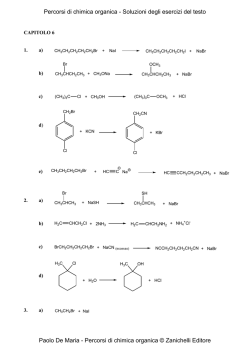
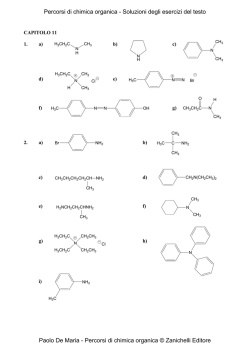
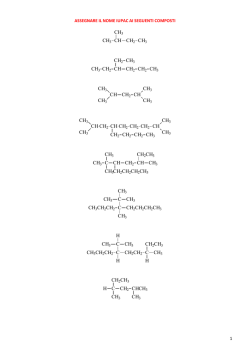
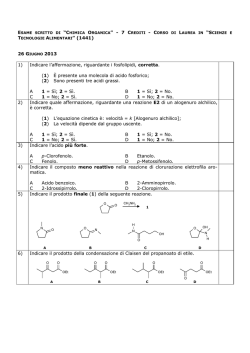
Capitolo 1: Catabolismo degli acidi grassi: digestione, mobilizzazione e trasporto; ossidazione; i corpi chetonici 2 Valore calorico degli alimenti • carboidrati = 4,1 kcal/g • lipidi = 9,3 kcal/g • proteine = 4,1 kcal/g • alcol = 7 kcal/g La chilocaloria (kcal) è l’energia richiesta per aumentare di 1°C (da 14,5 a 15,5°C) la temperatura di 1 kg di acqua ed equivale alla Caloria nutrizionale (Cal) e a 4,18 kJ. Destino dei lipidi della dieta nei vertebrati Nelson • Cox, I PRINCIPI DI BIOCHIMICA DI LEHNINGER, Zanichelli editore S.p.A. Copyright © 2014 17 | 4 5 Adipociti del tessuto adiposo bianco e bruno Nelson • Cox, I PRINCIPI DI BIOCHIMICA DI LEHNINGER, Zanichelli editore S.p.A. Copyright © 2014 23 | 6 Distribuzione del tessuto adiposo bruno in un neonato e in un adulto Nelson • Cox, I PRINCIPI DI BIOCHIMICA DI LEHNINGER, Zanichelli editore S.p.A. Copyright © 2014 23 | 7 Stato di digiuno: il fegato glucogenico Nelson • Cox, I PRINCIPI DI BIOCHIMICA DI LEHNINGER, Zanichelli editore S.p.A. Copyright © 2014 23 | 8 Stato di buona nutrizione: il fegato lipogenico Nelson • Cox, I PRINCIPI DI BIOCHIMICA DI LEHNINGER, Zanichelli editore S.p.A. Copyright © 2014 23 | 9 Punti di controllo nel mantenimento di una massa corporea costante Nelson • Cox, I PRINCIPI DI BIOCHIMICA DI LEHNINGER, Zanichelli editore S.p.A. Copyright © 2014 23 | 10 Un difetto nella produzione di leptina porta all’obesità Nelson • Cox, I PRINCIPI DI BIOCHIMICA DI LEHNINGER, Zanichelli editore S.p.A. Copyright © 2014 23 | 11 12 Mobilizzazione dei triacilgliceroli depositati nel tessuto adiposo Nelson • Cox, I PRINCIPI DI BIOCHIMICA DI LEHNINGER, Zanichelli editore S.p.A. Copyright © 2014 17 | 13 Altri induttori di lipolisi: • • • • tumor necrosis factor (TNF)-alfa glicocorticoidi ormone della crescita (GH) ormoni tiroidei (T3/T4) più grande è l’adipocita, più intensa è la lipolisi obesità: • Ipertrofica (aumento della grandezza delle singole cellule con conseguente aumento del volume) • Iperplastica (aumento del numero delle cellule con conseguente aumento del volume) 14 Metabolismo degli acidi grassi nel fegato Nelson • Cox, I PRINCIPI DI BIOCHIMICA DI LEHNINGER, Zanichelli editore S.p.A. Copyright © 2014 23 | 15 La via di ingresso nella glicolisi del glicerolo prodotto dalla demolizione dei triacilgliceroli (nel fegato) Nelson • Cox, I PRINCIPI DI BIOCHIMICA DI LEHNINGER, Zanichelli editore S.p.A. Copyright © 2014 17 | 16 Conversione di un acido grasso in un acil-CoA pirofosfatasi inorganica 2 Pi H2O O C O CoA-SH ATP PPi + AMP O C S-CoA acilCoA sintetasi (tiocinasi) acido grasso acilcoenzima A 17 Meccanismo d’azione Nelson • Cox, I PRINCIPI DI BIOCHIMICA DI LEHNINGER, Zanichelli editore S.p.A. Copyright © 2014 17 | 18 Ingresso degli acidi grassi nei mitocondri attraverso il trasportatore acil-carnitina Nelson • Cox, I PRINCIPI DI BIOCHIMICA DI LEHNINGER, Zanichelli editore S.p.A. Copyright © 2014 17 | 19 +N(CH ) 3 3 acil-CoA +N(CH ) 3 3 CoA CH2 H C CH2 OH CH2 H C O O C R CH2 - COO carnitina - COO acil-carnitina 20 -ossidazione mitocondriale (1): H3C (CH2 ) CH2 12 CH2 C SCoA O palmitoil-CoA FAD acil-CoA deidrogenasi FADH2 H H3C C (CH2 ) 12 C C H O SCoA trans-2 -enoil-CoA 21 22 -ossidazione mitocondriale (2): H H3C (CH2 ) C 12 C C H O SCoA trans-2 -enoil-CoA H2O enoil-CoA idratasi OH H H3C C (CH2 ) 12 H C C H O L- -idrossiacil-CoA SCoA 23 -ossidazione mitocondriale (3): OH H H3C C (CH2 ) 12 H C C H O SCoA L- -idrossiacil-CoA + NAD -idrossiacil-CoA deidrogenasi NADH + H+ H3C C (CH2 ) 12 O CH2 C SCoA O -chetoacil-CoA 24 -ossidazione mitocondriale (4): H3C (CH2 ) C 12 O CH2 C SCoA O -chetoacil-CoA CoA-SH acil-CoA acetiltransferasi (tiolasi) H3C (CH2 ) C SCoA 12 O miristoil-CoA CH3 C SCoA O acetil-CoA 25 Sequenza di reazioni conservata per l’introduzione di un gruppo carbonilico sul C beta Nelson • Cox, I PRINCIPI DI BIOCHIMICA DI LEHNINGER, Zanichelli editore S.p.A. Copyright © 2014 17 | 26 27 28 29 -ossidazione di un acido grasso monoinsaturo: 30 -ossidazione di un acido grasso poliinsaturo: 31 32 33 -ossidazione di un acido grasso con numero dispari di atomi di carbonio: O CH3 CH2 CH2 CH 2 CH 2 CH 2 CH 2 CH 2 CH 2 CH 2 CH 2 CH 2 CH 2 CH 2 CH 2 CH 2 C S-CoA 7 CoA-SH O 7 CH3 C S-CoA acetil-CoA H3 C CH2 C SCoA O propionil-CoA 34 latte piante organismi marini propionato (pane, cereali, etc.) NEFA C dispari valina isoleucina treonina metionina propionilCoA colesterolo succinilCoA gluconeogenesi glucoso ossalacetato ciclo del citrato malato enzima malico piruvato Pir DH acetilCoA 35 cellulosa glucoso fermentazione propionato assorbimento propionilCoA glucoso NEFA C dispari lattoso TAG latte 36 Metabolismo del propionilCoA (1): H3C CH2 C SCoA O propionil-CoA ATP HCO3 biotina propionil-CoA carbossilasi ADP + Pi H H3C C C C O SCoA O O D-metilmalonil-CoA 37 Meccanismo d’azione: ruolo della biotina nella reazione catalizzata dalla piruvato carbossiliasi, simile a quella della propionil-CoA carbossilasi. Nelson • Cox, I PRINCIPI DI BIOCHIMICA DI LEHNINGER, Zanichelli editore S.p.A. Copyright © 2014 16 | 38 Metabolismo del propionilCoA (2): H H3C C C C O SCoA O O D-metilmalonil-CoA metilmalonil-CoA epimerasi H H3C C C O O C O SCoA L-metilmalonil-CoA 39 Metabolismo del propionilCoA (3): H H H C C H C O O O C SCoA L-metilmalonil-CoA metilmalonil-CoA mutasi H O CoA-S C coenzima B12 (deossiadenosil-cobalamina) H C C H H C succinil-CoA O O 40 deossiadenosil-cobalamina: 41 Regolazione coordinata della sintesi e della demolizione degli acidi grassi 1: CO2 CH3 C O S-CoA acetilcoenzima A ADP + Pi ATP O O C biotina acetil-CoA carbossilasi (ACC) CH2 C