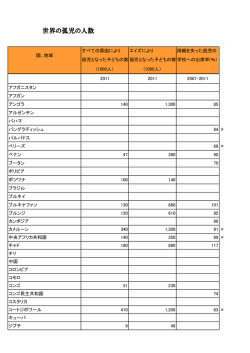
感染症の人口動態学
稲葉 寿1
1999 年 3 月 25 日
1
東京大学大学院数理科学研究科
Abstract
Population Dynamics of Infectious Diseases
Our main purpose of this paper is to provide ha brief review for recent developments in deterministic
structured models of the spread of infectious diseases.
We first review some basic ideas in mathematical epidemiology originates from the classical theory
of Kermack and McKendrick (1927). Most important key idea of mathematical epidemiology is the
basic reproduction number R0 by which the famous threshold theorem can be formulated. The basic
reproduction number is defined as the expected number of secondary cases produced, in a completely
susceptible population, by a typical infected individual during its entire period of infectiousness. The
threshold criterion states that the disease can invade into a population which is in a demographically
steady state with all individuals susceptible if R0 > 1, whereas it cannot if R0 < 1. For age-structured
SIR epidemic model, we show that the basic reproduction number is calculated as the spectral radius of
a certain positive integral operator called next generation operator. In this classic case, the threshold
condition also implies the existence and stability of endemic steady state.
Next we consider the variable susceptibility model which was also first developed by Kermack and
McKendrick in early 30th. It seems that Kermack and McKendrick’s work in 30th have been paid
relatively less attention in compare with their first paper in 1927 and their statements in those papers
have been so far not necessarily fully examined, maybe partly because their calculation was difficult
to follow and they did not clearly provide enough reason for which their complex modeling would be
truly needed to study infectious diseases in the real. However recently it is found that the variable
susceptibility model can be used to take into account the effect of genetic drift as observed in type A
influenza virus. We reformulate their model as partial differential equation system and discuss some
simple threshold phenomena and problems related to control of epidemic by vaccination.
Finally we pay our attention to HIV/AIDS models as a typical example of epidemic model with
variable infectivity. Since the incubation period of HIV is estimated to be about 10 years during which
the infectivity of infected individual is largely varying, the time elapsed from infection (disease-age)
should be taken as a structure variable. It is shown that we can directly formulate a linearized equation
describing the HIV invasion process by pair formation and calculate the basic reproduction number.
Since nonlinear dynamics of the spread of HIV infection in bisexual structured population is so complex,
existence of endemic steady state and stability of steady states are still open problems in mathematically
rigorous sense.
1
はじめに
感染症ないし伝染病の流行は人類史にきわめて大きなインパクトを与えてきた。14 世紀における世界的
なペスト流行は数年の間に中世ヨーロッパの三分の一から二分の一の人口を奪ったといわれている。また
今世紀においては第一次世界大戦末期に発生したスペイン風邪(インフルエンザ)は全世界で約6億人が
感染し、2300 万人が死亡した。日本においてもこのとき全人口の 40 パーセントが感染して 38 万人の死者
を出した。前工業化社会における死亡率の不安定性がしばしばこうした疫病の流行によるものであったこ
とは周知であるが、第二次大戦後における先進諸国を中心とした保健医療体制の充実によって感染症の制
圧を主因とする死亡率の劇的低下が実現され、一時的にはあたかも伝染病の撲滅は時間の問題であるかの
ような楽観すらもたらした。しかしながら 80 年代以降、抗生物質耐性菌等の出現、マラリア、結核等の従
来からある感染症の再興、エイズやエボラ等の新興感染症の相次ぐ出現等によって従来の楽観論は根拠を
失い、感染症の制圧がきわめて困難であることことが判明してくるにつれて、直接的医学的な対処法(治
療行為)とともに予防のための戦略研究が非常に重要であることが広く再認識されるようになってきてい
る。感染症流行の脅威は医学・公衆衛生学的な進歩にもかかわらず今日においても依然として極めて大き
いのである。
伝染病の流行は人々のコミュニケーションのあり方、生活の仕方、人口動態等にすぐれて依存する社会的
現象であり、どのように流行が発生し、予防のためにどのような介入行為が有効であるか、等を知るため
にはこれを定量的に数理モデルを用いて分析しておくことが非常に重要である。伝染病の流行過程の数理
的研究は人口論とともに長い研究の伝統があり、その端緒は 18 世紀のベルヌーイの研究に遡ると言われる
(Bernoulli 1760, Anderson 1991)。今世紀初頭のロス卿によるマラリア流行に関する閾値定理の発見、ケ
ルマックとマッケンドリックによる 1920 年代から 30 年代にかけての一連の業績は時代を超越した意義を
持ち続けている。一般人口モデルと同様に 1970 年代に至るまでその歩みは比較的遅々としたものであった
が、その後過去 20 年間にわたって、構造化人口モデルの発達や数理生態学と交流しながら、応用数理の一
分野として急速に発展してきている。とりわけ 80 年代におけるエイズの世界的流行は、それが欧米先進諸
国の中枢で発生しただけに学界にも大きなインパクトを与え、感染症流行の予測と効果的な予防や治療を
評価するための数理モデル研究は著しく促進されたのである。
感染症の数理モデルは大きく分けて、マクロな人口レベルにおける感染者人口の動態を考察するモデル
と、人体内の細胞レベルにおけるウィルス感染やウィルスと免疫系の相互作用を扱うミクロレベルのモデ
ルに分類できる。本稿ではマイクロレベルでのモデルには立ち入らず、専ら人口レベルにおける古典的な
SIR 型モデル(ケルマックーマッケンドリックモデル)から出発して、決定論的モデリングにおける最近
の問題を、特に感受性の変動と感染力の変動を考慮したモデルを取り上げて論ずることとしたい1 。
2
感染症流行の古典モデル:基本再生産数と閾値定理
