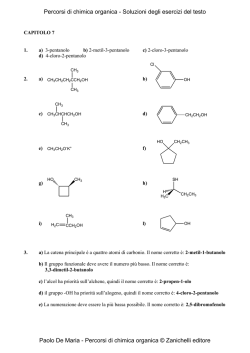
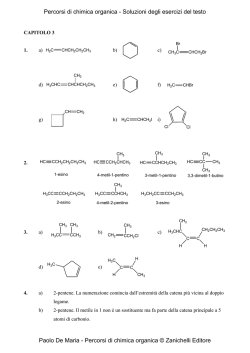
Polisaccaridi Proteine Le macromolecole biologiche Acidi nucleici Lipidi 2 CARBOIDRATI MONOSACCARIDI POLISACCARDI 3 La sintesi biologica dei carboidrati avviene attraverso il processo di fotosintesi Energia luminosa 6CO2+6H2O C6H12O6+6O2 Energia chimica 4 Struttura dei monosaccaridi O R H 1 H 2 H 3 C C O OH C OH C R2 Aldeide Chetone Aldosi Chetosi Funzione carbonilica ad una estremità della catena Funzione carbonilica non alle estremità della catena H O C R1 H H Gliceraldeide, , l’aldosio l’ aldosio più semplice 1 Catena lineare di atomi di carbonio H 2 HO 3 H 4 H 5 H 6 C O H H 1 C H OH C H C OH C OH H C OH Diidrossiacetone , il chetosio più semplice H Glucosio 2 H 3 C C O OH C OH 2 C O HO 3 C H H 4 C OH H 5 C OH H 6 C OH H OH C 1 Catena lineare di atomi di carbonio H Fruttosio 5 Nomenclatura dei monosaccarridi e numero dei loro possibili isomeri aldosi chetosi N° di atomi di C nello scheletro Nome N° di isomeri N° di isomeri ottici 3 3 aldotriosi aldotriosi 1 1 22 4 4 aldotetrosi aldotetrosi 2 2 44 5 5 aldopentosi aldopentosi 4 4 88 6 6 aldoesosi aldoesosi 8 8 16 16 3 3 chetotriosi chetotriosi 1 1 11 4 4 chetotetrosi chetotetrosi 1 1 22 5 5 chetopentosi chetopentosi 2 2 44 6 6 chetoesosi chetoesosi 4 4 88 6 Tutti i monosaccaridi tranne il diidrossiacetone contengono uno o più atomi di carbonio asimmetrici, cioè atomi di carbonio con quattro sostituenti diversi. Un composto che contiene un atomo di carbonio asimmetrico può essere presente sotto forma di due stereoisomeri, detti isomeri ottici o enantiomeri. 7 Gli stereoisomeri sono composti che pur avendo la stessa formula di struttura, ovvero la stessa sequenza di legami, differiscono per la diversa disposizione dei vari atomi o raggruppamenti di atomi nello spazio. Si definiscono enantiomeri le particolari coppie di stereoisomeri, correlati dalla caratteristica di essere l’uno l’immagine speculare dell’altro, con la peculiarità che le due immagini non sono sovrapponibili tra loro. 8 Le molecole che hanno la caratteristica di possedere immagini speculari non sovrapponibili sono dette molecole chirali Molecola Molecola con con quattro quattro sostituenti sostituenti diversi: diversi: la molecola è chirale la molecola è chirale Immagine speculare della molecola originale A Specchio La molecola ruotata non si sovrappone all’immagine speculare della molecola originale A B C D B A D C D Molecola Molecola con con tre tre sostituenti sostituenti diversi: diversi: la molecola non è chirale la molecola non è chirale Immagine speculare della molecola originale C B Molecola originale Molecola originale ruotata A Specchio La molecola ruotata si sovrappone all’immagine speculare della molecola originale A B C B B A B C B C B Molecola originale Molecola originale ruotata 9 Due enantiomeri differiscono tra loro per una specifica proprietà fisica: la capacità di ruotare il piano della luce polarizzata. Essa prende il nome di attività ottica. Quando la soluzione di uno degli enantiomeri ruota il piano della luce polarizzata verso destra, la soluzione dell’altro ruota tale piano verso sinistra. Polarizzatore 0 + º - a Campione Sorgente Analizzatore 10 La gliceraldeide contiene un solo centro chirale, costituito dall’atomo di carbonio centrale, essa possiede due diversi enantiomeri, che per convenzione vengono indicati l’uno con la lettera D, l’altro con la lettera L. 11 Gli stereoisomeri della gliceraldeide D- gliceraldeide L- gliceraldeide Specchio CHO CHO H OH CH 2 OH HO H CH 2 OH CH O H H C OH C HO HO C H CH 2O H C H2 OH CH O C HO C OH CH 2O H Modelli Modelli aa sfere palle ee bastoncini bastoncini HO C H Formule Formule in in prospettiva prospettiva Formule Formule di di proiezione proiezione di Fischer di Fischer C H 2 OH 12 Sempre per convenzione, gli stereoisomeri degli altri monosaccardi, indipendentemente dalla lunghezza della catena di atomi di carbonio e dalla loro configurazione, vengono divisi in due gruppi in base alla configurazione intorno all’atomo di carbonio chirale più distante dal gruppo carbonilico. I composti appartenenti al gruppo che ha una configurazione, a livello del centro chirale, identica a quella della D-gliceraldeide sono detti isomeri D. Quelli del gruppo con configurazione identica alla Lgliceraldeide, sono denominati isomeri L. 13 I monosaccaridi con una catena di cinque o più atomi di carbonio assumono in soluzione una forma ciclica ad anello. 