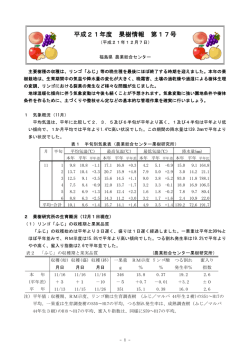
CODEN:HDNHFL ISSN 1344-8897 弘前大学農学生命科学部学術報告 第9号 BULLETIN OF THE FACULTY OF AGRICULTURE AND LIFE SCIENCE HIROSAKI UNIVERSITY No.9 付 研 究 業 績 目 録 2005年10月―2006年9月 Lists of Published Research Works of the Faculty of Agriculture and Life Science Hirosaki University 2005(October)–2006(September) 弘 前 大 学 農 学 生 命 科 学 部 2006年12月 FACULTY OF AGRICULTURE AND LIFE SCIENCE HIROSAKI UNIVERSITY HIROSAKI 036-8561, JAPAN December, 2006 弘前大学農学生命科学部学術報告 第9号 2006年12月 目 次 松岡教理・柳野克之:ウグイ地域集団間の遺伝的距離 1 松岡教理:アロザイム変異からみたヒトデ類4科の系統類縁関係 8 吉田 渉・大中臣哲也・石田幸子:マナマコ複合盤状体骨片の成長と体長と の関係 15 武田 潔・須田育江・殿内暁夫:リンゴ搾り粕のメタン発酵 ― メタン発酵 汚泥から分離したペクチン・キシラン分解菌の特徴 ― 21 浅田武典:リンゴ樹における発生部位別スパー構成に関する研究 28 宇野忠義:リンゴ果汁輸入の急増が日本のリンゴ経営に与えた影響 ― リン ゴ過剰恐慌輸出によってもたらされた経営危機 ― 68 宇野忠義・佐野輝男・田中和明:ニュージーランドのリンゴ生産と病虫害防 除 ― リンゴ火傷病とオタゴ大学訪問記録 ― 80 藤崎浩幸:安全性の高い野菜に対する消費者意識 ― みやぎ生協「産直野菜」 に対する来店者意識調査 ― 91 研究業績目録(2005 年 10 月 ― 2006 年 9 月) 101 1 Ge ne t i cDi s t a nc ebet we enPopul at i onsoft he Cypr i ni dFi s hTr i bol odonhak one ns i s Nor i ma s aMATSUOKA* a ndYo s hi yukiYANAGI NO Di v i s i o no fMo l e c u l a rEv o l u t i o n ,Fa c u l t yo fAg r i c u l t u r e& Li f eS c i e n c e , Hi r o s a k iUn i v e r s i t y ,Hi r o s a k i0 3 6 8 5 6 1 ,J a p a n (Rec ei vedf o rpubl i c a t i o nSe pt embe r2 5 ,2 0 0 6) I nt r oduct i on I npar al l elwi t hmol ec ul arphyl ogenyofec hi noder ms ,wehavebeens t udyi ngmo l ec ul arphyl ogenyof f i s heswhi c h have r i c hs pec i esdi ver s i t y. Unt i lno w,we r epor t ed mol ec ul arphyl ogeny ofs o me f i s h gr oupsby al l ozyme ormi t oc hondr i alDNA anal ys i s :genusOn (Ki t ano e ami l y c hor hy nc h us ta l . ,1997);f Hexagr a mmi dae (Mat s uoka e derCl upei f or mes (As anuma and Mat s uoka,2002) ;s ubor der ta l . ,2000);or Per c oi dei (Mat s uoka,2003) ;f ami l y Os mer i dae (Ma t s uoka and Ki t ano,2003) ;t wo body c ol or t ypes of (Mat s uoka and Sugi yama,2005) .Ho wever ,we have not yet r epor t ed mol ec ul ar S e b a s t e ri n e r mi s phyl ogenyoft her epr es ent at i vef a mi l yCypr i ni daei nc l udi ngmanyf r es hwat erf i s hs pec i esi nJ apan. sone ofc ommo ns pec i esoft he Cypr i ni dae i nJ apan and wi del y di s t r i but ed T r i b o l o d o nh a k o n e n s i si f r om Hokkai dot oKyus yu.I ti sal s odi s t r i but edi nt hes out her npar tofKor eanPeni ns ul aandSakhal i ni n t he As i an Cont i nent . The s pec i ess howst he r emar kabl e var i at i on i nt he mor phol ogy and ec ol ogy. Forexampl e,t he s pec i mensf r om r i verhave s mal lbody s i ze,butt hos ef r o m l ar ge l ake s uc h asLake Bi wa i nJ apan and s eas s ho w l ar ge body s i ze. Fur t her ,t he s pec i es di ver s i f i es ec ol ogi c al l y and i nhabi t si nt he s t r ong ac i di c wat er s (pH i sabout3)oft he l ake ofUs or i ko i n Shi moki t aPeni ns ul a of Aomor iPr ef . Al t hough i ts howswi degeogr aphi c aldi s t r i but i on,genet i cdi f f er ent i at i on bet we en di f f er ent ons i dert hats uc h l oc alpopul at i onsoft hes pec i eshasnotyetbeen exa mi ned atmol ec ul arl evel . Wec popul at i on genet i cs t udy i si mpor t antand val uabl et oc l ar i f yt hes pec i at i on off i s h. Asaf i r s ts t ep of s uc h popul at i on genet i cs t udi es ,wei nves t i gat ed t heext entofgenet i cdi f f er ent i at i on bet ween t wol oc al i b o l o d o nh a k o n e n s i s :o nei spopul at i on f r o ml ar gel akeofLakeBi wai nt hemi ddl epar tof popul at i onsofTr J apan and t heot herf r o mr i verofAomor iPr ef .i n nor t her nJ apan. I ti swel lkno wn t hatpopul at i on f r o m Lake Bi wa i sc har ac t er i s t i c mor phol ogi c al l y when c ompar ed wi t h ot her l oc alpopul at i ons . Na ka mur a(1969)r epor t ed t hatpopul at i on ofT r om Lake Bi wa s ho ws t he c har ac t er i s t i c .h a k o n e n s i sf mor phol ogy:Gener al l y,i n many l oc alpopul at i onsoft hes pec i es ,t hel engt h ofl o werj a wi ss hor t ert han t hatofupperj a w. However ,popul at i on f r om LakeBi wahast hepr oj ec t i ng l o werj a w,and t hust he upperand l o we rj awss ho wt hes amel engt h and t hes nouti ss har ppoi nt ed,whi c h ar enotobs er ved i n ot her l oc alpopul at i on (F s i g.1) .Fur t her mor e,popul at i on f r o m Lake Bi wa has mor es l ender body. Ther ef or e,i ti si nt er es t i ng t oi nves t i gat et heext entofgenet i cdi f f er ent i at i on bet ween popul at i onsf r o m heout gr oup i n anal ys i soft wopopul at i ons LakeBi waand ot herpopul at i onsatmol ec ul arl evel . Ast ofT ed t he c ypr i ni df i s h Ph I ti s al s o one of .h a k o n e n s i s ,we adopt o x i n u sl a g o ws k i is t e i n d a c h n e r i . c o mmo ns pec i esi nJ apanandwi del ydi s t r i but edi nHons huofJ apan,Kor ea,Chi na,Mongol i aandSi ber i a. I nt heuppers t r ea m ofmanyr i ver si nJ apan,t het woc ypr i ni ds pec i esar es ympat r i c . * Co r r e s po ndi nga ut hor Bul l .Fac .Agr i c .& Li f eSc i .Hi r os akiUni v.No.9:1 -7,2006 2 nd Yo s hi yukiYANAGI No r i mas aMATSUOKA a NO Fi g.1. T r om LakeBi wa . r i bol odonhak one ns i sf A:Adul tf i s h(upperi smal eand l owe rf ema l e )f r om La keBi wa,B:Fr yf r om La keBi wa,C: Adul tf i s ho fot herpopul at i ons ,a nd D:Ve nt r a lvi ew o fhe a do fa dul tf i s hs hown i n C.Thi s f i gur ewasquot ed f r om Na kamur a (1 9 6 9 ) . I nt hi spaper ,wer epor tt hegenet i cr el at i ons hi poft wol oc alpopul at i ons (f r om LakeBi waandr i ver i n Ao mor i Pr ef . )of T hat of t wo c ypr i ni df i s h,T .h a k o n e n s i s ,and t .h a k o n e n s i s and P .l a g o ws k i i nf er r edf r o m al l ozymevar i at i on. s t e i n d a c h n e r ii Al l ozymeVa r i a t i o ni nT .h a k o n e n s i s 3 Mat er i al sandMet hods T l ec t ed f r om t wo di f f er entl oc al i t i es :Lake Bi wa i n Shi ga Pr ef .att he r i b o l o d o nh a k o n e n s i swas col c ent r alpar tofJ apanandt her i verofHi r aka waatOhwa ni mac hii nAomor iPr ef .ofnor t her nJ apan. he s ame f ami l y Ci pr i ni dae wa sc ol l ec t ed f r o m t he r i ver of Ph o x i n u sl a g o ws k i is t e i n d a c h n e r i of t Shi madaga wa at Ohwa ni mac hii n Aomor iPr ef . The number ofi ndi vi dual s exa mi ned was 10 f or orP popul at i on f r o m LakeBi waand 8f orpopul at i on f r o m Hi r aka wai nT .h a k a o n e n s i s ,and 6f .l a g o ws k i i The t wo s pec i eswe r ec ol l ec t ed by f i s hi ng. Af t erc ol l ec t i on,t he whol e bodi eswer e s t e i n d a c h n e r i . s t or ed at -40 ℃ unt i lus e. Pr ot ei n el ec t r ophor es i s was per f or med on 7. 5 % pol yac r yl a mi de gelas des c r i bed i n Mat s uoka and Hat anaka(1991) :The mus c l e wasc utof ff r o m s pec i mens ,and about0. 2g phat e buf f erc ont ai ni ng 0. 1 M KCland mus c l e wasi ndi vi dual l y homogeni z ed wi t h 3 vol s .of20mM phos 1mM EDTA(pH 7. 0)by us i ng Pot t er El vehj em t ypehomogeni zeri n an i c ewat erbat h. Thet i s s ue ext r ac t was c ent r i f uged at 10, 000 r pm f or 5 mi n and t he c l ear s uper nat ant was us ed f or pr ot ei n el ec t r ophor es i s . El ec t r ode buf f er wa s Gl yc i net r i s buf f er ,pH 8 . 3. Af t er e l ec t r ophor es i s ,a l l ozyme var i at i onoft hef ol l owi ngei ghtdi f f er entenzymeswasanal yzed:al c oholdehydr ogenas ,gl uc os e6e(ADH) phos phat e dehydr ogenas e (G6PD) ,l ac t at e dehydr ogenas e (LDH) , not hi ng dehydr ogenas e (NDH) , xant hi ne dehydr ogenas e(XDH) ,αgl yc er ophos phat e dehydr ogenas e (αGPDH) ,f umar as e(FUM)and s uper oxi de di s mut as e(SOD) . Al l ozyme anal ys i s wa sc ar r i ed out a s des c r i bed i n Ma t s uoka and Hat anaka (1991) . Res ul t sandDi s cus s i on I n pr evi ous s t udi es ,we i ndi c at ed t hat al l ozyme el ec t r ophor es i si s muc h ef f ec t ualmet hod f or di s t i nc t i onori dent i f i c at i onofs i bl i ngs pec i esormor phol ogi c al l yver ys i mi l ars pec i esi nec hi noder ms . For exa mpl e,t hes eaur c hi n Ec c hi sc o mmo ns pec i esi nt hes easofs out her nJ apan i s h i n o me t r ama t h a e iwhi muc h var i abl ei n mor phol ogy and ec ol ogy,and t her e exi s tf ourdi f f er entt ypes (TypeA,B,C and D) wi t hi nt he s eaur c hi nf r o m Oki nawa. We i nves t i gat ed t he pr obl e m whet her t hes ef our t ypes ar e di s t i nc ts pec i esby al l ozymeanal ys i s . Ther es ul t sr eveal ed t hatt hey ar egenet i c al l y di f f er ent i at ed t o one anot her and f our t ypes ar e di s t i nc ts pec i es (Mat s uoka and Ha t anaka,1991) . Fur t her ,we r epor t ed t heal l ozymes t udy s howi ng t hatt hec ommo ns t ar f i s hAs r om J apanes ewat er s t e r i a sa mu r e n s i sf mi ghtc ons i s t soft hr ee genet i c al l y di f f er ents pec i es (Ma t s uoka and Ha t anaka,1998) . Mor er ec ent l y, we al s o demo ns t r at ed t hat t he pr i mi t i ve s eaur c hi n As r om J apan c ons i s t s oft wo t h e n o s o ma i j i ma if di s t i nc ts pec i es (one i s A.i he ot her f r o m Oki na wa i s A.i )(Mat s uoka e j i ma i and t j i ma iR. ta l . , 2004) . As des c r i bed above,we s howed t hat al l ozyme anal ys i si s muc h ef f ec t i ve i n di s t i nc t i on of s i bl i ngs pec i es . Rec ent l y,mt DNA anal ys i shasof t en been us ed i nt hef i el d ofmol ec ul arphyl ogeny. I n gener al ,i t i sc ons i der ed t hatpr ot ei n el ec t r ophor es i si snotmor es ens i t i ve t han mt DNA anal ys wever ,i n i s . Ho c ompar i s onbet weenal l ozymeandmt DNA anal ys i s ,Nei (1987)s ugges t edt hatt her es ol vi ngpo weroft he l at t eri snotnec es s ar i l y hi ghert han t hatoft he f or mer . Ac c or di ng t ot he es t i mat i on ofNe i (1987) , enzyme el ec t r ophor es i si s expec t ed t os ur vey about100 nuc l eot i des per l oc us . I fwe exa mi ned 30 genet i cl oc ibyel ec t r ophor es i s ,i ti sequi val entt os t udyi ng3, 000bas epai r satmt DNA l evel . Ther ef or e, t he r ys i s . Forexa mpl e,Neiand es ol vi ng powerofal l ozyme anal ys i si snotl owert han mt DNA anal Royc houdhur y (1993)s uc c eeded i n el uc i dat i on of evol ut i onar yr el at i ons hi ps of human popul at i ons by al l ozyme s t udy,and t he r es ul t wa s wel lc ons i s t ent wi t h mt DNA s t udy by Cann e (1 ta l . , 987). Fur t her mo r e,Mur phy e (1996)c l ai med t hat i n phyl ogenet i cs t udi es many mol ec ul ar c har ac t er s ta l . s houl d be us ed and t hatt he enzyme l oc iar et he i mpor t antmol ec ul ar c har ac t er s . The number of nd Yo s hi yukiYANAGI No r i mas aMATSUOKA a NO 4 Ta bl e1.Al l el e f r eque nc i e s at s i x po l ymor phi c l o c ii n t wo popul at i onsofT .h a k o n e n s i s Loc us Adh G6pd2 αGpdh1 Ldh1 Ldh2 Ndh Al l e l e ThB ThA a b a b a b a b a b a b 0 . 4 4 0 . 5 6 0 . 5 0 0 . 5 0 0 . 6 0 0 . 4 0 0 . 7 8 0 . 2 2 0 . 4 5 0 . 5 5 0 . 4 0 0 . 6 0 0 . 4 3 0 . 5 7 0 . 3 7 0 . 6 3 0 . 5 0 0 . 5 0 0 . 5 0 0 . 5 0 0 . 5 0 0 . 5 0 0 . 3 3 0 . 6 7 ThB=popul at i onf r om La keBi wao fT .h a k o n e n s i s , ThA=popul at i onf r om t her i ve ri nAomo r iPr e f .o fT .h a k o n e n s i s . Ta bl e2.Genet i ci dent i t i e s (a bo vedi a go na l )a nd ge ne t i cdi s t a nc e s (bel ow di agona l )amongt hr e epo pul a t i o nso fT .h a k o n e n s i s andP .l a gows k i is t e i n d a c h n e r i Popul at i ons 1.ThB 2.ThA 3.Pl s 1 2 3 0 . 0 1 5 0 . 7 1 5 0 . 9 8 5 0 . 6 0 6 0 . 4 8 9 0 . 5 4 6 - Genet i ci dent i t i es (I )andge ne t i cdi s t a nc e s (D)wer ec a l c ul a t e dby t he met hod ofNe i (19 7 2 ) .ThB=po pul a t i o nf r om La ke Bi wa o fT . hak one ns i s ,ThA=popul a t i o nf r om t he r i ve ri n Ao mor iPr e f .o fT . hak one ns i s ,Pl s =P .l a gows k i is t e i n d a c h n e r i mol ec ul ar c har ac t er s adopt ed i n pr ot ei n el ec t r ophor es i si s mor e enough t han t hatofmt DNA s t udy. Pr ot ei nel ec t r ophor es i si soneofpower f ult ec hni quesf ores t i mat i ngt hephyl ogenet i cr el at i ons hi ps . Byal l ozymeanal ys i s ,weat t empt edt oc l ar i f yt hepr obl em whet herpopul at i onf r o m LakeBi waofT . (i ts hows c har ac t er i s t i c e xt er nal mor phol ogy as evi dent f r om Fi g.1 )i s genet i c al l y ha k a n e n s i s gr oup wasP he s ame di f f er ent i at ed f r o m ot herl oc alpopul at i on. The out .l a g o ws k i is t e i n d a c h n e r ioft f a mi l y Cypr i ni dae whi c hi ss ympat i r i c wi t hT n many ar eas i nJ apan. Fr om al l ozyme .h a k o n e n s i si var i at i on ofei ghtdi f f er entenzymes ,12genet i cl oc iwer es c or ed:Ad h ,G6 p d 1 3 , L d h 1 2 , N d h , X d h ,α〜 〜 Oft hes e12genet i cl oc i ,s i xl oc i (Ad ) o d . h ,G6 p d 2 ,αGp d h 1 ,Ld h 1 h Gp d h 1 m andS 〜2andNd 〜2Fu r e mo nomor phi c . Tabl e1s ho ws t he al l el e we r e pol ymo r phi c ,a nd t he r emai ni ng s i x l oc i we f r equenc i esats i x pol ymor phi cl oc iofa t oat al12 genet i cl oc ii nt wo popul at i onsofT .h a k o n e n s i s .To quant i f yt heext entofgenet i cdi f f er ent i at i on bet ween t wopopul at i ons ,genet i ci dent i t i es (I )and genet i c di s t anc es (D)wer ec al c ul at ed f r om al l el ef r equenc i es dat ai n 12 genet i cl oc iby t he met hod ofNei (1972). Tabl e2s howst he mat r i x ofIand D val ues . Asa r e s ul t ,t he genet i ci dent i t y bet ween t wol oc alpopul at i onsofT =0. 985andt hegenet i cdi s t anc eD=0. 015. Thehi ghIval ue .h a k o n e n s i swasI was c o mpar abl et o Ival ues obs er ved bet ween c ons pec i f i cl oc alpopul at i ons i n many ot her ani mal s (Thor pe,1982) . Fur t her mor e,asevi dentf r om Tabl e1,t hes t at i s t i c al l ys i gni f i c antdi f f er enc esofal l el e f r equenc i esbet weent wol oc alpopul at i onswer enotobs er vedi nanyl oc i ,andt hedi agnos t i cl oc is ho wi ng t hedi f f er enc eofal l el ec ompos i t i on wer enotf ound. I n gener al ,s i bl i ng s pec i esorver yc l os el yr el at ed Al l ozymeVa r i a t i o ni nT .h a k o n e n s i s 5 s pec i es s how t he s i gni f i c ant di f f er enc es of al l el ef r equenc i es i ns o me genet i cl oc iand have s o me di agnos t i cl oc is ho wi ng t hedi f f er ental l el ec ompos i t i on(Mat s uokaand Hat anaka,1991) . Thepr es ent al l ozyme s t udy i ndi c at ed t hatt wo l oc alpopul at i ons ofT r o m Lake Bi wa and r i ver i n .h a k a o n e n s i sf Aomo r iPr ef .wer el i t t l egenet i c al l y di f f er ent i at ed. Thes i mi l aral l ozymer es ul t swer eal s oobt ai ned i n t wo di f f er entbody c ol ort ypes (Bl ac kt ype and Redbr ownt ype )oft he f i s hS (Mat s uoka e b a s t e si n e r mi s andSugi ya ma ,2005) . On t heot herhand,t heaver agegenet i ci dent i t y val uebet we en t wodi f f er ents pec i es ,T .h a k o n e n s i s and P =0. 518. I n gener al ,t he Ival ues bet ween c onf a mi l i aldi f f er ent .l a g o ws k i is t e i n d a c h n e r iwas I gener aar ei nt her angeofI =0. 20〜0. 35i nmanyani mal s (Thor pe,1982 ) . Thepr es ental l ozymes t udy s howe dt hatt heIval ue(I =0. 518)bet we en T abl e .h a k o n e n s i sand P .l a g o ws k i is t e i n d a c h n e r iwascompar t o I val ues obs er ved bet ween di f f er ent s pec i es bel ongi ng t os ame genus . Namel y,t hey ar e at c ongener ’ sl evel ,t hought axonomi cs ys t em c l as s i f i edt he mi nt ot wodi s t i nc tgener a. TheLakeBi wai sat r eas ur yoff r es hwat erf i s hi nJ apanandhasr i c hs pec i esdi ver s i t y. However , dur i ng t hel as tabout20year s ,t henumberofi ndi vi dual sofs o mes pec i ess uc h asPl e c o g l o s s u sa l t i v e l i s ct o Lake Bi wa(t hey ar e del i c i ousedi bl ef i s h)have r a s s i u sa u r a t u sg r a n d o c u l i sendemi a l t i v e l i sand Ca )and bl ac kbas s (Mi ext r e me l y dec r eas ed by t he i nt r oduc ed f i s h bl uegi l l (Le p o mi sma c r o c h i r u s c r o p t e r u s )f r om Nor t h Ame r i c a whi c hf eed eggs or young f i s h oft hes e ende mi cs pec i es f er oc i ous l y. s a l mo i d e s Thes ei nt r oduc ed f i s h have s t r ong r epr oduc t i ve po wer and ec ol ogi c alpr edo mi nanc e,and t hus t he at i on s i ze)i si nc r eas i ng dr as t i c al l y. Asar es ul t ,t heec ol ogi c als ys t em of numberofi ndi vi dual s (popul Lake Bi wa dr as t i c al l yc hanged by t hes ei nt r oduc ed f i s h. The number ofi ndi vi dual s ofc o mmo n s pec i esT s obeendec r eas i ngbyt hei nt r oduc edf i s hi nr ec entyear s ,andt husi ti smor e .h a k o n e n s i shasal di f f i c ul tt oc ol l ec ts pec i mens . Under s uc hs i t uat i on,popul at i on gene t i cs t udy o ft he s pec i es at mol ec ul arl evelwoul dpr ovi des omeus ef uli nf or mat i onf orel uc i dat i ngt hephyl ogenyorevol ut i onoff r es hwat erf i s h. I nc onc l us i on,t hepr es ental l ozymes t udy i ndi c at ed t hatpopul at i on f r o m LakeBi waofT .h a k o n e n s i s s howi ng c har ac t er i s t i c mo r phol ogy i sl i t t l e di f f er ent i at ed genet i c al l yf r om popul at i on f r om r i ver i n Aomo r iPr ef .i n nor t her nJ apan,and t hatT ghtbec l as s i f i ed .h a k o n e n s i sand P .l a g o ws k i is t e i n d a c h n e r imi asc ongener son ac c ountoft he hi gh genet i ci dent i t y ver i f i ed by t hi sal l ozyme s t udy. Fur t her ,t he evol ut i onar yr at eatmor phol ogi c all eveli nf i s h mi ghtbef as t ert han t hati n ot herani malgr oupswhen c ons i der edt her el at i onbet weengenet i ci dent i t yandt axonomi cr ank(mor phol ogi c aldi f f er enc es ) . Ack nowl edgment s Iam gr at ef ult o Dr .Sakai ,H. ,Nat i onalFi s her sUni ver s i t y and Dr .Sawa r a,Y. ,Hi r os akiUni ver s i t y f ort hei rval uabl eadvi c e,and t oMa t s uoka,K. ,Shi gaPr ef .f orpr ovi di ng s pec i mensofT r om .h a k o n e n s i sf LakeBi wa. Abs t r act T s one ofc o mmo nc ypr i ni df i s hs pec i es i nJ apan and wi del y di s t r i but ed i n r i b o l o d o nh a k o n e n s i si eas t er n As i an Cont i nent . Thel oc alpopul at i onsoft hes pec i esi nJ apan s ho wr emar kabl evar i at i on i n mor phol ogi c aland ec ol ogi c alc har ac t er s . I n par t i c ul ar ,popul at i on f r o m Lake Bi wa i n Shi ga Pr ef .i n mi ddl e par tofJ apan i sc har ac t er i s t i c atmor phol ogi c alvi e wpoi nt :The popul at i on has t he pr oj ec t i ng l ower j aw and s l ender body whi c h ar e notobs er ved i n ot her l oc alpopul t i ons . We exami ned t he ext entofgenet i cdi f f er ent i at i on bet ween t wol oc alpopul at i onsofT r o m LakeBi waand t he .h a k o n e n s i sf r i veri n Aomo r iPr ef .ofnor t her nJ apan by al l ozymeanal ys i s . Theoutgr oup wa sPh o x i n u sl a g o ws k i i 6 nd Yo s hi yukiYANAGI No r i mas aMATSUOKA a NO hes amef ami l y Cypr i ni dae. Twe l vegenet i cl oc iwer edet ec t ed by el ec t r ophor es i sof s t e i n d a c h n e r ioft ) ei ghtdi f f er entenzymes . Fr om al l el ef r equenc i esdat ai n 12 genet i cl oc is c or ed,genet i ci dent i t y(I andgenet i cdi s t anc e(D)wer ec al c ul at edbyt hemet hodofNei (1972) . Theal l ozymes t udyi ndi c at ed t hef ol l owi ngs : (1)Thegenet i ci dent i t y(I =0. 985)bet weent wol oc alpopul at i onswasmuc hhi ghval ueand c ompar abl et o Ival ues bet ween c ons pec i f i cl oc alpopul at i ons i n many ot her ani mal s ,but was not equi val entwi t ht heIval uesobs er ved bet ween s i bl i ng s pec i esorver yc l os el yr el at ed s pec i es . (2)The s t at i s t i c al l ys i gni f i c antdi f f er enc esofal l el ef r equenc i esbet ween t wo popul at i onswer e notf ound i n any l oc i . (3)Thedi agnos t i cl oc iwhi c hs how t hedi f f er enc eofal l el ec o mpos i t i onwer enotobs er ved. (4)The ue bet ween T =0. 518 and equi val entwi t ht hos e Ival .h a k o n e n s i sand P .l a g o ws k i is t e i n d a c h n e r iwas I bet weenc ongener si nmanyot herani mal s . Thes eal l ozymer es ul t sdemons t r at edt hatt wopopul at i ons el i t t l e genet i c al l y di f f er ent i at ed,and t hatt wo c ypr i ni df i s h,T ofT .h a k o n e n s i sar .h a k o n e n s i sand P . ghtbe c l as s i f i ed as c ongener s atmol ec ul ar l evel . The pr es ents t udy al s o l a g o ws k i is t e i n d a c h n e r imi s ugges t edt hatf i s hmi ghthavef as t erevol ut i onar yr at et hanot herani mal satmor phol ogi c all evel ,t hough t hemol ec ul arevol ut i onar yr at ei sc ons t ant . Ref er ences ASANUMA,T.and MATSUOKA,N. (2002)Phyl ogene t i cr e l a t i o ns hi ps amo ng s e ve nf i s hs pe c i e so ft he o r de r Cl upei f or me s i nf er r edf r o m al l ozymevar i at i on.Bul l .Fac .Agr i c .& Li f eSc i .Hi r o s a kiUni v.No. 4:1 1 5 . (i nJ a pa ne s e ) CANN,R. L. ,STONEKING,M.andWILSON,A. C. (1987)Mi t o c ho ndr i a lDNA a ndhumane vo l ut i o n.Na t ur e ,3 2 5:3 1 3 6 . c o r h y n c h u ss KITANO,T. ,MATSUOKA,N.andSAITOU,N. (1997)Phyl o ge ne t i cr e l a t i o ns hi po ft hege nusOn pe c i e si nf e r r edf r om nuc l earandmi t oc hondr i almar ker s .GenesGene t .Sys t . ,7 1:2 5 3 4 . Ma t s uoka,N. (2003 )Mol ec ul arphyl ogenyandal l oz ymeva r i a t i o no ft hef i vec ommonf i s hs pe c i e so ft hes ubo r de rPer c oi dei . Bul l .Fac .Agr i c .& Li f eSc i .Hi r os akiUni v.No. 5:1 7 2 2 . MATSUOKA,N.andHATANAKA,T. (1991)Mol ec ul are vi de nc ef o rt hee xi s t e nc eo ff o urs i bl i ngs pe c i e swi t hi nt hes e aur c hi n, Ec hi nome t r amat hae i ,f r om J apanes ewat er s .Zo o l .Sc i . ,8:1 2 1 1 3 3 . t e r i a sa mur e ns i s MATSUOKA,N.and HATANAKA,T. (1998)Genet i cdi f f e r e nt i a t i o n amo ng l o c a lpo pul a t i o nso ft hes t a r f i s h As i nf er r edf r o m al l ozymevar i at i on.GenesGenet .Sys t . ,7 3:5 9 6 4 . MATSUOKA,N.andKITANO,T. (2003)Genet i cdi f f er e nt i a t i o no ft hef ami l yOs mer i da eba s e do na l l o z yme .Bul l .Fa c .Agr i c .& Li f eSc i .Hi r os akiUni v.No. 6:4048. MATSUOKA,N. ,KOHYAMA,K. ,ARAKAWA,E.and AMEMIYA,S. (2 0 0 4)Mol e c ul a re vi de nc ef o rt he e xi s t e nc eo ft wo s i bl i ng s pec i eswi t hi nt heec hi not hur i oi dec hi noi dAs r om J a pa ne s ewat e r s .Zo o l .Sc i . ,2 1:1 0 5 7 1 0 6 1 . t h e n o s o mai j i ma if e b a s t e si ne r mi s MATSUOKA,N.andSUGIYAMA,T. (2005)Mol ec ul art a xo nomyo ft wodi f f e r e ntt ype so fbo dyc o l o ri nt hef i s hS f r om J apanes ewa t er s .Bul l .Fac .Agr i c .& Li f eSc i .Hi r o s a kiUni v.No. 7:1 8 . (i nJ a pa ne s e ) MATSUOKA,N. ,SUGIYAMA,T.and ASANUMA,T. (2 0 0 0 )Phyl o ge ne t i cr e l a t i o ns hi ps among f o ur f i s hs pe c i e so ft he f ami l y He xagr a mmi daef r om J apanes ewat er sbas edona l l o z ymes .Bul l .Fa c .Agr i c .& Li f eSc i .Hi r o s a kiUni v.No. 3:5 13. l e c u l a rS y s t e mat i c s MURPHY,R. W. ,SITES,J . W. ,BUTH,D. G.andHAUFLER,C. H. (1 9 9 6 )Pr o t e i ns :I s o z ymee l e c t r o pho r e s i s .I nMo (edi t edbyHi l l s ,D. M. ,Mo r i t z,C.andMabl e,B. K. ) ,pp. 5 1 1 2 0 ,Si na ue rAs s o c i a t e s ,Sunde r l a nd. pr i ni dFi s he sofJ apan.Res NAKAMURA,M. (1969)Cy e a r c hI ns t i t ut ef o rNa t ur a lRe s o uc e s ,To kyo . NEI,M. (1972)Genet i cdi s t anc ebet weenpopul at i ons .Am.Na t . ,1 0 6:2 8 3 2 9 2 . l e c ul arEv ol ut i onar yGe ne t i c s .Col NEI,M. (1987)Mo umbi aUni vPr e s s ,Ne w Yo r k. NEI,M.andROYCHOUDHURY,K (1 . 993)Evol ut i onar yr e l a t i o ns hi pso fhuma npo pul a t i o nso nagl o ba ls c a l e .Mo l .Bi o l .Evol . ,10: 9279 43. me r i c alT a x o n o my .Fr SNEATH,P. H. A.andSOKAL,P. R. (1973)Nu e eman,Sa nFr a nc i s c o . THORPE,J . P( .1982 )Themol ec ul arc l oc khypot hes i s :Bi o c hemi c a le vo l ut i o n,ge ne t i cdi f f e r e nt i a t i o n,a nds ys t ema t i c s .Ann.Rev. Ec ol .Sys t . ,13:139169. Al l ozymeVa r i a t i o ni nT .h a k o n e n s i s 7 ウグイ地域集団間の遺伝的距離 松岡 教理・柳野 克之 弘前大学農学生命科学部分子進化学研究室 ウグイ(T )は,日本では北海道 r i b o l o d o nh a k o n e n s i s =0 . 9 8 5と極めて高く(遺伝的距 集団間の遺伝的類似度は I から九州までの河川,湖,海洋に広く分布するコイ科魚 離は D=0 . 0 1 5と小さく),(2 )2地域集団間で対立遺伝子 類の普通種である。本種は形態・生態学的に地理的変異 頻度に統計的有意差を示す遺伝子座は見られず,(3 )対 が著しく,特に琵琶湖集団は他の集団と比較して形態学 立遺伝子組成が全く異なるDi a gno s t i cLo c i も観察されな 的に特徴的である(上顎と下顎との先端はほぼ同長で吻 かった。これらの結果より,ウグイ 2地域集団は形態学 端が尖る,体型が細長いなど) 。本研究では,このウグイ 的には異なるが,ほとんど遺伝的分化を遂げていないこ 琵琶湖集団が本州最北端の青森県河川集団と,どの程度 とが判明した。すなわち琵琶湖集団の形態学的差異は種 の遺伝的分化を遂げているのか?という集団遺伝学的問 内変異であると推察できる。またウグイとアブラハヤ 題をアロザイム分析により調査した。アウトグループに は,形態レベルからの分類体系では同科別属に分類され は,河川上流部でウグイとよく混生している同じコイ科 ているが,2種間の遺伝的類似度は I =0 . 5 1 8と比較的高 のアブラハヤ(Ph )を用い o x i n u sl a g o ws k i is t e i n d a c h n e r i く,他動物の同属別種間での値と同等であった。これら た。8酵素の電気泳動によるアロザイム分析から 1 2酵素 の結果は,魚類は他の動物群と比較して表現型レベルの 遺伝子座が検出された。1 2遺伝子座における対立遺伝 進化速度が速いことも示唆している。 子頻度から,Ne i (1 9 7 2 )の遺伝的類似度(I )と遺伝的 距離(D)を算出した。その結果, (1 )ウグイの 2地域 弘大農生報 No.9:1 -7,2006 8 Phyl o ge ne t i cRe l at i ons hi psofFo urFa mi l i esof t heAs t e r o i deabas edonAl l ozyme No r i ma s aMATSUOKA Di v i s i o no fMo l e c u l a rEv o l u t i o n ,Fa c u l t yo fAg r i c u l t u r e& Li f eS c i e n c e , Hi r o s a k iUn i v e r s i t y ,Hi r o s a k i0 3 6 8 5 6 1 ,J a p a n (Rec ei vedf o rpubl i c a t i o nSe pt embe r2 5 ,2 0 0 6) I nt r oduct i on Ec hi noder ms ar e a t t r ac t i ve i nver t ebr at es at t he e vol ut i onar y vi ewpoi nt . Fr om s ome nonmol ec ul arand mol ec ul arevi denc e,ec hi noder msar ec l os el yr el at ed t over t ebr at es ,and t hust hey ar eon t hekeypos i t i onofani malevol ut i on. Theevi denc ei sasf ol l o ws : (1 )Thever t ebr at esandec hi noder ms have s kel et on s ys t em. (2)The devel opment albi ol ogy s ho wed t hatt hey bel ong t o De ut er os t omi a. (3) They have hexos e6phos phat e dehydr uc os e6phos phat e dehydr ogenas e (G6PD) ogenas e(H6PD)ofgl i s ozymewhi c ht her edoesnotexi s ti ni nver t ebr at esexc eptec hi noder ms (Mat s uokaandHor i ,1980 ) . (4) Theas t er oi dshaves t r ong t r yps i nl i kedi ges t i veenzymewhi c hi sho mol ogoust ot r yps i n ofver t ebr at es (Hor ie (5)Themet amor phos i si nec hi noi dsi si nduc edbyt hyr oi dhor moneswhi c har ef ound ta l . ,1977). i nver t ebr at es (Chi noe t oe (6)Themol ec ul arphyl ogenet i ct r eec ons t r uc t edby ta l . ,1994;Sai ta l . ,1998). t he s equenc e anal ys i s of s ome pr ot ei ns demons t r at ed t he c l os e af f i ni t y bet ween ec hi noder ms and ver t ebr at es (Mi yat a,1994) . Ihave been i nves t i gat i ng t he phyl ogeny oft he c l as s Ec hi noi dea (ec hi noi ds ) ,whi c hi s one oft he ma j orgr oupsoft hephyl um Ec hi noder mat a,by us i ng al l ozymeanal ys i sand i mmunol ogi c almet hod(eg. , Mat s uoka,1986,1987;Mat s uokaand Hat anaka,1991;Mat s uokaand Suzuki ,1989;Mat s uokae ta l . ,2004). Anot herl ar gegr oup ofEc hi noder mat ai st hec l as sAs t er oi dea (as t er oi ds ) . Al t hough t het axono my of as t er oi ds has been ext ens i vel ys t udi ed f r om mor phol ogi c aland pal eont ol ogi c als t andpoi nt ,t her e ar e di s agr eeme nt sbet ween t axonomi s t s ,and t huss ever alt axono mi cs ys t e mshavebeen pr opos ed(Hayas hi , 1940;Spenc er and Wr i ght ,1966;Cl ar k and Downey,1992 ) . Namel y,many unr es ol ved pr obl e ms n par al l elwi t ht he mo l ec ul ar c onc er ni ng t he phyl ogeny and t axonomy ofas t er oi ds s t i l lr e mai n. I phyl ogenet i cs t udi esofec hi noi ds ,Ir epor t ed t hemol ec ul arphyl ogeny ofs o meas t er oi d gr oup (Ma s t s uoka s uokaand Hat anaka,1998;Mat s uokaand As ano,1999,2003) . Al t hough t hemol ec ul ar e ta l . ,1994:Mat phyl ogenet i cs t udi esofas t er oi dsats pec i esandgenusl evelhavebeenc ar r i edoutbys o mewor ker s ,t he i nt er f ami l i alr el at i ons hi psathi ghert axonomi cr ank havenotyetbeen r es ol ved atmol ec ul arl evelunt i l i r s ts t ep t oc l ar i f yt hepr obl em,Iadopt ed t hef ol l o wi ng f ourr epr es ent at i veand c o mmo n no w. Asaf f ami l i esast ar gett axa:t hef ami l i esAs t er i ni dae,Ac ant has t er i dae,Sol as t er i daeand Ec hi nas t er i dae. One oft hepur pos eoft hi ss t udy i st hemol ec ul arevol ut i on oft r adi t i onaland/orc l as s i c alt axono mi cs ys t e m hi ,1940;Spenc erand Wr i ght ,1966 ) i nc l uded bas ed on mor phol ogy:t he ol dt axonomi cs ys t em(Hayas t hes ef ourf ami l i esi nt ot hes ameor derSpi nul os i da,butt hene wt axonomi cs ys t em whi c h waspr opos ed by Cl ar k and Do wney(1992)c l as s i f i ed t hes ef ourf ami l i esi nt ot hr eedi f f er entor der s :t het wof ami l i es , As t er i ni daeandAc ant has t er i daear emember soft heor derVal vat i da,Ec hi nas t er i daei st hatoft heor der Spi nul os i da,and Sol f f er enc eoft wot axonomi cs ys t e ms as t er i daei st hatoft heor derVel at i da. Thedi Bul l .Fac .Agr i c .& Li f eSc i .Hi r os akiUni v.No.9:8 -14,2006 Bi oc hemi c a ls ys t ema t i c so fa s t e r o i ds 9 i s due t ot he di f f er ent es t i mat i on of t he mor phol ogi c alc har ac t er s adopt ed i nt axonomy. Ot her pur pos et hatIc hos et hes ef our f a mi l i es i st o exami ne t he phyl ogenet i c het er ogenei t y oft he f a mi l y As t er i ni daewhi c h wass ugges t ed by s o mewor ker s . Fr om mor phol ogi c alvi e wpoi nt ,t hedi s t i nc t i on of ar m anddi s ki sunc l earandt hear r angementofs kel et oni si r r egul ari nt hemember soft heAs t er i ni dae i nc t her mor e,i mmunol ogi c als t udy us i ng ant i body agai ns thexoki nas e ont r as twi t h ot herf ami l i es . Fur (HK)s ugges t ed t hatt hef ami l y As t er i ni daei shet er ogenousl i neage (Moc hi zukiand Hor i ,1980 ) . To c l ar i f yt hephyl ogenet i cr el at i ons hi psamong t hes ef ourf a mi l i es ,Iadopt ed t hef ol l o wi ng f ourt ypi c aland c ommo n s pec i es f r om e ac h f ami l y:As r o m As t er i ni dae, Ac r om t e r i n ap e c t i n i f e r af a n t h a s t e rp l a n c if Ac ant has t er i dae, Ec r om Ec hi nas t er i dae, and S r om Sol as t er i dae h i n a s t e rl u z o n i c u sf o l a s t e rd a ws o n if (Fi g.1) . As s muc hc ommo ns pec i es and one oft he t ypi c als eas hor e ani mal si n t e r i n ap e c t i n i f e r ai J apanes e wa t er s ,and has of t en been us ed i n devel opment albi ol ogy,phys i ol ogy or bi oc he mi s t r y as exper i ment alani mal . The t wo s pec i es (A.p )ar ef ound i ns hal l o w wa t er of l a n c iand E.l u z o n i c u s s ubt r opi c als easofRyukyu I s l ands (Oki nawa)i nJ apan. I ti sf amoust hatA.p st hepoi s onous l a n c ii as t er oi d wi t h ma ny s har ps pi nes and des t r oys t he c or alr eef by eat i ng t he pol yps of c or al s . d body and s hows t he ext ens i ve body c ol or var i at i on.S s Ec h i n a s t e rl u z o n i c u shas har o l a s t e rd o ws o n ii of t en f ound i n nor t her ns eas ofJ apan and i ti s one off er oc i ous as t er oi ds t hatt he s pec i es eat st he as t er oi dsandt heedi bl es c al l opi nMut s uBayofAomor iPr ef .ofnor t he r nJ apan. nt hi spaper ,Ir epor tt he al l ozyme s t udy oft he phyl ogenet i c and t axono mi cr el at i ons hi psa mong I t he f our f a mi l i es (As t er i ni dae,Ac ant has t er i dae,Ec hi nas t er i dae and Sol as t er i dae ) ,and t he mol ec ul ar eval uat i onoft axono mi cs ys t emsofas t er oi ds . Mat er i al sandMet hods The s t ar f i s h,As l ec t ed f r o m As amus hiofMut s u Bay i n Aomor iPr ef .by t e r i n ap e c t i n i f e r a was col s nor ker i ng.S l ec t ed f r om Yoko hamamac hiofMut s u Bay by s t ar mopmet hod o l a s t e rd a ws o n iwas col whi c hf i s her men havebeen us i ng i nt heext er mi nat i on ofas t er oi dst opr ot ec tt hes c al l op. Ac a n t h a s t e r ec ol l ec t ed f r om Ser agakii n Onnamur aofOki nawaPr ef .by s c uba h i n o a s t e rl u z o n i c u swer p l a n c iand Ec di vi ng and s nor ker i ng. I mme di at el y af t er c ol l ec t i on,t he pyl or i cc eac a wer ec utof ff r o m t he l i vi ng s pec i mens and f r ozen i n dr yi c e,and s t or ed at−40℃ unt i l us e. The number of i ndi vi dual s us ed i n al l ozyme anal ys i s was 12 f or A.p or S or A.p or E. e c t i n i f e r a ,10 f .d a ws o n i ,3 f l a n c iand 12 f As A.p s poi s onous and danger ous s t ar f i s h,i twas muc h di f f i c ul tt oc ol l ec tmany l a n c ii l u z o n i c u s . i ndi vi dual sand t hust henumberofs pec i menswass mal l . Al l ozymeanal ys i swasper f or med on 7. 5% pol yac r yl ami degelbyt hemet hoddes c r i bedi nMat s uokaandHat anaka(1991) :About0. 2gpyl or i cc a ec a phat e buf f er c ont ai ni ng 0. 1 M KCland 1mM was i ndi vi dual l y homogeni zed wi t h 3 vol s .of20mM phos EDTA(pH 7. 0)byus i ngPot t er El vehj em t ypeho mogeni zeri nani c ewat erbat h. Thet i s s ueext r ac t wa sc ent r i f ugedat10, 000r pm at5mi nandt hec l ears uper nat antwasus edf orpr ot ei nel ec t r ophor es i s . El ec t r ode buf f er wa s Gl yc i net r i s buf f er ,pH 8. 3. Af t er el ec t r ophor es i s ,al l ozyme var i at i on of t he f ol l o wi ng 11 di f f er ent enzymes was anal ,not hi ng dehydr ogenas e yzed:mal at e dehydr ogenas e (MDH) (NDH) , xant hi ne dehydr ogenas e (XDH) , gl uc os e6phos phat e i s o mer as e(GPI ) , hexoki nas e(HK) , s uper oxi dedi s mut as e(SOD) ,as par t at eami not r as f er as e(AAT) ,al kal i nephos phat as e (ALK) ,per oxi das e (PO) ,es t er as e(EST)and l euc i ne ami no pept i das e(LAP) . Al l ozyme anal ys i s and s t ai nr ec i pes of enzymeswe r eper f or medasdes c r i bedi nMat s uokaandHat anaka(1991) . Res ul t sandDi s cus s i on Fr om al l ozyme var i at i on i n 11 enzymes ,39 genet i cl oc iwer e det ec t ed.Of39 genet i cl oc i ,t he No r i ma s aMATSUOKA 1 0 Ta bl e1.Genet i c i dent i t i es (a bo ve di a go na l )a nd ge ne t i c di s t a nc e ( sbe l ow di agonal )amongf o ura s t e r o i ds . Spec i es 1.As t e r i nape c t i ni f e r a 2.Sol as t e rdaws oni 3.Ac ant has t e rpl anc i 4.Ec hi nas t e rl uz oni c us 1 2 3 4 0 . 9 0 4 0 . 9 5 5 0 . 9 0 4 0 . 4 0 5 0 . 6 6 7 0 . 8 1 6 0 . 3 8 5 0 . 5 1 3 0 . 8 7 5 0 . 4 0 5 0 . 4 4 2 0 . 4 1 7 - Genet i ci dent i t i esand genet i cdi s t a nc e swe r ec a l c ul a t e d by t hemet ho do fNe i (1972) . f ol l owi ng 17l oc iwer epol ymor phi c (Md h 2 ,Hk ,Po 3 ,Po 4 ,S o d 4 ,S o d 6 ,Es t 1 ,Es t 2 ,Es t 3 ,Es t 4 ,Es t 6 , ) . Ther emai ni ng22l oc iwer emonomor phi c . p 6 Al k 4 ,Al k 8 ,Al k 9 ,La p 1 ,La p 2andLa )and To quant i f yt he degr ee ofgenet i c di f f er ent i at i on a mong f ours pec i es ,t he genet i ci dent i t y(I genet i cdi s t anc e (D)bet ween eac hs pec i eswer ec al c ul at ed by t hemet hod ofNei (1972)f r om t heal l el e f r equenc i esdat ai n39genet i cl oc i . Tabl e1s ho wst hemat r i c esofIandD val uesbet weenal lpai r sof s pec i es . The hi ghes tIval ue(I =0. 513)wasf ound bet we en Ac a n t h a s t e rp l a n c iand S o l a s t e rd a ws o n i . Fi gur e1s howst he mol ec ul arphyl ogenet i ct r ee f ort he f ourf ami l i eswhi c h wasc ons t r uc t ed f r o m t he Ne i ’ sgenet i cdi s t anc emat r i xofTabl e1byUPGMA c l us t er i ngmet hodofSneat handSokal (1974) . The mo l ec ul arphyl ogenet i ct r eei ndi c at edt hef ol l owi ngs : (1)Oft he f ourf a mi l i es ,Ac ant has t er i dae and Sol oas t er i dae ar e mos tc l os el yr el at ed t o eac h ot her (D=0. 667) . (2)Ec hi nas t er i dae i smor ec l os el yr el at ed t ot he c l us t erofAc ant has t er i dae and Sol as t er i dae t han As t er i ni dae. (3)The As t er i ni dae i st he mos tdi s t antf ami l y oft he f ourf a mi l i es .The genet i cdi s t anc e bet we en As t er i ni daeandt hec l us t erofot hert hr eef ami l i eswasD=0. 