ケルマックとマッケンドリックの最も有名な第一論文 (Kermack and McKendrick 1927) において提起さ
れた伝染病流行モデルは、局地的な封鎖人口における伝染病の急速かつ短期的な流行に関するモデリング
であった。病気の流行期間が短いためホスト人口の出生、死亡等の人口動態は無視できると考える。S(t),
1 伝染病数理モデルについては Anderson and May (1991), Busenberg and Cooke (1993), Capasso (1993), Mollison (1995),
Isham and Medley (1996) 等がすぐれた解説である。またここでは専ら人間の感染症を念頭においているが、動植物や昆虫等の感
染症については Grenfell and Dobson (1995) がある。
1
I(t), R(t) をそれぞれ感受性人口 (susceptibles:感染する可能性のある人口)、感染人口 (infected/infectious:
感染していてかつ感染させる能力のある人口)、隔離された人口 (recovered/removed:病気からの回復によ
る免疫保持者ないし隔離者・死亡者)とする。このときケルマックーマッケンドリックの提起した最も単
純なモデルは以下のような常微分方程式システムによって表される:
dS(t)
= −βS(t)I(t)
dt
dI(t)
= βS(t)I(t) − γI(t)
dt
dR(t)
= γI(t)
dt
(2.1)
(2.2)
(2.3)
ここでβ は感染率、γ は隔離率(場合によっては病気による死亡率)である。βI(t) は感染力 (force of infection)
であり、感受性人口の瞬間的感染率(推移強度)を表す。この人口を三つのコンパートメントにわけた伝
染病モデルを SIR モデルという。もし感染しても感染性のない状態 (letent period/exposed) や感染後症状
の発症しない状態 (潜伏期間:incubation period) などを考慮すれば I−状態はさらに分割されるし (SEIR
モデル)、免疫性の獲得が恒久的なものでなければ R → S という状態変化が可能となり、そうした場合は
SIRS モデルなどと呼ばれる。
今全体が感受性人口からなる集団(サイズ N ) を考えて、ごく少数の感染者が発生した状況を考えると
(2.2) において S(t) ≈ N とおけるから、以下の線形化方程式を得る:
dI(t)
= βN I(t) − γI(t)
dt
(2.4)
したがって流行初期においては感染者人口は I(t) ≈ I(0)e(βN −γ)t というマルサス法則に従う。すなわち病
気が集団に侵入可能となる条件(閾値条件)はこのマルサス径数が正になる条件βN − γ > 0 にほかなら
ず、これは
R0 =
βN
>1
γ
と書き直せる。換言すれば臨界的な人口密度 Ncr =
(2.5)
γ
β が存在して、これ以下の密度では流行は発生しない。
この R0 は人口学と同様に基本再生産数(比)(basic reproduction number/ratio) と呼ばれるが、全体が感
受性である人口集団において典型的な一人の感染者が再生産する二次感染者の平均数に他ならない。した
がって直感的にいえば、R0 > 1 であれば病気の流行 (outbreak) が発生して感染者人口は初期には指数関数
的に増大するが、R0 < 1 であれば流行はおこらず感染者人口は自然に減衰すると考えられる。このような
現象を閾値現象 (threshold phenomena) とよぶ。
ケルマックーマッケンドリックモデルにおいては閾値条件 R0 > 1 が満たされていれば一回のピークをも
つ流行がおこるが、やがて自然に終息し、しかも一定の感受性人口が全く感染せずに残るという挙動を示
すことが示される。この未感染におわる人口 S(∞) は最終規模方程式 (final size equation)
¶
µ
N − S(∞)
S(∞) = S(0) exp −
Ncr
(2.6)
の正根として与えられる。すなわち伝染病流行は感染者人口の消滅によって終息するのであって感受性人
口の消滅によってではない。このモデルから得られる流行曲線は20世紀初頭のインドにおけるペストの
局地的流行のデータによく一致させることができた。こうしたモデルの特質は感受性人口の減少がそのま
ま感染力の減衰をもたらす mass action type の相互作用 SI を仮定しているためである。たとえば感染力が
人口サイズに独立であるような以下のモデルを考えよう。
2
dS(t)
I(t)
= −βS(t)
dt
S(t) + I(t)
(2.7)
dI(t)
I(t)
= βS(t)
− γI(t)
dt
S(t) + I(t)
(2.8)
dR(t)
= γI(t)
dt
(2.9)
これは性的接触による感染等のように人口規模の低下が感染力の低下をもたらさない場合には妥当な仮定で
あるが、この場合には R0 > 1 であれば感受性人口と感染人口の双方の消滅によって流行が終わる (Gleißner
1988)。
ケルマックーマッケンドリックモデルとしては上記のような単純化されたものだけが言及されることが多
いが、彼らは初めから感染者の感染年齢(感染してからの経過時間)を考慮した構造化モデリングをおこなっ
ていたことに注意すべきである。今 i(t, τ ) を時刻 t において感染年齢(感染からの)経過時間:disease-age)
τ である感染人口の密度、β(τ ) を感染年齢τ における感染率、γ(τ ) を隔離率とすると (2.1)-(2.3) のモデルは
以下のような偏微分方程式モデルになる。
dS(t)
= −λ(t)S(t) = −i(t, 0)
dt
µ
¶
∂
∂
+
i(t, τ ) = −γ(τ )i(t, τ )
∂t ∂τ
Z ∞
λ(t) =
β(τ )i(t, τ )dτ
(2.10)
(2.11)
(2.12)
0
これもまた常微分方程式モデルと同様な挙動を示すことがわかっている (Iannelli 1995)。このモデルにおい
てはホスト人口の動態(出生や死亡)が無視されているが、そうしたことが妥当であるような短いタイム
スケールにおける短期的、単発的流行では感染力の変動は重要な問題ではなく、感染者を感染年齢によっ
て構造化することの意義は疑わしい。現実にどのような伝染病に対してこのモデルが適用されるべきかに
ついてはこれまで議論がなかったのである。しかしながら後に述べるようにエイズのような潜伏期間が長
く、その間の感染力の変動が大きい場合にはホスト人口の動態を考慮することで、このモデルは重要な意
義をもつようになる。
子供の流行病(はしか、水疱瘡、おたふく風邪等)のように長期的に人口のなかに定着している病気を
表現するためには、ホスト人口の動態率を考慮する必要がある。いま b をホスト人口の出生率、µを自然死
亡率とすればモデル (2.1)-(2.3) は以下のように修正される:
dS(t)
= b − µS(t) − βS(t)I(t)
dt
(2.13)
dI(t)
= βS(t)I(t) − (µ + γ)I(t)
dt
(2.14)
dR(t)
= −µR(t) + γI(t)
dt
(2.15)
ただしここで R(t) は免疫保持者であり、一度得た免疫は生涯持続すると想定する。垂直感染はなく、病気
による死亡率の増加も無視できると仮定する。このとき総人口 N (t) = S(t) + I(t) + R(t) は µb を安定な平
衡値としているから、初めから全人口は一定値 N =
b
µ であると仮定すれば
3
R0 =
βN
γ+µ
(2.16)
となることはすぐにわかるが、さらに以下のような閾値定理が示される (Hethcote 1974):
命題 2. 1 R0 ≤ 1 であれば、定常状態としては自明な定常状態(病気のない定常状態:disease-free steady
state/ DFSS)(S ∗ , I ∗ ) = ( µb , 0) だけが存在して大域的に漸近安定である。また R0 > 1 であれば、自明な
定常解は不安定化して、病気と共存する定常状態(endemic steady state/ ESS)が唯一つ出現して大域的
に漸近安定になる。
一般に麻疹や水疱瘡は長期的に大規模人口集団に定着して周期的に流行することが知られている。上記
の SIR モデルでは長期的な共存状態の周囲での減衰振動はあるものの、持続的周期解は存在しない。そこ
で周期的流行現象を反映するような改良の試みが行われている。感染率の季節変動が周期解を導くことは
よく知られている (梯 1990)。また潜伏期間の存在や感染相互作用項の強い非線形性によっても周期性が現
れる。一方、ホスト人口の構造、特に年齢によって感染率は大きく異なると考えられ、年齢構造の導入は
より現実性のあるモデルを考えていく上で重要であり、また周期解などのより豊かな解の構造をを導くと
期待されている (Tudor 1985, Greenhalgh 1988, Inaba 1990)。年齢構造を持つSIRモデルは以下のよう
な偏微分方程式によって表される:
µ
¶
∂
∂
+
S(t, a) = −µ(a)S(t, a) − λ[a | I(t, ∗)]S(t, a),
∂t ∂a
µ
¶
∂
∂
+
I(t, a) = λ[a | I(t, ∗)]S(t, a) − (µ(a) + γ)I(t, a),
∂t ∂a
µ
¶
∂
∂
+
R(t, a) = γI(t, a) − µ(a)R(t, a),
∂t ∂a
S(t, 0) = B, I(t, 0) = R(t, 0) = 0,
(2.17)
(2.18)
(2.19)
(2.20)
ここで垂直感染は無視されていて、B は感受性人口の出生率、λ[a | I(t, ∗)] は感染力であり以下のように与
えられる。
Z
λ[a | I(t, ∗)] =
ω
β(a, σ)I(t, σ)dσ.