6 5 H 4 C 1 6 CHO 2 H C OH 3 HO C H 5 H 4 4 C 5 HO H C OH H C OH C OH H C C 2 1 H C C 2 H H O H OH 3 H OH 1 H C OH OH α -D-glucopiranosio C O 6 3 6 CH2OH C HO CH2OH CH2OH H OH CH2OH 5 H 4 D-glucosio C HO C H OH 3C H O 1 H OH C H C 2 OH β -D-glucopiranosio La chiusura della catena lineare in un ciclo avviene per formazione di un legame covalente intramolecolare tra il carbonio del gruppo carbonilico e l’atomo di ossigeno di un gruppo ossidrilico lungo la catena. 14 Le ragioni chimiche di questa reazione sono legate al fatto che aldeidi e i chetoni reagiscono con gli alcoli per dare come prodotti, chiamati rispettivamente, emiacetali ed emichetali. OH O R1 C + HO H Aldeide R2 Alcool R1 C O R2 H Emiacetale O R2 R 2OH R1 C OR2 H Acetale Emiacetali ed emichetali possono reagire con una seconda molecola di alcol producendo, rispettivamente, acetali e chetali. 15 Nel caso del glucosio, la reazione tra il gruppo ossidrilico sul C-5 con il carbonio aldeidico C- 1 conduce alla formazione di un anello a sei atomi caratterizzato da un legame emiacetalico intramolecolare. Tale forma è definita piranosica, dal momento che ricorda la struttura ciclica a sei atomi caratteristica del pirano. H C H C O C H H C C H H Pirano 16 La chiusura del ciclo comporta la formazione di due diversi stereoisomeri, che differiscono tra loro unicamente per la configurazione intorno all’atomo di carbonio emiacetalico C-1. 6 5 H 4 C 6 CH2OH CH2OH C 3 C H C O H OH HO 5 1 H 2 H C OH C OH α -D-glucopiranosio α H 4 C H OH HO 3 C H O 1 H OH C H β C 2 OH β-D-glucopiranosio Tale atomo è chiamato carbonio anomerico, mentre i due stereoisomeri sono chiamati anomeri e vengono indicati con le lettere α e β. I due anomeri del D-glucosio sono chiamati: α-D-glucopiranosio e β-D-glucopiranosio. 17 Le forme cicliche del D-glucosio subiscono in soluzione un processo di interconversione che comporta l’apertura e la successiva richiusura dell’anello piranosico, detto mutarotazione. 6 5 C H 4 C 6 5 H 4 C HO C H OH HO CH2OH C 3 OH H OH H C C 2 1 CH2OH H H O 1 H H C OH C 2 OH α -D-glucopiranosi glucopiranosioo C O 6 3 H OH CH2OH 5 C H 4 C HO H OH 3C H O 1 H OH C H C 2 OH β -D-glucopiranosio 18 Nel caso di un chetoesoso come il D-fruttosio, le due forme anomeriche α e β si formano per reazione tra il gruppo ossidrilico sul C-5 e il gruppo chetonico sul C-2. 6 HOCH2 1 O CH2OH 5 2 H H 4 OH HO OH 3 H α-D-fruttofuranosio 1 OH 6 H O H2C C 5 H H OH OH C3 OH 2 H H O HO CH OH 2 1 4 3 OH H H β-D-fruttofuranosio D-fr uttosi o Esse sono costituite da un anello emichetalico a cinque atomi che ricorda il furano, e viene pertanto denominato forma furanosica. O 5 C 2 C 4 6 HOCH2 C H2O H O H H H H Furano 19 Per rappresentare le forme ad anello dei monosaccaridi sono spesso usate formule di prospettiva dette proiezioni di Haworth. 6 6 CH2OH CH2OH 5 H OH 4 5 O H H 1 4 H HO OH 3 H OH OH 1 H HO 2 H O H H 3 OH 2 H α -D-gluco pirano sio OH β -D-gluco piranosio L’anello piranosico, tuttavia, non è esattamente planare, ma assume le due conformazioni a barca e a sedia. H H 4 6 HO 4 CH2OH 5 H HO 5 H H HO 2 OH H OH CH2OH O H 3 6 HO 1 O H H 3 H 2 OH 1 OH 20 Gli zuccheri degli acidi nucleici 5 CH2OH 4 H OH O 1 H H H 5 CH2OH 4 H H H 1 H 3 2 3 OH OH OH Ribosio OH O 2 H 2-Deossi-ribosio 21 Alcuni derivati del D-glucosio 1 6 CH2OH CHO 2 5 H C OH H 3 HO C H O OH H 4 4 1 OH H C OH H HO 5 H C OH H 3 6 H CH2OH 2 OH β -D-glucosio D-glucosio 6 CH2OH 6 6 CH2OH 5 H O 5 OH H 4 1 OH H HO H H 2 NH2 H O H 4 OH