921. The al l ozyme r es ul t sar e we l lc ons i s t entwi t ht he mor phol ogi c alevi denc e:Asevi dentf r o m Fi g.1, (1)A.p (Ac ant has t er i dae)andS (Sol as t er i dae )havemul t i ar ms (10 〜14)i nc ont r as twi t h l a n c i .d a ws o n i f i ve ar msofma ny as t er oi ds . (2)They ar es i mi l art o eac h ot heri n ext er nalmor phol ogy and have l ar gebody s i ze.On t heot herhand,A.p t er i ni dae )and E.l (Ec hi nas t er i dae )have e c t i n i f e r a(As u z o n i c u s s ma l lbody s i ze. (3)Thes pi neson l at er alambul ac r alpl at esar el ong and s har pi nt het wof ami l i es , Ac ant has t er i daeandSol as t er i dae. (4)Thedi s t i nc t i onbet weenar m anddi s ki sunc l eari nA.p e c t i n i f e r a (As t er i ni dae ) ,t hough i n ot hert hr eef ami l i esi ti sc l ear . (5)Thear r ange mentofs kel et alpl at es i s r egul ar i n A.p t er i ni dae), but t he ot her t hr ee s pec i es have i r r egul arar r ange ment . e c t i n i f e r a(As The mor phol ogi c al e vi denc e des c r i bed above i s wel lc ons i s t ent wi t h t he pr es ent mol ec ul ar phyl ogenet i cs t udy. Iwoul dl i ke t oc o mpar et he pr es ental l ozyme r es ul t st ot axono mi cs ys t e ms . I ns pi t e ofmany mo r phol ogi c als t udi es ,t axonomy ofas t er oi dsr e mai nst obeext r e mel yc ont r over s i aland s ever aldi f f er ent t axono mi cs ys t e ms ar e pr opos ed.Hayas hi (1940)and Spenc er and Wr i ght (1966)c l as s i f i ed t he f our f ami l i es (As t er i ni dae,Ac ant has t er i daeSol as t er i daeandEc hi nas t er i dae )i nt ot heor derSpi nul os i da. On t heot herhand,Cl ar kandDowney(1992)pr opos edne wt axonomi cs ys t e m andes t abl i s hedmanyor der s . Ac c or di ngt ot hei rt axono mi cs ys t em,t hef ourf ami l i ess t udi edher ear ec l as s i f i edi nt ot hef ol l o wi ngt hr ee di f f er entor der s : As t er i ni dae, Ac ant has t er i dae: Val vat i da Ec hi nas t er i dae: Spi nol os i da Sol as t er i dae: Ve l at i da The pr es ental l ozyme s t udy di d nots uppor tt he t axono mi cs ys t e m ofCl ar k and Downey(199 2) Bi oc hemi c a ls ys t emat i c so fa s t e r o i ds 11 Fi g.1. Fourt ypi c als pec i esoff ourf ami l i e so fAs t e r o i de aa do pt e di nt hepr e s e nta l l o z ymes t udy. 1=As t e r i nape c t i ni f e r a,2=Ec hi noas t e rl u z o n i c u s ,3 =Ac a n t h a s t e rp l a n c i ,4 =S o l a s t e rd a ws o n i . whi c hi swi del yadopt edbymanywor ker s . Thepr es entaut horc ons i der st hatt heyes t abl i s hedma ny t axono mi c gr oups (or der s ) ,and t hus t hat i ti sc ompl i c at ed t axono mi cs ys t e m. The phyl ogenet i c r el at i ons hi psa mong f ourf a mi l i es (Fi g.2)and genet i cdi s t anc edat a (Tabl e1)s ho wi ng t hatt hegenet i c di s t anc esamo ng f ourf ami l i esar ec ompar abl et ot hos ebet ween di f f er entf a mi l i esorgener aoft hes ame or deri nmanyot herani mal s (Thor pe,1980)s uppor tt het axono mi cs ys t e m pr opos edbyHayas hi (1940) andSpenc erandWr i ght (1966) . Fer gus on(1980)r epor t ed t hat t he al l ozyme anal ys i si s muc h ef f ec t i ve i n t he di s t i nc t i on of mo r phol ogi c al l y ver ys i mi l ar s pec i es (s i bl i ng s pec i es )or i nt he es t i mat i on ofgenet i cr el at i ons hi ps at s pec i esl evel ,andal s ot hati ti sef f ec t i vef orel uc i dat i ngr el at i ons hi psa mongdi f f er entgener aandf a mi l i es epor t ed t heal l ozymes t udy on t hedi s t i nc t i on off ours i bl i ng s pec i es athi ghert axonomi cr anks . Wer oft he ec hi noi d,Ec (Mat s uoka and Hat anaka,1991 )or oft wo s i bl i ng s pec i es oft he h i n o me t r a ma t h a e i ec hi oi d,As (Mat s uoka e Al t hough t he mol ec ul ar phyl ogenet i cs t udy of t h e n o s o ma i j i ma i ta l . ,2004). di f f er entf a mi l i esi st hef i r s tat t empt ,i twoul dpr ovi des o meus ef uli nf or mat i ont oas t er oi dt axono my. I ti si nt er es t i ng t oc ompar et he pr es ental l ozyme s t udy wi t h ot hermol ec ul arphyl ogenet i cs t udi es . Moc hi zukiandHor i (1980)exami nedt hephyl ogenet i cr el at i ons hi psa mong22as t er oi ds pec i esbel ongi ng t o di f f er entor der s by t he i mmunol ogi c alt ec hni que oft he enzyme i nhi bi t i on met hod us i ng ant i body agai ns thexoki nas epur i f i ed f r om As . Thei mmunol ogi c almet hod wasal s ous ed i nt he t e r i n ap e c t i n i f e r a phyl ogenet i cs t udy of ec hi noi ds by t he pr es ent aut hor and pr ovi ded s o me us ef uli nf or mat i on i nt o ec hi noi d phyl ogeny(Mat s uoka,1986) . Moc hi zukiand Hor i (1980)f ound t hatAs t e r i n ap e c t i n i f e r a of t hef ami l y As t er i ni daewasc ons i der abl edi f f er entl i neagef r o m ot heras t er oi ds ,and t hey s ugges t ed t hat ongs t o di ver gentl i neage f r om ot her as t er oi ds . The pr es ental l ozyme s t udy As t e r i n ap e c t i n i f e r a bel s ugges t i ng t hehet er ogeneousl i neageo fAs t er i ni daei swel lc ons i s t entwi t ht hei mmunol ogi c alr es ul t sof i (1980) . Mor e r ec ent l y, Mat s ubar a e (2004)r epor t ed t he mol ec ul ar Moc hi zuki and Hor ta l . phyl ogenet i cs t udy ofvar i ousas t er oi dsby nuc l earand mi t oc hondr i alDNA s equenc e anal ys i s . They No r i ma s aMATSUOKA 1 2 Fi g.2. Mol ec ul ar phyl ogene t i ct r e eo ff o ur f ami l i e so ft he As t e r o i de a bas ed on al l ozyme a na l ys i s . I t was c o ns t r uc t e d f r om Ne i ’ s genet i cdi s t anc esby t heUPGMA c l us t e r i ng met ho d. adopt ed t hr eemol ec ul arpar amet er soft henuc l ear18S r DNA,mi t oc hondr i al12S r DNA and 16S r DNA. As a r es ul t ,t hey s ugges t ed t he c l os er el at i ons hi p bet ween genus As a mi l y Sol as t er i dae t e r i n a and f bel ongi ngt ot hedi f f er entor der s . Asevi dentf r o m Fi g.2,t hepr es ental l ozymes t udywasi nc ons i s t ent wi t ht hei rmol ec ul arphyl ogenet i cs t udy. I nc onc l us i on,t hepr es ental l ozymes t udys uppor t st heol dt axono mi cs ys t em byHayas hi (1940)and Spenc erandWr i ght (1966)whi c hdi dnotes t abl i s hmanyt axono mi cgr oups . Ack nowl edgement s Iam gr at ef ult ot hef ol l owi ngwor ker s:Dr .Shi r as aki ,S. ,Oki na waPr ef . ,Dr .Shi nyas hi ki ,F. ,Oki nawa I nt er nat i onalUni ver s i t y,t he s t af fofYokohamamac hiFi s her men’ s Uni on,Aomo r iPr ef . ,t he s t af fof As amus hiMar i neSt at i on,TohokuUni ver s i t yandMat s uoka,K. ,Shi gaPr ef .f ort hei rhel pi nt hi ss t udy. Abs t r act The phyl ogenet i cr el at i ons hi psamong f ourf ami l i es (As t er i ni dae,Ac ant has t er i dae,Sol as t er i dae and Ec hi nas t er i dae )oft heAs t er oi deawer ei nves t i gat edbyal l ozymeanal ys i sof11di f f er entenzymes . As t ypi c als pec i esf r om eac hf ami l y,t he f ol l owi ng f ourc o mmo ns pec i eswer e adopt ed:As t e r i n ap e c t i n i f e r a f r om As t er i ni dae,Ac r om Ac ant hat er i dae,S r om Sol as t er i dae,and h a n t h a s t e rp l a n c if o l a s t e rd a ws o n if r o m Ec hi nas t er i dae. Fr om al l ozymevar i at i oni n11enzymes ,39genet i cl oc iwer e Ec h i n a s t e rl u z o n i c u sf det ec t ed and Nei ’ sgenet i cdi s t anc esamong f ourf a mi l i eswer ec al c ul at ed f r o m al l el ef r equenc i esdat ai n 39 genet i cl oc i .The mol ec ul ar phyl ogenet i ct r ee f or f our f ami l i es i ndi c at ed t hatAc ant has t er i dae and Sol as t er i dae ar e mos tc l os el yr el at ed t o eac h ot her ,and t hatAs t er i ni dae i smos tdi s t antf ami l y. The al l ozyme r es ul t sar e we l lc ons i s t entwi t h mo r phol ogi c alevi denc e. Fur t her mor e,t he genet i c di s t anc e val uesamong f ourf a mi l i eswer ec o mpar abl et ot hos ebet ween di f f er entf a mi l i esorgener aoft hes a me or deri not herani mal s . Thepr es ental l ozymes t udys uppor t st het axono mi cs ys t e m ofHayas hi (1940 ) and Spenc erand Wr i gh (1 t 964)t hatc l as s i f i ed t hes ef ourf a mi l i esi nt o one s a me or derSpi nul os i da,but Bi oc hemi c a ls ys t emat i c so fa s t e r o i ds 13 wer ei nc ons i s t entwi t ht hatofCl ar k and Downey(1992)whi c h es t abl i s hed ma ny or der s .The pr es ent al l ozymer es ul ts ugges t i ngt hatAs t er i ni daemi ghtbehet er ogeneousl i neagewaswel lc ons i s t entwi t ht he i mmunol ogi c als t udybyMoc hi zukiandHor i (1977) . Ref er ences t ar f i s he soft h eAt l a n t i c .ChapmanandHal CLARK,A. M.andDOWNEY,M. E. (1992)S l ,Lo ndo n. CHINO,Y. ,SAITO,M. ,YAMASU,K. ,SUYEMITSU,T.andI (1 9 9 4 )Fo r ma t i o no ft hea dul tr udi me nto fs e aur c hi nsi s SHI HARA,K. i nf l uenc edbyt hyr oi dhor mones .De v.Bi ol . ,161:1 1 1 . oc he mi c als y s t e mat i c sandEv o l u t i o n .Bl FERGUSON,A. (1980)Bi a c ki eGl a s gow. HAYASHI,R. (1940)Cont r i but i onst ot hec l as s i f i c at i o no ft hes e a s t a r so fJ a pa n.I .Spi nul o s a .J .Fa c .Sc i .Ho kka i doUni v.Ser . VI ,7:107204. HORI,S. H. ,TANAHASHI,K.andMATSUOKA,N( .1977)Mor pho l o gi c a la ndc yt o c hemi c a ls t udi e so nt hes e c r e t o r ygr a nul esoft he t e r i asamur e ns i s .Bi pyl or i cc eac aoft hes t ar f i s h,As o l .Bul l . ,1 5 2:6 4 7 4 . t e r i n a and Sol MATSUBARA,M. ,KOMATSU,M.and WADA,H. (200 4 )Cl o s er e l a t i o ns hi p be t we e n As a s t e r i da e(As t e r oi dea) s uppor t edbybot hnuc l earandmi t oc hondr i alge nemol e c ul a rphyl o ge ni e s .Zo o l .Sc i . ,2 1:7 8 5 7 9 3 . MATSUOKA,N. (1986)Fur t her i mmunol ogi c als t udy o nt he phyl o ge ne t i cr e l a t i o ns hi ps amo ng s e a ur c hi ns o ft he or der Ec hi noi da.Co mp.Bi oc hem.Phys i o l . ,84B :46546 8 . e u d o c e n t r o t u sd e p r e s s u s .Zool MATSUOKA,N. (1987)Bi oc hemi c als t udyont het axo nomi cs i t ua t i o no ft hes e a ur c hi n,Ps .Sc i . ,4 :339347. t e r i na MATSUOKA,N.and ASANO.H. (1999)Mol ec ul art a xo nomi cs t udy o nt he va r i a t i o no fbo dy c o l o ri nt he s t a r f i s h As pe c t i ni f e r a.Bul l .Fac .Agr i c .& Li f eSc i .Hi r os akiUni v. ,No. 1:1 8 . t e r i n ap e c t i ni f e r a. MATSUOKA,N.andASANO,H. (2003)Genet i cvar i at i o ni nno r t he r nJ a pa ne s epo pul a t i o nso ft hes t a r f i s hAs Zool .Sc i . ,20:985988. MATSUOKA,N. ,FUKUDA,K. ,YOSHIDA,K. ,SUGAWARA,M.a ndI ,M. (1 9 9 4 )Bi o c hemi c a ls ys t ema t i c so ff i vea s t er oi dsof NAMORI t hef a mi l yAs t er i i daebas edonal l ozymevar i at i o n.Zo o l .Sc i . ,1 1:3 4 3 3 4 9 . MATSUOKA,N.andHATANAKA,T. (1991)Mol ec ul are vi de nc ef o rt hee xi s t e nc eo ff o urs i bl i ngs pe c i e swi t hi nt hes e aur c hi n, Ec hi nome t r amat hae i ,f r om J apanes ewat er s .Zo o l .Sc i . ,8:1 2 1 1 3 3 . t e r i as MATSUOKA,N.and HATANAKA,T. (1998)Genet i cdi f f e r e nt i a t i o n among l o c a lJ a pa ne s epo pul a t i o nso ft hes t a r f i s h As amur e ns i si nf er r edf r om al l ozymevar i at i on.Gene sGe ne t .Sys t . ,7 3:5 9 6 4 . MATSUOKA,N.andHORI,S. H. (1980)I mmunol ogi c alr e l a t e dne s so fhe xo s e 6 pho pha t ede hydr o ge na s ea ndgl uc o s e 6 phos pat e dehydr ogenas ei nec hi noder ms .Comp.Bi oc hem.Phys i o l . ,6 5B :1 9 1 1 9 8 . MATSUOKA,N. ,KOHYAMA,K. ,ARAKAWA,E.and AMEMIYA,S. (2 0 0 4)Mol e c ul a re vi de nc ef o rt he e xi s t e nc eo ft wo s i bl i ng t he n o s o mai j i ma if s pec i eswi t hi nt heec hi not hur i oi dec hi noi dAs r om J a pa ne s ewat e r s .Zo o l .Sc i . ,2 1:1 0 5 7 1 0 6 1 . MATSUOKA,N.andSUZUKI,H. (1989)El ec t r ophor et i cs t udyo nt hephyl o ge ne t i cr e l a t i o ns hi psamo ngs i xs pe c i e so ft hes eaur c hi nsoft hef a mi l yEc hi nome t r i daef oundi nJ a pa ne s ewa t e r s .Zo o l .Sc i . ,6:5 8 9 5 9 8 . n v i t at i ont oMol e c ul arEv ol ut i on.Bl MIYATA,T. (1994)I ueBa c ks ,Ko da ns ha ,To kyo (i nJ a pa ne s e ) . MOCHIZUKI,Y.and HORI,S. H. (1980)I mmunol ogi c a lr e l a t i o ns hi pso fs t a r f i s h he xo ki na s e s :Phyl o ge ne t i ci mpl i c a t i o n.Comp. Bi oc he m.Phys i ol . ,65B :119125. NEI,M. (1972)Genet i cdi s t anc ebet weenpopul at i ons .Am.Na t . ,1 0 6:2 8 3 2 9 2 . l e c ul arEv ol ut i onar yGe ne t i c s .Col NEI,M. (1987)Mo umbi aUni vPr e s s ,Ne w Yo r k. SAITO,M. ,SEKI,M. ,AMEMIYA,S. ,YAMASU,K. ,SUYEMITSU,T.a ndI K. (1 9 9 8)I nduc t i o no fme t amor pho s i si nt hes and SHI HARA, e r one l l aj aponi c abyt dol l arP hyr oi dhor mones .De ve l o p.Gr owt hDi f f e r . ,4 0:3 0 7 3 1 2 . me r i c alT a x o n o my .Fr SNEATH,P. H. A.andSOKAL,P. R. (1973)Nu e eman,Sa nFr a nc i s c o . r e a t i s eo nI n v e r t e b r a t ePa l e o n t o l o g y SPENCER,W. K.andWRIGHT,C. W. (1966)As t er ozoans .I nT (Edi t e dbyMo o r e ,R. C. ) ,Par t U,pp.4107,Geol .Soc .Am & Uni v.Ka ns asPr e s s ,Ne w Yo r k. THORPE,J . P (1 . 982)Themol ec ul arc l oc khypot hes i s :Bi o c hemi c a le vo l ut i o n,ge ne t i cdi f f e r e nt i a t i o n,a nds ys t emat i c s .Ann.Rev. Ec ol .Sys t . ,13:139 169. 1 4 No r i ma s aMATSUOKA アロザイム変異からみたヒトデ類4科の系統類縁関係 松 岡 教 理 弘前大学農学生命科学部分子進化学研究室 棘皮動物ヒトデ類の系統分類学は議論が多く,研究者 である,(3 )イトマキヒトデ科は系統的に最も遠く,イ 間で見解の相違が大きい。特に上位の分類ランク(科・ トマキヒトデ科の系統的異質性が示唆される, (4 )4科 目)間の系統関係はほとんど解明されていないのが現状 間の遺伝的距離は,他動物の同目の別科・別属間で観察 である。本研究では,有棘目に属するヒトデ類 4科(イ される数値と同程度である。これらの結果は,ヒトデ類 トマキヒトデ科,オニヒトデ科,ニチリンヒトデ科,ル を多くの目に細分類し,本研究で調べた 4科を 3目に分 ソンヒトデ科)の系統類縁関係をアロザイム分析により 類して,イトマキヒトデ科とオニヒトデ科が近縁である 調査した。4科の代表種として,それぞれの科より普通 と主張した Cl a r ka ndDo wne y(1 9 9 2 )の分類体系とは 種であるイトマキヒトデ,オニヒトデ,ドーソンニチリ 全く一致せず,4科を系統的に近縁なグループと考えて ンヒトデ,ルソンヒトデを用いた。1 1酵素のアロザイム 1つの同じ目(有棘目:Spi nul o s i da )に分類した Ha ya s hi 分析から 3 9酵素遺伝子座が検出された。各酵素遺伝子 9 4 0 )や Spe nc e ra ndWr i ght (1 9 6 6 )の分類体系を強 (1 座における対立遺伝子頻度から,Ne i (1 9 7 2 )の方法に く支持する。またイトマキヒトデ科の系統的異質性を示 より 4種間の遺伝的距離を算出し,UPGMA法を用いて 唆した今回の結果は,Mo c hi z ukia ndHo r i (1 9 8 0 )の糖 4科の分子系統樹を作成した。その結果, (1 )多腕の体 代謝酵素ヘキソキナーゼの抗体を用いた酵素活性阻害法 型を持つオニヒトデ科とニチリンヒトデ科が最も近縁で による免疫学的研究ともよく一致する。 ある, (2 )このクラスターに近縁な科はルソンヒトデ科 弘大農生報 No.9:8 -14,2006 15 マナマコ複合盤状体骨片の成長と体長との関係 吉田 渉・大中臣哲也・石田幸子 弘前大学農学生命科学部遺伝情報科学講座 (2 0 0 6年 1 0月 1 2日受付) な骨片として釦状体骨片(b),網状体骨片(n)が確 緒 言 認された。鈎状骨片(h)はまれに含まれていた。 近年,ナマコの種苗生産が各地で行われ,ナマコの成 2 .触手 t e nt a c l e (Fi g.1 -2 ) 長速度と体長・体重との関係,年齢査定といった調査の 触手には棒状体骨片のみが存在していた。形状は弓 必要性が生じてきている。そこで,ナマコの骨片がそれ 状で彎曲している。触手に存在する棒状体骨片の大き らの指標となりうるのかを調査した。 さは様々である。骨片の周囲にみられる多数の小さい マナマコ骨片の形態と種類については,体の部位によ 突起が特徴的で,体壁の鈎状体骨片とは区別できる。 り触手骨片,水肺(呼吸樹)骨片,体背部骨片,体腹部 小さいものは突起が未発達である。アカナマコにおけ 骨片に類別され,触手骨片は一種,体背部骨片は 6種, 体腹部骨片は 5種で構成されていることが報告されてい る 1)。また形態的特徴をもとに,櫓状体骨片,棒状体骨 片,網状体骨片,釦状体骨片(楯状体骨片) ,鈎状体骨片, 1 ) る突起の発達はアオナマコに比べて顕著であった。 3 .呼吸樹 r e s pi r a t o r yt r e e (Fi g.1 -3 ) 骨片数は少ない。網状体骨片を有するが,体壁のそ れとは異なり,周囲が開口していて細い。形状は不定 針状体骨片,複合盤状体骨片に分類されている 。今回 型である。また,紐状,V字状の骨片も存在していた 我々は,様々な体長のマナマコから骨片を単離し,各部 (数は少ない)。 位の骨片の形態観察を行うと共に形態的特徴や数,大き 4 .管足 t ubef o o t (Fi g.1 -4 ) さを比較した。またマナマコ管足から単離した複合盤状 一管足当たり,一個の複合盤状体骨片が存在してい 体骨片の微細構造及びその成長過程についての知見が得 た。ほぼ円に近い形状を有する。また管足を形成して られたので報告する。 いる管には櫓状体骨片が確認された。 5 .消化器官 di ge s t i veo r ga n(Fi g.1 -5 ) 材 料 と 方 法 骨片数は少ない。網状体骨片とは異なる樹状骨片 (t r )が存在する。樹状骨片の形状は不定であり,樹状 陸奥湾から採集された体長の異なるマナマコ(アオナ の突起を有するのがこの骨片の特徴である。また,紐 マコ・アカナマコ)の各部位(触手,体壁,疣,呼吸樹, 状骨片も見られた。 消化管,管足)を,蒸留水で 5倍に希釈した漂白剤(ハ イターR)中に一晩つけ置きし,沈んだ残渣を骨片分画と 今回の各部位における調査結果から,管足と消化管に した。これを蒸留水で 2回洗い,観察に供した。骨片の 存在する骨片とその形状,種類が新たに明らかとなっ サイズは接眼,対物マイクロメーターを用いて測定し た。既に報告のある体腹部骨片のうち複合盤状体骨片 1) た。複合盤状体骨片の微細構造の観察試料には,体長と は管足のみに存在し,一つの管足の吸盤内に複合盤状体 骨片サイズの相関を求めるのに使用した骨片を用いた。 骨片が一個だけ存在することを明らかにした。また,消 これらの乾燥骨片はカーボンペースト試料台上で金コー 化器官においては樹状骨片が存在することを示した。ア ティングされ,走査型電子顕微鏡(J SM5 3 0 0 )により検 オナマコの櫓状体骨片においては,体長の大きい個体の 鏡された。 方が,櫓上部の突起部が顕著であるという傾向が認めら れた。同一殻重(消化管,水肺および体腔水を除いたカ 結 果 と 考 察 (1 )各部位における骨片の種類 1 .体壁 bo dywa l l (疣 pa pi l l a ) (Fi g.1 -1 ) 櫓状体骨片(t )が多数を占める。その他に,特徴的 弘大農生報 No.9:15 -20,2006 ラだけの重さ)のものを比較した場合,アオナマコの体 背部櫓状体骨片の底盤はアカナマコのそれより大きいと の報告がある 1)。また,殻重の増大とともに櫓状体骨片 の底盤長短径は減少する傾向を示し 1),その減少幅は殻 重1 0倍当たり 1 0 μm であると解釈されうる。崔と大島 1 6 吉田・大中臣・石田 Fi g.1. Theos s i c l esf r om t hevar i ouspar t sofA.j a p o n i c u s . 1:t abl e(t ) ,hookl i ke(h) ,but t on(b(s ) hi el dl i ke)a nd ne t l i ke (n)o s s i c l e sf r om t he pa pi l l a ,2 :s uppo r t i ng r o ds f r om t ent ac l es ,3:r od and net l i keos s i c l esf r om t her e s pi r a t o r yt r e e ,4 :c ompl e x pl a t e sf r om t het ubef e e t ,5 :t r e e l i ke(t r ) and r od os s i c l esf r o mt hedi ges t i veor gan. マナマコ複合盤状体骨片の成長と体長との関係 17 Fi g.2. Thec or r el at i on bet ween body l e ngt ha nd di amet e ro ft hec ompl e x pl a t eo s s i c l e s . c l os ed c i r c l es (n=88) :os s i c l esofgr e e nc o l o rt ype s pe c i me ns ,o pe nc i r c l e s (n=5 3 ) :o s s i c l e so f r edc ol or t ype s pec i mens ,x:di ame t e r (mm)ofa c ompl e x pl a t eo s s i c l ei n at ube f o o t ,y: body l engt h(c m)ofl i vi ng s pec i mens . の報告 1)はアオナマコとアカナマコにおける殻重当たり その有用性が示されている 5)。我々は現場での利便性を の櫓状体骨片の底盤の大きさの比較であるが,今回我々 考慮し,ナマコの体長には伸長時の体長(最大長)を採 が行った櫓状体骨片の大きさと体長に関しては,その関 用し,管足の複合盤状体骨片と体長との関係を調べた。 係を見い出すことはできなかった。本実験に用いたナマ 体長の異なる個体から無作為に管足を切り取り,複合盤 コは便宜的にアオナマコ,アカナマコと記載している 状体骨片を得た。測定の結果,体長の大きい個体の複合 が,形態的差異(色素の着色度合い)からは明確にアカ 盤状体骨片は大きい傾向にあった。複合盤状体骨片は円 ナマコとは特定できず,アオナマコとアカナマコの中間 に近い円盤状をしているため,最大直径を計測すること 型とも考えられる。マナマコは生態上の特徴からも区別 で骨片の大きさとした。測定した骨片の大きさは,アオ され,アカナマコは砂礫層/沖合いに生息する一方,ア ナマコ体長 2 5 c mで直径平均 0 . 9 5 mm(0 . 8 6−1 . 0 8 mm) , 2 ) オナマコは沿岸/砂泥に生息する 。また,Ka nno& 3 ) 体長 9 . 5 c m で平均 0 . 6 4 mm(0 . 5−0 . 7 6 mm)で,それぞ Ki j i ma のサンプリングデータからは陸奥湾産マナマ れのレンジは 0 . 2 2 mm,0 . 2 6 mm であった。アカナマコ コの全てがアオナマコあるいはクロナマコとしている。 9 . 3 c m においては,平均 0 . 6 1 mm(0 . 4 8−0 . 7 mm)であっ た。骨片直径と最大長の関係は,Fi g.2に示すような分 9 5 4 . 6 1 1 x1. 布が認められ,べき乗近似式 y= 2 (アオナマコ (2 )管足骨片サイズと体長との相関 ナマコの体長は,ナマコそのものが持つ伸長・収縮性 9 8 6 2 +アカナマコ),y= 2 5 . 2 6 7 x1. (アオナマコ)の関係式 によりかなり変化する。ナマコの体長測定基準はもっと を得ることができた。xは体長(c m),yは複合盤状体骨 も長くのびたサイズ最大長,もっとも短く縮んだ最短 片直径(mm)。いずれの場合に於いても,相関係数 R2= 長,自然長などがある。また,計測誤差もあるため,殻 0 . 9 4 3 9 ,0 . 9 6 0 3となり,体長は複合盤状体骨片の直径(半 重(前述)が適しているとの報告 4 ) もある。ナマコの麻 径)と高い相関がある。骨片直径に対する体長はアカナ 酔には KClやエタノールを用いてきたが,近年メントー マコとアオナマコを比較した場合,若干ではあるがアカ ルを用いた稚ナマコの体長測定用麻酔剤の報告があり, ナマコが大きい。これは,アカナマコ特有の伸長性が考 1 8 吉田・大中臣・石田 Fi g.3A.Ul t r as t r uc t ur eoft hec ompl ex pl at eos s i c l e si n A.j a p o n i c u s . Thet woos s i c l es (No . 1&4)wer ec ol l ec t ed f r om t het ubef e e to fagr e e nc o l o rt ypes pe c i men(bo dy l e ngt h,2 . 4 c m) .1: dor s alvi e w,2:magni f i c at i on ofa c ent r alr egi on i n Fi g.3A1 (bl a c k bo x) ,3 :ma gni f i c a t i o no fa mar gi na lr e gi on i n Fi g.3 A1 (whi t ebox) .4:vent r alvi ew,5,6:magni f i c a t i o no fFi g.3A4 (5 :bl a c k bo x,6 :whi t ebo x) . えられる。骨片直径からナマコの体長を推定する場合, 9 ,1 0 )。Fi g. 3 B 9 ,1 0の 上 側 は 管 足 水 腔(po di a l 体色(クロ,アカ,アオ)による伸張性を考慮する必要 hydr o c o e l )に通じていて,下側は接地面で,吸着できる があると考えられる。 構造になっている。体長 2 . 4 c m のナマコの複合盤状体 管足の複合盤状体骨片は櫓状体骨片に比べて,数は圧 骨片(Fi g.3 A)は多数の 1 0 μm内外の円状の間隙を有し, 倒的に少ない(実測:アオナマコ 9 . 5 c m;総数 1 6 1個, 中央部(1 5 0 μm)並びに外縁部(8 0 μm)からなる。中 アカナマコ 9 . 3 c m;総数 2 1 0個) 。また,その形状は比較 央部と外縁部の境界部は単純な枝分かれ構造をとってい 的単純である。さらに櫓状体骨片底盤より体長(殼重) る。体長 9 . 3 c m または 9 . 5 c m(Fi g.3 B 7〜 1 5 )の骨片 に対する骨片サイズの差異が顕著なことから測定作業が では中央部と外縁部の境界領域(境界部)は不明瞭とな 容易である。ナマコの成長に伴い,管足並びに管足内の るが,中央部と外縁部は単層構造を持つものとして識別 複合盤状体骨片も成長すると考察され,これらの関係式 できる。境界部はかなり大きく 1 0 0 μm にも達し,複雑 は,年齢査定や骨片直径からの体長推定などの指標とし に枝分かれした多層構造をもつ。体長 2 5 c m(Fi g.3 B て利用できると考えられる。 1 6〜 1 8 )になると中央部において明瞭な単層構造は見ら れなくなり,中央部と境界部は共に多層構造になってい (3 )管足骨片の微細構造 る。また,外縁部においても多層構造が見られる領域が ナマコ類の骨片は炭酸カルシウムの結晶からなり,骨 存在する。 片形成は体壁の表皮層に位置する s c l e r o c yt eの多核シン 以上の結果から,複合盤状体骨片は微細構造的に中央 シチウムの中で起ると報告されている 6)。しかし,複合 部,境界部,外縁部から成り立っていることが判明した。 盤状体骨片がどのように成長するかは知られていない。 境界部は骨片と管足水腔が結合する領域にあたると推測 そこで体長の異なる個体から採取した複合盤状体骨片の される。骨片の成長は管の成長(管径の増大)と共に境 微細構造(Fi g.3 A,B)を観察することにより,これら 界部から始まり,次第に径を増すにつれて外縁部が外側 の骨片の成長過程を推測した。 に移行していき,骨片が更に成長する過程で,中央部や マナマコ(アオナマコ,アカナマコ)の複合盤状体骨 外縁部においても縦方向の成長が進み,複雑な層構造に 片は,側面からみると片凸レンズ状をしている(Fi g.3 B 至ると考えられる。 マナマコ複合盤状体骨片の成長と体長との関係 19 Fi g.3B.Ul t r as t r uc t ur eoft hec ompl ex pl at eos s i c l e si n A.j a p o n i c u s . Theos s i c l esofNo . 79wer ei s ol at ed f r om agr een c o l o rt ype (bo dy l e ngt h,9 . 5 c m)and e a c ho fNo . 1 0 1 2wasf r om r ed t ype s pec i me n(body l engt h,9. 3c m) .The os s i c l eo fNo . 1 6 was c o l l e c t e df r om a gr e e nt ype s pe c i men(bo dy l engt h, 2 5c m) .7:dor s alvi ew,8:vent r alvi ew,9,10:l at e r a lvi ew,t he ups i de i spo di a lhydr o c o e l .1 1 :do r s a lvi ew,1 2 :vent r al vi ew,13:ma gni f i c at i on ofac ent r alr egi on i n Fi g.3 B8 (bl a c k bo x) ,1 4 :ma gni f i c a t i o no fabo unda r yr e gi o ni n Fi g.3B8 (gr ay box) ,t he boundar yr egi on wasi ndi c at ed by t he s a ndwi c he dl i ne s .1 5 :ma gni f i c a t i o no fa bo unda r yr e gi on i n Fi g. 3B11(gr ay box) , 16: dor s al vi ew, 17: ma gni f i c a t i o n of a c e nt r a lr e gi o n i n Fi g. 3 B1 6 (bl a c k bo x) ,1 8: ma gni f i c at i on ofama r gi nalr egi on i n Fi g.3B16(whi t ebo x) . 引 用 文 献 1.崔 相・大島泰雄:ナマコにみられる「アオ」と「アカ」 の形態及び生態的差異について.Bul l et i n ofJ a pa ne s e So c i e t yo fSc i e nt i f i cFi s he r i e s .2 7 (2) ,9 7 1 0 6 .1 9 61. 2.西村三郎:原色検索日本海岸動物図鑑 2,大阪,保育社, 1 9 9 5 . 3.KANnd KIJIMA, A. : Ge ne t i c di f f e r ent i at i on NO, M. a 吉田・大中臣・石田 2 0 a mong t hr ee c ol or var i ant s o f J apanes e s e a c uc umber St i c hopus j aponi c us .FI SHERI ES SCI ENCE. 69:806812.2003. 4 .崔 相:ナマコの研究,東京,海文堂 pp.3241 .1 9 6 3 . 5.畑中宏之・谷村健一:稚ナマコの体長測定用麻酔剤とし て の me (2 2 )2 2 1 nt holの 利 用 に つ い て.水 産 増 殖 4 225.1994. 6.STRICKER,S. A. :Thef i nes t r uc t ur eand devel opme nto f c al c i f i ed s kel et al e l ement s i n t he body wal lo f Ho l ot hur i an Ec hi node r ms .J .Mor phol ogy.188 :2 7 3 288.1986. 年度弘前大学学長指定重点研究費の助成を受けて行われ ました。 摘 要 マナマコ骨片を調査した結果,体腹部骨片のうちの複 合盤状体骨片は管足の吸盤に位置し,1管足当たり 1個 存在していること,管足の管壁には櫓状体骨片が存在し ていることが判明した。また,単離した管足骨片のう ち,複合盤状体骨片サイズと体長には相関関係が認めら 謝 辞 れた。我々は大きさの異なる複合盤状体骨片の微細構造 観察から,中央部,境界部,外縁部の 3部位から成り立っ マナマコの提供に御協力下さいました青森市産業部水 ていることを明らかにし,骨片の成長は管足管径の増大 産業課水産指導センター所長,長谷川清治氏並びに泉慎 と共に境界部から始まり,複雑な層構造を呈するに至る 也技師に深く感謝致します。この研究の一部は,平成 1 8 と考察した。 Re l a t i o nbe t we e nt hegr o wt ho fc o mpl expl at eos s i c l eandt hebodyl engt h o fs e ac uc umberApos t i c hopusj aponi c us Wat a r uYOSHI Tet s uyaÔNAKATOMIa ndSa c hi koI DA, SHI DA De p a r t me n to fBi o f u n c t i o n a lS c i e n c e ,Fa c u l t yo fAg r i c u l t u r ea n dLi f e S c i e n c e ,Hi r o s a k iUn i v e r s i t y ,Hi r o s a k i0 3 6 8 5 6 1 ,J a p a n As t he r es ul tofi nves t i gat i ng os s i c l es f r om Apos t i c hopusj aponi c us (Sel enka,1867 ) ,i twas c l ar i f i ed t hatt he s i ngl ec ompl ex pl at e os s i c l ei sl oc at ed i nt he s uc kerofone t ube f oot ,and al s oal otoft abl e os s i c l esexi s t ed i nt he t ube wal l .Fr om anal yzi ng t he s i ze ofos s i c l esi n var i ousbody l engt hsofl i vi ng s pec i mens ,t hec or r el at i on wasr ec ogni zed bet ween t hedi a met erofc o mpl ex pl at eos s i c l esand t hebody l engt h.I twasr eveal edt hatt hec ompl expl at eos s i c l ec ons i s t soft hr eer egi ons ,t hec ent r al ,t heboundar y and t he mar gi nal ,by s c anni ng el ec t r on mi c r os c opy.We c ons i der ed t hatt he gr o wt h ofc o mpl ex pl at e os s i c l es t ar t sf r om t he boundar yr egi on ac c or di ng t ot he gr o wt h oft he t ube f oot ,f i nal l yt he os s i c l e gr owsi nt ot hec o mpl exl ayers t r uc t ur e. 榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎Bul l .Fac .Agr i c .& Li f eSc i .Hi r os akiUni v.No.9:15 -2 0,2006 21 リンゴ搾り粕のメタン発酵 メタン発酵汚泥から分離したペクチン・キシラン分解菌の特徴 武田 潔・須田 育江・殿内 暁夫 応用生命工学科応用微生物学研究室 (2 0 0 6年 1 0月 1 2日受付) 緒 言 実験材料と方法 青森県は年間約 4 8万トンのリンゴを生産しているが, 1.リンゴ搾り粕試料 そのうち約 1 0万トンのリンゴがジュース用として利用 日本果実加工(株)(弘前市)のリンゴ搾り粕を使用し されている。リンゴジュースの製造過程で生成する搾り た。乾燥重量は試料 1 gを秤量瓶に入れ,1 0 5 ℃ で恒量 粕は約 3万トンであるが,その一部は肥料や家畜飼料に になるまで乾燥させ,計量した。 利用されているが大部分は廃棄処分されている。現在, 食,医薬,エネルギー等の資源としてリンゴ搾り粕を利 2.メタン発酵汚泥の製造 用する研究開発が進められている。リンゴ搾り粕は水分 メタン発酵リアクターは円筒状容量 3 L(直径 8 0 mm 含量が多いので廃棄物処理法としてはメタンと炭酸ガス ×高さ 6 0 0 mm)のスラッジブランケット型バイオリア に分解し,再生可能なエネルギー資源として利用するメ クター(Upf l o wAna e r o bi cSl udgeBl a nke t ,上向流式 タン発酵法が適している。青森県のジュース工場ではリ 嫌気汚泥床)を使用した(図 1 )。水田土壌をリンゴ搾り ンゴ搾り粕のメタン発酵はまだ実施されていないが,メ 粕で馴養し,メタン発酵汚泥を製造した。3 0 ℃ で約 3 タン発酵は嫌気性微生物によって有機物を分解する技術 月間馴養し,経時的にメタンと二酸化炭素を測定した。 で,生成するメタンを燃料にして温水や発電に利用する ことが出来る。微生物による嫌気性処理は好気性処理と 3.ペクチン・キシラン分解菌の培養と培地組成 比べて消費エネルギーが少なく,余剰汚泥の生成量が少 嫌気性細菌の培地,培養方法は土壌微生物実験法(2 ) ないことから省エネルギー,少コスト型の処理法であ に従って行った。リンゴ搾り粕で馴養したメタン発酵汚 る。近年はスラッジブランケット型バイオリアクター 泥5 mlをペクチン培地 1 0 0 mlの入った 1 2 5 ml容バイ (Upf l o wAna e r o bi cSl udge Bl a nke t ,上向流式嫌気汚 アル瓶に接種し,3 0 ℃ で集積培養した。気相は窒素ガ 泥床) (1 )が普及し,ビール工場や製糖工場などの製造 スで置換し,容器口はブチルゴム栓,アルミキャップで 廃水処理に利用されている。このリアクターは自己集塊 密栓し,静置培養した。集積培養を 3回行った後,ロー 作用を持つ嫌気性微生物を沈降性に優れた粒状汚泥とし ルチューブ法によりペクチン分解菌を分離した。試験管 てリアクター内に形成させ,上向流と発生ガスによる流 はブチルゴム中栓,スクリューキャップ付き加圧培養試 動状態の中で基質と汚泥を接触させてメタン発酵を効率 験管を使用した。 的に行わせることができるリアクターである。粒状汚泥 ペ ク チ ン 培 地 組 成 は Na HCO3,2 g;NH4Cl ,0 . 2 8 g; は増殖速度の遅いメタン生成菌を高い菌体濃度に維持す KH2PO4,0 . 2 4 g;(NH4) CO3,0 . 1 7 g;MgSO4・7 H2O,0 . 1 2 ると共に嫌気性菌相互の共生関係を効率的に行うことに g;Ca CO3,1 . 0 g;ペクチン,5 g;酵母エキス,0 . 1 g;シ より搾り粕の成分であるペクチンやヘミセルロースを最 ステイン塩酸塩,0 . 5 g;をイオン交換水 1 Lに溶かした。 も簡単な有機物,メタンにまで分解することができる。 pH 7 . 0に調整し,オートクレーブ滅菌した。寒天は必要 本研究は,リンゴ搾り粕のメタン発酵を効率的に行う粒 に応じて 3 % 加えた。糖類の試験にはペクチンの代わり 状汚泥の製造と粒状汚泥からペクチン・キシラン分解菌 に糖類を加えた。 を分離し,菌学的特徴を明らかにするとともにメタン生 水素利用メタン生成菌は水田土壌から分離した TM8 成菌との共生について検討した。 菌株(3 )を使用した。TM8菌株の培地と培養は Zo uら (3 )の方法に従って行った。AP8 1菌株と TM8菌株の 混合培養はペクチン培地を用いて行った。 弘大農生報 No.9:21 -27,2006 武田・須田・殿内 2 2 表 1 リンゴ搾り粕の成分(%) 水分 ペクチン リグニン ヘミセルロース セルロース その他(糖類,有機酸等) 6075 3. 5 1. 5 1. 0 2. 0 520 青森県産業技術開発センター「キープロジェ クト研究報告」(14) 4.ペクチンの測定 ペクチンの測定はカルバゾール硫酸法(4 )により行っ た。培養液 1 mlにメタリン酸溶液(2 5 %メタリン酸- 図1 上向流式嫌気性汚泥床リアクター(UASB) 5 N H2SO4)0 . 2 mlを加え,4 ℃,1 0 , 0 0 0 r pm,1 0分間遠 心分離し,上清を採取した。上清 0 . 5 mlを試験管に取 り,硫酸/ホウ酸ナトリウム 2 . 5 mlを加え,1 0分間煮 9.16S r DNAの塩基配列の分析 沸した。水冷後,カルバゾール液を加え,1 5分間煮沸し 6 Sr DNA 増 幅 用 プ ラ イ マ ー 1 6 Sr DNA は 細 菌 1 た。水冷後,波長 5 3 0 nm の吸光度を測定した。硫酸/ SDBa c 0 0 1 1 a s 1 7 (f o r wa r d)と SDBa c 1 4 9 2 bA1 9 ホウ酸ナトリウムは,Na2B4O7・1 0 H2O 0 . 9 5 gを濃硫酸 (r e ve r s e )を使用し,PCR法によって増幅後,サイクル 1 0 0 mlに溶かした溶液,カルバゾール液は,C12H9N 1 2 5 シ ー ケ ン ス を 行 っ た。塩 基 配 列 は Ge nBa nk,EMBL, gを無水エタノール 1 0 0 mlに溶かした溶液を用いた。 DDBJに登録した(アクセッション番号:AB2 7 3 7 3 0 ) 。 5.発酵生産物の測定 結 果 と 考 察 発酵生産物の測定は前法(5 )と同じ方法で行った。メ タン,水素,炭酸ガスの測定はリアクター,試験管の気 1.メタン発酵汚泥の製造 相をガスタイトシリンジで 3 ml採取し,熱伝導検出器 メタン発酵はスラッジブランケット型バイオリアク (TCD)のガスクロマトグラフ(カラム,WG1 0 0 )で測 ターを用いて行った。水田土壌を種汚泥としてリンゴ搾 定した。キャリアガスはアルゴン,カラムオーブン温度 り粕で馴養し,メタン発酵汚泥を製造した。リアクター 6 0 ℃ で分析した。培養液の発酵生産物,エタノールは にリンゴ搾り粕を 3日毎に 1 gを加え,生成されるメタ 水素炎イオン検出器(FI D)のガスクロマトグラフ(カ ンと炭酸ガスを経時的に測定した。リンゴ搾り粕の主成 ラム,クロマトパック 5 4 )で測定した。キャリアガスは 分は表 1に示すように多糖類ではペクチン,へミセル 窒素,カラムオーブン温度 1 8 0 ℃ で分析した。蟻酸,酢 ロースとセルロースであり,糖類ではフルクトース,グ 酸と乳酸は高速液体クロマトグラフで測定した。 ルコース,ショ糖であった。 馴養約 3ヶ月後に直径 0 . 5~ 5 . 0 mm のメタン発酵粒 6.微生物の同定 状汚泥が形成された。粒状汚泥は弾力性があり,容易に 分離菌株のグラム染色,嫌気度,カタラーゼ活性,イ 壊れなかった。図 2 -a ,bに示すように表面は密に絡み ンドール生成,硝酸塩還元能,硫酸塩還元能,ミルク凝 合った糸状性菌で覆われ,糸状性菌の周りには桿菌や球 固・消化試験,ゼラチン消化試験は土壌微生物実験法 菌が多数存在した。メタン発酵は嫌気環境下で起こる微 (2 )に従って行った。 生物作用であるが,多様な機能を持つ微生物によって有 機化合物が分解される過程で還元物質が生成される。こ 7.GC含量の測定 の還元物質は水素を利用してメタンを生成するメタン生 DNAの GC含量の測定はヤマサ(ヤマサ醤油,東京) 成菌によって酸化される。このような複数の微生物に の DNAGCki tを用い,高速液体クロマトグラフで行っ よって有機物が分解されるので多様な微生物が隣接する た(5 ) 。 汚泥の粒状化は沈降性が良いだけなく,微生物相互の栄 養共生関係が保持される場を形成している(6 )。フロッ 8.電子顕微鏡写真 ク表面に観察される糸状性菌は Me t h a n o s a e t aの細胞と 透過型電子顕微鏡写真と走査型電子顕微鏡写真は 類似していた。 Mi z uka miら(5 )と同じ方法で撮影した。 Me t h a n o s a e t aは酢酸のみを基質とするメタン生成菌で あり,Me t h a n o s a e t aが増殖することによって沈降性に優 リンゴ搾り粕のメタン発酵 23 図 2 リンゴ搾り粕で馴養したメタン発酵汚泥 a:Me t hanos ae t a様糸状性細菌でおおわれたフロックの表面 b:粒状汚泥内部の糸状性細菌,桿菌,球菌。これらの汚泥は走査型電子顕微鏡で撮影した。 CH4, CO(ml) 2 500 400 300 200 100 0 0 10 20 30 40 培養時間(日) CH4 CO2 図4 AP8 1菌株の細胞の電子顕微鏡写真 ネガティブ染色し,透過型電子顕微鏡で撮影した。 バーは 1μm を示す。 図 3 リンゴ搾り粕のメタン発酵 れた顆粒状汚泥が形成される。メタン生成量は経時的に Cl .x y l a n o l y t i c u m(8 )であった。 増加し,定常期には図 3に示すようにリンゴ搾り粕 1 g 本菌株はペクチン,キシランの多糖やグルコース,キ (乾燥重量)当たり 3 5 0 mlのメタンが生成された。この シロースなどの単糖など表 3に示すように広い範囲の糖 3 生産量から乾燥重量 1 tのリンゴ搾り粕から 3 5 0 m のメ 類を資化する菌種であった。本菌の広範囲の糖類を資化 タンが生産されることが期待される。以上の結果はリン する特徴は,Cl .a e r o t o l e r a n sと Cl .x y l a n o l y t i c u mに類似 ゴ搾り粕のメタン発酵は,スラッジブランケット型バイ r o t o l e r a n sは多糖類のペクチンとキシランを した。Cl .a e オリアクターを用いることにより沈降性の良い粒状汚泥 資化する点で同じであり,単糖類と 2糖類の資化性でも が容易に形成され,メタン収率が良いことを示唆してい 類似した。しかし本菌株はマンニトールを資化し,ラク る。 ト ー ス と ト レ ハ ロ ー ス は 資 化 し な か っ た 点 が Cl . a e r o t o l e r a n sと異なった。本菌株は Cl .x y l a n o l y t i c u mが 5 2.メタン発酵汚泥から分離したペクチン・キシラン分 解菌の特徴 単糖であるリボースとアラビノースを資化しない点で異 なった。また Cl .x y l a n o l y t i c u mはペクチンやデンプンな メタン発酵粒状汚泥をペクチン培地で集積培養し, どの多糖類の資化性について記載がないので比較できな ロールチューブ法によりペクチン・キシラン分解活性を かった。本菌株の発酵生産物は蟻酸,酢酸,乳酸,エタ 有する AP8 1菌株を分離した。本菌株は図 4に示すよう ノ ー ル,H2 と CO2 で あ っ た。発 酵 生 産 物 は Cl . に周鞭毛による運動性があり,グラム陰性,胞子形成の a e r o t o l e r a n sは 本 菌 株 と 同 じ で あ っ た が,Cl . 絶対嫌気性菌であり,またカタラーゼ活性,インドール x y l a n o l y t i c u mは蟻酸,酢酸と乳酸であるので異なった。 生成,硝酸還元能とゼラチン消化は陰性であった。これ 本 菌 株 の DNA の GC含 量 は 4 2 mo l% で あ っ た。Cl . らの結果を表 2に示す。本菌株の細胞形態と生理・生化 a e r o t o l e r a n sと Cl 0 mo l%であり,本菌 .x y l a n o l y t i c u mは 4 学的特徴を持つ類似の嫌気性菌は Cl .a e r o t o l e r a n s (7 )と 株の GC含量と近い GC含量値であった。 武田・須田・殿内 2 4 表 2 AP81菌株の細胞形態・生理生化学的特長 AP81 細胞形態(µm) 胞子形成 グラム染色 運動性 コロニーの色 カタラーゼ活性 インドール生成 ゼラチン消化 硝酸塩還元 ミルク消化 硫化水素生成 発酵生産物 GC含量 0. 8 -1. 2× 2. 0 -8. 0 + 陰性 +a 白色 - - - - + + 蟻酸,乳酸,酢酸, ノール,H2,CO2 42mol % Cl .a e r o t o l e r a n s Cl .x y l a n o l y t i c u m 0. 4 -0 . 6× 1 . 5 -3 . 0 0 . 5 -0 . 8× 1 . 8 -3 . 0 + + - - +a + 淡褐色 - - - - - - - - ND ND - - エタ蟻酸,乳酸,酢酸, エタ蟻酸,酢酸,乳酸 ノール,H2,CO2 4 0mo l % 4 0 -4 1mol % Cl .s a c c h a r o l y t i c u m 0 . 6× 3 . 0 + - - 白色 - + - - 凝固 - 酢酸,乳酸,エタノール, H2,CO2,ピルビン酸 2 8mo l % a周鞭毛 表3 AP8 1菌株の基質利用性 基質 グルコース フルクトース ガラクトース マンノース アラビノース キシロース リボース マンニトール イノシトール マルトース スクロース セロビオース ラクトース ラムノース トレハロース メリビオース ラフィノース エスクリン イヌリン デキストリン デンプン ろ紙 CMC ペクチン キシラン AP81菌株 Cl .a e r o t o l e r a n s Cl .x y l a n o l y t i c u m Cl .s a c c h a r o l y t i c um + + + + + + + + - + + + - + - + + + - - - - - + + + + + + + + ± - - + + + + + ± + + + - W + - ND + + + + ND + - + - - - + + + - + - ND + ND ND ND ND ND ND ND + + + + + - + + + + + + + + + + + + + - ND - - ND ND ND ND :No tDe t er mi ned,W :Weak AP8 1菌 株 の 1 6 S r DNA の 塩 基 配 列 は Cl . 学 的 特 徴,系 統 学 的 特 徴 か ら,AP8 1菌 株 は Cl . a e r o t o l e r a n s ,Cl .x y l a n o l y t i c u mと Cl .s a c c h a r o l y t i c u m(9 ) a e r o t o l e r a n sに最も近縁の菌種であると考えられる。 の1 6 Sr DNAに最も高い 9 7 . 9 %の相同性を示した。前 2者はキシランを分解する。Cl .s a c c h a r o l y t i c u mは広い 3.AP81菌株の生育の特徴 範囲の糖類を利用するが,ペクチン,キシランの資化性 本菌株はペクチン,キシラン,グルコースやキシロー の記載がない。またこの菌種の GC含量は 2 8 mo l %と スなど広範囲の基質を資化し,主要な発酵生産物として かなり異なっている。AP8 1菌株の細胞形態,生理生化 エタノール,酢酸,水素と炭酸ガスを生成し,微量の蟻 リンゴ搾り粕のメタン発酵 14 12 3 8 6 2 4 1 H2,CO2,CH(%) 4 10 25 酢酸,エタノール (mM) 4 ペクチン (g/L) 30 2 4 6 8 10 12 10 0 0 0 15 5 2 0 20 0 14 2 4 6 ペクチン (AP81+TM-8) 酢酸 (AP81) 酢酸 (AP81+TM-8) エタノール (AP81) エタノール(AP81+TM-8) 図5 -a AP81菌株の単独培養と TM8菌株との混合培養に よるペクチン分解と醱酵生産物 養に示す。培養開始後,約 7日間でペクチンをほぼ分解 と8 g/ L培地で培養した時のペクチンの消費と水素生成 を図 6に示す。ペクチン 8 g/ Lで培養した時,生成した 水素は気相の 8 %であったが,ペクチン 5 g/ Lと比べて 増殖は抑制されず,ペクチンを分解した。プロピオン酸 分解菌(1 0 )や,酪酸分解菌(3 ,1 1 )は極めて低い水素 ペクチン(g/L) 殖期にそれぞれ 5 %と 2 0 %であった。AP8 1菌株の増 CO(AP81) 2 CO(AP81+TM-8) 2 2 H(AP81) H(AP81+TM-8) 2 8 9 7 8 7 6 し,エタノール,酢酸,水素と炭酸ガスが主要な発酵生 殖は水素によって阻害されなかった。ペクチンを 5 g/ L 14 図5 b AP8 1菌株の単独培養と TM8菌株との混合培養に よるペクチン分解と醱酵生産物 で培養した時の増殖と発酵生産物を図 5 -a ,bの単独培 2 . 1の比率であった。気相中の水素と炭酸ガスは対数増 12 CH(AP81+TM-8) 4 酸と乳酸を生成した。AP8 1菌株を 0 . 5 %ペクチン培地 産物であった。エタノールに対する酢酸の比は 1 . 8~ 10 培養時間(日) 培養時間 (日) ペクチン (AP81) 8 6 5 5 4 4 3 H(%) 2 5 25 3 2 2 1 1 0 0 0 2 4 濃度で阻害されるが,AP8 1菌株は生成するかなり高い 6 8 10 12 14 培養時間(日) 水素濃度によっても阻害されないことを示唆している。 AP8 1菌株はペクチン分解にともない生成する水素濃 度の高い気相下で増殖が抑制されなかったが,水素消費 菌の共存によって発酵生産物がどのように変わるか検討 ペクチン(5g/L) ペクチン(8g/L) H(5g/L) 2 2 H(8g/L) 図6 ペクチン分解と水素生成 1菌株の細 した。本実験は,TM8菌株の細胞数を AP8 胞数に対して 1 0 0倍の細胞濃度にして接種し,培養を ことを示唆している。2者混合培養による主要な発酵生 行った。2者混合培養のペクチン分解と発酵生産物の経 産物である酢酸とエタノールは単独培養と比べ,エタ 時的変化を図 5 -a ,bの混合培養に示す。AP8 1菌株の ノールは減少し,酢酸が増加した。エタノールに対する 単独培養と TM8菌株との 2者混合培養はいずれもペク 酢酸の比は 3 . 5~ 4であった。この比率は AP8 1菌株単 チン分解にともない気相中に水素が検出されたが, 独培養の 2倍であった。AP8 1菌株と TM8菌株の混合 AP8 1菌株単独培養では最大 6 . 5 %であったのに対して 2 b u sと水 培養はルーメンから分離された Ru mi n o c o c c u sa l 者混合培養では 3日に最大 1 . 9 %であり,培養 1 0日以降 素利用メタン生成菌,Me t h a n o b r e v i b a c t e rr u mi n a n t i u mの には水素は検出されなかった。TM8菌株は単独ではペ 混合培養と似ている。Ru mi n o c o c c u sa l b u sは単独培養で クチン培地に増殖しなかったので AP8 1菌株との 2者混 は水素生成が低い水素分圧で阻害され,エタノールを生 合培養により本菌株は水素を消費し,メタンに転換した 成する菌種であるが,Me t h a n o b r e v i b a c t e rr u mi n a n t i u mと 武田・須田・殿内 2 6 の 共 生 培 養 で は 生 成 す る 水 素 は Me t h a n o b r e v i b a c t e r 酢酸を多く生成した。AP8 1菌株は水素利用メタン生成 r u mi n a n t i u mによって消費されるので水素は低い濃度に 菌との共生がエネルギー生成に有利に作用することを示 維持される。そのため Ru mi n o c o c c u sa l b u sは共生培養で 唆している。 はエタノールを生成せず,酢酸を生成することを Wo l i n と Mi l l e rは報告(1 2 ,1 3 )した。AP8 1菌株は気相に 8 % という高い水素濃度でもペクチン分解が阻害されない菌 種であったが,水素利用メタン生成菌と共生することに よって水素は水素利用メタン生成菌によって消費される ので,エタノール生成を抑制してエネルギー生成に有利 な酢酸生成の代謝系を優先させることを示唆している。 謝 辞 本研究の研究開発に協力頂いた青森県工業総合研究セ ンター弘前地域技術研究所生命科学部の方々に感謝致し ます。リンゴ搾り粕を提供していただいた日本果実加工 (株) に感謝致します。メタン発酵汚泥と微生物細胞の電 子顕微鏡試料の調製,撮影に関して指導していただいた 生物生産科学科の藤田隆助手に感謝致します。本研究の 一部は青森テクノポリス開発機構 ( 「環境循環型社会に対 応した未利用資源の高度利用技術の開発」 ) から支援して 頂きました。 摘 要 メタン発酵は上向流式嫌気汚泥床(UASB)リアクター を用いて行った。汚泥は水田土壌をリンゴ搾り粕で馴養 して製造した。3ヵ月後,球菌,桿菌や糸状性菌からな る粒状汚泥が形成された。粒状汚泥からペクチン,キシ ラン分解菌,AP8 1菌株を分離した。本菌株は周鞭毛に よる運動性があり,グラム陰性,胞子形成の絶対嫌気性 細菌であった。本菌株は広範囲の糖類を資化し,発酵生 産物として蟻酸,酢酸,乳酸,エタノール,H2と CO2を 生成した。DNAの GC含量は 4 2 mo l%であった。本菌 株の 1 6 Sr DNA の塩基配列は Cl .a e r o t o l e r a n sに最も近 縁で 9 7 . 9 %の相同性を示した。本菌株は,細胞形態,生 理・生 化 学 的 特 徴 と 1 6 Sr DNA の 解 析 結 果 か ら Cl . a e r o t o l e r a n sに最も近縁の菌種でると推察される。本菌 株はペクチン,キシランに加えてグルコースやアラビ ノースなど広範囲の糖類を資化し,主要な発酵生産物と してエタノール,酢酸に加えて著量の水素を生成した。 本菌株はペクチン,キシランの分解にともなって生成す る水素によって生育は阻害されなかったが,水素利用メ タン生成菌との混合培養によってエタノールは減少し, 引 用 文 献 1)上木勝司,永井史郎編著:嫌気微生物学,2 6 5 -284頁, 養賢堂,東京,1 9 9 3. 2)土壌微生物研究会:新編土壌微生物実験法,2 3 -35頁, 養賢堂,東京,1 9 9 2. 3)ZOU,B. ,K.TAKEDA,A.TONOUTI,S.AKADA,and T. FUJITA : Cha r a c t e r i s t i c s of ana e r o bi c ,s ynt r ophi c , but yr a t e de gr a di ng ba c t e r i um i n pa ddy f i el d s oi l : Bi o s c i .Bi o t e c h .Bi o c h e m.6 7:2 0 5 9 2 0 6 7 .2 0 0 3 . 4)BITTER,T.a nd H.M.MUIR :A modi f i c a t i o n ur oni c a s i dc a r ba z o l er e a c t i o n.:An a l .Bi o c h e m.4 :330334. 1 9 6 2 . 5)MIZUKAMI,S. ,K.TAKEDA,S.AKADA a nd T.FUJITA : I s o l a t i o na nd c ha r a c t e r i s t i c so fMe t h a n o s a e t ai n paddy f i e l ds o i l s .:Bi o s c i .Bi o t e c h .Bi o c h e m.7 0:8 2 8 8 3 5.2006. 6)STAMS,A.J .M.:Met a bo l i ci nt e r a c t i o ns bet we en a na e r o bi c ba c t e r i ai n me t ha no ge ni ce nvi r o nment s .: An t o n i ev a nLe e u we n h o e k6 6.:2 7 1 2 9 4 .1 9 9 4 . 7)VAN GYLSWYK,N. O.a nd J . J . T. K.VAN TOOM :Cl os t r i di um a e r o t o l e r a n ss p.No v. ,a xyl a no l yt i c ba c t e r i um f r o m c o r ns t o ve ra nd f r om t he r umi na o fs he e pf ed c or n s t o ve r .:I n t .J .S y s t .Ba c t e r i o l .3 7:1 0 2 1 0 5 .1 9 8 7 . 8)ROGERS, G. M. and A. A. W. BAECKER : Cl os t r i di um x y l a n o l y t i c u m s p. Nov. , an ana e r o bi c xyl anol yt i c ba c t e r i um f r om de c a ye d pi nus pa t ul a woo dc hi ps .: I n t .J .S y s t .Ba c t e r i o l .4 1:1 4 0 1 4 3 .1 9 9 1 . 9)MURRAY,W. D. ,A. W.KHAN a nd L.VAN DEN BERG : Cl o s t r i d i u m s a c c h a r o l y t i c u m s p.No v. ,a s a c c har ol yt i c s pe c i e sf r om s ewa ges l udge .:I n t .J .S y s t .Ba c t e r i ol .32 :1 3 2 1 3 5 .1 9 8 2 . 1 0)BOONE,D. R.a nd M. P.BRYANT :Pr o pi o na t e degr adi ng ba c t e r i um,S y n t r o p h o b a c t e r wo l i n i is p.No v.gen.nov. , f r om met ha no ge ni c ec o s ys t ems . : Ap p l . En v i r on. Mi c r o b i o l .4:6 2 6 6 3 2 .1 9 8 0 . 1 1)Mc J . ,M. P.BRYANT,R. B.HESPELL a nd J . W. I NERNEY,M. COSTERTON : S y n t r o p h o mo n a s wo l f e i ge n. No v. s p. No v. an ana e r o bi cs ynt r o phi c ,f a t t y ac i do xi di zi ng ba c t e r i um.:Ap p l .En v i r o n .Ba c t e r i o l .4 1 :1 0291039. 1 9 8 1 . 1 2)MILLER,T. L.a nd M. J .WOLIN :Fo r mat i o no fhydr ogen a nd f o r mat eby Ru mi n o c o c c u sa l b u s .:J .Ba c t e r i ol .116 . 8 3 6 8 4 6 .1 9 7 3 . 1 3)WOLIN, M. J . and T. L. MILLER : Mi c r o be Mi c r obe i nt e r a c t i o ns .I n t he r ume n mi c r o bi a le c os ys t e m. Ho bs o n, P. N. a nd C. S. St ewar t (Eds ) . Bl ac ki e Ac a demi c& Pr o f e s s i o na l .Lo ndo n.1 9 9 7 . 1 4)青森県産業技術開発センター:キープロジェクト研究報 告,リンゴ搾汁残渣の成分 p3~ 8 .1 9 9 2 リンゴ搾り粕のメタン発酵 27 Me t ha nef e r me nt at i onofanappl epo mac e Cha r ac t er i s t i c sofapec t i n・xyl a nde gr a di nga na e r o bi cba c t e r i um i s o l a t e df r o m me t hanef e r ment a t i o ns l udge s . Ki yo s hiTAKEDA,I kueSUDA a ndAki oTONOUCHI La b o r a t o r yo fAp p l i e dMi c r o b i o l o g y SUMMARY Met hanef er ment at i on wasc ar r i ed outwi t h an upf l o wanaer obi cs l udgebl anketr eac t or .Thes l udge wasmadeby ac c l i mat i zi ng paddy s oi ladded asas eed c ul t ur ewi t h an appl epo mac e.Gr anul at ed s l udge c ompos ed ofc oc c oi d,r odsand f i l ament ousbac t er i awasf or med af t er3mont hsi nc ubat i on.Pec t i n・xyl an degr adi nganaer obi cbac t er i a,s t r ai nAP81wasi s ol at edf r o mt hemet hanef er me nt at i ons l udge,whi c hwas ac c l i ma t i zedwi t happl epomac e.Thes t r ai nwasmot i l ewi t hl at er alf l agel l a,Gr a mnegat i ve,s por ef or mi ng s t r i c tanaer obes .The s t r ai n us ed a wi de r ange ofs ugar sc ont ai ni ng gl uc os e and xyl os ei n addi t i on t o pec t i n and xyl an and pr oduc ed f or mat e,ac et at e,l ac t at e,et hanol ,c ar bon di oxi de and hydr ogen as %.The16S r DNA s i mi l ar i t y ofs t r ai n AP81 f er ment at i on pr oduc t s .TheGC c ont entofDNA was42mol t o Cl .ae r ot ol e r answa s97. 9 %.The s t r ai ni sc ons i der ed t o be c l os el yr el at ed t o Cl .ae r ot ol e r anson t he bas i s ofi t sc el lmor phol ogy and t he phenot ypi c and phyl ogenet i cc har ac t er i s t i c s .St r ai n AP81 us ed a wi de r ange ofs ugar sc ont ai ni ng gl uc os e orar abi nos ei n addi t i on t o pec t i n and xyl an,and pr oduc ed a l ar ge amountofhydr ogen i n addi t i on t o et hanoland ac et at e ast he mai nf er ment at i on pr oduc t s .The gr owt h oft hes t r ai n wasnots uppr es s ed by hydr ogen pr oduc ed by degr adi ng ofpec t i n and xyl an.The pr oduc t i on ofet hanol ,however ,dec r eas ed,whi l et hatofac et at ei nc r eas ed by t he dualc ul t ur e wi t ha hydr ogenut i l i zi ng met hanogen.I ti ss ugges t ed t hatt he ener gy met abol i s m hasan advant ageousef f ec t ont hes t r ai nAP81byt heas s oc i at i onwi t hhydr ogenut i l i zi ngmet hanogens . l .Fac .Agr i c .