(2.21)
0
β(a, σ) は a 歳の感受性個体とσ 歳の感染個体の間における感染率である。全人口の年齢分布 N (t, a) =
S(t, a) + I(t, a) + R(t, a) は McKendrick 方程式
µ
¶
∂
∂
+
N (t, a) = −µ(a)N (t, a), N (t, 0) = B
∂t ∂a
(2.22)
を満たしているから、N (a) = B`(a) を定常解としてもっている。
全人口は既に定常的であると仮定して、i(t, a) :=
I(t,a)
N (a)
(N (a) = B`(a)) 等と定義して全人口を1に規格
化すると SIR モデルは以下のような (i, r) システムに還元される:
µ
¶
∂
∂
+
i(t, a) = λ[a | i(t, ∗)](1 − i(t, a) − r(t, a)) − γi(t, a),
∂t ∂a
µ
¶
∂
∂
+
r(t, a) = γi(t, a),
∂t ∂a
i(t, 0) = r(t, 0) = 0,
(2.23)
(2.24)
(2.25)
4
Z
ω
λ[a | ψ] =
β(a, σ)N (σ)ψ(σ)dσ.
(2.26)
0
ここで (i, r) システムの状態空間を閉凸集合Ω := {φ = (φ1 , φ2 ) ∈ X : 0 ≤ φ1 + φ2 ≤ 1} として X =
L1 (0, ω) × L1 (0, ω) 上の作用素 A と F を以下のように定める:
¶
¶µ
µ d
0
φ1 (a)
− da
, D(A) = {φ ∈ X : φ0 ∈ X, φ(0) = 0},
(Aφ)(a) =
d
0
− da
φ2 (a)
¶
µ
λ[a | φ1 ](1 − φ1 (a) − φ2 (a)) − γφ1 (a)
F (φ)(a) =
.
γφ1 (a)
µ ¶
i
このとき (i, r) システムは u =
とすると X 上の半線形のコーシー問題として定式化される:
r
u0 (t) = Au(t) + F (u(t)), u(0) = u0 .
(2.27)
(2.28)
(2.29)
作用素 A が C0 -半群 etA を生成することは容易に確かめられるから、この方程式の弱い解 (mild solution) は
以下の積分方程式の解として得られる(定数変化法の公式)
:
Z t
u(t) = etA u(0) +
e(t−s)A F (u(s))ds
(2.30)
0
この弱い解はΩを正方向に不変にするような流れ (semiflow) を定めることがわかる。
年齢構造のある SIR システム (2.17)-(2.20) の基本再生産数 R0 は正値積分作用素
Z ωZ ω
`(ξ) −γ(ξ−σ)
(T φ)(a) = N (a)
β(a, ξ)
e
dξφ(σ)dσ
`(σ)
0
σ
(2.31)
のスペクトル半径 r(T ) で与えられることが示される。このことは以下のように考えると理解しやすい。初
期侵入の状況においては B(t, a) := N (a)λ[a|I(t, a)] が年令 a 歳でちょうど感染した新感染者の分布となる
が、これは以下のような再生方程式を満たすことは容易に示される:
Z ∞
Z σ
`(σ) −γτ
B(t, a) = N (a)
β(a, σ)
e
B(t − τ, σ − τ )dτ dσ
0
0 `(σ − τ )
(2.32)
そこでいま積分作用素
Z
(K(τ )ψ)(a) = N (a)
∞
β(a, σ)
τ
`(σ) −γτ
e
ψ(σ − τ )dτ dσ
`(σ − τ )
を考えれば、(2.31) は抽象的なボルテラ方程式
Z ∞
(K(τ )B(t − τ, ∗))(a)dτ
B(t, a) =
(2.33)
0
と見なすことができる。B(t, a) = eλt ψ(a) とおけば
Z ∞
ψ=
K(τ )e−λτ dτ · ψ
(2.34)
0
すなわち線形化方程式の固有値は複素数の集合Λ := {λ ∈ C : 1 ∈ σ(K̂(λ))} で与えられる。ここでK̂ は
K のラプラス変換、σ(A) はスペクトル集合を表す。現実的な条件のもとでK̂(τ ) は実軸上で単調減少な正
値コンパクト作用素であり、そのスペクトル半径 r(K̂(λ)) もまた実軸上で単調減少になる。従って方程式
r(K̂(λ)) = 1 は唯一の実根λ0 をもち、λ0 ∈ Λとなる。しかも r(K̂(0)) = r(T ) > 1 であればλ0 > 0、r(T ) <
1 であればλ0 < 0 となり、さらにλ0 はΛの任意の他の要素の実部よりも大きいことが示される。そこでψ0
5
をK̂(λ0 ) の固有値1に属する正値固有ベクトルであるとすれば、初期侵入においては B(t, a) ≈ eλ0 t ψ0 とな
り、R0 = r(T ) であることがわかる。
上記の考え方は一般に各個体が異質的で様々なパラメータによって構造化されている場合に適用できる
ものであり、(2.31) のような作用素は初期の感染者の分布から二次感染者の分布を生み出す作用をすると
いう意味で、次世代作用素 (next generation operator) と呼ばれる (Diekmann, et al. 1990)。したがって
標語的に言えば、一般に伝染病流行モデルの基本再生産数は次世代作用素のスペクトル半径で与えられる。
モデルが複雑になれば R0 を決定することは一般に必ずしも容易ではない。伝染病モデルの研究動機の一
つは、流行を防ぐ方策の有効性を検討することであるが、流行を防止するということは R0 を1以下に押さ
えるようにパラメータを制御するということに他ならないから、R0 の計算方法を見いだすこと、およびそ
れへのパラメータ変化の影響を調べることは非常に重要である。SIRモデル (2.17)-(2.20) に関しては以
下のような閾値定理が得られている (Inaba 1990):
命題 2. 2 T は nonsupporting2 なコンパクト作用素とする。R0 = r(T ) ≤ 1 であれば、定常状態としては
DFSS だけが存在して大域的に漸近安定である。また R0 > 1 であれば、DFSS は不安定化して、ESS が少
なくとも一つ出現する。このときさらに全ての a, σ ∈ [0, ω] に対して
Z ω
`(ξ) −γ(ξ−σ)
β(a, σ) − γ
β(a, ξ)
e
dξ > 0
`(σ)
σ
であれば非自明な定常解は唯一つだけ存在する。またλ∗ (a) を定常状態における感染力とすれば
Rζ ∗
Z ζ
−γ(ζ−η)+
λ (z)dz
η
1>γ
e
dη
(2.35)
(2.36)
σ
がすべての 0 ≤ σ ≤ ζ ≤ ω で成り立てば、その定常状態は局所的に漸近安定である。
上記の主張における定常解の一意性と安定性のための十分条件 (2.35), (2.36) の生物学的な意義はよくわ
かっていない。感染率β(a, σ) が変数分離型、すなわちβ(a, σ) = β1 (a)β2 (σ) と書ける3 ならば ESS は唯一つ
であることは容易にわかる。また (2.36) から少なくともλ∗ が十分に小さい場合は定常解は安定であること
がわかるが、これは R0 = 1 においてそれまで安定であった自明定常解から非自明定常解が分岐して、安
定性の交換がおこることに対応している。しかし R0 が大きい一般の場合には定常解の個数や安定性は未知
である。Thieme (1991), Cha, Iannelli and Milner (1997) は定常解が不安定化する可能性を示しているが、
周期解が実際に存在するかどうかについてはβ = const.という単純な場合ですらまだわかっていない。
一方、もし感染からの回復が免疫性の獲得に至らない場合は (2.23)-(2.24) は単独の方程式になる(SIS モ
デル)
:
µ
∂
∂
+
∂t ∂a
¶
i(t, a) = λ[a | i(t, ∗)](1 − i(t, a)) − γi(t, a),
i(t, 0) = 0,
Z
λ[a | ψ] =
(2.37)
(2.38)
ω
β(a, σ)N (σ)ψ(σ)dσ.