H 3 5 COOH OH 4 1 HO H 1 OH H H 3 H 3 OH H H HO O H 2 NH 2 OH C O CH3 β -D-glucosammina Acido β -D-glucuronico N-acetil acetil--β -D-glucosammina 22 Alcuni derivati del D-galattosio 2 3 HO C H 4 HO C H 5 H C OH CH2OH CH2OH CHO H C OH 6 6 1 5 HO 4 H OH O H H D-galattosio 1 2 H OH β -D-galattosio HO 4 O H OH H 3 6 CH2OH 5 OH OH 1 H H H 3 H 2 NH2 β -D-galattosammina 23 Disaccaridi comuni 6 6 5 H 5 O 1 α 4 H 3 O 5 H H 1 α 4 OH H OH 3 2 H OH 6 CH2O H CH 2O H O HO H H H H OH 6 CH2O H CH 2O H H 5 O H H 1 α 4 OH H HO 2 OH H 1 β 4 OH H H H 3 OH O O H α -Maltosio 3 2 2 H OH OH β -Cellobiosio 6 6 CH2O H 6 CH2O H CH2OH 5 5 HO 5 O H H 1 β 4 OH H H O O OH H 4 OH H H 3 H H 3 2 OH β -Lattosio 1 β H 2 OH H 1 O H H 1 α 4 OH H HO 3 H HO CH2 β O 6 O CH2OH 5 2 H HO H 2 OH 3 OH 4 H Saccarosio 24 Amilopectina C H2 OH O O HO HO C H2 OH O O HO HO CH2 O H O O CH2 OH O O HO HO HO O C H2 OH O O C H2O H O HO HO HO O HO HO O C H2 OH O O CH2O H O HO HO HO HO C H2 O C H2 O H O O O HO HO HO CH2 OH O O HO HO Amilosio HO 25 CH2OH CH2OH O O HO OH CH2OH O O HO O O HO OH OH Cellulosa 26 Struttura tridimensionale della cellulosa 27 AMMINOACIDI E PROTEINE 28 Proteine Proteine strutturali glicoproteine, collagene, elastina, cheratina Proteine catalitiche (enzimi) Proteine di trasporto emoglobina, siero albumina Proteine di regolazione (ormoni) Proteine di protezione (anticorpi, trombina) 29 Dal punto di vista chimico, le proteine sono poliammidi composte da unità monomeriche rappresentate da circa 20 diversi α-amminoacidi. H α + H3N C CO O- R Nelle proteine, gli amminoacidi sono legati tra di loro da legami ammidici, chiamati legami peptidici. Legame peptidico R1 H3N CH C OH + H H R2 N CH O AA1 R1 CO O - H 3N CH C H R2 N CH COO - + H 2O O AA2 Dipeptide AA1- AA2 30 In tutti gli amminoacidi l’atomo di carbonio α è un centro chirale, dato che possiede quattro sostituenti diversi: essi sono dunque otticamente attivi e presenti in due enantiomeri denominati L e D. Fa eccezione la glicina, nella quale l’atomo di carbonio α possiede tre sostituenti diversi: COO - COO - + H3N α CH3 L-Alanina H + H NH3 α CH 3 D-Alanina 31 Le forme L e D hanno la stessa configurazione assoluta al carbonio asimmetrico rispettivamente della L- e DGliceraldeide. Solo la forma L- partecipa alla costruzione delle proteine. 32 Gli amminoacidi sono composti bifunzionali, dal momento che contengono sia la funzione amminica che quella carbossilica e possono pertanto esistere come ioni dipolari. H3N + CH R R R COOH + H3N CH COO H2N CH COO La concentrazione dello ione dipolare risulta massima in corrispondenza di un particolare valore di pH, detto punto isoelettrico, che è caratteristico di ciascun amminoacido. 33 Amminoacidi non polari 34 Amminoacidi polari non carichi 35 Amminoacidi polari carichi 36 Sequenze di un numero relativamente piccolo di amminoacidi sono chiamate oligopeptidi. Sequenze più lunghe, fino ad un peso molecolare di circa 5000 Da, sono dette invece polipeptidi o più semplicemente peptidi. Le unità monomeriche di un peptide sono in genere chiamate residui amminoacidici. I due residui alle estremità della catena hanno l’uno un gruppo amminico libero, l’altro un gruppo carbossilico libero; sono chiamati rispettivamente residui amminoterminale (N-terminale) e carbossi-terminale (Cterminale). 37 Livelli di struttura delle proteine La sequenza degli amminoacidi che danno origine alla catena proteica rappresenta la struttura primaria della proteina; è la struttura primaria che determina il modo e la forma secondo i quali la catena amminoacidica si organizza, sia localmente che globalmente, nello spazio, contribuendo ad assegnare ad ogni proteina la particolare struttura tridimensionale necessaria a svolgere le specifiche funzioni biologiche. La struttura secondaria è costituita dal modo in cui la proteina si organizza localmente, ad esempio, in tratti di α-elica o in segmenti estesi detti foglietti β. 38 Livelli di struttura delle proteine L’avvolgimento globale assunto dalla catena proteica in virtù delle interazioni che si stabiliscono tra i gruppi presenti in catena laterale degli amminoacidi, si chiama struttura terziaria. In alcuni casi, più catene primarie diverse, ciascuna dotata della sua particolare struttura secondaria e terziaria, si aggregano per formare complessi funzionali più grandi, nei quali si individua dunque una struttura quaternaria. 