 3.659 cm −1
12
1
B ∝
I
Be ≈ 10 −10 cm
D ≈ 10 −8 −10 −2 cm −1
C 16 O
α e ≈ 10 −4 −10 cm −1
8B̃
10B̃
12B̃
Corso di Biofisica, Università di Cagliari
38
Livelli rotazionali: rami P e R
Regole di selezione rotore rigido – oscillatore armonico (assorbimento):
Δn = +1, ΔJ = ±1
Ramo P
Ramo R
B1 < B 0 !!!
ν 0→0
ν 0→1 ( ΔJ = −1) = ν 0 − ( B 0 + B1 ) J + ( B1 − B 0 ) J 2
ν 0→1 ( ΔJ = +1) = ν 0 + 2 B1 + (3B1 − B 0 ) J + ( B1 − B 0 ) J 2
Corso di Biofisica, Università di Cagliari
39
Regola di selezione per il dipolo
Interpretazione classica
Perché il campo elettrico possa esercitare un momento torcente
sulla molecola occorre che essa sia dotata di momento di dipolo
elettrico (molecola polare).
Corso di Biofisica, Università di Cagliari
40
Regole di selezione: diagramma di Grotian
Corso di Biofisica, Università di Cagliari
41
Livelli vibrazionali molecole poliatomiche
• In assenza di campi di forze esterni, differenza di potenziale
rispetto alle posizioni di equilibrio è:
N
N
1 vib vib ∂2V
ΔV = V ( q1,..., qNvib ) −V ( 0,..., 0 ) = ∑ ∑
qi q j +...
2 i=1 j=1 ∂qi∂q j
• Diagonale in funzione dei modi normali {Qi},i=1,…,Nvib.
N
N vib
N
N
1 vib ∂2V 2
2 d 2 1 vib ∂2V 2 vib & 2 d 2 1 ∂2V 2 )
ΔV = ∑ 2 Qi → H vib = −∑
+ ∑ 2 Qi =∑(
+
Q
2
2
2 i +
2 i=1 ∂Qi
2 i=1 ∂Qi
2 ∂Qi
*
i=1 2 µi dQi
i=1 ' 2 µi dQi
Insieme di Nvib oscillatori armonici indipendenti:
N vib
Ψ vib = Πψ vib,i (Qi )
i=1
N vib
Evib =
∑(n + ) hν
i
1
2
i
i=1
Corso di Biofisica, Università di Cagliari
42
Livelli di H2O e CO2
Regole di selezione: dipolo varia in assorbimento del modo
vibrazionale (modo attivo nell’infrarosso).
H2O
Stiramento simmetrico
ν1 (attivo)
3650 cm-1
Stiramento asimmetrico
ν3 (attivo)
3760 cm-1
Piegamento
ν2 (attivo)
1600 cm-1
CO2
Stiramento simmetrico
ν1 (inattivo)
Stiramento asimmetrico
ν3 (attivo)
2349 cm-1
Piegamenti (doppia deg.)
ν2 (attivi)
667 cm-1
Corso di Biofisica, Università di Cagliari
43
Livelli rotazionali molecole poliatomiche
• Dipendono dai momenti principali di inerzia della molecola:
I A ≤ I B ≤ I C.
A =
• Tre costanti rotazionali:
…e tre tipi di rotore:
• Sferico: IA = IB = IC.
h
h
h
≥B= 2
≥C = 2
2
8π cI A
8π cI B
8π cI C
FJ = J ( J +1) B
gJ = ( 2J +1)
2
• Simmetrico:
– Oblato: IA = IB < IC.
FJ = J ( J +1) B + C − B K 2
– Prolato: IA < IB = IC.
FJ
( )
= J ( J +1) B + ( A − B ) K
2
J = 0,1, 2,...
K = 0, ±1, ±2,...
gJK = 2J +1
• Asimmetrico: spettro molto complesso.
Corso di Biofisica, Università di Cagliari
44
Linee spettrali rotore
Rotore simmetrico
Rotore asimmetrico
Corso di Biofisica, Università di Cagliari
45
Impieghi spettroscopia vibro-rotazionale
• Struttura molecolare (piccole molecole).
• Massa isotopica.
• Conformazioni molecolari e rispettive popolazioni.