(2.39)
0
この場合はたとえ垂直感染を考慮しても、R0 > 1 であれば ESS は唯一つ存在して大域的に安定となるこ
と、従って周期解は存在しないことが証明されている (Busenberg, Iannelli and Thieme 1991, 1993)。
2 正値錐 E 上の正値有界線形作用素 T が nonsupporting であるとは、任意のペアφ ∈ E \ {0}, f ∈ E ∗ \ {0} に対して自然数
+
+
+
p = p(φ, f ) が存在して、n ≥ p となるすべての自然数 n について< f, T n φ >> 0 となることである。
3 proportionate mixing assumption
6
3
感受性の変動:インフルエンザモデルについて
ケルマックとマッケンドリックは 1927 年の第一論文以降、1939 年までに5つの論文を伝染病数理モデル
に関して著しているが、第二、第三論文では感受性の変動という非常に重要な観点を導入した一般的モデ
ルを提起している (Kermack and McKendrick 1932, 1933)。ところがこのモデルは複雑で解析が困難であ
るためか、今日に至るまでほとんど研究がなされてきていない。本節ではこのモデルを現代的に再定式化
したうえで、その意義をあきらかにしておきたい。
いま S0 (t, τ ) を時刻 t における一度も感染したことのない感受性人口の密度、v(t, τ ) を感染者の密度、
S1 (t, τ ) を病気から回復して部分的な免疫性を得た感受性人口とする。ただしτ は各状態における滞在時間
(持続時間: duration)である。B を出生率(あるいは移入率)、µを自然死亡率、γ(τ ) を回復率、β(x, y) =
β1 (x)β2 (y) を感染年齢 y の感染者から持続時間 x の部分的感受性人口 S1 への再感染率とする。ここでβ1 (x)
は単調増大関数であるとする。すなわち回復してからの経過時間が長いほど再感染しやすくなると考える。
一度も感染したことのない感受性人口は最大の感染率をもつと想定されるから、感染年齢 y の感染者から
持続時間 x の完全感受性人口 S0 への初感染率はβ1 (∞)β2 (y) で与えられると想定できる。無論β1 (∞) < ∞
と仮定する。このとき変動する感受性を考慮したケルマックーマッケンドリックモデルは以下のように書
ける:
∂S0 (t, τ ) ∂S0 (t, τ )
+
= −µS0 (t, a) − S0 (t, τ )β1 (∞)
∂t
∂τ
Z
∞
β2 (τ )v(t, τ )dτ,
(3.1)
0
S0 (t, 0) = B
(3.2)
∂S1 (t, τ ) ∂S(t, τ )
+
= −µS1 (t, τ ) − S1 (t, τ )β1 (τ )
∂t
∂τ
Z ∞
S1 (t, 0) =
γ(τ )v(t, τ )dτ
Z
∞
β2 (τ )v(t, τ )dτ,
(3.3)
0
(3.4)
0
∂v(t, τ ) ∂v(t, τ )
+
= −(µ + γ(τ ))v(t, τ ),
∂t
∂τ
·
¸Z
Z ∞
Z ∞
v(t, 0) = β1 (∞)
S0 (t, τ )dτ +
β1 (τ )S1 (t, τ )dτ
0
0
(3.5)
∞
β2 (τ )v(t, τ )dτ,
ここでは簡単のため病気による死亡率の増加は無視している。N (t) を全人口サイズとすれば
Z ∞
Z ∞
Z ∞
S0 (t, τ )dτ +
S1 (t, τ )dτ +
v(t, τ )dτ.
N (t) =
0
0
(3.6)
0
(3.7)
0
このとき
N 0 (t) = B − µN (t)
となるから、全人口サイズは一定値 N =
(3.8)
B
µ であると仮定できる。
−µτ
このモデルは明らかに (S0 , S1 , v) = (Be
, 0, 0) という自明な定常解をもつから、その基本再生産数は
Z ∞
Rτ
−µτ −
γ(σ)dσ
0
R0 = N β1 (∞)
β2 (τ )e
dτ
(3.9)
0
7
で与えられる。ケルマックーマッケンドリックはこうした複雑なモデルに関して非自明な定常解の存在条
件を論じているが、それ以外の性質はほとんど不明である。しかしモデルの解析よりも前に考えておくべ
きことは、このモデルがいったいどのような具体的な感染症に対応しているかということであるが、その
点に関してケルマックーマッケンドリックはなにも語っていないのである。
感受性の変動という場合には少なくとも二つの要因が考えられる。一つはホストにおける免疫力の自然
減衰である。これは例えばワクチンによって誘導された免疫の場合にはよくみられることである。もう一
つはウィルスの側における抗原性の変異によって既存の免疫力が無効化される場合である。例えばA型イ
ンフルエンザウィルスの抗原性はアミノ酸置換によって連続的に変化しており、これが毎年の再帰小的流
行の要因となっていると考えられる(抗原ドリフト)。これに対して、数十年に一度発生する大流行は新し
いサブタイプの出現によると考えられる(抗原シフト)。A型インフルエンザについてはホストの感受性の
変化は専らウィルス側の抗原ドリフトの効果によることが実験的に確認されている。こうしたことを背景
にして、ペアゼ (Pease 1987) はA型インフルエンザの流行モデルとして以下のようなモデルを提案した:
∂S(t, a) ∂kS(t, a)
+
= −β(a)S(t, a)I(t),
∂t
∂a
Z ∞
dI(t)
= −γI(t) + I(t)
β(a)S(t, a)da,
dt
0
(3.10)
(3.11)
kS(t, 0) = γI(t),
(3.12)
ここで S(t, a) は感染経験のあるい感受性人口密度であり、パラメータ a はホスト個体が最後に感染したイ
ンフルエンザウィルスと時刻 t 時点において流行しているウィルスの(抗原としての)差異をアミノ酸置換
量を基準にして測ったものである。k はインフルエンザウィルスにおけるアミノ酸置換速度であり、β(a) は
再感染率、γ は回復率である。ホスト人口の動態は無視されており、総人口 N は保存される:
Z ∞
N=
S(t, a)da + I(t)
(3.13)
0
ペアゼのモデルの特徴はウィルスの変異をホスト人口の感受性の相対的変化として読み直す点にある。k
は定数であると仮定しているから、このモデルは感染経験のない人口や自然死亡等のホスト人口動態率を
考慮せず、感染者の回復率が感染年齢に独立であると仮定した場合のケルマックーマッケンドリックモデ
R∞
ル (3.1)-(3.6) の特別な場合と考えられる。このとき I(t) = 0 v(t, τ )dτ である。インフルエンザのように
感染年齢のタイムスケールが流行のタイムスケールに比べて非常に短い場合は、人口学的タイムスケール
に比べて短期的な流行期間を考える限りこれらは妥当な仮定である。
いま感染者のいない状態 I(t) = 0 を考えると、(3.10) からただちに以下を得る:
½
0,
t − ka > 0
S(t, a) =
S0 (a − kt), t − ka < 0
(3.14)
ただしここで S0 (a) は初期条件 S(0, a) = S0 (a) である。すなわちペアゼモデルは全てホスト人口に感染経
験があり、各人が感染した経験のあるウィルスタイプと現存するウィルスの相対距離は抗原ドリフトによっ
て常に拡大していくから、感染者のいない定常解 (DFSS) が存在しない。すなわち新たな感染からの回復
による免疫力のブーストがない限り、ホスト人口の「感染経験」は常に古びてゆき、その免疫構造は変化