39 Struttura primaria Lys Lys Gly Gly Leu Val Ala His Struttura Secondaria Struttura terziaria Struttura quaternaria 40 Strutture secondarie nelle proteine α-elica β-sheet 41 Struttura quaternaria dell’emoglobina β β α α Gruppo eme 42 ACIDI NUCLEICI 2 Acidi nucleici Acido desossiribonucleico (DNA) Conserva le informazioni ereditarie. Acido ribonucleico (RNA) Svolge ruoli fondamentali nei complessi processi di espressione dell’informazione genetica che portano alla sintesi proteica. 3 Struttura delle unità monomeriche degli acidi nucleici: i nucleotidi Gruppo fosfato O OH P OH 5' O Ba se e teroc ic lica CH2 O 4' H H 3' OH 1' H H 2' OH/H Zucchero 4 Gli zuccheri degli acidi nucleici 5 CH2OH 4 DNA OH O H 1 H H H 3 2 OH H 2-Deossi-ribosio 5 CH2OH 4 RNA H OH O 1 H H H 3 2 OH OH Ribosio 5 Le basi eterocicliche degli acidi nucleici Guanina (G) DNA Adenina (A) Citosina (C) Timina (T) Guanina (G) RNA Adenina (A) Citosina (C) Uracile (U) 6 Struttura delle basi eterocicliche Basi pirimidiniche Basi puriniche NH2 NH2 N N N N H O H Adenina N N O N N H N Citosina O N O N N H NH2 H3C N N H H O H Uracile O H Guanina Timina 7 Gli acidi nucleici sono poliesteri dato che le diverse unità monomeriche sono collegate le une alle altre mediante ponti fosfato di natura esterea tra il carbonio C-5' e il carbonio C-3' di due anelli zuccherini consecutivi. Essi sono caratterizzati da lunghe catene lineari con un ossidrile libero in posizione 5' ad un’estremità ed un ossidrile libero in posizione 3' all’altra estremità. La sequenza delle basi è variabile e questo aspetto risulta di grande importanza per le funzioni biologiche svolte dagli acidi nucleici. 8 DNA 9 Struttura di una catena di DNA O - O P O O CH2 Base O H H H H O P O O O CH2 Base O H H H H O - O H O P Nucleoside Nucleotide - H O CH2 H Base O H H H O H 10 La macromolecola di DNA consiste di due catene di nucleotidi antiparallele che si avvolgono a spirale attorno ad un asse centrale, dando origine ad una doppia elica destrorsa che rappresenta la struttura secondaria del DNA. Lo scheletro zucchero-fosfato-zucchero-fosfato è posto all’esterno. Le basi sono dirette verso l’interno e sono approssimativamente perpendicolari all’asse della molecola stessa. 11 Struttura secondaria del DNA A T 3 ' 5 ' G C A T T C A G A T 5 ' 3 ' Rappresentazione della doppia elica destrorsa formata dall’avvolgimento a spirale attorno ad un asse centrale di due catene antiparallele di nucleotidi 12 Struttura secondaria del DNA A C C T T C A 5' 3' T G A Rappresentazione di una singola catena di nucleotidi 13 La stabilità dell’intera macromolecola dipende dalle interazioni idrofobiche e dalle forze di van der Waals che si instaurano tra le basi impilate e dai legami ad idrogeno che si formano tra le basi delle catene complementari. L’insieme di queste forze tiene uniti i due filamenti. Queste forze sono tuttavia sufficientemente deboli da permettere alle due catene di DNA di separarsi facilmente durante lo svolgimento di alcune attività fondamentali. 14 La pirimidina di una catena è sempre appaiata con una purina di un’altra catena. La complementarietà (adenina-timina e guanina-citosina) comporta che quando è specificata la sequenza nucleotidica su un filamento, anche la sequenza sull’altro è fissata automaticamente. 15 Legami ad idrogeno tra le basi complementari H N N H O N H N CH3 O N O H H H N H N H H O O H H CH2 H Adenina-Timina H O CH2 O H N O H N N H N O H N O H H N H O H H N N H O H Guanina-Citosina O H H H CH2 H O CH 2 O 16 La struttura secondaria del DNA risulta determinante ai fini della comprensione dei meccanismi attraverso i quali le informazioni genetiche vengono conservate e trasmesse. La complementarietà tra le catene è la chiave mediante cui il DNA si duplica, trasferendo le informazioni dalla cellula madre alle cellule figlie. 