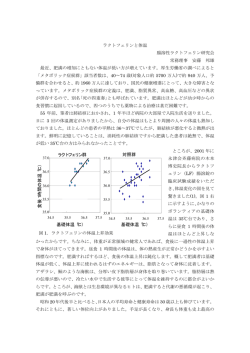
上原記念生命科学財団研究報告集, 28 (2014) 50. 哺乳類神経回路の発達と機能の遺伝学的研究 岩里 琢治 Key words:神経科学,神経回路,発達,細胞骨格, マウス 国立遺伝学研究所 形質遺伝研究部門 緒 言 我々は以前,両足をそろえて歩く突然変異マウスを偶然発見し,Rho ファミリーの主要メンバーである Rac を特異 的に不活化するタンパク質(RacGAP)である α キメリンの遺伝子が,トランスポゾン挿入により破壊されたことが, 突然変異の原因であることを明らかにした 1).また,この変異マウスでは,皮質脊髄路と脊髄中枢パターン発生器とい う二つの主要な運動系神経回路が,正中線で異常な交差をしていることを見つけた.さらに in vitro 実験を含む様々な レベルの解析を行い,以下のことを明らかにした.(1)α キメリンは,発達期の運動系神経軸索の先端(成長円錐) で膜蛋白質 EphA4 と結合している. (2)EphA4(受容体)は正中線グリアが発現する ephrin-B3(リガンド)に触れ ると,α キメリンの RacGAP 活性を介して Rac(アクチン重合を促進する蛋白質)を不活化する. (3)そして,それ により,正中線での成長円錐の前進が抑制され,不適切な軸索交差が防がれる.(4)その結果,正常な交代性歩行パ ターンが成立する 1).その後,ヒトの眼球運動の異常を引き起こす Duane's Syndrome の原因が α キメリンの過剰発 現による神経回路形成の異常であることが報告された 2).また,最近では α キメリンが大脳皮質の細胞移動に必要と いう報告もある 3).α キメリンは細胞骨格を制御する分子であり,形態変化を制御することにより脳高次機能にも重要 な役割をすることが期待されるが,α キメリンの脳高次機能における役割は未開拓のままである.本研究では,α キ メリンおよびそのイソフォームの各種ノックアウトマウスを作製し,その行動解析を行うことによって,成体での神経 回路機能における α キメリンの役割を初めて明らかにした. 方法、結果および考察 我々は以前,α キメリンの全身性ノックアウトマウスを作製し,そのマウスがウサギのように両足をそろえて歩く ことを報告した 1).本研究では,α キメリンの脳高次機能における役割を明らかにする目的で,α キメリンノックア ウトマウスの包括的行動解析を行った.その結果,このマウスは過活動など幅広い表現型を示したが(data not shown),特に我々が注目したのは海馬依存的学習である文脈型恐怖条件付けの亢進である(図 1A).一方,海馬機能 を必要としない手がかり型恐怖条件付けの成績には変化はなかった.これらの結果から,全身性 α キメリンノックア ウトマウスにおいて海馬依存的学習が亢進していることが示唆された. 次いで,α キメリンの flox マウスを Emx1Cre マウス 4)と交配し,背側終脳(海馬,大脳皮質など)の興奮性神経細 胞特異的に α キメリンをノックアウトした.このマウスは両足を交互に出す普通の歩行をし,活動量も正常であった (data not shown).一方,文脈型恐怖条件付けの亢進は再現された(図1 B).これらの結果は,全身性ノックアウトで 見られた文脈型恐怖条件付けの亢進が,過活動やウサギ様歩行の間接的な影響ではなく,背側終脳(海馬など)での α キメリン欠損の直接の影響であることを示すものである. 1 図 1. α キメリン変異マウスにおける文脈型恐怖学習の亢進. A) コントロールマウス(白)と比較して全身性 α キメリンノックアウトマウス(黒)では手がかり型恐怖学習 の成績には差はないが,文脈型恐怖条学習の成績は有意に亢進していた.B) コントロールマウス(白)と比較 して,背側終脳特異的 α キメリンノックアウトマウス(黒)は文脈型恐怖学習の成績が亢進していた.1:手 がかり型恐怖条件付け.2:文脈型恐怖条件付け.*p < 0.05, **p < 0.01.Unpaired two-tailed Student's t-test. α キメリンには α 1イソフォームと α 2イソフォームが存在するが,それぞれの in vivo での役割の違いは不明で ある.我々は α 1イソフォームと α 2イソフォームのそれぞれに特異的な全身性ノックアウトマウスを作製し,その 包括的行動解析を行った.その結果,それぞれ特徴的な表現型が得られた.恐怖条件付けに関しては,α 1キメリン 特異的ノックアウトマウスは正常であったが(図2 A),α 2キメリンノックアウトマウスでは,α キメリンノック アウトマウスと同様に文脈型に亢進が見られた(図2 B).Emx1Cre を用いて背側終脳特異的 α 2ノックアウトマウ スを作製し解析したところ,同様に恐怖学習の亢進が見られた(図2 C).これらの結果から,α 1ではなく α2 キメ リンが海馬依存的学習の調節に重要な働きをしていることがわかった.さらに,別のタイプの海馬依存的学習である空 間学習に関してもインテリケージシステムを用いて調べたが,α 2キメリンノックアウトマウスで,忌避学習の亢進 が見られた(data not shown). 図 2. 背側終脳特異的 α 2キメリンノックアウトマウスにおける文脈型恐怖学習の亢進. A) コントロールマウス(白)と比較して,全身性 α 1キメリンノックアウトマウス(黒)では,恐怖学習(手 がかり型,文脈型)の成績に差はなかった.B) コントロールマウス(白)と比較して,全身性 α 2キメリンノ ックアウトマウス(黒)では,文脈型恐怖学習の亢進が見られた.*p < 0.05.C) コントロールマウス(白)と 比較して,背側終脳特異的 α 2キメリンノックアウトマウス(黒)では,文脈型恐怖学習の亢進が見られた. *p < 0.05.1:手がかり型恐怖条件付け.2:文脈型恐怖条件付け.Unpaired two-tailed Student's t-test. 2 α 2キメリンは発達期の海馬で強く発現することから(data not shown),α キメリンが発達期において機能するこ とにより成体での学習機能を調節することが考えられた.