• Temperatura.
• Momento di dipolo molecolare.
Corso di Biofisica, Università di Cagliari
46
Impieghi spettroscopia vibro-rotazionale
Struttura molecolare
B può essere misurato con grande precisione
↓
I può essere ricavato con grande precisione
↓
Geometria Molecolare Accurata
(es. distanza di equilibrio)
B=
I
⇒I=
= µ req2 ⇒ req =
4π cI
4π cB
µ
Corso di Biofisica, Università di Cagliari
47
Impieghi spettroscopia vibro-rotazionale
Massa isotopica
Quale è la massa isotopica di 13C ?
B(12CO) = 1.921175 cm-1
B(13CO) = 1.836685 cm-1
µ(13 CO) 1.921175
1 1
=
= 1.04600
B ∝ ∝ ⇒ 12
µ( CO) 1.836685
I µ
(13 C) ×15.9994 12 + 15.9994
⇒ 1.046 = 13
×
( C) + 15.9994 12 ×15.9994
(13C) = 13.0006 amu
Corso di Biofisica, Università di Cagliari
48
Livelli elettronici
Per ogni livello elettronico, associato ad una superficie adiabatica, vi
sono diversi livelli vibrazionali (e rotazionali)
Transizioni elettroniche a energie E
molto maggiori delle vibrazionali
−
3
(Spettroscopia UV-VIS).
Σu
3
Σ
+
u
Born-Oppenheimer
Emol = ν el + Gn + FJ =
1
ν el +
Σ
+
g
1
2
(n + 12 )νe − xeνe (n + 12 ) +
J ( J +1) B − J ( J +1) D
2
2
1
Δ
Σ
−
g
O2
−
g
R
Corso di Biofisica, Università di Cagliari
49
Transizioni vibroniche
Transizioni vibrazionali in spettri elettronici
(includono anche transizioni elettroniche!)
E
Ignorando termini rotazionali, transizione
fra i livelli vibrazionali zero degli stati
fondamentale e eccitato avviene con
energia (no accoppiamento vibronico!):
n' 2’ 1’ 0’ G0'
v0",0' = ΔEel + G0' − G0"
G0i = 12 ν ei − 14 x eiν ei
n” 2” 1” 0” v0",0'
ΔEel
G0"
D0
De
R
Corso di Biofisica, Università di Cagliari
50
Transizioni vibroniche
Transizioni vibrazionali in spettri elettronici
(includono anche transizioni elettroniche!)
E
Quindi una transizione da 00” (stato
più popolato a Tamb) a 1n’ assorbirà
una energia (frequenza):
n' v
2’ 0",n'
1’ 0’ v0",n' = v0",0' + Gn' − G0' =
v0",0' + n'ν 'e − n'(n'+1) x 'e ν 'e
n” 2” 1” 0” Gn'
G0'
v0",0'
R
Corso di Biofisica, Università di Cagliari
51
Transizioni vibroniche
# 1
&
1
Δv0",n'/0",n'+1 = 2 x 'e ν 'e %
− (n'+1)( → n'max =
−1
2 x 'e
$ 2 x 'e
'
Al crescere di n’ separazione fra
i livelli vibronici converge a 0
Spettro discreto diventa
continuo, poi dissociazione (dallo
stato eccitato).
Corso di Biofisica, Università di Cagliari
52
Transizioni vibroniche
# 1
&
1
Δv0",n'/0",n'+1 = 2 x 'e ν 'e %
− (n'+1)( → n'max =
−1
2 x 'e
$ 2 x 'e
'
A seconda della T stati vibrazionali
eccitati possono essere popolati e
assorbire radiazione
spettri sovrapposti
Corso di Biofisica, Università di Cagliari
53
Parametri da spettri elettronici
Molecole biatomiche
è possibile ricavare le distanze di equilibrio e le energie di legame
Corso di Biofisica, Università di Cagliari
54
Livelli e transizioni molecolari
Richiami su stati di singoletto e tripletto
gS=2S+1=1
gS=2(1/2+1/2)+1=3
Corso di Biofisica, Università di Cagliari
55
Livelli e transizioni (molecola biatomica)
E
S1 • Assorbimento
T1 • Conversione
intersistema
• Fluorescenza
S0 • Fosforescenza
R
Corso di Biofisica, Università di Cagliari
56
Livelli e transizioni molecolari
Diagramma di Jablonski
S2 S1 T1 Q
S0 Corso di Biofisica, Università di Cagliari
57
Livelli e transizioni molecolari
Assorbimento
In genere avviene
senza cambiamenti
dello spin.