17 La sequenza delle basi è responsabile della conservazione delle informazioni genetiche Particolari sequenze di 3 basi nucleotidiche, triplette nel DNA, codoni nell’mRNA ed nell’rRNA, identificano e codificano i diversi amminoacidi. 18 RNA 19 I tre tipi principali di RNA presenti nelle cellule • RNA messaggero (mRNA) • RNA ribosomiale (rRNA) • RNA transfer (tRNA) 20 L’espressione del messaggio genetico che porta alla sintesi delle proteine si realizza in due fasi: • la prima, chiamata trascrizione, prevede che il messaggio contenuto nel gene sia trascritto in termini di mRNA. La sintesi di quest’ultimo è catalizzata da uno specifico enzima, la RNA polimerasi, che si serve di un filamento di DNA come stampo; • la seconda fase, detta traduzione, richiede l’intervento dell’mRNA che funge da stampo e del tRNA che trasporta i singoli amminoacidi sul ribosoma dove avviene la sintesi della catena proteica. 21 Ribosoma 3' 5' mRNA 3' 5' Codone dell’arginina tRNA Legame tra anticodone del tRNA e codone dell’mRNA Tyr Ser Leu tRNA Phe 5' 3' Scorrimento del ribosoma verso il codone successivo Leu Tyr Ser Phe Arg 5' 3' Phe Leu Tyr Ser Scorrimento del ribosoma verso il codone successivo Meccanismo della sintesi proteica che avviene a livello dei ribosomi Arg 22 Struttura dell’mRNA Legami ad idrogeno Basi Catena zucchero-fosfato 23 LIPIDI 24 Classificazione dei lipidi Lipidi di riserva (neutri) Triacilgliceroli Cere Fosfolipidi Sfingomieline Lipidi di membrana (polari) Sfingolipidi Cerebrosidi Gangliosidi Steroli 25 I lipidi di riserva o lipidi neutri Sono costituiti da esteri del glicerolo con acidi grassi, ossia con acidi carbossilici caratterizzati da lunghe catene idrocarburiche comprendenti da 4 a 36 atomi di carbonio. Quelli che a temperatura ambiente sono liquidi vengono generalmente denominati olii. Quelli che nelle medesime condizioni hanno consistenza solida vengono denominati grassi. 26 O O CH2 OH CH OH + Rx C CH2 O C O R1 CH O C O R2 CH2 O C R3 esterificazione OH CH2 OH Glicerolo Acido grasso Trigliceride I triacilgliceroli sono esteri del glicerolo con tre molecole di acidi grassi; dal momento che non contengono gruppi funzionali elettricamente carichi o gruppi fortemente polari, vengono spesso definiti trigliceridi o grassi neutri. 27 Gli acidi carbossilici che si ottengono per idrolisi dei grassi e degli olii hanno generalmente catene lineari con un numero pari di atomi di carbonio. I più comuni acidi carbossilici sono quelli con 14, 16 e 18 atomi di carbonio. Le proprietà fisiche, ed in particolare i punti di fusione degli acidi grassi e dei composti che li contengono, sono fortemente influenzate dalla lunghezza della catena idrocarburica e dal numero di doppi legami presenti nella molecola. 28 Testa polare Acidi grassi comuni - O O C H O C H H - C C H H H H H H C H H C C H H C H H C H H C H H C H H C H C H H C C H H C C H C H C H H C H C H C H H C H H H C C H C C H H H Configurazione cis Coda idrofobica H H C H H H H H C H C C C O H H C H H H H C H H H H H 29 Nei composti completamente saturi, la libera rotazione attorno ad ogni legame carbonio-carbonio conferisce alla catena idrocarburica una grande flessibilità; di conseguenza la conformazione più stabile risulta quella in cui le catene idrocarburiche sono completamente estese; ciò permette un migliore impaccamento tra catene vicine. Negli acidi grassi insaturi i doppi legami non sono quasi mai coniugati ed adottano la configurazione cis. Tale configurazione è responsabile del basso punto di fusione dei grassi in quanto la non linearità delle catene idrocarburiche, impedisce l’impaccamento compatto ed ordinato tipico dello stato solido. 