& Li f eSc i .Hi r os akiUni v.No.9:21 -27,2006 榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎Bul 浅 田 2 8 リンゴ樹における発生部位別スパー構成に関する研究 浅 田 武 典 園芸学講座 (2 0 0 6年 1 0月 1 3日受付) 目 次 第 1章 序論………………………………………………2 9 第 2章 せん定樹におけるスパーの発生部別構成割 合と栄養・生殖生長および果実生産性の関 係…………………………………………………3 0 緒言………………………………………………………3 0 材料および方法…………………………………………3 0 結果および考察…………………………………………3 1 1 .発生部位別スパーの構成割合と栄養生長お よび花芽形成との関係…………………………3 1 (1 )長さおよび発生部位別 1年枝の構成割合…3 1 (2 )発生部位別スパー率の相互関係と頂生ス パ-率を左右する要因………………………3 2 (3 )頂生スパー率と栄養生長および外部形態 の関係…………………………………………3 3 (4 )頂生スパー率と花芽形成の関係……………3 4 2 .スパーの発生部位別構成割合と果実生産性…3 5 )葉面積当たり果数(FLA, Fr ui t sperl e a f (1 ar ea )の樹冠内変異…………………………35 (2 )FLAに及ぼす太枝の枝齢,発出角度,発 出部位高,および栄養生長の強さの影響…3 5 (3 )FLAと頂芽増加率(BPR)および 1年枝 当たり葉面積の相互関係……………………3 6 (4 )1年枝当たり葉面積を左右する要因 ………3 7 (5 )FLAと頂生スパー率の関係 ………………3 7 (6 )FLAと頂芽増加率の相関関係における樹 間変異…………………………………………3 8 3 .発生部位別スパー形成に及ぼす着果の影響…3 8 摘要………………………………………………………4 0 第 3章 スパー枝齢と果実品質および果台枝形成の 関係………………………………………………4 1 緒言………………………………………………………4 1 材料および方法…………………………………………4 2 結果………………………………………………………4 2 1 .えき芽,1年生および 2年生以上のスパー の比較……………………………………………4 2 2 .1,2 ,3および 4年生以上のスパーの比較 … 4 3 考察………………………………………………………4 3 摘要………………………………………………………4 4 第 4章 無せん定樹におけるスパーの発生部位別構 成割合と栄養・生殖生長の関係………………4 4 緒言………………………………………………………4 4 材料および方法…………………………………………4 4 結果および考察…………………………………………4 5 1 .発生部位別スパー率の樹齢による変異およ び樹内変異………………………………………4 5 2 .スパーの発生部位別構成割合と栄養生長の 関係………………………………………………4 7 弘大農生報 No.9:28 -67,2006 (1)平均新しょう長,1・2年枝長および頂芽 増加率…………………………………………47 (2)発生部位別スパー率と弱小枝の発生との 関係……………………………………………48 3 .頂生スパー率と花芽形成の関係………………49 摘要………………………………………………………49 第 5章 スパー形成要因およびせん定によるスパー 率の調整…………………………………………49 緒言………………………………………………………49 材料および方法…………………………………………49 結果および考察………………………………………50 1 .スパー形成要因…………………………………50 (1)えき生スパーの形成要因……………………50 ①無せん定‘ふじ’5年生樹における発生 えき生枝数およびスパー数を左右する母 枝形質…………………………………………50 ②無せん定 9年生樹と成木せん定樹におけ る全えき生枝およびえき生スパーの発生 数と母枝長関係………………………………51 (2)頂生スパー 形成要因………………………52 ①えき生スパーから頂生スパーの形成………52 ②えき生スパーの頂芽から 2年後に頂生ス パーを形成する割合…………………………53 2 .せん定が発生部位別スパー率に及ぼす影響…54 (1)せん定樹と無せん定樹における枝長の経 年変化…………………………………………54 (2)1年枝の長さ別構成割合……………………5 4 (3)長枝の発生部位別構成割合…………………55 (4)スパーにおける発生部位別構成割合………56 3 .枝モデルによる発生部位別スパー率の調整 法の検討…………………………………………57 (1)基本枝形成モデル……………………………57 (2)頂生スパー率の増加方法……………………58 ①主軸枝からえき生の長枝が 1本も形成さ れず,スパーのみ形成される場合…………58 ②えき生枝の長枝数を減らした場合…………58 (3)えき生スパー率増加法………………………58 ①えき生枝における長枝・スパー比を高める…58 ②えき生枝のスパーの頂芽から長枝が発生 する……………………………………………58 摘要…………………………………………………59 第 6章 総合考察…………………………………………60 第 7章 総括………………………………………………63 謝辞…………………………………………………………64 引用文献……………………………………………………64 Summar y…………………………………………………66 リンゴ樹における発生部位別スパー構成 第 1章 序 論 果樹における枝形成に関する研究は,栽培管理と良品 29 ばえが果実品質や収量に影響すると考えられる。 スパー葉を摘除すると,収穫果の結実率,大きさ,Ca 含量が低下することが知られている(Abbo t t ,1 9 6 0 ; 生産にかかわる性質に関して行われてきた。短い節間を Fe r r e e ・Pa l me r ,1 9 8 2 ) 。樹冠内には,頂芽に花芽を付 もつ品種が花芽形成し易いこと(Br unne r ,1 9 9 0 ),開張 けたスパーでも,その性質に様々な変異がある。例え 性の品種が枝の直立性を示す品種に比べ管理がし易いこ ば,枝直径,休眠芽の大きさ,果台に着生する果そう葉 と(Qui nl a n・To but t ,1 9 9 0 ) ,頂芽優勢性と頂部優勢性の の枚数あるいは面積,果そう内花数,花弁の大きさなど 程度が果樹の種類や品種によって異なること(Lo r e t i , である。果そう葉面積は品種により 2倍以上の違いがあ 1 9 9 0 ),リンゴの早期収量を高めるためのフェザーの発 る(Ba r r i t t ・Sc ho nbe r g,1 9 9 0 ) 。これらの形質は,同じ 生のし易さ(Co s t e s ・Gue do n,2 0 0 2 ) ,隔年結果性とせん 品種内でも値が大きい方が果実品質にとって望ましい。 定の軽減に関係する発芽後の生長の継続性とその品種間 リンゴの休眠芽の大きさは,直径が 3 mm 以下を弱小芽 差異(La ur iら,1 9 9 5 ) ,枝の測定値からの樹冠の復元(山 とし,その割合を生産の指標にしている例がある(青森 本・伊藤,1 9 9 6 ) ,その他せん定技術を確立するための枝 県,2 0 0 0 )。これらの形質は,春の発芽の時点において 形成の諸性質(Ki kuc hi ,1 9 7 4 )などが主な研究の対象で は芽の素質であるが,それ以後は発育とともに変化す あった。 る。Ba r r i t tら(1 9 8 7 )は,樹冠上部のスパーは,葉乾 短果枝(以下スパーという)形成についても古くから 重,スパー当たり葉乾重,葉面積当たり乾物重(SLW) 注目されてきた。1 9 5 0年代に多くのスパータイプ品種 の他に,芽の直径,スパーの長さが樹冠下部のスパーよ が報告されている(Qui nl a n・To but t ,1 9 9 0 )。スパータ り優れると報告している。従って,生長期のスパーの生 イプは,標準タイプに比べ,えき生枝としてスパーを多 長条件が,光合成能力の優れたスパーを生長させ,良品 く着生する(Ar a s u,1 9 6 7 ) 。そのため,木が小型になる 果を生産すると同時に,翌年の素質のよいスパーを形成 ので,果実品質がよいことも評価されて,導入された。 すると考えられる。スパーの生長条件としては,光が主 北米におけるスパータイプを利用したリンゴ栽培の成功 な役割を果たすとする報告が多い(Ro bi ns o n,1 9 8 3 ; により,果樹の育種の目標として枝形成における品種特 Se e l e y,1 9 8 0 ) 。しかし,せん定がスパーと新しょうの葉 性が注目されるようになった。スパータイプの遺伝子は 面積に影響すること(La ks o ,1 9 8 4 )や“スパー枝齢” (第 同型接合的優性遺伝子 Coにより制御される(La pi ns , 3章 参 照)が 古 く な る に つ れ て 果 実 が 小 さ く な る 1 9 7 4 )。スパー形成の性質を育成する研究の成果の代表 (Ro bi ns o n,1 9 8 3 ;Vo l z ,1 9 9 4 )などの報告があり,ス 的なものの一つは, ‘Mc I nt o s hWi j c i k’のようなカラム パーのできばえを左右する要因については光以外の要因 ナータイプの作出であった(La pi ns ,1 9 7 4 ) 。カラムナー も考えられる。そのため,スパー形成を左右する要因に タイプでは長枝をほとんど形成せずにスパーを形成する ついて明らかにすることが重要である。 ため,樹が細い円筒状となり,省力と多収の実現,ある リンゴ樹の果実生産性はスパー率により影響される可 いは受粉樹として利用される可能性を認める報告がある 能性がある。浅田(19 9 9 )は,リンゴ樹の葉面積当たり (猪俣ら,2 0 0 4 )。一方,スパー形成能力の高い品種は 果数が頂芽増加率と負の相関関係にあることを示し,頂 隔年結果性が強いため,せん定を省力的にできる品種 芽増加率を左右する要因を検討した。その結果,スパー は,むしろ逆に長いえき生枝を発生しやすい品種とする の中で短い 2年枝の頂芽から発達するスパーの 1年枝に 考えもある(La ur iら,1 9 9 5 ) 。スパータイプやカラム 占める割合が高い方が頂芽増加率は低く,葉面積当たり ナータイプの果樹生産と育種資源における評価は分かれ 果数を多くする要因であることを見出した。この結果 ている。また,スパーを形成しやすい品種は頂部優勢性 は,同じスパーでもリンゴ樹の果実生産性に異なる役割 が 強 い と す る 報 告(Lo r e t i ,1 9 9 0 )が あ る 一 方 で, を果たすスパーが存在することを示している。そこで, Le s pi na s s e ・De l o r t (1 9 8 6 )のように,基部優勢を示す品 1年生スパーをスパー形成母枝としての 2年枝の長さ別 種に分類する報告もある。スパー性と頂部優勢性の関係 と 2年枝上の頂生・えき生別に分類し,それらの 1年枝 についても検討すべき課題が残されている。 に占める割合と栄養生長および生殖生長との関係を検討 リンゴにおけるスパーは,果実を生産する枝として最 した。その結果,スパーの頂生とえき生の違いが,樹全 も重要な枝である。Ki kuc hi (1 9 7 4 )は,無せん定樹とせ 体あるいは太枝の栄養生長と生殖生長の両者と密接な関 ん定樹について新しょうの長さ別度数分布を調べた結 係にあることを認めた(浅田,2 0 0 4 )。 果, ‘国光’では 5 c m, ‘スターキング・デリシャス’で これまでの研究をみると,スパーを長い枝のえき芽か は5 -1 0 c m に最少の分布域があり,二つの大きなグ ら形成される 1次スパーとすでに形成されたスパーの頂 ループに分かれることを示した。浅田(2 0 0 4 )は,5 c m 芽から形成される 2次スパーに分類した記述は存在する 0 %以上を占め,着果した 以下のスパーが全 1年枝の 7 が(Tho mps o n,1 9 6 6 ) ,分類したスパーの構成割合がど スパーが全 1年枝の 7 7 %以上に達することがあると報 ういう意味を持つのかを検討した報告はみられない。そ 告している。従って,スパーの形成過程やそれらのでき こで,スパーの発生部位別の構成割合と樹あるいは太枝 浅 田 3 0 の生長と結実の関係について詳細に検討することとし それらの断面積,樹冠容積,葉面積で表されることが多 た。 い(Fe r r e e ,1 9 8 0 )。 断面積は強いせん定が行われると 生産者は枝を観察し,良品果を安定生産するのに必要 樹あるいは枝の大きさとの相関が弱くなり,また,樹冠 な枝を確保するせん定を行っている。スパーの種類は眼 容積は測定が簡単でない。以上の理由により,果実生産 で識別できる形質の一つである。仮に,スパーの種類別 性を葉面積当たり果数で表した。 構成割合をせん定など人為的な操作により制御できると すれば,スパーの種類別構成割合の調整を通してせん定 材料および方法 を体系づけることが可能になる。本研究では,以下の 4 点を明らかにすることを目的として,データの収集と解 弘前大学農学生命科学部付属生物共生教育研究セン 析を行った。1 )発生部位別スパー構成割合と栄養生長, ター藤崎農場に植栽されている 1 6年生開心形‘王林’1 2 花芽形成,果実生産性との関係,2 )発生部位別スパーの 樹から選抜した 3 1本の太枝(生枝下直径が 5 c m 程度の 形成要因,3 )せん定による発生部位別スパー構成割合の 太さで,骨格を形成する枝でなく更新の対象となる枝) 変化,および 4 )発生部位別スパーの構成割合を調整する について,津軽地域の慣行栽培園 5園から選定した約 2 0 方法,である。 年生開心形‘ふじ’5樹(全数調査で,太枝数 1 6 3 )と なお,データの収集にあたって,スパーの定義付けが 1 0 a当たり 1 2 5本植えのマルバカイドウ付き M.2 6台 必要なので,考え方を整理しておく。結果枝とは果実を 主幹形園 7園から選んだ各園 2樹の約 1 0年生‘ふじ’計 直接形成する枝で,枝齢を満年齢で呼ぶ場合,リンゴで 1 4樹について解析した。主幹形‘ふじ’樹は,栄養生長 は 1年枝が該当する。1年枝は,短~長果枝,発育枝,徒 の強さが異なると推察される上・中・下の 3部位に分け, 長枝等に分けられる。そのうち結果枝はおもに短~長果 各部位毎に測定値をまとめた(全部位数 4 2 ) 。開心形樹 枝で構成され,リンゴでは,短果枝を 5 c m 以下,5~ は,両品種とも台木がマルバカイドウ(Ma l u sp r u n i f o r i a 1 0 c m を中果枝,1 0~ 2 0 c m を長果枝と分類されてき Bo r kh.v a r .r i n g oAs a mi ) ,栽植密度が 1 0 a当たり 1 8本 た(小原,1 9 8 4 ) 。外国の分類においても短い枝をスパー 植えであった。‘王林’樹は,地域でせん定指導を行って と呼ぶが,一般にその長さが 5~ 6 c m 以下とされてい いる生産者 6人がせん定した樹で,栄養生長の強さの違 るので(Vo l zら,1 9 9 4 ) ,以下 5 c m 以下の枝を短果枝と いを幅広く含むよう太枝を選定した。‘ふじ’の両樹形樹 し,スパーと称する。モモでは,短果枝が 1 0 c m 以下, は,該当する地域の農協より品質,収量とも優良と推薦 1 0~ 3 0 c m が中果枝,3 0 c m 以上が長果枝とされ,リン された園地の中央部の代表的な樹姿を有するものであっ 9 9 4 ) 。スパーは 1年枝と当年 ゴとは異なっている(柴,1 た。供試した‘王林’および‘ふじ’の開心形と主幹形 枝に該当する枝が存在する。せん定樹を供試する場合 樹 の 太 枝 は,頂 端 新 梢 長(平 均 ±SD)が そ れ ぞ れ は,1年枝のスパーはせん定の前後で全 1年枝に対する 1 1 . 1 ±8 . 9 ,1 5 . 1 ±5 . 5 ,1 0 . 3 ±3 . 1 c m で あ っ た。調 査 は 8 比率が異なる。そのため,スパーの年齢とせん定の前後 月から 9月にかけて新しょう伸長が停止した時点で行 を明記する必要がある。本研究では,特に記述しない限 い,当年生長した枝を新しょう,前年および前前年生長 り,1年枝のスパーをスパーとし,全 1年枝に対する割 した枝をそれぞれ 1年枝,2年枝とした。1年枝の分類 合をスパー率とした。 は,1年枝と 2年枝の長さ(短,中,長)の組合せおよ び頂生とえき生の違いに基づき 1 8種類とした。短,中 第2章 せん定樹におけるスパーの発生部位別 構成割合と栄養・生殖生長および果実 生産性の関係 緒 言 および長の枝長はそれぞれ 0 . 5~ 5 c m,5 . 1~ 1 0 c m, および 1 0 . 1 c m 以上とした。1年枝の頂芽については果 台と果実の有無および新しょう長を,えき芽については えき生花そう葉の有無と新しょう数および長さを調査し た。それらの調査は,枝の先端から基部に向かって太枝 の枝齢と垂直軸に対する発出角度を記録しながら,1樹 枝の発生部位別構成割合は,せん定によって変化する あるいは太枝全ての 1年枝について行った。葉面積は, 最も基本的な性質である。リンゴ樹におけるその仕組み 新しょう葉と花そう葉に分け,新しょう葉は新しょう長 を明らかにすることにより,せん定技術の理解に役立つ と新しょう上の葉面積の回帰式を各園について求め,そ ものと考えられる。本章では,スパーの発生部位別構成 の式を用いて総新しょう長から,また花そう葉は頂生花 割合に注目し,リンゴ樹の栄養と生殖生長にどのように そうとえき生花そうのそれぞれの 1花そう当たり平均葉 関係しているかをせん定樹について検討した。なお,太 面積を求め,各花そう数を乗じて算出した。なお,太枝 枝の果実生産性は,葉面積当たり果数で表した。樹や太 は,生枝下直径が 5 c m 以下の太さとし,更新対象の枝 枝の果実生産性は,収量と枝の大きさで決められるが, とした。また,太枝の発出角度は垂直を 0 °として表し 収 量 は 果 数 と 密 接 な 関 係 に あ る(As a da ,1 9 8 8 ; た。 Fo r s he ya nd El f vi ng,1 9 7 7 ) 。他方,木や枝の大きさは リンゴ樹における発生部位別スパー構成 31 60 ふじ-OC 分布割合 (%) 50 ふじ-CL 王林-OC 40 30 20 10 aL /L aM /L aS /L tL /M tM /M tS /M aL /M aM /M aS /M tL /S tM /S tS /S aL /S aM /S aS /S tS /L tL /L tM /L 0 1年枝 /2年枝 第2 -1 -1図 リンゴ‘ふじ’と‘王林’における 1年枝の発生部位別構成割合 横軸の文字は分子が 1年枝の発生部位と長さ,分母が 2年枝の長さを表す。tと aはそれぞれ頂生,えき 生を表し,L,M,Sはそれぞれ 10 c m < ,5~ 1 0 c m,5 c m ≧ の長さを表す。OCは開心形樹,CLは M.26台主幹形樹を示す。 結果および考察 1.スパーの発生部位別構成割合と栄養生長および花芽 形成との関係 (1)1年枝の長さおよび発生部位別構成割合 1 ,2年枝の長さと発生部位により 1年枝を 1 8種類に 分類し,それぞれの 1年枝の太枝における構成割合につ いて検討した(第 2 -1 -1図) 。最も大きな割合を占める 1年枝はスパーであり, ‘ふじ’で約 8 2 %, ‘王林’で 7 7 %を占めた。なお,リンゴ樹では,スパーは頂生(第 2 -1 -1図の t S/ L+ t S/ M +t S/ S)とえき生(同図の a S/ L+ a S/ M +a S/ S)の 2種類で構成される。 1 ,2年枝を長さと母枝上の発出位置により分類した 模式図を第 2 -1 -2図)に示した。 「頂生スパー」は,前 年枝の頂芽に形成されるスパーで,比較的短い前年枝に 形成されることが多い。一方, 「えき生スパー」 は前年枝 のえき芽に形成されるスパーで,通常は比較的長い前年 枝のえき芽から形成される。 両スパーの 1年枝に占める割合は,太枝によって大き く変異し,頂生スパーの場合,最低 1 . 5 %(開心形‘ふ じ’ )から最高 6 2 . 8 %(開心形‘王林’ )まで,またえき 第2 -1 -2図 生スパーの場合,最低 2 2 . 4 %(開心形‘ふじ’ )から 8 5 . 2 リンゴにおける頂生スパーとえき生スパーの 模式図 %(主幹形‘ふじ’ )まで認められた(第 2 -1 -4図)。 頂生スパーが 1年枝に占める割合の平均値は, ‘ふじ’2 5 %, ‘王林’3 0 %であった。同様に,えき生スパーの割 た(1 %水準で有意) 。‘ふじ’では樹形による違いはな 合は, ‘ふじ’5 7 %, ‘王林’4 7 %を占めた。また,1 0 かった。 c m 以上の長い 1年枝は 1 0 %弱を占め,5~ 1 0 c m の中 以上の結果,品種間で比率に違いはあるが,2種類の %と少なかっ 果枝は‘ふじ’で 5 %以下, ‘王林’で 7 スパーが 1年枝の主要な構成要素であることは,リンゴ た。頂生とえき生スパーの比率を品種で比較すると, の成木樹に共通する性質であることが認められた。 ‘王林’は‘ふじ’に比べ,えき生が少なく頂生が多かっ 果実生産の役割を担う主な 1年枝を明らかにするため 浅 田 3 2 70 60 ふじ-OC ふじ-CL 王林-OC 分布割合(%) 50 40 30 20 10 S aS / aM /S S aL /S tS / tM /S tL /S M aS / aM /M M aL /M tS / tM /M L tL /M aS / aM /L L aL /L tS / tM /L tL /L 0 1年枝 /2年枝 第2 -1 -3図 リンゴ‘ふじ’と‘王林’における着果した 1年枝の発生部位別構成割合 横軸の文字は分子が 1年枝の発生部位と長さ,分母が 2年枝の長さを表す。tと aはそれぞれ頂生,えき 生を表し,L,M,Sはそれぞれ 10 c m < ,5~ 1 0 c m,5 c m ≧ の長さを表す。OCは開心形樹,CLは M.26台主幹形樹を示す。 に,前述の 1 8種類に分けた 1年枝について着果してい 図) 。果実を着生したスパーは,両樹形の‘ふじ’で 8 9 %, ‘王林’で 7 7 %を占め,本調査樹における果実生産 を担う主要な 1年枝がスパーであった。2種類のスパー を比較すると,えき生スパーが‘ふじ’で 5 7~ 5 9 %, ‘王林’で 4 4 %と高く,頂生スパーが‘ふじ’で 3 0~ 70 頂生スパ−率(%) る枝の全着果数に占める割合を比較した(第 2 -1 -3 ふじOC y=74.20.91x (r=0.775**) 60 50 40 30 20 3 2 %, ‘王林’で 3 3 %と低かった。その他果実を生産す 10 る枝は,前述の 1年枝の分布割合とほぼ同じであり,全 0 いずれも少なかった。 以上のように,1年枝のせん定後の長さ別構成割合は スパーが大部分を占め,全果実の 8 0~ 9 0 %をスパーが 生産すること,平均するとえき生が頂生より多くを占め ることが認められた。この構造は,樹齢および品種によ りいくらか変化するが,成木樹に共通する性質と考えら 頂生スパ−率(%) 調査樹で長果枝が 6 . 7~ 1 3 . 5 %,中果枝が 3 . 3~ 9 . 6 %と リンゴ樹のスパーが頂生とえき生の 2種類で構成され るため,1年枝に占める全スパーの比率が高ければ,両 スパーの 1年枝に占める比率は直線関係になると考えら れる。そのことを確認するために,両スパーの比率にお ける相互関係を検討した結果,第 2 -1 -4図に示したよ うに,開心形‘ふじ’と‘王林’ ,さらに主幹形‘ふじ’ 50 40 30 20 0 頂生スパ−率(%) 右する要因 王林OC y=83.31.14x (r=0.768**) 60 10 れる。 (2)発生部位別スパー率の相互関係と頂生スパー率を左 70 70 ふじCL y=710.82x (r=0.860**) 60 50 40 30 20 10 0 0 のいずれにおいても,両スパー率は強い負の相関関係に 一定の割合で変化するのであれば,人為的な操作が関与 できる余地はないが,リンゴ‘ふじ’の短果枝率が 1 7年 40 60 80 100 えき生スパ−率 (%) あることを認めた。 頂生とえき生スパーの比率が,樹や枝の年齢とともに 20 第2 -1 -4図 リンゴ‘ふじ’と‘王林’における頂生スパー 率とえき生スパ-率の関係 横軸はえき生スパー率で縦軸は頂生スパー率,OCは 開心形樹,CLは M.2 6台主幹形樹を示す。 リンゴ樹における発生部位別スパー構成 70 第2 -1 -1表 リンゴ‘ふじ’と‘王林’における頂生ス パー率と 2年生長枝率,1年生長枝率,お よび頂生 1年枝あたり新しょう発生数との 各相関関係における決定係数 60 頂生スパ−率 (%) 33 50 品種-樹形 40 開心形ふじ 開心形王林 主幹形ふじ 30 20 2年生長 枝率 (%) 1年生長 枝率 (%) 頂生 1年枝当り 新しょう数(本) 0 . 7 6 0 . 8 1 0 . 7 6 0 . 4 4 0 . 6 8 0 . 1 3 0 . 6 3 0 . 8 1 0 . 5 7 注)各関係式はいずれも対数関数として求めた(第 2 -1 -6 図の回帰式参照) 10 0 4 6 8 10 12 14 16 太枝の年齢 第2 -1 -5図 リンゴ‘ふじ’樹における太枝の年齢(横軸) と頂生スパ-率(縦軸)の相互関係 生に達するまで樹齢とともに増加することが報告されて いる(塩崎・菊池,1 9 8 8 ) 。そこで,開心形‘ふじ’の頂 生スパー率と枝齢の関係を検討した(第 2 -1 -5図)。そ の結果,6年生までの若枝では,頂生スパー率が低く, 長い2年枝の全2年枝に占め る割合(%) 2 100 90 80 70 60 50 40 30 20 10 0 ふじOC 王林OC ふじCL 0 樹齢が進むほど頂生スパー率が高くなる傾向が認められ 10 20 30 40 50 60 70 頂生スパ−率 (%) た。しかし,全体としては,同じ齢でも両スパーの比率 に大きな変異があり,せん定などにより人為的に操作で きる性質であることが認められた。 (3)頂生スパー率と栄養生長および外部形態の関係 頂生スパー率とえき生スパー率は,全スパーに占める 割合が反比例する関係にあり,かつ変化する幅も 0~ 8 0 第2 -1 -6図 リンゴ‘ふじ’と‘王林’における頂生スパー 率と長い 2年枝率(1 0 c m <)の相互関係 OCは開心形樹,CLは M.2 6台主幹形樹。各品種の 回帰式は ふじ- OC:y=- 2 0 . 1 l o gex+ 8 9 . 3(R2= 0 . 7 6) 王林- OC:y=- 2 1 . 2 l o gex+ 8 8 . 5(R2= 0 . 8 1) ふじ- CL:y=- 2 0 . 3 l o gex+ 9 0 . 5(R2= 0 . 7 6) %と大きいため,太枝の性質も各スパー率の変化によっ て大きく異なると考えられる。そこで,頂生スパー率と 太枝の栄養生長との関係について検討した。 タに基づき,頂生スパー率の異なる太枝を模式的に再現 頂生スパー率と 1 0 c m より長い 1 ,2年枝の割合お -1 -7図である。 したものが第 2 よび頂生 1年枝当たり発生新しょう数との関係を示した 頂生 1年枝から発生する新しょうの数を目安にする のが第 2 -1 -1表である。主幹形‘ふじ’の長い 1年枝 と,頂生スパー率の低い方から高い方に向かって連続し 率を除き,いずれの関係も,0 . 5に近いかそれ以上の決 た外部形態の変化が認められ,その違いを判別すること 定係数が得られ,頂生スパー率が減少するにつれて,長 ができる。頂生スパー率の低い枝は,長い 1 ,2年枝が い1 ,2年枝の割合および頂生 1年枝上の発生新しょう 多いため,枝全体にわたり古い枝に対して新しょうが高 数が急激に増加する関係を示した。これらは,第 2 -1 - 密度に存在する状態を示し,逆に頂生スパー率が高くな 6図のように対数関数としてよく適合した。 るにつれ,古い枝が目立ち,新しょうの密度が低くなる 平均頂生新しょう長と頂生スパー率の間にも負の相関 特徴を示した。これらの外部形態的特徴は,品種と樹形 が得られた(図は省略) 。従って,頂生スパー率が低いほ にかかわらず一致した特徴であった。ただ,栄養生長は ど栄養生長が旺盛になることが認められた。なお,主幹 良果生産に適した強さの範囲が存在するため,頂生ス 形‘ふじ’では,頂生スパー率と長い 1年枝率との関係 パー率にも適正範囲が存在すると考えられるが,この点 が密接でなかった。このことは,主幹形樹では,2年枝 は今後の検討課題である。 上に発生する 1年枝の中でスパーを残し,長い,特に側 また,第 2 -1 -1表の結果は,えき生スパー率の高い 生の 1年枝をせん除する強いせん定が行われていること なり枝を作るための方法を示唆している。すなわち,え を示唆している。枝の年齢別に調査した新しょうのデー き生スパー率の高いなり枝では,比較的長い 2年枝が高 浅 田 3 4 100 着花率(%) 80 60 40 20 ふじOC y = 960.66x (r=0.600**) 0 100 王林OC 着花率(%) 80 60 40 20 0 第2 -1 -7図 リンゴ‘ふじ’開心形樹の測定値を用いて再 現した頂生スパ-率の異なる太枝の再現図 A,B,C,Dの頂生スパー率はそれぞれ 10. 5,2 4 . 2, 35. 2 ,61. 2 %である。図中の黒丸は果実を示す。 着花率 (%) 100 80 60 40 20 ふじCL y = 950.75x (r=0.762**) 0 0 10 20 30 40 50 60 70 頂生スパ−率 (%) 比率で存在しており,その 2年枝のえき芽には長い 1年 枝とスパーが着生し,また頂芽には比較的多くの新しょ うを発生させた長い 1年枝が着生するという枝形成の特 第2 -1 -8図 リンゴ‘ふじ’と‘王林’における頂生スパー 率と 1年枝頂芽の着花率の相互関係 OCは開心形樹,CLは M.2 6台主幹形樹 徴を示している。このことから,えき生スパー率の高い なり枝を作るには,適切な位置に比較的長い 1年枝をせ は,次のように考えられる。頂生スパーが形成される過 ん定で残し,そのえき芽から短い新しょうを発生させる 程においては,その基部の果台に果実が着生する場合が ことが必要になる。しかし,それには長い 2年枝,長い あるが,えき生スパーの場合は基部に果台が存在すると えき生 1年枝,さらに着生新しょう数の多い頂生 1年枝 してもえき芽花の果台であるため,その果台の果実は摘 の形成を伴う傾向があった。従って,太枝の枝葉空間の 除される。そのため,頂生スパーの方がより近い位置に 拡大につながらないえき生スパー形成法を検討する必要 果実が存在し,花芽形成が阻害されやすいためと考えら がある。 れる。本調査の結果,密植主幹形樹は開心形樹に比べ, 頂生スパー率が高くなると花芽分化がより強く阻害され (4)頂生スパー率と花芽形成の関係 たため,頂生スパー率を高くすることには限界があるこ 頂生スパー率と太枝の生殖生長との関係について検討 とを示唆している。ただ,頂生スパー率と花芽形成の関 した。1年枝の頂芽が花を着生した割合を着花率として 係については,再現性および品種間差異等さらに検討す 頂生スパー率との関係を示したのが第 2 -1 - 8図であ る必要がある。なお,‘王林’樹では,頂生スパー率と着 る。主幹形‘ふじ’樹の着花率は,いずれの頂生スパー 花率との間に有意な相関関係がみられなかったが,その 率でも 6 0 % 以 上 を 維 持 し た が, 負 の 相 関 関 係(r= 原因として前年の夏季の高温による花不足が関係してい -0 . 7 6 2**)を示した。開心形‘ふじ’でも,頂生スパー ると考えられた。 率が高くなるにつれ着花率の上限が低下する関係が認め 以上のように,頂生スパー率はその増加が花芽形成の ** られた(r=-0 . 6 0 0 ) 。しかし,開心形‘王林’では, 減少と関係し,一方で次項に述べるように葉面積当たり 頂生スパー率に関わらず着花率は大きく変動した。頂生 果数が増加することを伴うため,果実生産にとって望ま スパー率の高い太枝の方が着花率が劣る原因について しい適正範囲が存在すると考えられる。‘スパー枝齢’ が リンゴ樹における発生部位別スパー構成 35 60 Tree1 50 Tree2 頻度割合(%) Tree3 40 Tree4 Tree5 30 20 10 0 < 2.0 2.02.9 3.03.9 4.04.9 5.05.9 6.06.9 葉面積当たり果数 (果 7.07.9 8.08.9 9.0 < /m2) 第2 -2 -1図 リンゴ‘ふじ’における葉面積当たり果数の園地間と樹内変異 果実品質に影響すること(Ro bi ns o nら,1 9 8 3 ;J a c ks o n, せん定をより簡明に会得することに役立つものと考えら 1 9 6 7 ;Ro m・Ba r r i t t ,1 9 9 0 ;Vo l zら,1 9 9 4 ) ,また,果台 れる。 から果台枝が発生する力に品種間差異があること(La ur i ら,1 9 9 5 )も報告されており,頂生スパー率の適正範囲 について今後さらに検討する必要がある。また,実際栽 培では,品種によっては中果枝の方がスパーより果実品 2.スパーの発生部位別構成割合と果実生産性 eafar ea)の樹 (1)葉面積当たり果数(FLA,Fr ui t sperl 冠内変異 質が優れたり,整枝上都合がよい場合も見受けられるの Fe r r e e (1 9 8 0 )は,リンゴ‘ゴールデン・デリシャス’ で,この点についても検討する必要がある。 では,樹冠を高さで 3等分した上部における FLAが下 本項では,スパーの発生部位別構成割合と栄養および 部と中部より大きいと報告している。 本調査では, 生殖生長との間に存在する基本的な関係を明らかにし FLAは全供試樹において 0 . 7から 1 3 . 3まで変異し,平均 た。この関係の成立には,スパーの 1年枝に占める割合 5 . 0であった(第 2 -2 -1図) 。この平均値は Fe r r e eに が過半を占め,極端に低くないことが前提となる。本調 よって報告された樹冠下部の FLAと一致する。本研究 査では,幅広い強さの栄養生長を示す太枝を選んで検討 で得られた比較的低い FLA値については,品種と望ま したが,いずれの品種及び樹形においても全スパー率 5 0 しい果実の大きさ(浅田,19 8 9 )の違いによると考えら %以下の太枝は存在しなかった。また,スパー率が極端 れる。 に低い樹は栽植直後の若木あるいは強樹勢の樹であっ て,実際栽培上極めて非生産的な木である。以上のこと から,この基本的関係は,リンゴ樹が有する生長特性に (2)FLAに及ぼす太枝の枝齢,発出角度,発出部位高お よび栄養生長の強さの影響 基づくもので,栽培に供されているリンゴ樹に広く成り FLAの樹間および樹冠内変異はかなり大きく,その 立つものと考えられる。 ため変異の原因について発生部位別スパー率を中心に検 スパーの種類別構成割合は,せん定によって制御でき 討した。FLAと太枝の諸測定値との相関関係を示した る可能性がある。それには,各スパーを人為的に増減さ のが第 2 -2 -1表である。FLA は枝齢が進むにつれ増 せるせん定法が必要になるが,えき生スパーが長い 2年 加する傾向を示し(r= 0 . 4 4 6**),6年生になるまでの 枝に着生することと,そのえき生スパーから頂生スパー 若枝ではとくに小さかった(第 2 -2 -2図)。しかし,太 が形成されることがスパー形成の起点となることに注目 枝の発出角度,高さ,栄養生長の強さは FLAと密接な する必要がある。そして,えき生スパー率を高めるせん 関係を示さなかった。これらの性質がリンゴの果実品質 定方法および長い 1年枝の比率を減少させながら頂生ス に影響を及ぼすことを示す報告がある(Le s pi na s s e ・ パー率を増加させる方法等について検討することによ De l o r t ,1 9 8 6 ;Tus t i nら, 1 9 8 8 ;Mye r s ・Fe r r e e ,1 9 8 3 ) 。 り,スパーの発生部位別構成割合を制御するせん定法を しかし,本研究では,枝齢を含むこれらの性質は太枝間 確立できると考えられる。また,スパーの発生部位別構 の FLAの違いをほとんど説明しなかった。従って,調 成割合の違いが眼で判別できることから,その判別法を 査した成り枝は集約的なせん定によって望ましい生理状 明らかにすることにより習熟に多くの経験を必要とする 態に調整されていると考えられる。 浅 田 第2 -2 -1表 リンゴ‘ふじ’における葉面積当たり果数 (FLA)と太枝の枝齢, 発出角度,発出部 位高,樹勢との相関関係 太枝の性質 相関係数(r ) 枝齢 発出角度 発出部位高 樹勢 0. 446** 0. 305** - 0. 051 - 0. 314** 14 葉面積当たり果数(果 / ㎡) 3 6 Tree 1 12 Tree 2 10 Tree 3 8 Tree 4 6 Tree 5 4 2 0 1.5 0 Tree 1 Tree 2 Tree 3 Tree 4 5 Tree 5 15 枝齢 (年) 第2 -2 -2図 リンゴ‘ふじ’における葉面積当たり果数 (FLA)と太枝の年齢との関係 1 0.12 Tree 1 Tree 2 0.5 Tree 3 Tree 4 Tree 5 0.1 0 −0.5 0.5 1 1.5 2 2.5 3 BPR(新しょう数/芽) 3.5 第2 -2 -3図 リンゴ‘ふじ’における葉面積当たり果数 (FLA)と 1年枝当たり新梢発生数(BPR) の関係 各樹の回帰式は, Tr ee1 : y= 1. 33- 0. 28x,r=- 0. 821** Tr ee2 : y= 1. 21- 0. 34x,r=- 0. 800** Tr ee3 : y= 1. 18- 0. 33x,r=- 0. 873** Tr ee4 : y= 1. 12- 0. 33x,r=- 0. 624** Tr ee5 : y= 1. 02- 0. 35x,r=- 0. 663** (3)FLAと頂芽増加率(BPR:Budpr oduct i onr at e)お よび 1年枝当たり葉面積の相互関係 BPRは 1年枝当り発生新しょう数で表した。FLAの 常用対数値は BPRと負の直線関係を示し,それぞれの 1年枝当たり葉面積 (㎡) log FLA 10 0.08 0.06 0.04 0.02 0 0.5 1 1.5 2 2.5 3 BPR (新しょう数/芽) 3.5 第2 -2 -4図 リンゴ‘ふじ’における 1年枝当たり葉面積 と 1年枝当たり新しょう発生数(BPR)の関 係 各樹の回帰式は, Tr e e1: y=- 0 . 0 0 4+ 0 . 0 3x,r= 0 . 9 0 9** Tr e e2: y= 0 . 0 0 3+ 0 . 0 2x,r= 0 . 8 0 7** Tr e e3: y=- 0 . 0 1 3+ 0 . 0 3x,r= 0 . 9 6 0** Tr e e4: y=- 0 . 0 0 0 2+ 0 . 0 3x,r= 0 . 9 0 8** Tr e e5: y=- 0 . 0 0 8+ 0 . 0 3x,r= 0 . 8 6 7** 樹で固有の回帰式を示した(第 2 -2 -3図) 。この関係が 成立した背景には,次の条件が存在している。(1 )手摘 するために BPRと 1年枝当たり葉面積を小さくするこ 果により 1年枝の頂芽に果実を着生させている,(2 )花 と が 重 要 で あ る こ と を 示 唆 し て い る。Po r t e ra nd 芽形成の抑制を避けるため(Ko i ke ,1 9 9 0 ) ,1年枝当たり Ll e we l yn(1 9 8 4 )は,せん定で残された芽の割合とその の着果数に上限を設けていること, (3 )1年枝当たり葉 後の BPRの常用対数値との間に負の直線関係があるこ 面積が BPRの増加とともに大きくなる,の条件が存在 とを報告した。彼らの試験では,切り返しせん定が行わ している。本研究では,各供試樹において BPRと 1年 れているので,BPRや 1年枝当たり葉面積が増加し, 枝当り葉面積の間に密接な相関が認められた(第 2 -2 - FLAを大きくするのには適切とは考えられない。ただ, 4図) 。 果実肥大が劣るような弱い生長状態にある太枝では当て FLAと BPRの相関関係は,太枝の果実生産性を改善 はまらない。 リンゴ樹における発生部位別スパー構成 1 ,2年枝の種類 年齢 長さ 1 1 1 1 2 2 2 2 長 長 短 短 長 長 短 短 z 長:> 0.08 リンゴ‘ふじ’の太枝における 1年枝当た り葉面積と 1,2年枝の種類別構成割合の 関係 z 相関係数(r ) 発生部位 頂生 えき生 頂生 えき生 頂生 えき生 頂生 えき生 - 0. 031 0. 547** - 0. 597** 0. 188 - 0. 301** 0. 524** - 0. 469** - 0. 066 0.07 1年枝当たり葉面積(㎡) 第2 -2 -2表 37 0.06 0.05 0.04 0.03 0.02 0.01 0 10 c m,短:5 c m≧ 10 20 30 40 50 60 70 頂生スパ−率 (%) 第2 -2 -5図 (4)1年枝当たり葉面積を左右する要因 前述のように,1年枝当たり葉面積は BPRと正の相 関関係にあるため,それを左右する要因について検討し た。 リンゴ‘ふじ’の太枝における頂生スパ-率 と 1年枝当たり葉面積の関係 頂生スパ-率 3 0 %以下の回帰式: y= 0 . 0 7- 0 . 0 0 1x(r=- 0 . 7 2 9**) 頂生スパ-率 3 0 %以上の回帰式: y= 0 . 0 4- 0 . 0 0 0 4x(r=- 0 . 4 0 9**) 第 2 -2 -2表は分類した各 1 ,2年枝の割合と 1年枝 当たり葉面積の相関関係を示している。1年枝当たり葉 16 面積は,長いえき生の 1 ,2年枝の割合と正の,また短い 頂生の 1 ,2年枝の割合と負の相関を示している。長い 14 頂生と短いえき生の 1 ,2年枝の割合は 1年枝あたり葉 面積と強い関係は認められなかった。長いえき生枝が少 12 なく,短い頂生枝が多い成り枝は,高い FLAをもつこ 10 Ma ggs (1 9 5 9 )は,若い樹では,せん定が短い新梢の 割合を減少させることを報告した。しかし,樹が果実を 生産し始めるときには,剪定で除かれる枝のタイプと場 所によって,せん定は短い頂生の枝の割合を増やす可能 性がある。そのようなせん定とは,“長いえき生枝を除 去するせん定”であると考えられる。すなわち,前年に 作られた枝のえき芽から発出した長い 1 ,2年枝が過剰 FLA とが示唆された。 8 6 4 2 な栄養生長を刺激しない程度に取り除かれるせん定であ る。長いえき生枝は 1本かあるいはそれ以上の長い新梢 0 を発出させ,その基部にある短い枝の生長を弱くする。 そして,長い頂生枝に比べれば,その枝を除去しても芽 の生長を強く刺激することはない。従って,短い 1 ,2 年枝の割合をこのようなせん定方法により比較的高く維 持するのがよいと考えられる。 1年枝当たり葉面積は,短い頂生 1年枝の割合が 3 0 % 以下になると,急激に増加する(第 2 -2 -5図) 。従っ 0 %になることが適正な剪定 て,頂生スパーの比率が 3 の目安であると考えられる。 0 10 20 30 40 50 60 頂生スパ−率 (%) 70 ●Tree 1, ○Tree 2, △Tree 3, □Tree 4, *Tree 5 第2 -2 -6図 リンゴ‘ふじ’の太枝における頂生スパ-率 と葉面積当たり果数(FLA)の関係 各樹の回帰式は, Tr e e1:y= 4 . 4 8+ 0 . 1 5x,r= 0 . 5 4 3** Tr e e2:y= 1 . 6 7+ 0 . 0 9x,r= 0 . 7 0 1** Tr e e3:y= 2 . 8 4+ 0 . 1 0x,r= 0 . 4 3 0** Tr e e4:y= 2 . 2 9+ 0 . 1 1x,r= 0 . 4 5 2** Tr e e5:y= 3 . 0 6+ 0 . 0 1x,r= 0 . 1 3 0 (5)FLAと頂生スパー率の関係 各樹における FLAと頂生スパー率の関係を第 2 -2 - Tr e e5を除いて,頂生スパー率と FLAの間には正の 6図に示した。 有意な相関が認められた。この結果は,Tr e e5が 3 . 4 9 浅 田 3 8 第2 -2 -3表 リンゴ‘ふじ’の葉面積当たり果数,結果率,着花,スパー率,および樹勢における樹間差異 項 目 葉面積当たり 果数(果/ ㎡) 結果率(%) 開花率(%) スパー率(%) 樹勢(c m)y 樹の No . 1 2 3 4 5 z 7. 97a 32. 3a 82. 6ab 74. 8b 26. 7c 4. 7 2 b 15. 7 d 68. 2 c 81. 0 a 35. 3 a b 4 . 8 2 b 2 1 . 3 b 8 4 . 4 a b 7 5 . 5 b 3 6 . 9 a 4 . 6 4 b 1 7 . 8 c 8 9 . 1 a 8 1 . 1 a 3 0 . 0 bc 3 . 4 9 c 1 0 . 9 d 8 0 . 6 b 8 2 . 1 a 3 0 . 1 bc Z 異なる文字の付いた樹間の数値には,S hef f e’ sF t es tにより Y 5 %水準で有意差がある 頂生の 1年枝の頂芽から生長した新梢の平均長 と小さい FLAをもつ木(次項(6 )第 2 -2 -3表を参照) 第2 -3 -1表 で着果不足とみられるので,充分な着果状態にある樹で 1年枝の長さ は,頂生スパー率が増加するとともに FLAが大きくな ることを示している。 (6)FLAと BPRの相関関係における樹間変異 FLAの対数値と BPRの回帰直線は有意な樹間変異が 認められたが,傾きは同じであった(第 2 -2 -3図)。 No .1の樹の太枝は,BPRのどの値においても FLAが Z 他の樹より高かった。すなわち,回帰直線の Y切片の Y 値が他の樹より有意に大きかった。このことは,No .1 リンゴ‘ふじ’における着果が頂端新しょ う長に及ぼす影響 頂端新しょう長(c m) (c m) 着果 ≦0 . 5① Z ≦0 . 5② 5 . 1~ 1 0 . 0 >1 0 . 0 1 5 . 0 Y 1 3 . 7 a 2 1 . 9 3 1 . 0 c 不着果 (花) 1 5 . 4 9 . 1 b 2 4 . 2 3 8 . 8 b 葉芽 1 5 . 5 3 . 8c 2 4 . 6 4 7 . 8a ①は頂生スパー,②はえき生スパー 異なる文字の付いた 1年枝頂芽の種類間には Shef f e ’ sF t e s tにより 5 %水準で有意差がある の樹が他の樹に比べて FLAを高くする要素を備えてい ることを示唆している。そこで,結実に関係する要因に とを見出したので整理した。調査した枝は,‘ふじ’ 成木 ついて樹と樹の比較を行った(第 2 -2 -3表) 。No .1の 5樹(材料および方法を参照)の全 1年枝である。 樹は,他の樹に比べ結果率(1年枝当たり着果数)が有 果実はリンゴ樹の中で最も強いシンクである(望月, 意に高かった。しかし,スパー率,開花率,樹勢には違 1 9 6 2 ;Ma ggs ,1 9 6 3 ) 。着果により新しょう生長が抑制 いが認められなかった。発育と着色のよい果実は細い枝 され,スパー形成が影響されると考えられる。リンゴに より太い枝で生産されることが,一般的に知られている おける新しょうは貯蔵養分と同化養分により生長する s t o phe r ,1 9 7 4 ) 。No .1の木の高結果率は太枝に (Chr i が,どちらの養分がどの程度影響するかは明らかでな 着生するスパーが太いことも影響していると推察され い。従って,着果が新しょうへの同化養分の供給を抑制 る。 するとしても,新しょう生長がどの程度阻害されるかは 以上のように頂生スパー率の増加は,花芽形成を減少 不明である。隔年結果と新しょう生長の関係から,新 させ,一方で葉面積当たり果数を増加させるように働く しょう生長がなり年に旺盛であるとする報告(Fo r s he y, ため,果実生産にとって望ましい適正範囲が存在すると 1 9 8 2 b;Cur r y・Lo o ne y,1 9 8 6 )や着果過多の影響が翌年 考え ら れ る。ス パー枝齢が果実品質に影響する こ と に現れるとする報告(Mo c hi z uki ,1 9 6 2 )が存在する。着 (Ro bi ns o nら,1 9 8 3 ;J a c ks o n, 1 9 6 7 ;Ro m・Ba r r i t t , 果と当年の新しょう生長の関係を明らかにするには,貯 1 9 9 0 ;Vo l zら,1 9 9 4 ) ,また,果台から果台枝が発生す 蔵養分が同じ条件で着果と不着果の影響を検討する必要 る力に品種間差異があること(La ur iら,1 9 9 5 )も報告 がある。しかし,これまでの報告では,若木を供試した されており,頂生スパー率の適正範囲について今後さら 樹全体で着果と不着果処理区を設ける試験が多い(Er f ・ に検討する必要がある。なお,実際栽培では,品種に Pr o c t o r ,1 9 8 7 )。その場合は着果が根の発達を抑制し, よって中果枝の方がスパーより果実品質が優れたり,整 樹全体の生長を阻害する。一方,実際栽培では,樹全体 枝上都合がよい場合も見受けられるので,品種間差異に では適正着果量で管理されており,根への同化養分の供 ついても検討する必要がある。 給が充分でありながら,果実の着生する周辺,例えば果 台枝などの新しょうのみが果実との同化養分の競合に遭 3.発生部位別スパー形成に及ぼす着果の影響 遇することになり,着果の影響の仕方が異なると考えら スパー形成要因については第 5章で取り上げるが,本 れる。そこで,実際栽培園において,樹内の着生条件の 章の調査資料からスパー形成要因について検討すべきこ 異なる新しょう,すなわち,葉芽,着果している果台, リンゴ樹における発生部位別スパー構成 39 第2 -3 -2表 リンゴ‘ふじ’の新しょう生長に及ぼす着果の影響 1年枝の長さ 1年枝の頂芽の種類 (c m) 着 果 ≦ 0. 5① Z ≦ 0. 5② 5. 1~ 10. 0 > 10. 0 1. 0 0. 7bY 1. 6b 3. 7b ≦ 0. 5① ≦ 0. 5② 5. 1~ 10. 0 > 10. 0 12. 9 12. 0a 20. 1 50. 2b 不着果(花) 葉 芽 1年枝当たり新しょう発生数(本) 1 . 1 0 . 4 b 1 . 8 b 4 . 6 b 1年枝当たり発生新しょう総長(c m) 1 3 . 5 7 . 1 a b 2 8 . 3 7 1 . 6 a b 1 . 5 1 . 0 a 2 . 7 a 8 . 4 a 1 6 . 0 4 . 3 b 2 7 . 1 9 6 . 8 a Z ①は頂生スパー,②はえき生スパー Y 異なる文字の付いた 1年枝の種類間には Shef f e’ sF t e s tにより 5 %水準で有意差がある 不着果の果台から発生する新しょうの生長比較を行っ た。 第2 -3 -3表 1年枝を長さにより 3つに分類し,スパーはさらに頂 1年枝の長さ 生とえき生に分けた。また,1年枝の頂芽の状態により 化しなかった葉芽枝の 3種類に分けた。従って,着果枝 と不着果枝から発出する新しょうは果台枝となる。1年 枝の頂芽から生長した頂端新しょう長に及ぼす着果の影 響を第 2 -3 -1表に示した。 生スパーと 1 0 c m 以上の 1年枝であった。1 0 c m 以上の 1年枝は,着果枝が最も短く,葉芽枝が最長であり,1 6 . 8 Z Y 不発生率 (%) (c m) 着果 不着果 (花) 葉芽 ≦0 . 5① Z ≦0 . 5② 5 . 1~ 1 0 . 0 >1 0 . 0 Y 1 4 . 7 a 3 0 . 8 a 4 . 0 a b 4 . 0 a 1 8 . 7 a 3 4 . 1 a 4 . 8 a 1 . 8 a b 0 . 7b 0 . 2b 0 . 1b 0 . 1b 着果枝,開花後果実を着けなかった不着果枝,花芽を分 着果により頂端新しょう長に違いがあったのは,えき リンゴ‘ふじ’における頂芽の新しょう不 発生率に及ぼす着果の影響 ①は頂生スパー, ②はえき生スパー 異なる文字の付いた結果枝の種類間には She f f e’ sF t es t により 5 %水準で有意差がある c m の違いがあった。しかし,えき生スパーでは,逆に 葉芽,不着果,着果の順で新しょうが長くなった。頂生 を発生させない枝が葉芽より多かった。えき生スパーが スパーは着果に関係なく同じ新しょう長であった。 3 0 %以上と最も多く,次いで頂生スパーが 1 5ないし 1 9 次に,えき生新しょうを含めた 1年枝当たり新しょう % で 多 く,5 c m よ り 長 い 1年 枝 は,5 %以下と少な 数と総長について検討した。結果を第 2 -3 -2表に示し かった。ただ,着果と不着果の間で違いがみられなかっ た。発生新しょう数は,頂生スパーを除いて,葉芽枝が た。 着果枝と不着果枝より多かった。着果と不着果では違い 1年枝上に形成される 5 c m 以下の新しょう(当年生の がなかった。頂生スパーの葉芽からは,スパーにもかか スパー)について検討した。結果を第 2 -3 -4表に示し わらず平均 1 . 5本発生したが,5 c m 以下の長さでも葉 た。えき生スパーは,着果枝と不着果枝の方が葉芽枝よ 芽であればえき生新しょうを発生させる場合もあること り2 . 5倍多く当年生頂生スパーを形成した。1 0 c m 以上 を示した。 の 1年枝では,着果枝が葉芽枝より約 3倍多く当年生頂 1年枝当たり発生新しょう総長は,えき生スパーと 1 0 生スパーを形成した。頂生スパーと 5~ 1 0 c m では,当 c m 以上の 1年枝において,着果枝と葉芽枝との間に違 年生頂生スパーの形成に着果の影響はみられなかった。 いがみられた。1 0 c m 以上の 1年枝では,着果枝が葉芽 また,1年枝のえき芽から形成される当年のスパー(当 6 c m もの違いがあった。 枝より短く,1年枝当たり 4 6 . 年生えき生スパー)について検討した(第 2 -3 -4表) 。 えき生スパーでは,逆に葉芽枝が 4 . 3 c m と着果枝より 0 . 1本以上の当年生えき生スパーが形成されるのは,5 も短かった。頂生スパーでは,着果にかかわらず同じ長 c m 以上の 1年枝であり, 着果の影響もその枝のみで さであった。 みられた。当年生頂生スパーとは反対の結果が得られ, 1年枝における新しょうの不発生率に及ぼす着果の影 葉芽の方が着果と不着果枝より多くの当年生えき生ス 響について検討した。 結果を第 2 -3 -3表に示した。 パーを形成した。 葉芽は 1年枝の長さにかかわらずほとんど全て新しょう 着果が新しょう生長に及ぼす影響について検討するた を発生させた。しかし,着果および不着果枝は新しょう め,1年枝の頂芽が着果,不着果,葉芽の 3種類につい 浅 田 4 0 第2 -3 -4表 リンゴ‘ふじ’における当年生スパー(5 c m 以下の新しょう)数に及ぼす着果の影響 1年枝の長さ 1年枝の頂芽の種類 (c m) 着 果 ≦ 0. 5① Z ≦ 0. 5② 5. 1~ 10. 0 > 10. 0 0. 16 Y 0. 14a 0. 19 0. 07a < 0. 5① < 0. 5② 5. 1~ 10. 0 > 10. 0 0. 02 0. 0 0. 03 0. 23b 不着花(果) 葉 芽 1年枝当たり当年生頂生スパー数(本) 0 . 1 3 0 . 1 4 a 0 . 1 4 0 . 0 4 a b 1年枝当たり当年生えき生スパ-数(本) 0 . 0 3 0 . 0 0 . 0 5 0 . 4 0 b 0 . 1 1 0 . 0 6 b 0 . 1 1 0 . 0 2 b 0 . 0 4 0 . 0 0 . 1 3 0 . 6 7 a Z ①は頂生スパー,②はえき生スパー Y 異なる文字の付いた結果枝の種類間には Shef f e’ sF t e s tにより 5 %水準で有意差がある て 1年枝上の新しょう生長を比較した。このうち着果と ている。特に基部周辺のえき芽は小さい芽が多く,それ 不着果は明らかな違いがみられなかった。不着果も摘果 らから作られるスパーは小さい。そのような芽が葉芽に されるまでは着果していたのであり,生育の中・後期に なった場合は,花芽を形成したスパーより性質が劣るこ 果実が存在しないという違いとみられる。調査項目が生 とになると考えられる。 育の前半に決定される性質であれば,着果と不着果は同 スパー形成に及ぼす着果の影響は,1年生えき生ス じ条件となる。したがって,着果の影響は,葉芽との違 パーと 1 0 c m より長い 1年枝からの当年生頂生スパー いを吟味するのが妥当と考えられる。 数において大きいが,同えき生スパーには小さい。結 着果・不着果が葉芽と新しょう生長において異なった 局,両者の差し引きになるので,全体として着果は翌年 点は,1 0 c m より長い 1年枝の頂端新しょう長が短いこ のスパー形成を抑制することを本調査は示唆している。 と,1年枝当たり新しょう発生数が少ないこと(頂生ス パーを除く) ,1年枝から新しょうが全く発生しない割 摘 要 合が高いこと,当年生頂生スパー数が増えること,1 0 c m より長い 1年枝に形成される当年生えき生スパー数 リンゴのマルバカイドウ台開心形‘ふじ’および‘王 が少ないことである。以上の性質が着果により影響され 林’樹と M.2 6台主幹形‘ふじ’樹を供試し,冬季せん ることを本調査は示唆した。したがって,本調査は,着 定直後の長さ 5 c m 以下の短果枝(スパー)を,発生部 果により当年の新しょう生長は抑制されることを示して 位によって頂生とえき生の 2種類に分け,それらの全 1 いる。ただ,以上の着果の影響が現れない 1年枝もあっ 年枝に占める割合と栄養・生殖生長や果実の生産性との た。それは,1年生の頂生スパーであった。1年生頂生 関係および両スパーの形成に及ぼす着果の影響について スパーの頂端新しょう長,新しょう総数,総長,頂生ス 検討した。 パー数では,1年生頂生スパーの頂芽に果実がなって 頂生スパーは主として 5 c m 以下の 2年枝の頂芽に, も,あるいは葉芽でも同じ数と長さの新しょうが発生し えき生スパーは 1 0 c m 以上の 2年枝のえき芽から形成 た。一般に,果実がなると新しょうの生長が阻害される されたが,両者の 1年枝に占める比率は,開心形と主幹 と考えられているが,この観察は頂生スパーに当てはま 形‘ふじ’,開心形‘王林’でそれぞれ 2 4 . 7 %と 5 6 . 9 %, らなかった。着果した 1年生頂生スパーから頂端新しょ 2 4 . 6 %と 5 6 . 7 %,3 0 . 2 %と 4 6 . 5 % で あ っ た。頂 生 ス うが平均 1 5 c m 伸びていることをみると,せん定等の管 パー率は最低 1 . 4 %から最高 6 2 . 8 %まで,えき生スパー 理により比較的強い生長力をもつ頂生スパーを育成して 率は同 2 2 . 4 %から 8 5 . 2 %までの範囲で変化し,両者の いるため,着果の影響がみられなかったと考えられる。 比率は互いに反比例に類似した関係を示した。長い 2年 さらに,1年生えき生スパー上の新しょう形成は特異 枝の割合が高いほどえき生スパー率が高く,頂生 1年枝 的であった。すなわち,他の 1年枝とは異なり,葉芽の 当たり発生新しょう数が増加し,長い新しょうが密度高 頂端新しょう長と 1年枝当たり新しょう総長が着果・不 く樹冠を形成する外部形態を示した。主幹形‘ふじ’で 着果より短く,着果の影響とは反対の結果を示した。こ は,頂生スパー率と 1年枝の頂芽における着花率の間に れは,えき生スパーの性質に原因があると考えられる。 有意な負の相関が認められた(R2= 0 . 5 8 )。以上の結果, すなわち,えき生スパーは長い 1年枝のえき芽から形成 樹冠の外部形態から識別できるスパーの発生部位別構成 されるが,えき芽は位置により大きさがさまざま異なっ のあり方を,せん定等における樹体調節の目安として利 リンゴ樹における発生部位別スパー構成 41 第3 -1表 リンゴにおけるスパーの種類が果実品質に及ぼす影響 スパーの種類 果 重 (g) 横 径 (mm) 硬 度 (l b) 糖度含量 (°Br i x) 滴定酸 (g/ 1 0 0ml ) えき芽 1年 2年以上 268. 4bZ 372. 1a 370. 5a 86. 8b 95. 1a 94. 3a ふ じ 1 4 . 5 a 1 3 . 1 b 1 2 . 5 c 1 4 . 2 1 3 . 9 1 3 . 9 0 . 3 1 0 . 3 2 0 . 3 2 6 4 . 5 6 8 . 3 7 0 . 8 えき芽 1年 2年以上 320. 4ab 333. 5a 309. 1b 86. 7ab 87. 2a 85. 2b 王 林 1 2 . 2 1 2 . 4 1 2 . 5 1 5 . 0 1 4 . 8 1 4 . 5 0 . 2 0 0 . 2 1 0 . 2 1 - - - えき芽 1年 2年以上 345. 2b 425. 7a 395. 7ab 93. 4b 98. 9a 96. 8a ジョナゴールド 1 0 . 6 9 . 6 1 0 . 4 1 3 . 4 1 3 . 9 1 3 . 8 0 . 4 4 0 . 4 5 0 . 4 5 3 1 . 4 b 5 0 . 2 a 4 5 . 7 ab Z 異なる文字の付いた年数の数値間には 着色割合 (%) Tukeyの多重検定により 5 %水準で差がある 用可能なことが示唆された。 リンゴ‘ふじ’における太枝の果実生産性,すなわち 葉面積当たり果数(FLA)は,0 . 7から 1 3 . 3まで分布し, 平均値は 5 . 0であった。太枝の年齢,発出部位高,発出 角度,栄養生長の強さは FLAの変異を説明する主要な 第3章 スパー枝齢と果実品質および果台枝形 成の関係 緒 言 要因ではなかった。FLAと芽の増加率(BPR)の間には 頂生スパーは,2年枝の頂端に形成されるものからス 負の相関関係が認められた。BPRの増加は 1年枝当た パーが連年継続して生長して形成されるもの,また太い り葉面積の増加を伴う(r> 0 . 8 3 1**)ため,1年枝当た 枝から直接発生するスパーなど違いがみられる。スパー り葉面積を増加させる要因について検討した結果,1 ,2 を年齢別に分類した報告(Ro bi ns o nら,1 9 8 3 )によると, 年生えき生長枝(> 1 0 c m)の割合が低いほど,また 1 , 太枝を枝齢で区分し,各区分に形成されているスパーに 2年生頂生短枝(≦ 5 c m)の割合が増加するほど 1年枝 対して区分の年齢を当てはめ,スパー枝齢(s pura ge ) 当たり葉面積が大きくなることを認めた。1年生頂生短 としている。この定義によると,同じスパー枝齢であっ 枝(頂生スパー)の 1年枝に占める割合が 3 0 %以下にな ても,その年に太枝に直接形成されたスパーか,何年か ると,1年枝当たり葉面積が急激に増加した。FLA と スパー生長が継続して形成されたスパーか不明である。 頂生スパー率との間に着果率の低い木を除いて正の直線 一方,スパーの年齢は全て 1年枝あるいは当年枝しか存 関係が認められた。FLAと BPRの関係は供試樹間で違 在しない,という考えもある。しかし,「スパー枝齢」と いがみられ,BPRに対し常に高い FLAを有する木は着 いう用語は,その枝の年齢を意味するのでなく,「ス 果率(全 1年枝に対する着果した 1年枝の割合)が高かっ パー生長を繰り返した年数」と定義することも可能であ た。 る。本研究では,スパー生長の継続年数を「スパー枝齢」 1年枝を頂芽の状態によって,着果,不着果,葉芽の と称する。従って,古い太枝に直接潜芽から形成された 3種類に分け,1年枝から発生する新しょうの生長を比 1年生のスパーは,1年生スパーとする。 較することによりスパー形成に及ぼす着果の影響を検討 スパー枝齢の違いは,貯蔵養分,近辺に着生する葉量, した。着果あるいは不着果の 1年枝が葉芽の 1年枝と異 太い骨組み枝との距離など生長条件が異なるとみられ なる点は,頂端新しょう長が短いこと,えき生をも含む る。スパーに着果させた場合,それらの条件の違いが果 1年枝当たり新しょう数と総長が劣ること,新しょうの 実品質や花芽形成に影響を及ぼす可能性がある。リンゴ 不発生率が高いこと,頂生スパー数が多いこと,および では,スパーの着生位置と果実品質の関係は,主にえき えき生スパー数が少ないことであった。ただ,1年枝で 芽果とスパー果で比較されており(Vo l zら,1 9 9 4 ) ,ス も頂生スパーは,頂芽に着果してもしなくても,また葉 パー枝齢の違いについては不明な点が多い。なお,ニホ 芽であっても新しょうの数と長さに違いはなかった。ま ンナシでは果実肥大のすぐれる側枝年齢について明らか た,えき生スパーは他の 1年枝と異なり,葉芽の方が着 にされている(田村ら,19 9 0 )。そこで,スパー枝齢の違 果・不着果より頂端新しょうと新しょう総長が短かった。 いによって果実品質及び周辺の葉量,さらに果台枝形成 にどのような違いがあるか検討した。 浅 田 4 2 第3 -2表 スパーの種類が果そう葉数と果そう最大葉の 大きさに及ぼす影響 スパーの種類 果そう葉数 大きさ(mm) 縦 径 横 径 48. 6b 56. 4a 55. 3a 29. 1 b 39. 4 a 38. 5 a えき芽 1年 2年以上 ふ じ 5. 1bZ 6. 5a 6. 4a えき芽 1年 2年以上 4. 7b 5. 6a 5. 6a 52. 0b 62. 6a 60. 8a 31. 5 b 40. 3 a 37. 7 a えき芽 1年 2年以上 ジョナゴールド 4. 9 53. 7b 6. 4 58. 9a 6. 4 59. 1a 38. 4 b 48. 4 a 47. 4 a 王 林 Z 異なる文字の付いた種類の数値間には Tukeyの多重検定 第3 -3表 スパー枝齢の違いが果台枝の発生率,長さおよ び太さに及ぼす影響 スパーの種類 ふ じ 王 林 ジョナゴールド えき芽 1年 2年以上 果台枝の発生率(%) -* 1 0 0 . 0 1 0 0 . 0 8 6 . 6 9 8 . 9 9 4 . 1 9 0. 0 8 3. 2 8 5. 8 えき芽 1年 2年以上 果台枝の長さ(c m) - 9 . 5 1 1 . 0 1 0 . 7 9 . 8 7 . 6 8. 3 1 1. 4 1 4. 4 えき芽 1年 2年以上 果台枝の太さ(mm) - 3 . 2 2 . 7 a 3 . 3 2 . 1 b 3 . 3 3. 4bZ 4. 1a 4. 3a Z 異なる文字の付いた種類間の数値には Fi s he r ’ sPLSDの検 定により 5 %水準で有意差がある * 測定せず により 5 %水準で有意差がある 第3 -4表 材料及び方法 弘前大学農学生命科学部付属生物共生教育研究セン スパー枝齢 ター藤崎農場に植栽されている 1 5年生の‘ふじ’ ,‘王 林’ , ‘ジョナゴールド’各 5樹を供試した。いずれもマ ルバカイドウ台開心形樹である。各品種各樹当たり 6本 の太枝を選び,5月末,着果しているスパー(長さ 5 c m 以下)と 1年枝のえき芽(以下えき芽という)に分けラ 1年 2年以上 1年 2年以上 スパー枝齢が翌春における果台枝の開花率に 及ぼす影響 ふじ 王林 頂芽の開花率(%) 3 4 . 7 7 8 . 6 3 5 . 7 6 3 . 9 えき生芽の開花率(%) 2 2 . 5 4 5 . 2 1 4 . 3 2 5 . 0 ジョナゴールド 6 6 . 7 6 9 . 0 3 3 . 3 2 4 . 1 ベルを付けた。2年枝にえき生するスパーを 1年生ス パーとする。 試験 1では,摘果は,各果実が 2 0~ 3 0 c m の距離を 穫直前に果実周辺の葉量として果そう葉,果実から枝の 維持するように行った。7月中旬には,果そう葉数と最 長さで 1 0 c m 以内に着生する葉を採取し,葉面積計測用 大葉の大きさを調査した。収穫期に果台枝の長さ及び太 ndo ws 9 5 )を用 ソフト(山本一清製作所,LI A3 2f o rWi さ(枝の中位における直径)を測定した。果実は当農場 いて葉面積を算出した。 における各品種の収穫期にあわせて採取し,果重,硬度, 糖度,滴定酸含量,着色割合などの果実品質を調査した。 結 果 着色割合は観察により果実全体に対する赤色面積割合と 1.えき芽,1年生および 2年生以上のスパーの比較 した。翌年開花期に,前年ラベルを付けた果台から生長 スパー枝齢 1年生及び 2年生以上のスパーとえき芽に した枝の開花率,さらに収穫期に果実品質を測定した。 着生した果実の品質を第 3 -1表に示した。1年生と 2 なお,枝の年齢は満年齢で表し,春のせん定時における 年生以上のスパーで違いのみられた果実品質は,‘ふじ’ 昨年生長した枝を 1年枝とした。 の硬度,‘王林’の果実横径であり,1年生スパーが 2年 試験 2では,試験 1と同じ藤崎農場における 1 6年生 生以上のスパーより優れていた。えき芽果はスパー果に マルバカイドウ台開心形‘ふじ’ 6樹を供試した。スパー 比べ,‘ふじ’と‘ジョナゴールド’において果実横径が の分類は試験 1と同じであるが,えき芽果を含めず,1 劣り,ふじで硬度が高く,‘ジョナゴールド’で着色が劣 年生から 4年生以上の 4種類のスパーについて比較を ることが認められた。 行った。摘果等の管理は試験 1と同じである。果実は 果実品質の違いをもたらす要因として芽の素質が考え 1 1月初めに採取し,試験 1の果実品質の他に Ca含量を られるため,果そう葉の枚数と大きさに違いがあるかど 調査した。Ca含量の測定は,コルクボーラーで果心部 うか検討した(第 3 -2表) 。その結果,どの品種におい を除いて採取した果肉片を乾燥後乾式灰化し,日立偏光 ても 1年生スパーと 2年生以上のスパーの間に違いがみ ゼーマン原子吸光光度計を用いて行った。また,果実収 られなかった。ただ,えき芽の果そう葉はスパー葉に比 リンゴ樹における発生部位別スパー構成 43 第3 -5表 リンゴ‘ふじ’におけるスパー枝齢が果実品質に及ぼす影響 z スパー枝齢 果実重 (g) 硬度 (kg) 糖度 (°Br i x) 滴定酸含量 (g/ 1 0 0ml ) Ca含量 (mg/ 1 00g) 1年 2年 3年 4年以上 257. 4 265. 0 268. 9 236. 7 6. 7 6. 7 6. 5 6. 7 1 4 . 5 a bz 1 4 . 9 a 1 5 . 0 a 1 3 . 9 b 0 . 3 1 0 . 3 1 0 . 3 0 0 . 3 0 0 . 6 7 b 0 . 6 9 b 0 . 6 8 b 0 . 8 0 a 異なる文字の付いた処理間には Fi s her ’ sPLSDの多重検定により 5 %で有意差がある べ枚数が少なく,小さかった。 翌年あるいは翌々年の結果枝を確保するには,果台か 第3 -6表 ら新しょうが発生することが望ましい。スパー枝齢と果 台枝の発生,長さ,太さの関係について検討した(第 3 スパー枝齢 一果そう葉面積 (c m2) 1年 2年 3年 4年以上 4 6 . 8 5 3 . 3 4 3 . 3 4 3 . 8 -3表)。果台枝の発生割合はいずれのスパーにおいて も8 0 %以上であり,スパー枝齢によって発生率に違い はなかった。また,えき芽の果台からも同程度の果台枝 が発生した。果台枝の長さと太さは,長さの平均が‘ふ じ’9 . 8 c m, ‘王林’9 . 1 c m, ‘ジョナゴールド’1 0 . 7 c m, 太さの平均が‘ふじ’2 . 2 mm, ‘王林’3 . 0 mm, ‘ジョナ ゴールド’2 . 7 mm であった。果台枝の長さは全ての品種 Z Y リンゴ‘ふじ’におけるスパー枝齢と一果そう 葉及び果実周辺の葉面積の関係 果実周辺の葉面積 Z (c m2) Y 3 0 7 . 1 a 2 5 0 . 0 a 2 5 0 . 0 a 1 3 0 . 0 b 果台から 1 0 c m 以内の距離にある枝の総葉面積 異なる文字の付いた処理間には Fi s he r ’ sPLSDの多重検 定により 5 %で有意差がある においてスパー枝齢により違いは認められなかった。果 台枝の太さは, ‘ふじ’ では年齢の進んだ果台の方が細く なったが, ‘王林’と‘ジョナゴールド’では違いが認め みられる。そこで,一果そう葉と周辺の葉の量について られなかった。 比較した。なお,果実周辺の葉は果台から枝の長さで 1 0 スパーに着果させ,その果台から発生した果台枝の頂 c m までの距離に着生している当年枝に着生している葉 芽とえき芽における花芽形成にスパーの継続年数の違い とした。結果を第 3 -6表に示した。果そう葉面積はス が影響するかどうかを検討した。花芽形成の割合を開花 パー年齢により違いが認められなかった。果実周辺の葉 率で表した(第 3 -4表) 。果台枝頂芽の開花率は, ‘王 面積は,4年以上のスパーが 1 3 0 c m2と少なく,他の年 林’と‘ジョナゴールド’は 6 0 %以上で,平年並みの値 齢のスパーの約半分の葉面積しか存在していなかった。 が得られたが, ‘ふじ’は 3 5 %程度で低かった。しかし, 継続年数が 1年と 2年以上のスパーの間には,いずれの 考 察 品種とも違いは認められなかった。果台枝にえき生する 結果枝となるスパーは,太枝の中でさまざまな形で存 芽の開花率をみると,枝齢 1年のスパーが高く,2年以 在する。強く伸長した若枝にえき生する場合があり,何 上のスパーが低い傾向がみられた。 年も短い生長を繰り返した後に形成される場合もある。 そのため,スパーに着生する果実の生長条件が,スパー 2.1,2,3および 4年生以上のスパーの比較 に養分を供給する通導組織とのつながり,あるいは同化 1では,スパー枝齢が 2年以上のスパーを平均して 1 養分を供給する葉の存在などに違いがあると考えられ 年のスパー及びえき芽と比較したが,樹内にはさらに多 る。それらの違いが果実の生長と品質に影響するとすれ くの年数を経て形成されたスパーが存在している。そこ ば,良品果の生産にとって好ましいスパーを増加させる で,2年以上のスパーを 3 ,4年以上に分けて比較した。 樹体管理をする必要がある。本研究では,えき芽果の果 果実品質における違いを示したのが第 3 -5表である。 実品質がスパー果より果実肥大が劣ることが認められた。 果実重,硬度,滴定酸含量はスパー枝齢によって違いは この結果は,これまでの報告と一致する(J a c ks o n, みられなかった。ただ,果実重では,4年以上のスパー 1 9 6 7 ;J a c ks o n,1 9 7 0 )。果実肥大が劣る理由としては, に小さい果実が多い傾向がみられた。糖度は,2年と 3 果そう葉が少なく,小さいことが関係していると考えら 年のスパーが 4年以上のスパーより高かった。しかし, れた。‘王林’のスパー着生果のなかでは,1年生スパー Ca含量は 4年以上のスパーがそれより年数の少ないど 果が 2年生以上のスパー果より大きかった。しかし, のスパーより高かった。 ‘ふじ’と‘ジョナゴールド’では,スパー枝齢により スパー枝齢の多いスパーは,太い枝に近い位置で他の 果実肥大が影響されなかったが,4年生以上の果実が小 若い枝と離れて存在しており,果実周辺の葉が少ないと さくなる傾向がみられた。4年生以上の果実の周辺に存 浅 田 4 4 在する葉面積が小さいことが関係していると推察され 果台枝の発生率と果台枝頂芽の開花率は,3品種ともス る。しかし, 「スパー枝齢」 の古い果実は光条件が劣ると パー枝齢による違いがなかった。果台枝のえき芽の開花 されるため(Ro bi ns o nら,1 9 8 3 ) ,違いをもたらす要因 率は,スパー枝齢 1年が同 2年以上より高かった。‘ふ についてはさらに検討する必要がある。‘ふじ’では,4 じ’のスパー枝齢 4年以上の果実は,同 3年以下のス 年生以上の古いスパーの果実の糖度が低くなったが,平 パーより糖度が低く,Ca含量が多かった。また,果そ 均1 3 . 9 %を含んでおり,販売上問題はないと考えられ う周辺(1 0 c m 以内)の葉面積も,同 4年以上のスパー る。 のみ小さかった。 Ca含量は 4年以上のスパーで多くなった。Vo l zら (1 9 9 4 )は,果実周辺の葉面積が増大するほど,果実の Ca含量が増加することを示し,本試験とは逆の結果を 得ている。果実への Caの移行は蒸散流の強さだけでな く,貯蔵形態の Caの動きについても考慮する必要があ 第 4章 無せん定樹におけるスパーの発生部位 別構成割合と栄養・生殖生長および果 実生産性の関係 り,今後さらに検討を要する。 緒 言 本研究の結果は, 「スパー枝齢」 が増すにつれて果実肥 大と糖度が減少するとした Ro bi s o nら(1 9 8 3 )の報告と 第 2章では,せん定樹における種類別スパーの構成割 は類似した結果を得た。また,ニホンナシでは,ある程 合が栄養生長と生殖生長に密接に関係していることを明 度年数を経た側枝上の短果枝の方が良品果を生産すると らかにした。しかし,種類別スパーの構成割合と両生長 される(田村ら,1 9 9 0 ) 。リンゴでは,枝齢の多いスパー との関係は,せん定などの人為的操作が加えられていな でもいわゆるショウガ芽ほどのスパーの集塊を形成しな いリンゴ樹において確認しておくことと,どのような基 い。同じ結果枝齢でも,リンゴとナシでは果実周辺にお 本的関係が存在するかを明らかにしておく必要がある。 ける枝葉の着生や養水分の供給体制が異なると考えられ 最初に,発生部位別スパー率の枝齢による変化を明ら る。 かにしようとした。一つは若木における栽植時からのス 連年安定生産のためには,毎年枝を発生させ,それに パー率の変化であり,もう一つは無せん定主幹形樹の側 花芽を形成させることが必要である。スパー枝齢の多い 枝の位置と枝齢に伴う変化を調査した。スパー率が増加 スパーは太枝の先端部に位置する若いスパーに比べ,生 する要因は,樹齢が進むこと,受光量の減少,着果過多, 長力が劣ると考えられる。従って,果実を着生すること 無せん定にすること,土壌の過剰な乾燥など多数存在す によって果台枝や周辺の芽の発生率が抑制されること, る。これらの要因は,樹勢を低下させる要因として共通 また発芽しても花芽形成が抑制される可能性がある。し している。Tho mps o n(1 9 6 6 )は,リンゴの樹が年を経 かし,本試験の結果,果台枝の発生率とその頂芽の花芽 て樹勢が低下するとき,ほとんど全ての新しょうがス 分化率はスパー枝齢によって違いが認められなかった。 パーになると説明している。しかし,スパー率の増加 2年以上のスパーは周辺の葉面積が小さいにも関わら が,頂生スパーとえき生スパーのどのような変化を伴っ ず,若いスパーと同様の果台枝発生と花芽形成を示した て生じるかは明らかでない。そこで,枝齢による全ス のは,同化養分が比較的広い範囲から供給されているた パー率,頂生スパー率,えき生スパー率の相互関係につ めと考えられた。以上の結果は,スパー枝齢が 3年以内 いて検討した。 のスパーであれば,果実品質にほとんど差がなく,連年 発生部位別スパー率と栄養生長および花芽形成の関係 生産が可能であることを示すものである。 については,せん定樹と同様に検討した。無せん定樹で は,樹冠内部に極めて弱い新しょう生長がみられる。原 摘 要 スパーを形成する年数(スパー枝齢)の違いが,果実 因は加齢と日照不足が考えられる。加齢に伴う生長の減 少は植物に共通する性質である(Ga t s ukら,1 9 8 0 ) 。果 樹においても,分枝能力が年齢を経るとともに衰退して 品質と果台枝の発生及び花芽形成に及ぼす影響を明らか いくことが知られている(Oue l l e t t e ・Yo ung,1 9 9 5 ) 。頂 にするため, ‘ふじ’ , ‘王林’ , ‘ジョナゴールド’を供試 生とえき生の各スパー率が,生長の衰退とどのように関 し,検討した。 係にあるかを知るために,3年間継続した頂端生長の総 スパー枝齢が 1年と 2年以上のスパーで違いのみられ 長が 1 . 5 c m の芽を弱小枝とし,両者の関係を検討した。 た果実品質は, ‘ふじ’の硬度, ‘王林’の果実肥大であ り,1年スパーが 2年以上のスパーより優れていた。え き芽果はスパー果に比べ, ‘ふじ’ と ‘ジョナゴールド’に 材料および方法 おいて果実肥大が劣り,ふじで硬度が高く,‘ジョナ 弘前大学農学生命科学部生物共生教育研究センター付 ゴールド’で着色が劣った。果そう葉数と大きさは,3 属藤崎農場植栽のリンゴ‘ふじ’, ‘ジョナゴールド’, 品種ともえき芽果そうがスパー果そうより小さかった。 ‘スターキング・デリシャス’(以下,‘SD’と称す)の リンゴ樹における発生部位別スパー構成 100 100 ふじ 全スパ−率 割合(%) 80 ふじ 80 えき生スパ− 率 頂生スパー率 60 60 40 40 20 20 0 若若下上下 上上下上上 中中下中 中上下下下 下下中中中 1 2 3 4 5 100 全スパ−率 ジョナゴールド 80 えき生スパ− 率 頂生スパー率 60 40 全1年枝に占める割合 (%) 0 割合(%) 45 100 ジョナゴールド 80 60 40 20 0 下 下 中 下 中 若 下 下 下 中 中 下 20 100 0 1 2 3 80 4 60 樹齢(年) 第4 -1 -1図 40 リンゴ無せん定樹における全スパー率と種類 別スパー率の経年変化 20 0 3品種を供試した。供試樹数は, ‘ふじ’と‘ジョナゴー スターキング・デリシャス 下下下下下中下下下上中 中上上下中中上中中下下 なり枝の着生部位 ルド’が 9年生無せん定樹 1樹,5年生無せん定樹 2樹, ‘SD’は 9年生無せん定樹 1樹である。台木はいずれも 第4 -1 -2図 マルバカイドウで,土壌は沖積土である。無せん定樹は 植栽以来せん定を行わず無摘果としたが,その他の管理 リンゴ 3品種の無せん定樹における全スパー 率と発生部位別スパ-率の相互関係。若は 5 年生樹の平均値,上中下は主幹上の着生位 置,シンボルは第 4 -1 -1図を参照。 はせん定樹と同様に青森県の標準的な管理を行った。調 査用の枝については,主幹に着生する全ての側枝,調査 は新しょう生長の停止した 8月以降に行った。 結果および考察 側枝の発出角度については,垂直を 0度とし,0~ 6 0 度を上向き,60~ 1 2 0度を水平,12 0~ 1 8 0度を下向き とした。枝齢と長さの区分けについては,1年枝を前年 生長した枝とし,1 0 . 1 c m 以上,5 . 1~ 1 0 c m,5 c m以 1.発生部位別スパー率の樹齢による変異および樹内変 異 5年生樹における栽植後の 4年間の全スパー率,頂生 下に分け(小原,1 9 8 4 ) ,それぞれ長枝(長果枝より長い スパー率,えき生スパー率の経年変化を第 4 -1 -1図に 発育枝を含む) ,中果枝,スパーと呼称することにした。 示した。2年目から全スパー率が形成され始め,5年目 また,調査枝の各長さ別と発生部位別の 1年枝数の割合 には約 8 0 %に達した。2年目までは,えき生スパーだけ は 1年枝総数で除して算出した。2~当年生の枝長は, が存在し,頂生スパーは存在しなかった。