しつづける。したがって通常の意味における固定した基本再生産数も定義できないが、そのかわりに時間
に依存する基本再生産数 R0 (t) が定義される:
8
1
R0 (t) =
γ
Z
∞
kt
1
β(a)S0 (a − kt)da =
γ
Z
∞
β(a + kt)S0 (a)da
(3.15)
0
R0 (t) は仮定から単調増大であるから、初期には流行が不可能(R0 (0) < 1)であっても R0 (∞) > 1 であれ
ば有限時間のうちに侵入条件が満たされて再び流行が可能になる。さらに以下が示される (Inaba 1998):
命題 3. 1 R0 (∞) < 1 であればシステム (3.10)-(3.12) は定常状態を持たない。一方 R0 (∞) > 1 であれば
少なくとも一つの定常状態が存在する。
定常状態の安定性はβ(0) > 0 である場合には十分条件が得られているが、そうでない場合についてはペ
アゼ自身が扱ったβ(a) = βa という特殊な場合に局所安定となることがわかってるだけで、一般的には未
解決の問題である (Inaba 1998)。周期解の存在も不明であるが、しかしペアゼが指摘するように、少なく
とも定常解の摂動が減衰振動を導くのであれば、抗原シフトの間における再帰的小流行が抗原ドリフトに
よって引き起こされうるという考えは支持される。
ペアゼモデルにホスト人口の動態を導入するためにはケルマックーマッケンドリックと同様に感染経験
のない人口というカテゴリーを付加するのが妥当であろう。いま S0 (t) を t 時刻における感染経験のない感
受性人口数、b を出生率、µを死亡率、β0 を初感染率とすれば、(3.10)-(3.12) は以下のように書き直せる:
dS0 (t)
= b − µS0 (t) − β0 S0 (t)I(t)
dt
(3.16)
∂S(t, a) ∂kS(t, a)
+
= −µS(t, a) − β(a)S(t, a)I(t),
∂t
∂a
·
¸
Z ∞
dI(t)
= −(µ + γ)I(t) + β0 S0 (t) +
β(a)S(t, a)da I(t),
dt
0
(3.17)
(3.18)
kS(t, 0) = γI(t),
(3.19)
ただしここでは病気による死亡率の増加はないと仮定している。この場合は病気の存在しない人口学的定
常状態 (S0 , S, I) = ( µb , 0, 0) が得られるから、(3.18) において S0 =
b
µ,
S = 0 とおくことによって I(t) の線
形化方程式が得られ、基本再生産数が以下のように定義できる。
R0 =
bβ0
µ(µ + γ)
(3.20)
このとき以下がなりたつ:
命題 3. 2 R0 > 1 であれば少なくとも一つの非自明な定常解 (ESS) が存在する。一方、β0 ≥ β(a) かつ
R0 ≤ 1 であれば自明な定常解 (DFSS) は大域的に安定である。
証明 定常を (S0∗ , S ∗ (a), I ∗ ) とおけば以下を得る:
S0∗ =
b
µ + β0 I ∗
S ∗ (a) =
γ ∗ − µk a− Ik∗
I e
k
全人口サイズは一定値 N =
N=
b
γ
+ I∗
µ + β0 I ∗
k
(3.21)
Ra
0
β(σ)dσ
(3.22)
b
µ と仮定してよいから、
Z
∞
∗
e
I
−µ
k a− k
Ra
0
β(σ)dσ
da + I ∗
0
9
(3.23)
いま
F (x) :=
b
γ
+ x
µ + β0 x k
Z
∞
x
−µ
k a− k
e
Ra
0
β(σ)dσ
da + x
0
とおけば、方程式 F (x) = N が区間 (0, N ] に根をもつことが非自明な定常解の存在する必要十分条件であ
る。F (N ) > N , F (0) = N であり、さらに
F 0 (0) = −
bβ0
µ+γ
γ
(1 − R0 )
+ +1=
2
µ
µ
µ
となることから、R0 > 1 であれば F (x) = N は少なくとも一つの実根を区間 (0, N ) にもつ。従って前半は
証明された。後半はβ0 ≥ β(a) と仮定できることから、
µ
¶
Z ∞
Z ∞
β0 S0 (t) +
β(a)S(t, a)da ≤ β0 S0 (t) +
S(t, a)da = β0 (N − I(t))
0
0
を得る。したがって
¶
¸
·
¸
·µ
µ+γ
µ+γ
I 0 (t) ≤ β0 N −
− I(t) I(t) = β0
(R0 − 1) − I(t) I(t)
β0
β0
となるから、R0 ≤ 1 であれば t → ∞ で I(t) → 0 となる。従って非自明な定常解が大域的に安定となるこ
とがわかる。2
ホストの人口動態の導入とは別に、ワクチン接種によるホスト人口の継続的な免疫化も病気のない定常
状態を可能にすることに注意しておこう。いま最も単純なワクチン導入の仕方として、単位時間あたり一
定の率²で感受性人口がワクチン接種によって免疫化したとすると、新たに免疫化された人口は感染から回
復したと同様であるから4 、以下のようにシステム (3.10)-(3.12) を修正できる:
∂S(t, a) ∂kS(t, a)
+
= −²S(t, a) − β(a)S(t, a)I(t),
∂t
∂a
Z ∞
dI(t)
= −γI(t) + I(t)
β(a)S(t, a)da,
dt
0
Z ∞
kS(t, 0) = γI(t) + ²
S(t, a)da,
(3.24)
(3.25)
(3.26)
0
従って容易にわかるように以下のような自明な定常解が出現する:
S ∗ (a) =
²N − ² a
e k ,
k
I∗ = 0
(3.27)
これは各時点において流行しているウィルスタイプと正確に一致するようなワクチン接種によって一定率
の感受性人口の免疫力が絶えずブーストされて、感受性人口における定常的な免疫力分布が維持されてい
る場合に相当している。従ってこのような人口における基本再生産数は
Z
Z
1 ∞
²N ∞ − ² a
R0 =
β(a)S ∗ (a)da =
e k β(a)da
γ 0
kγ 0
(3.28)
であり、以下が成り立つ (Inaba 1998):
命題 3. 3 R0 < 1 であれば自明な定常解は局所安定であり、R0 > 1 であれば不安定になる。
4 実際には、ワクチンによって誘導される免疫力は実際の感染によるものよりも弱く、持続性がないと考えるべきであろうが、こ
こではそのことは無視しておく。
10
もしβ(0) = 0、すなわちあるタイプのウィルスによる感染から回復した直後は同一タイプのウィルスに
よる感染はおこらないということを仮定すれば、十分におおきな²に対して R0 < 1 となるから、原理的に
はワクチンによる流行抑止は可能になる。しかし R0 は免疫化率²の単調減少関数ではないから、あるパラ
メータ領域においては免疫化率の増加は基本再生産数の減少をもたらさないことに注意すべきであろう。
このような抗原性変異モデルにおけるワクチンによる流行抑止の可能性は、前節でみたような感受性の
変化のない小児伝染病のような場合に比べるとかなり実現は困難であろうと考えられる。現実においては
変化しつつあるウィルスタイプにを正確に同定することが困難であることと、それ故にワクチンによって
誘導された免疫力は非常に弱いか無効である可能性がある。その場合ワクチンを導入されている人口の再
感染率をβ̂(a) とすれば、これは実際に感染から回復した場合における再感染率β(a) よりも高く、min β̂ =
β̂(0) > 0 である可能性は小さくない。そうした場合には免疫化率²がどのように大きくとも
R0 ≥
N
β̂(0)
γ
となるから、人口密度が十分に大きく、N ≥