30 Le possibilità d’impaccamento delle catene degli acidi grassi dipende dal grado d’insaturazione Acidi saturi Acidi insaturi 31 Lipidi di membrana Le membrane biologiche sono costituite da un doppio strato formato da lipidi strutturalmente caratterizzati di una testa polare idrofilica e da una coda non polare idrofobica. 32 L’orientamento delle regioni idrofobiche ed idrofiliche determina la disposizione e l’impaccamento dei lipidi che caratterizzano il doppio strato della membrana. Le molecole nelle quali sono contemporaneamente presenti proprietà antitetiche come l’ idrofilicità e l’idrofobicità sono dette anfipatiche. Pertanto, a differenza dei lipidi di riserva, i lipidi di membrana sono, in generale, anfipatici. 33 Classificazione dei lipidi di membrana Fosfolipidi Sfingomieline Lipidi di membrana (polari) Sfingolipidi Cerebrosidi Gangliosidi Steroli 34 GLICEROFOSFOLIPIDI 35 I glicerofosfolipidi sono caratterizzati dalla presenza di una molecola di glicerolo, le cui funzioni alcoliche sono esterificate rispettivamente con una molecola di acido fosforico e da due code apolari, rappresentate da acidi grassi a lunga catena. O CH 2 O C Acido grasso saturo O Glicerolo CH O Acido grasso insaturo C O CH 2 O P O X Gruppo alcolico polare OAcido fosforico L’acido fosforico, a sua volta, può essere esterificato con particolari alcoli (serina, etanolammina, colina o ancora glicerolo, inositolo) dando così origine ai diversi glicerofosfolipidi. 36 Struttura generale dei glicerofosfolipidi O CH2 O C Acido grasso saturo O Glicerolo CH O C Acido grasso insaturo O CH2 O P O X Gruppo alcolico polare X OAcido fosforico X HO CH2 CH2 NH3 HO Etanolammina HO Acido Acido fosfatidico fosfatidico OH Etanolammina Etanolammina Fosfatidiletanolammina Fosfatidiletanolammina Glicerolo Colina Colina Fosfatidilcolina Fosfatidilcolina Serina Serina Fosfatidilserina Fosfatidilserina Glicerolo Glicerolo Fosfatidilglicerolo Fosfatidilglicerolo Inositolo Inositolo Fosfatidilinositolo Fosfatidilinositolo CH COO Serina OH H OH CH2 CH NH3 CH2 OH OH CH2 CH2 N(CH3)3 Colina HO H H H H H HO H OH - OH H Inositolo 37 code non polari teste polari OH OH H H C O C O CH2 CH2 CH2 CH2 CH2 CH2 CH2 CH2 CH2 CH2 CH2 CH2 CH2 CH2 CH3 O O P O O CH2 C H O C O CH2 CH2 CH2 CH2 CH2 CH2 CH2 CH CH CH2 CH2 CH2 CH2 CH2 CH3 Acido fosfatidico H H C O C O CH2 CH2 CH2 CH2 CH2 CH2 CH2 CH2 CH2 CH2 CH2 CH2 CH2 CH2 CH3 NH3 + CH2 CH2 O O P O O CH2 C H O C O CH2 CH2 CH2 CH2 CH2 CH2 CH2 CH CH CH2 CH2 CH2 CH2 CH2 CH3 Fosfatidiletanolamina H H C O C O CH2 CH2 CH2 CH2 CH2 CH2 CH2 CH2 CH2 CH2 CH2 CH2 CH2 CH2 CH3 N(CH3 )3+ CH2 CH2 O O P O O CH2 C H O C O CH2 CH2 CH2 CH2 CH2 CH2 CH2 CH CH CH2 CH2 CH2 CH2 CH2 CH3 Fosfatidilcolina H H C O C O CH2 CH2 CH2 CH2 CH2 CH2 CH2 CH2 CH2 CH2 CH2 CH2 CH2 CH2 CH3 COO CH NH3+ CH2 O O P O O CH2 C H O C O CH2 CH2 CH2 CH2 CH2 CH2 CH2 CH CH CH2 CH2 CH2 CH2 CH2 CH3 Fosfatidilserina H H H C O C O CH2 CH2 CH2 CH2 CH2 CH2 CH2 CH2 CH2 CH2 CH2 CH2 CH2 CH2 CH3 H OH H O H O P O O CH2 OH H OH C H O C O CH2 CH2 CH2 CH2 CH2 CH2 CH2 CH CH CH2 CH2 CH2 CH2 CH2 CH3 Fosfatidilnositolo 38 Il composto capostipite dei glicerofosfolipidi é l’acido fosfatidico, che non ha testa alcolica. Si trova solo in piccole quantità in forma libera ma costituisce un importante intermedio nella biosintesi dei glicerofosfolipidi. Gli acidi grassi presenti nei glicerofosfolipidi hanno, in genere, da 16 a 18 atomi di carbonio. Solitamente uno degli acidi grassi è saturo, mentre l’altro è insaturo ed è sempre esterificato al gruppo ossidrilico in posizione 2 del glicerolo. L’atomo di carbonio 2 del glicerolo dei glicerofosfolipidi è un centro chirale di configurazione L. 39 A pH fisiologico, tutti i glicerofosfolipidi hanno una carica negativa sul gruppo fosforico; inoltre, sempre intorno a pH 7, il gruppo alcolico di testa può fornire una o più cariche elettriche. I glicerofosfolipidi, per riscaldamento in presenza di acidi o basi, si idrolizzano liberando i componenti di base: acidi grassi, glicerolo, acido fosforico ed alcol. Possono anche venire idrolizzati enzimaticamente da specifiche fosfolipasi, che catalizzano l’idrolisi di particolari legami nella molecola di fosfogliceride. 