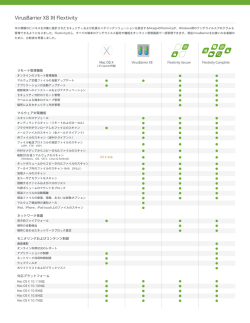
695 原 著 Mycobacterium avium complex 感染マクロファージにおける フォスフォリパーゼ A2 の細胞内動態 1) 島根大学医学部微生物免疫学,2)同 耳鼻咽喉科学 佐野 安元 千晶1)2) 清水 剛1) 冨岡 利朗1) 治明1) 多田納 豊1) (平成 19 年 4 月 12 日受付) (平成 19 年 7 月 23 日受理) Key words : Mycobacterium avium complex, macrophage, phospholipase A2 要 旨 著者らは,これまでの一連の検討で,マクロファージ(MΦ)内での Mycobacterium avium complex(MAC) 殺菌には,細胞質 phospholipase A2(cPLA2)が重要な役割を演じている可能性を示唆する成績を得ている が,今回は cPLA2 がどのような形で MΦ 内での MAC 殺菌メカニズムに関わっているのかをさらに詳しく 調べる目的で,MAC を貪食した MΦ 内での cPLA2 の動態について若干の検討を行った.その結果,(1)腹 腔 MΦ 内では,cPLA2 は MAC 感染の有無にかかわらず構成的に発現しており,MAC 感染による発現増強 は認められないこと,他方,(2)RAW264.7 MΦ に MAC を感染させた場合では,cPLA2 の一部は細胞内に 貪食された MAC 菌体と一致して局在していること,(3)MAC 感染により RAW264.7 MΦの細胞質内での cPLA2 の発現とリン酸化反応が有意に増強されることが明らかになった.これらの成績は,RAW264.7 MΦ では,MAC 感染に伴って cPLA2 の活性化と細胞内に取り込まれた MAC 菌体周辺への translocation が起 こることを示唆している. 〔感染症誌 序 81:695∼699,2007〕 る活性化シグナルにより誘導される MΦ アポトーシ 文 マクロファージ(MΦ)の細菌に対する増殖阻害・ スに連動して,MΦ 内に局在する結核菌や BCG に対 殺菌作用に関わるエフェクター分子としては,活性酸 する殺菌作用が強く誘導されることが報告されている 素分子種(ROI) ,活性酸化窒素分子種(RNI) ,リソ が,この MΦ 殺菌能も ROI や RNI には依存しないも ソーム内の殺菌ペプチド・蛋白およびリソソームの加 のとされている5).また最近,結核菌により誘導され 水分解酵素などが知られている1)2).ところが,代表的 る肉芽腫に強く発現するケモカインであるオステオポ な細胞内寄生菌である結核菌,Mycobacterium bovis ンチンは MΦ の BCG に対する抗菌活性発現に重要な BCG,Mycobacterium avium complex(MAC)などの 役割を演ずるが,このオステオポンチンによって up- 抗酸菌についてみると,一般に MΦ 殺菌能には RNI regulate される MΦ 抗菌活性もまた RNI には依存し 依存性メカニズムのかかわりが重要であると考えられ ていないことが報告されている6).こうしたことから, ており,MΦ 殺菌能の発現における ROI の関与につ 結核菌,BCG 菌,MAC などの抗酸菌に対する MΦ 1) 3) いては否定的な報告が多い .然しながら,最近 nitric 殺菌能・増殖阻害能にも ROI や RNI に依存しないメ oxide synthase 2 欠損マウスの MΦ の MAC に対する カニズムの寄与が考えられる. 抗菌活性は,wild type マウスの MΦ のそれとは変わ これまで著者らは,特に結核菌や MAC などの病原 らないことが報告されており,MAC の殺菌には RNI 性抗酸菌に対する MΦ の抗菌活性の発現における遊 の関与も部分的なもの過ぎないという可能性が示唆さ 離脂肪酸(FFA)のかかわりについて検討して来て 4) れている .さらに,ATP の P2X7 レセプターを介す 別刷請求先:(〒693―8501)出雲市塩冶町 89―1 島根大学医学部微生物免疫学 冨岡 平成19年11月20日 いるが,MΦ 内抗酸菌殺菌メカニズムにおいては,RNI と FFA との協同作用の果たす役割が最も大きいこと 治明 を示唆する成績が得られている7)∼9).すなわち,MΦ 696 佐野 千晶 他 のファゴソーム内局在 MAC に対するアラキドン酸の なわち,抗 MAC マウスモノクローナル抗体(Chemi- 殺菌作用発現には IV 型細胞質フォスフォリパーゼ A2 con International 社,米国) (1 : 500)で一次染色後, (cPLA2)が重要であること,さらに抗酸菌に感染し Alexa Fluor 546 標識抗マウス IgG1 ヤギ抗体(Mo- た MΦ では,実際に共焦点レーザー顕微鏡での観察 lecular Probes 社,米国)で二次染色を行い,共焦点 で cPLA2 のファゴソーム膜への translocation が認め レーザー顕微鏡下での細胞内蛍光強度(励起光,543 られること,またこうした反応は,ATP による purino- nm ; emission band pass,585 nm)の観察を行った. 