O S-CoA malonilcoenzima A 42 Regolazione coordinata della sintesi e della demolizione degli acidi grassi 2: 43 Regolazione coordinata della sintesi e della demolizione degli acidi grassi 3: AMP AMPK 44 OH H H3C (CH2 ) C 12 H C C H O SCoA H3C CH2 C SCoA O -chetoacil-CoA L- -idrossiacil-CoA + NAD -idrossiacil-CoA deidrogenasi (CH2 ) C 12 O CoA-SH acil-CoA acetiltransferasi (tiolasi) NADH + H+ H3C C (CH2 ) 12 O CH2 C O SCoA H3C (CH2 ) C SCoA 12 O miristoil-CoA CH3 C SCoA O acetil-CoA -chetoacil-CoA 45 46 -ossidazione perossisomale (1): H3C (CH2 ) CH2 12 CH2 C SCoA catalasi O palmitoil-CoA H 2O2 FAD H 2O acil-CoA ossidasi FADH2 O2 H H3C (CH2 ) C 12 C C H O SCoA trans-2 -enoil-CoA 47 -ossidazione perossisomale (2): H2O H H3C (CH2 ) C 12 OH H C C H O trans-2 -enoil-CoA H3C SCoA enoil-CoA idratasi (CH2 ) C 12 H C C H O L- -idrossiacil-CoA + NAD -idrossiacil-CoA deidrogenasi H3C C SCoA (CH2 ) 12 O miristoil-CoA CH3 C SCoA O acetil-CoA CoA-SH NADH + H+ H3C acil-CoA acetiltransferasi (tiolasi) SCoA (CH2 ) C 12 O CH2 C SCoA O -chetoacil-CoA 48 Confronto tra la -ossidazione dei mitocondri, dei perossisomi e dei gliossisomi: tiocinasi shuttle carnitina tiocinasi shuttle carnitina 49 Alcuni ruoli dei perossisomi: • ossidazione acidi grassi con C > 20 • ω-ossidazione degli acidi grassi • ossidazione degli acidi grassi ramificati (α-ossidazione) • conversione colesterolo in acidi biliari • sintesi dei plasmalogeni sindrome di Zellweger: impossibilità di produrre i perossisomi. Adrenoleucodistrofia legato al cro. X: manquanza di ossi. degli a.g a 50 catena molto lunga, assenza del trasportatore. ω-ossidazione: citocromi P450 reticolo endoplasmatico (fegato e rene) 51 α-ossidazione: perossisomi citocromi P450 malattia di Refsum: accumulo di acido fitanico (derivato dal fitolo della clorofilla) per deficit di un enzima perossisomale: gravi problemi neurologici, cecità e sordità 52 I corpi chetonici: 53 Chetogenesi (1): 54 Chetogenesi (2): 55 Chetolisi: 56 Nelson • Cox, I PRINCIPI DI BIOCHIMICA DI LEHNINGER, Zanichelli editore S.p.A. Copyright © 2014 17 | 57 58 A digiuno: catecolammine glucagone lipolisi adipocita NEFA plasmatici epatocita acetil-CoA glucoso corpi chetonici ossalacetato citrato ciclo del citrato 59 Dopo un pasto: adipocita lipolisi insulina NEFA plasmatici acetil-CoA glucoso epatocita corpi chetonici ossalacetato citrato piruvato ciclo del citrato insulina glucoso 60 Diabete e sovrapproduzione di corpi chetonici • L’aumento di acetoacetato e di D-β-idrossibutirrato abbassa il pH del sangue (acidosi). • I corpi chetonici possono raggiungere, nel sangue e nelle urine dei soggetti diabetici non trattati, concentrazioni molto elevate: – Nel sangue 90 mg/mL rispetto al livello normale inferiore a 3 mg/mL – Un escrezione urinaria di 5000 mg/24 ore rispetto alla quantità normale inferiore o uguale a 125 mg/24 ore.
© Copyright 2026 Paperzz