Paper
zz
Explore Categories
Log in
Create new account
No category
Large Subunit from Sweet Potato
Download
Report
Festive Brunch ¥4500 - Hyatt Regency Kyoto
New Year`s Brunch ¥5000
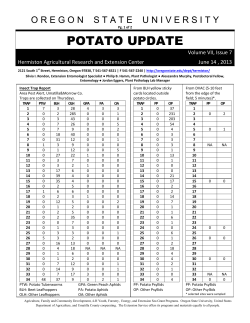
June 14
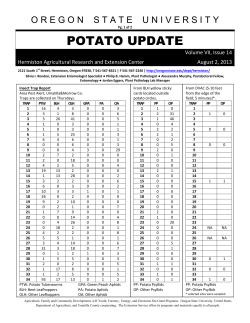
August 2
Potato Processing
2014年1月9日(木) コンチネンタルダイニング 伊藤
© Copyright 2026 Paperzz
About Paperzz
DMCA / GDPR
Report