頂生スパーは 3年継続して頂端生長した枝の各年枝の平均値で表し えき生スパーの形成開始から 1年遅れて,すなわち 3年 た。頂生とえき生については,それぞれ 2年枝の頂芽と 目から形成され始めた。両スパーとも数は次第に増加し ,1 9 9 9 ) 。なお, えき芽から発生した枝とした(As a da たが,全スパーの中では,えき生スパー率の増加率が低 ‘ふじ’と‘ジョナゴールド’の 9年生無せん定樹では, 下して頂生スパー率が増加した。 隔年結果性の現象がみられ,2 0 0 0年と 2 0 0 2年に多くの 9~ 1 0年生無せん定樹の主幹から発生している太枝 果実が着果した。 ‘SD’については隔年結果性がみられ を単位として,全スパー率の低い枝から高い枝までの頂 なかった。 生スパー率とえき生スパー率の変化について検討した。 結果を第 4 -1 -2図に示した。 全スパー率は, ‘ふじ’と‘ジョナゴールド’では約 5 0 %から,‘SD’では約 8 5 %から約 1 0 0 %までの幅で 浅 田 30 20 ふじ 平均1年枝長(cm) 平均新しょう長(cm) 4 6 0.7844 y = 42.012x R2 = 0.643 15 10 5 平均1年枝長(cm) 平均新しょう長(cm) y = 86.718x 0.8444 R2 = 0.8691 20 15 10 5 0 0 20 ジョナゴールド 15 y = 6.5807e 0.0126x R2 = 0.5438 10 5 0 40 ジョナゴールド 30 y = 41.384e 0.0453x R2 = 0.6317 20 10 0 30 20 18 16 14 12 10 8 6 4 2 0 スターキング・デリシャス 平均1年枝長 (cm) 平均新しょう長 (cm) ふじ 25 y = 1.799e 0.0022x R2 = 0.0075 スターキング・デリシャス 25 y = 4.181e 0.0075x R2 = 0.053 20 15 10 5 0 0 20 40 60 80 100 0 頂生スパ−率(%) 第4 -2 -1図 リンゴ 3品種の無せん定樹における頂生ス パー率と平均新しょう長の関係。 変異があった。ただ, ‘ジョナゴールド’ は最高全スパー 20 40 60 80 100 頂生スパ−率 (%) 第4 -2 -2図 リンゴ 3品種の無せん定樹における頂生ス パー率と平均 1年枝長の関係 は,樹齢が進むにつれ,栄養生長力の低下とともに樹冠 率が 9 0 %と低かった。各スパー率の変化をみると,‘ふ 内の光環境が悪化する。9~ 1 0年生樹になると,樹冠内 じ’と‘ジョナゴールド’では,全スパー率が低い枝で の太枝は,さまざまな全スパー率をもつ枝で構成され はえき生スパー率が頂生スパー率より高く,全スパー率 る。本調査の結果では,すでに‘ふじ’と‘SD’で全ス が高い枝では逆になった。全スパー率の増加に伴ってえ パー率 1 0 0 %近い太枝が存在した。それらの枝に共通す き生スパー率より頂生スパー率が高くなった。両者が逆 る性質は,全スパーのうち頂生スパーが大半を占めるこ 転する全スパー率は, ‘ふじ’で約 9 0 %, ‘ジョナゴール とであった。この結果は,樹齢の進行に伴う衰弱が頂生 ド’で約 8 0 %であった。 ‘SD’では,他の 2品種と異な スパーの形成とともに生じることを示している。 り,ほとんどの太枝においてえき生スパー率より頂生ス また,本調査は,無せん定樹の下枝では,栽植後の若 パー率の方が高かった。しかし,全スパー率約 9 5 %以 木時代に頂生スパー率がえき生スパー率を越える転換期 上になると,両スパー率の違いがより大きくなった。全 が存在することを示している。この転換期は,全スパー 品種に共通する変化は,全スパー率が高くなると,えき 率が 8 0~ 9 0 %に達した時期に生じると考えられるが, 生スパー率が低くなり,頂生スパー率が高くなることで ‘ふじ’と‘ジョナゴールド’で違いがみられたので, あった。 さらに検討する必要がある。 栽植後の若木は栄養生長が強いために,全スパー率が ‘SD’については,栽植直後の各スパー率の変化を調 ゼロから徐々に増加する。種類別スパー率をみると,最 査していないため,えき生スパー率の高い期間について 初に形成されるのがえき生スパーで,そのえき生スパー は不明であるが,他の 2品種に比べ,全スパー率と頂生 から 1年遅れで頂生スパーが形成される。頂生スパーの スパー率が極めて高い品種特性を有するとみられた。こ 増加率がえき生スパーのそれより大きいため,全スパー の違いは,スパー形成能力(Wa r r i ngt o nら,1 9 9 0 )に基 に占める頂生スパー率が次第に増加する。無せん定樹で づくと考えられたが,さらに詳細な検討が必要である。 リンゴ樹における発生部位別スパー構成 6 ふじ 20 頂芽増加率(%) 平均 2 年枝長(cm) 25 0.0314x y = 20.569e R2 = 0.8942 15 10 5 y = 2.8063e 0.0138x R2 = 0.7235 4 3 2 1 6 30 25 頂芽増加率(%) ジョナゴールド y = 19.464e 0.0287x R2 = 0.5422 20 15 10 5 ジョナゴールド 4 y = 5.9778e 0.0234x R2 = 0.6782 3 2 5 1 0 0 25 6 スターキング・デリシャス 20 0.038x 頂芽増加率 (%) 平均 2 年枝長(cm) ふじ 5 0 0 平均 2 年枝長 (cm) 47 y = 35.506e R2 = 0.7387 15 10 5 0 スターキング・デリシャス 5 y = 1.8068e 0.005x R2 = 0.198 4 3 2 1 0 0 20 40 60 80 0 100 20 頂生スパ−率(%) 第4 -2 -3図 リンゴ 3品種の無せん定樹における頂生ス パー率と平均 2年枝長の関係 2.スパーの発生部位別構成割合と栄養生長の関係 (1)平均新しょう長,1・2年枝長および頂芽増加率 第4 -2 -4図 第4 -2 -1表 年枝長および頂芽増加率との関係について検討した。 80 100 均新しょう長の関係をそれぞれ,第 4 -2 -1~ 3図に示 した。これらの図にみられるように,頂生スパー率と栄 養生長の間には負の相関関係が確認された。ただ,品種 弱いか,相関のみられない関係もあった。その理由とし ては, ‘SD’は品種の特性として高い頂生スパー率を有 する(第 2章第 2項参照)ために,平均 1年枝長と 2年 枝長の変異の幅が極めて小さいことによると考えられ た。 第 4 -2 -4図に頂生スパー率と頂芽増加率(BPR)の 関係を示した。BPRは栄養生長の強さの指標でもある Z が,第 2章第 2項で検討したように葉面積当たり果数, * リンゴ無せん定樹とせん定樹における弱小 枝の発生 榎榎榎榎榎榎榎榎榎榎榎榎榎(%) 無せん定 せん定* 若木 上向き 水平 下向き ふじ 0 . 0 Z 9 . 6±13 . 6 2 0 . 0±10 . 6 3 0 . 8±17 . 3 - 0 . 0 1 3 . 1±7. 4 1 2 . 3±1 0. 2 若木 上向き 水平 下向き ジョナゴールド 1 . 9±2. 3 4 . 2±4 . 2 1 . 3±3 . 3 4 . 3±2 . 8 - 0 . 0 1 . 4±1. 7 0 . 0 上向き 水平 下向き スターキング・デリシャス 1 1 . 7±15 . 8 1 4 . 4±5. 5 1 0 . 4±8. 0 0 . 0 1 . 1±1 . 7 0 . 0 によって両者の関係の強さは異なった。 ‘ふじ’ が最も強 い関係を示し,次いで‘ジョナゴールド’ , ‘SD’は最も リンゴ 3品種の無せん定樹における頂生ス パー率と頂芽増加率の関係 太枝の発出角度 頂生スパー率と平均 2年枝長,平均 1年枝長および平 頂生スパー率の増加とともに低下した。ただ, ‘SD’は 60 頂生スパ−率 (%) 1年枝の頂生スパー率と平均新しょう長,1年枝長,2 すなわち果実の生産性と関係する要素である。BPRは 40 平均 ± 標準偏差 せん定の供試樹については,第 5章の材料および方法を参 照 浅 田 4 8 70 頂生スパー率 3年継続頂端生長した 枝に占める割合(%) 60 えき生スパ−率 50 40 30 20 10 0 0 10 20 30 40 50 60 70 80 0 10 20 頂生スパー率(%) 第4 -2 -5図 30 40 50 60 70 80 えき生スパ−率 (%) ●:ふじ,○:ジョナゴールド,△:SD リンゴ無せん定樹における種類別スパ-率と 3年継続頂端生長した枝の総長が 1 . 5 c m の枝の割 合との関係 60 他の 2品種と異なり,頂生スパー率が増加しても BPR ふじ がほとんど変化しなかった。 3年 間 伸 長 し た 枝 の 総 長 は 1 . 5 c m(弱 小 枝)か ら 2 4 5 c m の長い枝まで広い範囲で存在した。そのなかの 弱小枝の割合を第 4 -2 -1表に示した。 ‘ふじ’と‘ジョ ナゴールド’の 5年生樹では,無せん定でも弱小枝は存 在しなかった。無せん定 9年生樹の弱小枝率をみると, 花芽形成率 (%) (2)発生部位別スパー率と弱小枝の発生との関係 JG SD 50 40 30 20 10 ‘ふじ’は,上向き枝から下向き枝へと次第に増加し, 下向き枝で約 3 0 %に達した。弱小枝率の増加は全体の スパー率に比例した。 ‘ジョナゴールド’では,弱小枝率 0 0 は5 %以下と低く,太枝の角度による違いはなかった。 全体のスパー率が 8 5 %と高かった下向き枝も弱小枝率 が4 . 3 %と低かった。 ‘SD’では,太枝の角度にかかわら ず1 0~ 1 5 %と前 2品種の中間の弱小枝率を示した。弱 小枝の発生の仕方に品種間差異があるとみられた。 ‘ふ じ’の無せん定樹では,独立樹であるにもかかわらず樹 20 40 60 頂生スパ−率 (%) 80 100 第4 -3 -1図 リンゴ 3品種の無せん定樹における頂生ス パ ー 率 と 花 芽 分 化 率 の 関 係。J Gは ジ ョ ナ ゴールド,SDはスターキング・デリシャス ふじ:= 1 1 . 9- 0 . 2 2 X(r=- 0 . 5 7**) , J G:Y= 3 9 . 9- 0 . 5 9 X(r=- 0 . 6 9 4**) , SD:Y= 2 6 . 9- 0 . 2 3 X(r=- 0 . 4 4*) 冠内が暗く,頂生スパーが何年も継続して形成された後 に,小さな葉だけを着生する貧弱な頂生スパーが多くみ ルド’と‘SD’では,弱小枝率が低く,両者の間に明確 られた。菊池(1 9 7 5 )は, ‘国光’の無せん定樹が‘SD’ な関係がみられなかったが,‘ふじ’では,弱小枝率が頂 より過繁茂による光不足になりやすいと報告している。 生スパー率の増加とともに増え,逆にえき生スパー率と ‘国光’は‘ふじ’の片親であるため,‘ふじ’も‘国 ともに低下する関係が認められた。 光’と類似した特性を有すると考えられた。一方,せん 頂生スパーとえき生スパーの相反する弱小枝率との関 定樹では, ‘ふじ’の場合,弱小枝率が水平枝と下向き枝 係は,前述の栄養生長との関係と同じ結果になった。す 3 %存在したが,無せん定樹より少なかった。 で1 2~ 1 なわち,栄養生長が強いえき生スパー率の高い枝では, ‘ジョナゴールド’と‘SD’の弱小枝率は,せん定樹で 弱小枝が少なかった。 ほとんど認められなかった。以上の結果は,いずれの品 以上の結果は, リンゴ樹では, 頂生スパーが連続し 種においても,せん定が弱小枝率の発生を抑制している て形成されることが,栄養生長の減衰の結果であるこ ことを示している。 と, 同じスパーでもえき生スパー率が高いと強い栄養 無せん定樹における弱小枝の割合と頂生・えき生のス 生長の状態にあることを示している。 パー率との関係を示した(第 4 -2 -5図) 。 ‘ジョナゴー リンゴ樹における発生部位別スパー構成 3.頂生スパー率と花芽形成の関係 1年枝の頂生スパー率と 1年枝頂芽の花芽形成率との 関係をみたのが,第 4 -3 -1図である。 49 第 5章 スパー形成要因およびせん定によるス パー率の調整 3品種とも隔年結果の状態にあり,花芽形成率が低 緒 言 かった。従って,頂生スパー率と花芽形成の関係を検討 するのに適切な状態とは考えられなかったが,頂生ス えき生スパーと頂生スパーの形成に必要な条件を明ら パー率の増加に伴って最大花芽形成率が低下するという かにし,両スパーの構成割合を調整するせん定方法を確 両者の関係は,せん定樹と同様に確認された。 立することが大切である。そのため,無せん定樹とせん 定樹を供試し,各スパーを形成する母枝の形質につい 摘 要 て,また,せん定樹の枝構成を無せん定樹のそれと比較 することにより,せん定が発生部位別スパー率にどのよ リンゴ 3品種の無せん定樹を供試し,栽植から 5年ま うに影響したか,そしてどのような枝をせん除すれば各 でと,9~ 1 0年生樹の主幹に着生するさまざまな年齢 スパー率が変化するか等についてについて検討した。全 の太枝における全スパー率,えき生スパー率,頂生ス スパー率は,毎年 5 c m より長い枝のみが形成されると パー率の相互関係について検討した。 0 %になるため,長枝の発生により左右される。えき生 ‘ふじ’と‘ジョナゴールド’では,2年生樹から全ス スパーも,長枝がなければ形成されないので,長枝の発 パー率が形成され始め,5年生樹では約 8 0 %に達した。 生により左右される。頂生スパーは,えき生スパーの頂 えき生スパーは 2年生樹から,頂生スパーは 3年生樹か 芽から形成されるのが最初で,その後の主な形成位置 ら形成された。全スパー率の増加につれてえき生スパー は,頂生スパーの頂芽になる。従って,頂生スパーの形 率が減少し,頂生スパー率が増加した。えき生スパー率 成を左右する要因は,えき生スパー形成数と頂芽からの と頂生スパー率の逆転する全スパー率は,‘ふじ’で約 スパー形成率である。頂芽からのスパー形成率はスパー 9 0 %, ‘ジョナゴールド’で約 8 0 %であった。9年生 になるか長枝になるかの割合と頂芽から新しょうが発生 ‘SD’では,どの全スパー率の太枝でもえき生スパー率 しない割合によって左右されるので,(1 0 0-(長枝率+ より頂生スパー率の方が高かった。全品種に共通する変 生長停止率))の式により求められる。ただ,各種スパー 化は,全スパー率が高くなると,えき生スパー率が低く 率は相互に関連して変化するため,1種類のスパー率の なり,頂生スパー率が高くなることであった。 み変化させることはできない。従って,どのような操作 無せん定樹における頂生スパー率と栄養生長との関係 により全体として各種スパー率がどう変化するかを把握 については, ‘ふじ’と‘ジョナゴールド’において頂生 する必要がある。 スパー率と平均新しょう長の間に負の強い指数関係が認 これらの考えに基づき,枝モデルを使用し,全,頂生 められ,頂生スパー率が増加するにつれて平均新しょう およびえき生スパー率の経年変化を推定し,各スパー率 長が減少した。同 2品種では,頂生スパー率は平均 2年 を左右する要因を確認するとともに,各種スパー率を調 枝長,1年枝長および頂芽増加率とも同様の指数関係を 整するせん定方法を確立するための知見を得ようとし 示し,同スパー率の増加に伴って各枝長および頂芽増加 た。 率が減少した。 ‘SD’では,頂生スパー率の増加ととも に平均 2年枝長が減少する関係が認められた。3年継続 頂端生長の総長が 1 . 5 c m の枝(弱小枝)は, ‘ふじ’と 材料および方法 ‘ジョナゴールド’の 5年生樹では無せん定でも弱小枝 供試した無せん定樹は第 4章と同じである。無せん定 は存在しなかった。9年生樹では, ‘ふじ’は,上向き枝 樹との比較を行うため,無せん定樹と同じ弘前大学農学 から下向き枝へと次第に増加し,下向き枝で約 3 0 %に達 生命科学部生物共生教育研究センター付属藤崎農場植栽 した。弱小枝の増加は全体のスパー率と正の相関が認め のリンゴ‘ふじ’と‘ジョナゴールド’および同農場に られた。 ‘ジョナゴールド’では,弱小枝は 5 %以下と低 隣接する S氏園植栽の‘スターキング・デリシャス’ (以 く,太枝の角度による違いはなかった。‘SD’では,太 下,‘SD’と称す)の 3品種を供試した。‘ふじ’と‘ジョ %と前 2品種の中間の弱 枝の角度にかかわらず 1 0~ 1 5 ナゴールド’は 2 2年生開心形各 4樹で,‘SD’は約 3 0 小枝を示した。ふじ’の無せん定樹では,弱小枝率が頂 年生開心形 3樹である。発出角度の異なる太枝を 1樹当 生スパー率の増加とともに増え,逆にえき生スパー率と たり 3本選出した。調査方法は第 4章と同じである。な ともに低下した。 ‘ジョナゴールド’と‘SD’では,弱 お,当年枝および 1年枝の形成における母枝というの 小枝率が低く,両者の間に明確な関係がみられなかっ は,それぞれ 1年枝および 2年枝を指している。 た。 花芽形成率は頂生スパー率の増加とともに最大形成率 が減少する傾向が 3品種に共通して認められた。 浅 田 5 0 第5 -1 -1表 発生えき生枝数と母枝長,母枝直径,母枝角度の相関関係における決定係数 樹番号 母枝の 生長年 1 1 2 2 平均 母枝長 単相関 母枝直径 母枝角度 重相関 3変数 0. 8 6 1 0. 8 0 1 0. 8 1 0 0. 8 1 1 0. 8 2 1 0 . 6 7 9 0 . 3 5 5 0 . 6 7 7 0 . 3 4 6 0 . 5 1 4 0 . 0 3 5 0 . 0 5 3 0 . 0 1 0 0 . 0 3 1 0 . 0 3 2 0 . 8 6 9 0 . 8 0 5 0 . 8 5 6 0 . 8 8 3 0 . 8 5 3 2001 2002 2001 2002 第5 -1 -2表 発生スパー数と母枝長,母枝直径,母枝角度の相関関係における決定係数 樹番号 母枝の 生長年 1 1 2 2 平均 母枝長 単相関 母枝直径 母枝角度 重相関 3変数 0. 7 0 9 0. 6 9 7 0. 7 9 6 0. 8 7 8 0. 7 7 0 0 . 5 2 1 0 . 3 4 1 0 . 6 1 8 0 . 3 3 4 0 . 4 5 4 0 . 0 2 1 0 . 0 5 1 0 . 0 0 3 0 . 0 3 0 0 . 0 2 6 0 . 7 1 1 0 . 7 7 8 0 . 8 2 5 0 . 8 7 9 0 . 7 9 8 2001 2002 2001 2002 25 結果および考察 Y=−0.21+0.19X(r=0.937**) 1.スパー形成要因 20 えき生スパーは主に 2年枝のえき芽に形成される。 従って,2年枝のスパーを形成する形質について検討し た。せん定の有無がスパー形成に影響すると考えられる ため,無せん定樹とせん定樹の両樹について比較検討し た。 ①無せん定‘ふじ’5年生樹における発生えき生枝数お よびスパー数を左右する母枝形質 母枝の形質として,長さ,母枝直径,発出角度をとり えき生スパ−数 (本) (1)えき生スパーの形成要因 15 10 5 0 0 あげ,全えき生枝およびスパー発生数との関係について 検討した。単相関および重相関分析の結果を第 5 -1 -1 50 100 150 −5 表に示した。 母枝長 (cm) 全えき生枝発生数との関係では,母枝の長さ,直径, 発出角度のなかで最も密接に関係する変数は母枝長で あった。3変数を用いた重回帰分析を行った結果,母枝 第5 -1 -1図 リンゴ‘ふじ’の無せん定若木における母枝 長とえき生スパ-発生数の関係(2 0 0 2) 長との単相関における決定係数 0 . 8 2を大きく上回るこ とはなかった。この結果は,全えき生枝発生数が主に母 第5 -1 -3表,第 5 -1 -4表のようであった。 枝の長さにより左右されることを示している。 全えき生枝 1本と 5本を形成するのに必要な母枝長 次に,発生スパー数と母枝の形質の関係を検討した。 は,それぞれ 8 c mと 3 4 c m であった。また,えき生ス 結果を第 5 -1 -2表に示した。最も寄与率の高い変数 パー 1本と 5本を形成するのに必要な母枝長は,それぞ は,全えき生枝発生数と同様母枝長であった。重回帰分 れ9 c m,4 0 c m であった。 析では,母枝長との単相関で得られた関係とほぼ同じ決 母枝の発出角度は,全えき生数およびえき生スパー数 定係数しか得られなかった。従って,発生スパー数は母 と単相関で密接な関係は得られなかったが,両者の散布 枝長で決まると考えられる。第 5 -1 -1図に母枝長とえ 図(第 5 -1 -2図)をみると,発出角度が大きくなり, き生スパー発生数の単相関関係を示した。 枝が下を向くほどえき生枝の最大発生数が減少した。母 得られた回帰式から,1本と 5本の全えき生枝および 枝の発出角度がえき生枝発生の上限を決定する要因であ えき生スパーを形成するのに必要な母枝長は,それぞれ る可能性が認められた。しかし,母枝の発出角度が大き リンゴ樹における発生部位別スパー構成 51 第5 -1 -3表 リンゴ‘ふじ’の無せん定 5年生樹における母枝長と発生えき生枝数の回帰係数 樹番号 母枝の 生長年 定数項 傾き 決定係数 n 1 1 2 2 平均 2001 2002 2001 2002 - 0. 023 - 0. 116 - 0. 460 - 0. 212 - 0. 203 0 . 1 3 5 0 . 1 5 3 0 . 1 3 3 0 . 1 9 4 0 . 1 5 4 0 . 8 6 0 0 . 8 0 1 0 . 8 5 4 0 . 8 7 8 0 . 8 4 8 1 9 0 6 5 0 1 7 2 6 8 9 母枝長(c m) スパー本数 1本 5本 7 . 6 7 . 3 1 1 . 0 6 . 2 8 . 0 3 7. 2 3 3. 4 4 1. 1 2 6. 9 3 3. 8 第5 -1 -4表 リンゴ‘ふじ’の無せん定 5年生樹における母枝長と発生えき生スパー数の回帰係数 樹番号 母枝の 生長年 定数項 傾き 決定係数 n 1 1 2 2 平均 2001 2002 2001 2002 0. 127 - 0. 084 - 0. 251 - 0. 189 - 0. 099 0 . 0 9 0 0 . 1 4 0 0 . 1 0 7 0 . 1 7 1 0 . 1 2 7 0 . 7 1 0 0 . 7 7 5 0 . 8 2 4 0 . 8 7 6 0 . 7 9 6 1 9 0 6 5 0 1 7 2 6 8 9 9 . 7 7 . 7 1 1 . 7 7 . 0 9 . 0 5 4. 1 3 6. 3 4 9. 1 3 0. 3 4 0. 2 140 120 20 100 母枝長 (cm) えき生スパー数 (本) 25 母枝長(c m) スパー本数 1本 5本 15 10 5 80 60 40 20 0 0 0 50 100 150 200 0 リンゴ‘ふじ’の無せん定若木における母枝 の発出角度とえき生スパ-発生数の関係 100 150 200 母枝角度 (°) 母枝の発出角度(°) 第5 -1 -2図 50 第5 -1 -3図 リンゴ‘ふじ’の無せん定若木における母枝 長と母枝の発出角度の関係 くなると,母枝長が短くなるためにえき生枝の発生が減 木樹においても母枝長とえき生枝発生数の関係が 5年生 少することも考えられるため,母枝の発出角度と母枝長 の若木と同様であるかどうかを検討した。母枝長と全え の関係を検討した結果,第 5 -1 -3図にみるように,母 き生枝発生数およびえき生スパー数との関係をそれぞれ 枝角度が大きくなるにつれて母枝長の最大値が減少し 第5 -1 -5表,第 5 -1 -6表に示した。母枝長と全えき た。 生枝発生数の関係は,いずれの品種においても,せん定 以上の結果から,母枝角度はえき生枝発生数の上限を の有無にかかわらず 2次曲線としてよく当てはまった 決める要因ではなく,母枝長の上限を決める要因である (第 5 -1 -5表) 。決定係数は 0 . 8 1から 0 . 9 6と高く,母 ことが認められた。 枝長によりえき生枝発生数が決定されることを示した。 母枝長とえき生スパー発生数との関係は,若木の場合 ②無せん定 9年生樹と成木せん定樹における全えき生 枝およびえき生スパーの発生数と母枝長の関係 と同様,直線関係としてよく当てはまった(第 5 -1 -6 ) 。 ‘SD’は全えき生枝発生数と同程度の決定係数を示した 前項において,えき生枝の発生数に最も密接な関係を が,‘ふじ’と‘ジョナゴールド’では決定係数が 0 . 7 0 有する要因が母枝長であることを 5年生無せん定樹の調 から 0 . 8 1と小さかった。回帰式の傾きをみると,‘SD’ 査により明らかにした。そこで,無せん定とせん定の成 の傾きが‘ふじ’と‘ジョナゴールド’の傾きより大き 浅 田 5 2 第5 -1 -5表 リンゴ 3品種の 9年生無せん定樹と成木せん 定樹における 1年枝長と全えき生枝発生数の 関係 第5 -1 -6表 リンゴ 3品種の 9年生無せん定樹と成木せん定 樹における 1年枝長とえき生スパー発生数の関 係 品種 せん定 の有無 回帰式 決定係数 品種 せん定 の有無 回帰式 決定係数 1本発生に要 する枝長(cm) ふじ 無 有 無 有 有 無 y= 0. 20+ 0. 20x- 0. 0003x2 y=- 0. 11+ 0. 16x- 0. 0005x2 y= 0. 35+ 0. 15x- 0. 0006x2 y= 0. 11+ 0. 17x+ 0. 0003x2 y=- 0. 28+ 0. 30x- 0. 0015x2 y=- 0. 32+ 0. 13x- 0. 0011x2 0 . 8 1 0 . 9 4 0 . 9 0 0 . 8 5 0 . 8 6 0 . 9 6 ふじ 無 有 無 有 無 有 y= 0 . 2 6+ 0 . 1 9 x y= 0 . 0 5+ 0 . 1 1 x y= 0 . 1 6+ 0 . 1 6 x y= 0 . 2 5+ 0 . 1 3 x y=- 0 . 0 7+ 0 . 2 4 x y=- 0 . 6 0+ 0 . 2 5 x 0 . 7 8 0 . 7 0 0 . 8 1 0 . 7 1 0 . 8 4 0 . 9 5 5. 0 8. 4 4. 8 5. 2 4. 5 0. 9 J Gz SD z J Gはジョナゴールド,SDはスターキング・デリシャス 第5 -1 -7表 リンゴ‘ふじ’と‘ジョナゴールド’の無せ ん定 9年生樹と 5年生若木および‘スターキ ング・デリシャス’の 9年生無せん定樹にお けるえき生スパ-頂芽からの 1年目の生長 榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎(%) ≧ 5 c m 不発芽 z 54. 2 29. 2 13. 3 14. 2 18. 9 0 . 0 0 . 0 0 . 0 0 . 0 0 . 0 ジョナゴールド 92. 9 7. 1 64. 2 35. 8 89. 3 9. 4 76. 8 22. 6 0 . 0 0 . 0 1 . 3 0 . 7 スターキング・デリシャス 97. 4 1. 0 99. 5 0. 0 97. 7 2. 3 98. 2 1. 1 1 . 6 0 . 5 0 . 0 0 . 7 木(枝)の種類 頂生スパー ふ じ 45. 8c 70. 8b 86. 7a 85. 8a 81. 1 若木 上向き枝 水平枝 下向き枝 平均 x 若木 水平枝 下向き枝 平均 x 上向き枝 水平枝 下向き枝 平均 z y x 芽の不発芽と枯死を含む 異なる文字の付いた木(枝)間には Fi s her ’ sPSLDにより 5 %水準で有意差がある 若木を除く平均値 J Gz SD z J Gはジョナゴールド,SDはスターキング・デリシャス 第5 -1 -8表 ≧5 c m 不発芽 z 2 8 . 0 2 9 . 9 2 5 . 7 2 7 . 9 0. 0 0. 0 0. 0 0. 0 上向き枝 水平枝 下向き枝 平均 ジョナゴールド 8 0 . 0 2 0 . 0 7 1 . 4 2 8 . 6 6 7 . 6 2 9 . 7 7 3 . 0 2 6 . 1 0. 0 0. 0 2. 7 0. 9 上向き枝 水平枝 下向き枝 平均 スターキング・デリシャス 9 1 . 6 8 . 4 9 5 . 6 4 . 4 9 5 . 0 5 . 0 9 4 . 1 5 . 9 0. 0 0. 0 0. 0 0. 0 太枝の種類 上向き枝 水平枝 下向き枝 平均 z リンゴ 3品種のせん定樹におけるえき生ス パー頂芽からの 1年目の生長榎榎榎榎榎(%) 頂生スパー ふ じ 7 2 . 0 7 0 . 1 7 4 . 3 7 2 . 1 芽の不発芽と枯死を含む しょうはすべてスパーになるとは限らず,その長さを決 める要因があると考えられる。そこで,無せん定樹とせ ん定樹におけるえき生スパーと頂生スパーの頂芽に形成 される長さ別新しょうの割合について検討した。 い傾向を示した。この違いは,‘SD’が他の 2品種より ①えき生スパーからの頂生スパーの形成 スパーを形成しやすい性質を有していることによると考 リンゴ‘ふじ’,‘ジョナゴールド’および‘SD’の木 えられた。えき生スパー 1本を発生させるに要する母枝 あるいは太枝におけるえき生スパー頂芽の 1年目の生長 長は, ‘ふじ’のせん定樹を除いて約 5 c m あるいはそれ を第 5 -1 -7表に示した。 以下( ‘SD’のせん定樹)と短かった。若木より成木の 頂芽が発芽しないえき生スパーはいずれの品種におい 方がスパーを形成しやすいかどうかについてさらに検討 てもほとんどみられなかった。頂生スパーを形成するえ する必要がある。 き生スパーの割合の平均値は, ‘ふじ’, ‘ジョナゴール ド’,‘SD’でそれぞれ,8 1 . 1 ,7 6 . 8 ,9 8 . 2 %であった。 (2)頂生スパー形成要因 ‘ふじ’では,5年生樹,9年生上向き枝,同水平枝と下 頂生スパーは,いろいろな長さを持つ母枝の頂芽から 向き枝の順で次第に高くなった。水平枝と下向き枝で 形成されるが,主としてえき生スパーの頂芽から形成さ は,約 8 5 %が頂生スパーになった。すなわち,頂生ス れるものと頂生スパーの頂芽から形成されるものが大部 パーの形成率は樹勢の弱い太枝ほど高くなった。 ‘ジョ 分を占める(第 2章) 。しかし,両頂芽から形成される新 ナゴールド’では,水平枝を除いて,8 9 %以上と高かっ リンゴ樹における発生部位別スパー構成 53 第5 -1 -9表 リンゴ 3品種におけるせん定の有無とえき生スパー頂芽からの 1 ,2年後における生長のタイプ(%) 2年目 の生長 の生長 NPy P NP P NP P TSx 〃 〃 ≧5 c m 〃 〃 せん定の有無 u TS ≧ 5 c m なし v TS ≧ 5 c m なし w 76. 4a 4. 4bc 0. 3c 13. 4b 5. 4bc 0. 1c ** 3 1 . 5 a 1 1 . 2 bc 2 9 . 5 a 7 . 8 c 1 7 . 2 b 2 . 8 d 6 7 . 9 a 7 . 4 b 7 . 3 b 2 . 9 b 1 3 . 2 b 1 . 4 b 3 1 . 4 a 2 2 . 0 a b 1 2 . 5 b 6 . 2 b 1 5 . 7 b 4 . 2 b 8 5 . 1 a 5 . 5 b 8 . 2 b 0 . 1 c 0 . 3 c 0 . 8 c * 6 1 . 0a 2 7 . 5b 5 . 5c 4 . 1c 1 . 9c 0 . 0c z y x w v u ふじ SDz 1年目 ジョナゴールド SDはスターキング・デリシャス NPは無せん定,Pはせん定あり TSは頂生スパー 異なる英文字はタイプ間に Fi s her ’ s PLSDにより 5 %水準で有意差がある 芽の不発芽+枯死 **,*はそれぞれ 1 %,5 %水準で有意差がある た。水平枝は,本章次項の第 5 -2 -1表で示すように, 成する場合とがあり,前者が頂生スパーの主要な形成過 平均枝長が特に長い枝であったことを考慮すると,同じ 程であった。3品種のうち‘ふじ’と‘SD’においてせ 5年生樹でも‘ふじ’より頂生スパー率が高いことから, ん定樹の方が無せん定樹より頂生スパー形成率が明らか ‘ジョナゴールド’は‘ふじ’より頂生スパーを形成し に低かった。また,せん定樹では,えき生スパーから 1 やすい品種特性を有していると推察される。 ‘SD’では, 年目に頂生スパーが形成された後,次の年に長い新しょ 他の 2品種と異なり,どの発出角度の太枝でも 9 7 %以 うを発生させる割合が無せん定樹より高かった。この結 上の頂生スパー形成率を示し,特に頂生スパーを形成し 果は,せん定が頂生スパー形成率を左右する要因の 1つ やすい品種の性質を有していると考えられた。 であることを示している。 第 5 -1 -8表にせん定樹におけるえき生スパー頂芽か 2年目に新しょうが発生しない枝の割合(TS+≧ 5 ら形成される頂生スパーの割合を示した。頂芽から新 c m)は,‘ふじ’,‘ジョナゴールド’,‘SD’それぞれ,無 しょうが形成されない不発芽率はどの品種においてもほ . 6 せん定樹で 0 . 4 ,8 . 7 ,9 . 0 %,せん定樹で 3 2 . 3 ,1 6 . 7 ,5 とんどみられなかった。全枝の平均頂生スパー形成率 %であった。‘ふじ’と‘ジョナゴールド’では,無せん は, ‘ふ じ’ ,‘ジ ョ ナ ゴ ー ル ド’, ‘SD’で そ れ ぞ れ, 定樹よりせん定樹の方が生長を停止する頂芽が高い傾向 7 2 . 1 ,7 3 . 0 ,9 4 . 1 %であった。頂生スパーの形成率は, を示した。なお,タイプ別分布割合は品種間に有意な違 ‘ふじ’と‘ジョナゴールド’ではほぼ等しく,両品種 いが認められなかった。 とも太枝の発出角度による差異は認められなかった。せ 頂生スパーは,主にえき生スパーあるいは頂生スパー ん定樹の頂生スパー形成率は,無せん定樹の値と比較し の頂芽から形成される。頂芽からはスパーの他に 5 c m て,低い傾向がみられた。‘SD’では,せん定樹におい 以上の新しょうが伸びる場合と新しょうが伸びず生長を ても太枝の発出角度にかかわらず 9 0 %以上頂生スパー 停止する場合がある。従って,5 c m 以上伸びる新しょ を形成し,他の 2品種より頂生スパーを形成しやすい品 うの割合と生長を停止する割合により頂生スパー形成率 種特性を示した。 が左右される。 ②えき生スパーの頂芽から 2年後に頂生スパーを形成 る生長は,新しょうを発出させない芽が‘ふじ’, ‘ジョ 本研究の結果では,えき生スパー頂芽の 1年目におけ する割合 ナゴールド’,‘SD’の 3品種ともせん定の有無にかかわ 結果を第 5 -1 -9表に示した。2年後に頂生スパーを らずほとんど無く,5 c m 以上の新しょうになるかどう 形成する割合は,‘ふじ’ , ‘ジョナゴールド’ , ‘SD’が かで頂生スパー形成率が左右された。5 c m 以上の新 9 . 8 %,7 0 . 8 %,8 5 . 2 %,せん定 それぞれ無せん定樹で 8 しょうになる割合は,‘ふじ’の 5年生樹と 9年生無せん 樹で 3 9 . 3 %,3 7 . 6 %,6 5 . 1 %であった。無せん定樹の 定樹の上向き枝および‘ジョナゴールド’の 9年生無せ えき生スパーの頂芽からは 2年後に極めて高い割合で頂 ん 定 樹 の 水 平 枝 で,そ れ ぞ れ 5 4 . 2 ,2 9 . 2 ,3 5 . 8 %と高 生スパーが形成された。 かったが,他は 1 5 %以下であった。5 c m 以上の新しょ 頂生スパー形成の過程をみると,えき生スパーから連 うを多く形成した木あるいは太枝は樹勢が強かった(第 続して頂生スパーを形成する場合と,えき生スパーから 5章第 2項の平均枝長参照)。従って,えき生スパーから 5 c m 以上の新しょうを生長させた翌年頂生スパーを形 頂生スパーを形成することに影響する要因は栄養生長の 浅 田 5 4 第5 -2 -1表 リンゴ 3品種のせん定樹と無せん定樹の平均枝長 z(%) 品種 せん定樹 2年枝 1年枝 無せん定樹(9年生) 当年枝 分散 2年枝 1年枝 当年枝 分析 1 1 . 5 9 . 1 6 . 6 1 0 . 8 a 1 3 . 7 a 4 . 6 b 6 . 7 a 8 . 0 a 2 . 3 b ** w * * 5 . 2 5 . 5 5 . 1 4 . 6 b 1 9 . 3 a 2 . 9 b 3 . 2 b 1 2 . 8 a 2 . 0 b ** ns ** 5 . 2 4 . 5 6 . 5 5 . 2 a 2 . 8 b 3 . 8 b 4 . 0 3 . 5 2 . 4 ** ** ns 上向き枝 y x ふじ J Gv SD 35. 2a 21. 6a 4. 2b 31. 4 27. 0 7. 7 37. 7 35. 5 19. 5 ふじ J G SD 9. 2ab 12. 9a 4. 3b 10. 6a 13. 5a 4. 7b 12. 9 a b 18. 3 a 7. 7 b ふじ J G SD 6. 8 8. 6 4. 8 9. 1a 10. 5a 4. 3b 7. 6 11. 3 5. 7 ふじ J G - - 水平枝 下向き枝 z y x w v (5年生) - - 1 8 . 6 4 3 . 5 - - 2 6 . 1 4 0 . 5 4 3 . 7 2 4 . 8 平均枝長は,せん定樹と 9年生無せん定樹では太枝・側枝単位,5年生無せん定樹では樹単位に算出 垂直を 0°として,上向き枝は 0~ 60° ,水平枝は 6 0~ 1 2 0 ° ,下向き枝は 1 2 0~ 1 8 0 °に発出した枝 異なる文字の付いた品種間に,Fi s her ’s PLSDにより 5 %水準で有意差あり せん定樹と 9年生無せん定樹の間に,**は 1 %,*は 5 %で有意差あり,nsは差なし J Gはジョナゴ-ルド,SDはスターキング・デリシャスの略 強さと考えられた。ただ, ‘SD’は太枝の発出角度にか のし方と種類別スパーの構成割合の関係を明らかにする かわらず 1 . 0~ 2 . 3 %と低く,えき生スパー頂芽からほ ことが必要と考えられる。 とんど全て頂生スパーを形成する性質を有していた。さ らに,せん定樹における 5 c m 以上の新しょう発生率は, 太枝の発出角度にかかわらず, ‘ふじ’と‘ジョナゴール 2.せん定が発生部位別スパー率に及ぼす影響 (1)せん定樹と無せん定樹における枝長の経年変化 ド’で 2 0~ 3 0 %と,無せん定樹より高かった。一方, 本研究で用いたせん定樹と無せん定樹は樹齢が異なっ ‘SD’は 4 . 4~ 8 . 4 %と無せん定樹よりは高いが,他の ている。樹齢の違いに伴って樹勢が異なると,枝構成に 2品種より低く‘SD’固有の性質と考えられた。以上の も影響を及ぼすと考えられるため,3年間にわたる枝長 結果は, ‘ふじ’と‘ジョナゴールド’では,せん定が樹 の変化を比較した。せん定樹では, ‘SD’の平均枝長が 勢を強くし,頂生スパー形成率を減少させることを示唆 ‘ふじ’と‘ジョナゴールド’より短く,品種による違 している。 いがみられた(第 5 -2 -1表)。この差異は,次項で述べ 次に,えき生スパー頂芽の 2年目の生長,すなわち頂 るように, ‘SD’が他の 2品種よりもスパー率が高いこ 生スパーから頂生スパー形成の要因についてみると,1 とによると考えられた。無せん定樹では,5年生全樹と 年目に頂生スパーを形成した場合,2年目の頂生スパー 9年生‘ジョナゴールド’を除いて,2年枝から当年枝へ 形成率は 5 c m 以上の新しょうになる割合だけでなく, と樹齢が進むにつれて長さが減少し,無せん定樹の 1年 生長を停止する割合によっても左右された。2年目に 枝および当年枝の長さは,せん定樹と比較して同程度か は,せん定樹の場合‘ふじ’ , ‘ジョナゴールド’ ,‘SD’ あるいは短かった。‘ふじ’と‘SD’については,9年 でそれぞれ 2 9 . 5 ,1 2 . 5 ,5 . 5 %の高い割合で生長を停止 生無せん定樹が樹齢の高いせん定樹と同程度からやや弱 した。特に‘ふじ’と‘ジョナゴールド’はせん定樹の い樹勢を有しており,枝構成の比較に供し得ると判断し 方が無せん定樹より生長停止率が高かった。よって,せ た。ただ,9年生‘ジョナゴールド’については,樹勢 ん定が 5 c m 以上の新しょうを増加させ,さらに生長停 の強さを考慮し,上向き枝と下向き枝は結果に含め,水 止率をも高くし,頂生スパー形成率を低下させることが 平枝は参考とした。 示唆された。 以上の結果,えき生スパーと頂生スパーの形成に最も (2)1年枝の長さ別構成割合 強く影響する要因は樹勢と品種の性質であり,特に,せ 1年枝の本数における長さ別構成割合を第 5 -2 -2表 ん定のし方が種類別スパーの構成割合に重要な役割を果 に示した。せん定樹における長枝率とスパー率の全平均 たすことが認められた。よって,各品種についてせん定 は,‘ふじ’では 2 5 . 3 %および 6 6 . 3 %, ‘ジョナゴール リンゴ樹における発生部位別スパー構成 55 第5 -2 -2表 リンゴにおける 1年枝の長さ別分布に及ぼす品種とせん定の影響(%) 品種 せん定樹 長枝 y 中果枝 検定 z 無せん定樹(9年生) スパー 長枝 中果枝 スパー 2 0 . 8 b 3 6 . 7 a 1 0 . 3 c 5 . 0 a 4 . 9 a 2 . 5 b 7 4 . 2 b 5 8 . 5 c 8 7 . 2 a ns ** * 1 5 . 4 b 3 5 . 3 a 7 . 4 c 6 . 1 a 2 . 3 b 1 . 9 b 7 8 . 6 b 6 2 . 4 c 9 0 . 6 a ns ** ns 1 0 . 3 9 . 6 5 . 9 3 . 2 3 . 6 1 . 7 8 6 . 8 8 5 . 8 9 2 . 4 ns ** * 1 5 . 5 b 2 9 . 0 a 8 . 8 c 4 . 9 3 . 5 1 . 7 7 9 . 6 b 6 7 . 6 c 8 9 . 5 a ** ** ** 上向き枝 x w ふじ J Gv SD 28. 8 21. 6 20. 8 10. 0a 2. 1b 2. 8b 61. 2 76. 3 76. 4 ふじ J G SD 21. 3a 21. 0ab 11. 2b 5. 6 2. 8 5. 3 73. 1 b 76. 2 a b 83. 4 a ふじ J G SD 18. 5a 23. 7a 7. 1b 7. 0 4. 7 6. 4 74. 5 a b 71. 6 b 86. 5 a ふじ J G SD 25. 3a 26. 0a 13. 5b 8. 3 3. 1 5. 4 66. 3 b 70. 9 a b 81. 1 a ふじ J G - - 水平枝 下向き枝 全平均 z y x w v (5年生) - - 4 7 . 6 5 . 0 - - 3 3 . 8 8 . 9 4 7 . 8 5 7 . 3 せん定の有無による結果枝の長さ別分布割合のχ 2検定,**,*はそれぞれ 1 %,5 %水準で有意差あり,nsは有意差なし 長枝は 10. 1 c m 以上,中果枝は 5. 1~ 10 c m,スパ-は 5 c m 以下 垂直を 0°として,上向き枝は 0~ 60° ,水平枝は 6 0~ 1 2 0 ° ,下向き枝は 1 2 0~ 1 8 0 °に発出した枝 異なる文字の付いた数値間に, Fi s her ’ sPLSDにより*は 5 %水準で有意差あり,nsは有意差なし J Gはジョナゴールド,SDはスターキング・デリシャスの略 ド’では 2 6 . 0 %および 7 0 . 9 %, ‘SD’では 1 3 . 5 %および パ ー 率 を 低 下 さ せ て い る こ と が 示 唆 さ れ た。Ma ggs 8 1 . 1 %となり,スパー率の割合が高かった。せん定樹に (1 9 5 9 )は,1年生苗木ではせん定が強いほどスパーが おける品種間比較では,‘SD’は‘ふじ’よりも水平枝 減少することを報告しており,本研究でもせん定が成木 と下向き枝で長枝率が低く,水平枝でスパー率が高かっ でも同様の現象を起こすことを示している。 た。 ‘ふじ’と‘ジョナゴールド’の間には有意な違いは みられなかった。無せん定樹では, ‘SD’のスパー率が (3)長枝の発生部位別構成割合 極めて高く,水平枝と下向き枝で 9 0 %を越えた。 えき生枝の数と長さは,スパータイプ品種の特徴や頂 La ur iら(1 9 9 5 )は,枝の生長と結実により分類した 部優勢性の違いを表す重要な要素である。また,えき生 リンゴの 3品種群の特性比較を行い,スパータイプは短 枝の密度は樹冠内の光環境などを左右するため,せん定 果枝結実性品種,‘ふじ’は頂端結実性品種,‘ジョナ の対象になる重要な枝である。そこで,長枝の頂生とえ ゴールド’ は両者の中間の性質を有する品種としている。 き生枝の本数における構成割合を太枝の発出角度別に検 本研究では, ‘SD’はスパータイプでないものの短果枝 討した。 結実性を示し,他の 2品種よりもスパー率が高かった。 結果を第 5 -2 -3表に示した。 ‘ふじ’, ‘ジョナゴー ‘ふじ’と‘ジョナゴールド’については両者に違いは ルド’,‘SD’におけるせん定樹の頂生とえき生別長枝率 みられず,さらに詳細な検討が必要と考えられる。 の全平均は,それぞれ 1 3 . 0 %と 1 2 . 4 %,1 3 . 2 %と 1 2 . 8 せん定樹と無せん定樹の比較では, ‘ふじ’ は全ての枝 %および 9 . 8 %と 3 . 7 %であった。せん定樹の水平枝と で, ‘ジョナゴールド’は下向き枝で, ‘SD’は上向き枝 無せん定樹の上向き枝では品種間差異が認められ, と下向き枝でせん定樹の方が無せん定樹よりもスパー率 ‘SD’は他の 2品種と比べて,頂生とえき生の長枝率が が低かった。しかし, ‘ジョナゴールド’ の上向き枝と水 低かった。‘ふじ’と‘ジョナゴールド’については,せ 平枝では逆にせん定樹の方がスパー率が高かった。 ん定樹の下向き枝と無せん定樹の上向き枝で‘ジョナ ‘ジョナゴールド’のこれらの枝は前項で述べたように ゴールド’の方がえき生枝率が高かったが,この違いは 特に平均枝長の長い枝であった。 品種の違いによるものかさらに検討が必要である。 以上の結果から,せん定作業は結果枝構成においてス 前項で述べたように,せん定によってスパー率は低下 浅 田 5 6 第5 -2 -3表 リンゴにおける 1年生長枝数の発生部位別分布割合に及ぼす品種とせん定の影響(%) 品種 せん定樹 無せん定樹(9年生) 頂生 えき生 ふじ J Gw SD 8. 2 10. 5 13. 3 20. 6 11. 1 7. 5 ふじ J G SD 13. 9a 13. 3a 8. 3b 7. 4a 7. 7a 2. 9b ふじ J G SD 14. 9 14. 0 6. 5 3. 6b 9. 8a 0. 6b ふじ J G SD 13. 0 13. 2 9. 8 ふじ J G - - χ 2検定 z 頂生 えき生 x 1 1 . 7 a 1 0 . 2 a 7 . 4 b 9 . 1 b 2 6 . 5 a 2 . 9 c ** ** ns 8 . 9 b 2 0 . 3 a 5 . 8 b 6 . 5 b 1 5 . 0 a 1 . 6 b ns ns ns 5 . 9 5 . 9 5 . 2 4 . 4 3 . 7 0 . 7 ** ns ns 9 . 4 a b 1 3 . 0 a 6 . 6 b 6 . 1 b 1 6 . 0 a 2 . 2 b ns ** ns 上向き枝 y 水平枝 下向き枝 全平均 z y x w 12. 4 12. 8 3. 7 (5年生) - 2 2 . 1 - 1 0 . 2 2 5 . 5 2 3 . 6 せん定樹と 9年生無せん定樹の間に**は 1 %,*は 5 %で有意差あり,nsは有意差なし 垂直を 0°として,上向き枝は 0~ 60° ,水平枝は 6 0~ 1 2 0 ° ,下向き枝は 1 2 0~ 1 8 0 °に発出した枝 異なる文字の付いた数値間に, Fi s her ’ sPLSDにより*は 5 %水準で有意差あり,nsは有意差なし J Gはジョナゴールド,SDはスターキング・デリシャスの略 し,長枝率は増加したが,頂生とえき生のどの長枝率が 頂生スパー率が高く,えき生スパー率が低い傾向がみら 増加するか検討した。樹勢の強い 5年生樹では,えき生 れた。無せん定樹でも,せん定樹と同様に,‘SD’が他 長枝率が‘ふじ’で 2 5 . 5 %, ‘ジョナゴールド’で 2 3 . 6 の 2品種より頂生スパー率が高かった。しかし,えき生 %と極めて高い割合を示した。せん定は樹勢を強くする スパー率については太枝の発出角度により異なった。こ ため,えき生長枝率を増加させると推察される。せん定 の結果は, ‘SD’の生長特性がえき生スパーよりも頂生 の有無による違いをみると, ‘ふじ’ の上向き枝と下向き スパーの形成に特徴があることを示唆している。 枝および‘J G’の上向き枝以外では有意な差はなく,太 スパーの発出部位別割合をせん定の有無で比較した結 枝の発出角度によってもえき生長枝率も異なった。この 果,‘SD’の水平枝以外の太枝で有意差が認められた。 理由としては,せん定によってえき生長枝が選択的に除 ‘ふじ’と‘ジョナゴールド’では,頂生スパー率につ 去されていることなどが考えられるが,今後の検討課題 いては,せん定樹の方が無せん定樹よりも低く,えき生 である。 スパー率については,‘ふじ’の上向き枝以外でせん定樹 の方が高かった。 ‘SD’では,下向き枝で他の 2品種と (4)スパーにおける発生部位別構成割合 同じ結果だったが,上向き枝と水平枝では異なった。せ 頂生スパーとえき生スパーの本数における構成割合の ん定樹の方が無せん定樹よりも頂生スパー率が低く,え 品種間およびせん定の有無による違いについて検討し, き生スパー率が高かった理由として,一つは,強勢の 5 その結果を太枝の発出角度別に第 5 -2 -4表に示した。 年生樹や 9年生の上向き枝は水平枝や下向き枝より頂生 ‘ふじ’ , ‘ジョナゴールド’ , ‘SD’におけるせん定樹の スパー率が低いことにみられるように(第 5 -2 -4表) , 頂生スパーとえき生スパーの構成割合の全平均は,それ せん定が樹勢回復の手段として行われていることが考え ぞれ 1 8 . 3 %と 4 8 . 1 %,2 1 . 9 %と 4 9 . 3 %および 4 6 . 1 %と られる。もう一つは,頂生スパーよりもえき生スパーに 3 5 . 1 %であった。せん定樹における両スパーの構成は品 多く着果させるためと考えられる。 種により異なり, ‘ふじ’と‘ジョナゴールド’では,太 えき生スパーは主に長い 2年枝のえき芽から発達し, 枝の発出角度にかかわらず頂生スパー率の方がえき生ス 一方,頂生スパーの大部分はスパーの頂芽から形成され パー率より低かった。しかし,‘SD’では,逆に頂生ス る。そのため,えき生スパーは“スパー枝齢”では若く, パー率の方が高かった。また,‘SD’は他の 2品種より 頂生スパーは若いスパーから高齢のスパーまで存在する。 リンゴ樹における発生部位別スパー構成 57 第5 -2 -4表 リンゴにおける 1年生スパー数の発生部位別分布割合に及ぼす品種とせん定の影響(%) 品種 せん定樹 頂生 無せん定樹(9年生) えき生 χ 2検定 z 頂生 えき生 2 4 . 1 b 2 4 . 5 b 4 2 . 3 a 5 0 . 1 a 3 4 . 0 b 4 4 . 9 a ** ** * 3 2 . 7 b 3 3 . 8 b 5 4 . 9 a 4 5 . 9 2 8 . 6 3 5 . 7 ** ** ns 4 6 . 1 b 5 3 . 8 b 6 3 . 6 a 4 0 . 7 a 3 2 . 0 a b 2 8 . 8 b ** ** * 3 4 . 8 b 3 7 . 2 b 5 3 . 0 a 4 4 . 9 a 3 3 . 1 b 3 6 . 5 b ** ** ns 上向き枝 y x ふじ J Gw SD 12. 3b 20. 0b 46. 2a 48. 9 56. 3 30. 2 ふじ J G SD 21. 2b 21. 0b 53. 2a 51. 9a 55. 2a 30. 2b ふじ J G SD 28. 8b 23. 2b 50. 6a 45. 7 48. 4 35. 9 ふじ J G SD 18. 3b 21. 9b 46. 1a 48. 1a 49. 3a 35. 0b ふじ J G - - 水平枝 下向き枝 全平均 (5年生) - 5 . 1 - 6 . 1 4 2 . 7 5 1 . 2 注は,第 5 -2 -3表を参照 一般に, “スパー枝齢”は果実品質に影響し,枝齢が高 パー率およびえき生スパー率の相互変化がどうなるかを くなると果実重が減少する(Ro bi ns o nら,1 9 8 3 ;Vo l z 調べるために,枝モデルを作り検討した。基本枝形成モ ら,1 9 9 4 ) 。そのため,リンゴ生産現場では,若いスパー デルを次のように設定した。 で生産する方がよいという考えがせん定者にあることが 推察される。ニホンナシでは,4~ 5年生側枝上の短果 (1)基本枝形成モデル 枝に形成される果実が最もよく発育するとされる(田村 枝形成の基本的条件を以下のようにする。 ら,1 9 9 0 )。本研究の結果は,リンゴでは若いスパーを ア.長枝の頂端から長枝を形成する。 重視したせん定がなされていることを示しており,ニホ イ.スパーの頂端からスパーを形成する。 ンナシとは異なっている。この違いをもたらす原因につ ウ.えき生枝を 1 0本発生する。 いては今後の課題である。 著者(20 0 5 )は,マルバカイドウ台開心形‘ふじ’の リンゴのせん定作業では,上向き枝などの強勢の枝で せん定樹の全スパー率,えき生スパー率および頂生ス はスパーの着生を促し,下向き枝などの弱勢の枝では結 パー率が,それぞれ約 8 0 %,5 0 %および 3 0 %程度の値 果枝の伸長を図るなど樹勢の調節を目的として行うが, になると報告した。1 0本のえき生枝のうち長枝とス 翌年の果実品質の向上や均質化も同時に意図して行って パーの本数をさまざま検討した結果,それぞれ 2本と 8 いる。本研究では,せん定作業には,長枝とスパーの発 本にした場合に一般的なせん定樹の構成割合になること 生部位別の構成割合の調節も含まれ,せん定が長枝率を が認められた。 増加させ樹勢を維持させていることを明らかにした。ス 枝モデルの樹姿と全,えき生および頂生の各スパー率 パーについては, ‘ふじ’と‘ジョナゴールド’では全ス の樹齢に伴う変化を第 5 -3 -1図に示した。この枝モデ パー率を減少させる場合,頂生スパーを減らし,えき生 ルは,全スパー率,えき生スパー率および頂生スパー率 スパーを増加させるようせん定を行っていることを明ら %,5 3 . 3 %および 2 6 . 7 %で,枝齢が進ん がそれぞれ 8 0 かにした。 ‘SD’では,頂生スパー率が極めて高いため, でもほとんど変化しない。樹冠を構成する枝は,発出角 下向き枝でのみ他の 2品種と同様の操作を行っているこ 度が異なっているためえき生と頂生のスパー率が枝によ とが認められた。今後さらに詳細な理論的解析を行う必 り違いがある。しかし,長枝とスパーの構成割合は, 要がある。 2 :8の割合が供試した‘ふじ’の平均値に近似した。 3.枝モデルによる発生部位別スパー率の調整法の検討 せん定の方法を変えた場合に,全スパー率,頂生ス 浅 田 5 8 A A 100 90 80 70 60 50 40 30 20 10 0 B スパ−率(%) スパ−率(%) B えき生 頂生 全 100 90 80 70 60 50 40 30 20 10 0 0 1 2 3 4 5 6 7 8 9 10 11 12 13 えき生 基本枝モデルにおける全,えき生および頂生 の各スパー率の経年変化。A:樹姿,B:各ス パー率の変化。えき生枝のえき生長枝 2本, スパー 8本 (2)頂生スパー率の増加方法 ①主軸枝からえき生の長枝が 1本も形成されず,スパー のみ形成される場合 全 0 1 2 3 4 5 6 7 8 9 10 11 12 13 年齢 第5 -3 -1図 頂生 年齢 第5 -3 -2図 枝モデルの樹姿と全,えき生および頂生の各 スパー率の経年変化。A:樹姿,B:各スパー 率の変化。えき生枝のえき生長枝 0本,ス パー 1 0本 両スパー率はほぼ同じ比率になり,約 4 5 %で一定に なった。第 5 -3 -2図と異なり,樹冠は立体化し,かつ 頂生スパー率が増加した。この結果は,「主軸から発生 枝モデルの樹姿と全,えき生および頂生の各スパー率 するえき生長枝を発達させ,えき生枝のえき生長枝を減 の樹齢に伴う変化を第 5 -3 -2図に示した。 らす方法」が頂生スパー率の増加法として有効であるこ 主軸からえき生長枝が発生しない場合は,側方に伸び とを示している。 る枝が無く全てスパーになるため,1 2年を経過しても 樹姿は一本棒状になる。これは,カラムナータイプと言 われる樹姿である。全スパーと頂生スパーの割合が 1 0 0 (3)えき生スパー率の増加法 ①えき生枝における長枝・スパー比を高める %に,えき生スパー率が 0 %に漸近する。頂生スパー率 えき生枝の長枝数を 4本,スパー数 6本にした場合の は増加するが,樹冠を立体的に拡大することができな 枝モデルの樹姿と全,えき生および頂生の各スパー率の い。 -4図に示した。 樹齢に伴う変化を第 5 -3 長枝・スパー比率が増加すると,全スパー,えき生ス ②えき生枝の長枝数を減らした場合(主軸枝のえき生枝 は不変) パーおよび頂生スパー率がそれぞれ 6 4 . 3 %,4 2 . 9 %, 2 1 . 5 %となった。えき生スパーを形成する長枝の比率を 主軸枝のえき生枝は発生するが,えき生枝のえき生枝 高くすると,全スパー率が低下し,えき生と頂生の両ス において長枝が少ない場合である。長枝数を 1本,ス パー率とも減少する。また,第 5 -3 -4図 Aのように樹 パー数を 9本とする。枝モデルの樹姿と全,えき生およ 冠内の枝密度が極めて高い枝構造になる。この結果は, び頂生の各スパー率の樹齢に伴う変化を第 5 -3 -3図に 長枝上のえき芽からの長枝増加が,単純にえき生スパー 示した。 率を増加させることにならないことを示している。 第 5 -3 -1図に比べ,樹冠内の枝の密度が低下してい る。全スパー率が 1 0 %増加し,約 9 0 %に達した。ま ②えき生枝のスパーの頂芽から長枝が生長する た,頂生スパー率も増加し,えき生スパー率が減少した。 頂生スパーからはスパーが発生することが多いが,第 リンゴ樹における発生部位別スパー構成 59 A A 100 90 80 70 60 50 40 30 20 10 0 B スパ−率(%) スパ−率(%) B えき生 頂生 全 100 90 80 70 60 50 40 30 20 10 0 えき生 年齢 枝モデルの樹姿と全,えき生および頂生の各 スパー率の経年的変化。A:樹姿,B:各ス パー率の変化。えき生枝のえき生長枝 1本, スパー 9本 全 0 1 2 3 4 5 6 7 8 9 10 11 12 13 0 1 2 3 4 5 6 7 8 9 10 11 12 13 第5 -3 -3図 頂生 年齢 第5 -3 -4図 枝モデルの樹姿と全,えき生および頂生の各 スパー率の経年的変化。A:樹姿,B:各ス パー率の変化。えき生枝のえき生長枝 4本, スパー 6本 1項で検討したようにせん定樹では長枝が発生すること 整法について検討した。 も普通にみられる。そこで,8本の頂生スパーのうち 2 無せん定の 5年生‘ふじ’における 2年枝のえき芽か 本から長枝が出る場合について検討した。樹姿と全,え ら生長する 1年枝の総数と 5 c m 以下のスパー数は,2 き生および頂生スパー率の経年変化を第 5 -3 -5図に示 年枝の長さ,直径,発出角度の 3変量との重相関関係と した。 2年枝長との単相関関係においてほぼ等しい決定係数が 全スパー率は 7 4 . 7 %で第 5 -3 -4図の 6 4 . 3 %と比べ えられ,2年枝長により決まることを示した。2年枝長 て減少が少ない。また,頂生スパー率は 1 7 . 9 %に減少す とえき生スパー数の関係は,Y=- 0 . 0 9 9+ 0 . 1 2 7 X(R2 るが,えき生スパー率が 5 6 . 8 %と第 5 -3 -4図の 4 2 . 9% =0 . 8 0 )であった。スパー 1本形成するのに必要な母枝 より大きく増加している。また, 第 5 -5 -4図 Aに比 長は 9 . 0 c m で あ っ た。 ‘ふ じ’, ‘ジ ョ ナ ゴ ー ル ド’ , べ,樹冠内の枝密度が低い。 ‘SD’の 9年生無せん定樹と成木せん定樹においても, 以上の結果は, 「スパーから長枝発生」が「長枝上のえ 母枝長と全えき生数およびスパー数の間に強い関係が認 き芽からの長枝増加」より,えき生スパー率を増加させ められた。 ‘SD’は回帰式の傾きが大きくスパー形成し る方法として有効であることを示している。スパーから やすい性質を示した。3品種がスパー 1本形成する母枝 長枝の発生を促すせん定は,いわゆる頂部優勢性を弱く 長の長さは,約 5 c m あるいはそれ以下であり,5年生無 し,枝の基部の生長を刺激するせん定と考えられる。栄 せん定樹の長さより短かった。 養生長の弱い枝では,切り戻しなどのせん定が有効と考 ‘ふじ’, ‘ジョナゴールド’,‘SD’の頂生スパーは, えられるが,その切り方等についてさらに検討する必要 えき生スパーの頂芽から形成される割合の平均値がそれ がある。 ぞれ,無せん定樹で 8 1 . 1 ,7 6 . 8 ,9 8 . 2 %で,せん定樹で 7 2 . 1 ,7 3 . 0 ,9 4 . 1 %であった。頂生スパー形成率は,無 摘 要 せん定樹がせん定樹より,また‘SD’が他の 2品種より 高かった。‘ふじ’と‘ジョナゴールド’の無せん定樹で リンゴ 3品種の無せん定樹とせん定樹を供試し,えき は,上向き枝より下向き枝の方が頂生スパー形成率高 生スパーと頂生スパーの形成要因およびせん定による調 く,樹勢に影響されることを示した。頂生スパーから形 浅 田 6 0 では下向き枝において頂生スパー率が低くえき生スパー A 率が高い構成になった。せん定作業が,長枝率を高める ことにより樹勢を維持し,また頂生スパー率を下げてえ き生スパー率を高めるように行われていることが示唆さ れた。 全スパー率,えき生スパー率および頂生スパー率を調 整する方法を明らかにするため,枝モデルを用いて検討 した。全スパー率が 8 0 %で,えき生スパー率と頂生ス パー率がそれぞれ 5 3 . 3 %,2 6 . 7 %の枝は,えき生枝の長 枝とスパーが 8 :2の比率で構成される枝であった。頂 生スパー率を増加させるには,主軸枝からえき生長枝を スパ−率(%) B 90 80 70 60 50 40 30 20 10 0 生長させ,それ以外の枝のえき生枝の長枝を減少させる ことが有効であった。えき生スパー率を増加させるに は,えき生長枝の発生を増加させることは全スパー率を 低下させるとともにえき生スパー率がわずかしか増えな いので有効でなく,頂生スパーから長枝を発生させるこ とが有効であった。 えき生 頂生 全 0 1 2 3 4 5 6 7 8 9 10 11 12 13 年齢 第5 -3 -5図 枝モデルの樹姿と全,えき生および頂生の各 スパー率の経年的変化。 A:樹姿,B:各ス パー率の変化。えき生枝のえき生長枝 2本, えき生スパー 8本のうち 2本のスパーから長 枝が出る 第 6章 総合考察 リンゴ生産において利用される主要な結果枝は,長さ 5 c m 以下のスパーである。スパーは発生部位により頂 生とえき生の 2種類に分けることができる。1年枝に占 めるスパーの割合が高ければ,1年枝に占める頂生とえ き生スパーの比率は相反する関係になる。実際のリンゴ 樹に着生する太枝を単位として,1年枝に占める頂生と えき生スパーの比率の相互関係を調べた結果,両者は直 成される頂生スパーの割合は, ‘ふじ’ , ‘ジョナゴール 線関係を示し,頂生スパー率が高くてえき生スパー率の ド’ , ‘SD’それぞれ,無せん定樹で 8 9 . 8 ,7 0 . 8 ,8 5 . 2 %, 低い枝からその逆の枝まで幅広く存在していることを認 せん定樹で 3 9 . 3 ,3 7 . 6 ,6 5 . 1 %であった。せん定がえき めた。これまで,スパーの分類については,着生する部 生スパーの頂芽から 2年後に形成される頂生スパーの割 位(セクション)の枝齢による分類がなされているが 合を低下させた。 (Ro bi ns o nら,1 9 8 3 ;田村ら,1 9 9 0 ;Vo l zら,1 9 9 4 ) , 無せん定樹とせん定樹を供試し,スパーを中心とする 発生部位別構成割合に関する報告はみられない。そこ 結果枝の長さ別および発生部位別構成割合に及ぼす品種 で,本研究では,両スパーの構成比率の違いが太枝およ とせん定の影響について検討した。せん定樹における長 び木全体の生長とどのような関係にあるかを検討した。 枝(1 0 c m <)とスパー(5 c m ≧)の構成割合は,‘ふ 第 2章では,せん定樹における両スパーの比率と枝の じ’ , ‘ジョナゴールド’ , ‘SD’でそれぞれ 2 5 . 3 %と 6 6 . 3 生長,花芽形成率および果実の生産性との関係を検討し %,2 6 . 0 %と 7 1 . 0 %および 1 3 . 0 %と 8 1 . 1 %であった。 た。しかし基本的な関係についてはせん定等の人為的操 ‘SD’のスパー率は他の 2品種より高く,無せん定樹で 作の加わらない無せん定樹で明らかにしておく必要があ は約 9 0 %に達した。 ‘ふじ’と‘SD’では,スパー率は ると考え,無せん定樹についても検討した(第 4章) 。栄 せん定樹の方が無せん定樹より低かった。長枝の頂生と 養生長との関係では,頂生スパー率が増加するにつれて え き 生 別 構 成 割 合 は,‘ふ じ’ , ‘ジ ョ ナ ゴ ー ル ド’, 栄養生長が弱くなるという関係が存在した。‘ふじ’ のせ ‘SD’でそれぞれ 1 3 . 0 %と 1 2 . 4 %,1 3 . 2 %と 1 2 . 8 %お ん定樹の場合,頂生スパー率と 2年枝の長枝率(1 0 c m よび 9 . 8 %と 3 . 7 %であった。 ‘SD’では,他の 2品種よ <)の間に決定係数 0 . 7 6~ 0 . 8 1で密接な負の相関関係が り頂生とえき生の長枝率とも低い傾向を示した。せん定 認められた。無せん定樹では,栄養生長の強さを平均新 樹における頂生スパーとえき生スパーの構成割合は, しょう長のほかに,1 ,2年枝長についても検討した結 ‘ふじ’,‘ジョナゴールド’, ‘SD’でそれぞれ 1 8 . 3 % 果,頂生スパー率との間にはせん定樹よりさらに密接な と4 8 . 1 %,2 1 . 9 %と 4 9 . 3 %および 4 6 . 1 %と 3 5 . 1 %で 負の相関関係が得られている。例えば,せん定樹の頂生 あった。せん定樹は無せん定樹に比べ, ‘ふじ’と‘ジョ スパー率と 1年枝の長枝率との関係では,決定係数が ナゴールド’では,水平枝と下向き枝において,‘SD’ 0 . 1 3~ 0 . 4 4と低く,かつ太枝によって変異が大きかった リンゴ樹における発生部位別スパー構成 61 のに対し,無せん定樹の頂生スパー率と平均 1年枝長の 性があるが,今後の検討課題である。 間では 0 . 8 7と高かった。頂生スパー率と平均新しょう 次に,発生部位別スパー率と果実生産性との関係につ 長との関係においても,無せん定樹の場合 0 . 6 4の決定係 いて検討した。果実の生産性は葉面積当たり果数で表し 数を得ている。また,無せん定樹において,毎年 0 . 5 c m た。葉面積当たり果数は,頂生スパー率が高くなるにつ 程度の極めて生長の弱い枝の割合と両スパー率の関係を れて増加する関係が認められた(第 2章第 2項)。すなわ みたが,頂生スパー率の増加とともに枝の生長が弱くな ち,頂生スパー率が高くなると,一定の範囲においては る結果が得られている。一方,同弱小枝率とえき生ス 果実の生産性が向上するという関係である。この関係が パー率の間では逆の関係を示し,えき生スパー率が増加 成り立つ理由は次のように考えられる。頂生スパー率が するにつれてその割合が減少した。 増加することは,1年枝当たりの新しょう数(芽の増加 以上の結果は,リンゴ樹の両スパー率と栄養生長の間 数)が減少することであり,1年枝当たり葉面積の減少 に,頂生スパー率が増加すると栄養生長が弱くなり,え をもたらすことになる。以上の理由から,1果当たり頂 き生スパー率が増加すると栄養生長が強くなるという基 芽数が同程度に維持される我が国の栽培では,頂生ス 本的な関係が存在することを示している。従来,スパー パー率が高くなると葉面積あたりの果数が増加する。す 率の増加は,樹が衰弱に向かう変化として知られていた なわち果実の生産性が向上することになる。 (Tho mps o n,1 9 6 6 ) 。しかし,本研究の結果によると, 発生部位別スパー率と果実品質の関係について検討し 同じスパーでもえき生スパー率の増加は栄養生長の強化 た(第 3章)。果実品質は,頂生スパー率の増加につれて を伴うため,従来の理解が充分でないことが判明した。 向上する形質と低下する形質が存在すると考えられる。 従って,リンゴ樹は頂生スパー率の増加に伴って衰弱に スパーは毎年短い生長を継続し,何年かスパー形成を継 向かうと表現すべきものと考えられる。 続しても全体として短い結果枝状の枝になる場合が多い 頂生スパー率とえき生スパー率の高い枝で栄養生長に ので,スパーが着生している領域(s e c t i o n)の年齢を 違いが生じる理由は次のように考えられる。長い新しょ )と呼ぶ報告がある(Ro bi ns o n “スパー枝齢”(s pura ge うは長い 1年枝から発生する場合が多い。えき生スパー ら,1 9 8 3 ;Vo l zら,1 9 9 4 ;田村ら,1 9 9 0 ) 。スパー枝齢 は長い 1年枝とともに形成されるので,長い新しょうと の違いと果実品質の関係を調べた結果,スパー枝齢が果 豊富な葉群に近い距離に位置する。従って,えき生ス 実の生長,糖度,酸度,Ca含量と密接な関係がみられて パー率の高い枝は,長い新しょうの多い栄養生長の盛ん いる。Vo l zら(1 9 9 4 )は,1~ 2年生スパーの果実が 3 な枝となる。一方,頂生スパーは長い新しょうとは遠い 年枝以上のスパーの果実より大きいこと,果実内 Caと 距離に位置する。スパー枝齢が高いほどその距離は大き Mg含量が 1年生枝の頂生果で最も多く,その他の年枝 くなる。そのため,頂生スパー率の高い枝は長い新しょ の 果 実 間 に 違 い は な か っ た と 報 告 し て い る。ま た, うの少ない栄養生長の弱い枝となる。 Ro bi ns o nら(1 9 8 3 )は,スパー枝齢が増すにつれて果実 花芽形成と両スパー率との間には,負の相関関係が認 の長さと幅,重量,可溶性固形物含量が減少し,果実の められた。とくに, ‘ふじ’の M.2 6台主幹形樹におい 硬さと滴定酸度が増加すると報告した。しかし,一方で て負の直線関係がみられた。開心形樹では,各頂生ス 3~ 4年枝に形成される‘Co x’ sOr a ngePi ppi n’リンゴ パー率における最大花芽形成率が頂生スパー率の増加と 果実はそれより若いか,あるいは古い枝に形成される果 ともに減少するという関係であった。これらの結果は, ks o n,1 9 6 7 ) 。本研 実より大きいとする報告もある(J a c 頂生スパーが増えるにつれて花芽形成が困難になること 究では,4年生より古いスパーの果実が 3年生より若い を示している。前述のように,頂生スパー率の増加が樹 スパーの果実より可溶性固形物含量が少なく,果実が小 勢の減少を伴うとすれば,頂生スパー率の増加によって さくなる傾向を示したのに対し,Ca含有量が増加する 花芽形成が増加すると推察される。しかし,結果は逆で ことが認められた。 あった。この理由の一つとしては,頂生スパーとえき生 以上の報告に共通する関係は,5年生より古いスパー スパーの花芽形成に異なる条件が存在するためと考えら では果実の発育や可溶性固形物含量が劣ることである。 れる。えき生スパーが発出する芽はえき生であり,花芽 枝齢の大きいスパーで構成する太枝は頂生スパー率が高 を形成しても果実品質が劣るため,我が国では摘果す いとみなされるため,品質のよい果実を生産するには適 る。従って,えき生スパーの頂芽における花芽形成は着 度な頂生スパー率に維持することが必要と考えられる。 果の影響を受けにくい。しかし,頂生スパーは 2 0 %か 発生部位別スパー率と果実品質の基本的な関係は,長い ら3 0 %の芽が着果するため,着果の花芽形成阻害影響 新しょうが着生する群葉の豊富な部位との距離で説明す を受けるスパーが存在することになる。ただし,諸外国 ることができる(Te ngら,1 9 9 8 )。栄養生長の盛んな部 では,えき芽果も収穫の対象にされているので,えき生 位に位置するえき生スパー果は,同化養分の豊富な供給 スパー率が高くても花芽形成率の低い場合がみられるこ と新しょう生長との強い競合により発育のよい果実と不 とも考えられる。頂生スパー率の増加に伴う花芽形成率 良な果実が幅広く存在する。一方,頂生スパー果は貯蔵 の減少については,着果以外の要因も関与している可能 養分と展開の早いスパー葉に恵まれ,速度は遅いがよく 浅 田 6 2 今後さらに検討が必要である。 平均新しょう長 頂生スパー率は人工的に調整され得る性質である。調 花芽形成率 整手段の一つはせん定であり,せん定が強くなると頂生 平均2年枝長 スパー率の低下,えき生スパー率の増加につながる。せ 葉面積当たり果数・果実 Ca ん定以外にも,施肥,着果程度,光条件などが影響する と考えられるが,さらに検討が必要である。 頂生スパー率がせん定により調整が可能であること, またスパーの着生状況を眼で識別できることから,頂生 指数 スパー率を用いて種々のせん定技術を分析することや初 心者用せん定マニュアルの作成などに利用することがで きると考える。そのため,せん定技術と密接に関係する と考えられる発生部位別スパー形成要因および各スパー 率の調整法について検討した。 えき生スパーの形成は,前年枝の長さが最も重要な要 0 10 20 30 40 50 60 70 頂生スパ−率(%) 因であった。‘ふじ’の無せん定若木の場合,えき生ス パーを形成するために必要な前年枝の長さは 9 c m 以上 であった。前年枝の長さが増すにつれてえき生スパーの 90 ←えき生スパー率(%)→ 20 本数が増加した。前年枝の太さについても検討したが, ← 強せん定・・無せん定 → 長さほど密接な関係はみられなかった。発出角度が広 ← 上向き枝・・下向き枝 → ← 多窒素・・無施肥 → ← 無着果・・過着果 → い,すなわち前年枝が下を向くほどえき生スパーの最大 発生数が減少するが,その理由は下を向くほど前年枝の 長さの最大長が減少するためであった。これらのことか 第 6図 頂生スパー率と栄養生長,花芽形成率および果実生 産効率との相互関係を示す模式図 ら,えき生スパーの本数を増加させるためには,長い枝 を形成することが条件となることが認められた。 頂生スパーは,様々な長さの前年枝から形成される 揃った発育をすると推察される。 が,とくにスパーから形成される頂生スパーが多くを占 以上の結果を模式的に示したものが第 6図である。頂 める。‘ふじ’の開心形せん定樹では,全頂生スパーのう 生スパー率の増加に伴って増加する形質と減少する形質 ち平均 8 3 . 3 % が ス パ ー か ら 形 成 さ れ て い た。頂 生 ス とがある。前者は栄養生長に関する要素と花芽形成であ パーは,えき生スパーに形成されるものから何年もス り,後者は葉面積当たり果数と果実内 Ca含量である。 パーを継続した後に形成されるものまで存在する。いず なお,Ca含量との関係についてはさらに詳細な検討を れにしてもスパー形成の条件は短い枝を形成することで 必要とする。これらの関係は,リンゴ生産においては, あるが,短い枝の頂芽の全てが頂生スパーを形成するわ 頂生スパー率の望ましい範囲が存在することを示唆して けではない。長枝を発生したり,枯死するスパーなど変 いる。その範囲については,頂生スパー率の最低限界点 異がある。えき生スパーから頂生スパーが形成されるの を葉面積当たり果数との関係でみるのが適切と考えられ は,せん定樹の場合,‘ふじ’と‘ジョナゴールド’で る。葉面積当たり果数(Y)と頂生スパー率(X)の関 9 . 3 %である。2 0 . 7~ 2 9 . 9 %が 5 c m より長 は,7 0 . 1~ 7 係が,開心形樹‘ふじ’の場合,Y= 0 . 1 X+ 3が平均的 い枝になり,頂芽が枯死するものがほとんど無いためで な関係となることから,葉面積当たり果数を 4以上必要 ある。 この割合は太枝の発出角度によって違いはみら と考えると,頂生スパー率は 1 0 %以上が望ましくなる。 れなかった。また,えき生スパーから頂生スパーを形成 頂生スパー率 1 0 %は,栄養生長を平均新しょう長でみ した後,さらに翌年頂生スパーを形成する枝の割合は, ると,第 4 -2 -1図のように急激に増加する転換点に当 無せん定樹とせん定樹で違いが認められた。無せん定樹 たっている。一方,頂生スパー率の最高限界点は,弱小 の方が 7 6 . 4~ 8 5 . 1 %とせん定樹の 3 1 . 4~ 3 1 . 5 %に比べ 枝率との関係から推定できる。‘ふじ’の無せん定樹で 高かった。この違いは,せん定により 5 c m より長い枝 は,老衰枝率が頂生スパー率の増加とともに増加する と枯死する頂芽が 3 4 . 5~ 4 0 . 7 %と増加するためである。 (第 4章第 2項)が,頂生スパー率が 5 0 %を越えると急 なお,このうち枯死する頂芽の割合は品種により違いが 増する。 大きくさらに検討を要するが,約 1/ 2を占めると考え 以上より,‘ふじ’の頂生スパー率は,大枠として 1 0 られた。リンゴにおいてせん定が芽の枯死率を増加させ %から 5 0 %の範囲が考えられたが,望ましい果実品質 ることは La ur iら(1 9 9 5 )も報告している。 などの条件によりさらに限定された範囲が存在すること 以上の結果は,頂生スパーを減少させる方法として, も予想される。その他の品種における適正範囲も含めて 頂生スパーを除去する方法の他に,スパーから長枝を発 リンゴ樹における発生部位別スパー構成 63 生させるせん定が行われていることを示唆している。長 ることができる。そのため,発生部位別スパーの全 1年 枝を発生させるせん定は,同時にえき生スパーを増加さ 枝に占める比率をそれぞれ頂生スパー率,えき生スパー せることになる。また,頂生スパー率を高くする方法と 率とすると,スパー率がある段階に達したリンゴ樹で しては,長枝を減らすせん定が考えられる。ただし,樹 は,高えき生スパー率・低頂生スパー率の太枝から低え 勢を強くすると全スパー率が低下する可能性があるた き生スパー率・高頂生スパー率の太枝まで幅広くさまざ め,間引きせん定による長枝の減少が主なせん定方法に まな枝が存在することになる。そこで,発生部位別ス なると考えられる。すなわち,主軸になる長枝でなく, パー率の異なる太枝が,栄養と生殖の両生長においてど えき生の長枝をせん去する間引きせん定である。なお, のような違いを示すかを明らかにし,両生長のメカニズ ‘SD’はスパー形成率が極めて高く,前 2品種とは違っ ムに関する情報を得ようとした。また,種類別スパーの て頂生スパー形成にせん定の影響は認められなかった。 形成要因について検討し,その成果に基づいて種類別ス 前述のように,頂生スパー率とえき生スパー率には適 パー率の調整方法を確立しようとした。 正範囲があると考えられるが,樹の生長状態により適正 栄養生長との関係では,頂生スパー率が高くなると栄 範囲に近づける方法が異なる。例えば,若木や若い枝で 養生長が弱くなり,えき生スパー率が増加すると逆に栄 は頂生スパー率を速やかに増加させるような,高齢樹や 養生長が強くなる関係が認められた。頂生スパーが多い 年数を経た枝ではえき生スパー率を高めるせん定が必要 ことは,主に短い枝の頂芽に形成されるので,短い生長 となる場合が多い(第 5章第 1項) 。その際には,全ス を年々繰り返し,次第に衰弱することを意味する。一 パー率の維持および増加を前提として,頂生スパー率と 方,えき生スパーが多いことは,主に長い 2年枝のえき えき生スパー率の増減を計らなければならない。しか 芽から形成されるので,長い新しょうが距離的に近い位 し,頂生スパーはえき生スパーから形成されるものが含 置に存在し,生長の盛んな部位が多いことを意味する。 まれることから,両スパー率は連動して変化する。その 以上の形態形成上の違いが,両スパー率による栄養生長 ため,全,頂生およびえき生スパーの相互変化をみなが の相反する性質を特徴付けている。 ら種々のせん定法が各スパー率をどう変化させるかを知 生殖生長との関係では,頂生スパー率が増加するにつ る必要がある。そこで,枝モデルを用いていくつかのせ れ,花芽形成率が低下する関係が認められた。また,果 ん定方法の比較を行った。長枝からえき生枝が,長枝 2 実生産性との関係については,頂生スパー率がある程度 本,スパー 8本計 1 0本発達するモデルを基準として,長 高い方が効率よく果実生産できることを明らかにした。 枝を 1本に減らすと,えき生スパーが減少する一方で頂 果実生産性,すなわち葉面積当たり果数は,1年枝当た 生スパーが増加し,両スパー率が約 4 5 %で一定となっ り新しょう発生数(頂芽増加率)あるいは同葉面積が少 た。主軸となる長枝からのえき生長枝を無くすとカラム ない方が大きくなるが,頂生スパー率の増加とともにそ ナータイプに近い形となるが,主軸に着生するえき生枝 れらが減少することが果実生産性向上の理由と考えられ から発達するえき生長枝を減らすせん定は,頂生スパー た。 率を増加させるのに有効である。また,えき生スパー率 果実品質との関係では,頂生スパー率が高くなり,4 を高めるために,えき生枝における長枝/スパーの比を 年以上スパー形成を繰り返した頂生スパーが増加すると 高めるせん定とえき生スパーの頂芽から長枝を発生させ 果実の大きさや可溶性固形物含量などの果実品質が低下 るせん定を比較した。その結果,長枝/スパー比を高め するが,Ca含量は増加することが認められた。 るせん定は,えき生スパー率は高く維持できるが,枝の 発生部位別スパー率と栄養・生殖生長の関係は,無せ 密度が急増するとともに全スパー率が低下することにな ん定樹とせん定樹について得られた結果であり,リンゴ る。それに対し,一部のスパーから長枝を発達させるせ 樹において基本的に備わった法則性であることを明らか ん定は,全スパー率が高く維持され,えき生スパー率も にした。頂生スパーとえき生スパーは,どちらか一方が 増加させたため,えき生スパー率を高くするせん定法と 良品生産のために貢献するのでなく,両スパー率ともあ して有効であると考えられた。 る幅をもって維持される必要がある。その望ましい両ス 本研究で得られた成果は,発生部位別スパー率を調整 パー率の範囲は,品種,樹齢,土地条件および品質基準 するせん定方法についてさらに詳細に検討することによ などによって違いがあるが,今後の課題としたい。 り,初心者用のせん定マニュアルを作成すること,また, 太枝を単位としてみると,頂生スパー率とえき生ス 生産者の間で工夫されている種々のせん定技術を科学的 パー率は,樹内変異が大きいことから,せん定により調 に説明することにも利用できると考えられる。 整可能な性質と考えられる。