S2 S1 T1 S0 Corso di Biofisica, Università di Cagliari
58
Livelli e transizioni molecolari
S2 S1 S0 transizione non radiativa
all’interno dello stato
T1 elettronico
(in seguito a
urti con altre molecole).
Processi molto rapidi
rispetto alle transizioni
tra stati elettronici.
Corso di Biofisica, Università di Cagliari
59
Livelli e transizioni molecolari
Conversione interna
S2 S1 S0 T1 decadimento nonradiativo tra stati
vibrazionali relativi a
stati elettronici diversi
e aventi stessa
molteplicità di spin.
Meccanicisticamente
identico al
rilassamento
vibrazionale.
Corso di Biofisica, Università di Cagliari
60
Livelli e transizioni molecolari
Conversione
intersistema
S2 S1 S0 transizione non-radiativa
tra stati elettronici aventi
diversa gS, se c’è
sovrapposizione fra livelli
T1 vibrazionali dei due stati.
Non dovrebbe avvenire,
stando alle regole di
selezione elettroniche, ma
l’accoppiamento con i
livelli vibrazionali lo
rende poco probabile
(più lento della
conversione interna).
Corso di Biofisica, Università di Cagliari
61
Livelli e transizioni molecolari
Fluorescenza
S2 S1 S0 Sistema si trova nello stato
vibrazionale più basso
dello stato eccitato S1.
Poco probabile che
T1 molecole circostanti
assorbano la grossa
quantità di energia
necessaria per rilassare
elettronicamente la
molecola.
Emissione di radiazione a
λfluor ≥ λass.
Corso di Biofisica, Università di Cagliari
62
Principio di Franck-Condon
Tante transizioni possibili, quali con maggiore probabilità?
Principio di Franck-Condon
• me>>MN transizioni elettroniche
avvengono a nuclei quasi fermi.
• Transizione verticale più intensa è quella
verticale (maggiore sovrapposizione funzioni
d’onda).
Intensità
∝ fattori di
Franck-Condon
Pn"→n' = Ψ n" Ψ n'
Corso di Biofisica, Università di Cagliari
63
2
Fluorescenza
Assorbimento
Fluorescenza
λ ν’
1’
0’
ν”
1”
0”
• Scala temporale >> processi non radiativi
fluorescenza da stato vibrazionale fondamentale.
A seconda della molecola, tempi simili a
conversione intersistema (competizione).
• Più probabile dallo stato fondamentale del primo
livello eccitato (popolazione, altri processi favoriti).
• Dato alto numero di transizioni vibroniche possibili,
emissione su range di frequenze.
• Distanza fra righe assorbimento dipende da Δν’
assorbimento rileva struttura caratteristica dello
stato eccitato.
• Distanza fra righe fluorescenza dipende da Δν”
fluorescenza rileva struttura caratteristica dello
stato fondamentale. È spostata a frequenze più
basse.
• Intensità righe da principio di Franck-Condon.
Corso di Biofisica, Università di Cagliari
64
Livelli e transizioni molecolari
Fosforescenza
S2 S1 S0 Sistema si trova nello stato
vibrazionale zero di T1.
Interazione LS permette
transizione T1S0 (P << 1)
molecola irradia
T1 e la debolmente.
Dato che (regola di Hund)
ET < ES λfosf ≥ λfluor.
Diminuzione di T ostacola
processi di rilassamento
competitivi (quenching),
aumento fosforescenza.
Corso di Biofisica, Università di Cagliari
65
Livelli e transizioni molecolari
Smorzamento e
autosmorzamento
S2 S1 Q
S0 trasferimento di energia
ad altra molecola
(quenching agent Q).
Avviene se c’è
T1 sovrapposizione spettri di
fluorescenza della
molecola che emette e
assorbimento di Q. Può
essere dovuto a molecole
della stessa specie
(autosmorzamento).