2+ ceptor の刺激で細胞内 Ca 濃度を増加させることに 8) 9) よって誘導されることが明らかになっている .そこ 4.cPLA2 のリン酸化プロフィール RAW264.7 MΦ の 1×107 個を 1% FBS-RPMI培地 で,cPLA2 がどのような形で MΦ 内での MAC 殺菌 に浮遊させたものを径 90mm の細胞培養ディッシュ メカニズムに関わっているのかをさらに詳しく調べる に植えて 1 夜培養後,2% FBS-HBSS で洗浄し,MΦ 目的で,MAC を貪食した MΦ 内での cPLA2 の動態 単層培養を得た.これを供試菌(1×108)を浮遊させ について若干の検討を行った. た 1% FBS-RPMI 培地中で 37℃,2 時間培養後,PBS 材料と方法 で洗浄し,次いで,Lysis buffer(20 mM Tris-HCl, 1.供試菌 pH 8.0,0.1 M NaCl,1 mM EDTA,0.5% わが国の MAC 症患者より分離された Mycobacterim P-40,30 mM sodium pyrophosphate,0.2 mM バナ Nonidet intracellulare N-260 株(血清型 16)の 7H9 培地培養菌 ジウム酸,10µg! mL を供試した. その 15,000 x g,15 分遠心上清(cell lysate)を得た. 2.MΦ 単層培養 leupeptin)で細胞を溶解して, これを SDS-PAGE(7% アクリルアミド)にかけ,セ BALB! c マウスより採取したザイモザン A 誘導腹 腔 滲 出 細 胞(PEC)(5×106)ま た は RAW264.7 ミドライ式ブロッティング装置を用いてニトロセル マ ロース膜への blotting を行った.次いで,Qiu ら10)の ウス MΦ 細胞株(1×106)を浮遊させた 5% 牛胎児血 ゲルシフトアッセイ法に準じて,抗 cPLA2 ポリクロー 清(FBS)加 RPMI 1640 培地(腹腔 MΦ)または 1% ナル抗体(Santa Cruz 社)による染色と ECL 法によ FBS 加 RPMI 培 地(RAW264.7 MΦ)5mL を,予 め る発光により cPLA2 バンドとリン酸化 cPLA2 バンド カバーグラス(18×18mm)を沈めた組織培養用 6 ウェ を検出した. ルカルチャープレートに加え,炭酸ガスフラン器内, 結 果 37℃ で 1 夜 培 養 を 行 い,2% FBS 加 ハ ン ク ス 氏 液 まず,MAC 感染 MΦ での細胞内 cPLA2,sPLA2 発 (HBSS)で洗浄して非付着細胞を除くことにより MΦ 現プロフィールについて検討した.Fig. 1a,1bに示 すように,マウス腹腔 MΦ の場合,cPLA2 は非感染 単層培養を得た. 3.cPLA2 の MΦ 内局在 対照 MΦ でも細胞質全体に均等にかつ構成的に発現 mL)を浮遊させた MΦ 単層培養を供試菌(1×10 ! 7 しており,MAC 感染により cPLA2 発現レベルがさら 5%(腹 腔 MΦ)ま た は 1%(RAW264.7 MΦ)FBS に増強されると言った現象はみられなかった.また, 加 RPMI 培地中で 37℃,2 時間培養し,リン酸緩衝 この場合には細胞内感染 MAC 菌体へ局在する傾向も 生理食塩水(PBS)で洗浄後,4% パラホルムアルデ 認められなかった(Fig. 1c,1d) .他方,Fig. 1e,1f ヒドで固定した.固定後のカバーグラスを 50 mM NH4 に示すように,マウス腹腔 MΦ での sPLA2 発現は,非 Cl-PBS でクエンチング処理,次いで 0.2% Triton X- 感染 MΦ ではほとんど認められなかったが,MAC 感 100-PBS で処理後,さらに 2% 牛血清アルブミン―PBS 染により軽度の発現誘導が起こることが分かった.な でブロッキングを行った.次に,抗 cPLA2 マウスモ おこの場合,sPLA2 は細胞質全体に均一に発現してお ノクローナル抗体(Santa Cruz 社,米国) (1 : 100)ま り,特定のオルガネラに局在する傾向は認められな たは抗 IIa 型分泌性 PLA2(sPLA2)マウスモノクロー かった(Fig. 1g,1h) . ナル抗体(Santa Cruz 社)で一次染色を行い,0.05% 次に,RAW264.7 MΦ を用いた系で,MAC 感染後 Tween 20-PBS で洗浄後,さらに FITC 標識抗マウス の cPLA2 の細胞内局在プロフィールについて検討し IgG2b 抗体(Cybus Biotechnology Hants 社,米国) た.Fig. 1i,1jに 示 す よ う に,MAC 感 染 MΦ で は, (1 : 100)で二次染色を行い,洗浄後,共焦点レーザー cPLA2 は細胞質内に均等に分布していたが,一部は細 顕微鏡(Olympus Fluoview FV300,オリンパス光学, 胞内に貪食された MAC の菌体と一致して局在してい 東京)下での MΦ 細胞内の蛍光強度(励起光,488 nm ; る像(矢印)が観察された.このことは,MAC 感染 emission band pass,505-530 nm)の観察を行った. 