そこで,無せん定樹とせん 定樹の枝構成の違いを分析し,各スパーの形成要因を明 第 7章 総 括 らかにすることにより,せん定による各スパー率調整法 を明らかにしようとした。せん定樹と無せん定樹の比較 リンゴの生産を担う主要な結果枝はスパーである。ス より,せん定樹はえき生スパー率を高くするよう調整し パーは,発生部位により頂生とえき生の 2種類に大別す ていることが示唆された。一般的には,えき生スパー形 浅 田 6 4 成を促すことにより,若い枝で結果部位を構成するせん 定が行われていると推察される。えき生スパーの形成を 促すには,母枝の性質の中で枝長が最も重要であり,あ る程度長い母枝を形成する必要がある。しかし,長い母 枝を増加させると,樹勢が強くなり,全スパー率そのも のを低下させる可能性がある。そこで,枝モデルを使用 して,せん定方法と全,頂生およびえき生の各スパー率 との相互関係を検討した結果,えき生スパー率を高める には,頂生スパーから頂生スパーを形成する割合を低下 させ,頂生スパーから長枝をある比率で発達させるせん 定が有効であることが判明した。一方,若い樹などにお いて頂生スパーを増加させるには,主軸となる枝でな く,側方に発達するえき生長枝を減らす方法が有効で あった。 これらの研究成果は,いろいろなせん定技術の特徴を 発生部位別スパー率から解析することを可能にし,初心 者用せん定マニュアル作成に活用できると考えられる。 謝 辞 本研究を遂行するに当たり,貴重な助言と校閲の労を 賜った弘前大学農学生命科学部荒川修教授には衷心より 感謝の意を表します。学位審査委員として本論文のご精 査を賜った岩手大学農学部寿松木章教授,山形大学農学 部平智教授,さらに本論文の審査ならびにリンゴ樹の調 査に便宜を図って頂いた弘前大学農学生命科学部附属生 物共生教育研究センター塩崎雄之輔教授に深甚なる謝意 を表します。本研究で行った調査は,弘前大学農学部果 樹園芸学教室所属の学生であった駒井秋浩氏,工藤智 氏,三上義勝氏,小笠原理高氏,鳴海東之氏,田端聡子 氏,藤井亨氏,堀田拓人氏,佐藤透氏および佐々木拓氏 の方々のご協力を得て行った。以上の方々に衷心より感 謝を申し上げる次第である。 引 用 文 献 ABBOTT,D. L.1960.The bour s es hootas a f ac t or i nt he gr o wt hofappl ef r ui t s .Ann.Appl i edBi ol .48:4 3 4 4 3 8 . 青森県.2000 .平成 12年りんご生産指導要項.青森県農林 部りんご課.青森. ARASU, N. T. 1968. Spur t i pe s por t si n appl e. Rpt . E. Ma l l i ng Res .St a.1967.p.113119. 浅田武典.1988 .開心形リンゴ樹の果実生産性に関する研 究.第 2報 葉面積指数,単位葉面積当たり果数,収量 の相互関係.弘大農報.50:4654. 浅田武典.1989 .開心形リンゴ樹の果実生産性に関する研 究.第 3報 収穫果の果径別分布と発育良果の多収につ いて.弘大農報.51:6979. ASADA,T.1 999.St udi es o n yi el de f f i c i enc yo ff r ui t i ng uni t si n‘Fuj i ’Appl et r ees .J .J apan.Soc .Ho r t .Sc i .6 8 :108 41089. 浅田武典.2004 .短果枝の発生部位別構成割合がリンゴ樹の 栄養および生殖生長に及ぼす影響.園学雑.73 :1 4 3 - 1 4 9 . BARRITT,B. H.C. M.ROM,K. R.GUELISH,S. R.DRAKE,a nd M. A. DILLEY.19 8 7 .Ca no py po s i t i o n and l i ght ef f ec t so n s pur ,l e a f ,a nd f r ui tc ha r a c t e r i s t i c so f ‘De l i c i ous ’ a ppl e .Ho r t Sc i e nc e2 2:4 0 2 4 0 5 . BARRITT, B. H. a nd B. J . SCHONBERG. 19 9 0 . Cul t i var a nd c a no py po s i t i o ne f f e c t so ns e a s o na lde ve l o pme nt of ve ge t a t i ves pur so fa ppl e .Ho r t Sc i e nc e2 5:6 2 8 631. BRUNNER, T. 19 9 0 . Phys i o l o gi a lf r ui tt r e et r a i ni ng f or i nt e ns i vegr owi ng.Aka demi a iKi a do ,Buda pe s t . CAIN,J . C.1 9 7 1 .Ef f e c t so fmec ha ni c a lpr uni ng ofappl e he dge r ows wi t h as l o t t i ng s aw o nl i ghtpe net r at i on a ndf r ui t i ng.J .Amer .So c .Ho r t .Sc i .9 6:6 6 4 6 67. COSTES,E. ,C.GODIN a nd Y.GUEDON.1 9 9 7 .A me t hodol ogy f o rt he e xpl o r r a t i o no ff r ui tt r e es t r uc t ur e s .Ac t a Ho r t i c .4 5 1:7 0 9 7 1 5 . CHRISTOPHER,E. P.1 9 7 4 .The pr uni ng manua l .Mac mi l l an Publ i s hi ng,Ne w Yo r k,pp.7 8 7 9 .De nne ,M. P.1963. Fr ui tde ve l o pmenta nd s omet r e ef a c t o r sa f f e c t i ng i t . Ne w Ze a l a nd J o ur .Bo t .1:2 6 5 9 4 . CURRY,E. A.a nd N. E.LOONEY.1 9 8 6 .Ef f e c to fc r o ppi ng on s ho o t gr owt ho fs pur t ype‘Go l de n De l i c i o us ’t r ees . Ho r t Sc i e nc e2 1:1 0 1 5 1 0 1 7 . DENNE, M. P. 1 9 6 3 . Fr ui t de ve l o pme nt a nd s ome t r ee f a c t o r sa f f e c t i ngi t .Ne w Ze a l a ndJ o ur .Bo t .1:265294. ERF,J . A.a nd J . T. A.PROCTOR.1 9 8 7 .Cha nge si na ppl el eaf wa t e rs t a t usa nd ve ge t a t i vegr owt ha si nf l ue nc ed by c r o pl o a d.J .Amer .So c .Ho r t .Sc i .1 1 2:6 1 7 6 2 0 . FERREE, D. C. 1 9 8 0 . Ca no py de ve l o pment and yi el d e f f i c i e nc yo f ‘Go l de n De l i c i o us ’a ppl et r e e si nf our o r c ha r d ma na geme nt s ys t ems .J .Amer .So c .Ho r t . Sc i .1 0 5:3 7 6 3 8 0 . FERREE, D. C. a nd J . W. PALMER. 1 9 8 2 . Ef f e c to fs pur de f o l i a t i o na nd r i ngi ng dur i ng bl o om o nf r ui t i ng,f r ui t mi ne r a ll e ve l ,a nd ne t pho t o s ynt he s i so f ‘Gol den De l i c i o us ’a ppl e .J .Amer .So c .Ho r t .Sc i .1 0 7 :11821 1 8 6 . FORSHEY,C. G.a nd D. C.ELFVING.1 9 7 7 .Fr ui tnumbe r s ,f r ui t s i z e ,a nd yi e l dr e l a t i o ns hi ps i n‘Mc I nt o s h’a ppl es .J . Amer .So c .Ho r t .Sc i .1 0 2:3 9 9 4 0 2 . FORSHEY C. G.a nd D. C.ELFVING.1 9 7 9 .Es t i mat i ng yi el d and f r ui tnumbe r so fa ppl et r e e sf r om br a nc hs ampl es .J . Amer .So c .Ho r t .Sc i .1 0 4:9 8 7 9 0 0 . FORSHEY, C. G. 19 8 2 b. Ef f e c t so ff r ui t i ng, pr uni ng, a nd ni t r o ge nf e r t i l i z a t i o no ns ho o t gr owt ho f ‘Empi r e ’ a ppl et r e e s .J .Amer .So c .Ho r t .Sc i .1 0 7:1 0 9 2 1097. GATSUK L. A. ,SMIRNOVA O. V,VORONTSOVA L. I . ,ZAUGOLNOVA L. B. ,ZHUKOVA L. A.1 9 8 0 .Age s t a t e s of pl ant s of va r i o usgr owt hf o r ms:ar e vi ew.J .Ec o .6 8:6 75696. 猪俣雄司・工藤和典・和田雅人・増田哲男・別所英男・鈴木 邦彦.2 0 0 4.カラムナータイプリンゴ‘メイポール’の 樹体生育・果実生産効率・乾物生産特性に及ぼす整枝法 の影響.園学研.3:3 8 7 3 9 2 . J . E. 1 9 6 7 . Va r i a bi l i t y i n f r ui ts i z ea nd c ol or ACKSON. J wi t hi ni ndi vi dua lt r e e s .Rpt .E.Mal l i ng Re s .St a.1966. p.1 1 0 1 1 5 . J . E.1 9 7 0 .I ndi vi dua lf r ui ts i z ei nr e l a t i o nt o age ACKSON,J o f be a r i ng wo o do n‘La xt o n’ s Supe r b’a ppl et r ees . Rpt .E.Mal l i ngRe s .St a .1 9 6 9 .P.8 3 8 5 . J . E.a nd J . W.Pa l me r .1 9 7 7 .Ef f e c t so fs hade on ACKSON,J t he gr owt h a nd c r o ppi ng of a ppl e t r e es .Ⅰ . Expe r i me nt a l de t a i l s a nd ef f e c t s on ve get at i ve リンゴ樹における発生部位別スパー構成 gr owt h.J .Ho r t .Sc i .52:245252. KIKUCHI,T.1974.Gr owt h and s t r uc t ur e of t he s ho o t s ys t e m o fa ppl e t r ees a s c har ac t er i zed by t he f r equenc y di s t r i but i on of s hoot l engt hs .Bul l .Fa c . Agr i c .Hi r os akiUni v.12:5065. 菊池卓郎.1975 .せん定と生長・結実.青木二郎編,新編リ ンゴの研究.p.273293.津軽書房.青森. 小林章・吉村不二男.1953.遮光が果樹の苗木の生育に及ぼ す影響.園芸学研究集録.6:6468. 小池洋男・吉沢しおり・塚原一幸.1990.リンゴわい性台樹 の適正着果量と乾物生産の分配.園学雑.58:82 7 8 3 4 . 熊代克巳・西村昇.1984.受光量がリンゴ樹の生育に及ぼす 影響.第 2報.遮光が樹体生育,花芽形成および果実品 質に及ぼす影響.園学要旨.昭 61春 :5051. LAKSO, A. N.1 984. Leaf ar ea devel opment pat t e r ns i n young pr uned and unpr uned appl et r ees .J .Amer . Soc .Ho r t .Sc i .109:861865. LAPINS,K. O.1974.Spur t ype gr owt h habi ti n 60 a ppl e pr ogeni es .J .Amer .Soc .Ho r t .Sc i .99:568572. LAURI,P. E. ,E.TEROUANNE,J . M.LESPINASSE,J . L.REGNARD a nd J . J .KELNER,1995.Genot ypi cdi f f er enc esi nt hea xi l l a r y bud gr o wt h and f r ui t i ng pat t er n of appl ef r ui t i ng br anc hes o ver s ever al year s : An appr oa c h t o r egul at i on off r ui tbear i ng.Sc i ent i a Ho r t i c .64 :2 6 5 281. LAURI, P. E. ,J . M. LESPINASSE and E. TEROUANNE. 19 9 7 . Ve get at i ve gr owt h and r epr oduc t i ve s t r at egi e si n appl ef r ui t i ng br anc hes -an i nves t i gat i on i nt o va r i o us c ul t i ver s .Ac t aHo r t i c .451:717724. LAURI,P. E. ,J . M.LESPINASSE and F.LAURENS.1997.Wha t ki nd of mo r phol ogi c al t r ai t ss houl d be s ought i n appl es eedl i ng pr ogeni es i n or der t os el ec tr e gul a r bear i ngc ul t i var s?Ac t aHo r t i c .451:725729. LAURI,P. E.and J . J .KELNER.2001.Shoott ype demo gr a phy and dr y mat t er par t i t i oni ng : a mor pho me t r i c appr oac hi n appl e (Mal us ×dome s t i c a) .Can.J .Bo t .7 9 :12701273. LESPINASSE. J . M. a nd J . F. DELORT. 1 986. Appl e t r e e ma nage me nt i n ver t i c al a xi s :a ppr ai s al a f t e rt e n year sofexper i ment s ,Ac t aHo r t i c .160:139155 . LORETI, F. 1 990. St r uc t ur al ma ni pul at i on f or i mpr o ve d per f or ma nc ei nwoodypl ant s .Ho r t Sc i enc e.25:6 4 7 0 . MAGGS,D. H.1959.Thepr uni ngr es pons eofoneyeara ppl e t r ees .Ann.Bot .23:319330. MAGGS,D. H.1960.The s t abi l i t y oft he gr owt h pat t e r no f young appl et r ees under f our l evel s ofi l l umi na t i o n. Ann.Bot .24:434450. MAGGS,D. H.1963.Ther educ t i on i n gr owt h ofappl et r e e s br oughtaboutbyf r ui t i ng.J .Ho r t .Sc i .38:1191 2 8 . 望月武雄.1962 .りんご樹において惹起される樹勢衰弱現象 の解明に関する研究.弘大農報.8:40126. MULLINS,M. G.1965.The gr avi t at i onalr es pons esofyo ung appl et r ees .J .Ho r t .Sc i .40:237247. MYERS S. C.and D. C.FERREE.1983.I nf l uenc e oft i me o f s ummerpr uni ng and l i mb or i ent at i on on yi el d,f r ui t s i ze,and qual i t y ofvi l i c i ous ’appl et r e e s . gor ous ‘De J .Amer .Soc .Ho r t .Sc i .108:630633. 小原信実.1984 .整枝剪定.p.5388.津川力編著.リンゴ栽 培技術.養賢堂.東京. PALMER,J . W.1 989.Canopy mani pul at i on f or opt i mum ut i l i zat i on ofl i ght .p.245262.I n :V.J .Wr i ght (e d) . Mani pul at i onoff r ui t i ng.But t er wo r t hs ,London. 65 PORTER,J . R.a nd F. W. M.LLEWELYN.1 9 8 4 .Thea r c hi t ec t ur al dynami c so fpr une da ppl et r e e s .J .Ho r t .Sc i .59:3133 2 1 . PRIESTLEY,C. A.1 9 6 9 .Some a s pe c t so ft he phys i ol ogy of a ppl er o o t s t o c k va r i e t i e sunde rr e duc e di l l umi nat i on. Ann.Bo t .3 3:9 6 7 9 8 0 . QUELLETTE D. V. And E. YOUNG. 19 9 5 . La t e r a ls hoot de ve l o pme nt i ns i x di ve r s es e e dl i ng po pul a t i ons of a ppl e .Fr ui tVa r i e t i e sJ o ur na l .4 9:2 4 8 2 5 4 . QUINLAN,J . D.a nd K. R.TOBUTT.1 9 9 0 .Mani pul a t i ng f r ui t t r e e s t r uc t ur e c hemi c a l l y and ge ne t i c a l l y f or i mpr o ve dpe r f o r ma nc e .Ho r t Sc i e nc e .2 5:6 0 6 4 . ROBINSON,T, L. ,E. J .SEELEY,a nd B. H.BARRITT.1 9 8 3.Ef f ec t o fl i ght e nvi r o nme nt a nd s pur a ge o n‘De l i c i ous ’ a ppl ef r ui ts i z e .J .Amer .So c .Ho r t .Sc i .1 0 8:8 55861. RUBINSTEIN, B. a nd M. A. NAGAO. 1 9 7 6 . La t e r al bud o ut gr owt h and i t sc o nt r o l by t he ape x.Bot ani c al Re vi ew 4 2:8 3 1 1 3 . ROM,C. R.a nd B. H.BARRITT.1 9 9 0 .Spur de ve l o pment of ‘De l i c i o us ’a ppl ea si nf l ue nc e dbypo s i t i o n,woodage, s t r a i na ndpr uni ng.Ho r t Sc i e nc e1 8:5 8 5 5 8 7 . SEELEY, E. J . , W. C. MICKE, a nd R. KAMMERECK. 1 980. ‘De l i c i o us ’a ppl ef r ui ts i z ea nd qua l i t ya si nf l uenc ed by r a di a t e gr o wi ng a nt f l ux de ns i t yi nt he i mmedi e nvi r o nment .J .Amer .So c .Ho r t .Sc i .1 0 5:6 4 5 657. 柴壽.1 9 9 4.整枝せん定の基本用語.p.3 9 5 2 .農耕と園芸編 集部編.落葉果樹の整枝せん定.誠文堂新光社.東京. 塩崎雄之輔・菊池卓郎.1 9 8 8.開心形リンゴ園の収量経過と 葉面積指数,樹勢.園学要旨.昭 63秋 :1 2 0 1 2 1. SUZUKI,T.1 9 9 0 .Api c a lc o nt r o lo fl a t e r a lbud de ve l opment a nd s ho o tgr owt hi n mul be r r y(Mo r u sa l b a) .Phys i ol . Pl a nt .8 0:3 5 0 3 5 6 . 田村文男・田辺賢二・林真二・伴野潔.1 9 9 0.ニホンナシ ‘二 十世紀’の側枝年齢と果実発育との関係.園学雑.59: 9 1 9 8 . 田村太郎.1 9 7 5.全糖の定量法.作物分析法委員会編:栄養 診断のための栽培植物分析測定法.p.2 8 9 2 9 3 .養賢堂. 東京. TENG,Y. ,K.TANABE,F.TAMURA a nd A.I .1 9 9 8.Ef f ec t TAI o fs pur a ge on t r a ns l o c a t i o n of as s i mi l a t es f r o m s pur so f ‘Ni j i s s e i ki ’Pe a r .J .J a pa nSo c .Ho r t .Sc i .67: 3 1 3 3 1 8 . THOMPSON, C. R. 1 9 6 6 . Pr uni ng appl et r e e s . Fa ber a nd Fa be rLi mi t e d.Lo ndo n. TUSTIN D. S. , F. M. HIRST a nd I . J . WARRINGTON. 1 988. I nf l ue nc e of o r i e nt a t i o n a nd po s i t i o n of f r ui t i ng l a t e r a l so nc a no py l i ght pe ne t r a t i o n, yi e l d,a nd f r ui t qua l i t yo f ‘Gr a nny s mi t h’a ppl e .J .Amer .Soc .Ho r t . Sc i .1 1 3:6 9 3 6 9 9 . TOBUTT,K. R.19 8 5 .Br e e di ng c o l umna ra ppl e s at Eas t Ml l i ng.Ac t aHo t .1 5 9:6 3 6 8 . VOLZ,R. K. ,I . B.FERGUSON,E. W.HEWETT a nd D. J .WOOLLEY. 1 9 9 4 .Wo o da gea nd l e a fa r e ai nf l ue nc ef r ui ts i zeand mi ne r a lc ompo s i t i o no fa ppl ef r ui t .J .Ho r t .Sc i . ,69 (2 ) :3 8 5 3 9 5 . WARRINGTON,I . J . ,D. C.FERREE,J . R.SCHUPP,F. G.DENNIS J r . , a nd T. A.BAUGHER.1 9 9 0 .St r a i na nd r o o t s t o c k ef f ec t s o ns purc ha r a c t e r i s t i c sa nd yi e l do f ‘De l i c i o us ’appl e s t r a i ns .J .Amer .So c .Ho r t .Sc i .1 1 5:3 4 8 3 5 6 . 山本隆儀・伊藤博祐.1 9 9 6.広角レンズを用いた直角 2側面 写真による密植わい性リンゴ樹の樹形・葉群構造の計測. 園学雑.6 4:7 2 9 7 3 9 . 6 6 浅 田 A St udyo fDi f f e r e nt i al l yLoc at edAppl eTr eeSpur s Ta ke no r iASADA La b o r a t o r yo fPo mo l o g y SUMMARY Fr ui t i ngt wi gl et s ,ors pur s ,ar ees s ent i alf ort hepr oduc t i onoft hef r ui t soft heappl et r ee.Dependi ng on t hei rl oc at i on on t he br anc hes ,s pur sc an be c l as s i f i ed i nt ot wo t ypes :t er mi nals pur sand axi l l ar y s pur s .Looki ng att hec ompar at i veper c ent agesoft er mi nalt oaxi l l ar ys pur sa mong t het ot als pur son 1year ol d br anc hesofappl et r eesi n whi c ht henumber sofs pur shaver eac hed as pec i f i cs t age,i tc an be s een t hatt her ei swi de var i et y,wi t hs ome l ar ge br anc heshavi ng a hi gh per c ent age ofaxi l l ar ys pur s and al o w per c ent ageoft er mi nals pur s ,and s omehavi ng al o w per c ent ageofaxi l l ar ys pur sand ahi gh per c ent ageoft er mi nals pur s .It her ef or ei nves t i gat ed t heveget at i vegr o wt h and r epr oduc t i vegr o wt hi n l ar gebr ac heswi t h di f f er i ng t er mi nal /axi l l ar ys purr at i osi n or dert oel uc i dat et heme c hani s msoft hos e t wot ypesofgr owt h.If ur t heri nves t i gat ed t hef or mat i on oft het wot ypesofs pur sand bas ed on t hos e r es ul t s ,t r i edt oes t abl i s hmet hodsf orc ont r ol l i ngt hei rr at i os . Regar di ng t he r el at i ons hi p ofveget at i ve gr owt h wi t hs purt ype,t he c ur r ents t udy c onf i r med t hat veget at i ve gr o wt hi sdec r eas ed when t he per c ent age oft er mi nals pur si shi gher ,whi l ei ti si nc r eas ed when t he per c ent age ofaxi l l ar ys pur si s hi gher .Hi gh number s oft er mi nals pur sf or m mo s t l y on t er mi nalbudsofs hor tbr anc hes ,s how r epeat ed annuals hor t l engt hsofgr o wt h,and event ual l y weaken. Ont heot herhand,axi l l ar ys pur sf or m mos t l yonaxi l l ar ybudsonl onger2year ol dbr anc heswher el ong ne w s hoot s ar el oc at ed at r el at i vel yc l os e pos i t i ons ;t her ef or e,t hey ar e us ual l yl oc at ed i n ar eas of vi gor ousgr owt h.Thes edi f f er enc esi nc o mpos i t i o nandf or mat i oni mpar tt hedi s par at eveget at i vegr o wt h t hatdependsont het er mi nal /axi l l ar ys purr at i os . or epr oduc t i ve gr owt h,t hi ss t udy s ho wed t hat hi gher per c ent ages of t er mi nals pur s ar e As t ac c ompani ed by adec r eas ei nt hef or mat i on off l ower i ng buds ,and al s oc l ar i f i ed t hatar el at i vel y hi gher per c ent age ofapi c albuds l eads t o mor e ef f i c i entpr oduc t i on off r ui t s .The f r ui tpr oduc t i vi t y,i . e.t he numberoff nc r eas edr at eof r ui t sperl eafar ea,i shi gherwhent her ei sal owe rnumberofne ws hoot s (i api c albuds ) ,oral owe rl eafar ea,on 1year ol d br anc hes .Aswi t h an i nc r eas ed r at eoft er mi nals pur s , l owe r ednumber sofs hoot sors mal l erl eafar easar et houghtt ol eadt ohi gherf r ui tpr oduc t i vi t y. I nf r ui tqual i t y,Ic onf i r med t hatas per c ent ages oft er mi nals pur s bec o me hi gher ,and t her e ar e t er mi nals pur st hathaver epeat edl yf or med overt hec our s eoff ouryear sormor e,i ndi c at or sofqual i t y s uc hass i zeands ol ubl es ol i dsc ont ent sbec omel ower ,wher east hec al c i um c ont enti nc r eas es . el at i ons hi p bet ween s purt ype r at i osand veget at i ve and r epr oduc t i ve gr o wt h wass i mi l ari n The r bot h pr uned and unpr uned t r ees ,i ndi c at i ng t hatt hi si san i nnat ec har ac t er i s t i cofappl et r ees .I ti snot t r ue t hatonl yt er mi nals pur s ,oronl y axi l l ar ys pur s ,c ont r i but et ot he pr oduc t i on ofgoodqual i t yf r ui t , r at her ,ac ont i nuous l y mai nt ai ned r at i obet we en t het wot ypesi snec es s ar y.Whatt heopt i mum r angeof r at i osi sdependsonvar i ousf ac t or ss uc hast hepar t i c ul arc ul t i var ,ageoft het r ee,ands oi lc ondi t i ons ,as wel last hequal i t ys t andar dsf ort heappl evar i et yi n ques t i on,and t hes ewi l lbet hes ubj ec toff ur t her s t udy. Lo oki ng ati ndi vi duall ar gebr anc hes ,t het er mi nal /axi l l ar ys purr at i osvar y gr eat l ywi t hi n eac ht r ee, i ndi c at i ng t hati two ul d bepos s i bl et oc ont r olt her at i ost hr ough pr uni ng met hods .It her ef or eanal yzed リンゴ樹における発生部位別スパー構成 67 t he f or mat i on ofpr uned and unpr uned br anc hest oc l ar i f yt he f ac t or si nf l uenc i ng t he devel opmentof t het wot ypesofs pur sand t r i ed t oes t abl i s h pr uni ng met hodst oc ont r olt hes purr at i os .Co mpar i s on of pr uned and unpr uned br anc hes s ugges t ed t hatpr uned br anc hes t end t o devel op a hi gher r at i o of axi l l ar ys pur s .Gener al l ys peaki ng,pr uni ng t o enc our age t he devel opmentoff r ui t i ng s pur s on young br a nc hest hr ough t hepr omot i on oft hedevel opmentofaxi l l ar ys pur si sc o mmo nl y pr ac t i c ed.I nr egar ds t ot hi spr omot i on,t hemos ti mpor t antamongt hec har ac t er i s t i c soft hemot herbr anc hi st hel engt h;i ti s , t her ef or e,nec es s ar yt ogr ow mot herbr anc hesofs omel engt h.Ho wever ,i ft henumberofl ong mo t her br anc hes i si nc r eas ed,t he t r ee bec omes mor e vi gor ous ,and t he t ot alnumber ofs pur si t s el fma y be i ng di f f e l ower ed. Us hods i n a br anc h mo del ,t he t ot al number o fs pur s ,t he r ent pr uni ng met t er mi nal /axi l l ar ys purr at i o,and t he r ec i pr oc alr el at i ons hi ps ho wed t hatt o pr oduc e hi gherper c ent ages ofaxi l l ar ys pur s ,t he per c ent age oft er mi nals pur sdevel opi ng f r o m t er mi nals pur ss houl d be l o wer ed, andt hatapr uni ngmet hodt hatf ac i l i t at esac er t ai nr at i oofdevel opmentofl ongbr anc hesf r o mt er mi nal s pur si sef f ec t i ve.On t heot herhand,t oenc our agegr eat ernumber soft er mi nals pur si n young t r ees ,i t wasmor e ef f ec t i ve t o dec r eas e axi l l ar yl ong br anc hest hatdevel op l at er al l yr at hert han t he br anc hes t hatwi l lbec o mel eader s . es ul t s of t he c ur r ent s t udy make pos s i bl e anal ys i s of c har ac t er i s t i c s of var i ous pr uni ng The r met hodst hr ough t her at i osoft hes pur st hatdevel op on di f f er entpar t soft het r ees ,and s houl d pr ove us ef uli nt hepr oduc t i o nofpr uni ngmanual sf orbegi nner s . l .Fac .Agr i c .& Li f eSc i .Hi r os akiUni v.No.9:28 -67,2006 榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎Bul 6 8 リンゴ果汁輸入の急増が日本のリンゴ経営に与えた影響 − リンゴ過剰恐慌輸出によってもたらされた経営危機 − 宇 野 忠 義 地域資源経営学講座 (2 0 0 6年 1 0月 1 0日受付) は じ め に 近年における中国のリンゴ生産の増大は世界の果実生 産の歴史において驚異的なものであり,その輸出の激増 によって,世界のリンゴ生産とリンゴ経営に大きな影響 を及ぼしている。本論文では,中国のリンゴ生産の激増 と生産過剰の下でリンゴ輸出の急増の背景を探る。 つぎに,日本における果汁輸入の増大がリンゴ価格の 低下などを通じてリンゴ経営に極めて深刻な影響を及ぼ しており,恐慌状態にあることを分析する。 最後に,りんご経営とりんご産業が当面している課題 と今後の展望について,青森県の代表的なりんご地域相 馬村の農協を中核とした取り組みによるリンゴ産業構造 立体化の事例によって検討したい。 1 中国のリンゴ生産の激増とその背景 表 1は,世界の主要国のリンゴ生産量の変化を示した ものである(統計表の原典が明示されていないが,最新 のデータが掲載されているので,概要を把握する意味で 表1 リンゴの主要国別生産量の変化と変化率(1 9 93~2 00 5) 平均生産量(千 ㌧) 国 名 1 9 9 3 -5 2 0 0 3 -5 割合 (%) 変化率 (%) 世界計 4 9 , 2 6 5 6 1 , 8 9 2 1 0 0 . 0 125. 6 米 国 チ リ 中 国 トルコ イラン インド 日 本 韓 国 フランス イタリア ドイツ ポーランド ニュージーランド 4 , 9 5 4 8 3 3 1 1 , 4 1 1 2 , 0 9 2 1 , 8 7 4 1 , 2 2 3 9 8 8 6 4 9 2 , 5 5 7 2 , 1 0 5 1 , 7 7 0 1 , 5 2 4 4 8 2 4 , 3 2 3 1 , 3 0 0 2 3 , 2 6 4 2 , 4 1 7 2 , 4 0 0 1 , 4 7 0 8 2 2 3 6 8 2 , 1 5 9 1 , 9 8 1 1 , 5 9 0 2 , 3 3 3 4 8 8 7 . 0 2 . 1 3 7 . 6 3 . 9 3 . 9 2 . 4 1 . 3 0 . 6 3 . 5 3 . 2 2 . 6 3 . 8 0 . 8 87. 3 156. 0 203. 9 115. 5 128. 1 120. 2 83. 3 56. 6 84. 4 94. 1 89. 8 153. 1 101. 3 注:中央果実基金『海外の果実生産・貿易状況 2 0 0 6年版』 (2 0 0 6 .3)1 0頁より作成。 以下この統計を利用する) 。 2 0 0 3 -5年平均の世界のリンゴ生産量は,6 , 1 8 9万 tで あり,1 0年間に 2 6 %も増加した。地域別にはアジアの 伸びが著しく,ことに中国は 2倍以上の驚異的な伸びを 記録している。中国は,今や,2 , 3 2 6万 tに達し,世界の 3 8 %を占めるに至った。他方では,米国,ヨーロッパ, 日本,韓国などでは生産がかなり減少している。日本 は,9 9万 tから 8 2万 tへと 1 7 %も減少している。ヨー ロッパではポーランド,南米ではチリが中国に次ぐ高い 伸びを示し,注目される。 中国における生産の増大は,短期間のうちに 4つの段 (4 ) 階を経ていることが,図 1に示されている。 すなわち, ①1 9 8 0年代前半の栽培面積の急増と生産の緩やかな増 大期,② 1 9 8 8年から 9 1年にかけての停滞期,③ 9 2年か 図1 中国リンゴ栽培面積と生産量の推移 資料:中国統計年鑑 (1 9 8 1 -1 9 9 1 ) , 中国農業年鑑 (1 9 9 0 -2 0 0 3 ) ら9 6年にかけての栽培面積と生産量の大躍進期,④ 9 7 年以降の栽培面積の減少と生産量の増加・停滞・減少期 特徴的なことは永年木の果実生産であるにもかかわら という具合にめまぐるしく変化してきている。 ず,面積,収量が短期間に急激な増減を記録しているこ 弘大農生報 No.9:68 -79,2006 リンゴ果汁輸入の急増が日本のリンゴ経営に与えた影響 69 表 2 リンゴ園 10アール当たりの粗収入及び純収入(中国) 榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎(単位:元/ 1 0アール) 1999年 2 0 0 0年 2 0 0 1年 地域 粗収入 純収入 粗収入 純収入 粗収入 純収入 山西省臨猗 2, 466 1, 6 4 0 2 , 0 6 7 1 , 3 6 1 1 , 4 3 0 9 6 5 陝西省白水 3, 339 2, 3 4 5 1 , 9 6 7 9 7 1 1 , 6 4 0 6 8 3 山東省栖霞 3, 710 2, 5 7 1 4 , 3 6 4 3 , 0 0 0 4 , 8 9 2 3 , 3 1 1 河北省順平 4, 640 3, 3 3 5 4 , 4 1 9 3 , 1 8 5 3 , 7 0 2 2 , 3 4 2 河南省霊宝 2, 102 8 4 2 1 , 5 8 0 5 6 7 5 8 2 -2 , 9 5 6 平均 3, 251 2, 1 4 6 2 , 8 7 9 1 , 8 1 7 2 , 4 4 9 1 , 4 0 1 注:前掲『中国におけるリンゴの生産・流通事情調査報告書』2 0 ,2 1頁より引用。 表 3 リンゴ農家一人当たりの年収入(中国) 榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎 (単位:元/人) 地域 1999年 2 0 0 0年 2 0 0 1年 平均 山西省臨猗 4, 3 4 0 3 , 6 0 2 2 , 5 5 3 3 , 4 9 8 陝西省白水 4, 2 6 7 1 , 7 6 6 1 , 2 8 6 2 , 4 3 9 山東省栖霞 3, 6 8 6 4 , 3 0 0 4 , 7 4 5 4 , 2 4 4 河北省順平 3, 0 8 6 2 , 9 4 8 2 , 1 6 8 2 , 7 3 4 河南省霊宝 9 0 0 5 5 6 -2 9 7 3 8 6 3, 25 5 . 8 2 , 6 3 4 . 4 2 , 0 9 1 . 0 2 , 6 6 0 . 2 1 0 0 8 1 6 4 平均 1999年比 注:同上 21頁より引用。 とである。第 3段階には面積の増大よりも生産量の増大 均栽培面積は 1戸当たり 4ムー= 2 6 . 7 a~ 5ムー= 3 3 . 3 a が大きく,単位面積あたり収穫量が顕著に増大してきて (2 ) と言われる) 。 いる。これは,剪定,摘花,摘果,袋かけ,摘葉などの 中国農業大学農学院 陳青雲教授による『中国におけ 着色管理といった生産技術がリンゴ生産に広く導入さ るリンゴの生産・流通事情調査報告書』には山西省,陝 れ,良品種がいっそう普及したことの結果である。第 4 西省,山東省,河北省,河南省の五カ所の調査結果が報 段階の 9 8年以降も労働集約的な生産技術の改良が進展 告されている。19 9 9年~ 2 0 0 1年の平均値で 1 0 a当たり し,単位面積あたり収量の増大が目立つ。 投下労働日数が 1 5 0日に及んでいる。青森県の約 5倍に この第 3段階において,生産の増大に国内の需要の増 達する極めて労働多投的な栽培といえる。5 4日~ 2 4 5 加が追いつかず,たちまち過剰生産を引き起こすことに 日という大きな幅で地域差があるが,摘み花,摘果,袋 なった。その結果,リンゴ及び果汁の輸出増大を迫ら かけ,摘葉などの作業労働に多くを費やしているところ れ,低価格生産を梃子として,米国やアジア諸国に輸出 が投入量が多く,逆に,労働投入の少ない地域は労働集 を急激に伸ばしている。 約的技術が導入されておらず,伝統的な技術段階に止 もう一つの特徴は,山東省,陝西省への生産の拡大・ (3 ) まっていると指摘されている。 集中と旧産地遼寧省の相対的な後退といった地域的分化 1 0 a当たりの粗収入は表 2で示したように,5地域平 を伴っていることである。 均で 1 9 9 9年= 3 2 5 1元,2 0 0 0年= 2 8 7 9元,2 0 0 1年= そして最大の特徴と言うべき点は,農業請負生産責任 2 4 4 9元へと 2年間で 2 4 . 7 %も減少している。これは過 制の下で,典型的な零細農家経営によるきわめて労働集 剰基調の下での価格低下が影響している。 約的な生産であることである(零細分散耕地に基づく平 粗収入から農林特産税(10 a当たり 3 3 1元であり,経 宇 野 7 0 表 4 生食用リンゴ(一級着色系「ふじ」 )の流通段階別の価格動向 榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎(単位:元/ kg) 1999年 生産地 2 0 0 1年 2 0 0 3年 生産者 価 格 卸売 価格 小売 価格 生産者 価 格 卸売 価格 小売 価格 生産者 価 格 卸売 価格 小売 価格 山東省煙台 5. 5 6. 5 10. 5 5 . 6 6 . 1 1 1 4 . 9 5 . 5 1 0 河北省高陽 6 6. 5 10. 5 5 . 8 6 . 4 1 1 5 6 1 0 注:中央果実基金 海外果樹農業情報 No . 84『中国におけるリンゴの生産・流通事情調査報告書』 (2 0 0 5 .6)2 9頁より 引用。 営費の 3 1 %を占める) ,農薬,化学肥料,光熱水道費, (陝西省果樹管理局長王振興氏の説明による)。 袋代,技術サービス料などを差し引いた純収入は,19 9 9 このように,今後も成園化が進み,技術向上とともに 年= 2 1 4 6元,2 0 0 0年= 1 8 1 7元,2 0 0 1年= 1 4 0 1元へと 供給過剰が予想されるもとで,価格低下と経営の恐慌的 2年間で 3 4 . 7 %も減少している。所得率は 6 6 . 0 %から 状態が現れている。零細経営の下で増産を図ればますま 5 7 . 2 %へと大きく低下した。これは粗収入の減少の中で す過剰が激化し,悪循環を招くことになろう。そのはけ 農薬,肥料などの購入資材費などの増加によって経営の 口がリンゴと原果汁の急激な輸出の増大となっている。 悪化がより進んだことを意味している。 また,高品質リンゴの生産こそが供給過剰の下で激しい そのため,農家一人当たりの年間純収入は表 3で示し 市場競争を勝ち抜いていく基礎と考えられている。それ たように,5地域平均で 1 9 9 9年が 3 2 5 6元(1元 1 5円と 故,国内の競争力上昇のための集約的技術の導入,高品 して 4 8 ,8 4 0円) ,2 0 0 0年 が 2 6 3 4元(同 3 9 , 5 1 0円), 質生産への努力は,同時に国際的な市場競争力を高める 2 0 0 1年が 2 0 9 1元(同 3 1 , 3 6 5円)へと 2年間で 3 5 . 8 %も ことになり,輸出圧力を強めることにもなる。 大幅に減少している。 ただし,高品質生産のための労働集約的技術の導入の また,販売価格について見ると,表 4のように,生食 ためにさらなる労働投入を図ればいかに低賃金とはい 用リンゴ(一級着色系「ふじ」 )の流通段階別の価格は,山 え,コスト高要因となってくる。その意味では中国にお 東省煙台および河北省高陽における調査では 1 kg当た いては,リンゴ品種の適切な選択,栽培管理技術さらに り生産者価格が 1 9 9 9年には 5 . 5~ 6元,卸売価格が 6 . 5 は貯蔵,輸送技術やそれらのシステムの改善が引き続き 元,小売価格が 1 0 . 6元である。2 0 0 3年では生産者価格 重要な課題となっている。また,経営費増大をいかに低 が4 . 9~ 5元に低下し,卸売価格が 5 . 5~ 6元,小売価格 下させていくか,そのための組織的な取り組みも重要な は1 0元に低下している。低下率は生産者価格が一番大 課題である。 きく,ついで卸売価格,小売価格の順となっており,生 2 0 0 4年から農業税,農林特産税は廃止されたが,きび 産地サイドに影響,しわ寄せがより大きく現れている。 しい産地間競争は続いており,それは輸出圧力ともなっ 小売価格の低下は小幅といえる。 て,海外のリンゴ産業に大きな影響を与えることとな なお,「中国農業情報ネット 2 0 0 4年果実市場形勢分 る。 析」によれば,2 0 0 2年以降「ふじ」系リンゴ価格及び 「ふじ」 (等級なし) 「国光」 の卸売価格は低下傾向にあり 2 中国産リンゴ及び果汁の輸出の激増 の価格は 2 0 0 3年から 2 0 0 4年では,2元~ 3元の範囲で 2 0 0 4年における世界のリンゴ生産量は 6 , 3 2 1万 tであ 推移している。また, 「国光」 (等級なし)の 2 0 0 3年から り,そのうち 1 0 %の 6 4 3万 tが輸出されている。表 5 2 0 0 4年の卸売価格はおおよそ 1~ 2元の間で推移して のごとく,9 0年代まではフランス,米国が最大の輸出国 いる。品質・等級差があるので単純に比較は出来ないが, であったが,輸出量は停滞,減少傾向にあり,逆に, 2 0 0 2年以降も価格が低下傾向にあり,ことに下級品価格 2 0 0 4年には輸出新興国の中国,チリが上位に躍進し,7 0 の下落が見て取れる。 万トン台の輸出量となっている。ヨーロッパではイタリ なお,加工用原料価格は統計がないので不明である ア,ポーランドが輸出を増大しており,ニュージーラン が,一部農家の聞き取りでは 2 0 0 1~ 2 0 0 3年の調査では ドも増加傾向にある。輸出を巡る各国の盛衰,競争の激 (4 ) 1 kg当たり 0 . 3元から 0 . 6元で販売している。 化が伺える。 また,近年の価格については,需要が供給を上回るほ つぎに,リンゴ果汁の輸出について検討したい。表 6 ど評判の良い陝西省における 2 0 0 5年の農家販売価格は, は,濃縮果汁の輸出量の推移を示したが,世界計では 5 0 リンゴ 1 kg当たり 3から 3 . 5元(同 4 5円~ 5 2 . 5円)で 万 tから 8 7万 tへと年率 5 . 5 %の高い増大率が注目され あり,加工用価格はその 1 0分の 1程度と言われている る。ヨーロッパ諸国が主要な地位を占めており,ことに リンゴ果汁輸入の急増が日本のリンゴ経営に与えた影響 71 表 5 主要国のリンゴ輸出量の推移(1 9 9 5~ 2 0 0 4年) 榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎(千 ㌧,%) 国 名 世 界 計 中 国 チ リ フランス イタリア 米 国 ポーランド オランダ ニュージーランド 1995 1 9 9 8 2 0 0 1 2 0 0 4 年成長率 5, 187 5 , 1 1 8 5 , 3 2 6 6 , 4 2 6 2 . 3 0 109 433 768 499 635 139 412 302 1 7 0 5 7 6 7 6 6 5 4 0 5 8 2 1 6 9 3 3 9 2 9 2 3 0 4 6 1 5 7 7 8 5 3 3 7 1 5 2 4 6 2 3 4 2 6 2 7 7 4 7 3 9 6 2 8 5 4 2 4 9 2 4 0 7 3 8 8 3 5 8 2 2 . 1 8 5 . 1 7 -1 . 3 2 3 . 3 1 -1 . 8 8 1 4 . 8 7 -1 . 9 8 1 . 4 4 注:中央果実基金『海外の果実生産・貿易状況 2 0 0 6年版』 (2 0 0 6 .3)1 0 8頁より作成。 表 6 主要国のリンゴ果汁(濃縮)輸出量の推移(1 9 9 5~ 2 0 0 4年) 榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎(千 ㌧,%) 国 名 世 界 計 ポーランド スイス オーストリア チ リ ハンガリー ドイツ アルゼンチン トルコ 1995 1 9 9 8 2 0 0 1 2 0 0 4 年成長率 503 5 5 3 7 0 0 8 7 1 5 . 5 2 85 8. 8 43 31 0 91 67 30 1 0 4 9 . 1 5 8 3 4 0 7 3 5 5 5 1 1 9 2 8 . 3 7 0 6 2 0 7 3 7 9 3 7 2 2 2 8 0 7 5 5 4 5 5 4 8 4 4 4 3 1 1 . 6 0 3 4 . 9 9 5 . 4 9 5 . 5 0 9 . 9 9 -3 . 5 1 1 . 6 6 注:同上 147頁より作成。年成長率は 2 0 0 4年の数値と 1 9 9 5年の数値の対比である。 表 7 主要国のリンゴ果汁(非濃縮)輸出量の推移(1 9 9 5~ 2 0 0 4年) 榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎(千 ㌧,%) 国 名 世 界 計 中 国 ドイツ オーストリア イタリア 1995 1 9 9 8 2 0 0 1 2 0 0 4 年成長率 409 5 7 9 8 0 8 1 , 1 3 9 1 1 . 7 2 18 78 6 29 8 1 8 4 1 2 1 9 2 2 9 1 1 6 1 8 2 5 4 8 7 2 8 3 6 8 5 8 4 8 . 4 9 1 6 . 0 3 3 0 . 2 7 9 . 8 4 注:同上 148頁より作成。 ポーランドが 2 2万 tと際だっている。 している。 ここでは中国は登場しないが,非濃縮果汁の輸出で 1 9 9 5年にはドイツや米国が果汁輸出の主要国であっ は,表 7に示すように,1 9 9 8年以降中国の輸出量が激増 たが,9 0年代末以降新興輸出国に取って代わられた。 し,2 0 0 4年には 4 9万 tに及び,世界の 4 3 %を占めてい 1 9 9 8年には,米国は中国産果汁の米国への低価格輸出に る。図 1で示したように,1 9 9 7年以降中国の栽培面積 対し反ダンピング訴訟を起こし,5 1 . 7 4 %の追加賦課を は減少しているが,結果樹面積は増大しており,他方で 課したが,一時減少後も輸入が増大している。米国では は,中国国内の生食用消費量は,1 9 9 9年に 1 8 6 0万 tと 果汁価格が 1 9 9 5年の 1ガロン(3 . 8ℓ)7 . 6 3 ㌦(約 2 2 1 ピークになり,以後 1 7 0 0万 tから 1 8 0 0万 t前後に減少 円/ℓ)から 9 8年には 3 . 5 7 ㌦(約 1 0 3円/ℓ)へと 5 3 しており,増加した生産過剰分が加工仕向けと輸出に %も低下しており,米国のりんご生産者団体は危機感を 回っている。2 0 0 3年の加工仕向け量は 2 3 0万 t ,輸出量 強め,ダンピング提訴に踏み切り,あるいは日本への輸 は6 1万 tに及ぶ。こうして,9 8年以降果汁輸出が激増 出圧力を強めた (5)。また,これ以降,中国は日本への果 宇 野 7 2 表 8 主要国のリンゴ果汁(濃縮)輸入量の推移(1 9 9 5~ 2 0 0 4年) 榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎(千 ㌧,%) 国 名 1995 世 界 計 米 国 ドイツ 日 本 英 国 オーストリア ロシア 1 9 9 8 2 0 0 1 2 0 0 4 年成長率 444 5 1 6 7 3 2 1 , 0 1 9 9 . 9 1 193 90 * 75 34 29 0 2 2 0 1 2 4 *6 5 3 6 2 3 0 2 6 8 2 0 2 *1 0 0 4 2 4 7 0 3 2 2 1 5 1 9 4 8 6 5 0 4 0 4 . 4 7 5 . 2 6 1 2 . 5 1 1 6 . 7 3 注:同上 147頁より作成。日本の 2 0 0 2.2 0 0 3年の輸入量は 7 5千 ㌧ となっている。 なお,日本の 1995~ 2001年は,誤記入と思われる「非濃縮」果汁輸入量として示されて いる数値を「濃縮」果汁輸入量としてここに表示した。 表 9 リンゴ 1 ㎏ 当たりの産地、消費地市場平均価格の推移並びに果汁用加工原料価格の推移(青森県) 榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎(円) 1990 1991 1992 1993 1 9 9 4 1 9 9 5 1 9 9 6 1 9 9 7 1 9 9 8 1 9 9 9 2 0 0 0 2 0 0 1 2 0 0 2 2003 産地市場価格 消費地市場価格 果汁用加工原料価格 181 354 35 264 463 33 162 295 42 172 264 36 1 7 6 3 1 9 2 2 1 6 0 2 8 8 3 5 1 7 7 2 7 7 4 2 1 4 0 2 2 7 1 9 1 6 1 3 3 7 1 2 1 6 4 2 7 4 1 9 1 7 7 3 2 2 3 2 1 3 1 2 1 8 1 8 1 2 4 2 2 9 9 158 268 24 注:青森県りんご果樹課『りんご流通対策要項』各年産版による。2 0 0 4年度はそれぞれ,1 9 6円,3 1 2円,3 0円に回復した。 汁輸出を増加させることになる。 できた。 しかしながら,リンゴ果汁輸入の激増によるリンゴ産 3 日本のリンゴ及び果汁の輸入状況 業への影響は非常に大きい。1 9 9 0年 4月にリンゴ果汁 1 9 7 1年,日本では,リンゴの輸入が自由化された。し の輸入自由化が決定され,それ以降輸入量が増加を続け かしながら,日本では未発生のコドリン蛾,火傷病など た。表 8及び脚注に示したように,1 9 9 5年には 7万 5 は植物防疫法に規定された輸入禁止品に該当し,発生国 千t (生果換算で約 6 6万 t ,国産果実の 6割を超える量) からは許可されず,当初は,未発生国の韓国からのみ少 に達し,以後も変動を続けながらも増加基調にあり,台 量が輸入された。その後,米国,ニュージーランドから 風被害のあった 2 0 0 4年には 9 4千 tの輸入量となってい 輸入解禁の申し入れが続いたが,日本は禁止してきた。 る。生果・果汁合計の生果換算輸入量が 8 0万 t ,国内加 1 9 9 3年 6月,植物防疫法の改正により,火傷病の発生 工仕向け量の 5倍に達し,リンゴ全体の自給率は 6 0 % 国ニュージーランドからの輸入が 6品種について解禁さ を割り込むところまで低下している。 れた。続いて,9 3年 8月,米国の火傷病など 3病害虫の 果汁の輸入先は,米国産果汁がトップであったが, 防除データの提出を受け,同法施行規則の一部改正によ 1 9 9 8年から中国産果汁が最多となり,リンゴ果汁輸入量 り,1 9 9 4年 8月,米国産リンゴ 2品種の輸入が解禁され の5 0 %前後を中国産が占めるようになった。 た (6)。 世界第 2位の生産・輸出国である米国からは,1 9 9 5年 4 リンゴ果汁輸入の増大がリンゴの価格に与えた影響 に9 4年産リンゴが 8 , 9 3 5 t ,1 4 . 6億円の輸入がなされた。 リンゴ果汁輸入の増大が日本のリンゴの市場価格及び ところが,米国からの輸入量が多かったのはその 1年の 加工用リンゴ価格に及ぼした影響について,日本のリン みであり,以後は,4 0 0~ 1 0 0 t台に激減し,2 0 0 2年以 ゴ生産の 5 0 %以上を占める青森県を対象にして検討し 降は輸入が途絶えている。 たい。 9 5年産以降の米国リンゴ輸入の減少理由は,①食味が 表 9に示すように,リンゴ価格は 1 9 9 2年以降低下傾 劣る割に価格が安くない,②日本で人気のない品種,③ 向にあるが,収量・需給変動を反映して不安定である。 日本で使用禁止の農薬防燻剤 TBZが 9 4年産から検出 1 9 9 7年には,生果の産地市場価格が 1 ㎏ 当たり 1 4 0円 されたことなどである。 へと 2 1 %も低落し,また,果汁用価格は 1 ㎏1 9円へと こうして,1 9 9 5年を除き,日本のリンゴ生果実市場に 5 5 %も激落した。その後,生果の価格は少し回復する おいては,生産者の品種転換と高品質生産並びに販売努 が,果汁用価格は低下したままであり,採算割れ価格で 力や消費者の志向等により,海外産生果実の輸入を防い ある。ことに,2 0 0 2年産に至っては 9円という投げ売 リンゴ果汁輸入の急増が日本のリンゴ経営に与えた影響 73 表 10 青森県リンゴ農家の 1 0 ㌃ 当たり収益性の推移 榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎 (千円) 粗収益 所 得 1990 1995 199 9 2 0 0 0 2 0 0 1 2 0 0 2 2 0 0 3 2 0 0 4 502. 8 317. 1 420 243. 1 347 . 5 157 . 3 3 5 2 1 6 7 . 6 3 4 3 . 3 1 6 1 . 4 3 3 3 . 3 1 4 8 . 6 3 0 8 1 2 5 3 6 1 1 7 2 注:1990,1995年は農水省の生産費調査による。9 6年以降は経営動向調査による。 り同然の大赤字価格となってしまった。 万円に 3 0 %も減少している。またそれ以上に注目され この背景には,リンゴ果汁輸入量の増大がある。リン ることは,所得の減少がいっそう大きいことである。 ゴ加工業は,生食リンゴの下級品に付加価値をつけるこ 1 9 9 0年の 1 0 a当たり 3 1 . 7万円に対し,9 5年には 2 4 . 3 と,並びに生食リンゴの価格安定,需給調整という役割 万円に 2 3 %も減少し,さらに 1 9 9 9年には 1 5 . 7万円へと を担っていたが,その役割を果たせなくなった。19 9 0年 5 0 %も減少している。2 0 0 3年の所得低下はさらにひど のリンゴ果汁の輸入自由化以降安価な海外果汁の流入に く,1 0 a当たり 1 2 . 5万円に激減し,リンゴ経営の困窮状 よって国内産加工リンゴの需要が減り,生産農家は収入 態をもたらした。 維持のためそれまで加工に振り向けていたリンゴを生果 1日(8時間労働)当たり家族労働報酬は,1 9 9 0年に 市場にまわすと,生果市場がダブつき価格低迷へとつな は1 1 ,0 4 7円であり,地域の現業労働平均賃金を 7 0 %ほ がる。 ど上回っていたが,1 9 9 5年には 1日当たり 6 8 1 1円に低 また,円高を利用して,加工業者や大手ボトラーは輸 下し,地域の平均賃金 9 0 0 0円を 2 4 %も下回るように 入原果汁への依存を強め,あるいは,すぐ販売できる製 なった。表示していないが,それ以後も更に低下し, 品の輸入や,海外の加工場で生産し,逆輸入するメー 2 0 0 0年以降 5 0 0 0円台以下となり,平均賃金の半分以下 カーも増えてきており,青森県内の加工場に対する大手 に激減している。そして,2 0 0 3年には 4 2 7 2円という最 ボトラーからの原果汁の注文が減ってきている。 低賃金水準以下に落ち込んでいる。学生のアルバイト賃 青森県りんご果樹課の資料によれば,青森県内の果汁 金をもかなり下回る驚くべき低水準である。2 0 0 4年に 加工用リンゴ販売量は,輸入自由化以前の 5年間の平均 は台風被害による収量減少,価格回復により収益性は改 で年間 1 2万 5 2 0 0 tであったが,自由化以降減少し, 善したが,2 0 0 5年はまた悪化している。 2 0 0 0年からの 5年間では年平均 8万 5 2 0 0 tとなり,1 0 リンゴ経営が比較的に順調であった果汁の自由化以前 (7) 年間で 3 2 %も減少している。 の労働報酬に比較して,その 4 0 %以下という「ワーキン こうして,生果需要自体の減少傾向の下で,供給の増 グ・プアー=働いても,働いても楽にならざる」状態に 加は大幅な価格低下をもたらすことになる。さらにま 追い込まれている。リンゴ史上かってない状態である。 た,加工用リンゴ需要が減少し,供給がダブついて価格 リンゴ生産者は専業的従事者が多いが,その労働の対 が激落している。 価・評価のあまりの低落と価値実現条件の劣悪化は驚異 このようにリンゴ農家はダブルパンチを受けることに 的であり,悲惨とも言うべき状態である。この数値をみ なる。 れば恐慌的な価値破壊とさえ言えよう。 また,流通構造の変化についても指摘しておきたい。 1 0 a当たり生産費では,農薬,光熱動力費,建物及び 需要の停滞傾向の下で,1 9 9 9年の卸売市場法の改正に 土地改良設備費,諸材料費が増加している。粗収益の低 より公認された「先取り相対取引」により,量販店主導 下,減少にも関わらず,これらの生産費の増大が持続し の値決めに移行し,市場の建値による価格形成が機能低 ているところに経営収益に関わる構造的問題がある。昭 下していること,また,量販店の競争激化で,低価格競 和初期の恐慌の際のシェーレ現象を思い起こさせるもの (8 ) 争に拍車がかかっていることも注目すべき点である。 (9 ) である。 5 青森県のリンゴ農家の経営の悪化による恐慌的状態 生産費調査及び経営動向調査によって,青森県のリン つぎに,青森県のリンゴ農家の経営の悪化について検 ゴ経営について,仮に企業的会計計算をすれば,1 9 9 0年 討し,今後の対策について触れたい。 では,10 a当たり 1 0万円の利潤があったが,1 9 9 5年に 表 1 0は,農水省の生産費調査及び経営動向調査に は1 0 a当たり 8万円の赤字となり,2 0 0 0年以降では, よって青森県のリンゴ園 1 0 a当たりの収益性の推移を 1 5~ 1 6万円の赤字へとさらに悪化している(1 9 9 5年以 示しているが,粗収益は収量・価格の変動・低下を反映 降は公表数値がないので,労賃水準及び家族労働の評価 して,1 9 9 0年以降,ことに 9 7年以降減少が著しい。 については著者の試算である) 。企業経営であれば倒産 1 9 9 0年の 5 0 . 3万円から 9 5年の 4 2 . 0万円,9 9年の 3 4 . 8 状態が続いている。もちろん,家族経営でも経営のまと 宇 野 7 4 表 11 青森県りんご経営農家の経営成果(2 0 0 0~ 2 0 0 4年 平成 1 2~ 1 6年) 榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎(kg;円) 項 目 作付面積 榎榎榎榎榎榎榎榎榎榎榎榎榎(a) 収入 16年平均 10 a当たり 15年平均 10 a当たり 14年平均 10 a当たり 13年平均 10 a当たり 12年平均 10 a当たり 16年の 前年対比 16年の 12年対比 179 178 173 172 169 101 106 2, 101 2, 493 2, 427 2, 332 1, 884 84 112 2 経営仕向量 0 26 0 0 0 0 3 家計仕向量 18 21 21 26 30 88 60 216 164 154 166 206 132 105 458, 000 411, 305 376, 014 391, 428 394, 284 111 116 109 137 776 0 8, 253 79 1 458, 109 411, 442 376, 790 391, 428 402, 537 111 114 1 販売量 4 販売単価 5 販売・家計金額((1+ 3)× 4) 6 副産物等販売額 7 粗収益合計(5+ 6 ) 費用 種苗費 2, 593 1, 724 568 1, 269 1, 768 150 147 肥料費 農薬費 15, 226 41, 136 14, 731 39, 558 13, 986 41, 402 15, 095 36, 446 16, 033 36, 683 103 104 95 112 諸材料費 27, 396 32, 811 36, 134 33, 305 22, 766 83 120 光熱水費 13, 003 10, 739 10, 076 11, 437 10, 802 121 120 租税公課 5, 401 4, 918 5, 424 5, 928 4, 529 110 119 14, 547 13, 156 11, 460 12, 159 8, 758 111 166 小農具費 3, 031 2, 451 4, 364 3, 709 5, 210 124 58 土地改良水利費 1, 355 602 5, 691 1, 294 1, 315 225 103 農業共済掛金 8, 701 7, 983 9, 006 8, 993 10, 034 109 87 賃料料金 5, 240 5, 857 8, 489 14, 411 9, 644 89 54 40, 200 41, 672 32, 748 26, 358 23, 823 96 169 修繕費 雇用労働費 借地料 支払利子 農用被服費 その他 建物施設却費 機械償却費 動物・植物償却費 0 70 63 30 57 0 0 2, 005 1, 737 2, 034 4, 380 1, 430 115 140 3, 132 2, 835 2, 338 2, 119 3, 164 110 99 15, 507 18, 866 18, 536 25, 972 13, 892 82 112 7, 119 7, 822 11, 022 7, 596 7, 480 91 95 21, 107 23, 774 21, 882 17, 771 18, 702 89 113 1, 112 1, 121 1, 076 802 697 99 160 61, 594 69, 558 78, 397 69, 285 79, 193 89 78 289, 405 301, 986 314, 696 298, 359 275, 980 96 105 231, 488 237, 608 251, 467 271, 002 264, 343 97 88 費用合計 10a当たり家族労働時間 520, 892 185 539, 594 190 566, 163 201 569, 361 215 540, 323 225 97 97 96 82 10a当たり農業所得 168, 704 109, 456 62, 094 93, 069 126, 557 154 133 出荷経費 経営費合計 家族労賃見積額 分析指標 所得率榎榎榎榎榎榎榎榎榎 (%) 農業純収益 家族 8時間当たり所得 37 27 16 24 31 138 117 - 62, 784 7, 288 - 128, 152 4, 607 - 189, 373 2, 469 - 177, 933 3, 463 - 137, 786 4, 500 49 158 46 162 注:1 青森県農林総合研究センター資料による。 2 小規模 4経営,中規模 5経営,大規模 5経営(計 1 4経営)の調査結果である。 3 担当した普及センターは五所川原,弘前,黒石,鰺ヶ沢,三戸である。 4 家族労働費は 1時間当たり 1, 250円で計算した。 5 生産コスト=経営費+家族労働費である。 もな再生産ができる状態ではない。異常なまさに危機的 分析をしておこう。表 1 1は調査農家の 5年間の経営成 な恐慌状態といえよう。 果の 1戸平均値の推移を示している。 その要因について,青森県の資料を利用して経営内部 暦年で集計しているので,当該年産のリンゴの販売が からもう少し分析しておきたい。 翌年にまたがり,また,農協出荷などではプール計算に 2 0 0 0年以降,青森県が県内のリンゴ地域の記帳農家か より翌年に精算されるので,それらの集計事情には注意 ら抽出された 1 4戸のリンゴ農家を継続して調査してい を要する。 る。その結果によって,2 0 0 4年度までの 5年間の経営 調査したリンゴ農家の 1戸平均では 1 . 7 haの作付け規 リンゴ果汁輸入の急増が日本のリンゴ経営に与えた影響 75 模である。総販売単価が 2 0 0 0年の 1 kg 2 0 6円から 2 0 0 2 村(2 0 0 6年 2月に合併して現在は弘前市に含まれる)の 年の 1 5 4円,2 0 0 3年の 1 6 4円へと暴落したにもかかわ 事例によって検討しておきたい。(11) らず,経営費は逆に増大している。表に見られるごと 旧相馬村は津軽平野の南方,弘前市の西隣,秋田県境 く,農薬費,諸材料費,雇用労働費,機械償却費の増大 の山岳地帯に接し,三方を国有林に囲まれた農山村地帯 が目立っている。また,出荷経費が極めて大きいことが である。最初に,2 0 0 0年センサスにより相馬村の農業 経営悪化に拍車を掛けている。その結果 1 0 a当たり所 概況に触れておきたい。農家戸数 5 8 2戸,耕地面積 1 0 4 0 得は 2 0 0 0年の 1 2 . 7万円から 2 0 0 2年の 6 . 3万円へと半減 ha (1戸あたり 1 7 9 a ),うち 8 6 %が樹園地で 8 9 5 ha ,1 し,所得率も 3 1 %から 1 7 %というかって経験したこと 戸あたり 1 6 5 aで県下で 1位の樹園地規模である。水田 のない低さに低落している。家族労働 8時間当たり所得 は1 3 2 ha (所有農家 2 8 9戸,1戸あたり 4 6 a )にすぎな は2 0 0 0年の 4 5 0 0円という最低賃金水準からさらに低下 い。 し,2 5 0 0円(2 0 0 2年)という恐るべき低水準に到ってい 所有耕地面積の規模別分布をみると,1 ha未満が 1 6 3 る。 戸,2 8 %,1~ 2 haが 1 9 5戸,3 4 %,2~ 2 . 5 haが 8 0 なお,2 0 0 4年には,収穫期に 2度の台風襲来があり, 戸,1 3 . 7 %,2 . 5~ 3 haが 6 2戸,1 0 . 7 %,3~ 4 haが 収穫量が 1 0 a当たり 2 1 0 1㎏ と前年比で 1 6 %も減少し, 5 4戸,9 . 3 %,4~ 5 ha が 1 5戸,2 . 6 %,5 ha以上が 他産地の収量減と競合青果物の収量減,品薄からリンゴ 1 1戸,1 . 9 %を占めている。2 . 5 ha以上層がやや増加傾 価格が 1㎏ 当たり 2 1 6円に高騰し,数年前の水準に回復 向にある。 した。その結果,収量の減少よりも価格の上昇がより大 農業労働力についてみると,2 0 0 0年の販売農家 5 4 7 きく,粗収益が増大した。また,収量の減少により,出 戸のうち専従者がいる農家は 4 9 5戸,9 0 %に達する。 荷経費や諸材料費が減少したことも経営収益にプラスに そのうち男女の専従者がいる農家は 3 8 4戸,7 7 %に及 作用した。 ぶ。基幹的農業従事者の平均年齢は 5 6 . 4歳であり,県平 2 0 0 4年には,経営費合計に対する出荷経費の比率は 均の 5 9歳より 2歳ほど若い。農業就業人口 1 3 6 5人(1 2 1 . 3 %にまで低下した。2 0 0 0年には 2 9 %と言う高い出 戸あたり 2 . 5 0人)を年齢別にみると,男子 6 4 2人の中, 荷経費比率であったが,減収が流通経費の減少に好影響 2 0歳 代 2 1人,3 0歳 代 7 1人,4 0歳 代 9 7人,6 0歳 代 をもたらしている。それでも支出項目の中で最大の額を 1 9 6人,7 0歳 以 上 1 2 2人 と な っ て お り,6 5歳 以 上 が 占めている。出荷経費の削減対策の重要性が指摘でき 5 0 %を占めている。 る。農協や生産者団体を挙げて取り組むべき課題の一つ 耕地規模と労働力を対比すれば,耕地 1 . 5 ha以上の農 である。 家全戸が,また,リンゴ栽培農家 1 ha以上の 3 7 0戸全部 つぎに,家族労働 8時間当たり所得についてみると, に男女の専従者がいることになる。リンゴ園は傾斜地, 前述の生産費調査結果よりも更に低くなっている。20 0 0 山間地がほとんどであり,収穫は手作業であり,一人当 年の 4 5 0 0円という最低賃金水準からさらに低下し, たり千箱が限度である。それ故,リンゴ 2 ha以上の栽 2 0 0 2年には 2 5 0 0円という恐るべき低水準に到り,もっ 培農家においては通常 2世代,4人の労働力を必要とし とも危機的な状態であった。2 0 0 3年には 4 6 0 0円と最低 ている。 賃金水準に戻り,台風被害のあった 2 0 0 4年には 7 2 8 8円 相馬村では,2 0 0 0年当時,リンゴ経営については,4 0 へと数年ぶりに 7千円台に復帰した。それでも地域平均 歳代以上の労働力は確保されているが,3 9歳未満では 賃金水準には及ばない。所得率も 3 7 %に回復した。 手薄になってきている。これは,1 9 8 0年代前半までは しかしながら,翌年の 2 0 0 5年には平年作水準を回復 リンゴ経営が現在ほど厳しくなく,若い後継者が確保で し,価格が前年比で 2 0 %以上も低落し,再度経営が劣悪 きていたが,その後は次第に困難となってきており,品 化している。 「豊作貧乏」から更に「平年作貧乏」とも言 質を維持,高めつつ省力栽培を採用している。無袋栽 うべき状態に追い込まれている。荒廃園の出現や協同防 培,葉採らずリンゴ,わい化栽培,低農薬栽培などに集 除の維持存続が現実の問題となり,また,果樹の伐採が 団で取り組み,また,農協が仲介して農協間の労働力交 進行している。リンゴの再生産基盤は果樹園・樹体にお 換やアップルヘルパー制度を導入している。さらに,村 いても,労働力においても確実に掘り崩されてきてお レベルで青年農業者の会が組織されており,リンゴ農家 り,農家の懸命の努力も限界に来つつある。耕作放棄が の自主的組織であるりんご協会主催の研修会などに積極 出ても不思議ではないが,リンゴ農家はじっと耐えてい 的に参加し,高い技術水準を維持し,その継承を図って る (1 0 ) 。 いる。その結果,相馬村では優良銘柄の確立と多くの受 賞を記録し,篤農家を輩出してきた。 6 今後の課題と展望 −相馬村における農業を核とし た産業構造立体化の取り組み− しかしながら,長期不況と農産物需要の頭打ち,後退, 他方での農産物輸入の増大と農産物価格の低落といった 最後に,今後のリンゴ経営及びリンゴ産業の課題と展 経済状況の下で,農業予算も削減され,農業所得の減少 望について,津軽の代表的なリンゴ生産地域である相馬 は顕著である。その克服のためには,農業を核として, 宇 野 7 6 農産物加工や農業の多面的機能を発揮させるような二次 目で分かるというシステムである。これについては, 産業,三次産業への展開とそれらを包摂した産業構造の 「トレーサビリティを確立して安全を確保し,消費者に (1 2 ) 立体化が必要となっている。 信頼してもらうというのは今では当たり前の話であり, 相馬村における産業構造立体化の取り組みは,農協を 信頼を裏切る産地は成り立ちません。これからは,更に 中核としたリンゴ,米などの生産,加工,販売の指導体 信頼を超えた感動を実感してもらうことを考えていま 制と高い系統利用率による立体化が特徴的である。 す。」と J A相馬村事業推進部長兼販売本部長の田沢俊則 農協は非合併を決断し, 「小さな J A,大きな貢献」を 氏は語っている。