(3.29)
γ
である限りウィルスの侵入を防ぐことはできない。
β̂(0)
ペアゼモデルにおいてはウィルス変異はランダムドリフトによって発生して、その変化速度は一定であ
ると想定されているが、もし遺伝子間の選択によって進化がおこるのであれば、変化速度は密度依存的と
なり非常に複雑なモデルになりうる。このような場合に関してもまったく未解明である。
4
感染力の変動:エイズ流行モデルについて
はじめに述べたように80年代におけるエイズの世界的流行は伝染病数理モデルの研究に対して大きな
インパクトを与えた。その社会的影響の大きさから欧米を中心に定量的モデルによるエイズ流行の予測・
分析に多くの人的・物的資源が投入されたために、伝染病モデルの研究全体が活性化されたからであるが、
同時にエイズはその特異な感染経路、発病機構のために古典的モデルとは非常に異なったモデルを要求し
たことも大きなチャレンジとなった (Castillo-Chavez 1989, Jewell, et al. 1992, Kaplan, et al. 1994)。
エイズは HIV(ヒト免疫不全ウィルス)の感染によっておこる症候群であるが、その感染から発病に至
るまでに平均的には10年近い時間がかかるとされる。しかもその潜伏期間においてはほとんど自覚症状
はないものの、感染者の感染力は大きく変化していると考えられている。また感染経路は体液交換を伴う
ような行為(性交渉、輸血、出産、注射針の共有等)に限られているが、リスク集団によって流行の様相
は多様であり、一律なモデル化になじまない。
従ってエイズのモデル化における重要なポイントは、潜伏期間が長くその間に感染力が大きく変化する
ため感染年令をパラメータとする必要があること、死亡率が高いためにホスト人口の変動との長期的相互
作用が無視できないこと、感染経路に応じた感染力の形態をどのように考慮するかということ等である。
もっとも単純な場合、例えば同質的な同性愛者の集団におけるランダムな性交渉による HIV 流行を想定す
ると、ケルマックーマッケンドリックモデルをもとに以下のように書ける:
dS(t)
= b − (µ + λ(t))S(t)
dt
µ
¶
∂
∂
+
i(t, τ ) = −(µ + γ(τ ))i(t, τ )
∂t ∂τ
(4.1)
(4.2)
i(t, 0) = λ(t)S(t)
(4.3)
11
λ(t) =
C(P (t))
P (t)
Z
ω
β(τ )i(t, τ )dτ
(4.4)
0
ここでτ は感染年令、P (t) = S(t) +
Rω
0
i(t, τ )dτ はリスク集団のサイズであり、C(P ) は単位時間あたりの
接触数を表す増加関数である。C(x)/x が非増加であれば、R0 < 1 のとき自明な定常解 ( µb , 0) は大域的に
安定であり、R0 > 1 では非自明な定常解が現れるが、その不安定化とともに周期解が現れることが示唆さ
れている (Thieme and Castillo-Chavez 1993, Iannelli 1995)。このモデルはイタリアにおける薬物濫用者
の集団におけるエイズ流行予測に用いられている (Iannelli, Loro, Milner, Pugliese and Rabbiolo 1992)。
モデル (4.4) において新規感染者 i(t, 0) について注目すると以下が成り立つ:
¸
·Z t
Z ∞
C(P (t))
Γ(τ )
i(0, a − τ )dτ
i(t, 0) = S(t)
β(τ )Γ(τ )i(t − τ, 0)dτ +
β(τ )
P (t)
Γ(a − τ )
0
t
(4.5)
ただしここでΓ(τ ) は感染状態における生残率で、
Rτ
−µτ −
γ(σ)dσ
0
Γ(τ ) := e
である。さらに HIV の侵入初期を考えるために、P = S =
b
µ とおけば (4.5) は線形のボルテラ方程式にな
る。従って
µ ¶Z ∞
b
R0 = C
β(τ )Γ(τ )dτ
µ
0
(4.6)
であり、流行初期段階では感染人口はロトカの安定人口モデルで記述されることがわかる。
エイズ流行の主要な経路は現在では異性間の性的接触と薬物濫用(注射針の共有)に限定されつつある
が、特に前者はほとんどすべての成人男女がリスク集団であるために最も重大な問題である。この場合は
数理モデルとしては両性の非線形モデル (pair formation model) になり、関連するパラメータは非常に多
く解析は困難であるが、初期侵入状態においては線形化されたモデルを考えることによって R0 を計算でき
る場合がある (Knolle 1990, Diekmann, et al. 1991, Inaba 1997)。
この点を単純化したモデルで説明してみよう。いま性別による差異を無視して、感染個体の状態を「感
染者とペアを形成している」、「シングル」、「未感染者とペアを形成している」の3状態に分類して、感染
年令 a において各状態に見いだされる確率をそれぞれ `1 (a), `2 (a), `3 (a) としよう。さらにρ, σ をペアの形
成率と解消率、µを自然死亡率(あるいは隔離率)、β(a) を感染時間 a において一回の性交渉当たりの感染
率、η をペア内における単位時間当たりの性交頻度とする。ρ, σ, µ, η は一定値であると仮定する。いまペ
ア形成は性交渉によって開始される(すなわち非性的なペア形成を無視)と仮定して、さらに HIV 侵入初
期のためにエイズの発症による隔離は無視できると想定する。このとき以下が成り立つ:
d`1 (a)
= −(2µ + σ)`1 (a) + ρβ(a)`2 (a) + β(a)η`3 (a)
da
(4.7)
d`2 (a)
= (σ + µ)`1 (a) − (ρ + µ)`2 (a) + (σ + µ)`3 (a)
da
(4.8)
d`3 (a)
= (1 − β(a))ρ`2 (a) − (β(a)η + σ + 2µ)`3 (a)
da
(4.9)
新たな感染はペア形成の瞬間ないしはペア内での持続的交渉による感染によってのみ発生するから、初期
データは `1 (0) = 1, `2 (0) = `3 (0) = 0 と選ぶ。このとき解析解が以下のように求まる:
`1 (a) = e−µa − `2 (a) − `3 (a)
(4.10)
12
σ + µ −µa
e
(1 − e−(σ+ρ+µ)a ),
σ+ρ+µ
Z a Ra
ρ(σ + µ)
−
(β(ζ)η+σ+2µ)dζ
`3 (a) =
e s
(1 − β(s))e−µs (1 − e−(σ+ρ+µ)s )ds
σ+ρ+µ 0
`2 (a) =
一方、新しく感染した個体がひきおこす二次感染の総数は
Z ∞
R0 =
[β(a)ρ`2 (a) + β(a)η`3 (a)]da
(4.11)
(4.12)
(4.13)
0
であるから、以下のようにぺア形成による HIV 流行の基本再生産数が計算される。
Z ∞
R0 =
S(a)φ(a)da
(4.14)
0
ただしここで
Z
∞
β(τ )ηe
S(a) := β(a) + (1 − β(a))
−
Rτ
a
(β(ζ)η+σ+2µ)dζ
dτ,
a
φ(a) :=
ρ(σ + µ)
e−µa (1 − e−(σ+ρ+µ)a )
µ(σ + ρ + 2µ)
であり、S(a) は感染年令 a で未感染者とペアを形成した場合のペア当たりの感染確率であり、φ(a) は感染
年令 a においてペアを形成する確率である。ここで陽に計算された基本再生産数は非常に単純化された仮
定のもとで得られたものであるが、パラメータ変化の影響や感染力変動の要因を考えていく上で重要な手