後の MΦ では,細胞質内に分布していた cPLA2 の一 なお実験によっては,上記の染色に加えて,抗 MAC 部は MAC の局在するファゴソームに translocate し 抗体による二重染色を以下の如くの手順で行った.す ていくことを示唆している. 感染症学雑誌 第81巻 第 6 号 MAC 感染 MΦ とフォスフォリパーゼ A2 動態 697 Fi g.1 I nt r a c e l ul a re xpr e s s i o na nddi s t r i but i o no fc PLA2a nds PLA2i nmur i nepe r i t o ne a lma c r o pha ge s( MΦs )a ndRAW 2 6 4 . 7 at oh: Pe r i t o ne a lMΦswi t h( c , d, g, h)o rwi t ho ut( a , b, e , f )MAC MΦsme a s ur e dbyc o nf o c a li mmuno f l uo r e s c e nc emi c r o s c o py. i nf e c t i o nwe r es t a i ne dwi t ha nt i c PLA2a nt i bo dy( a , c )o ra nt i s PLA2a nt i bo dy( e , g) . I npa ne l sb, d, f , a ndh, c e l lmo r pho l o gyi s a s t( DI C)i ma gi ng.ia ndj:MACi nf e c t e dRAW 2 6 4 . 7MΦswe r es t a i ne dwi t he i t he r s ho wni ndi f f e r e nt i a li nt e r f e r e nc ec o nt r a nt i c PLA2a nt i bo dy( i )o ra nt i MACa nt i bo dy( j ) . Fi g.2 Pr o f i l e so fi nt r a c e l l ul a re xpr e s s i o na ndpho s pho r yl a t i o no fc PLA2i nRAW2 6 4 . 7ma c r o pha ge swi t h( l a ne s2 a nd4 )o rwi t ho ut( l a ne s1a nd3 )MAC i nf e c t i o nme a s ur e d by We s t e r n bl o ta na l ys i s .I nl a ne s3a nd 4 ,t he a mo unto fpr o t e i nss ubj e c t e dt oe l e c t r o pho r e s i swa s t wi c et ha ti nl a ne s1a nd2 . I nl a ne s3a nd4 , ba ndsc o r r e s po ndi ng t o pho s pho r yl a t e d c PLA2 ( Pc PLA2) a r e c l e a r l yde t e c t a bl e . 増強されることが分かった(lane 4) . 考 察 今回は,MAC を貪食し た MΦ 内 で の cPLA2 の 動 態について若干の検討を行った.その結果,(1)腹腔 MΦ 内では,cPLA2 は MAC 感染の有無にかかわらず 構成的に発現しており,MAC 感染による発現増強は 認められないこと,他方, (2) RAW264.7 MΦ に MAC を感染させた場合では,cPLA2 の一部は細胞内に貪食 された MAC 菌体と一致して局在していること,(3) MAC 感 染 に よ り RAW264.7 MΦの 細 胞 質 内 で の cPLA2 の発現とリン酸化反応が有意に増強されること 次に,MAC 感染後の MΦ での cPLA2 の リ ン 酸 化 が明らかになった.これらの成績は,少なくとも RAW プロフィールについて検討した.Fig. 2に示すように, 264.7 MΦでは,MAC 感染に伴って,cPLA2 発現増強 RAW264.7 MΦ を用いた系では,非感染 MΦ でも構 と cPLA2 のリン酸化,すなわち活性化が起こり,ひ ,これ 成的な cPLA2 の発現が認められるが(lane 1) いては細胞内に取り 込 ま れ た MAC 菌 体 へ translo- は MAC 感 染 で 有 意 に 増 強 さ れ た(lane 2) .他 方, cate することを示唆している. PAGE ゲルへの cell lysate タンパクの apply 量を 2 倍 先に著者らは,3H 標識アラキドン酸(3H-AA)で に増してみると,明確な cPLA2 バンドのシフト(リ ラベルした MΦ では,結核菌感染後の時間を追って , ン酸化 cPLA2 のバンド)が認 め ら れ た が(lane 3) 放射活性が細胞内局在結核菌の周辺に translocate し このリン酸化 cPLA2 のバンドは MAC 感染で有意に ていくこと,さらに,この現象は MΦ の PLA2 活性と 平成19年11月20日 698 佐野 千晶 他 貪食能に依存性であることを見い出している8).