(13) 目標にし, 「みんなが集まる J A」を基本理念にして,地 こうして,J A相馬村の 2 0 0 4年度の事業総利益は 6億 域と一体となった取り組みを特徴としている。J Aの米 1 6 9 8万円,事業管理費は 4億 9 9 3 0万円,事業収益は 1 取り扱い率 1 0 0 %,リンゴの取扱量 9 3 %,その他主要事 億1 7 6 9万円を実現している。これに事業外収益を加え, 業ともおおむね 8 0 %を超える高い利用率を実現してい 法人税など諸課税を差し引いた当期剰余金は 1億 2 2 0 0 る。 万円の実績を残している。農協は経営の健全性を図る上 そして,産業構造の立体化としては,農協がリンゴ で内部留保を増強し,正組合員一人当たり 1 0 6万円の出 ジュースの加工など 2次産業面に取り組んでおり,さら 資となっており,自己資本比率は実に 3 8 . 8 %に達する。 に農協運営による農産物直売所を開設している。その他 県内トップクラスを堅持している。全国的に見ても非常 に CA貯蔵庫,冷蔵庫,選果場,農産物加工施設,リン に優れた実績を上げている。 ゴジュース加工場,育苗施設,ライスセンター,石油ス 前述したように,価格低下などを反映した経営の悪化 タンド,機械化センター,事務所などを揃え,2 0 0 5年度 が進行しているので,かかる農協の経営余剰をも利活用 末では,正職員 4 4名,常傭的臨時雇用者 2 7名で運営さ した特別の経営安定対策などの取り組みが更に必要であ れている。農家家族や地元民の雇用の機会を提供してい る。 る。また,何よりも,特徴を持った農産物の生産と付加 相馬村における農業を核とした産業構造の立体化はま 価値を付けた有利な販売に工夫を重ねて取り組んでお だ展開途上にあるが,多くの困難な重い課題を抱えてい り,その結果として高い系統利用率と健全な収益性を実 る。中でも重要なことは,リンゴ農家の経営安定対策で 現している。 ある。1 9 9 7年のリンゴ価格の激落を契機に青森県の単 他地域同様に,農産物の販売額はこの 2 0年近く,停 独事業で 9 9年に発足した「青森県生食用りんご価格安定 滞,減少してきているが,リンゴジュースの加工を拡大 制度」は,20 0 1年からは国も加わった「果樹経営安定対 し,ほぼ順調な軌道に乗っている。これは,品質の良さ, 策事業」として継続している。緊急不可欠のものであり, 積極的販路の拡大,農協内での生産者・加工部門・販売 不十分ながらも価格の暴落に対して一定の補填がなさ 所の三者にそれぞれ収益があがるような価格調整・作業 れ,激減している所得の一部が補償されている。20 0 1 , 調整,バランスのとれた管理運営を農協担当指導者が計 0 2年産の価格暴落時に合計 6 4億円の支払いがあり,損 画・勘案し,経験を積み上げてきた成果である。 失の一部補填がなされ,ひと息つくことが出来た。しか また,直売所も売り上げを伸ばし,1 9 9 7~ 9 9年度に し,金額・量などでまだ不十分であり,また,何よりも は 3億円を超え,3億 4 4 0 0万円に達し,組合員の農産 加工用リンゴが全く対象とされておらず,底に穴が空い 物・加工品の売り上げも 9 0 0 0万円に近づいた。2 0 0 0年 ているバケツのようなものである。それは,前述したよ 度以降は,リンゴの宅配とリンゴジュースの特産販売を うに,生果の価格下落に強く連動しているからである。 別事業に移行したため, 「りんごの森」売上高は,形式的 しかも,その措置も 2 0 0 5年度より廃止されることにな には後退したように見えるが,2 0 0 0年度の 1 . 8 7億円か り,日本のリンゴ生産の将来は不安と暗雲が立ちこめて 4年度には 2億 5 4 2 3万円の販売額へと順調に増大 ら2 0 0 (1 4 ) いる。 している。9 4年の発足当初は余った農産物を売るとい リンゴ農家の後継者の確保,農家労働力の再生産を保 う意識であったが,現在では「売れる農産物,買ってく 証するものとして経営安定対策の継続・拡充は緊急不可 れる商品づくり」を目指している。いわば,需要に対応 欠のことである。また,付加価値生産の拡大を図る産業 したまた需要創造型の生産・販売を目指しているといえ 構造の立体化を実現していく上でも,密接な関係を持つ る。 リンゴ果汁の生産・流通に対する価格保証を含めた行政 そのためにも,全農家が「農作業記録簿」に記帳して 施策も緊急不可欠となっている。銘記すべきことであ おり,トレーサビリティを確立し,また薬剤散布回数の る。 削減や減農薬なども含めて「J A相馬村農産物信頼シス テム」を独自に構築していることが特筆すべきことであ る。これは,2 0 0 6年 1月からりんごでは全国初めてと 要 約 なる 2次元バーコード(QRコード)を導入しており, 本論文では,中国のリンゴ生産の激増と果汁輸出の急 携帯電話で 2次元バーコードを読み取ると生産履歴が一 増の背景を探った。農業請負生産責任制の下で,極めて リンゴ果汁輸入の急増が日本のリンゴ経営に与えた影響 零細な経営による労働集約的栽培により供給過剰・価格 低下となり,リンゴと果汁の輸出の激増となっている。 また,日本では,輸入自由化後に果汁輸入が増大し,日 本のリンゴ経営に極めて深刻な打撃を与えており,いま や恐慌状態にあることを分析した。最後に,青森県相馬 村のリンゴ産業構造立体化による懸命な取り組みを紹介 した。 2 0 0 3 -5年平均の世界のリンゴ生産量は 6 , 1 8 9万 tで あるが,中国は 1 0年間に 2倍以上の驚異的な伸びを記 録し,世界の 3 8 %を占める。2 0 0 4年には中国がリンゴ の最大輸出国となり,また,非濃縮果汁の輸出では世界 の4 3 %を占めている。 日本では,1 9 9 0年にリンゴ果汁の輸入自由化が決定 され,2 0 0 4年には 9万 4千 tの濃縮果汁輸入量となっ た。生果と果汁合計の生果換算輸入量が 8 0万 tに達し た。果汁の輸入先は,5 0 %前後を中国産が占めている。 日本のリンゴ生産の半分を占める青森県では,リンゴ 生産の約 2 0 %が低価格で加工用に回されている。その 加工用リンゴ価格が,1 0年間で 6 0 %以上も低下し,リ ンゴ農家は赤字経営となった。加工用リンゴの低価格が 生果市場にも影響し,リンゴ価格も低下した。リンゴ園 1 0㌃ 当たりの所得は,1 9 9 0年の 3 1 . 7万円に対し,1 9 9 9 . 7万円へと 5 0 %も減少し,リンゴ経営の困窮 年には 1 5 状態をもたらした。 注 (1)中央果実基金・海外果樹農業情報 No .84『中国におけ るリンゴの生産・流通事情調査報告書』 (2005年 6月) 13~ 14頁より引用。原著者は,中国農業大学農学院, 陳青雲教授である。なお,1 ha当たり収量を 199 6年と 2005年について主要国で示せば,米国 24. 9 t ,26. 6 t 。中 国 5. 7 t ,11. 4 t 。日本 19. 1 t ,21. 2 t 。韓国 14. 9 t ,1 4 . 1 t 。 フランス 31. 4 t ,36. 6 t 。ポーランド 12. 0 t ,12. 3 t 。イタ リア 29. 0 t ,35. 6 tである(中央果実基金・海外果樹農業 情報 No . 87 『海外の果実生産・貿易状況 2006年版』に よる) 。中国では 9年間で単収が倍増しており,驚異的 な伸長である。これには日本のリンゴ生産技術の普及も 反映している。品種も着色系「ふじ」が 2002年で 5 7 %, 国光が 7 %に達している。 (2 )前掲『中国におけるリンゴの生産・流通事情調査報告書』 , 黄孝春「中国におけるりんごの生産・流通に関する調査 報告」(りんご振興研究会『国際化・自由化段階における 青森県りんご産業の活性化に関する研究』(20 0 3年 6 月),中央果実基金『中央果実基金通信・海外果樹農業 ニュース No .79』頁 53~ 55等を参照。 『中国におけるリンゴの生産・流通事情調査報告書』によ れば,中国国内の生産と流通における問題点として,分 散した小規模少量生産であり,かつ流通上の諸問題が指 摘されている。①生産量は増加し続けたが,高品質の果 実が少ない。②技術が標準化されておらず,品質が悪 く,ばらつきが大きい。③果実の流通機構,業者,組合, 流通ルートなどが未整備であり,形成されてなく,流通 が順調ではない。④果実の包装,貯蔵に大きな問題を抱 えており,流通過程に乗らないことがある。⑤果実加工 業の処理能力,技術及び流通上の問題を抱えており,適 時に加工処理されない。 77 ただし,気象条件などにおいては大規模なリンゴ生産適 地を擁しており,また,交易上の地理的条件も有利性を 備えており,低価格を武器にして輸出の増加の可能性は 高いとみられる。 (3)前掲『中国におけるリンゴの生産・流通事情調査報告書』 頁1 9による。 (4)同上頁 2 8~ 2 9による (5 )農民運動全国連合会機関紙『農民』2 0 0 5年 8月 1/ 8/ 1 5日号,インドのジャーナリスト,デビンダ・シャルマ 氏の記事と中央果実基金『中央果実基金通信・海外果樹 農業ニュース No .7 9』5 3~ 6 0頁参照。また,宇野忠義 「最重要病害リンゴ火傷病の日米検疫問題」 ( 『弘前大学 ,2 0 0 5年 1 2月)頁 89 農学生命科学部学術報告第 8号』 ~9 1も参照。 (6)リンゴの最重要病害である火傷病の日米検疫問題につい ては,宇野忠義「リレーエッセイ 時代を読む・重大病 害リンゴ火傷病に万全の対策を望む」(『現代農業』2005 年 7月号) ,宇野「最重要病害リンゴ火傷病の日米検疫問 題」 (『弘 前 大 学 農 学 生 命 科 学 部 学 術 報 告 第 8号』, 2 0 0 5年 1 2月) ,宇野「最重要病害リンゴ火傷病の日米検 疫問題の交渉経過と残された問題」 (農業・農協問題研究 所『農業・農協問題研究』第 3 5号,2 0 0 6年 8月)を参 照されたい。 (7)青森県『平成 1 7年産りんご流通対策要項』(2 006年, 1 0 2頁)による。 (8)古いデータであるが,中央果実生産安定基金協会『果実 の市場外流通実態調査報告書』 (1 9 9 6年 3月)によれば, 9 5年 産 の 収 穫 量 9 6万 3 3 0 0 t ,生 食 向 け 出 荷 量 72万 0 8 6 0 tである。そのうち,卸売市場仕向け 5 4万 6800 t (7 5 . 9 %) ,市場外仕向け 1 7万 4 0 6 0 t (2 4 . 1 %)となって いる。市場外仕向けのうち主要な経路は,生協・消費者 グループ= 3 1 %,直売所・観光農園= 1 7 %,宅急便・ ゆうパック= 1 7 %,量販店・小売店= 1 4 %である。当 時は,市場外流通の中では量販店の比重が小さかった が,その後比重を高めており,また,市場流通を介した 中での量販店・スーパーマーケットの販売比重が高まっ ている。 (9 『平成 1 ) 6年度食料・農業・農村白書』 (原資料は「農業物 価統計」 )によれば,2 0 0 0年を基準として農産物及び,農 業生産資材並びに農業交易条件指数が付表 1のように示 されている。 それによれば,2 0 0 0年に対する 2 0 0 4年の農産物総合指 数は 1 0 6 . 3,内訳では,米 1 0 4 . 2,野菜 1 1 5 . 6,果実 98. 3と なっている。また,生産資材総合の物価指数では,2004 年1 0 1 . 6であり,4年間の資材価格はほとんど上昇して おらず,この限りでは交易条件の悪化はみられない。し かしながら,WTOの発足した 1 9 9 5年を物価動向の基 準年にすると,様相は激変する。1 9 9 5年から 2 000年に かけて農産物総合では物価指数が 1 4 . 4ポイント低下し ており,そのうち米の低下が 2 6 . 7でもっとも激しく。次 いで果実が 2 2 . 1ポイントも下落している。逆に,1995 年の農業生産資材総合の物価指数は,2 0 0 0年よりも 1. 9 ポ イ ン ト 低 い も の で あ っ た。す な わ ち,1 9 9 5年 か ら 2 0 0 0年にかけて生産資材価格は 1 . 9ポイント上昇し,逆 に,農産物総合物価は 1 4 . 4ポイントも下落しており,明 白なシェーレ現象がみられる。この短期間に農業の交易 条件は 1 6 . 6ポイントも悪化している。この農業生産物 に極めて不利な劣悪な交易条件がその後も基本的に改善 されていないところに構造的な問題を指摘できる。 (『農 業と経済』 (2 0 0 5年 9月別冊号,1 0 7頁参照) (1 0)ここで,調査データが得られた 20 0 1年の中南津軽リン ゴ専業農家 6戸の農業所得について触れておこう。いず 宇 野 7 8 付表 1 農業物価指数と農業交易条件指数の動向(2 0 0 0年= 1 0 0) 指 数 農産物総合 うち 米 野菜 果実 畜産物総合 農業生産資材総合 うち 種苗及び苗木 畜産用動物 肥料 飼料 農業薬剤 諸材料 光熱動力 農機具 建築資材 農業の交易条件指数 1995年 2 0 0 0年 2 0 0 2年 2 0 0 4年 (概算) 114. 4 1 0 0 . 0 9 7 . 0 1 0 6 . 0 126. 7 116. 4 122. 1 101. 1 1 0 0 . 0 1 0 0 . 0 1 0 0 . 0 1 0 0 . 0 9 8 . 2 1 0 1 . 2 8 4 . 2 9 8 . 6 1 0 4 . 2 1 1 5 . 6 9 8 . 3 1 0 4 . 9 98. 1 1 0 0 . 0 9 9 . 5 1 0 1 . 6 95. 2 92. 9 95. 3 98. 5 102. 6 99. 7 101. 9 97. 0 103. 2 1 0 0 . 0 1 0 0 . 0 1 0 0 . 0 1 0 0 . 0 1 0 0 . 0 1 0 0 . 0 1 0 0 . 0 1 0 0 . 0 1 0 0 . 0 1 0 0 . 7 9 2 . 4 1 0 0 . 4 1 0 6 . 3 9 8 . 3 9 7 . 8 9 7 . 7 9 8 . 3 9 7 . 3 1 0 1 . 2 1 1 1 . 7 1 0 0 . 8 1 1 5 . 2 9 6 . 8 9 6 . 8 1 0 3 . 3 9 7 . 8 9 5 . 5 116. 6 1 0 0 . 0 9 7 . 5 1 0 4 . 3 資料:農林水産省「農業物価統計」 注:1)品目については,主要なもののみ表示した。 2)農業交易条件指数は,農業生産資材価格指数(総合)に対する農産物価格指数(総合)の 比率である。 3)「平成 16年度 食料・農業・農村の動向」より引用。 れも市町村の上位に位置する先導的経営といえるが,経 営耕地,リンゴ成園面積,農業所得,耕地 10 a当たり所 得について列記すると,A:416 a,322 a,446万円, 10. 9万 円。B:515 a,513 a,327万 円,6. 3万 円。C: 329 a,296 a,645万 円,1 9. 6万 円。D:386 a,3 8 6 a, 442万円,11. 5万円。E:362 a,150 a,281万円,7 . 8万 円。F:300 a ,250 a,321万 円,10. 7万 円 と な っ て い る。 いずれも家族労働力 3~ 4人が就業しており,成果のや や良い Cを除いて,他の 5戸は相当厳しい経営内容と なっている。 稲作とリンゴは輸入の理由と形態,方法は異なるが,年 々数十万トンの輸入が十年以上に亘ることによって,国 内生産者の息の根が止められようとしている。 (1 1)相馬村の農業については,宇野忠義「全村ぐるみの稲作 生産組織化とりんご経営-青森県相馬村-」 (田代洋一 編『21世紀の農業農村・第 3巻・日本農業の主体形成』 筑波書房,2004年,150 -164頁)を参照のこと。全村統 一した稲作生産組織の形成とその成果を分析し,併せて 農業経営状態も分析した。 (1 2)津軽地域の産業構造の立体化については,笹森 正著 『りんご加工業発達史-青森県を中心に』(北の街社発 行,1998年)において,青森県産業の構造変革として, 農業・りんごを起点とした地域産業の立体化の必要性を 提起した上でりんご加工業の分析をしており,重要な文 献である。なお,それに関連して私たちが調査した報告 書に,弘前大学農学部農業生産流通学講座『転機に立つ りんご果汁-津軽地方におけるりんご産業の行方-』 (1 9 9 9年)がある。また,産業構造の立体化を視点として 青森県名川町,相馬村を対象として分析した調査報告書 (渋谷長生編)として,青森県農林部『農業を核とした産 業構造立体化の検証』 (2 0 0 0年)が参考となる。 (1 3) 『青 森 ト ゥ モ ロ ウ No .6 3』(財 団 法 人 日 本 立 地 セ ン ター,2 0 0 6年 2月)1~ 2頁を参照。 (1 4)価格・所得政策の課題については,田代洋一著『新版 農業問題入門』 (大月書店,2 0 0 3年)1 9 0~ 1 9 6ページを 参照。なお,家族経営による日本の農業と農山村の維 持・再生の課題については,同上『新版 農業問題入門』 及び石井啓雄「農業・農山村の再生と家族経営(上), (下)」 (新日本出版社『経済』NO.9 7 .NO.9 8,2 0 0 3年 10月号, 1 1月号)を参照。その他,市場問題及び価格政策につい ては,日本農業市場学会編『講座 今日の食料・農業市 場 全 5巻』 (筑波書房) ,ことに村田武・三島徳三編『農 政転換と価格・所得政策』 (2 0 0 0年) ,滝澤昭義・細川允 史編『流通再編と食料・農産物市場』 ,三國英實・来間泰 男編『日本農業の再編と市場問題』 (2 0 0 1年) ,三島徳三 『規制緩和と農業・食料市場』 (日本経済評論社,2001 年)を参照。 リンゴ果汁輸入の急増が日本のリンゴ経営に与えた影響 79 TheRa pi dI nc r e a s ei nI mpo r tofAppl eJ ui c eHa sSt r uc kaHea vyBl o w a tJ a pa nes eAppl eFar mi ng Ta da yo s hiUNO La b o r a t o r yo fRe g i o n a lRe s o u r c eMa n a g e me n t Thi ss t udyanal yzedt her eas onsf ort her api di nc r eas esi nChi nes eappl epr oduc t i onandappl ej ui c e expor t s ,and ho wt her api di nc r eas eofChi nes eappl ej ui c eexpor t si nt oJ apan hass t r uc k aheavy bl o w att heJ apanes eappl ei ndus t r y. Wor l d pr oduc t i on ofappl esr os et o 61, 890, 000 t onson t he aver age bet ween 2003 and 2005.Chi na ’ s s har e ofwor l d pr oduc t i on r oc ket ed t o 38 %.Chi na i st he wor l d’ sl ar ges texpor t er ofappl es and t he s har eoft heappl ej ui c eexpor tmar kethasr i s ent o43% i n2004. n 1990,t he J apanes e gover nments anc t i oned unl i mi t ed i mpor t s ofappl ej ui c e.I n 2004,J apanes e I i mpor t sofc ondens ed appl ej ui c eamount ed t o94, 000t ons ,and i mpor t sofappl esand appl ej ui c ewer e equi val entt o800, 000t onsofr a w appl es .50% oft heJ apanes ei mpor t sofc ondens ed appl ej ui c ec o me f r om Chi na. Ao mor igr owshal foft he appl espr oduc ed i nJ apan,and about20 % oft hes e appl esar e us ed f or pr oc es s i ngandar es ol datver yl ow pr i c es .Thepr i c ef orappl esus edf orpr oc es s i nghasf al l enar ound60 %,and many appl ef ar mi ng bus i nes s es ar e oper at i ng i nt he r ed.The l o w pr i c es oft hes e pr oc es s i ng appl eshave af f ec t ed t he pr i c esoff r es h appl est hathave al s o dr opped s har pl y.Thus ,t he i nc o mesof appl ef ar mi ngbus i nes s e shavedec r eas edabout50%,f r o m 317, 000yenper10ar ei n1990t o157, 000yen i n1999.Thi shasc aus edgr avehar ds hi pf orf ar mer s . n addi t i on,Ianal yzed how t heappl ef ar mi ng bus i nes shasmadeef f or t st oover c omet heec onomi c I c r i s i s ,ast hec as es t udyonSōmavi l l agei nAomor ipr ef ec t ur e. 榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎Bul l .Fac .Agr i c .& Li f eSc i .Hi r os akiUni v.No.9:68 -79,2006 8 0 ニュージーランドのリンゴ生産と病虫害防除 − リンゴ火傷病とオタゴ大学訪問記録 − 宇野 忠義・佐野 輝男・田中 和明 地域資源経営学講座・環境生物学講座 (2 0 0 6年 1 1月 2日受付) は じ め に 表1 リンゴの主要国別生産量の変化と変化率(1 9 9 3~ 2005 ) 平均生産量(千 ㌧) ニュージーランドは南半球においてはチリに次ぐリン 国 名 ゴの生産国である。また,アメリカ大陸以外では初め て,1 9 1 9年にリンゴ火傷病が発生した国としても有名 である。 2 0 0 5年 7月,日本と米国の間で長年懸案事項となって いた米国産リンゴの火傷病に係る日本の植物検疫措置に 関する紛争処理小委員会で日本が敗訴し,WTOパネル 報告書が採択されたことにより,日本は火傷病検疫措置 の大幅な緩和を余儀なくされた。それゆえ,火傷病の日 本への伝播の危険性が非常に高まったと言える。それに 備えるために,この報告は,2 0 0 6年 3月,ニュージーラ ンドのリンゴ生産と火傷病など病虫害の発生,防除につ いてオタゴ大学,ダニーデン植物園及び果樹研究所クラ イド支所を訪問調査,収集したことを記録にとどめるも (1 ) のである。 なお末尾に,「ドイツにおける火傷病の発生と防除対 策並びに火傷病防疫条令」を参考記録として掲載してお 1 9 9 3 -5 2 0 0 3 -5 割合 (%) 変化率 (%) 世 界 計 4 9 , 2 6 5 6 1 , 8 9 2 1 0 0 . 0 125. 6 米 国 チ リ 中 国 トルコ イラン インド 日 本 韓 国 フランス イタリア ドイツ ポーランド ニュージーランド 4 , 9 5 4 8 3 3 1 1 , 4 1 1 2 , 0 9 2 1 , 8 7 4 1 , 2 2 3 9 8 8 6 4 9 2 , 5 5 7 2 , 1 0 5 1 , 7 7 0 1 , 5 2 4 4 8 2 4 , 3 2 3 1 , 3 0 0 2 3 , 2 6 4 2 , 4 1 7 2 , 4 0 0 1 , 4 7 0 8 2 2 3 6 8 2 , 1 5 9 1 , 9 8 1 1 , 5 9 0 2 , 3 3 3 4 8 8 7 . 0 2 . 1 3 7 . 6 3 . 9 3 . 9 2 . 4 1 . 3 0 . 6 3 . 5 3 . 2 2 . 6 3 . 8 0 . 8 87. 3 156. 0 203. 9 115. 5 128. 1 120. 2 83. 3 56. 6 84. 4 94. 1 89. 8 153. 1 101. 3 注:中央果実基金『海外の果実生産・貿易状況 2 0 0 6年版』 (2 0 0 6 .3)1 0頁より作成。 きたい。 1 ニュージーランドのリンゴ生産と日本のリンゴ輸入 状況 している。また,ニュージーランドの果汁生産と輸出に ついてみると,2万 2千 ㌧ ~ 2万 3千 ㌧ が生産されて ニュージーランドのリンゴ生産について,世界の主要 おり,そのおおよそ半分の 1万 1千 ㌧ が輸出されてい 国と対比して示したのが第 1表である。2 0 0 3 -5年平均 る。 の世界のリンゴ生産量は,6 , 1 8 9万 tであり,1 0年間に 日本への輸入は,1 9 9 3年 6月,日本の植物防疫法の改 2 6 %も増加した。地域別にはアジアの伸びが著しく,こ 正により,火傷病の発生国ニュージーランドから 6品種 とに中国は 2倍以上の驚異的な伸びを記録している。中 について解禁された。1 9 9 4年から 9 6年にかけて毎年 国は,今や,2 , 3 2 6万 tに達し,世界の 3 8 %を占めるに 2 0 0 ㌧ 前後が 3年間の平均 CI F価格 1 ㎏ 当たり 2 6 6円で 至った。他方では,米国,ヨーロッパ,日本,韓国など 輸入されたが,その後減少し,19 9 9年以降は輸入されて では生産がかなり減少している。ヨーロッパではポーラ いない。ちなみに,米国からのリンゴ輸入は 1 9 9 5年に ンド,南米ではチリが中国に次ぐ高い伸びを示し,注目 開始され,8 9 3 5㌧ が同 1㎏ 当たり 1 6 4円で輸入された される。ニュージーランドのリンゴ生産は 4 9万 ㌧ 前後 が,品種,品質と価格,残留農薬問題などによりその後 を維持しており,2 4番目の位置にあるが,リンゴの輸出 は激減している。 量は 3 0万 ㌧ 前後と約 6 0 %の生果を輸出しており, リンゴの有力な輸出国であった両国からは輸入解禁後 2 0 0 4年には 3 6万 ㌧ に達し,世界の 8番目の位置を維持 も日本の火傷病の検疫措置が厳しすぎる,それが輸出の 弘大農生報 No.9:80 -90,2006 ニュージーランドのリンゴ生産と病虫害防除 81 表 2 ニュージーランドの果樹病害リスト(Summer f r ui ti nNe w Ze a l a ndより抜粋) 害虫 Pl ant f eedi ng mi t es ,Pr edat or y mi t es ,Thr i ps (アザミウマ) ,Aphi ds (アブラムシ) ,Le a f r o l l e r s (ハマキガ) , Or i ent alf r ui tmot h(ナシヒメシンクイ) ,Co dl i ngmot h(コドリンガ) ,Sc a l e s (カイガラムシ) ,Me a l ybug(イ ボタムシ) ,Wheatbug,#Mi r i d, Ear wi g(ハサミムシ) ,Che r r ys l ug,Gr a s sgr ub o rgr a s sgr ub be e t l e ,Br onze beet l e,Vi negarf l y[ Fal s ef r ui tf l y] (ショウジョウバエ(スバエ) ) ,Bo o kl i c e(チャタテムシ) 線虫 Rootl es i onnemat ode(ネグサレセンチュウの 1種) ,Ri ngnemat o de(ワセンチュウの 1種) ,Da gge rnemat ode (ハリセンチュウの 1種) ,Root knotnemat o de(ネコブセンチュウの 1種) ,Pi nenemat o de(ピンセンチュウの 1 種) ウイルス類病害 Pr unusnec r ot i cr i ngs pot ,Pr unusdwar fvi r usa nd pe a c hr o s e t t e di s e a s e ,Pl um l i ne pa t t e r n vi r us ,Pl um mo t t l el eaf ,Apr i c otc hl or ot i cl eafmo t t l e ,Pe a c h da r kgr e e ns unke n mot t l e ,Pe a c h Ca l i c o ,Pe a c h ye l l ow mo t t l e,Peac hc hl or ot i cs pot ,Cher r yr a s pl e a fvi r us ,Che r r y ne c r o t i cr us t y mo t t l e ,St r awbe r r yl at ent r i ngs pot ,Cher r yr us t ys pot ,Cher r ygr e e nr i ngmo t t l evi r us ,Li t t l ec he r r yvi r us 菌類・細菌病 Peac hl eafc ur l (モモ縮葉病) ,Cher r yl e a fc ur l (オウトウてんぐ巣病) ,Bl a dde rpl um(スモモふくろみ病) , St onef r ui tbr own r ot (核果類灰星病) ,Pe a c hr us t (モモ褐さび病) ,Apr i c o tgr e e nf r ui tr o t ,Pr e a nd pos t har ves tf r ui tdec aysand bl emi s hes ,Ba c t e r i a ls po t (アンズ・モモせん孔細菌病) ,Bc t e r i a lbl a s ta nd de c l i ne, Di ebac k di s eas e,Peac hs c ab(モモ黒星病) ,Va l s ai nj ur y(モモ類胴枯病) ,Eut ypac a nke r ,Si l ve rl e a f (リン ゴ・ナシ・モモ銀葉病) ,Powder ymi l dew(モモうどんこ病) ,Rus t ys po t 土壌病害 Ar mi l l ar i ar ootandc r ownr ot (ならたけ病) ,Phyt o pht ho r ar o o ta ndc r ownr o t s (疫病) ,Ve r t i c i l l i um wi l (半 t 身 萎 ち ょ う 病) ,Fus ar i um c r own and r o o tr o t ,Ro s e l l i ni ar o o tr o t (白 紋 羽 病) ,Amyl o s t e r e um r o o tr ot , Cr wo ngal l (根頭がんしゅ病) 妨げになっているとして,毎年のように日本政府に緩和 を申し入れてきた。さらにまた,2 0 0 2年に,米国は,日 本の検疫措置について SPS協定違反として WTOへの 提訴を行った。 2.ニュージーランド南島オタゴ地区の果樹生産と果樹 病害 (1)オタゴ大学とその周辺 1)オタゴ大学: ところで,ニュージーランドは,隣国オーストラリア Kni ght先 生(Bus i ne s s学 部 Ma r ke t i ng学 科)は との間で 8 0年以上に渡るリンゴ火傷病の検疫措置・輸入 J o ur na lo fPubl i cAf f a i r sに「Adva nc eAus t r a l i aFa i r ? 禁止 を 巡 っ て 対 立が続いている。オーストラリ ア は The Ana t o my a nd Pa t ho l o gy o fa n8 4 Ye a r Tr a de ニュージーランドで 1 9 1 9年にリンゴ火傷病が発生した Di s put e 」と題する論文を発表したばかりだった。ニュー ため,1 9 2 1年からニュージーランドからのリンゴ輸入 ジーランド(NZ)とオーストラリア間に生じたリンゴの は禁止してきた。ところが 1 9 9 7年 5月にオーストラリ 輸出入と火傷病検疫に関わる貿易問題を論じたもので, アで火傷病が「発見」されるという出来事が起こり,大 NZは火傷病の発生国でオーストラリアにリンゴを輸出 問題となった。豪州政府は自国でのバクテリアの検査を するという立場からの意見である。日本と米国の同問題 行なったが,同時にドイツの植物検査研究所へもサンプ の関係に当てはめると,米国側の立場の意見ということ ルを送付して第三者の検査を依頼した。検査の結果,バ になる。Kni ght先生によれば,① WTOパネル(火傷病 クテリアは Er wi n i aa my l o v o r aという比較的稀な無毒性 検疫紛争)では日本に火傷病があるかないかは問題にさ のタイプであることが判明した。リンゴ火傷病ではな れなかった,②生果実の輸出入が火傷病の拡散の原因と (2 ) かったのである。 なるかどうかが議論の中心であった,また,③ NZでは オーストラリアと韓国はリンゴ生産国でありかつ輸入 火傷病は同国内に長い間(記録では 1 9 1 9年以来)存在し 国でもあるが,リンゴ火傷病は未発生であり,また両国 ているのでコントロール可能と考えている,ということ ともリンゴ火傷病の発生国からのリンゴ輸入は禁止して であった。①と②に関しては日本でもそのように聞いて いる点で共通している。火傷病に最も厳重な輸入禁止措 いる。③に関しては,後述のように特にオタゴ地区では 置で臨んできたことで知られている。参考のために, そうかもしれない。オタゴ大学内の書店で見たバイエル オーストラリアが輸入解禁に当たって検討しているリン (株)が出版している農作物の病害防除マニュアルでは, ゴ火傷病の検疫措置について注(3 )に記しておこう。ま よく管理された園地では火傷病はコントロール可能と記 た,国内の黒星病対策についても注(3 )に示しておこ 載されていた。また後述の中央オタゴ高原クライデ町の う (3 ) Ho r t Sc i e nc e (園芸試験場)で戴いた NZの果樹類病害に 関する本「Summe r f r ui ti nNe w Ze a l a nd:Ma na ge me nt 8 2 宇野・佐野・田中 o fPe s t& Di s e a s e s 」には,火傷病の記載がなかった(第 (写真 3 )。また,民家の庭木にはコトネアスターが好ん 2表) 。 で植えられている風があり,枝枯れ症状も散見された。 その他,Kni ght先生から,④ NZではリンゴのエチレ ただ,これらを持ち帰って調べることはできず,その症 ンの発生をコントロールして北半球のリンゴ端境期に輸 状が何に起因するものか確認することはできなかった。 出したいと考えていること,⑤輸出に関する調査の結 果,中国産リンゴが将来 NZの脅威となると考えている (2)ENZA(エンザ) こと,⑥ NZからオーストラリアへのリンゴの輸出に関 ENZA(エンザ)は 1 9 9 1年に組織された「ニュージー して,オーストラリア側は小さな問題と考えているが, ランドリンゴ・ナシ輸出販売協会」の名称である。国内 NZのリンゴ産業にとっては,リンゴをオーストラリア 4箇所に拠点を持ち,今回訪問したダニーデンの ENZA に輸出できるかどうかは大変大きな問題であること,⑦ はオタゴ地区の 2 0戸の生産者と毎年栽培契約して,世 遺伝子組換え作物(GMO)の問題(ただし,これはリン 界4 0カ国にリンゴ(1 0品種) ,ナシ(3品種),アプリコッ ゴ以外の話)に興味を持っていること,などを伺った。 ト,サクランボ,ネクタリン,スモモなどを輸出してい 特に GMO農産物の拡大が NZの農業に大きな影響を与 る。収穫物はここの貯蔵庫に集められ,リンゴの場合, えそうだという話は興味深かった。2 0 0 1年 GMO禁止 2 0 Kg入ケース(9 0 ,1 0 0 ,1 1 0 ,或は 1 2 0個入)に詰め のモラトリアムが全面解禁された後,GMOが NZ農業 られて輸出される。今回倉庫で見たものは英国向けで に与える影響を分析した結果,GMOの利益が危険性よ あった(写真 4 )。この地区の出荷量は,大きな生産者で り大きいという評価が得られ,認可申請等にはケースバ 3 0万ケース,小さいところで 1 0万ケース,摘み取り期 イケースで対応していること,今後 1~ 2年で中国とイ には 2 0 0~ 4 0 0人の季節労働者・アルバイトを募集する ンドの GMO農産物が飛躍的に増加して世界の市場が大 という。因みに,NZ最大の生産地 Ha wk’ sBa y(北島) きく変化するかもしれないというお話であった。 では 8 0 0万ケースという生産者もあるという。 2)ダニーデン植物園 NZでは今リンゴの国際価格が安くて利益が少ないた 火傷病は北米起源の病気で,最初の報告は 1 7 8 0年 めリンゴ生産者が減少している。代わりにブドウが増え ニューヨーク州のハドソン渓谷で記載された。もともと ているという。ENZAでは NZで育種した新品種 J AZZ リンゴやナシの病気だったわけでなく,米国土着のサン e bur n)に力を入れており, (ジャズ:Ro ya lGa l a ×Br a ザシ(Cr a t a e gus ) ,自生のクラブリンゴ(Ma l us属), これから生産調整しながら独占販売してゆく予定である ナナカマド(So r bus )などバラ科植物を宿主として進化 という。 してきたものが,栽培果樹類の導入により同じバラ科の 貯蔵施設や品質管理の様子を見学した。抜き取り検査 リンゴやナシに宿主域を拡げ,栽培地域の拡大と共に北 ではリンゴをコルクボーラーで打ち抜き,糖度と硬度を 米大陸全体に拡散し,終には大洋を渡って NZ,ヨー 測定する。糖度はフラックス 8 -1 0度で合格。「ふじ」は ロ ッ パ 全 域,西 ア ジ ア ま で 拡 が っ た も の で あ る 蜜入りが問題になるようで,判別用の蜜入りの程度を示 (Va nne s t e ,2 0 0 0 ) 。前述のように NZの果樹病害に関 した写真が壁に貼ってあった。また,施設の有効利用の する専門書には火傷病の記載がなかったが,オタゴ大学 ために,隣の貯蔵庫には輸出用のチョコレート(ダニー 内或は市内の書店に並べられているガーデニングの一般 デンに有名なチョコレート会社の工場がある)や魚類な 書のコトネアスターやピラカンサ(共にバラ科の庭木) どが冷凍保管されていた。 の解説欄には必ず「火傷病に感染する可能性がある」或 は「火傷病の被害を受けやすい」という記述があった (Mul l e r ,2 0 0 4 ,Br ya nt ,2 0 0 5 ) 。商業的果樹栽培では栽 (3)園芸試験場クライデ支所(中央オタゴ高原クライデ 町)とその周辺 培者の意識も高く防除を徹底することによりコントロー 園芸試験場クライデ支所 (Hor t Sci ence, Cl y deBr anch) ル可能かもしれないが,一般家庭の庭先の防除まで徹底 園芸試験場クライデ支所は,ダニーデンから北へ車で することは難しい。火傷病が定着してしまった国では, 約 3時間,セントラルオタゴ・クライデ町にある。1 9世 このような一般向けのガーデニング解説書で注意を促す 紀の半ばから金鉱山として栄えた町であるが,現在は冷 ことにそれなりの意義があることを理解すると共に, 涼で乾燥した気候を利用して果樹類の栽培が盛んな地帯 NZでは既にその根絶は困難で火傷病と共存してゆかな である。かなりの乾燥地帯で,年間降水量は 3 5 0 mm 程 ければならない状況であることを実感した。 度,NZ最大のリンゴ栽培地域 Ha wk’ sBa yの 3分の 1 火傷病発生国におけるリンゴやナシ以外の火傷病菌の である。降水量の多い順に Ha wk’ sBa y,Ne l s o n,Cl yde 宿主植物の状況を観察するため,オタゴ大学のあるダ で,リンゴ生産量の順番と同じである。 ニーデン市の植物園とその周辺を散策した。ダニーデン i l lMa c La r e n 支所長で,唯一の研究者格のスタッフ,J 植物園ではクラブリンゴに黒星病と枝枯れ症状が見られ さんに案内していただいた。昆虫学者である。この地帯 た(写真 1 ,2 ) 。植物園近くの墓地を取り巻く生垣のサ の果樹栽培面積は 6 0 0 ha ,農家の平均作付面積は 4 0 - ンザシ(Cr a t a e gus )の新梢には葉枯れ症状が見られた 5 0 ha 。そのうちリンゴは 1 0 -2 0 ha程度である。ここ ニュージーランドのリンゴ生産と病虫害防除 83 1 2 3 4 図1 -4. 1.ダニーデン植物園 クラブリンゴ黒星病,2.ダニーデン植物園 クラブリンゴ枝枯れ, 3.ダニーデン植物園近く サンザシ枝枯れ,4.ENZA 倉庫 英国向けリンゴ でもリンゴ栽培は減少しているという。火傷病は現在こ 剤,蛾の生長促進のために砂糖(シュガー)液,紫外線 の地域では問題になっておらず,場内で発生することも 焼けの防止に乳脂肪(ミルクファット)液を散布してい あるが,伐採して今は残っていないということであっ る。病害虫では黒星病と葉巻蛾(Le a fr o l l e rmo t h)が問 た。火傷病の流行が春先,開花期の高温・多湿とそれに 題という。また火傷病と共に日本未発生の重要病害虫コ 続く激しい降雨などを引き金として発生することを考慮 ドリンガの被害もある。見学中に作業員がコドリンガの すると,この地域で火傷病が問題になっていないのは, 被害果実を見つけて持ってきてくれた(写真 5 )。有機栽 多分に乾燥した気候条件に拠るところが大きい。ただこ 培でも今のところ火傷病の問題はないが,Ro ya lGa l a のような乾燥地帯でも,火傷病菌が定着している限り, と Br a e bur nの血を引く J AZZは火傷病に弱いかもしれ 開花期の高温・多湿など,ひとたび火傷病菌の繁殖に好 ず,J AZZ栽培普及による将来的な懸念はあるようだ。 都合な異常気象に見舞われると,瞬く間に火傷病が大流 研究棟で葉巻蛾(Le a fr o l l e rmo t h)の抵抗性検定試験 行する恐れがある。話しを伺っていて,クライデでも火 とリンゴ黒星病(Sc a b)の発生予察の説明を受けた。黒 傷病の発生・流行を常に気にかけている雰囲気が感じら 星病の発生予察は,降雨と気温(積算温度)のデーター れた。 から病気の発生危険度を予測して 3段階で表示し, 場内の有機栽培リンゴ試験園地を見学した。ちょうど e;www.ho r t pl us .c o m(有料)で生産者に情報 We bs i t 有機栽培 J AZZの収穫時期であった。収穫物はまだ若 提供している。栽培者はこのデーターを見て薬剤散布の 干早取りの感じであったが,酸味と甘みがあり美味し 判断を行う。3段階の危険度のいずれかを示す警報が出 い。小粒だったが,それは英国人が米国人よりさらに小 た時,例えば 2 -3日前に散布していれば再度散布しな さいリンゴを好むためということであった。薬剤散布 いが,1週間前なら散布する,という具合に生産者が判 は,銅-石灰硫黄(ライムサルフェート)を ha当たり 断して散布するかしないかを決定するという。他にナシ 3 0 Kgまで散布できる。その他に,殺虫のために BT 灰星病の発生予察システムも稼働中であった。 8 4 宇野・佐野・田中 5 6 7 8 9 1 0 図5 -10. 5.クライデ園芸試験場 J AZZコドリンガ被害果実,6.クライデ町の商業りんご園, 7.クライデ町 コトネアスター枝枯れ,8.クライデ町 ピラカンサ枝枯れ, 9.クライデ町 コドリンガ被害リンゴ,1 0.クライデ町 マルメロの果実腐敗 商業リンゴ園 期には 2 0 -4 0人程度の季節労働者が世界中から集まる 近隣のリンゴ園を見学した(写真 6 ) 。ちょうど収穫し という。もちろん日本人もいる。リンゴの農薬散布は最 たリンゴを運搬して来た園主の奥さんから話しを聞くこ 低 2回,対象はコドリンガと黒星病である。 とができた。リンゴ 5品種,サクランボ 1 3品種,アプリ 薬剤で摘果しているが,風向きによって散布ムラがで コットなどを栽培している。通常の作業員は 8人,サク きることがある。効きすぎると落果が多くなり,大きい ランボの摘み取り時期には 2 0 0人程度,リンゴの収穫時 のができて困るらしい。私たちには程よい大きさの立派 ニュージーランドのリンゴ生産と病虫害防除 なリンゴに見えたが,欧米では好まれないためである。 また,NZでは来年から日本の正月時期に合わせてサク ランボ(黒い品種)を輸出する予定があるという。もち ろん日本の「佐藤錦」がすばらしい品種であることを認 識している。 クライデ町の住宅の庭木 クライデは開拓時代の雰囲気を残す静かな田舎町で, よく手入れされた庭木に囲まれた平屋の綺麗な家が多 い。ちょうど初秋の紅葉の始まりの時期で,どこの家の 庭にもコトネアスターやピラカンサ,クラブリンゴなど 火傷病菌の宿主となる庭木が植えられ,綺麗な実をいっ ぱいつけていた。最も近い商業リンゴ園でも 1 -2 km の距離にあるとはいえ,特に火傷病菌の宿主植物を庭木 に植えることに気をつけている風には感じられなかっ た。数軒の庭先を見て回ったが,コトネアスターやピラ カンサに気になる枝枯れと果実腐敗が散見された(写真 7 ,8 ) 。 町の中心,嘗ての鉄道駅の近くに町の歴史博物館があ り,その裏庭のリンゴ樹にコドリンガが大発生していた (写真 9 ) 。被害果はほとんどが落果しており,残ったも のも穴だらけであった。また,そのすぐ脇のマルメロの 木にはかなりの数の黒く干からびた未熟果が見られ,一 部が茶色に変色する果実腐敗がたくさん発生していた (写真 1 0 )。私達が火傷病かもしれないと写真を撮って いると,隣家のご主人がやってきて Br o wn r o t (灰星 病)だろうと教えてくれた。昔,園芸試験場で働いてい たことがあるそうで,意外と詳しい。 参考のために,NZ南島のリンゴ生産の中心地である ネルソン地域における火傷病対策として垣根のサンザシ (4 ) の排除計画を注記で示しておきたい。 謝 辞 オタゴ大学は平成 1 2年以来弘前大学の姉妹校として 順調な学生・教員の交流が続いている。2 0 0 6年 3月末の 視察調査もオタゴ大学国際交流部門を中心としたスタッ フの方々の熱心で誠意あるご協力に拠るところが大き かった。ニュージーランド滞在中のスケジュールから訪 問先への連絡,バスやホテルの予約まで全てをお世話く ださった Al i s o n Fi ni ga nさん,Sa ndy Ma c Andr e wさ ん,日本語学科の郭南燕先生には心から感謝の意を表す る。 なお,本研究及び NZ視察調査は,平成 1 7年度及び 1 8年度弘前大学学長指定緊急重点研究,弘前大学農学生 命科学部 5 0周年記念地域振興支援研究助成,りんご協 会及び津軽農民組合の呼びかけによる研究費募金を受け て実施したものである。記してお礼を申し上げたい。 また,後掲付記のドイツ語文献の翻訳については弘前 大学人文学部原田悦雄先生のご協力に感謝したい。 85 注 (1)火傷病については,To m Va n De rZwet& Ha r r y L. Ke i l" FI RE BLI GHT" , USDA, 1 9 7 9 ,J . L. Va nnes t e " Fi r eBl i ght " ,CABIPubl i s hi ng,2 0 0 0 ,水野明文・佐藤成 良他「日本における火傷病発生の疑義」 (横浜植物防疫所 『植物防疫所調査研究報告』第 3 9号,2 0 0 3年等を参照。 また,ニュージーランドの火傷病に関しては,畔上耕児 他「ニュージーランドにおける火傷病の発生生態と防除」 ( 『植物防疫』第 6 0巻 3号,2 0 0 6年)が詳細に報告して いる。なお,火傷病の日米検疫問題については,宇野忠 義「リレーエッセイ 時代を読む・重大病害リンゴ火傷 病に万全の対策を望む」 ( 『現代農業』2 0 0 5年 7月号) ,宇 野「最重要病害リンゴ火傷病の日米検疫問題」 ( 『弘前大 学農学生命科学部学術報告第 8号』 ,2 0 0 5年 12月), 宇野「最重要病害リンゴ火傷病の日米検疫問題の交渉経 過と残された問題」 (農業・農協問題研究所『農業・農協 問題研究』第 3 5号,2 0 0 6年 8月)を参照されたい。 (2)ニュージーランドとオーストラリアとの間のリンゴ火傷 病の検疫を巡る永年の対立を象徴するような出来事が, 1 9 9 7年 5月に発生した。メルボルンの植物園で,ニュー ジーランドの研究者が「火傷病の発生を発見」して大騒 ぎになった事件である。長い引用になるが,紹介した い。 Sub:豪州,971220,火傷病の調査結果が白に <要約> 今年 5月にニュージーランドの科学者によっ て発見されたリンゴの火傷病について,豪州政府は国外 の研究機関に検査を依頼したが,バクテリアは無毒性の ものであることが判明した。また,春秋の実地調査にお いても病原菌は発見されなかった。これにより,これは 豪州にリンゴの輸入解禁を迫るニュージーランド側の意 図的な仕掛けではないかとの疑惑が再燃した。 <本文> 今年(1 9 9 7年)の 5月,リンゴに大被害を及 ぼすバクテリア性病原菌火傷病(Fi r e bl i ght )がメルボル ンとアデレードの植物園で発見される事件があった。火 傷病は果実の口蹄疫にたとえられる深刻な病原菌である が豪州には存在しない。豪州政府は,発見以来,秋と春 の 2回にわたって全国の果樹産業地域で集中的な検査を 行なってきた。1 2月の初めに発表された第 2回目の春 の調査結果でもいずれの州でも火傷病は全く認められな かった。この事件が原因でタスマニア産リンゴの日本へ の輸出申請手続きが中断されているが,リンゴ・ナシ生 産者協会は近いうちに再開されることを期待している。 火傷病は旅行で来豪中の隣国ニュージーランドの科学 者によって発見された。火傷病原菌の植物園での発見と 同時に,豪州政府は自国でのバクテリア菌の検査を行 なったが,同時にドイツの植物検査研究所へもサンプル を送付して第三者の検査を依頼した。検査の結果,バク テリアは e r wi naamyl o vo r aという比較的稀な無毒性の タイプであることが判明した。植物園で確認された火傷 病に感染している潅木は検疫・隔離をして観察したが, 病気が広がる形跡は認められなかった。そして今回の 2 回にわたる豪州国内のリンゴ,ナシの各果樹園での集中 検査へと至った。 豪州政府は火傷病を理由にニュージーランドからのリ ンゴの輸入を長年禁止してきている。火傷病の発見者が 豪州政府に解禁を迫っている隣国ニュージーランドの科 学者 2名であったことから「意図的な仕掛け」ではない かと騒ぎを一層大きくした。さらに,この発見が輸入解 禁を迫るニュージーランド政府の新規申請書に対して豪 州政府が輸入禁止維持を勧告した直後に起こったため, 疑惑が一層強まり,今回の調査結果ではその疑惑が再燃 8 6 宇野・佐野・田中 する形となった。産業界はこの調査結果をまたとないク リスマスのプレゼントになったと歓迎している。[ヤ フーの「リンゴ火傷病」のサイトより原文のまま引用] なお,輸出の解禁を迫るニュージーランド側からの オーストラリアの輸入禁止・火傷病の検疫措置に対する 論評として,オタゴ大学教授 J ohn Kni ght “Adva nc e Aus t r al i af ai r ?Anat omy and pat hol ogy ofan 8 4 ye a r t r adedi s put e ”(J .Publ .Af f .5:112123,2005)がある。 私たちはオタゴ大学において,ナイト教授とニュー ジーランドとオーストラリア間及び日米間のリンゴ火傷 病の検疫問題と WTOのパネルでの日本の敗訴などにつ いて意見を交わした。双方の論文の交換も行った。火傷 病発生国であり,かつ主要輸出国である国ニュージーラ ンドと未発生の輸入国日本との立場の違いが大きいこと を改めて感じた。また,米国が日本のリンゴ火傷病の検 疫措置を提訴した際の裁定委員の一人がニュージーラン ド人であったことが思い出された。 (3)参考までに,オーストラリアの病害虫・植物検疫につい て紹介しておこう。 < NZ産リンゴの改定輸入リスク分析報告書(案) を公表> 豪州がニュージーランド(NZ)産リンゴの輸入を禁止 したのは 1921年のことで,その後 NZが同国産リンゴ の輸入を認めるように申し入れてからも既に 20年近い 年月が過ぎたが,輸入解禁の是非を巡るせめぎ合いはさ らに続きそうである。 2005年 12月 1日,NZ産リンゴの輸入リスクの分析 を行っているバイオセキュリティーオーストラリア (BA)は,2004年 2月に公表した分析結果報告書(案) の改訂版を公表した。 <輸入解禁のための植防条件> このほど公表された改訂版報告書(案)では,NZ産 リンゴの輸入解禁は,①輸入に先立って NZで行われる あらゆるリスク管理措置に際して豪州の植防・検査機関 担当官の立会いを義務付け,これには NZ当局が輸出承 認に際して行う諸手続きの仕組みや運用状況に関し聴取 することを含むこと,②リンゴを輸出しようとする果樹 園は火傷病に犯されていないこと,また,火傷病の症状 が認められたリンゴ園で栽培されたリンゴ園の輸出は認 めないこと,③火傷病菌による汚染を防止するために選 果・荷造り施設で塩素等による消毒を行うこと,④秋か ら 冬 に か け て 落 葉 後 の リ ン ゴ 園 を 検 査 し,Eur o pe a n c anke r (がんしゅ病)に犯されていないことを確認し, 犯されていると認められた場合,その園地で栽培された リンゴの輸出は認めないこと,⑤ NZで,無作為に収集 されたリンゴ 3000個について appl el eafc ur l i ngmi dge (タマバエの一種でリンゴだけを宿主とし,葉を巻いて 中に生息する)がいないことを確認し,もし appl el e a f c ur l i ngmi dgeの存在が認められた時は輸出差し止めか 消毒を行うこと,⑥これら以外の病害虫についても検査 を行い,要すれば然るべき措置を施すこと等の厳しい植 物衛生上のリスク管理システムの下で認めるとしてい る。 同報告では NZ産リンゴのウエストオーストラリア (WA)州への輸入を禁止するとしている。これは,これ まで WA州ではリンゴ黒星病の発生が見られず,有効な リスク評価が確立していないことによるものである。ち なみに WA 州以外の州では既に黒星病は存在している ことから,他州から WA州へリンゴを持ち込むことは禁 じられている。 [中央果実基金『中央果実基金通信・海外果樹農業 ニュース No .8 4』 (2 0 0 6 .1) (現地報告,トニー・ムー ディ「豪州における最近の果樹産業を巡る状況」36~ 37 頁)より引用。 ] (4)ニュージーランドのネルソン地域における火傷病対応策 ネルソンを中心とするリンゴ栽培地域のタスマン地区 協議会(Ta s ma nDi s t r i c tCo unc i l )は,火傷病を撲滅す るため,サンザシの垣根を取り除く活動を展開してい る。サンザシは本病原菌のホスト植物であり,その垣根 を取り除くことは感染の危機を減らすことになる。これ と同様の排除計画は,最近,ブレンハイム地区でも実施 されている。 [中央果実基金『中央果実基金通信・海外果樹農業 ニュース No .8 4』 (2 0 0 6 .1) (現地報告,ビッキー・ハイ ド「ニュージーランドにおける最近の果樹産業を巡る状 況」3 3頁) 。 ] 引用文献と参考資料 ① BRYANT G. :Ga r de ni ng e a s y Tr e e s .Ra ndom Ho us e.96 p.2 0 0 5 . ② KNIGHT, J . : Adva nc e Aus t r a l i a f a i r ? Ana t omy a nd pa t ho l o gy o fa n8 4 ye a rt r a dedi s put e .J .Publ .Af f .5: 1 1 2 1 2 3 ,2 0 0 5 . ③ MCLAREN,G. F. ,GRANDISON,G. ,WOOD,G. A. ,TATE,G. , HORNER,I . :Summer f r ui ti n Ne w Ze a l a nd:Ma nage me nt o f Pe s t & Di s e a s e s . Ho r t i Sc i e nc e AGMARDT / Summer f r ui tNe w Ze a l a nd I nc .U.o fOt a goPr es s .136 p.1 9 9 9 . ④ MULLER,J . :Pl a ntpr uni ng A t oZ.Da vi d Ba t eman Lt d. 1 6 0p.2 0 0 4 . ⑤ VANNESTE,J . L. :Fi r e Bl i ght ,CABI Publ i s hi ng,370 p. 2 0 0 0 . ニュージーランドのリンゴ生産と病虫害防除 87 Appl ePr o duc t i o na ndPes t sMa nage menti nNe w Zeal and Tada yo s hiUNO,Te r uoSANO a ndKa z ua kiTANAKA La b o r a t o r yo fRe g i o n a lRe s o u r c eMa n a g e me n t ,La b o r a t o r yo fPh y t o p a t h o l o g y “Fi r ebl i ght ”i soneoft hemos tdes t r uc t i vedi s eas esf orappl eandpearpr oduc t i on.Thedi s eas ehas l ong been r es t r i c t ed i n Nor t h Ame r i c a,butes c aped and di s s e mi nat ed amo ng near l y 40 c ount r i es i n Eur ope,Mi ddl e Eas tand Oc eani a.Amo ng t hem,New Zeal and was r ec or ded i n 1919 as t he f i r s tf i r e bl i ghtepi de mi cc ount r youts i det heNor t hAme r i c a. J apan had i nt er di c t ed t hei mpor tofappl esf r o m USA and New Zeal and bec aus eoft hequar ant i ne pr obl e m c onc er ni ng Fi r e bl i ght .I n 1994,however ,J apan had r e moved t he ban on t wo ki ndsofappl es f r om USA by t hei rs t r ong pr es s ur et o open t he mar ket .The s t r ai ned negot i at i onson t he quar ant i ne ys t em.Af t erl ong met hodsf orFi r ebl i ghts t ar t ed s i nc et hen bet ween J apan and USA,undert heWTO s negot i at i ons ,J apanhasbeendef eat edi nt heWTO dec i s i onsi nJ ul y2005andt hegover nmentdec i dedt o al l evi at es i gni f i c ant l yt hequar ant i nemet hodsf orFi r ebl i ght .Ta ki ng t hes ec hangesi nc ons i der at i on,t he t hr eatofFi r ebl i ghti nvas i on i nt oJ apan hasi nc r eas ed mor eorl es sf ort hef i r s tt i mei nt hel as ts ever al dec ades . ms o ft hi sr epor t ar et oi nt r oduc ec ur r ent s t at us of appl e pr oduc t i on and t he pes t The ai ma nageme nt i nt he c ount r y wher e Fi r e bl i ght has l ong been es t abl i s hed,i . e. ,New Zeal and,Ot ago di s t r i c t ,asac as es t udy. l .Fac .Agr i c .& Li f eSc i .Hi r os akiUni v.No.9:80 -90,2006 榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎Bul 宇野・佐野・田中 8 8 付記:ドイツにおける火傷病の発生と防除対策並びに火傷病防疫条令 宇 野 忠 義 リンゴ火傷病はヨーロッパでは 1 9 5 7年に英国で初め くなっていく。病原体は花の他に傷から寄生植物に入り て発生し,その後,海峡を渡って 1 9 6 6年にポーランド, 込むこともある。真夏の雹が降った後で,このようにし オランダに伝播し,さらに 6 8年にデンマーク,1 9 7 1年 て新たな感染が起こり得る。病原体は老木に入り込ん にドイツ,7 2年にフランス,ベルギーへと感染地域を広 で,その樹皮の中で生き続けたりする。Ca nke rと言わ げていき,現在では域内 2 0カ国に及んでいる。 れるこの部分はややくぼんでおり,褐色になっている。 ドイツでは南部のりんご生産地帯で 1 9 9 3年から 9 6年 春になり気温が上昇すると,細菌は Ca nke rの周縁部で にかけて発生し,大きな被害を受けた。注目すべきこと 非常に早く繁殖し,新たな病原体の蔓延が始まることが は,リンゴやナシだけでなく,観賞植物や庭木にも多く ある。 発生していることである。それらの事情をドイツで入手 危険にさらされた庭園の樹木を綿密に監視することが した専門書の翻訳によって紹介しておこう。 最も重要な対策である。火傷病の症状が発見されたら, また,ドイツで火傷病防疫対策として 1 9 8 5年に制定 羅病した部分は大規模に取り除かねばならない。加えて された“Fe ue r br a ndve r o r dnung” (火傷病防疫条令)の 枝は目に見える傷から下部を少なくとも 3 0ないし 5 0 翻訳文を掲載しておく。 c m 除去しなければならない。これが必要なのは病原体 がまだ健康に見える木に侵入している可能性があるから 火傷病 だ。罹病した木を適切に廃物処理することが重要であ 火傷病菌は 2 0世紀半ばになって漸く北米から持ち込 る。病原体が伐採した木に残り続け,庭園内にしつらえ まれたが,とかくする間に欧州の大部分に蔓延した。最 た堆肥の山では病原菌を殺すに十分な高い温度が生じな 近の大きな感染波及は気象が原因となって 1 9 9 3年から いので,罹病した物は焼却するか家庭廃棄物として処理 1 9 9 6年にかけて南ドイツで発生した。 しなければならない。専門的堆肥産業プラントではそこ 自然の中で感染するのはナシ状花を結ぶバラ科だけで で発生する高い温度によって殺菌が保証されている。裁 ある。これにはリンゴ属,ナシ属の果樹と観賞植物の他 断工具は他の寄宿植物に使う前に例えば沸騰した湯で殺 にマルメロ属も入る。ナシ属の西洋果仲間にはコトネア 菌しなければならない。 スター属,サンザシ属にはサンザシとアカサンザシ,ナ 序でに言えば,危険地域ではコトネアスター種のよう ナカマド属にはナナカマドが含まれるのが目につく。栽 な感染率が高い寄宿植物の植樹は完全に断念すべきであ i ni a da vi di a naが火 培が稀な Funke nbl a t tすなわち Pho t ろう。 傷病菌に非常に感染しやすい寄生植物と見られている。 リンゴ種とナシ種についてもいくつかの種類は感染率 時には観賞用マルメロとその雑種,そしてとげのあるベ が低いことが観察されている。リンゴ種で火傷病の感染 ニバナが感染することがある。 率が低いものとして,例えば Fl o r i na ,Li be r t y,Ont a r i l , 茎頂が松葉杖状に湾曲するのが典型的症状と見なされ Re a nda ,Re mo ,Re ne ,Re we naが あ る。ナ シ の Gut e ている。外見はかなりの程度に的中するとはいえ,木本 s e種も感染率が低いと見られている。とはいえ,梨 Lui 植物は水の移動でも他の障害に対して同じように反応を 果の種別感染率に関してこれまで長期にわたって蓄積さ するのだ。唯一の確かな認知目印は,湿度の高い日に新 れた知識はなく,決定的な推薦をすることはできない。 芽に,リンゴやナシでは果実の上にも,現れる粘液状の 細菌病撲滅のための植物予防薬は出ていない。しかし 滴である。この樹齢と気象によって,ほとんどガラスの 1 9 9 3年 以 降,ス ト レ プ ト マ イ シ ン 硫 酸 塩 ように透明なものから琥珀色をした粘液状滴が,数百万 St r e pt o myc i ns ul f a tを持つ抗生物質を投与することが, の極小細菌を含んでいる。鳥,虫,そして果樹園を営む その都度非常に限られた条件下で特別許可を得ながらご 人間もが,この細菌を他の寄主植物へと広めるのであ く限られた地域に行える。しかしこの例外的許可は果実 る。 と観賞用植物の経営栽培にだけ出されてきた。ところが 細菌の集団的蔓延は春の梨花の開花時に起こる。この 若干の殺真菌性抗生物質は細菌に対して副作用を持って 時期に湿度があって気温が高ければ,細菌は特に繁殖が いるのだ。銅を含む殺真菌薬を短い間隔で定期的に散布 うまくゆき,病気の爆発的な蔓延が見込まれる。細菌は すると,新芽の感染を防げるだろうが,この薬剤適用指 虫によって開いた花へと運ばれ,そこから花柄を通って 示は目下の所は出ていない。とはいえ,銅入りの薬剤は 新芽に突き進んで行く。感染した花は黒褐色に見える。 植物の表面だけを感染から防げるのであり,作用物質は 病原体が侵入しつつある葉は中央葉脈隆起から同様に黒 植物内に入り込むことはない。例えば保護皮膜が雨,植 ニュージーランドのリンゴ生産と病虫害防除 Ch a e n o me l e sLi ndl . Zi e r qui t t e 物の生長あるいは傷によって破壊されると,再び感染可 別名ボケ Sc he i nqui t t e 能となるのだ。火傷病撲滅のために植物や鉱物を基盤に した薬剤とか自然に存在する敵対物について様々な試み Co t o n e a s t e rEhr h. シャリントウ Cr a t a e g u sL. サンザシ We i β do r nおよび Cy d o n i aMi l l . マルメロ Qui t t e Ma l u sMi l l . リンゴ Apf e l Zwe r gmi s pe l が始められたが,未だに最終的な結論が出ていない。火 傷病の重大性は火傷病規定に反映されている。この中に 89 報告義務から栽培禁止に至るまでの,対策に対する法的 t do r n ヒトシベサンザシ Ro 根拠が示されている。 細菌火傷 Py r a c a n t h aM.Ro e m.タチバナモドキ 火 傷 病 細 菌 に よ る も の と 非 常 に 似 通 っ た 症 状 が Fe ue r do r n Ps e u d o mo n a ss y r i n g a eの感染後に観察されることがあ Py r u sL. ナシ Bi r ne る。細菌火傷は非常に多くの広葉樹を襲う。特に感染し S o r b u sL. ナナカマド Ebe r e s c he やすいのはリラ,レンギョウ,ナシ,サクランボである。 S t r a n v a e s i aLi ndl . St r a nva e s i e 感染は湿度があって冷たい気象状況で促進される。晩霜 2 .特別感染しやすい宿主植物:品種が火傷病に特別 の後で病原体がどっと現れるのは,霜が原因となって若 感染しやすい宿主植物。農林業生物連邦研究所は い組織についた小さな傷のために,特に効力ある感染が 特別感染しやすい宿主植物を連邦官報に載せてい 可能となるからである。サクランボにあっては,病原体 る。 が秋にはもう葉柄の傷に入り込み,樹皮組織の中で蔓延 第 2条 していることがよくある。これは樹皮火傷症状とゴム質 (1 )処分権所有者と宿主植物の所有者は所管官庁に火 化異常樹液分泌病の反応を示し,罹病した新芽は死ぬこ 傷病の発生と発生疑惑を直ちに報告する義務を負 と が あ る。リ ラ,レ ン ギ ョ ウ,ナ シ の 場 合 は, う。報告には植物の種類,林の場所と規模,なら Ps e u d o mo n a s発病による被害は柔らかな新芽に限られ, びに長くて 2年間その場所にある植物の出所が記 木質化した組織には及ばない。 されているべきこと。 罹病した新芽は除去して焼却しなければならない。銅 )宿主植物を移植,増殖あるいは除去するために根 (2 を含む殺真菌薬を,特に秋の落葉時にサクランボの葉に をつけたまま動かす,つまり移動させる人は,こ 傷がつくことから守るために,投入することは,証明を の宿主植物の種類,出所,場所あるいは生育地を 欠いているので目下の所不可能である。基本的に春の新 指令に従って所管官庁に報告する義務を負う。 芽時に,しかるべき散布をすることで新芽感染を減らせ 第 3条 るかもしれない。とはいえ,繊細な組織はこの散布で傷 (1 )火傷病の発生が確認されたら所管官庁は発症した つけられることもあるのだ。 土地,発症疑惑のある土地,5 ㎞ 圏内にある発症 [原 典 は, J o c he n Ve s e r “Pf l a nz e nkr a nkhe i t e n” の危険に曝された土地もしくは耕地を隔離できる 9に掲載された (Ve r l a gEuge nUl me r ,2 0 0 3 )p.1 6~ 1 “Fe ue r br a nd” , “Ba kt e r i e nbr a nd”である。翻訳は,弘 (隔離地)。 (2 )再調査の結果,発症が認められず,最後の火傷病 前大学人文学部原田悦雄助教授の協力による。 ] 発生から 2年経過している場合,所管官庁は土地 の隔離を止めること。 火傷病防疫条令(火傷病条令) 第 4条 (1 )発症しているか発症疑惑のある宿主植物もしくは その一部は,その現在地から動かすことを禁止す 1985年 12月 20日(BGBI .I S.2551) る。 1 9 7 5年 1 0月 2日発令条文中の植物保護法第 3条 1項 No .1 ,3 ,8ないし 1 1 ,1 4 ,1 5に基づき食糧・農業・山 (2 )境界地に生育している宿主植物もしくはその一部 林連邦大臣により,また植物保護法第 3条 1項 No .5お は,所管官庁の許可を得てのみ地域から移すこと よび 6に基づき食糧・農業・山林連邦大臣により青少年・ が許される。 家庭・健康連邦大臣の協力を得て,連邦参議院の承認を 第 5条 受けて以下の条令が発令される。 火傷病防疫が必要とされる限り,所管官庁は長くて 5年 第 1条 間以下のことを禁止できる。 本条令の主旨に含まれるもの 1 .火傷病:Er wi n i aa my l o v o r a (Bur r i l l )Wi ns l o we t a l .を病原体として引きおこされる下記種類の植 物(宿主植物)病 Ame l a n c h i e rMe di k. ザイフリボク Fe l s e nbi r ne 1 .特別感染しやすい宿主植物を植えること 2 .特別感染しやすい宿主植物もしくは発症疑惑のあ る宿主植物を栽培材料として売買すること この禁止は反復可能である。 宇野・佐野・田中 9 0 第 6条 1 5 0 5 )第 4 0条第 1項 No .5の文言の意味するとこ (1 )火傷病防疫が必要とされる限り,処分権所有者と ろに従い,意図的もしくは不注意によって以下の 行為をなす人は違反行為をしている。 持ち主は所管官庁の指令に従い,以下のことを行 う義務を負う。 1 .第 2条 1項に違反し,報告をしない,適切にしな い,完全にしない,あるいは適切な時にしない 1 .特別感染しやすい宿主植物と感染した宿主植物を 現在地もしくはその隣接地で処分すること 2 .第 4条第 1項に違反し,宿主植物もしくはその一 部を現在地から移す 2 .宿主植物の存在する隔離地で火傷病防疫を行うこ と 3 .第 4条第 2項に違反し,宿主植物もしくはその一 部を許可なく隔離地から移す 3 .隔離地内で地所もしくは栽培地から宿主植物を除 去すること 4 .第 8条第 1項に違反し,火傷病の病原体を培養す る,あるいは保持する,あるいはそれを扱う 4 .発症した,発症疑惑のある,発症危険のある地所 を5 0 0 m の圏内まで種苗栽培園,増殖施設から離 (2 )1 9 8 6年 9月 1 5日発令の植物保護法第 4 0条第 1項 すこと,もしくは梨果栽培地から特別感染しやす No .2の文言 Cの意味するところに従い,意図的 い宿主植物を除去すること もしくは不注意によって以下の行為をなす人は違 反行為をしている。 (2 )所管官庁は 1項 No .1のケースで,火傷病拡大の 危険が起こらない限りにおいて,宿主植物を別の ,6条第 1項あるいは第 7条 1 .第 2条第 2項,第 5 によって実行可能な指令に逆らう行為あるいは 場所で処分することを許可できる。 2 .許可を得たことにより,第 6条第 2項あるいは第 第 7条 所管官庁は火傷病防疫が必要とされる限りにおいて,以 8条第 2項と結びついている実行可能な命令に逆 下のことを指令できる。 らう行為 1 .ミツバチを飼うこと 第1 1条 2 .養蜂業者が隔離地に出入りすること 本条令は 1 9 8 6年 1月 1日に発効する。 第 8条 (1 )火傷病病原体の培養と保持,ならびにこの有害有 機体を取り扱うことを禁止する。 (2 )火傷病防疫の妨げにならず,病気拡大の危険が生 榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎 改正は以下のとおり: ● 1 9 8 8年 5月 2 0日の条令(BGBI .I S.6 4 0 ) じない限りにおいて,所管官庁は調査,実験,培 ● 1 9 9 2年 1 1月 1 0日の条令(BGBI .I S.1 8 8 7 ) 養計画に関わる個別ケースで,この禁止の例外を ● 1 9 9 9年 1 0月 2 7日の植物保護法改革のための最初の 許可できる。 条令(BGBI .I S.2 0 7 0 ) 第 9条 植物保護法第 1項 No .5と結びつく第 3条 3項に基づく [グーグルによって「Fe ue r br a ndve r o r dnung」を検索 法規命令により,州政府は所管官庁が火傷病を防疫する し(2 0 0 6年 1 0月 1 7日),ドイツ語文章を弘前大学人文学 よう指令する権限を独自に持ち続ける。 部原田悦雄助教授によって日本語に翻訳されたものであ 第1 0条 る。] 5日発令の植物保護法(BGBI .I S. (1 )1 9 8 6年 9月 1 91 安全性の高い野菜に対する消費者意識 − みやぎ生協「産直野菜」に対する来店者意識調査 − 藤 崎 浩 幸 地域環境計画学講座 (2 0 0 6年 1 0月 1 3日受付) Ⅰ はじめに Ⅱ 「 産直野菜」 生産農家からの聞き取り 近年,消費者の食料の安全性や環境保全に対する意識 来店者意識調査に先立ち 2 0 0 3年 1 0月に宮城県角田市 が高まってきていると言われている。安全な食料が広く の「産直野菜」栽培農家へ聞き取りを行った。 供給されることは,食料を食べて体内に取り込む消費者 農家によると,農薬の使用量を半分以下に減らして栽 の健康はもとより,食料の生産地における生態系保全, 培しているために病虫害が増え,売り物にならない野菜 土壌汚染防止,域内物質循環システム構築といった環境 が 3割程増えた,そのために生産者としては現状の 2倍 保全のためにも望ましいことである。 くらいの値段で売りたいのが本音だが,消費者が離れて しかし,多くの食品売り場では,まだまだ「有機栽培」 しまうのでそれはできない状況だ,さらに農薬使用回数 とか「無(減)農薬栽培」という言葉を冠さない食品が を守らないなど,ガイドラインに違反した場合の罰則が 主流となっている。安全性に配慮した食料については, あり,野菜生産部会全体で連帯責任を負わせられるの 生産に手間がかかるため価格が高くなったり,農薬を使 で,産直野菜の生産を断念し部会を抜ける人も出てきて 用しないことで害虫や病気の影響を受けやすく見た目が いる,とのことだった。 悪くなったりするため,その普及があまり進んでいない ことが推測される。 Ⅲ 来店者調査方法 「有機栽培」や「無(減)農薬栽培」といった用語につ いては,消費者の食料の安全への意識の高まりに対応し 調査は,来店者が買い物を終え店を出る際に質問紙を て,一時,農産物生産者や流通業者により濫用されたこ 配布し,家で回答後,後日,店内にある回収箱の中に入 とがある。 れてもらうこととした。複数人で来店している場合に そこで農林水産省は安全な農産物の生産や流通を円滑 は,野菜購入に対し主に決定権を持っている者に,回答 化すると同時に,消費者の混乱を防止するため,19 9 2年 を依頼した。 に特別栽培農産物に係る表示ガイドラインを定めた(そ 調査項目は回答者の年齢,性別,職業,生協への来店 の後,何度か改正されている)。これを受けて各県など 頻度,野菜の購入状況,野菜購入時における重視事項, で,特別栽培農産物認証制度が設けられている。さら 通常の野菜と比較した安全性の高い野菜の価格増加限 に,1 9 9 9年には,持続農業法に基づくエコファーマー認 度,産直野菜ブランドの認識度,産直野菜の詳細内容の 定制度や,J AS法を改正し,J AS(日本農林規格)にお 認知度,イメージや購入後の感想についてである。 ける有機農産物等の検査認証制度が設けられている。 調査店舗は,みやぎ生協の担当者と相談の上,生協来 本研究では,みやぎ生協が「宮城県有機農産物表示認 店者の平均的な姿が取れるよう,仙台市街地にあり一日 証制度」(1 9 9 9年制定)に基づいて「産直野菜」というブ あたりの来客者数が平均 1 3 0 0人で産直野菜の売上げが ランド名で販売している野菜を取り上げ,来店者を対象 良い八木山店と一日あたりの来客者数が平均 2 0 0 0人の に消費者の安全性の高い野菜購入に対する意識を調べる 割には売上げが少ない幸町店にした。 こととした。 配布は 2 0 0 3年 1 2月の平日,午前・午後・夕方とわけ ここでみやぎ生協「 産直野菜」 を選定したのは,東北一 て両店舗ともに 3 0 0枚ずつ,合計 6 0 0枚配布した。回収 の消費地である仙台を拠点としていることと,生協が食 枚数は 2 2 0枚であり回収率は 3 7 %であった。 料の安全性や消費者と生産者との交流に対し積極的であ るからである。 弘大農生報 No.9:91 -96,2006 藤 崎 9 2 男 8% 週1回未満 4% 週1∼2回 34% 女 週3回以上 62% 92% 図4.回答者の生協への来店頻度 図1.回答者の性別 無回答 1% 39歳以下 14% 70歳以上 15% 1∼3割 12% 40-49歳 22% 60-69歳 23% 4∼6割 19% 7∼9割 35% 50-59歳 26% 図2.回答者の年齢 学生 1% ほとんど全て 34% 図5.回答者の生協での野菜購入比率 自営業 5% 勤め 5% 年金・無職 18% パート 16% 農薬使用状況 肥料使用状況 生産地 栽培方法 価格 主婦 55% 新鮮さ 見ばえ 図3.回答者の職業 0 Ⅳ 回答者の概要 回答者は女性が 9割を占め,年齢では,5 0歳代を中心 20 40 とても重視 やや重視 全く 重視しない 無回答 60 80 100 % あまり 重視しない 図6.野菜購入時の重視事項 に幅広い年代からの回答を得た(図 1 ,図 2 ) 。 職業は図 3に示すように「主婦」が 5 5 %で,次いで 「年金生活・無職」1 8 %, 「パート」1 6 %と続いている。 図 4が生協への来店頻度で,6 2 %が「週 3回以上」で あるのに対し, 「週 1回未満」は 4 %とわずかで,多くの 「とても重視する」,「やや重視する」,「あまり重視しな 回答者が頻繁に来店している。 い」,「全く重視しない」の 4段階で問うた結果が図 6で そして野菜を生協で購入する比率については,図 5の ある。 ように「7~ 9割」と「ほとんど全て」がともに 3分の 「とても重視する」が最も多いのが「新鮮さ」8 0 %で, 1程度となっており,多くの回答者が野菜を主に生協で 次いで「生産地」5 5 %,「農薬使用状況」3 7 %,「価格」 購入している。 3 5 %,「肥料使用状況」27 %という順になり,これらの 項目については「やや重視」というものを加えると,い Ⅴ 野菜購入時の重視事項 ずれも 9割の回答者が重視する事項となっている。 逆に「あまり重視しない」が多かったのが「見ばえ」 野菜購入時に重視することを 「農薬使用状況」 「肥料使 4 6 %と「栽培方法」3 9 %で,この 2つの項目については, 用状況」 「生産地」 「 (露地栽培やハウス栽培といった)栽 重視するものとあまり重視しない者とが相半ばしてい 培方法」 「価格」 「新鮮さ」 「見ばえ」の 7項目について る。 安全性の高い野菜に対する消費者意識 93 安全優先 28% その他 37% 安全×価格 10% 図 9 「産直野菜」の値札 価格優先 25% 図7.安全と価格の重視状況による回答者区分 39歳以下 40−49歳 50−59歳 60−69歳 図 10 生産者の説明板 70歳以上 0 20 40 60 80 安全優先 安全×価格 価格優先 その他 100 % 図8.安全と価格の重視状況と回答者の年齢 る産直」という帯がコーナー脇の棚の上から下へ掛けら れていて,値札も「顔と暮らしの見える産直品」(図 9 ) と 書かれた専用のものが用いられている。また,棚に上に は,図 1 0のような生産者の顔写真とメッセージが記載 された説明板が掲示され,看板には「メンバーのみなさ んへのお約束」として,「①おいしい野菜の味を知ってい る私たちは,取りたての一番おいしい野菜をお届けしま す。②安全な野菜作りを知っている私たちは,農薬を使 ここで安全と価格の重視状況を比較して回答者を区分 わない努力をし産直の精神を守ります。③産直活動をす する。「農薬使用状況」が「とても重視」で「価格」が る私たちは,メンバーのみなさんの要望する野菜作りに 「やや重視」以下の者を「安全優先」と呼び,逆に「価 はげみます。」