それを確かめるために,誘導性 Cre マウス(SLICK-H マウ ス)と α 2キメリン flox マウスを交配して,そのマウスに 1 ヶ月齢から薬剤(タモキシフェン)を与えることによ り,成体特異的 α2 キメリンノックアウトマウスを作製した.このマウスで恐怖条件付けのテストをしたところ,学習 に異常は見られなかった(図3).この結果から,成体での α 2キメリン発現は成体での学習能力に必須ではないこと がわかった. 図 3. 成体特異的 α 2キメリンノックアウトマウスにおける正常な文脈型恐怖学習. 1 ヶ月齢以降で α 2キメリンをノックアウトしたマウス(黒)ではコントロールマウス(白)と比較して,恐 怖学習に差は見られなかった.1:手がかり型恐怖条件付け.2:文脈型恐怖条件付け.Unpaired two-tailed Student's t-test. 以上の結果をまとめると,α キメリンの α 2イソフォームが発達期で働くことが,成体での海馬依存的学習の調節 に重要な役割を担うことが明らかとなった.α キメリンは細胞骨格を制御する分子であることから,発達期における 軸索誘導あるいはスパイン形成制御を通じて,脳高次機能の調節に働いていることが考えられる.学習制御の基盤とな る回路レベルの機構の解明が今後の課題である.なお,α キメリンの α 1イソフォームと α 2イソフォームが in vivo において異なった役割を持つことも,本研究によりはじめて明らかになった.本研究成果は論文として最近発表 した 5,6). 共同研究者 本研究の主な共同研究者は,国立遺伝学研究所の岩田亮平博士,理化学研究所脳科学総合研究センターの糸原重美博士 および大阪大学の橋本亮太博士である.本研究の遂行に当たり多大なご支援をいただきました上原記念生命科学財団 に深謝いたします. 文 献 1) Iwasato, T., Katoh, H., Nishimaru, H., Ishikawa, Y., Inoue, H., Saito, Y. M., Ando, R., Iwama, M., Takahashi, R., Negishi, M. & Itohara, S. : Rac-GAP α-chimerin regulates motor-circuit formation as a key mediator of EphrinB3/EphA4 forward signaling. Cell, 130 : 742-753, 2007. 2) Miyake, N., Chilton, J., Psatha, M., Cheng, L., Andrews, C., Chan, W. M., Law, K., Crosier, M., Lindsay, S., Cheung, M., Allen, J., Gutowski, N. J., Ellard, S., Young, E., Iannaccone, A., Appukuttan, B., Stout, J. T., Christiansen, S., Ciccarelli, M. L., Baldi, A., Campioni, M., Zenteno, J. C., Davenport, D., Mariani, L. E., Sahin, M., Guthrie, S. & Engle, E. C. : Human CHN1 mutations hyperactivate α2-chimaerin and cause Duane's retraction syndrome. Science, 321 : 839-843, 2008. 3) Ip, J. P., Shi, L., Chen, Y., Itoh, Y., Fu, W. Y., Betz, A., Yung, W. H., Gotoh, Y., Fu, A. K. & Ip, N. Y. : α2chimaerin controls neuronal migration and functioning of the cerebral cortex through CRMP-2. Nat. Neurosci., 15 : 39-47, 2011. 4) Iwasato, T., Datwani, A., Wolf, A. M., Nishiyama, H., Taguchi, Y., Tonegawa, S., Knopfel, T., Erzurumlu, R. S. & Itohara, S. : Cortex-restricted disruption of NMDAR1 impairs neuronal patterns in the barrel cortex. Nature, 406 : 726-731, 2000. 3 5) Mizuno, H., Luo, W., Tarusawa, E., Saito, Y. M., Sato, T., Yoshimura, Y., Itohara, S. & Iwasato, T. : NMDARregulated dynamics of layer 4 neuronal dendrites during thalamocortical reorganization in neonates. Neuron, 82 : 365-379, 2014. 6) Iwata, R., Ohi, K., Kobayashi, Y., Masuda, A., Iwama, M., Yasuda, Y., Yamamori, H., Tanaka, M., Hashimoto, R., Itohara, S. & Iwasato, T. : RacGAP α2-chimaerin function in development adjusts cognitive ability in adulthood. Cell Rep., 8 : 1257-1264, 2014 . 4
© Copyright 2026 Paperzz