40 SFINGOLIPIDI 41 Gli sfingolipidi, come i glicerofosfolipidi, sono anch’essi caratterizzati da una testa polare e da due code non polari. Diidrosfingosina HO CH CH CH (CH2)12 CH3 O Acido grasso CH N C CH2 O X Testa polare A differenza dei glicerofosfolipidi, invece del glicerolo, contengono un particolare amminoalcol, caratterizzato da una lunga catena idrofobica che presenta una insaturazione, la diidrosfingosina. 42 CH CH (CH2)12 CH3 CH2 HO CH2 CH OH Glicerolo, Glicerolo, un alcol trifunzionale CH OH HO CH2 CH OH NH2 Diidrosfingosina , Diidrosfingosina, un amminoalcol che contiene una lunga catena idrocarburica insatura 43 Struttura generale degli sfingolipidi Diidrosfingosina HO CH CH CH (CH2 )12 CH3 O Acido grasso CH N C CH2 O X Testa polare X H H Ceramide Ceramide Fosfocolina Fosfocolina Sfingomielina Sfingomielina Galattosio Galattosio Galattocerebroside Galattocerebroside Lattosio Lattosio Lattosilceramide Lattosilceramide Oligosaccaride Oligosaccaride complesso complesso Ganglioside Ganglioside O HO P O CH2 CH2 N(CH3)3 O Fosfocolina 44 La formazione di un legame ammidico tra la funzione amminica della diidrosfingosina ed il gruppo carbossilico di un acido grasso da luogo alla formazione del ceramide; esso rappresenta l’unità comune fondamentale che serve a costruire gli sfingolipidi più complessi: le sfingomieline, i cerebrosidi ed i gangliosidi. HO CH CH CH (CH2 )12 CH3 O Sfingomielina Ceramide CH N C O CH2 O HP O CH2 CH2 N(CH3)3 O L’esterificazione della funzione idrossilica del ceramide con fosfocolina o fosfoetanolammina, porta alla formazione delle sfingomieline. O HO P O CH2 CH2 N(CH3)3 O Fosfocolina 45 Le sfingomieline, a differenza di cerebrosidi e gangliosidi, possiedono teste polari, contengono fosforo e rappresentano la classe più semplice ed abbondante di sfingolipidi. La guaina mielinica che circonda alcune cellule nervose è molto ricca di sfingomieline. 46 La glicosilazione del gruppo ossidrilici del ceramide porta invece alla formazione di cerebrosidi e gangliosidi che non contengono fosforo e hanno gruppi di testa neutri; poiché il gruppo di testa consiste di una o più unità glucosidiche, i cerebrosidi ed i gangliosidi sono spesso chiamati glicosfingolipidi. CH 2 OH OH O O H OH H H H H OH C H2 H H O C N C acido grasso a catena lunga Galattocerebroside H C C H C H C H2 CH2 C H2 C H2 C H2 CH2 C H2 C H2 C H2 CH2 C H2 C H2 C H3 OH I galattocerebrosidi contengono D-galattosio e sono caratteristici delle membrane cellulari del cervello, mentre i glucocerebrosidi contengono D-glucosio e sono presenti nelle membrane cellulari dei tessuti non neurali. 47 I cerebrosidi possono contenere fino a 6 unità saccaridiche, dando luogo alla formazione di cerebrosidi più complessi, localizzati sulla superficie esterna della membrana cellulare, che interagiscono con enzimi, tossine, virus e batteri e sono inoltre coinvolti in specifici processi di adesione. 48 I gangliosidi, contengono teste polari molto grandi formate da numerose unità glucosidiche, solitamente derivatizzate con acido N-acetilneuramminico, detto anche acido sialico; quest’ultima molecola che è anche presente nelle porzione saccaridica di alcune glicoproteine di membrana. CH2 O H OH O O H OH H H H OH CH2 O H CH 2 O H CH2 O H O O O OH H CH2 C O O O O OH H H H H H H H C NH H H H H H OH H C OH H NH H OH C O H C CH2 O H C H3 C H H C OH H C OH O CO O H O OH H CH3 C N H H O acido stearico sfingosina H Ganglioside GM 49 I gangliosidi, di gran lunga gli sfingolipidi più complessi, sono importanti componenti dei siti recettoriali specifici e sono coinvolti nei processi di riconoscimento molecolare. I gangliosidi sono particolarmente abbondanti nella materia grigia del cervello, ma sono pure presenti, anche se in minore quantità, sulla superficie delle membrane cellulari della maggior parte dei tessuti. Essi si trovano, ad esempio, in siti specifici sulle terminazioni nervose cui si legano i neurotrasmettitori durante la trasmissione di impulsi da una cellula nervosa alla successiva. 