と書かれている。 格」が「とても重視」で「農薬使用状況」が「やや重視」 まず,回答者の「産直野菜」ブランドに対する認知度 以下の者を「価格優先」 ,そして「農薬使用状況」「価格」 を見てみると,図 1 1のように認知度は高く,8 3 %は産 共に「とても重視」する者を「安全 × 価格」と呼んで区 直野菜が安全を売り物にしたブランドだと知っていて, 分すると,図 7のように「安全優先」が 2 8 %, 「安全 × 「名前も知らない」というものはごくわずかだった。 価格」1 0 %, 「価格優先」が 2 6 %で,安全も価格もとも また,図 1 2は,生協で買う野菜のうち産直野菜の購入 に「とても重視」ではない「その他」が 3 7 %となった。 比率を示したものである。「ほとんど全て」というものは この安全と価格の重視状況と回答者の年齢との関係を 6 %で,「7~ 9割」と「4~ 6割」がそれぞれ 3分の 1で 見たのが図 8である。「3 9歳以下」では「安全優先」が あった。 1 6 %に対し「価格優先」6 3 %と多い。逆に「5 0 -5 9歳」 ここで,生協での野菜購入時における「産直野菜」の 「6 0 -6 9歳」 「7 0歳以上」では「安全優先」が 3分の 1 購入比率を,安全と価格の重視状況別に見てみたのが図 を占めるのに対し「価格優先」は 1 5~ 1 8 %にとどまっ 1 3である。 「価格優先」は最も「産直野菜」を購入する ている。若年層では家計に余裕がないことから価格を重 比率が低く,「安全優先」や「安全 × 価格」の方が「産 視するのに対し,高齢層では家計に余裕が生まれると同 直野菜」を購入する比率は高い。しかし「安全優先」 「安 時に健康への関心が高まることからへの安全を重視する 全 × 価格」という考えの回答者であっても,「ほとんど 傾向がうかがえ,野菜購入時の安全と価格への重視状況 全て」「産直野菜」を購入するというものは 1割にとどま は年齢層により異なることがわかる。 り,「7~ 9割」を加えて 5 ,6割に達する状況である。 次に,「産直野菜」について,「ガイドラインに従い栽 Ⅵ 「産直野菜」に対する意識 培(している)」「農薬使用を(慣行の)半分以下に(減 らしている)」「栽培の手間が増(加している)」「出荷で 生協店内での「産直野菜」の販売は,他の野菜と明確 きない作物が増(加している)」「頻繁に抜き打ち検査 (が に区分された一角で行われている。 「顔と暮らしの見え ある)」 「違反すると罰則あり」 「ネットで栽培履歴確認 藤 崎 9 4 名前も知らない 2% 無回答 3% ほとんどない 5% 無回答 名前は 知っている 15% ほとんど全て 6% 7∼9割 31% 1∼3割 22% 安全が売り と知っている 83% 4∼6割 33% 図11.「産直野菜」ブランドの認知度 ガイドラインに従い栽培 安全優先 農薬使用を半分以下に 安全×価格 栽培の手間が増 価格優先 出荷できない作物増 その他 0 図12.生協で野菜購入時における 「産直野菜」の購入比率 頻繁に抜き打ち検査 20 40 60 80 ほとんど全て 7∼9割 4∼6割 1∼3割 ほとんどない 無回答 100 % 図13.安全と価格の重視状況別 生協での野菜購入時における 「産直野菜」の購入比率 違反すると罰則あり ネットで栽培履歴確認可 0 安全優先 いた」 「そんな感じがしていた」 「知らなかった」の 3段 安全×価格 全体に認識度は低く,最も認識度が高い「栽培に手間 が増」 「ネットで栽培履歴確認可」でも「知っていた」は 4割で,「そんな感じがしていた」というものを加える と, 「ガイドラインに従い栽培」 「農薬使用を半分以下 知っていた そんな感じ がしていた 知らなかった 無回答 図14. 「産直野菜」の詳細内容の認識度 可(能) 」という 7項目の詳細な内容について, 「知って 階での認知度を問うた。その結果が図 1 4である。 20 40 60 80 100 % 価格優先 その他 0 20 40 罰則あり」では「知っていた」は 1割を越える程度に過 ぎず, 「知らなかった」ものが 6 ,7割である。 図 1 5はこのうち最も認識度が高い「栽培の手間が増」 について,安全と価格の重視状況別に,その認知度を示 80 100 % 知っていた そんな感じが していた 知らなかった 無回答 に」 「出荷できない作物が増」についても 5 ,6割に達す るようになる。逆に 「頻繁に抜き打ち検査」 「違反すると 60 図15.安全と価格の重視状況別 「産直野菜」の「栽培の手間が増」 に対する認識度 したものである。 「価格優先」は「知っていた」というも のが 3割で認知度が最も低いものの, 「安全優先」「安全 答者は経験がない,という結果だった。 × 価格」でも「知っていた」という回答は 5割程度で, 最後に,「産直野菜」について,普通の野菜と比較して 野菜購入時に安全を「とても重視」しているものが,必 イメージが良いのか悪いのかを,「新鮮さ」「おしいさ」 ずしも野菜の生産状況について,いろいろと情報を収集 「見ばえ」「価格」「虫の危険性」の 5項目について, 「良 しているわけではないことがわかる。 い」 「やや良い」 「変わりない」 「やや悪い」「悪い」の 5 また,栽培履歴が確認可能なことについて認識度は高 段階で問うた。また,「産直野菜」を購入した経験がある かったが,実際に「産直野菜」について栽培履歴を確認 ものについては,購入後の感想についても同様に問う した経験については,図 1 6に示したように,8 8 %の回 た。その結果が図 1 7である。 安全性の高い野菜に対する消費者意識 無回答 3% 生協関係者から 5% 95 上段:イメージ 下段:購入後の感想 生産者から 3% インターネットで 1% 新鮮さ おいしさ 経験無し 88% 見栄え 価格 図16.「産直野菜」の栽培履歴確認経験 虫の危険性 0 「新鮮さ」 「おいしさ」について通常の野菜より優れた イメージを持っている者が多く,5割が「良い」 ,「やや 20 良い やや 良い 悪い 無回答 良い」を加えると 8割のものが「産直野菜」の方を良く 評価していて,悪く評価するものはほとんどない。逆に 40 60 80 100 % 変わら ない やや 悪い 「価格」については「変わらない」が 2割,ごく一部に「や や良い」というものも存在するが, 「やや悪い」というも のが 6割と多くなっている。 「虫の危険性」については, 図17.普通の野菜と比較した 「産直野菜」のイメージ(上段)と 購入後の感想(下段) 半数のものが「変わらない」としているものの,良いに 比べて悪いと評価する者の方が多くなっている。 平均 また,イメージと購入後の感想を見比べると, 「虫の 危険性」や「見ばえ」で「やや悪い」とするものが減少 全体 1.29 している。また「新鮮さ」と「おいしさ」においても 安全優先 1.36 「良い」がやや減少している。イメージというものは, 安全×価格 1.34 良いにしろ悪いにしろ,実際よりもやや大きくとらえら 価格優先 1.19 その他 1.31 れる可能性があるのかもしれない。 0 20 40 60 80 100 % Ⅶ 安全に対する価格増加限度 増加なし 1割増 2割増 3割増 5割増 2倍 3倍 無回答 図 1 8は,安全性の高い野菜に対し,どのくらいまで価 格が増加しても購入するかを示したもので,全体と合わ せて,安全と価格の重視状況別の結果も示してある。 全体では,生産者の希望する 2倍でも購入するのは 5 %にすぎず,5割増でも 2割くらいの回答者にすぎない 図18.安全に対する価格増加限度 ものの,2割増であれば 8割の回答者が購入する。平均 値では 1 . 2 9倍であった。 「安全優先」では他に比べ価格増加限度が高いものの, 逆に,特に若年層を中心とする「価格優先」の者にお それでも価格が 5割増では 3割には達さず,3割増であ いては,価格増への抵抗は強いが,産直野菜を購入しな れば 6割で,平均 1 . 3 6倍であった。逆に, 「価格優先」で いというわけでもなく,産直野菜についての理解が著し は増加なしという回答が 2割存在し,平均 1 . 1 9倍であっ く欠ける,というわけでもなかった。 た。 今回の結果から,これまで以上に,安全な野菜を普及 させようとする場合,販売価格上昇を 1 .2倍程度に抑 Ⅷ おわりに えることが,重要なことの一つと考えられる。このため には,政策的に安全な野菜の生産や販売に対する優遇制 今回の結果から,中高年層を中心とする野菜購入時に 度を導入することや,より低コストで安全な野菜を生産 安全性を重視する者であっても,価格増加への許容幅は するための技術開発の推進が望まれる。 あまり大きくないし, 「産直野菜」 のみを購入するわけで また,安全な野菜生産に対する消費者の理解は,安全 はなく, 「産直野菜」 の詳細内容について詳しく知ってい を気に掛けている者であっても,まだまだ不足気味であ る,というわけでもなかった。 るので,安全な食料を生産するための努力や手間やコス 9 6 藤 崎 ト,さらには,そのことによる地域環境保全への効果等 調査に際し,専攻生渡邊了さんの甚大なる協力を得 について,より一層消費者に伝達していく努力が,望ま た。また,みやぎ生協,産直野菜生産農家の関係者にも れる。 協力を得た。付記して謝意を表する。 Co ns ume r ’ sAt t i t udet o wa r dSaf eVe get abl es − Byanat t i t udes ur ve yo fc us t ome ra tMI YAGI COOP. t owa r di t ss t or ebr and:" SANCHOKUYASAI " (Di r e c t l yDel i ve r e dVege t a bl es ) − Hi r o yukiFUJI SAKI La b o r a t o r yo fRe g i o n a lEn v i r o n me n t a lPl a n n i n g SUMMARY Rec ent l y,t hec ons umer ’ sc onc er nf ors af ef ood r i s es .ButFar m pr oduc t swi t hs af et y ar enotwi del y s pr ead.So,aboutt he" SANCHOKUYASAI " (Di r ec t l y Del i ver ed Veget abl es )whi c hi st hes t or ebr and of MI YAGI COOP.a nd whi c hi s pr oduc ed bas ed on t he s af ef ar m pr oduc t s at t es t at i on s ys t e m,t he c us t omer ' sat t i t udewass ur veyedi nDec ember ,2003. Ast her es ul t ,aquar t erofc us t omer ss ett hes af et y abovet hepr i c ewhen t hey buy veget abl e s ,but f ort hos epeopl ewhomakemuc hoft hes af et y,t hepr i c ei nc r eas el i mi ti sabout1. 3t i mes . l .Fac .Agr i c .& Li f eSc i .Hi r os akiUni v.No.9:91 -96,2006 榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎Bul 弘前大学農学生命科学部 研 究 業 績 目 録 2 0 0 5年10 月-2006年 9月 Li s t sofPubl i s hed Res ear c h Wor ksoft heFac ul t y ofAgr i c ul t ur eand Li f eSc i enc e Hi r os akiUni ver s i t y 2005(Oc t ober ) - 2006(Sept ember ) 弘前大学農学生命科学部 2006年 12月 Fac ul t y ofAgr i c ul t ur eand Li f eSc i enc e Hi r os akiUni ver s i t y Hi r os aki036 -8561,J apan Dec ember ,2006 は し が き 本号の「研究業績目録」には,2005年 10月から 2006年 9月までの業績を掲載しました。 業績の区分は,a -研究論文,b-学術図書,c -その他の著書・訳書,d-学会発表,e -調査・実験報告書, f -その他とし,各自の申請にもとづいています。 各講座の教員組織(2006年 10月 1日現在)は以下の通りですが,研究業績目録は各学科・講座あるいは研 究室でとりまとめて掲載してあります。 生物機能科学科 (生命理学講座) 鮫島正純,黒尾正樹,松岡教理,福澤雅志 (遺伝情報科学講座) 小原良孝,石田幸子,原田竹雄,石川隆二,吉田 渉 (植物エネルギー工学講座) 青山正和,葛西身延,齊藤 寛,大河 浩,千田峰生 応用生命工学科 (生体機能工学講座) 戸羽隆宏,長田恭一,殿内暁夫 (生体情報工学講座) 宮入一夫,橋本 勝,姫野俵太,牛田千里,高田 晃 (細胞工学講座) 片方陽太郎,大町鉄雄,菊池英明,石黒誠一,吉田 孝,畠山幸紀 生物生産科学科 (園芸学講座) 嵯峨紘一,荒川 修,浅田武典,本多和茂,張 樹槐 (農業生産学講座) 鈴木裕之,杉山修一,松山信彦,福地 博,松崎正敏 (環境生物学講座) 佐原雄二,比留間潔,佐野輝男,城田安幸,東 信行,田中和明,藤田 隆 地域環境科学科 (地域環境工学講座) 万木正弘,工藤 明,佐々木長市,萩原 守,泉 完,角野三好,加藤 幸 (地域環境計画学講座) 谷口 建,高橋照夫,桧垣大助,藤崎浩幸 (地域資源経営学講座) 高橋秀直,宇野忠義,神田健策,武田共治,澁谷長生,泉谷眞実 生物共生教育研究センター 澁谷長生(併任),塩崎雄之助,村山成治,伊藤大雄 1 00 研 究 業 績 目 録 目 次 生物機能科学科 ……………………………………………………………………… 101 生命理学講座 ……………………………………………………………………… 101 遺伝情報科学講座 ………………………………………………………………… 102 植物エネルギー工学講座 ………………………………………………………… 103 応用生命工学科 ……………………………………………………………………… 106 生体機能工学講座 ………………………………………………………………… 106 生体情報工学講座 ………………………………………………………………… 107 細胞工学講座 ……………………………………………………………………… 110 生物生産科学科 ……………………………………………………………………… 113 園芸学講座 ………………………………………………………………………… 113 農業生産学講座 …………………………………………………………………… 114 環境生物学講座 …………………………………………………………………… 115 地域環境科学科 ……………………………………………………………………… 119 地域環境工学講座 ………………………………………………………………… 119 地域環境計画学講座 ……………………………………………………………… 120 地域資源経営学講座 ……………………………………………………………… 120 生物共生教育研究センター ………………………………………………………… 122 研 究 業 績 目 録 101 業 績 目 録 生 物 機 能 科 学 科 生 命 理 学 講 座 * * * a 0 1 . SHIRAI,Y. ,N.SASAKI* ,Y.KISHI** ,A.I K.I M.SAMESHIMA,T.KOBAYASHI* ,K.MURAKAMIZUMI , TOH , MUROFUSHI* :Re gul a t i o no fl e ve l so fa c t i nt hr e o ni ne pho s pho r yl a t i o n dur i ng l i f ec yc l e of Ph y s a r u m p o l y c e p h a l u m.Ce l lMo t i l i t ya nd t he Cyt o s ke l e t o n6 3 :7 7 8 7 ,2 0 0 6 . (* De pa r t me nto fBi o l o gy,Oc ha no mi z u Uni ve r s i t y,** I naI ns t i t ut e ,Me di c a la ndBi o l o gi c a lLa bo r a t o r i e sCo .Lt d) a 0 2 . KANZAWA N* ,Y.HOSHINO* ,M.CHIBA* ,D.HOSHINO* ,H.KOBAYASHI* ,N.KAMASAWA** ,Y.KISHI*** ,M. OSUMI** ,M.SAMESHIMA,T.TSUCHIYA* :Cha ngei nt hea c t i nc yt o s ke l e t o ndur i ngs e i s mo na s t i cmo ve me nto f Mi mo s ap u d i c a .Pl a nt Ce l lPhys i o l .4 7 :5 3 1 5 3 9 ,2 0 0 6 . (* Fa c ul t yo f Sc i e nc ea nd Te c hno l o gy,So phi a Uni ve r s i t y,** Fa c ul t yo fSc i e nc e ,J a pa n Wo me n’ s Uni ve r s i t y,*** I na I ns t i t ut e ,Me di c a la nd Bi o l o gi c a l La bo r a t o r i e sCo .Lt d) a 0 3 . YOSHIMURA,A. ,A.NAKATA,M.KUROl e c ul a rc yt o ge ne t i cc ha r a c t e r i z a t i o na nd O,Y.OBARA,Y.ANDO :Mo c hr o mo s o ma l di s t r i but i o n o ft he s a t e l n t he ge no me of Ox y ah y l ai n t r i c a t a (Or t ho pt e r a : l i t e DNA i Ca t a nt o pi da e ) .Cyt o ge ne t .Ge no meRe s . ,1 1 2:1 6 0 1 6 5 ,2 0 0 6 . a 0 4 . 松岡教理:進化学について(2 ) .蒲生野,3 7:7 1 2 (2 0 0 5 ) a 0 5 . 松村行栄・五十嵐聖貴・松岡教理:日本産アゲハチョウ科の分子系統学的研究.弘前大学農学生命科学部学術報 告,8:1 8 ,2 0 0 5 . a 0 6 . MATSUOKA,N.:Ge ne t i cva r i a t i o ni nt wos t a r f i s h,Ac a n t h a s t e rp l a n c ia n dEc h i n a s t e rl u z o n i c u s ,f r o m Oki na wa . Bul l .Fa c .Agr i c .& Li f eSc i .Hi r o s a kiUni v.8:9 1 6 ,2 0 0 5 . d0 1 . 佐々木和也*,嵯峨紘一,鮫島正純:GA3およびウニコナゾール処理がデルフィニウム鉢物の生育に及ぼす影響. 園芸学会平成 1 7年度秋季大会.2 0 0 5 .1 0 .1 3 .仙台(*青森県農林総合研究センターフラワーセンター 2 1 ) d0 2 . 井沼道子・小原良孝・黒尾正樹:ヒメネズミの Che tに関する研究 ― QM 蛍光遅延に伴う染色体 DNAの変化―. 染色体学会 2 0 0 5年度(第 5 6回)年会,2 0 0 5 . d0 3 . 片山 亮・長谷川雅美*・森 哲**・森部絢嗣***・黒尾正樹・小原良孝:mt DNAを指標としたシマヘビの系統 地理(2 ) .日本爬虫両棲類学会第 4 4回大会,2 0 0 5 .(*東邦大・理,**京大・理,***名大・生命農学) d0 4 . 佐々木和也*,嵯峨紘一,鮫島正純:夏季および冬季における GA3処理がシネンシス系デルフィニウムの生育に 及ぼす影響.園芸学会東北支部平成 1 8年度大会.2 0 0 6 .8 .2 2 2 3 .八戸(*青森県農林総合研究センターフラワー センター 2 1 ) d0 5 . 佐々木和也*,嵯峨紘一,鮫島正純:日長及び温度がシネンシス系デルフィニウムの抽台に及ぼす影響.園芸学 会平成 1 8年度秋季大会.2 0 0 6 .9 .2 3 2 5 .長崎(*青森県農林総合研究センターフラワーセンター 2 1 ) d0 6 . 高畑恵介,柴田大輔,福澤雅志,鮫島正純:細胞性粘菌胞子に出現する G-アクチンを含む構造の解析.日本植 物形態学会第 1 8回大会.2 0 0 6 .9 .1 3 .熊本 d0 7 . 田中浩平,夏目茉莉子,鮫島正純,福澤雅志:細胞性粘菌での発生におけるサイトカイニンの役割について.日 本植物形態学会第 1 8回大会.2 0 0 6 .9 .1 3 .熊本 d0 8 . 阿部広和,鮫島正純,福澤雅志:細胞性粘菌の 1 Rmyb解析について.日本植物形態学会第 1 8回大会.2 0 0 6 .9 . 1 3 .熊本 d0 9 . 江田真毅*,**・黒尾正樹・長谷川博***・樋口広芳****・小池裕子**:ミトコンドリア DNAの重複を考慮したア ホウドリの集団構造の再検討.日本鳥学会 2 0 0 6年度大会,2 0 0 6 .(*日本学術振興会 特別研究員,**九大・比 文,***東邦大・理,****東大・農) f 0 1 . KATAYAMA,A. ,M.HASEGAWA* ,A.MORI** ,J .MORIBE*** ,M.KUROyl o ge o gr a phy o f O,Y.OBARA :Ph El a p h eq u a d r i v i r g a t ao nt he ba s i so fmt DNA(2 ) .Bul l .He r pe t o l .So c .J pn,2 0 0 6 :6 3 ,2 0 0 6 . (* Fa c ul t yo f Sc i e nc e ,To c ul t yo fSc i e nc e ,Kyo t o Uni ve r s i t y,*** Gr a dua t e Sc ho o lo fBi o a gr i c ul t ur a l ho Uni ve r s i t y,** Fa Sc i e nc e ,Na go yaUni ve r s i t y) f 0 2 . I ,Y.OBARA,M.KUROus a t i o no fde l a ye d QMf l uo r e s c e nc ei nt he Cbl o c ks o fAp o d e mu s NUMA,M. O :Ca a r g e n t e u s- St r uc t ur a lc ha nge so fChe t e r o c hr o ma t i cDNA.Chr o mo s o meSc i . , (i npr e s s ) . 研 究 業 績 目 録 1 02 遺伝情報科学講座 * a 0 1 . YOSHIMURA,A. ,A.NAKATA,M.KUROl e c ul a rc yt o ge ne t i cc ha r a c t e r i z a t i o na nd O,Y.OBARA,Y.ANDO ; Mo c hr o mo s o ma l di s t r i but i o n o ft he s a t e l l i t e DNA i n t he ge no me o f Ox y ah y l ai n t r i c a t a (Or t ho pt e r a : Ca t a nt o pi da e ) .Cyt o ge ne t .Ge no meRe s . ,1 1 2:1 6 0 1 6 5 ,2 0 0 6 . (*弘前大学名誉教授) a 0 2 . WAKASA Y* ,H KUDO ,R I SAKADA* ,M SENDA* ,M NIIZEKI,T HARADA :Lo we xpr e s s i o no fa n SHI KAWA, e ndo po l yga l a c t ur o na s e ge ne i na ppl ef r ui t wi t hl o ngt e r m s t o r a ge po t e nt i a l .Po s t ha r ve s t Bi o l o gy and Tec AS,** Ge neRe s e a r c hCe nt e r ,Hi r o r a kiUni v. ) hno l o gy3 9:1 9 3 1 9 8 .2 0 0 6 (* NI * a 0 3 . SAKAMOTO S. ,R.I .NAKAMURA** ,YI .SATO*** ,S.SHIMAMOTO**** :Spe c i e si de nt i f i c a t i o no f SHI KAWA,I 6 0 0 0 ye a r s o l dbe a nsf r o m Sa nna i Ma r uya mas i t e ,Ao mo r i ,J a pa n.J o ur na lo ff o s s i lRe s e a r c h3 9:1 6 ,2 0 0 6 . ( *太 ** *** **** 子食品 , 千葉大学大学院 , 総合地球環境学研究所 , 北海道大学名誉教授) a 0 4 .I , S. YAMANAKA* , Y. FUKUTA** , S. CHITRAKON*** , C. BOUNPHANOUSAY**** , K. SHI KAWA, R. KANYAVONG*** ,LH.TANG**** ,I .NAKAMURA***** ,T.SATO****** ,YI .SATO******* :Ge ne t i ce r o s i o n f r o m mo de r n va r i e t i e si nt ot r a di t i o na lupl a nd r y z as a t i v aL. )i n no r t he r n Tha i l a nd.Ge ne t . i c ec ul t i va r s (Or Re s o ur .Cr o pEvo l .5 3:2 4 5 2 5 2 ,2 0 0 6 . (*独立行政法人生物資源研究所,**独立行政法人国際農林水産業研究セ ンター,***元タイ王国パトンタニ試験場イネ研究所,***ラオス国立農業研究センター,****中国江蘇省農業 科学院,*****千葉大学大学院,******東北大学大学院,*******総合地球環境学研究所) d0 1 . 長井和哉・川上和人*・黒尾正樹・小原良孝:アホウドリ類における分子的性判定法.平成 1 7年度日本動物学会 東北支部大会(弘前大学)2 0 0 5 . (*森林総研・多摩森林科学園) d0 2 . 井沼道子・小原良孝・黒尾正樹:ヒメネズミの C-he tに関する研究 ― QM 蛍光遅延に伴う染色体 DNAの変 化.2 0 0 5年度(第 5 6回)染色体学会年会(弘前大学)2 0 0 5 . d0 3 . 中田章史・吉村 文・吉田光明*・黒尾正樹・小原良孝:ヒミズから単離された SI NE様配列の構造と染色体上で の分布について.2 0 0 5年度(第 5 6回)染色体学会年会(弘前大学)2 0 0 5 .(*放射線医学総合研究所・緊急被爆 医療研究センター) d0 4 .I S. ,M.FUKUSHIMA,F.TATEBAYASHI,M.MORIKAWA,M.YAMAZAKI,K.HANATA,H.ORII* ,W.YOSHIDA : SHI DA, The i nt e s t i na lc e l l s ha ve pl ur i po t e nc y i n Ps e u d o s t y l o c h u si n t e r me d i u s (Po l yc l a di da ) . 10 t h I nt e r na t i o na l Sympo s i um o nFl a t wo r m Bi o l o gy.2 0 06 a dua t es c ho o lo fLi f eSc i e nc e ,Uni ve r s i t yo f .I nns br uc k,Aus t r i a . (* Gr Hyo go ) d0 5 . NISHITANI,S. ,S.I W.YOSHIDA,W.TESHIROGI:Po l ypl o i dsa ndc hr o mo s o ma le vo l ut i o ni nt hef r e s hwa t e r SHI DA, pl a na r i a n,S e i d l i aa u r i c u l a t a .1 0 t hI nt e r na t i o na lSympo s i um o nFl a t wo r m Bi o l o gy.2 0 0 6 .I nns br uc k,Aus t r i a . d0 6 . SHOJIMA,K. ,K.YUKITA,S.I Thedi f f e r e nt i a t i o npr o c e s so ft hepr o t o ne phr i di um f r o ms pe c i f i ca l ka l i ne SHI DA : pho s pha t a s ee xpr e s s i ve c e l l s dur i ng e mbr yo ge ne s i so ft he po l yc l a d Ps e u d o s t y l o c h u si n t e r me d i u s .1 0 t h I nt e r na t i o na lSympo s i um o nFl a t wo r m Bi o l o gy.2 0 0 6 .I nns br uc k,Aus t r i a . d0 7 . YOSHIDA,W. ,H.YAMAYA,M.OWADA,Y.SHINOZAKI,S.I pr e s s i o n pa t t e r no fa nt e r i o rHOM/ HOX SHI DA :Ex ge ne s dur i ng e mbr yo ge ne s i si n po l yc l a ds .1 0 t hI nt e r na t i o na lSympo s i um o n Fl a t wo r m Bi o l o gy.2 0 0 6 . I nns br uc k,Aus t r i a . d0 8 . 福島 誠・橋爪 力*・吉田 渉・石田幸子:淡水棲プラナリアにおけるテストステロン検出と生殖器官の発達度 合による量的変動.日本動物学会第 7 7回大会(島根大学),2 0 0 6 .(*岩手大・農学) d0 9 . 山野弘幸・高久未央・藤井 武・中村哲郎・吉田 渉・石田幸子:ナツドマリヒラムシの生殖細胞発達過程での v a s a ,n a n o sホモログの発現.日本動物学会第 7 7回大会(島根大学),2 0 0 6 . d1 0 . 王 愛徳・原田竹雄 リンゴ‘ふじ’と芽条変異体‘ひろさきふじ’の果実硬度低下速度の違いに関する分子機 構の解明. 「見てみて,聞いてみて,触ってみて,弘前大学」へのポスター発表 2 0 0 6年 9月 d1 1 . 工藤久幸・千田峰生*・石川隆二・赤田辰治*・原田竹雄 Meトマトを台木にしたジャガイモ穂木葉の形質転換 東北育種談話会(山形大学)2 0 0 6年 8月(*弘前大学遺伝子実験施設) d1 0 0 6年 9月 2 . 原田竹雄 リンゴの科学: ‘ふじ’の優良遺伝子とは? 弘前大学シニアサマーカレッジ 2 d1 3 . WAKASA Y,WANG A,HARADA T.2 0 0 6 The e xpr e s s i o na na l ys i so fr i pe ni ngr e l a t e d ge ne sa mo ng a ppl e c ul t i va r se xhi bi t i ng de f f e r e nt f r ui ts t o r a ge pt e nt i a l .The Se c o nd I nt e r na t i o na l Sympo s i um o n Appl e Pr o duc t i o n.Ya nTa i ,Chi na d1 4 . 工藤久幸・千田峰生*・石川隆二・赤田辰治*・新関 稔**・原田竹雄:Meトマトを台木にしたジャガイモ穂木 葉の形質転換 育種学研究 8 (別 1 (東京農工大) ) 2 0 0 6年 3月(*弘前大学遺伝子実験施設,**弘前大学名誉教授) 研 究 業 績 目 録 103 d1 5 . 原田竹雄:リンゴについての遺伝子研究.平成 1 7年度東北農業試験研究推進会議・生物工学推進部会.重点検討 事項の検討『今後の生物工学研究の展開方向』 盛岡市 2 0 0 6年 2月 d1 6 . 原田竹雄:リンゴの遺伝子研究.農学生命科学部公開講座『りんごを科学する』.板柳町 2 0 0 5年 1 2月 * ** *** d1 7 .I , K.I ,A.KAWASAKI :As s o c i a t i o n ma ppi ng t oc ha r a c t e r i z e SHI KAWA ,R. MAI ,N.TOKI ,T.AOKI di a gno s t i cma r ke r sf o ri ndi c a j a po ni c adi f f e r e nt i a t i o n.Fi f t hRi c eGe ne t i cSympo j i um. (Phi l i ppi ne s )2 0 0 6 . (*岩 手大学連合大学院農学研究科,**独立行政法人生物資源研究所,***栃木県小山市) * d1 8 .I ,K. ,Y.CHIBA,M.MURAI** ,R.I ppi ng o fUn d u l a t e dr a c h i s 1o r i gi na t e df r o m mut a bl e MAI SHI KAWA :Ma s t r a i n,Aka ge ,o namo l e c ul a rma p.Fi f t hRi c eGe ne t i cSympo j i um. (Phi l i ppi ne s )2 0 0 6 . (*岩手大学連合大学院 農学研究科,**高知大学) d1 9 . 石川隆二・今井克則*・川崎顕子・信田清夏**・千葉悠貴・本間照久・千田峰生***・赤田辰治***・原田竹雄・ 新関 稔****:イネ連鎖不平衡マッピングの応用 V.集団構造の解析と系統分化に関連した候補遺伝子座の特 定.日本育種学会第 1 0 9回講演会(東京農工大)2 0 0 6 .(* 岩手大学連合大学院農学研究科,** 千葉大学大学 院,***弘大遺伝子実験施設,****弘前大学名誉教授) d2 0 . 石川隆二・千葉悠貴・川崎顕子・今井克則*・小林伸哉**・福田善通***:インド型品種,I R2 4派生系統から得ら れた易変異性系統の解析:根の異常性を呈する新規変異体.日本育種学会第 1 1 0回講演会(愛媛大)2 0 0 6 . ( *岩 手大学連合大学院農学研究科,** I RRI ,***独立行政法人国際農林水産業研究センター) d2 1 . 今井克則*・千葉あや乃・千葉悠貴・高牟禮逸朗**・村井正之***・千田峰生****・赤田辰治****.原田竹雄・ 石川隆二:イネ枝梗湾曲- 1遺伝子,Ur 1の分子選抜マーカーの特定.日本育種学会第 1 1 0回講演会(愛媛大) 2 0 0 6 . (*岩手大学連合大学院農学研究科,**北大院農,***高知大農,****弘大遺伝子実験施設) d2 2 . 千葉悠貴・石川隆二・今井克則*・千葉あや乃・高牟禮逸朗**:赤毛自殖後代で生じた新規突然変異体の遺伝解 析.日本育種学会第 1 1 0回講演会(愛媛大)2 0 0 6 .(*岩手大学連合大学院農学研究科,**北大院農) d2 3 . 本間照久・波多野佑一・千田峰生*・赤田辰治*.原田竹雄・石川隆二:日印交雑で生じた易変異系統の特性解 析:クロリナ変異とキメラ制御遺伝子の解析.日本育種学会第 1 1 0回講演会(愛媛大)2 0 0 6 .(*弘大遺伝子実験 施設) d2 4 . 石川隆二:日本文化と農耕の起源―三内丸山遺跡にみる縄文農耕の可能性.農学生命科学部 5 0周年記念地域支 援事業公開シンポジウム.2 0 0 6 (弘前) e 0 1 . 小原良孝(哺乳類担当)青森県外来種対策学術調査報告書 ―青森県外来種リスト―.青森県 2 0 0 6 . e 0 2 . 小原良孝(哺乳類担当)青森県の希少な野生生物 ―青森県レッドリスト (2 0 0 6年 改訂増補版)―.青森県 2 0 0 6 . e 0 3 . SATO,YI ,R.I H,TANG :I I I .RI CE Si kki m(I ndi a ) ,no r t hwe s tVi e t na m,Phi l i ppi ne s ,Tha i La o s SHI KAWA,LCa mbo di a ,Luc kno w(I ndi a ) , (Pr o j e c tl e a de r .K.Ta ke da ) ,i nGe ne t i ca s s a ya nds t udyo fc r o pge r mpl a s mi n a nda r o undChi na (3rd) ,A r e po r to fGr a nt i nAi df o rSc i e nt i f i cRe s e a r c h(A) (2 (2 ) 0 0 3 2 0 0 5 ) ,pp.5 2 0 ,2 0 0 6 . f 0 1 . 小原良孝:野生動物学特論.東京農業大学農学特別講義.(東京農大),2 0 0 5 . f 0 2 . 石川隆二 縄文農耕を考える(対談集) .縄文ファイル 1 3 2 ,pp3 7 ,2 0 0 6 ,NPO法人三内丸山縄文発信の会,青森. f 0 3 . 石川隆二 三内丸山遺跡にみられる植物の利用~ヒトは食べられる植物を選抜していたのか~ 三内丸山から縄 文ムーブメント.隔月間あもり草子,pp.1 2 1 4 ,2 0 0 6年 1 0月 1日発行,企画集団ぷりずむ,青森. 植物エネルギー工学講座 a 0 1 . 青山正和:有機成分,部門別進歩総説 第 2部門 土壌化学・土壌鉱物,日本土壌肥料学雑誌,7 6:6 9 8 7 0 1 ,2 0 0 5 . a 0 2 . AOYAMA,M. ,B.ZHOU* ,M.SAITOH** a nd N.YAMAGUCHI** :Mi c r o bi a lbi o ma s si ns o i l s wi t hc a l c i um a c c umul a t i o na s s o c i a t e dwi t ht hea ppl i c a t i o no fc o mpo s t e dl i me t r e a t e ds e wa ges l udge .S o i lS c i e n c ea n dPl a n t Nu t r i t i o n5 2:1 7 7 1 8 5 ,2 0 0 6 . (* So i la nd Fe r t i l i z e rI ns t i t ut e ,He i l o ngj i a ng Ac a de my o fAgr i c ul t ur a lSc i e nc e s , ** Ao mo r iPr e f e c t ur a lAgr i c ul t ur ea ndFo r e s t r yRe s e a r c hCe nt e r ) a 0 3 . AOYAMA,M.:Pr o pe r t i e so fne ut r a lpho s pha t ebuf f e re xt r a c t a bl eo r ga ni cma t t e ri ns o i l sr e ve a l e dus i ngs i z e e xc l us i o nc hr o ma t o gr a phya ndf r a c t i o na t i o nwi t hpo l yvi nyl pyr r o l i do ne .S o i lS c i e n c ea n dPl a n tNu t r i t i o n5 2:3 7 8 3 8 6 ,20 0 6 . *1 a 0 4 . MATSUMURA,H. ,S.WATANABE*2,K.HARADA*2,M.SENDA,S.AKADA*3,S.KAWASAKI*4,E.DOUBOUZET*5, N.MINAKA*6 a nd R.TAKAHASHI*5 :Mo l e c ul a rl i nka ge ma ppi ng a nd phyl o ge ny o ft he c ha l c o ne s ynt ha s e mul t i ge nef a mi l yi ns o ybe a n.Th e o r e t i c a la n dAp p l i e dGe n e t i c s1 1 0:1 2 0 3 1 2 0 9 ,2 0 0 5 . (*1Uni ve r s i t yo fTs ukuba , *2 Chi ba Uni ve r s i t y,*3 Ge ne Re s e a r c h Ce nt e r ,Hi r o s a kiUni ve r s i t y,*4 Na t i o na lI ns t i t ut eo fAgr o bi o l o gi c a l 研 究 業 績 目 録 1 04 Sc i e nc e s ,*5Na t i o na lI ns t i t ut eo fCr o pSc i e nc e ,*6Na t i o na lI ns t i t ut eo fAgr o Envi r o nme nt a lSc i e nc e s ) a 0 5 . WAKASA,Y. ,H.KUDO,R.I S.AKADA* ,M.SENDA,M.NIIZEKIa ndT.HARADA :Lo we xpr e s s i o no fa n SHI KAWA, e ndo po l yga l a c t ur o na s ege nei na ppl ef r ui twi t hl o ngt e r ms t o r a gepo t e nt i a l .Po s t h a r v e s tBi o l o g ya n dT e c h n o l o g y 3 9:1 9 3 1 9 8 ,2 0 0 6 . (* Ge neRe s e a r c hCe nt e r ,Hi r o s a kiUni ve r s i t y) * a 0 6 . OSADA,K. ,T.SUZUKI,Y.KAWAKAMI,M.SENDA,A.KASAI,M.SAMI* ,Y.OHTA* ,T.KANDA* a ndM.I KEDA : Do s e de pe nde nthypo c ho l e s t e r o l e mi ca c t i o nso fdi e t a r ya ppl epo l yphe no li nr a t sf e dc ho l e s t e r o l .Li p i d s4 1: 1 3 3 1 3 9 ,2 0 0 6 . (* As a hiBr e we r i e s ) a 0 7 . 山本英樹*・千田峰生:秋田県のキクにおけるキク微斑ウイルスの発生状況.北日本病虫研報 5 6:7 9 8 0 ,2 0 0 5 . (*秋田農試) a 0 8 . KEREN,N* . ,H.OHKAWA,E. A.WELSH** ,M.LIBERTON** a nd H. B.PAKRASI** :Ps b2 9 ,ac o ns e r ve d2 2 kD pr o t e i n,f unc t i o nsi nt hebi o ge ne s i so fPho t o s ys t e mI Ic o mpl e xe si nSyne c ho c ys t i sa ndAr a bi do ps i s .Pl a n tCe l l 1 7:2 7 6 8 2 7 8 1 ,2 br e w Uni ve r s i t yo fJ e r us a l e m,** Wa s hi ngt o nUni ve r s i t y) 0 0 5 . (* He *1 a 0 9 . KAHLON,S. ,K.BEERI*1,H.OHKAWA,Y.HIHARA*2,O.MURIK*1,I .SUZUKI*3,T.OGAWA*4 a ndA.KAPLAN*1 : A put a t i ve s e ns o r ki na s e ,Hi k3 1 ,i si nvo l ve di n gl uc o s es e ns i ng i n Syne c ho c ys t i ss p.s t r a i n PCC6 8 0 3 . *2 *3 Mi c r o b i o l o g y1 5 2 :6 4 7 6 5 5 ,2 0 0 6 . (*1 He br e w Uni ve r s i t yo fJ e r us a l e m, Sa i t a ma Uni ve r s i t y, Na t i o na l *4 I ns t i t ut ef o rBa s i cBi o l o gy, Na go yaUni ve r s i t y) b0 1 . 青山正和:高速サイズ排除クロマトグラフィーによる堆肥腐熟度の推定,東北の農業と土壌肥料,1 8 9 -1 9 0頁, 日本土壌肥料学会東北支部,2 0 0 6 . c 8 6 -1 9 1頁,農文協,2 0 0 6 . 0 1 . 青山正和:堆肥と腐植,堆肥とことん活用読本,1 c 0 2 . 千田峰生・増田 税*:ダイズ種子の着色メカニズムに迫る! 黄ダイズの種皮着色抑制に転写後型ジーンサイレ ンシングが関与.化学と生物 4 3:2 2 0 2 2 1 ,2 0 0 5 . (*北海道大学大学院農学研究科) c 0 3 . 千田峰生:ウイルスが大豆の上に絵を描く.RNANe t wo r kNe ws l e t t e r4:2 8 3 0 ,2 0 0 6 . d0 1 . 青山正和・村山美穂:カラシナによる土壌からの銅と鉛の吸収に及ぼす腐植物質の影響,日本腐植物質学会第 2 1 回講演会講演要旨集,1 2 1 3 ,2 0 0 5 . d0 2 . 青山正和:バイオス醗酵過程における腐植物質の生成機構と農業生産性向上,日本腐植物質学会第 2 1回講演会 講演要旨集,1 8 1 9 ,2 0 0 5 . d0 3 . 青山正和・山谷すみれ:土壌微生物による植物遺体の分解過程で生成する水溶性タンパク質の性状,日本土壌肥 料学会講演要旨集 5 2:2 7 ,2 0 0 6 . d0 4 . 高木陽子・青山正和:リンゴ園土壌表層への堆肥施用が土壌微生物バイオマスに及ぼす影響,日本土壌肥料学会 講演要旨集 5 2:2 9 ,2 0 0 6 . d0 5 . 保嶋敬子・青山正和:乾燥リンゴ搾り粕の土壌改良資材としての利用―植物生育ならびに土壌の物理性と生物性 に及ぼす影響―,日本土壌肥料学会講演要旨集 5 2:1 5 9 ,2 0 0 6 . d0 6 . 福田悠希・葛西身延:ダイズのソース・シンク比と光合成の制御の関係.第 7 0回日本植物学会,2 0 0 6 . d0 7 . 小口祐輔・葛西身延:ダイズのシンク・リミット状態における光合成とプロトンポンプ活性.第 7 0回日本植物学 会,2 0 0 6 . d0 8 . 千田峰生・葛西宏介 *・葛西厚史:黄ダイズの種皮着色をつかさどる I遺伝子の構造的特徴.日本育種学会第 1 0 9 回講演会(東京農工大学)2 0 0 6 . (*弘前大学医学部保健学科) d0 9 . 葛西厚史・赤田辰治*・石川隆二・原田竹雄・新関 稔・千田峰生:I遺伝子転写産物は黄ダイズ種皮において CHS遺伝子の 2本鎖 RNAを形成するか? 日本育種学会第 1 0 9回講演会(東京農工大学)2 0 0 6 .(*弘前大学遺 伝子実験施設) d1 0 . 大久保喜光・葛西厚史・千田峰生:トランスジェニックアラビドプシスを用いたダイズ種皮着色抑制遺伝子(I遺 伝子)の組織発現に関する研究.東北育種研究集会(山形大学)2 0 0 6 . d1 1 . 徳富(宮尾)光恵*,大河 浩,谷口 洋二郎*,深山 浩*:ゲノムから見た炭素代謝関連遺伝子.日本植物学 会第 7 0回大会シンポジウム,2 0 0 6 . (*農業生物資源研究所) e 0 1 . 青山正和:環境修復への利用をめざした腐植物質のキャラクタリゼーション,平成 1 5年度~平成 1 7年度科学研 究費補助金(基盤研究 C)研究成果報告書,1 -6 0頁,2 0 0 6 . e 0 2 . 葛西身延:光合成物質生産の新規制御物質の分離同定とソース・シンク制御機構の総合的研究.平成 1 6~ 1 7年 度科学研究費補助金研究成果報告書.2 0 0 6 . f 0 1 . 青山正和:りんご剪定枝の堆肥利用の課題,第 1回未利用バイオマスセミナー「未利用バイオマスとしてのりんご 剪定枝の利用―その課題と地域内連携―」 ,弘前大学農学生命科学部付属未利用バイオマス研究センター,2 0 0 6 . 研 究 業 績 目 録 105 f 0 2 . 青山正和:リンゴ搾り粕の有機質土壌改良資材としての利用,第 2回未利用バイオマスセミナー「バイオマスと してのリンゴ絞り粕の活用方策―その課題と地域内連携―」 ,弘前大学農学生命科学部付属未利用バイオマス研 究センター,2 0 0 6 . f 0 3 . FUKUDA,Y.a nd KASAI,M.:Thee f f e c to fpo dr e mo va lo n pho t o s ynt he s i si ns o ybe a n pl a nt s .J .Pl a ntRe s . Suppl .:9 8 ,2 0 0 5 . f 0 4 . 千田峰生:ダイズ種皮着色抑制遺伝子によるアントシアニン生合成の抑制機構.ミヤコグサ・ダイズワークショッ プ(理化学研究所)2 0 0 5 . f 0 5 . 千田峰生:黄大豆の種皮着色抑制にはたす e ndo ge no usRNA s i l e nc i ngの役割について.山形県立園芸試験場バ イオ育種部セミナー 2 0 0 5 . f 0 6 . 千田峰生:ウイルスが大豆の上で RNAiをさけぶ ―大豆が黄色い理由について― 弘前大学遺伝子実験施設竣 工1 0周年記念シンポジウム(弘前大学創立 5 0周年記念会館)2 0 0 6 . f 0 7 . 姫野俵太・武藤あきら・牛田千里・佐野輝男・原田竹雄・鬼島 宏 *・種田晃人**・千田峰生:機能性 RNAを基 盤とする新技術開発のための基礎研究とその展開. 「見てみて,聞いてみて,触ってみて,弘前大学」 (弘前シ ティホテル)2 0 0 6 . (*弘前大学医学部,**弘前大学理工学部) 研 究 業 績 目 録 1 06 応 用 生 命 工 学 科 生体機能工学講座 食品安全学研究室 a 0 1 . HORIE,M. ,MURAKAMI,T. ,SATO,T. ,TARUSAWA,Y. ,NAKAMURA,S. ,TOBA,T.:Ana e r o bi ci nduc t i o no f a dhe r e nc et ol a mi ni ni nLa c t o b a c i l l u sg a s s e r is t r a i nsbyc o nt a c twi t hs o l i ds ur f a c e .Cu r r .Mi c r o b i o l .5 1:2 7 5 2 8 2 , 2 0 0 5 . b0 1 . 戸羽隆宏:乳酸菌発酵食品の製造と生理機能.齋藤忠夫・西村敏英・松田 幹編 最新畜産物利用学 5 5 -6 6頁, 朝倉書店,2 0 0 6 . d0 1 . 村上恵光・高杉美波・青木裕史 1)・片方陽太郎・佐藤 征 2)・大友良光 2)・戸羽隆宏:乳酸菌による皮膚有害菌の 付着阻止.平成 1 8年度日本酪農科学シンポジウム要旨.p.1 9 ,2 0 0 6 . (1) 昭和電工(株)研究開発センター,2) 本 学医学部保健学科) 食品栄養化学研究室 a 0 1 . OSADA K,SUZUKI T,KAWAKAMI Y,SENDA M,KASAI A,SAMI M,OHTA Y,KANDA T,I s e KEDA M.:Do de pe nde nthypo c ho l e s t e r o l e mi ca c t i o nso fdi e t a r ya ppl epo l yphe no li nr a t sf e dc ho l e s t e r o l .Li pi dsVol .4 1: 1 3 3 1 3 9 ,2 0 0 6 . a 0 2 . OSADA K,FUNAYAMA M,FUCHIS,SAMIM,OHTA Y,KANDA T,I f e c t so fdi e t a r y pr o c ya ni di ns KEDA M.:Ef a ndt e apo l yphe no l so na di po s et i s s uema s sa ndf a t t ya c i dme t a bo l i s mi nr a t so nahi ghf a tdi e t .J o ur na lo f Ol e oSc i e nc eVol .5 5:7 9 8 9 ,2 0 0 6 . a 0 3 . OHTA Y,SAMIM,KANDA T,SAITO K,OSADA K,KATO H.:Ge nee xpr e s s i o na na l ys i so ft hea nt i o be s i t ye f f e c t bya ppl epo l yphe no l si nr a t sf e dahi ghf a tdi eto rano r ma ldi e t .J o ur na lo fOl e oSc i e nc eVol .5 5:3 0 5 3 1 4 . 2 0 0 6 . a 0 4 . 山口 彩,藤田恵子,和田哲哉,長田恭一,山谷修.脂質代謝と抗酸化機能に与えるサメ肝油由来高純度スクア レンの摂取による影響の検証.脂質生化学研究 Vol .4 8:1 5 2 1 5 5 .2 0 0 6 . a 0 5 . 佐々木文之,石川絵理,長利 卓,佐藤玲子,竹本成孝,本田真也,藤兼洋一,鈴木喬士,長田恭一.茶葉飲料 のポリフェノールレベルとフリーラジカル捕捉機能,ならびにプロシアニジン類の抗酸化機能の検証.弘前大学 農学生命科学部学術報告,Vol .8:2 4 2 8 ,2 0 0 6 . b0 1 . 長田恭一.脂質の栄養:エッセンシャル基礎栄養学,宮本賢一ら編,第 5章 医歯薬出版(東京),p5 9 6 6 .2 0 0 5 . d0 1 . 長田恭一.シンポジウム:りんご未熟果実のプロシアニジン化合物による脂質代謝調節機能に関する基礎研究. 平成 1 7年度 日本栄養食糧学会東北支部会(盛岡).2 0 0 5 d0 2 . 山口 彩・鈴木喬士・藤兼洋一・長田恭一・山谷 修.肝油由来スクアレン投与による脂質代謝調節作用の解明. 日本栄養食糧学会東北支部会(盛岡) .2 0 0 5 d0 3 . 藤兼洋一・佐々木輔・清澤朋子・長田恭一・葛西厚史・千田峰生・菅野道廣.食事由来酸化コレステロールによ る脂質代謝攪乱作用の機序解明.平成 1 7年度 日本栄養食糧学会東北支部会(盛岡).2 0 0 5 . d0 4 . 鈴木喬士・山口 彩・長田恭一・神田智正・佐見 学・太田 豊・池田満雄.りんご未熟果実由来プロシアニジ ン摂取によるコレステロール代謝調節作用の機序解明.平成 1 7年度 日本栄養食糧学会東北支部会(盛岡). 2 0 0 5 . d0 5 . 鈴木喬士・山口 彩・長田恭一・神田智正・佐見 学・太田 豊・池田満雄.りんご未熟果実由来プロシアニジ ン摂取による肥満抑制作用と脂質代謝調節機能.2 0 0 5年度日本農芸化学会東北支部会.2 0 0 5 .(鶴岡). d0 6 . NAKAMURA,S,OSADA K,FUNAZAWA.R.:Ant i o xi da nta c t i vi t yo fwa t e r s o l ubl ef r a c t i o no fChi ne s eI ndi go .第 1 7回日本 MRS学術シンポジウム(東京)2 0 0 5 . d0 7 . 藤兼洋一・川岸 由・長田恭一・菅野道廣.食餌由来酸化コレステロール摂取濃度に応じた代謝変動の解明.2 0 0 6 年度日本農芸化学会(京都)2 0 0 6 . d0 8 . 山口 彩・藤田恵子・鈴木喬士・藤兼洋一・長田恭一・山谷 修.サメ肝油スクワレンと DHAリッチ油脂コン ビネーションによる脂質代謝調節作用追究.2 0 0 6年度日本農芸化学会(京都)2 0 0 6 . d0 9 長田恭一・船山 誠・清野博威・太田 豊・砂川忠広・佐見 学・神田智正・大竹康之.糖尿病モデルラットに 対するリンゴポリフェノール摂取の影響.2 0 0 6年度日本農芸化学会(京都)2 0 0 6 . d1 0 . 山口 彩・藤田恵子・鈴木喬士・藤兼洋一・長田恭一・山谷 修.脂質代謝および抗酸化機能に与えるサメ肝油 研 究 業 績 目 録 107 スクワレン+ DHAリッチ油脂の投与効果.第 6 0回日本栄養食糧学会 2 0 0 6 (静岡) d1 1 . 長田恭一・船山 誠・清野博威・太田 豊・砂川忠広・佐見 学・神田智正・大竹康之.リンゴポリフェノール 摂取による糖尿病予防および抗酸化強化作用.第 6 0回日本栄養食糧学会 2 0 0 6 (静岡) d1 2 . 長田恭一.特別講演 内蔵脂肪減らして防ごう生活習慣病:りんごポリフェノールの効用.20 0 6年度日本フラボ ノイド研究会近畿大会(大阪) .2 0 0 6 . d1 3 . 長田恭一.特別講演 内蔵脂肪減らして防ごう生活習慣病:りんごポリフェノールの効用.20 0 6年度日本フラボ ノイド研究会中部北陸大会(岐阜) .2 0 0 6 . d1 4 . 長田恭一.招待講演 鮫肝油由来高純度スクアレンの脂質代謝および抗酸化機能に与える作用.2 0 0 6年度油化 学セミナー(日本油化学会主催) (函館) .2 0 0 6 . 応用微生物学研究室 a 0 1 . MIZUKAMI,S. ,TAKEDA,K. ,AKADA,S. ,FUJITA,T.I s o l a t i o na ndCha r a c t e r i s t i c so fMe t h a n o s a e t ai npa ddyf i e l d s o i l s .Bi o s c i .Bi o t e c hno l .Bi o c he m.7 0 ,8 2 8 8 3 5 (2 0 0 6 ) a 0 2 . SUKTHAI,P. , ・佐々木長一・松山信彦・野田香織・殿内暁夫:カドミウム汚染水田の浸透型が物質動態および水稲 に及ぼす影響.土壌の物理性 1 0 1:1 7 2 6 (2 0 0 5 ) a 0 3 . 武田 潔:白神山地の窒素固定菌の研究.白神研究 3:2 6 3 1 (2 0 0 6 ) a 0 4 . 佐々木長市・松山信彦・本多和茂・殿内暁夫・野田香織・佐瀬 隆:白神山地における土壌(3 ) .白神研究 3 :4 5 5 2 (2 0 0 6 ) a 0 5 . 村中文人・岩間直子・斉藤知明・殿内暁夫・赤田辰治・武田 潔:カプロン酸エチル高生産株を用いたリンゴ搾 り粕からのフレーバーアルコール生産:日本醸造協会誌 1 0 1:7 1 1 7 1 6 (2 0 0 6 ) d0 1 . 殿内暁夫・野呂剛司:水田土壌から分離した新規な糸状菌について.第 2 1回日本微生物生態学会講演要旨集.p. 4 8 . (2 0 0 5 ) d0 2 . 舘山明日佳・犀川陽子 1・橋本貴美子 2・中田雅也 1・殿内暁夫・奥野智旦 3・宮入一夫:毒キノコ、オオシロカラ カサタケの毒性メタロエンドペプチダーゼの精製と切断特異性.日本農芸化学会東北支部第 1 4 0回大会(鶴岡) (2 0 0 5 ) d0 3. 岩間直子・斉藤知明 1・殿内暁夫・宮入一夫・武田 潔:麹菌 As p e r g i l l u so r y z a eにおける糖化酵素高発現および褐 変性に関する研究.2 0 0 6年度日本農芸化学会講演要旨集 p.8 3 . (2 0 0 6 ) d0 4 . 村中文人・岩間直子・斉藤知明・殿内暁夫・赤田辰治・武田 潔:りんご搾汁残渣からのフレーバーアルコール に関する研究.2 0 0 6年度日本農芸化学会講演要旨集 p.2 7 1 . (2 0 0 6 ) d0 5 . 佐々木長市・川島一就・松山信彦・野田香織・殿内暁夫:カドミウム汚染水田模型の浸透型が物質動態および水 稲に及ぼす影響.農業土木学会講演要旨集.pp8 2 2 8 2 3 . (2 0 0 5 ) d0 6 . 松山信彦・工藤予志夫・境谷栄二・殿内暁夫・工藤啓一:施肥により強酸性化したアロフェン質黒ボク土の特徴, 日本土壌肥料学会(2 0 0 6 ) d0 7 . 大久保敦史・殿内暁夫・杉山修一:ナタネとレンゲ根圏における土壌微生物群集構造の比較.日本作物学会第 2 2 1 講演会(東京大学) (2 0 0 6 ) d0 8 . 大久保敦史・殿内暁夫・杉山修一:根圏微生物群集の作物間差異- PCR- DGGE法による解析.植物微生物研 究会第 1 6回研究交流会(北海道大学) (2 0 0 6 ) 生体情報工学講座 生化・有機化学研究室 a 0 1 . 橋本 勝:ピラノース環酸素を硫黄原子で置換したチオオリゴサッカリドの合成研究.有機合成化学協会誌,6 4 (7 ) ,7 6 6 7 7 7(2 0 0 6 ) . 1 a 0 2 . SHIMIZU,T1. ,ARAYA,T. ,SHIBATA,H2. ,NAKATSU,T1. ,MIYAIRI,K. ,OKUNO,T. ,KATO,H. :Expr e s s i o n, pur i f i c a t i o na nd c r ys t a l l i z a t i o no fe ndo po l yga l a r a c t ur o na s ef r o m apa t ho ge ni cf ungus ,S t e r e u mp u r p u r e u m,i n Es c h e r i c h i ac o l i . Pr o t e i n Ex r ma c e ut i c a lSc i e nc e ,Kyo t o 0 0 5 ) . ( 1 Pha p r e s s i o na n d Pu r i f i c a t i o n .4 4 ,1 3 0 1 3 5 (2 Uni ve r s i t y,2RI KEN Ha r i maI ns t i t ut e ) c 0 1 . 村上貴宣,橋本 勝:マイコパラサイトの化学,リンゴ果実夏念珠病に関わるホスト菌にダメージを与える La mbe r t e l l o l類.化学と生物,4 (4 4 ) ,2 1 7 (2 0 0 6 ). c 0 2 . 橋本 勝:分子モデリングソフトの講義への利用 コンピュータ分子設計学を例として.弘前大学総合情報処理 研 究 業 績 目 録 1 08 センター広報 HI ROI N,2 3 ,6 3 7 0 (2 0 0 6 ) . c 0 3 . 橋本 勝:菌代謝産物の新展開.岩手大学連合大学研究科広報 RURUKA NEWS,6 ,Ma y(2 0 0 6 ). c 0 4 . 橋本 勝,山崎裕子:ナフタレノン化合物とその植物病害防除用途.特開 2 0 0 5 -2 0 6 2 5 9 . c 0 5 . 橋本 勝,姫野俵太,村上貴宣,奥野智旦:新規環状チオペプチドおよびその取得方法.特願 2 0 0 6 -0 5 8 0 6 8 0 . c 0 6 . 志村華子,高田 晃,幸田泰則:ランと菌の生存を賭けた闘争 レブンアツモリソウの共生発芽実験から見た 「共生」の実体.化学と生物,4 (4 4 ) ,2 6 1 2 6 4 (2 0 0 6 ). d0 1 . 山本和範,渡辺直樹,新谷智行,橋本 勝,宮入一夫,奥野智旦,岡崎 功,斉藤 稔,清水哲也,加藤博章: エンドポリガラクツロナーゼ反応機構解明を目指した硫黄置換オリゴガラクツロン酸の設計と合成.4 7回天然 有機化合物討論会(徳島) . d0 2 . 荒添圭朗,山本和範,佐藤 佑,橋本 勝,奥野智旦:e n d o PG 1の反応機構解明を目指した遷移状態アナログ の合成.日本農芸化学会東北支部第 1 4 0回大会(鶴岡). d0 3 . 村上貴宣,橋本 勝,奥野智旦:安定同位体取り込みを利用した ne o l a mbe r t e l l i nの同定とその構造決定.日本農 芸化学会東北支部第 1 4 0回大会(鶴岡) . d0 4 . 村上貴宣,橋本 勝,奥野智旦:Cyc l o t hi a z o myc i n Bの構造研究 ―立体化学の決定―.日本農芸化学会東北 支部第 1 4 0回大会(鶴岡) . d0 5 . 山本和範,新谷智行,橋本勝,宮入一夫,奥野智旦,岡崎 功,斉藤 稔:e n d o PG 1反応機構解明を目指した 硫黄置換オリゴガラクツロン酸の設計と合成.日本農芸化学会東北支部第 1 4 0回大会(鶴岡). d0 6 . MURAKAMI,T.;SASAKI,A.;FUKUSHI,E.;KAWABATA,J .;HASHIMOTO,M.;OKUNO,T.:Opt i mi z a t i o no fi s o t o pe l a be l i ng c o ndi t i o nsf o rl a mbe r t e l l i n ba s e do ni s o t o pe pa t t e r nso bs e r ve d by ma s ss pe c t r o me t r y.The 2 0 0 5 I nt e r na t i o na lChe mi c a lCo ngr e s so fPa c i f i cBa s i nSo c i e t i e s (ホノルル) . d0 7 . YAMAMOTO,K. ,MIYAIRI,K. ,OKUNO,T. ,SAITO,M. ,HASHIMOTO,M.:De s i gn a nd s ynt he s i so fa s ul f ur s ubs t i t ut e da na l o gue o ft r i ga l a c t ur o ni ca c i da s a mo l e c ul a r pr o be f o re n d o PG I .The 2 0 0 5I nt e r na t i o na l Che mi c a lCo ngr e s so fPa c i f i cBa s i nSo c i e t i e s (ホノルル) . d0 8 . MURAKAMI,T. ;HASHIMOTO,M. ;FUKUSHI,E. ;KAWABATA,J . ;OKUNO,T. :I s o l a t i o na nd bi o s ynt he s i s o fl a mbe r t e l l o l sf r o m myc o pa r a s i t i cf ungiLa mb e r t e l l as p.1 3 4 6 .The2 0 0 5I nt e r na t i o na lChe mi c a lCo ngr e s so f Pa c i f i cBa s i nSo c i e t i e s (ホノルル) . l o t hi a z o myc i nB1 ,B2の構造とその立体化学.日本農芸化学会 d0 9 . 村上貴宣,橋本 勝,奥野智旦,姫野俵太:Cyc 2 0 0 6年度大会(京都) . d1 0 . 木村天胤,村上貴宣,橋本 勝,奥野智旦,武藤 昱,姫野俵太:大腸菌リボソーム小サブユニットで活性化さ れる新規 GTPa s e (Rs gA)に作用する Cyc l o t hi a z o myc i nB1の作用.日本農芸化学会 2 0 0 6年度大会(京都) . d1 1 . 村上貴宣,橋本 勝,奥野智旦:マイコパラサイトと l a mbe r t e l l o l類の生産性との関連性について.日本農芸化 学会 2 0 0 6年度大会(京都) . d1 2 . 山本和範,荒添圭朗,橋本 勝,奥野智旦:e n d o PG 1基質遷移状態モデルの合成研究.日本農芸化学会 2 0 0 6年 度大会(京都) . d1 3 . 村上貴宣,奥野智旦,木村天胤,姫野俵太,橋本 勝:Cyc l o t hi a z o myc i nB1 ,B2の単離・構造決定・Rs gAに対 する活性.新規素材探索研究会 第 5回セミナー(静岡). d1 4 . MURAKAMI,T.;KIMURA,T.;OKUNO,T.;HIMENO,H.;HASHIMOTO,M.:I s o l a t i o n,s t r uc t ur ede t e r mi na t i o n,a nd bi o l o gi c a lpr o per t i e so fc yc l o t hi a z o myc i nsB1a nd B2 .I COB5& I SCNP2 5I UPAC I nt e r na t i o na lc o nf e r e nc e o nBi o di ve r s i t ya ndNa t ur a lPr o duc t s (京都) . d1 5 . HSHIMOTO,M.;YAMAMOTO,K.;SAITO,M.;MIYAIRI,K.:OKUNO,T.:Synt he t i cSt udi e so fOl i go ga l a c t ur o na t e a sMo l e c ul a rPr o be sf o re n d o PG 1 .I COB5& I SCNP2 5I UPAC I nt e r na t i o na lc o nf e r e nc eo nBi o di ve r s i t ya nd Na t ur a lPr o duc t s (京都) . d1 6 . 橋本 勝:リンゴ果実におけるマイコパラサイトの化学的研究.サントリー生物有機科学研究所月例コロキウム (大阪) . d1 7 . 橋本 勝:ピラノース環酸素を硫黄原子で置換したオリゴサッカリドの合成研究.関西学院大学理工学部講演会 (兵庫) . d1 8 . 舘山明日佳,犀川陽子 1,橋本貴美子 2,中田雅也 1,殿内暁夫,奥野智旦,宮入一夫:毒キノコ,オオシロカラカサ タケの毒性メタロエンドペプチダーゼの精製と切断特異性.日本農芸化学会東北支部第 1 4 0回大会(鶴岡) ( 1慶 応大・理工,2京薬大) . d1 9 . 清水哲哉 1,中津 亨 1,清水伸隆 2,佐藤 衛 2,栗原和男 3,宮入一夫,奥野智旦,新村信雄 4,山本雅貴 2,加藤 研 究 業 績 目 録 109 博章 1:リンゴ銀葉病菌由来エンドポリガラクツロナーゼの超高分解能 X線結晶構造解析と高分解能中性子結晶 構造解析.日本結晶学会(姫路) ( 1京大院薬,2理研播磨,3横浜市大院総合理・原研機構,4茨城大院理工) . d2 0 . 柴田泰成,工藤真紀子,奥野智旦,宮入一夫:Pl e u r o t u se r y n g i iの 2成分性ヘモリシン A,Bの単離と性質.日本 農芸化学会 2 0 0 6年度大会(京都) d2 1 . 新谷智行,清水哲哉 1,中津 亨 1,加藤博章 1,吉田 孝,奥野智旦,宮入一夫:S t e r e u mp u r p u r e u m由来エンド ポリガラクツロナーゼ Iにおける As n9 2結合糖鎖の熱安定性への関与.日本農芸化学会 2 0 0 6年度大会(京都) ( 1京都大薬) . d2 2 . 石井絵里子,数馬恒平 1,宮入一夫,奥野智旦:ベニバナ花弁由来のグルコース転移酵素について.日本農芸化学 会2 0 0 6年度大会(京都) ( 1青森グリーンバイオ). d2 3 . 堀部雅史 1,小林夕香 1,道羅英夫 1,松浦正憲 2,山田美奈 2,犀川陽子 2,宮入一夫,奥野智旦,橋本貴美子 3,中 田雅也 2,河岸洋和 1:ドクヤマドリ由来の毒性レクチンについて.日本農芸化学会 2 0 0 6年度大会(京都) ( 1静 岡大農,2慶応大・理工,3京都薬科大) . d2 4 . 山谷 聡,木皿正人,浅野純平,宮入一夫,奥野智旦 1:リンゴ花粉タンパクと花柱 SRNa s eの相互作用について. 日本農芸化学会 2 0 0 6年度大会(京都) (1秋田看福大). d2 5 . 岩間直子 1,斉藤知明 1,殿内暁夫,宮入一夫,武田 潔:麹菌 As p e r g i l l u so r y z a eにおける糖化酵素高発現および 褐変性に関する研究.日本農芸化学会 2 0 0 6年度大会(京都)(1青森工総セ・弘前技研). d2 6 . 堀部雅史 1,小林夕香 1,茶山和敏 1,道羅英夫 1,松浦正憲 2,山田美奈 2,犀川陽子 2,宮入一夫,奥野智旦,橋本 貴美子 3,中田雅也 2,河岸洋和 1:ドクヤマドリ由来の毒性レクチンの精製と性質第 7 8回日本生化学大会(神戸) ( 1静岡大農,2慶応大・理工,3京都薬科大). d2 7 . 高田 晃 1,松野純子 1,幸田泰則 1:ダイズの一回結実性老化を制御する内生因子に関する研究.4 7回天然有機化 合物討論会(徳島) ( 1北大院農) . d2 8 伴戸香月 1,高田 晃 1,幸田泰則 1:コムギの座死を引き起こす内生因子.平成 1 7年度北海道談話会(札幌) ( 1北 大院農) . d2 9 伴戸香月 1,高田 晃 1,幸田泰則 1:コムギの座死に関与する内生因子.日本農芸化学会 2 0 0 6年度大会(京都) (1北大院農) . d3 0 松浦真弓 1,志村華子 1,高田 晃 1,幸田泰則 1:レブンアツモリソウ幼植物が生産する抗菌物質.日本農芸化学 会2 0 0 6年度大会(京都) ( 1北大院農) . d3 1 . 木村天胤,村上貴宣,橋本 勝,奥野智旦,武藤 昱,姫野俵太:新規抗生物質による Rs gAの GTP加水分解 阻害メカニズムの解析,第 8回日本 RNA学会年会(淡路). f 0 1 . 宮入一夫:未利用資源活用の魔法の技術.市民公開講座.2 0 0 5. f 0 2 . 宮入一夫:リンゴペクチンの機能性.付属生物共生教育センター藤崎農場公開講座.りんごを科学する(板柳) . f 0 3 . 宮入一夫:リンゴ絞り粕からのペクチンオリゴ糖の簡易製造法とその利用.第 2回未利用バイオマス・セミナー. f 0 4 . 宮入一夫,水島伸治,土岐幸子,合田紫乃:リンゴ搾汁残渣からペクチンオリゴ糖の簡易製造法とその性状.産 学官連携フェア i n弘前『見てみて,聞いてみて,触ってみて,弘前大学』(弘前). f 0 5 . 橋本 勝:天然物化学の面白さ.弘前大学ドリーム講座 i n五所川原(青森). f 0 6 . 橋本 勝:天然物化学の面白さ.弘前大学ドリーム講座 i n三沢(青森). f 0 7 . 高田 晃,竹本成孝:i nv i t r o抗抽だい評価法の開発.産学官連携フェア i n弘前『見てみて,聞いてみて,触っ てみて,弘前大学』 (弘前) . 遺伝子工学研究室 a 0 1 . NAMEKI,N. ,SOMEYA,T. ,OKANO,S. ,SUEMASA,R. ,KIMOTO,M. ,HANAWASUETSUGU,K. ,TERADA,T. , SHIROUZU,M. ,HIRAO,I . ,TAKAKU,H. ,HIMENO,H. ,MUTO,A. ,KURAMITSU,S. ,YOKOYAMA,S.& KAWAI,G. I nt e r a c t i o na na l ys i sbe t we e nt mRNA a ndSmpB f r o m Th e r mu st h e r mo p h i l u s .J .Bi o c h e m.1 3 (2 8 0 0 5 )7 2 9 7 3 9 . a 0 2 . HOKII,Y. ,KUBO,A. ,OGASAWARA,T. ,NOGI,Y. ,TANEDA,A. ,ARAI,R. ,MUTO,A. ,USHIDA,C.Twe l veno ve lC. e l e g an a ndi da t e si s o l a t e dbyt wo di me ns i o na lpo l ya c r yl a mi dege le l e c t r o pho r e s i s .Ge n e3 6 (2 5 0 0 6 )8 3 8 7 . sRNA c b0 1 . 牛田千里, 「線虫にみる RNA機能の多様性」,機能性 No nc o di ngRNA(河合剛太,金井昭夫 編),5 5 -6 8頁, 2 0 0 6年. c 0 1 . 姫野俵太: 「1 0 SaRNAと共に過ごした 1 3年」,RNA Ne t wo r kNe ws l e t t e r ,(1 5 ) ,3 1 3 3 (2 0 0 6 ). c 0 2 . 橋本 勝,姫野俵太,村上貴宣,奥野智旦:新規環状チオペプチドおよびその取得方法.特願 2 0 0 6 -0 5 8 0 6 8 0 . c 0 3 . 牛田千里:低分子 RNAの分析方法.外国出願特許 PCTJ P2 0 0 6 / 3 0 0 0 7 8 . 研 究 業 績 目 録 1 10 d0 1 . 栗田大輔,今野貴之,佐々木瑠美,浅野クリスナ,高田一馬,武藤 昱,姫野俵太:Hydr o xyr a di c a lf o o t pr i nt i ng による SmpBとリボソームの相互作用の解明,第 8回日本 RNA学会年会(淡路)(2 0 0 6 ). d0 2 . 木村天胤,村上貴宣,橋本 勝,奥野智旦,武藤 昱,姫野俵太:新規抗生物質による Rs gAの GTP加水分解 阻害メカニズムの解析,第 8回日本 RNA学会年会(淡路)(2 0 0 6 ). d0 3 . 高田一馬,竹本千重,川添将仁,今野貴之,浅野クリスナ,白水美香子,横山茂之,武藤 昱,姫野俵太:高度 好熱菌の無細胞翻訳系を用いたトランス・トランスレーション,第 8回日本 RNA学会年会(淡路)(2 0 0 6 ) . d0 4 . 木村天胤,村上貴宣,橋本 勝,奥野智旦,武藤 昱,姫野俵太:大腸菌リボソーム小サブユニットで活性化さ れる新規 GTPa s e (Rs gA)に作用する Cyc l o t hi a z o myc i nB1の作用.日本農芸化学会 2 0 0 6年度大会(京都) (2 0 0 6 ) . d0 5 . 村上貴宣,奥野智旦,木村天胤,姫野俵太,橋本 勝:Cyc l o t hi a z o myc i nB1 ,B2の単離・構造決定・Rs gAに対 する活性.新規素材探索研究会 第 5回セミナー(静岡)(2 0 0 6 ). d0 6 . MURAKAMI,T. ,KIMURA,T. ,OKUNO,T. ,HIMENO,H.& HASHIMOTO,M.:I s o l a t i o n,s t r uc t ur ede t e r mi na t i o n,a nd bi o l o gi c a lpr o pe r t i e so fc yc l o t hi a z o myc i nsB1a nd B2 .I COB5& I SCNP2 5I UPAC I nt e r na t i o na lc o nf e r e nc e o nBi o di ve r s i t yandNa 0 0 6 ). t ur a lPr o duc t s (京都)(2 d0 7 . 牛田千里,小笠原隆広,保木井悠介,武藤 昱:線虫低分子 RNAのカタログ化,第 2 8回日本分子生物学会年会 (福岡) (2 0 0 5 ) . d0 8 . Yus ukeHOKII,Ma s a yo s hiSHIMOYAMA,Aki r aMUTO,Chi s a t oUSHIDA :Cha r a c t e r i z a t i o no fCe n2 1 / Ce R2RNA, s ma l lnc RNA l o c a l i z e di nCa e n o r h a b d i t i se l e g a n snuc l e o l i .第 2 0回国際生化学・分子生物学会議(京都)(2 0 0 6 ) . d0 9 . USHIDA,C. ,HOKII,Y. ,SUGAWARA,Y. ,MIZUKI,Y. ,ENDO,Y.& MUTO,A.:Expr e s s i o no fC.e l e g a n sno ve l nc RNAs .Re gul a r o t yRNAs ,t heLXXICo l dSpr i ngHa r bo rLa bo r a t o r ySympo i um(Co l d Spr i ng Ha r bo r ) (2 0 0 6 ) . d1 0 . 保木井悠介,笹野由未,藤原俊伸,坂本 博,種田晃人,武藤 昱,牛田千里:C.e l e g a n s低分子 RNA Ce R2RNA の細胞内局在と生合成,第 8回日本 RNA学会年会(淡路)(2 0 0 6 ). d1 1 . 遠藤優子,保木井悠介,牛田千里:C.e l e g a n s新規 H/ ACA型 RNAの発現解析.RNA若手の会 2 0 0 6 (三島) (2 0 0 6 ) . d1 2 . 牛田千里:C.e l e g a n ss ma l lRNAs .新しい nc RNA/ RNPを見つける会(お台場)(2 0 0 6 ). d1 3 . 菅原由起,保木井悠介,遠藤優子,牛田千里:ホールマウント RNAFI SHによる線虫低分子 RNAの発現解析. 新しい nc RNA/ RNPを見つける会(お台場)(2 0 0 6 ). f 0 1 . 姫野俵太:t mRNAによる t r a ns t r a ns l a t i o nの分子メカニズム:mo l e c ul a rmi mi c r y? ,弘前大学遺伝子実験施設第 1 2回(竣工 1 0周年記念)シンポジウム(弘前)(2 0 0 6 ). f 0 2 . 姫野俵太,武藤 昱,牛田千里,佐野輝男,原田竹雄,鬼島 宏,種田晃人,千田峰生:機能性 RNAを基盤と する新技術開発のための基礎研究とその展開.産学官連携フェア i n弘前『見てみて,聞いてみて,触ってみて, 弘前大学』 (弘前) (2 0 0 6 ) . f 0 3 . 牛田千里:機能未知新規低分子 RNAの機能解析にむけて,機能性 RNAプロジェクトキックオフミーティング (お台場) (2 0 0 5 ) . f 0 4 . 牛田千里:線虫新規低分子 RNAの局在,機能性 RNAプロジェクトコロキウム(お台場)(2 0 0 5 ). f 0 5 . 牛田千里:機能未知新規低分子 RNAの機能解析,機能性 RNAプロジェクト成果報告会(お台場)(2 0 0 6 ) . f 0 6 . 牛田千里:Func t i o na lRNo mi c si nC.e l e g a n s .