がかりになる (Inaba 1997)。
モデル (4.7)-(4.9) は侵入初期の流行過程が一種のマルコフ過程によって表現されるであろうことを示唆
しているが、この過程は流行の全体像を記述する非線形の基本モデルの線形化によって得られている。従っ
て厳密に言えば、もとの非線形システムに立ち戻って定常解の線形安定性解析が正しいかどうかを示さな
ければならないが、ペア形成による HIV 流行の非線形年齢構造化モデルをそのまま扱うのは非常に困難で
あり、自明平衡解の安定性、非自明平衡解 (endemic state) の存在とその安定性については理論的にはほと
んどなにもわかっていない。シュミレーションによる研究については例えば Kakehashi (1998a, b) がある。
実用的な問題としては HIV 感染者数の推定問題が重要である。実際、HIV 感染は自覚症状に乏しく感染
者が抗体検査を受ける可能性は低いために感染規模を直接に知ることはできない。一方、AIDS を発症し
た場合は医療機関を来訪すると考えられるから患者数については信頼性が高いデータが得られるであろう。
いま i(t, 0) = b(t) とおけば感染者数 I(t) と累積患者数 C(t) は以下のように書ける:
Z ∞
b(t − τ )Γ(τ )dτ,
I(t) =
(4.15)
0
Z
t
Z
C(t) =
∞
b(t − τ )Γ(τ )γ(τ )dτ dx
−∞
(4.16)
0
初期侵入の状態を想定して b(t) = b0 eλ0 t と仮定できれば
R ∞ −λ τ
e 0 Γ(τ )γ(τ )dτ
λ0
C(t), κ = 0 R ∞ −λ τ
I(t) =
κ
e 0 Γ(τ )dτ
0
(4.17)
を得る。生存率Γ(τ ) は臨床データから推定されているから (4.17) から I(t) を求めることができる。日本の
エイズデータは HIV/AIDS の指数関数的拡大の傾向を強く示唆しており、(4.17) 式のそれへの適用によれ
ば、現行のサーベイランスによる感染者の捕捉率は高々3割から5割であると推定された (稲葉 1995)。ま
た A(t) を単位時間あたりの新規患者数とすれば
13
Z
∞
A(t) =
b(t − τ )Γ(τ )γ(τ )dτ
(4.18)
0
であるから、ボルテラ積分方程式 (4.18) の反転問題として b(t) の推定問題が定式化できる。この問題は結果
が不安定な ill-posed な問題であることが知られており、データが少なければ b(t) としてあらかじめ関数型を
想定するパラメトリックな方法を用いる他ないが、もし A(t) に関する信頼できる長期時系列データがあれば
ノンパラメトリックな数値的解法も可能であり、感染規模推定の標準的手法になっている (back-calculation
method: Brookmeyer and Gail 1994, Kakehashi 1998a)。
また HIV はヒトの体内において長期にわたって変異を繰り返しながら免疫系との闘争を生き延びて、最
終的にエイズの発病に至ると考えられるが、こうした体内でのウィルスのダイナミクスに関する数理モデル
も、発病過程を理解して薬剤投与の効果を最適化するための手段として臨床的治療への応用が考えられてき
ていることに注意しておこう (Asachenkov, et al. 1994, Nowak and May 1991, Kirschner 1996, Kirschner
and Webb 1997)。
5
おわりに
20 世紀の初頭に、ロス卿はインドにおけるマラリア流行研究においてマラリアの感染ルートを特定する
とともに、その流行過程を数理モデルを用いて解析することではじめて閾値現象を見出した。その後の数
理伝染病学 (mathematical epidemiology) の発達は著しいものがあるが、現在においてもマラリア流行の
抑止にはまったくめどがたっていない。マラリアのように伝染のための中間媒介者が存在する場合、それ
を生態系を損なうことなく撲滅することは困難であるし、その行動が介入行為などの環境変動に対して適
応的に進化してしまうという問題がある。すなわち安易な薬剤散布はウィルスや媒介者の薬剤耐性を進化
させてしまう危険がある。インフルエンザのようにウィルス自体が絶えず変化している場合や、HIV のよ
うに進化速度が速い場合も一般にはワクチンによる流行抑止は困難ならざるを得ない。こうした進化的機
構を考慮にいれたモデルの開発は今後の最も重要な課題のひとつであろう。
効果的なワクチンや治療薬が開発されるかどうかは医学的な問題であるが、仮にそうした薬剤が存在し
ても実行可能な接種計画やリスク行為の回避によって流行が抑止しうるかどうかは別の問題である。エイ
ズの場合のように個人レベルでの発症予防治療が潜伏期間を延長し、人口レベルにおける基本再生産数
を増加させてしまうという可能性も存在する。また現実には感染経路が生活習慣と深く絡み合っていたり
(Nakazawa, et al. 1998)、治療薬や医療施設へのアクセスが経済的に不可能であれば感染症流行抑止政策
の有効性は限られたものでしかないかもしれない。しかしそうした場合ですら実行可能な最良政策を検討
するためには数理モデルによる分析をぬきにしては考えることができない。
これまで伝染病疫学における数理モデルの開発はもっぱら欧米諸国においておこなわれてきており、わ
が国からの貢献はきわめて少ない。ここには既存の領域をこえた研究が育ちにくいことや、社会・人間現
象に関わる応用数理が未発達であるというわが国の学問体制のもつ問題点が伏在している。しかしながら
はじめに述べたようにコミュニケーション密度の高い現代社会は感染症流行に対してきわめて脆弱であり、
国際的にも人口爆発と稠密な居住環境、環境破壊の進展は新たな感染症の世紀を導くことは予想されると
ころである。次々に現れる感染症に対してつねに「魔法のワクチン」の出現を期待することは必ずしもで
きないのであり、予防・防疫のためのソフトウェアの開発によって対抗することが不可欠であろう。広い
意味における医学領域における数学的分析手法の充実と応用が期待される所以である。
14
参考文献
[1] A. Asachenkov, G. Marchuk, R. Mohler and S. Zuev (1994), Disease Dynamics, IIASA, Birkhäuser,
Boston.
[2] R. M. Anderson (1991), Discussion: The Kermack-McKendrick epidemic threshold theorem, Bull.
Math. Biol. 53(1/2): 3-32.
[3] R. M. Anderson and R. M. May (1991), Infectious Diseases of Humans: Dynamics and Control,
Oxford UP, Oxford.
[4] Bernoulli, D. (1760), Essai d’une nouvelle analyse de la mortalite causee par la petite verole et des
avantages de l’incubation pout la prevenir, Mem. Math. Phys. Acad. R. Sci. Paris: 1-45.
[5] R. Brookmeyer and M. H. Gail (1994), AIDS Epidemiology: A Quantitative Approach, Oxford UP,
New York Oxford.