さら に,こ の 放 射 活 性 の translocation は,(1)cPLA2 阻 害剤である arachidonyl trifluoromethylketone や, (2) MΦ 貪食能の阻害剤 colchicine によって強く抑制され るが,(3)sPLA2 の阻害剤である manoalide によって 3 Hはそのような抑制は認められないこと,さらに, (4) AA と結核菌とを混合して incubate すると,経時的 に 3H-AA の菌体への binding が認められることなど も明らかになっている8)9).これらの知見は,MΦ のファ ゴソーム膜のリン脂質から cPLA2 の酵素作用により ファゴソーム内に遊離されたアラキドン酸がファゴ ソーム内に局在する結核菌に対して攻撃的に働くこと を強く示 唆 す る も で あ る が,MAC 感 染 に 伴 っ て, cPLA2 発現増強と cPLA2 のリン酸化,すなわち活性 化が起こると言う今回の成績は,このような考え方と 良く符合している.なお,腹腔 MΦ の場合は,RAW 264.7 MΦ の場合とは異なり,MAC 感染に伴う cPLA2 発現の増強は認められなかったが,これが供試 MΦ の種類の違いに起因するものなのか否か,あるいは他 の理由によるものなのか否かについては今後の検討が 必要である. ところで,今回の腹腔 MΦ を供試して行った検討 では,MAC 感染による sPLA2 の発現増強が認められ た(Fig. 1) .元 々,sPLA2 は cPLA2 に 比 べ て 基 質 特 異性が広いが,近年,工藤らによって,sPLA2 はその アイソタイプごとに異なった基質特異性や発現プロ フィールを示すことが報告されており11),最近の著者 らの検討でも V 型 sPLA2 が MΦ に強く発現している が,他方,IIa 型 sPLA2 は炎症部位に高発現してくる ことなどが明らかになって来ている9).こうした知見 を勘案すると,cPLA2 よりも広い基質特異性を有する sPLA2 もまた MΦ の抗 MAC 活性に対して何らかの 作用をもつ可能性が高いように思われる. 現在,MAC 感染 MΦ 内での cPLA2 の MAC が局在 するファゴソームへの translocation を直接的に証明 す る た め に,CR3 と GFP 標 識 cPLA2 を 発 現 さ せ た CHO 細 胞 に MAC を 感 染 さ せ た 場 合 に GFP 標 識 cPLA2 がどのような細胞内動態を示すのかについての 検討を進めているが,その成績も含めて別の機会に報 告したい. 文 献 1)冨岡治明:抗酸菌感染症が難治性である理由を 探る.日細菌誌 1995;50:687―701. 2)冨岡治明:マクロファージの殺菌作用における 遊離脂肪酸の役割.臨床免疫 2003;39:85―92. 3)Hingley-Wilson SM, Sly LM, Reiner NE, MaMas- ter WRl:The immunobiology of the mycobacterial infected macrophage. Mod Asp Immunobiol 2000;1:96―101. 4)Gomes MS, Florido M, Pais TF, Appelberg R: Improved clearance of Mycobacterium avium upon disruption of the inducible nitric oxide synthase gene. J Immunol 1999;162:6734―9. 5)Lammas DA, Stober C, Harvey CJ, Kendrick N, Panchalingam S, Kumararatne DS : ATPinduced killing of mycobacteria by human macrophages is mediated by purinergic P2Z (P2 X7) receptors. Immunity 1997;7:433―44. 6)Nau GJ, Liaw L, Chupp GL, Berman JS, Hogan BL, Young RA:Attenuated host resistance against Mycobacterium bovis BCG infection in mice lacking osteopontin. Infect Immun 1999; 67:4223―30. 7)Akaki T, Sato K, Shimizu T, Sano C, Dekio S, Tomioka H, et al.:Effector molecules in expression of the antimicrobial activity of macrophages against Mycobacterium avium complex roles of reactive nitrogen intermediates, reactive oxygen intermediates, and free fatty acids. J Leukoc Biol 1997;62:795―804. 