Compete con
fluorescenza.
Corso di Biofisica, Università di Cagliari
66
Livelli e transizioni molecolari
Fluorescenza ritardata
S2 S1 Q
S0 Se conversione
intersistema ST è
seguita non da
T1 fosforescenza ma da
seconda conversione
TS e da fluorescenza.
Compete con
fosforescenza.
Corso di Biofisica, Università di Cagliari
67
Livelli e transizioni molecolari
S2 S1 Q
T1 Altri processi
non radiativi
S0 Corso di Biofisica, Università di Cagliari
68
Livelli e transizioni molecolari
Processo
Transizione
Scala temporale
Assorbimento
S0S1
10-15 s
Collisionale
Conversione interna
Conversione intersistema
Q
S0 Fluorescenza
S1S0
10-9-10-7 s
Fosforescenza
T1S0
10-6-102 s
T1 ILLUMINAZIONE
S1 10-11-10-14 s
S1T1/T1S0 10-12-10-6/10-8-10-3 s
INTENSITÀ DI EMISSIONE
S2 Collisionale
FOSFORESCENZA
10-6 – 102 s
FLUORESCENZA
10-10 – 10-7 s
Tempo
Corso di Biofisica, Università di Cagliari
69
Larghezza e intensità delle righe spettrali
Larghezza delle righe influenzata da:
• Indeterminazione QM (larghezza naturale)
• Urti (allargamento di Lorentz)
• Effetto Doppler
Corso di Biofisica, Università di Cagliari
70
Larghezza naturale
Principio di indeterminazione di Heisenberg
"1%
hc
ΔE Δt = hΔν Δt = hcΔ $ ' Δt = 2 Δλ Δt ≥
#λ&
λ
λ2
Δλ ≥
2 π c Δt
Per una riga a λ = 450 nm ed una vita media di Δt = 10-8 s,
la minima larghezza di riga è Δλ = 0.01 pm.
Corso di Biofisica, Università di Cagliari
71
Effetto Doppler e allargamento di Lorentz
Effetto Doppler
• Dipende dalla velocità e direzione di spostamento degli atomi
assorbenti (possono dirigersi verso o in direzione opposta rispetto al
rivelatore).
• Provoca allargamento di ~10-2 Å.
Effetto di pressione (allargamento di Lorenz)
• Urto tra particelle a temperatura elevata porta a cambiamenti dei
livelli energetici degli atomi. Allargamento varia da circa 0.01 a 1 Å.
Larghezza riga atomica (FWHM, Full Width at Half Maximum) data da:
Δλtot = Δλ Natural + ΔλDoppler + ΔλLorentz
Corso di Biofisica, Università di Cagliari
72
Intensità e temperatura
Intensità relative varie transizioni dipende principalmente da
popolazione relativa dei livelli, data da distribuzione Boltzmann:
N i gi −ΔEi 0
= e
N 0 g0
k BT
• kB = R/NA = 1.38x10-23 J/K
• gi fattore di degenerazione
T cambia il rapporto Nj/N0!!!
• Importante per spettri di emissione, assorbimento non molto influenzato
poiché N0 non cambia sensibilmente.
• Es. a 2500°C aumento di 10°C provoca incremento 3% numero di atomi
Na aventi elettrone più esterno nello stato 3p, mentre numero di atomi con
l’elettrone nello stato fondamentale (3s) diminuisce dello 0,002%.
Corso di Biofisica, Università di Cagliari
73
Intensità e temperatura
Corso di Biofisica, Università di Cagliari
74
Intensità e temperatura: rotore rigido
gi
Ni
= 2J +1 →
= ( 2J +1) e−ΔEJ 0
g0
N0
k BT
Corso di Biofisica, Università di Cagliari
75
Intensità: rotore rigido
Intensità relative varie transizioni dipendono anche dal momento
di dipolo di transizione:
µij = ∫ ψ *j µψi dτ
• Dipende dal momento di dipolo della molecola, µ.
µCO = 0.10 D
µHCN = 3.00 D
• N quasi indipendente da J.
Corso di Biofisica, Università di Cagliari
76
Spettroscopia risolta nel tempo
Corso di Biofisica, Università di Cagliari
77
© Copyright 2026 Paperzz