岩手大学 COEフォーラム(盛岡)(2 0 0 6 ). 細 胞 工 学 講 座 微生物工学研究室 a 0 1 . 片方陽太郎:非上皮細胞におけるケラチンの発現. 生化学,7 8 ,7 7 0 7 7 4 .2 0 0 6 . a 0 2 . 片方陽太郎・渡辺名歩・加藤陽治:血管肉腫細胞株での Hs p4 7の再現性と他の HSP分子について.第 2 0回角 化症研究会記録集,pp.8 1 1 .2 0 0 5 . a 0 3 . TATARA,Y. ,YOSHIDA,T. ,I i ngl ef r e ec yc t e i ner e s i duea nddi s ul f i debo ndc o nt r i but et ot he CHI SHI MA,E.:A s t he r mo s t a bi l i t yo fAs p e r g i l l ss a i t o i1 ,2 αma nno s i da s e .Bi o s c i e n c e ,Bi o t e c h n o l o g y ,a n dBi o c h e mi s t r y ,6 9 ,2 1 0 1 2 1 0 8 , 2 0 0 5 . a 0 4 . AKAO,T. ,YAMAGUCHI,M. ,YAHARA,A. ,YOSHIUCHI,K. ,FUJITA,H. ,YAMADA,O. ,AKITA,O. ,OHMACHI,T. , ASADA,Y. ,YOSHIDA,T.:Cl o ni ng a nd e xpr e s s i o no f1 ,2 αma nno s i da s e ge ne (f ma n I B)f r o m f i l a me nt o us 研 究 業 績 目 録 111 f ungusAs p e r g i l l u so r y z a e:i nv i v ovi s ua l i z a t i o no ft heFma nI BpGFP f us i o n pr o t e i n.Bi o s c i e n c e ,Bi o t e c h n o l o g y , a n dBi o c h e mi s t r y ,7 0 ,4 7 1 4 7 9 ,2 0 0 6 . d0 1 . 田島 勇・大町鉄雄:Ps e u d o mo n a ss p.ON4 a株の新規 2 -アミノチアゾリン-4 -カルボン酸加水分解酵素:大 腸菌形質転換体から ATC加水分解酵素の精製とその性質.第 7 8回日本生化学会大会(神戸)2 0 0 5 . d0 2 . OCHIAI,H. ,TAKEDA,K. ,FUKUZAWA,M. ,KATO,A. ,SAITO,T. ,OHMACHI,T.:ThePKB/ AKT r e l a t e d ki na s e ma y bee nga ge di nt het r a ns i t i o nf r o mt hef i r s tf i nge rs t a get ot hes l ug s t a ge .I nt e r na t i o na lCo nf e r e nc eo f Di c t yo s t e l i um(Di nt aFe ,Ne w Me xi c o ,USA) ,2 0 0 6 . c t y2 0 0 6 ) , (Sa d0 3 . KATAGATA,Y. ,OTSU,K. ,HIRAYAMA,T.:Expr e s s i o no fHSP(He a tSho c kPr o t e i n)i nc ul t ur e dhuma nt umo r c e l ll i ne s .2 0 t hI UBMB I nt e r na t i o na lCo ngr e s so fBi o c he mi s t r ya nd Mo l e c ul a rbi o l o gy a nd 1 1 t h FAOBMB Co ngr e s s , (Kyo t o )2 0 0 6 .6 .2 1 . d0 4 . 片方陽太郎・小田桐弓芽乃・高橋和久:植物細胞に中間径線維タンパク質は存在するか?(第 1報).第 2 1回角 化症研究会(東京)2 0 0 7 .8 .5 . d0 5 . 吉田 孝・笠島勇也・平井伸明・山口雅子・大町鉄雄:麹菌のタンパク質糖鎖に対する Gl c NAc転移酵素発現の 影響.日本農芸化学会大会,2 0 0 6 .3 . d0 6 . 新谷智行・清水哲哉・中津 亨・加藤博章・吉田 孝・奥野智旦・宮入一夫:S t e r e u mp u r p u r e u m由来エンドポリ ガラクツロナーゼ Iにおける As n9 2結合糖鎖の熱安定性への関与.日本農芸化学会大会,2 0 0 6 .3 . d0 7 . 多田羅洋太 *・難波 剛*・吉田 孝・一島英治*:酸性条件で活性化するプロチロシナーゼのユニークな機構.日 本農芸化学会大会,2 0 0 6 .3 . (*創価大学工学部) d0 8 . 小泉英誉・高橋 礼*・吉田尚生*・戸谷一英*・北本則行**・吉田 孝:La c縮合活性を有するセルラーゼの精 製.FCCAグライコサイエンス若手フォーラム,2 0 0 6 .8 . (*一関高専,**愛知食品工技) e 0 1 . 吉田 孝:食品・医薬品開発を目的とした新規な酵素の探索.平成 1 7年度青森県学術国際振興基金助成事業研究 結果報告,2 0 0 6 . f 0 1 . 佐々木文之,片方陽太郎:リンゴ鹿角霊芝の水溶性成分による腫瘍細胞の生育抑制について.産学官連携フェア i n弘前・ 「見てみて聞いてみて触ってみて弘前大学」(弘前)2 0 0 6 .9 .1 9 . f 0 2 . 吉田 孝:オリゴ糖をつくる酵素・削る酵素.産学官連携フェア i n弘前「見てみて,聞いてみて,触ってみて弘 前大学」 (弘前)2 0 0 6 .9 .1 9 . 細胞工学研究室 a 0 1 . SUGANO,E. ,TOMITA,H. ,I . ,I ,a nd TAMAI,M.:Ni t r i co xi de i nduc e da c c umul a t i o no f SHI GURO,S SAGO,H. l i po f us c i nl i kema t e r i a l si sc a us e dbyi nhi bi t i o no fc a t he ps i nS.Cur r .EyeRe s .3 1:6 0 7 6 1 6 ,2 0 0 6 . d0 1 . 尾崎 卓・富田浩史*・玉井 信*・石黒誠一:ミトコンドリア膜間スペースのμ-カルパイン様プロテアーゼの 性質.第 7 8回日本生化学大会(神戸国際会議場)2 0 0 5 .1 0 . (*東北大学先進医工学研究機構) * * d0 2 . OZAKI,T. ,TOMITA,H. ,Ma ko t o TAMAI,M. ,I .:Pr e s e nc eo fμc a l pa i nl i ke pr o t e a s ei nt he SHI GURO,S mi t o c ho ndr i a li nt e r me mbr a nes pa c eo fs wi nel i ve r .2 0 t hI UBMB I nt e r na t i o na lCo ngr e s so fBi o c he mi s t r ya nd ngr e s s .(Kyo t oI nt e r na t i o na lCo nf e r e nc eHa l l )(第 2 0回国際生化学会) J une 1 9 ,2 0 0 6 . 1 1 t hFAOBMB Co (* Bi o me di c a lEngi ne e r i ngRe s e a r c hOr ga na i z a t i o n,To ho kuUni ve r s i t y,Se nda i ,J a pa n) * * d0 3 . MIZUKOSHI,S.OZAKI,T. ,I . ,NAKAZAWA,M. ,OHGURO,H. :Thec ha ngeo fc a l pa i na c t i vi t yi nt he SHI GURO,S r e t i na so ft heRCS r a t st r e a t e d wi t h Ca2+ a nt a go ni s t ,Ni l va di pi ne .TheFo ur t h As i a n Co nf e r e nc eo n Vi s i o n. (く に び き メ ッ セ,松 江) ,J ul y2 8 Augus t1 .2 0 0 6 . (* De pa r t me nto fOpht ha l mo l o gy,Hi r o s a kiUni ve r s i t y Sc ho o lo fMe di c i ne ) d0 4 . 澁谷基世・石黒誠一:ブタ網膜における局所特異的なタンパクの解析.平成 1 8年度日本動物学会東北支部大会 (山形県立博物館講堂) 2 0 0 6 .8 .5 . d0 5 . 服部安孝・浅井信晴・石黒誠一:脳由来神経栄養因子投与によりブタ網膜から放出されるタンパク質の解析.第 7 7回日本動物学会大会(島根大学)2 0 0 6 .9 .2 1 2 4 . d0 6 . KUDO,K. ,SASAMORI,E. ,EBINA,M. ,KIKUCHI,H.:I de nt i f i c a t i o no fa mi noa c i dr e s i due si nvo l ve di nt hel i ga ndbi ndi ngo fmo us eAhr e c e pt o r .第 7 8回日本生化学会(神戸)2 0 0 5 . d0 7 . 上野紀子*・武谷 立*・宮野 佳*・菊池英明・住本英樹*:NADPHオキシダーゼ No x3は p2 2 pho x依存性に恒 常的にスーパーオキシドを生成する:オキシダーゼ o r ga ni z e rと a c t i va t o rによる調節.第 2 8回日本分子生物学 会年会(福岡)2 0 0 5 . (*九州大学医学部) d0 8 . 石田真人・So he lAhme d・菊池英明:LMAT細胞における TCDD誘因性アポトーシスでの PKCɵと Va vの相互 1 12 研 究 業 績 目 録 作用の変化.日本生化学会東北支部第 7 2回例会(弘前)2 0 0 6 .5 .1 3 . d0 9 . KOBAYASHI,D. ,AHMED,S. ,I ,KASAI,S. ,KIKUCHI,H.:I nt r a c e l l ul a rc a r c i uli sa s s o c i a t e d wi t h TCDD SHI DA,M. i nduc e da po pt o s i si n LMAT,a huma nl ympho bl a s t i c Tc e l ll i ne .2 0 t hI UBMB I nt e r na t i o na lCo ngr e s so f Bi o c he mi s t r ya ndMo l e c ul a rBi o l o gya nd1 1 t hFAOBMB Co ngr e s s . (Kyo t o ) .J une1 8 2 3 .2 0 0 6 . d1 0 . 畠山幸紀・雨森道紘*:Ti meLa psVi de o撮影によるマウス白血病細胞の運動解析.第 3 5回日本免疫学会総会・ 学術集会(パシフィコ横浜)2 0 0 5 .1 2 . (*弘前大学理工学部) d1 1 . 畠山幸紀・雨森道紘*:動画解析プログラムを用いたマウス白血病細胞の運動能の解析.第 6 5回日本癌学会学術 総会(パシフィコ横浜)2 0 0 6 .9 . (*弘前大学理工学部) f 0 1 . 畠山幸紀*:科学番組「細胞力覚~生命を支える力の感覚」 (曽我部正博 監修,科学技術振興機構,NTV映像 センター制作)J STサイエンスチャンネル.2 0 0 5 . (*資料協力) f 0 2 . 畠山幸紀*:科学番組「生命に秘められたアポトーシスの謎」 (長田重一 監修,科学技術振興機構,NTV映像 センター制作)J STサイエンスチャンネル.2 0 0 6 . (*資料協力) 研 究 業 績 目 録 113 生 物 生 産 科 学 科 園 芸 学 講 座 a 0 1 . BAKHSHI D.a nd ARAKAWA O.:Ef f e c t so fUVBi r r a di a t i o no n phe no l i cc o mpo und a c c umul a t i o na nd a nt i o xi da nta c t i vi t yi n‘J o na t ha n’a ppl ei nf l ue nc e dbyba ggi ng,t e mpe r a t ur ea ndma t ur a t i o n.J .Fo o d,Agr i .& Envi r o n.(1 4 ):7 9 8 3 .2 0 0 6 a 0 2 . 荒川 修:リンゴの着色と紫外線.太陽紫外線防御研究委員会学術報告第 1 5巻第 1号 2 1 2 4 .2 0 0 5 a 0 3 . 浅田武典・堀田拓人:リンゴ樹の結果枝構成に及ぼす品種とせん定の影響.園芸学研究 4 (4 ):4 1 7 4 2 1 .2 0 0 5 . a 0 4 . 本多和茂・北藤吉浩 *:低温処理がデルフィニウム園芸品種‘サマースカイズ’の発芽および開花におよぼす影 響.専修大学北海道短期大学環境科学研究所報告 第 1 2号 :1 1 3 1 1 5 .2 0 0 5 . (* J A美唄 苗工房) a 0 5 . Mi ng SUN* , Shuhua iZHANG, Ya o gua ng WEI* ,Do ng AN* :No nde s t r uc t i ve Me a s ur e me nt o f Fr ui t Fi r mne s s Us i ng a La s e r Do nt e r na t i o na lCo nf e r e nc eo nI mpul s i ve ppl e r Vi br o me t e r , Pr o c e e di ngs t he 3rd I Dyna mi cSys t e msa ndAppl i c a t i o n,1 4 0 5 1 4 0 8 ,2 0 0 6 (*中国農業大学) a 0 6 . Ke nt a r oMATSUO* ,Ke nt a r oNISHIWAKI* ,Shuhua iZHANG,Mi ki oYASHIRO* :De ve l o pme nto fExpe r i me nt a l Se t upf o rDi s t i nc t i o no fDi s e a s ePl a nt ,ASABE Pa pe rNo .0 6 3 0 1 6 ,1~ 5 ,2 0 0 6 (*東北農研センター) a 0 7 . 張 樹槐:携帯電話ネットワークによる農業施設の監視・制御システムの構築 ―システムの構築及び動作確認 実験―,弘前大学総合情報処理センター広報 HI ROI N No.2 3 ,4 1 4 4 ,2 0 0 6 a 0 8 . 加藤弘道,張 樹槐,福地 博:氷温冷蔵に関する研究,弘前大学農学生命科学部学術報告 No.8 ,3 7~ 4 5 ,2 0 0 5 a 0 9 . 武田純一*,鳥巣 諒*,張 樹槐,陳 軍*,朱 忠祥*:中国における大学の研究動向について,農業機械学会 東北支部報 No.5 2 ,2 5~ 2 8 ,2 0 0 5 (*岩手大学) b0 1 . ARAKAWA O.a nd KOMORI S.:Appl e ,Hor t i c ul t ur ei nJ a pa n2 0 0 6 ,Edi t e d by t he J a pa ne s e So c i e t yf o r Ho r t i c ul t ur a lSc i e nc e ,Sho uka do hPubl i c a t i o n,De pt .o fPubl i s hi ngo fNa ka ni s hiPr i nt i ngCo . ,Lt d.3 4 4 22 0 0 6 d0 1 . マナスィカン タマウォン,荒川 修:リンゴ 4品種における果実の生長と成熟に伴うデンプン,アミロースお よびアミロペクチン含量の変化の比較,園学雑 7 (別 5 2 ):1 2 9 .2 0 0 6 d0 2 . ARAKAWA O.a nd BAKHSHI D.:Po l yphe no l i c sa c c umul a t i o ni n’ Fuj i ’a sa f f e c t e d by e nvi r o nme nt a la nd phys i o l o gi c a lf a c t o r s .TheSe c o ndI nt e r na t i o na lSympo s i um o nAppl ePr o duc t i o n,2 0 0 6 d0 3 . 長岐正彦*,伊丸岡大斗*,嵯峨紘一,槙 雄二**,古山種俊***:カボチャの組織培養を利用したイソプレノイ ド合成.日本化学会第 8 6春季年会. (*弘前大理工,**山形大理,***東北大多元研) d0 4 . 長岐正彦*,伊丸岡大斗*,嵯峨紘一,槙 雄二**,古山種俊***:カボチャの組織培養を利用したゲラニルゲラ ニオール(GGOH)の Bi o t r a ns f o r ma t i o n.日本化学会第 8 6春季年会.(*弘前大理工,**山形大理,***東北大多 元研) d0 5 . 嵯峨紘一,相馬志穂,柴谷奈津子:トウガラシ果肉乾燥粉末の貯蔵過程における内容成分の変化.園学雑 7 5別 1:3 4 9 .2 0 0 6 . d0 6 . 長岐正彦*,伊丸岡大斗*,嵯峨紘一,西野徳三**,佐上 博***,大場 亮***,古山種俊***:Bi o t r a ns f o r ma t i o n による抗ガン剤開発の試み.平成 1 8年度化学系学協会東北大会.(*弘前大理工,**東北大院工,***東北大多元 研) d0 7 . 嵯峨紘一,相馬志穂:弘前在来’トウガラシ果実の緑色期における辛味成分の発現と果色の変化.園芸学会東北 支部平成 1 8年度大会研究発表要旨.1 2 .2 0 0 6 . d0 8 . 佐々木和也*,嵯峨紘一,鮫島正純:夏季および冬季における GA3処理がシネンシス系デルフィニウムの生育に 及ぼす影響.園芸学会東北支部平成 1 8年度大会研究発表要旨.4 5 4 6 .2 0 0 6 . (* 青森県農林総研フラワーセン ター) d0 9 . 佐々木和也*,嵯峨紘一,鮫島正純:日長および温度がシネンシス系デルフィニウムの抽台に及ぼす影響.園学 雑7 5 .別 2:3 6 7 .2 0 0 6 . (*青森県農林総研フラワーセンター) d1 0 . 本多和茂・北藤吉浩*:低温処理がデルフィニウムの発芽および開花におよぼす影響.園学雑 7 4別 2:2 2 6 .2 0 0 5 . (* J A美唄 苗工房) . d1 1 . 本多和茂・澁谷真実・佐藤 恵・嵯峨紘一・小林 勝*:弘前公園におけるサクラの花芽分化・休眠および開花に 関する研究.園芸学会東北支部大会平成 1 8年度大会 研究発表要旨:4 9 5 0 . (*弘前市商工観光部公園緑地課) d1 2 . 片平光彦*1,田村 晃*1,張 樹槐,大泉隆弘*2,後藤恒義*2:画像処理によるエダマメの選別方法に関する研究 (第 1報)―傷害の発生頻度と選別基準の策定―,農業環境工学関連 7学会 2 0 0 6年合同大会講演要旨集,2 7 , 研 究 業 績 目 録 1 14 2 0 0 6 (*1秋田県農業試験場,*2㈱山本製作所) d1 3 . 張 樹槐,片平光彦*1,田村 晃*1,大泉隆弘*2,後藤恒義*2:画像処理によるエダマメの選別方法に関する研究 (第 2報)―エダマメの熟度計測及び傷害検出方法について―,農業環境工学関連 7学会 2 0 0 6年合同大会講演要 旨集,2 7 ,2 0 0 6 (*1秋田県農業試験場,*2㈱山本製作所) d1 4 . 上野有穂,張 樹槐:分光反射特性を利用したリンゴ果実表面の打撲傷の検出,農業環境工学関連 7学会 2 0 0 6年 合同大会講演要旨集,3 5 ,2 0 0 6 d1 5 . 張 樹槐,川田将司,片平光彦*1,後藤恒義*2,大泉隆弘*2:画像処理によるサヤエンドウの熟度計測,平成 1 8 年度農業機械学会東北支部研究発表会講演要旨,5~ 6 ,2 0 0 6 (*1秋田県農業試験場,*2㈱山本製作所) d1 6 . 上野有穂,張 樹槐:分光反射特性を利用したリンゴ果実表面の打撲傷の検出,平成 1 8年度農業機械学会東北支 部研究発表会講演要旨,7~ 8 ,2 0 0 6 f 0 1 . 張 樹槐:リンゴ果実の内部・外部品質の非破壊的測定方法,平成 1 7年藤崎農場公開講座「リンゴを科学する」 テキスト,4 2~ 4 7 ,2 0 0 5 農業生産学講座 作物学教室 a 0 1 . 松山信彦・高橋 正*:黒ボク土のペドロジーとエダホロジー,(2 ) 分類,東北の農業と土壌肥料,p7 7~ 8 0 ,日 本土壌肥料学会東北支部編,2 0 0 6 . (*東北大学大学院) a 0 2 . 村山成治・松山信彦・齋藤 司・小田桐正英・工藤啓一:低投入型稲作に関する研究,第 6報 アイガモ除草を 導入した不耕起移植栽培における雑草発生量と水稲の生育・収量,日本作物学会東北支部会報,4 8 ,2 1~ 2 3 ,2 0 0 5 . a 0 3 . 佐々木長市・松山信彦・本多和茂・殿内暁夫・野田香織・佐瀬 隆*:白神山地の土壌に関する研究(3 ),白神研 究,3 ,4 5~ 5 2 ,2 0 0 6 . (*北方ファイトリス研究室) a 0 4 . PONGPATTANASIRI,S. ・佐々木長市・松山信彦・野田香織・殿内暁夫:カドミウム汚染水田の浸透型が物質動態お よび水稲に及ぼす影響,土壌の物理性,1 0 1 ,1 7~ 2 6 ,2 0 0 5 . d0 1 . 松山信彦・工藤予志夫*・境谷栄二*・殿内暁夫・工藤啓一:施肥により強酸性化したアロフェン質黒ボク土の特 徴,日本土壌肥料学会,2 0 0 6 . (*青森県農林総合研究センター) d0 2 . 岩渕伸雄・村山成治・松山信彦・工藤啓一:低投入型稲作に関する研究,第 7報 リンゴジュース搾り粕の施用 方法と抑草効果の関係,日本作物学会東北支部講演会,2 0 0 6 . 植物遺伝生態学研究室 a 0 1 . SUGIYAMA S.:Re s po ns e so fs ho o tgr o wt ha nd s ur vi va lt owa t e rs t r e s sgr a di e nti n di pl o i da nd t e t r a pl o i d po pul a t i o nso fLo l i u m mu l t i f l o r u ma ndL.p e r e n n e .Gr a s s l a ndSc i e nc e5 2:1 5 5 1 6 0 .2 0 0 6 . d0 1 . 杉山修一・二戸奈央子:気候温暖化に対する牧草の適応ポテンシャル I V.オーチャードグラス自然集団の分布中 央と南限集団のアイソザイムによる遺伝変異量の比較.日本草地学会第 6 1回講演会(筑波農林団地)2 0 0 6 d0 2 . HOSSAIN Z. ,S.SUGIYAMA :Pl a ntl i t t e re f f e c t so nt hes o i lmi c r o bi a lc o mmuni t ys t r uc t ur e PLFA a na l ys i s .The 2nd Sc i e nt i f i cCo ngr e s so fEa s tAs i a nFe de r a t i o no fEc o l o gi c a lSo c i e t i e s . (Ni i ga t aUni ve r s i t y) .2 0 0 6 d0 3 . SUGIYAMA S. ,Z.HOSSAIN :Gr a z i ng i nt e ns i t ya nd s o i lmi c r o bi a lc o mmuni t ys t r uc t ur ei ns e mi na t ur a l gr a s s l a nds ,J a pa n. The2nd Chi na J a pa nSo ut hKo r e aj o i nts ympo s i um o ngr a s s l a nda gr i c ul t ur a la ndl i ve s t o c k pr o duc t i o n. (La nz ho uUni ve r s i t y,Chi na )2 0 0 6 d0 4 . 大久保敦史・殿内暁夫・杉山修一:ナタネとレンゲ根圏における土壌微生物群集構造の比較.日本作物学会第 2 2 1 講演会(東京大学)2 0 0 6 d0 5 . 大久保敦史・殿内暁夫・杉山修一:根圏微生物群集の作物間差異-PCR-DGGE法による解析.植物微生物研究 会第 1 6回研究交流会(北海道大学)2 0 0 6 f 0 1 . 杉山修一:科学研究費補助金 基盤研究(B)「気候温暖化による牧草種の衰退原因と適応ポテンシャルの解明」 研究成果報告書.2 0 0 6 畜産学研究分野 1) a 0 1 . SUZUKI,H. ,SATOH,M. a nd TOYOKAWA,K.:Cha nge si n di s t r i but i o no fa c t i vemi t o c ho ndr i adur i ng o o c yt e ma t ur a t i o na ndf e r t i l i z a t i o ni nt heha ms t e r .J .Ma mm.OvaRe s .2 2:1 6 3 1 6 9 .2 0 0 5 . a 0 2 . SUZUKI,H.a ndSAITO,Y.2) :Cumul usc e l l sa f f e c tdi s t r i but i o na ndf unc t i o no ft hec yt o s ke l e t o na ndo r ga ne l l e 研 究 業 績 目 録 115 i nt hepo r c i neo o c yt e s .Re pr o duc t i veMe di c i nea ndBi o l o gy,1 8 3 1 9 4 .2 0 0 6 . a 0 3 . 松崎正敏:初期成長期の栄養制御によるプログラミングを利用した家畜の生産能力の向上.Bi o九州 1 8 0:6 8 ,2 0 0 6 . * * * a 0 4 . WALLACE,J . M. ,MATSUZAKI,M. ,MILNE,J . S. ,a ndAITKEN,R. P. :La t ebutno te a r l yge s t a t i o na lma t e r na l gr o wt h ho r mo ne t r e a t me nt i nc r e a s e sf e t a la di po s i t y i n o ve r no ur i s he d a do l e s c e nt s he e p.Bi o l o gy o f Re pr o duc t i o n7 5:2 3 1 2 3 9 ,2 0 0 6 . (* Ro we t tRe s e a r c hI ns t i t ut e ) a 0 5 . 常石英作*,中西雄二*,平野 清*,小路 敦*,松崎正敏,柴 伸弥**:放牧牛の半棘筋における機能性成分と 遊離アミノ酸の含有量.西日本畜産学会報 4 9:1 0 3 1 0 5 ,2 0 0 6 . (*九州沖縄農業研究センター,**東北農業研究 センター) d0 1 . 鈴木裕之:成熟中のブタ卵における細胞骨格の再配置―とくに卵丘細胞の付着程度の影響―.第 5回 RMB(生 殖医学・生物学)研究会シンポジウム(東京都).2 0 0 6 . * * ** *** * ,WATANABE,A. ,SUZUKI,T. ,TSUNEISHI,E. ,MATSUZAKI,M. ,HIGUCHI,M. ,a ndI d0 2 . SHIBA,N. WAMOTO, **** H. :I nt r a mus c ul a rc o l l a ge nc ha r a c t e r i s t i c so fdo ubl e mus c l e da nd no r ma lJ a pa ne s es ho r t ho r ns t e e r s . 5 2 nd I nt e r na t i o na l Co ngr e s so f Me a t Sc i e nc e a nd Te c hno l o gy(Dubl i n, I r e l a nd) ,2 0 0 6 . ( * Na t i o na l Agr i c ul t ur a lRe s e a r c h Ce nt e rf o r To ho ku Re gi o n,** I wa t e Agr i c ul t ur a lRe s e a r c h Ce nt e r ,*** Na t i o na l Agr i c ul t ur a lRe s e a r c hCe nt e rf o rKyus huOki na waRe gi o n,**** Kyus huUni ve r s i t y) 榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎 1 ) 佐藤 学 現在は I VF大阪 2) 齋藤陽介 現在は宮城県職員 生産管理システム学 a 0 1 . 加藤弘道・張 樹槐・福地 博:氷温冷蔵に関する研究.弘前大学農学生命科学部学術報告,8:3 7 4 5 ,2 0 0 5 . 環境生物学講座 植物病理学研究室 a 0 1 . HARADA,Y. ,SASAKI,M. ,SASAKI,Y. ,I ,Y. ,SANO,T.:Mo n i l i n i as s i o r is p.no v.i nt heSc l e r o t i ni a c e a e , CHI HASHI c a us i ngl e a fbl i ghta ndyo ungf r ui tr o to fPr u n u ss s i o r ii nJ a pa n.Myc o s c i e ne4 (6 6 ):3 7 6 3 8 0 .2 0 0 5 . *1 *1 a 0 2 . LI,S. ,QIAN,S. ,GUO,R.*1,TSUJI,M. ,SANO,T.;Thef i r s tr e po r to fCo l e usbl ume ivi r o i df r o mc o l e usi n Chi na .Pl a ntPa t ho l o gy5 5:5 6 5 .2 0 0 5 . *1 *1 a 0 3 . LI,S. ,GUO,R. ,TSUJI,M. ,SANO,T.:Fi r s tr e po r t so ft wogr a pe vi nevi r o i dsi n Chi naa nd t hepo s s i bl e de t e c t i o no fat hi r d.Pl a ntPa t ho l o gy5 5:5 6 4 .2 0 0 5 . *2 a 0 4 . TAKAKI,F. ,SANO,T. ,YAMASHITA,K. :Thec o mpl e t enuc l e o t i des e que nc eo fa t t e nua t e dOni o nye l l o w dwa r f vi r us :ana t ur a lpo t yvi r usde l e t i o nmut a ntl a c ki ngt heNt e r mi na l9 2a mi noa c i dso fHCPr o .Ar c h.Vi r o l .1 5 1: 1 4 3 9 1 4 4 5 .2 0 0 6 . *1 *1 *1 *1 a 0 5 . ZHOU,Y. ,GUO,R. ,CHENG,Z. ,SANO,T. ,LI,S. :Fi r s tr e po r to fho ps t untvi r o i df r o m pe a c h(Pr unus pe r s i c a )wi t hda ppl ef r ui ts ympt o msi nChi na .Pl a ntPa t ho l o gy5 5:5 6 4 .2 0 0 5 . *1 *1 *1 *1 a 0 6 . QIAN. ,GUO,R. ,CHENG,Z. ,SANO,T. ,LIS. :De t e c t i o na ndmo l e c ul a rc ha r a c t e r i z a t i o no fCo l e us FU,S bl ume ivi r o i di n Chi na .Ac t a Phyt o pa t ho l o gi c a Si ni c a3 (3 6 ):2 2 6 2 3 1 .2 0 0 6 (i n Chi ne s e wi t h Engl i s h s umma r y) . *1 *1 *1 *1 *1 a 0 7 . YANG,Y. A. ,WANG,H. Q. ,GUO,R. ,CHENG,Z. ,M.LI,S. ,SANO,T.:Fi r s tr e po r to fHo ps t untvi r o i di n Apr i c o ti nChi na .Pl a ntDi s .9 0:8 2 8 .2 0 0 6 . a 0 8 . TANAKA,K. ,HATAKEYAMA,S. ,HARADA,Y.:Thr e e ne wf r e s hwa t e ra s c o myc e t e sf r o m r i ve r si n Akke s hi , Ho kka i do ,no r t he r nJ a pa n.Myc o s c i e nc e4 6:2 8 7 2 9 3 .2 0 0 5 . a 0 9 . TANAKA,K. ,HARADA :Ba mbus i c o l o usf ungii nJ a pa n(6 ):Ka t u mo t o a ,ane w ge nuso fpha e o s pha e r i a c e o us a s c o myc e t e s .Myc o s c i e nc e4 6:3 1 3 3 1 8 .2 0 0 5 . a 1 0 . ARAI,S. ,FUJITA,T. ,HARADA,Y.:Ro l eo fCy l i n d r o s p o r i u mt ypec o ni di ao fMy c o s p h a e r e l l ap o mi (Pa s s . )Li nda u: c a us eo fBr o o ksf r ui ts po to fa ppl e ,a sa ni nf e c t i o ns o ur c ei na ppl eo r c ha r ds .J .Ge n.Pl a nt .Pa t ho l .7 (5 1 ):3 5 7 3 5 9 .2 0 0 5 . *15 a 1 1 . MIZUKAMI,S. ,TAKEDA,K. ,AKADA,S. ,FUJITA,T.:I s o l a t i o na ndc ha r a c t e r i s t i c so fMe t ha no s a e t ai npa ddy f i e l ds o i l s .Bi o s c i .Bi o t e c hno l .Bi o c he m.7 (4 0 ):8 2 8 8 3 5 .2 0 0 6 . 研 究 業 績 目 録 1 16 d0 1 . 磯野清香・佐野輝男・上森隆司*3・向井博之*3:等温遺伝子増幅法(I CAN法)を用いたキク矮化ウイロイドの診 断.日本植物病理学会大会 2 0 0 6年 6月,札幌市 d0 2 . 對馬由記子*4・佐野輝男・向井博之*3・上森隆司*3:リンゴさび果病ウイロイド(Appl es c a rs ki nvi r o i d)の I CAN (I s o t he r ma la ndChi me r i cpr i me r i ni t i a t e dAmpl i f i c a t i o no fNuc l e i ca c i ds )法による検出.日本植物病理学会 大会 2 0 0 6年 6月,札幌市 d0 3 . 磯野清香・佐野輝男・忠 英一*5・上森隆司*3・向井博之*3:温遺伝子増幅法(I CAN法)を用いたキク矮化ウイ ロイドの発生圃場調査.日本植物病理学会東北部会 2 0 0 6年 9月,鶴岡市 d0 4 . 星 朱香*6・関根健太郎*6・佐藤加奈子*6・本堂大祐*6・長谷 修*6・佐野輝男・高橋英樹*6:ダイコンモザイクウ イルスのゲノム RNA構造解析と東北地方における発生分布.日本植物病理学会東北部会 2 0 0 6年 9月,鶴岡市 d0 5 . 原田幸雄・北 真弓・高橋由紀子・飯島 勉*7・難波成任*8・塩崎雄之輔・藤田 隆・佐野輝男:東京都立川市で 発見されたイチョウ樹のてんぐ巣症 6状と 2 ,3の原因調査試験.日本植物病理学会東北部会 2 0 0年 9月,鶴岡 市 d0 6 . 平田行正*9・山下裕子*9・佐野輝男・福井博一*10:I ns i t uRTLAMP法を用いたキクわい化ウイロイド(CSVd) RNA視覚化の試み.日本園芸学会大会,2 0 0 5年度 d0 7 . 佐野輝男: 「ウイロイドとは」 新潟市民大学「人類とウイルスの共存を考える」.2 0 0 5年 1 1月,新潟市 d0 8 . 佐野輝男: 「リンゴ,ナシの火傷病と果実異常症状について」(財)長野県果樹研究会 2 0周年記念会員大会,記念 講演,2 0 0 6年 3月,千曲市 d0 9 . SANO,T.:Tr a ns ge ni cpo t a t or e s i s t a nta ga i ns tmul t i pl a ntvi r usI nf e c t i o nme di a t e dbypa c 1 .I ns t i t ut eo fPl a nt Pr o t e c t i o n,Chi ne s eAc a de myo fAgr i c ul t ur a lSc i e nc e s .Be i j i ng,Chi na ,Se pt e mbe r2 0 0 6 d1 0 . SANO,T.:Vi r o i da nd i t sde t e c t i o n,Pl a ntVi r usi ns pe c t i o nc e nt e ro fChi naEnt r ya nd Exi tI ns pe c t i o na nd Qua r a nt i neBur e a u.Be i j i ng,Chi na ,Se pt e mbe r2 0 0 6 d1 1 . 鈴木陽裕・畠山 聡・田中和明・原田幸雄:シウリザクラ紅点病について.日本菌学会 2 0 0 6年 6月.千葉市 d1 2 . 和泉恵介・長尾智萌・原田幸雄:ササ・タケ類赤衣病菌 S t e r e o s t r a t u mc o r t i c i o i d e sの生活環について.日本菌学会 2 0 0 6年 6月.千葉市 d1 3 . 田中和明:日本産プレオスポラ目菌の分類学的研究.日本菌学会 2 0 0 6年 6月.千葉市 d1 4 . TANAKA,K.:Ta xo no my a nd phyl o ge ny o fpl e o s po r a l e a nf ungii nJ a pa n.Myc o l o gi c a lSo c i e t yo fJ a pa na nd Br i t i s hMyc o l o gi c a lSo c i e t yJ o i ntSympo s i um.Chi ba ,J a pa n,J une2 0 0 6 . *11 *12 *13 *14 d1 5 . OKADA,G. ,TANAKA,K. ,HUANG,J W. ,CHANG,HS. ,KAKISHIMA,M. :Synne ma t o usf ungii nTa i wa n (1 ) .Ei ght hI nt e r na t i o na lMyc o l o gi c a lCo ngr e s s ,Ca i r ns ,Aus t r a l i a ,Augus t2 0 0 6 . *11 d1 6 . TANAKA,K,OKADA,G. :Us e f ulme t ho ds f o ri nduc i ng a na mo r ph / t e l e o mo r ph (s )i nc ul t ur e .Ei ght h I nt e r na t i o na lMyc o l o gi c a lCo ngr e s s ,Ana Sa t2:Fr om Spo r et oc ul t ur e .Ca i r ns ,Aus t r a l i a ,Augus t2 0 0 6 . d1 7 . 荒井茂光*2・藤田 隆・原田幸雄:リンゴ黒星病菌偽子のう殻世代の越冬罹病落下果実上での確認.日本植物病 理学会大会 2 0 0 6年 6月,札幌市 e 0 1 . 佐野輝男: 「ウイロイド」植物病原アトラス―目で見るウイルス・細菌・菌類の世界.米山勝美他編,分担執筆, ソフトサイエンス社,2 6 1ページ,2 0 0 6年,ASI N :4 8 8 1 7 1 1 1 4 8 榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎 * 1. 中国農科院植物保護研究所 * 2. 青森県農総研県南果樹 * 3. タカラバイオ(株) * 4. 青森県農総試リンゴ試験場 * 5. 青森県農総試フラワーセンター * 6. 東北大学院農 * 7. 東京都立川市 * 8. 東京大学院農生 * 9. 和歌山県アグリバイオセンター * 10 .岐阜大農 * 11 .理化学研究所バイオリソースセンター * 12 .台湾国立中興大学 * 13 .台湾中央研究院植物研究所 * 14 .筑波大学 * 15 .農業技術総合研究所つくばセンター 研 究 業 績 目 録 117 昆虫学・進化生態学研究室 a 0 1 . TRUMAN,J . W. ,HIRUMA,K. ,ALLE,J . P. ,Ma c WHINNIE,S. G. B. ,CHAMPLIN,D. T. ,a nd RIDDIFORD,L. M. (2 0 0 6 ) . J uve ni l eho r mo nei sr e qui r e dt oc o upl ei ma gi na ldi s cf o r ma t i o n wi t h nut r i t i o ni ni ns e c t s .S c i e n c e3 1 2 ,1 3 8 5 1 3 8 8 . a 0 2 . KESHAN,B. ,HIRUMA,K. ,a nd RIDDIFORD,L. M. (2 0 0 6 ) .De ve l o pme nt a le xpr e s s i o na nd ho r mo na lr e gul a t i o no f di f f e r e nti s o f o r mso ft het r a ns c r i pt i o nf a c t o rE7 5i nt het o ba c c oho r nwo r m,Ma n d u c as e x t a .De v .Bi o l . ,2 9 5,6 2 3 6 3 2 . d0 1 . HIRUMA,K. (2 0 0 5 ) .Re gul a t i o no ft he e c dys o ne i nduc e dc a s c a de o ft r a ns c r i pt i o nf a c t o r st ha tl e a dt ot he i nduc t i o no fdo pade c a r bo xyl a s ee xpr e s s i o n.NI AS/ COE I nt e r na t i o na lSympo s i um:Pr e s e ntSt a t uso fSt udi e s f o rUt i l i z a t i o no fI ns e c tPr o pe r t i e s .Abs t r a c tp.3 1 .Oc t o be r4 5 ,2 0 0 5 .Ts ukubaI nt e r na t i o na lCo ngr e s sCe nt e r (招待講演) . d0 2 . KESHAN,B. ,HIRUMA,K. ,a ndRIDDIFORD,L. M. (2 0 0 5 ) .Thee c dys o ne i nduc e de a r l yge neE7 5i nMa n d u c as e x t a: i t sde ve l o pme nt a lr e gul a t i o na ndho r mo na lc o nt r o li ne pi de r mi sa ndde ve l o pi ngwi ng.Fr o nt i e r si nMo l e c ul a r Endo c r i no l o gy.Abs t r a c tp.2 7 .De c e mbe r1 2 1 5 ,2 0 0 5 .Hyde r a ba d,I ndi a (招待講演) . d0 3 . HIRUMA,K. (2 0 0 6 ) .An o ve r vi e w o ft he mo l e c ul a ra s pe c to fMa n d u c ae ndo c r i no l o gy.J a pa nUSA J o i nt Sympo s i um.Ho r nwo r msa sExpe r i me nt a lI ns e c t s:I nRe t r o s pe c ta ndPr o s pe c t .Abs t r a c tp.5 7 .J a nua r y2 8 , 2 0 0 6 .To kyoSi l kHa l l (招待講演) . d0 4 . 比留間潔,RIDDIFORD,L. M. (2 0 0 6 ) .エクダイソンに誘導される核レセプター,MHR3と E7 5 B,は転写因子 MHR4の調節因子であると共に,do pa de c a r bo xyl a s e発現の阻害因子である.日本応用動物昆虫学会第 5 0回 大会.講演要旨 p.3 3 .2 0 0 6年 3月 2 7 -2 9日.筑波大学. d0 5 . 金城輝則,比留間潔,三田和英,篠田徹郎(20 0 6 ).カイコのアラタ体における J H生合成酵素と転写因子 E7 5の 発現解析.日本応用動物昆虫学会第 5 0回大会.講演要旨 p.3 3 .2 0 0 6年 3月 2 7 -2 9日.筑波大学. d0 6 . TRUMAN,J . W. ,HIRUMA,K. ,ALLEE,J . P. ,Ma c WHINNIE,S. G. B. ,CHAMPLIN,D. ,a nd RIDDIFORD,L.M. (2 0 0 6 ) .A di r e c tr o l eo fJ Hi nt he c o nt r o lo fi ma gi na ldi s cf o r ma t i o na nd gr o wt hi n Ma n d u c a .Fl y Me e t i ng.2 0 0 6 Ec dys o neWo r ks ho pWe dne s da y,Ma r c h2 9 ,2 0 0 6 ,La ni e rGr a ndBa l l r o o m A,Hi l t o nAme r i c a s ,Ho us t o n,TX. d0 7 . TRUMAN,J . W. ,HIRUMA,K. ,ALLEE,J . P. ,Ma c WHINNIE,S. G. B. ,CHAMPLIN,D. ,a nd RIDDIFORD,L. M. (2 0 0 6 ) .A di r e c tr o l eo fJ Hi nt hec o nt r o lo fi ma gi na ldi s cf o r ma t i o na nd gr o wt hi n Ma n d u c a .The5 t hI nt e r na t i o na l Sympo s i um o nMo l e c ul a rI ns e c tSc i e nc e .Ma y2 0 2 4 ,2 0 0 6 .Tuc s o n,AZ. d0 8 . HIRUMA,K. ,a ndRIDDIFORD,L. M. (2 0 0 6 ) .Thec o o r di na t i o no ft hes e que nt i a la ppe a r a nc eo fMHR4a nddo pa de c a r bo xyl a s e dur i ng t he de c l i ne o ft he e c dys t e r o i dt i t e ra tt he e nd o ft he mo l t .The 1 6th I nt e r na t i o na l Ec dys o neWo r ks ho p.Abs t r a c tp.6 4 .J ul y1 0 1 4 ,2 0 0 6 .Ghe nt ,Be l gi um. d0 9 . SHINODA,T. ,KINJOH,T. ,MITA,K. ,a nd HIRUMA,K. (2 0 0 6 ) .Cl o ni ng a nd e xpr e s s i o na na l ys i so fj uve ni l e ho r mo ne bi o s ynt he t i ce nz yme sa nd E7 5i s o f o r ms i nt he s i l kwo r m,Bo mb y x mo r i .The 7th I nt e r na t i o na l wo r ks ho po n Mo l e c ul a r Bi o l o gy a nd Ge ne t i c so ft he Le pi do pt e r a .Augus t2 0 2 6 ,2 0 0 6 .Ko l ympa r i ,Cr e t e , Gr e e c e . d1 0 . 水口智江可,比留間潔,RIDDIFORD,L. M. (2 0 0 6 ).Ma n d u c as e x t a皮膚における mo l i n g遺伝子の発現誘導機構. 昆虫ワークショップ 0 6金沢.2 0 0 6年 9月 1 3 -1 5日.石川県青少年総合研修センター. d1 1 . 城田安幸,阿部練平 1):りんご(Ma l u sd o me s t i c a )の抗腫瘍効果(6 )母親がリンゴジュースを飲むことで,子供, 孫世代の免疫力が高まる.日本癌学会第 6 5回総会,2 0 0 6 .( 1)東北大学大学院 農学研究科 生物産業創成) d1 2 . 城田安幸・井戸川基規・加藤 彰・松井佳之・金子悠太:無農薬りんご園の害虫管理(2 ) 交信撹乱法によるモ モシンクイガの管理.第 5 0回日本応用動物昆虫学会大会,2 0 0 6 . f 0 1 . 城田安幸: 「免疫力を高めてガンの発生を防ぎ血糖値まで下げるとわかったリンゴジュース」壮快 1月号 P1 8 4 1 8 5 ,2 0 0 6 . f 0 2 . 城田安幸:免疫力を高める,無農薬未熟果汁入りリンゴジュース.アグリビジネス創出フェア 2 0 0 5 (ブース展示 及びプレゼン発表) ,2 0 0 5 . f 0 3 . 城田安幸:リンゴはヒトの免疫力を高める.弘前市農村活性化推進協議会主催 第 5回「食を考える集い」基調 講演,2 0 0 6 . f 0 4 . 城田安幸:リンゴと健康.青森県リンゴ対策協議会主催「在京マスコミ関係者との懇談会」基調講演, 2 0 0 6 . f 0 5 . 城田安幸:空飛ぶ目玉クワイコ- 2 5年の選択実験から見えてきたこと.日本昆虫学会 東北支部 青森県談話会 特別講演,2 0 0 5 . 研 究 業 績 目 録 1 18 動物生態学・野生生物管理学研究室 a 0 1 . SAWARA,Y.a nd K.SATOU :Fe e di ng a c t i vi t yr hyt hm o fj uve ni l ego bi i df i s h,Ch a e n o g o b i u sg u l o s u s ,a tt i da l l y di f f e r e ntl o c a l i t i e s .Bul l .Fa c .Agr .Li f eSc i . ,8 ,2 9 3 6 .2 0 0 5 a 0 2 . ENDO,N. ,Y.SAWARA,R.KOMATSU a ndM.OHTSUBO :Di e la c t i vi t ypa t t e r nso fpr e s e nc ea nda bs e nc ef r o ma he r o nr y a nd po s t br e e di ng r o o s t so bs e r ve d i n r a di o t a gge d Bl a c kc r o wne d Ni ght He r o ns Ny c t i c o r a x n y c t i c o r a x .Or ni t ho l .Sc i . ,5 ,1 1 3 1 1 9 .2 0 0 6 a 0 3 . KASAI,A. ,H.TOYOHARA,A.NAKATA,T.MIURA a nd N.AZUMA :Fo o ds o ur c ef o rt he bi va l e ve Co r b i c u l a j a p o n i c ai nt hef o r e mo s tf i s hi ng l a ke se s t i ma t e df r o ms t a bl ei s o t o pea na l ys i s ,Fi s he r i ss c i e nc e7 2:1 0 5 1 1 4 , 2 0 0 6 a 0 4 . 泉 完・矢田谷健一・東 信行・工藤 明:河川流下水を用いたスタミナトンネルによるウグイの突進速度につ いて.農業土木学会論文集 2 4 4巻,1 7 1 1 7 8 ,2 0 0 6 b0 1 . 佐原雄二:谷内ほか(編) 「魚の科学事典」3章 1節 食性.1 5 8 -1 7 1ページ.朝倉書店.2 0 0 5 b0 2 . 佐原雄二:日本陸水学会(編) 「陸水の事典」講談社.2項目,2 0 0 6 c 0 1 . 佐原雄二:青森県のメダカの歴史と現状.市史研究あおもり,8 ,1 1 2 . ,2 0 0 6 c 0 2 . 佐原雄二:汽水・淡水魚類.「青森県の希少な野生生物―青森県レッドリスト(2 0 0 6年 改訂増補版) ―」5 6 -5 8 ページ.青森県,2 0 0 6 c 0 3 . 佐原雄二:汽水・淡水魚類.「青森県外来種対策学術調査報告書―青森県外来種リスト―」3 2 -3 3ページ,6 7 - 7 1ページ.青森県,2 0 0 6 d0 1 . SAWARA,Y. ,T.ADACHI,Y.TAKATSUJIa ndM.I c l i ne si ns ma l lpi s c i vo r o usbi r dsduet oe xo t i cf i s h SOGAI:De i nJ a pa n,a nd t hei mpo r t a nc eo fhypo xi cpo ndsa saf e e di ng e nvi r o nme nt .2 4 t hI nt e r na t i o na lOr ni t ho l o gi c a l Co ngr e s s . (Ha mbur g) ,2 0 0 6 d0 2 . NUMATA,S. ,K.TAKEUCHI a nd N.AZUMA :Br e e di ng s i t epr e f e r e nc e so ft heJ a pa ne s eMa r s h Wa r bl e r si n ma na ge dr e e dbe dsi nJ a pa n.2 4 t hI nt e r na t i o na lOr ni t ho l o gi c a lCo ngr e s s . (Ha mbur g) ,2 0 0 6 d0 3 . AZUMA,N. ,A.I T.OZAKI,S.SATO a ndH.EBISAWA :Co l o ni z a t i o no fa ppl eo r c ha r dsbyUr a lowl s .2 4 t h NADA, I nt e r na t i o na lOr ni t ho l o gi c a lCo ngr e s s . (Ha mbur g) ,2 0 0 6 d0 4 . 東 信行・東 美苑・壱岐康生・水野直樹:北と南のメダカの成長と摂餌・遊泳行動の比較,平成 1 8年度日本水 産学会大会,2 0 0 6 d0 5 . 東 信行・五十嵐勇気・佐藤孝司・石井健太郎・佐原雄二:小河川における魚類群集の密度変化は温暖化による ものか?,平成 1 8年度日本水産学会大会,2 0 0 6 d0 6 . 笠井亮秀・豊原治彦・三浦常廣・東 信行:安定同位体による汽水湖におけるヤマトシジミの食性の推定,平成 1 8年度日本水産学会大会,2 0 0 6 d0 7 . 和田 実・七里朋子・古林千春・東 信行・塚本久美子・水野直樹・鈴木 譲:青森県十三湖で採集されたヒイ ラギ稚魚の発光器官における P .l e i o g n a t h iについて,平成 1 8年度日本水産学会大会,2 0 0 6 d0 8 . 伊藤 忍・東 信行:温暖化に伴うカジカ大卵型(Co t t u sp o l l u x )の成長特性の変化と個体群崩壊,応用生態工学 会 第 1 0回研究発表会,2 0 0 6 d0 9 . 山谷彩子・小西あや・百瀬麦子・東 信行:ため池における魚類および魚食性鳥類の生息場選択と環境構造,応 用生態工学会 第 1 0回研究発表会,2 0 0 6 d1 0 . 東 信行:偶然から必然へ ~偶然成り立つ野鳥との共生~ 日本鳥学会 2 0 0 6年大会公開シンポジウム「野鳥の 保護は農林業と共存可能か」 ,2 0 0 6 e 0 1 . 東 信行:平成 1 7年度岩木川左岸農業水利事業地区内生態系調査業務報告書,農林水産省東北農政局津軽農業水 利事務所,2 0 0 6 f 0 1 . 山田裕之・東 信行:津軽地方における森林性小型哺乳類の巣箱利用,青森自然誌研究 1 1 ,6 9 7 3 ,2 0 0 6 f 0 2 . 東 信行:岩木川・河川環境フォーラム記録集 ~津軽ダムで何が変わるか~,国土交通省東北地方整備局, 7 4 pp,2 0 0 5 研 究 業 績 目 録 119 地 域 環 境 科 学 科 地域環境工学講座 a 0 1 . 山下祐介*・澤田信一・田中重好**・工藤 明:河川流域社会における共通理解の構築と合意形成―岩木川流域で の試み―,環境科学会誌 1 (4 9 ):3 0 9 3 1 8 ,2 0 0 6 .7 . (*弘前大学人文学部,**名古屋大学大学院環境学研究科) a 0 2 . 管 化冰・工藤 明・泉 完:循環灌漑地区における降雨の有効利用に関する研究,日本雨水資源化システム学 会誌 1 2 1:2 5 3 0 ,2 0 0 6 .7 . a 0 3 . 泉 完・工藤 明:水クッション型落差工の流水音調査について,農業土木学会論文集,No.2 4 0:8 1 8 8 ,2 0 0 5 . a 0 4 . 泉 完・矢田谷健一*・ 東 信行・工藤 明:河川流下水を用いたスタミナトンネルによるウグイの突進速度につ いて,農業土木学会論文集,No.2 4 4:1 7 1 1 7 8 ,2 0 0 6 . (* (株)建設技術研究所東北支社) a 0 5 . 佐々木長市・松山信彦・本田和茂・殿内暁夫・野田香織・佐瀬 隆*:白神山地の土壌に関する研究(3 ),白神研 究,第 3号 ,4 5 5 2 ,2 0 0 6 . (*北方フ ァ イ ト リ ス 研究室) a 0 6 . 佐藤幸一*・徳永光一**・佐々木長市・佐瀬 隆***:岩手県北部火山灰累層の中部,下部更新統における根成孔 隙の劣化とその分級,農土論集,2 4 3号 ,1 0 1 1 1 2 ,2 0 0 6 . (*北里大学,**岩手大学名誉教授,***北方ファイトリ ス研究室) a 0 7 . Cho i c hiSASAKI :Expe r i me nt a ls t udy o nt he i nf l ue nc eo fpe r c o l a t i o n pa t t e r nso nt he r e mo va l ? o fs o l ubl e e l e me nt si ns t r a t i f i e da l l uvi a l s o i lpa ddyf i e l dsf o rr i c e ,Pa ddyWa t e rEnvi r o n,4 ,pp.8 9 9 9 ,2 0 0 6 . a 0 8 . 加藤 幸・片山寿伸*; 「食の安全」への取組みと農業土木分野の関連性,農業土木学会誌,Vol7 3/No.1 2 ,1 0 9 3 1 0 9 7 ,2 0 0 5 .1 2 . (*片山りんご有限会社・日本 GAP協会) a 0 9 . 谷口建・佐藤正彦**・藤崎浩幸・加藤 幸;地域資源と人材による町おこし,農業土木学会誌,Vol7 4/No.1 , 1 1 1 5 ,2 0 0 6 .1 . (** NPO法人尾上蔵保存利活用促進会) c 0 1 . 工藤 明・管 化冰:降雨時の流出負荷量に関する研究,東北地域災害科学研究 第 4 2巻 :3 5 4 0 ,2 0 0 6 .3 . d0 1 . 管 化冰・工藤 明・泉 完:循環灌漑地区における降雨の有効利用に関する研究,日本雨水資源化システム学 会第 1 3回研究発表会講演要旨 :8 1 8 6 ,2 0 0 5 .1 1 . d0 2 . 管 化冰・工藤 明・泉 完:水田地帯における降雨時の流出負荷について,平成 1 8年度農業土木学会大会講演 要旨 7 3 4 ,2 0 0 6 .8 . d0 3 . 伊東竜太・矢田谷健一・ 泉 完・ 工藤 明・ 東 信行:全面越流型階段式魚道における魚類等の遡上特性―岩木川取 水堰魚道の事例―,平成 1 7年度農業土木学会応用水理研究部会講演集,1 6 ,2 0 0 5 .1 2 . d0 4 . 矢田谷健一・ 伊東竜太・泉 完・ 工藤 明・ 東 信行:自然河川水を用いた現地実験における淡水魚の突進速度と遊 泳距離,平成 1 7年度農業土木学会応用水理研究部会講演集,7 1 2 ,2 0 0 5 .1 2 . d0 5 . 泉 完・ 工藤 明・ 東 信行・矢田谷健一・ 伊東竜太:自然条件下におけるスタミナトンネルを用いた淡水魚の突進 速度,平成 1 8年度農業土木学会大会講演要旨,1 1 6 1 1 7 ,2 0 0 6 .8 . d0 6 . 泉 完・ 工藤 明・ 東 信行・伏見佳奈子:粗石付き斜路型魚道における魚類等の遡上実態とその行動,平成 1 8年 度農業土木学会大会講演要旨,6 5 2 6 5 3 ,2 0 0 6 .8 . d0 7 . 佐々木長市・野田香織・成田瑞季・坂口良介・川島一就:鉱物由来の Cd汚染水田土壌モデルの浸透型が稲の重 金属吸収に及ぼす影響,農業土木東北支部大会,1 1 8 1 1 9 ,2 0 0 5 d0 8 . 佐々木長市・B.K.MITRA・江成敬次郎*・藤田光則*・山本太平**:屏風山砂丘畑における地下水の水質変動,日 本砂丘学会第 5 3回全国大会,2 2 2 3 ,2 0 0 6 . d0 9 . Cho i c hiSASAKI,To mo ka TAKIZIRI,No buhi ko MATUYAMA,Bi j o n Kume r MITRA,Le pi ng YUE* ,Ko hi c hi TOKUNAGA** ,Ko ui c hiTAKEYAMA*** :St udi e so nphys i c a la ndc he mi c a lpr o pe r t i e so fa e r o l i a ns o i la tXi ’ a n i nChi na ,中日拠点大学交流シンポジウム要旨,3 1 3 6 ,2 0 0 6 . (*西北大学,**岩手大学名誉教授,***島根大学) d1 0 . 佐々木長市・上西訓生・野田香織・川島一就・松山信彦・成田瑞季:カドミウム汚染水田模型の浸透型が物質動 態および水稲に及ぼす影響(Ⅳ) ,平成 1 8年度農業土木学会講演要旨,9 7 4 9 7 5 ,2 0 0 6 . d1 1 . 角野三好,五十嵐悠也,吉田和史*,ヘルショーモデルによる傾斜遮水ゾーン型フィルダムの浸潤線について(2 ) , 農業土木学会大会,2 0 0 6 .8 . (*福島県土地改良連合会) d1 2 . 加藤 幸・片山寿伸 ほか,映像によるトレーサビリティ情報を活用した食の安全の確保に関する研究,農業土 木学会大会,2 0 0 6 .8 . (*片山りんご有限会社・日本 GAP協会) d1 3 . 万木正弘・渡部均*・渡邉賢三**,炭酸イオンを含む溶液のモルタル侵食性試験,農業土木学会大会,2 0 0 6 .8 . (* 東 北農政局,**鹿島技術研究所) 研 究 業 績 目 録 1 20 e 0 1 . 工藤 明:新城下堰浄化施設の効果検証,全 4 8頁,2 0 0 6 .3 . e 0 2 . 工藤 明:環境負荷軽減水管理技術確立調査報告書,全 3 7頁,2 0 0 6 .3 . e 0 3 . 泉 完:岩木川左岸幹線水路落差工の流水音調査報告書,全 5 4頁,2 0 0 6 .3 . e 0 4 . 万木正弘:循環型社会の構築に向けた水利構造物の維持管理,平成 1 7年度東北農政局管内農業農村整備事業推 進方策検討業務報告書,農業土木学会,2 0 0 6 . f 0 1 . 佐々木長市:オーストラリアの世界遺産を見て,白神研究,第 3号,1 1 1 3 ,2 0 0 6 . 地域環境計画学講座 a 0 1 . 谷口 建・佐藤正彦・藤崎浩幸・加藤 幸:地域資源と人材による町おこし,農業土木学会誌,7 4 1 ,1 1 1 5 ,2 0 0 6 . a 0 2 . 藤崎浩幸・山路永司:標準区画から脱却し創造的な大区画圃場を目指せ!,農業土木学会誌,74 9 ,8 0 1 8 0 4 ,2 0 0 6 . a 0 3 . 藤崎浩幸:認定農業者親子間の就農誘導と就農意欲―岩手県盛岡市での意識調査―,弘前大学農学生命科学部学 術報告,8 ,5 2 5 7 ,2 0 0 5 . a 0 4 . 小笠原康雄・加藤陽治・藤崎浩幸:青森県の転作水田におけるアピオス栽培の可能性,弘前大学農学生命科学部 学術報告,8 ,4 6 5 1 ,2 0 0 5 . a 0 5 . 小原嬢子・森屋 洋・桧垣大助:秋田県谷地地すべり地における地すべりの発達過程からみた微地形と内部構造. 地形,2 (3 7 ) ,3 0 1 3 1 8 ,2 0 0 6 . a 0 6 . GHIMIRE,S.K. ,HIGAKI,D.a ndBHATTARAI,T. P. ,2 0 0 6:Gul l ye r o s i o ni nt heSi wa l i kHi l l s ,Ne pa l:Es t i ma t i o no f s e di me ntpr o duc t i o nf r o ma c t i vee phe me r a lgul l i e s .Ea r t hSur f a c ePr o c e s s e sandLa ndf o r ms ,3 1 ,1 5 5 1 6 5 ,2 0 0 6 . a 0 7 . 桧垣大助・宮城豊彦・八木浩司・千葉則行・森屋 洋:2 0 0 6年大舟木地区で発生した地すべり災害.地すべり, 4 (2 3 ) ,3 8 4 0 ,2 0 0 6 . d0 1 . 高橋照夫:ステレオ視法によるリンゴ園果実群の三次元位置計測(第 2報).農業環境工学関連 7学会 2 0 0 6年合 同大会講演要旨集,CDROM,2 0 0 6 .9 . d0 2 . ガウリ・マハルジャン・高橋照夫:画像特徴を利用した植物病名の検索システムに関する研究―イネの病気の色 特徴による分類と検索法に関する検討―.農業環境工学関連 7学会 2 0 0 6年合同大会講演要旨集,CDROM, 2 0 0 6 .9 . d0 3 . 桧垣大助・小原嬢子・高橋明久:秋田県砥沢地すべり地における地形と移動体内部構造.第 4 5回日本地すべり学 会研究発表会講演集,6 3 6 6 .2 0 0 6 . d0 4 . 小原嬢子・桧垣大助:秋田県南部珪質泥岩地域における地すべり地の微地形と内部構造,第 4 5回日本地すべり学 会研究発表会講演集,6 7 7 0 .2 0 0 6 . e 0 1 . 小林裕志・谷口 建・他 4名:干拓地を対象に農業と野生生物との共生を意図した環境教育牧場創出の実証実験, 平成 1 5 -1 7年度科学研究費補助金(基盤研究(B) (2 )),1 0 8 1 8 1 ,2 0 0 6 . e 0 2 . 谷口 建・藤崎浩幸:弘前市のグリーン・ツーリズムに関する調査研究,平成 1 7年度弘前市委託研究報告書,1 4 3 ,2 0 0 6 . e 0 3 . 谷口 建・田村義夫・田村えり子・佐々木義博:グリーン・ツーリズム情報受発信コアセンター・地域サテライ トネットワークモデル構築研究,平成 1 7年度青森県農林水産部構造政策課委託研究報告書,1 3 2 ,2 0 0 6 . e 0 4 . 谷口 建・つがる里山体験塾:北のまほろば ぐるっと青森グリーン・ツーリズムマップ,2 0 0 6 . f 0 1 . 小原嬢子・桧垣大助:平成 1 7年度砂防学会ワークショップ成果概要報告「不安定斜面の自然環境の多様性とそれ を考慮した対策計画」 ,砂防学会誌,5 (1 9 ) ,6 1 ,2 0 0 6 . 地域資源経営学講座 a 0 1 . 宇野忠義:最重要病害リンゴ火傷病の日米検疫問題.弘前大学農学生命科学部学術報告 第 8号 :8 8 1 0 1 ,2 0 0 5 . 1 2 . a 0 2 . 宇野忠義:最重要病害リンゴ火傷病の日米検疫問題の交渉経過と残された問題.農業・農協問題研究 第 35号 :7 5 8 9 (農業・農協問題研究所,2 0 0 6 .8 ) a 0 3 . 高橋秀直:住宅解体材リサイクルの現状と課題.弘前大学経済研究 第 2 8号 :1 2 3 2 ,2 0 0 5 .1 2 . a 0 4 . 小林俊介・澁谷長生:大鰐温泉もやしに関する研究.弘前大学農学生命科学部学術報告,第 8号,6 4 -8 7頁, 2 0 0 5 ,1 2 . a mi I a 0 5 . Mas a r a c t e r i s t i c s of t he Re c yc l i ng Cha nne l sf o r Appl e Po ma c e .BULLETI N of t he ZUMI YA :Ch 研 究 業 績 目 録 121 FACULTY OF AGRI CULTURE AND LI FE SCI ENCE HI ROSAKIUNI VERSI TY(8):5 8 6 3 ,2 0 0 5 . b0 1 . 神田健策: 「新編 弘前市史」編纂委員会編集:新編 弘前市史 通史編 4 (近・現代 1 )』(農業史担当) ,弘前 市,2 0 0 5 .1 0 . b0 2 . 神田健策: 「新編 弘前市史」編纂委員会編集:新編 弘前市史 通史編 5 (近・現代 2 )』(農業史担当) ,弘前 市,2 0 0 5 .1 1 . b0 3 . 神田健策:特産物(こかぶ)生産で地域農業の活性化をはかる小規模農協―青森県野辺地町農協の事例―.吉田・ 渡辺・大木・西山編:食と農を結ぶ協同組合,5 3 7 1 ,筑波書房,2 0 0 6 .6 . b0 4 . 澁谷長生:宮城県における産直の新しい挑戦.吉田・渡辺・大木・西山編:食と農を結ぶ協同組合,5 3 7 1 ,筑波 書房,2 0 0 6 .6 . b0 5 . 平石学*・志賀永一**・泉谷眞実・板橋 衛***:十勝農業の変動と地域農業の対応.牛山敬二・岩崎 徹編著: 北海道農業の地帯構成と構造変動.北海道大学図書刊行会,3 0 7 3 2 3 ,2 0 0 6 .*北海道立中央農業試験場,**北海 道大学,***広島大学 d0 1 . 宇野忠義:中国産りんご果汁輸出の急増が日本のリンゴ経営に与えた影響.第 3回中日学術研究会,中国内蒙古 フフホト市,2 0 0 6 .8 . d0 2 . 神田健策:戦後日本資本主義と農業 その現段階と協同組合運動.瀋陽農業大学学術講演会,中国瀋陽市,2 0 0 5 . 1 1 . d0 3 . 宋 暁凱・神田健策:生産過剰期における中国りんご生産・流通構造に関する研究 ~山東省栖霞市の事例~ 日本農業市場学会 2 0 0 6年度大会個別報告,弘前大学 2 0 0 6 .7 . d0 4 . 宋 暁凱・神田健策:中国におけるりんご流通と卸売市場の設立~山東省・栖霞市の事例~.日本流通学会北海 道・東北部会研究例会,宮城学院女子大学,2 0 0 6 .9 . d0 5 . K.KANDA ;Re c e ntc ha nge si nt heJ a pa ne s es o c i e t ya ndt her o l eo fno npr o f i ta ndc o o pe r a t i veo r ga ni z a t i o ns . Co o pe r a t i veSe mi na r ,Mi dSwe de nUni ve r s i t y.2 0 0 6 .9 . d0 6 . K.KANDA ;Cur r e ntChi naa ndt hes i t ua t i o no fCo o pe r a t i ve s .Co o pe r a t i veSe mi na r ,Mi dSwe de nUni ve r s i t y. Os t e r s und,2 0 0 9 .9 . d0 7 . 森 久綱*・泉谷眞実:食品加工残渣の飼料利用におけるリサイクル経路に関する考察.日本流通学会,名古屋大 学,2 0 0 5 .1 0 .*三重大学 d0 8 . 泉谷眞実:農業静脈市場における需給調整様式の多段階性.日本農業市場学会個別報告,弘前大学,2 0 0 6 .7 . d0 9 . 杉村泰彦*・泉谷眞実:青果物卸売市場における残品発生の実態とその発生要因.2 0 0 6年度日本農業市場学会大会 個別報告,弘前大学,2 0 0 6 .7 .*酪農学園大学 e 0 1 . 青森県 6次産業研究会(代表 神田健策)『青森県農林水産振興のための第 6次産業ビジョン策定事業報告書』 (平成 1 7年度 むつ小川原地域・産業振興プロジェクト事業),1 5 8 ,2 0 0 6 .4 . e 0 2 . 武田共治・谷田親彦:弘前大学 4年生の視点に基づく 2 1世紀教育の成果と課題.2 1世紀教育フォーラム.創刊 号,4 1 5 2 ,弘前大学 2 1世紀教育センター,2 0 0 6 .3 . e 0 3 . 泉谷眞実:農業静脈市場に関する主要文献と論点.玉真之介研究代表:農業市場の制度問題と分析モデルに関す る統合的研究,基盤研究(B)報告書(初年度).8 9 1 0 0 ,2 0 0 6 . e 0 4 . 泉谷眞実:農業(雇用)労働市場に関する主要文献と論点.玉真之介研究代表:農業市場の制度問題と分析モデ ルに関する統合的研究,基盤研究(B)報告書(初年度) .1 0 8 1 1 9 ,2 0 0 6 . f 0 1 . 宇野忠義:火傷病防疫指針(案)にもの申す.りんごニュース第 2 6 6 5号:青森県りんご協会,2 0 0 6 .1 .2 5 . f 0 2 . 宇野忠義:最重要病害リンゴ火傷病の日米検疫問題の交渉経過と残された問題.りんご火傷病研究報告会報告資 料,1 1 1 ,(津軽農民組合,藤崎町文化センター,2 0 0 6 .6 ) f 0 3 . 神田健策:中国三農問題と農産物流通・販売の課題.りんご振興研究センター発足記念シンポジウム・弘前大学 農学生命科学部,2 0 0 6 .1 2 . f 0 4 . 泉谷眞実:農業未利用資源をもちいた地域づくりの方法と課題.弘前大学農学生命科学部公開講座,20 0 5 .10 ,弘 前大学. 研 究 業 績 目 録 12 2 生物共生教育研究センター a 0 1 . 村山成治・松山信彦・齋藤 司:低投入型稲作に関する研究 第 6報 アイガモ除草を導入した不耕起移植栽培 における雑草発生率と水稲の生育・収量.日本作物学会東北支部会報 4 7:2 1 2 3 ,2 0 0 5 . a 0 2 . 伊藤大雄・杉浦俊彦 1・黒田治之 2:ニホンナシ園における蒸発散速度の解明と日蒸発散量の推定.農業気象 6 2 (1 ) ,2 3 3 2 (2 0 0 6 ) .( 1農業・生物系特定産業技術研究機構,2鹿児島大学農学部) d0 1 . Takur oKIKUCHIa nd Yuno s ukeSHIOZAKI:Fl a t Ca no pi e d Ope n Ce nt e rTr e e s :A Ne w Di r e c t i o nf o rAppl e Or c ha r dSys t ems .2 7 t hI nt e r na t i o na lHo r t i c ul t ur a lCo ngr e s sa ndExhi bi t i o n.Wo r ks ho p1 7:Or c ha r dSys t ems . 7 1 .Se o ul ,Ko r e a (2 0 0 6 ) . d0 2 . 塩崎雄之輔・藤田知道・笹亜希子・木村錦昭:リンゴの白根と新梢の生長に及ぼす台木の種類と着果の影響.園 芸学会東北支部平成 1 8年度大会 研究発表要旨 1 9 2 0 (2 0 0 6 ) . d0 3 . 塩崎雄之輔・村木裕俊・藤田知道:豪雪によるリンゴの枝折損被害と樹の仕立て方の関係,並びにネズミの食害 誘発.園芸学会東北支部平成 1 8年度大会 研究発表要旨 2 1 2 2 (2 0 0 6 ) . d0 4 . 岩渕伸雄・村山成治・松山信彦・工藤啓一:低投入型稲作に関する研究 第 7報 リンゴジュース搾り粕の施用 方法と抑草効果の関係.日本作物学会東北支部講演会,2 0 0 6 . d0 5 伊藤大雄・小笠原清訓・藤田 隆・澤田 勲:農薬不使用リンゴ園における転換 2~ 3年目の病害虫発生と収量・ 品質.園学要旨平 1 8東北支部,2 3 2 4 (2 0 0 6 ) . f 0 1 . 塩崎雄之輔:リンゴ 3 0年余の施肥試験と草生栽培から学んだもの.藤崎農場公開講座「リンゴを科学する」平成 1 7年度テキスト.5 2 5 9 . (2 0 0 5 ) f 0 2 . 塩崎雄之輔:これからのリンゴの栽培方式について考える(記念講演要旨).第 5 3回全国りんご研究大会 北海 道大会プログラム.2 1 2 6 (2 0 0 6 ) . f 0 3 . 伊藤大雄:リンゴの減農薬・無農薬栽培に向けたあの手この手.「りんごを科学する」平成 1 7年度テキスト,1 8 (2 0 0 5 ). 榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎榎 農学生命科学部学術報告編集委員会 委 員 長: 姫 野 俵 太 ( 応 用 生 命 工 学 科 ) 原 田 竹 雄 ( 生 物 機 能 科 学 科 ) 藤 田 隆 ( 生 物 生 産 科 学 科 ) 泉 完 ( 地 域 環 境 科 学 科 ) 2006年12月20日 印刷 2006年12月26日 発行 編集兼発行者 弘前大学農学生命科学部 〒036−8561 弘 前 市 文 京 町 3 印 刷 所 やまと印刷株式会社 〒036−8061 弘前市神田4丁目4ノ5 Published by Faculty of Agriculture and Life Science, Hirosaki University 3 Bunkyo-cho, Hirosaki-shi, Aomori-ken 036-8561, Japan 26 December 2006 Printed by Yamato Printing Co. 4-4-5 Kanda, Hirosaki-shi, Aomori-ken 036-8061, Japan 20 December 2006 BULLETIN OF THE FACULTY OF AGRICULTURE AND LIFE SCIENCE, HIROSAKI UNIVERSITY Number 9 December, 2006 CONTENTS Norimasa MATSUOKA and Yoshiyuki YANAGINO : Genetic distance between populations of the cyprinid fish Tribolodon hakonensis 1 Norimasa MATSUOKA : Phylogenetic relationships of four families of the Asteroidea based on allozyme 8 Wataru YOSHIDA, Tetsuya ONAKATOMI and Sachiko ISHIDA : Relation between the growth of complex plate ossicle and the body length of sea cucumber Apostichopus japonicus 15 (20)* Kiyoshi TAKEDA, Ikue SUDA and Akio TONOUCHI : Methane fermentation of an apple pomace Characteristics of a pectin・xylan degrading-anaerobic bacterium isolated from methane fermentation sludges 21 (27) Takenori ASADA : A study of differentially located apple tree spurs 28 (66) Tadayoshi UNO : The rapaid increase in import of apple juice has struck a heavy blow at Japanese apple farming 68 (79) Tadayoshi UNO, Teruo SANO and Kazuaki TANAKA : Apple production and pests management in New Zealand 80 (87) Hiroyuki FUJISAKI : Consumer's attitude toward safe vegetables By an attitude survey of customer at MIYAGI-COOP. toward its store brand : "SANCHOKU-YASAI" (Directly Delivered Vegetables) 91 (96) *English Summary Lists of Published Research Works of the Faculty of Agriculture and Life Science, Hirosaki University, 2005 (October) ― 2006 (September) 101
© Copyright 2026 Paperzz