8)Akaki T, Tomioka H, Shimizu T, Dekio S, Sato K:Comparative roles of free fatty acids with reactive nitrogen intermediates and reactive oxygen intermediates in expression of the antimicrobial activity of macrophages against Mycobacterium tuberculosis. Clin Exp Immunol 2000; 121:302―10. 9)Tomioka H, Sano C, Sato K, Ogasawara K, Akaki T, Shimizu T, et al.:Combined effects of ATP on the therapeutic efficacy of antimicrobial drug regimens against Mycobacterium avium complex infection in mice and roles of cytosolic phospholipase A2-dependent mechanisms in the ATP-mediated potentiation of antimycobacterial host resistance. J Immunol 2005;175:6741―9. 10)Qiu ZH, Gijon MA, de Carvalho MS, Spencer DM, Leslie CC:The role of calcium and phosphorylation of cytosolic phospholipase A2 in regulating arachidonic acid release in macrophages. J Biol Chem 1998;273:8203―11. 11)Kudo I, Murakami M:phospholipase A2 enzymes. Prostaglandins Other Lipid Mediat 2002;68−69:3―58. 感染症学雑誌 第81巻 第 6 号 MAC 感染 MΦ とフォスフォリパーゼ A2 動態 699 Profiles of Intracellular Expression, Distribution, and Activation of Phospholipase A2 in Host Macrophages Infected with Mycobacterium avium complex Chiaki SANO1)2), Toshiaki SHIMIZU1), Yutaka TATANO1), Ko YASUMOTO1) & Haruaki TOMIOKA1) 1) Department of Microbiology and Immunology and 2)Department of Oto-Rhino-Laryngology, Shimane University School of Medicine Our recent studies have shown that type IV cytosolic phospholipase A2 (cPLA2) plays an important role in the expression of macrophage (MΦ) antimicrobial activity against the Mycobacterium avium complex (MAC). To clarify the modes of cPLA2 participation in MΦ anti-MAC antimicrobial function in detail, we studied intracellular profiles of phospholilase A2, focusing on cPLA2, in MAC-infected MΦs, with the following findings : In murine peritoneal MΦs, cPLA2 was constitutively expressed even in uninfected MΦs, and MAC infection did not increase intramacrophage cPLA2 expression. In an RAW264.7 mouse MΦ cell line (RAW264.7 MΦs) infected with MAC, a portion of intracellular cPLA2 was concentrated in MAC organisms residing within MΦ phagosomes. MAC infection upregulated intramacrophage cPLA2 expression and induced its phosphorylation. These findings suggest that MAC infection of RAW264.7 MΦs may induce the activation of intracellular cPLA2, translocating it to phagosomes engulfing infected MAC organisms. 平成19年11月20日
© Copyright 2026 Paperzz