50 STEROIDI 51 Gli steroidi sono lipidi strutturali presenti nella membrana di molte cellule eucariotiche. 1 Chimicamente gli steroidi sono molecole complesse caratterizzate da quattro anelli: tre esaatomici ed uno pentaatomico, condensati tra loro. 2 3 4 52 Il nucleo steroideo, derivato dal ciclopentanoperidrofenantrene , è quasi planare e relativamente rigido in quanto gli anelli condensati non consentono alcuna rotazione attorno ai legami C-C. 53 Gli steroidi più abbondanti sono gli alcoli steroidei, detti steroli. Il colesterolo è il principale sterolo dei tessuti animali. Il colesterolo avendo una testa polare, rappresentata dal gruppo ossidrilico in posizione 3, ed una coda idrocarburica non polare costituita dal nucleo steroideo e dalla catena laterale alchilica legata al C-17, è anfipatico. HO CH3 CH3 CH3 CH CH2 CH2 CH2 CH CH3 CH3 Colesterolo 54 Il colesterolo ed i suoi esteri con acidi grassi a lunga catena sono componenti importanti delle lipoproteine plasmatiche e della membrana cellulare esterna. 55 I glicerofosfolipidi, gli sfingolipidi e gli steroli sono composti anfipatici, praticamente insolubili in acqua; a contatto con questo solvente formano microscopici aggregati lipidici in una fase separata dall’ambiente acquoso circostante. Le molecole lipidiche si raggruppano in modo che le parti idrofobiche siano in contatto tra loro, mentre i gruppi idrofilici interagiscono con la fase acquosa. Il raggruppamento dei lipidi ha lo scopo di ridurre la quantità di superficie idrofobica esposta all’acqua e di minimizzare il numero di molecole di acqua che si trovano nello strato organizzato all’interfaccia acqua-lipide, producendo un aumento di entropia. 56 AGGREGATI DI LIPIDI ANFIPATICI 57 Quando lipidi anfipatici vengono mescolati con acqua, si possono ottenere tre tipi di aggregati, a seconda delle condizioni e della natura chimica dei lipidi utilizzati: Micelle Doppi strati Liposomi 58 Le micelle sono strutture sferiche relativamente piccole nelle quali le molecole di lipidi disposte in modo che le loro parti idrofobiche siano all’interno della struttura, nascoste all’acqua, e le parti idrofiliche siano esposte sulla superficie, a contatto con l’acqua. La formazione di micelle è favorita quando l’area della sezione trasversale della testa polare è superiore a quella della catena idrofobica, come accade nei sali degli acidi grassi, negli isofosfolipidi (che mancano di un acido grasso) e nel detergente sodio dodecilsolfato (SDS). 59 Nel doppio strato i due strati lipidici si combinano in modo da formare un foglietto bidimensionale; le porzioni idrofobiche dei due monostrati interagiscono tra loro escludendo l’acqua; le teste idrofiliche si dispongono sulle due superfici esterne del doppio strato a contatto con l’acqua. La formazione del doppio strato ha luogo quando le aree delle sezioni trasversali delle teste polari e delle catene idrofobiche sono simili, come per i glicerofosfolipidi e gli sfingolipidi. 60 Il terzo tipo di aggregato lipidico si forma quando un doppio strato lipidico si richiude su se stesso generando una sfera chiamata liposoma o vescicola. Mediante la formazione delle vescicole, i doppi strati perdono l’idrofobicità dei bordi che nella struttura aperta restano scoperti e acquistano la massima stabilità nell’ambiente acquoso. Queste vescicole a doppio strato includono acqua formando un compartimento acquoso separato dall’ambiente circostante. 61 Come si vedrà più avanti, i doppi strati lipidici svolgono un ruolo fondamentale per la struttura di tutte le membrane biologiche. I doppi strati fosfolipidici e i liposomi sono molto studiati sia perché le loro proprietà sono molto simili a quelle delle membrane naturali sia per le crescenti applicazioni in campo farmaceutico. 62
© Copyright 2026 Paperzz