[6] S. Busenberg, M. Iannelli and H. Thieme (1991), Global behaviour of an age-structured S-I-S epidemic model, SIAM J. Math. Anal. 22: 1065-1080.
[7] S. Busenberg, M. Iannelli and H. Thieme (1993), Dynamics of an age-structured epidemic model,
In Dynamical Systems, Nankai Series in Pure, Applied Mathematics and Theoretical Physics Vol.
4, Liao Shan-Tao, Ye Yan-Qian and Ding Tong-Ren (eds.), World Scientific, Sigapore: 1-19.
[8] S. Busenberg and K. Cooke (1993), Vertically Transmitted Diseases: Models and Dynamics,
Biomathematics 23, Springer, Berlin.
[9] V. Capasso (1993), Mathematical Structures of Epidemic System, Lec. Notes in Biomath. 97,
Springer, Berlin.
[10] C. Castillo-Chavez (ed.) (1989), Mathematical and Statistical Approaches to AIDS Epidemiology,
Lecture Notes in Biomathematics 83, Springer-Verlag, Berlin.
[11] Y. Cha, M. Iannelli and F. A. Milner (1997), Stability change of an epidemic model, preprint.
[12] Y. Cha, M. Iannelli and F. A. Milner (1998), Existence and uniqueness of endemic states for the
age-structured S-I-R epidemic model, Math. Biosci. 150: 177-190.
[13] O. Diekmann, J. A. P. Heesterbeak and J. A. J. Metz (1990), On the definition and the computation
of the basic reproduction ratio R0 in models for infectious diseases in heterogeneous populations, J.
Math. Biol. 28: 365-382.
[14] O. Diekmann, K. Dietz and J. A. P. Heesterbeek (1991), The basic reproduction ratio for sexually
transmitted diseases I. Theoretical considerations, Math. Biosci. 107: 325-339
[15] W. Gleißner (1988), The spread of epidemic, Applied Mathematics and Computation 27: 167-171.
15
[16] D. Greenhalgh (1988), Threshold and stability results for an epidemic model with an age-structured
meeting rate, IMA J. Math. Appl. Med. Biol. 5: 81-100.
[17] B. T. Grenfell and A. P. Dobson (eds.) (1995), Ecology of Infectious Diseases in Natural Populations,
Cambridge University Press, Cambridge.
[18] H. W. Hethcote (1974), Asymptotic behaviour and stability in epidemic models, In Mathematical
Problems in Biology, P. van den Driessche (ed.), Lec. Notes Biomath. 2, Springer, Berlin: 83-92.
[19] M. Iannelli, R. Loro, F. Milner, A. Pugliese and G. Rabbiolo (1992), An AIDS model with distributed
incubation and variable infectiousness: Applications to IV drug users in Latium, Italy, Eur. J.
Epidemiol. 8(4): 585-593.
[20] M. Iannelli (1995), Mathematical Theory of Age-Structured Population Dynamics, Giardini Editori
e Stampatori in Pisa.
[21] H. Inaba (1990), Threshold and stability results for an age-structured epidemic model, J. Math.
Biol. 28: 411-434.
[22] 稲葉 寿 (1994), 日本のエイズの動向と HIV 感染者数の推定について, 人口問題研究 50(4), 31-44.
[23] H. Inaba (1997), Calculating R0 for HIV infection via pair formation, In Advances in Mathematical
Population Dynamics -Molecules, Cells and Man, O. Arino, D. Axelrod and M. Kimmel (eds.),
World Scientific, Singapore: 355-382.
[24] H. Inaba (1998), Mathematical analysis for an evolutionary epidemic model, to appear in the Proceedings of the Conference on Mathematical Models in Medical and Health Sciences, Vanderbilt
University Press.
[25] V. Isham and G. Medley (eds.) (1996), Models for Infectious Human Diseases: Their Structure and
Relation to Data, Cambridge University Press, Cambridge.
[26] N. P. Jewell, K. Dietz and V. T. Farewell (eds.) (1992), AIDS Epidemiology: Methodological Issues,
Birkhäuser, Boston.
[27] 梯 正之 (1990), 数理モデルによる麻疹予防接種の効果分析, 日本公衆衛生学雑誌 7, 481-489.
[28] M. Kakehashi (1998a), Mathematical analysis of the spread of HIV/AIDS in Japan, IMA Journal
of Mathematics Applied in Medicine and Biology, in press.
[29] M. Kakehashi (1998b), Validity of simple pair formation model with realistic parameter setting,
Paper presented at the IUSSP Seminar on Measurement of Risk and Modelling the Spread of AIDS,
Copenhagen, 2-4 June.
[30] E. H. Kaplan and M. L. Brandeau (eds.) (1994), Modeling the AIDS Epidemic: Planning, Policy,
and Prediction, Raven Press, New York.
16
[31] W. O. Kermack and A. G. McKendrick (1927), Contributions to the mathematical theory of
epidemics-I, Proceedings of the Royal Society 115A: 700-721. (reprinted in Bulletin of Mathematical
Biology 53(1/2): 33-55, 1991)
[32] W. O. Kermack and A. G. McKendrick (1932), Contributions to the mathematical theory of
epidemics-II. The problem of endemicity, Proceedings of the Royal Society 138A: 55-83. (reprinted
in Bulletin of Mathematical Biology 53(1/2): 57-87, 1991)
[33] W. O. Kermack and A. G. McKendrick (1933), Contributions to the mathematical theory of
epidemics-III. Further studies of the problem of endemicity, Proceedings of the Royal Society 141A:
94-122. (reprinted in Bulletin of Mathematical Biology 53(1/2): 89-118, 1991)
[34] D. Kirschner (1996), Using mathematics to understand HIV immune dynamics, Notices of the AMS,
43(2): 191-202.
[35] D. E. Kirschner and G. F. Webb (1997), A mathematical model of combined drug therapy of HIV
infection, J. Theoretical Medicine 1(1): 25-34.
[36] H. Knolle (1990), Age preference in sexual choice and the basic reproduction number of HIV/AIDS,
Biom. J. 32(2): 243-256.
[37] D. Mollison (ed.) (1995), Epidemic Models: Their Structure and Relation to Data, Cambridge
University Press, Cambridge.
[38] M. Nakazawa, H. Ohmae, A. Ishii and J. Leafasia (1998), Malaria infection and human behavioral
factors: A stochastic model analysis for direct obserbation data in the Solomon islands, Amer. J.
Human Biology 10: 781-789.
[39] M. A. Nowak and R. M. May (1991), Mathematical biology of HIV infections: Antigenic variation
and diversity threshold, Math. Biosci. 106(1): 1-21.
[40] C. M. Pease (1987), An evolutionary epidemiological mechanism, with applications to type A influenza, Theor. Poul. Biol. 31: 422-452.
[41] D. W. Tudor (1985), An age-dependent epidemic model with applications to measles, Math. Biosci.
73: 131-147.
[42] H. R. Thieme (1991), Stability change for the endemic equilibrium in age-structured models for the
spread of S-I-R type infectious diseases, In Differential Equation Models in Biology, Epidemiology
and Ecology, Lec. Notes in Biomath. 92, Springer, Berlin: 139-158.
[43] H. R. Thieme and C. Castillo-Chavez (1993), How may infection-age-dependent infectivity affect
the dynamics of HIV/AIDS ?, SIAM J. Appl. Math. 53(5): 1447-1479
17
© Copyright 2026 Paperzz