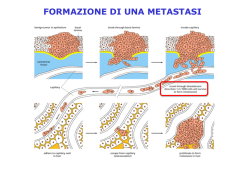
Antibody-targeted cell fusion Takafumi Nakamura, Kah-Whye Peng, Sompong Vongpunsawad,Mary Harvey, Hiroyuki Mizuguchi, Takao Hayakawa, Roberto Cattaneo & Stephen J Russell Anna Raganato Paola Saeli Veronica Rainone Leonardo Mirandola 1 La fusione cellulare • • • • • fertilizzazione sviluppo di placenta, muscolo, osso infezioni virali produzione di anticorpi monoclonali terapia antitumorale genica o citoriduttiva 2 Measles virus • Paramixovirus • tropismo: epitelio respiratorio, leucociti, sistema reticoloendoteliale. • caratteristica dell’infezione è la presenza di cellule giganti multinucleate, o sincizi, che si formano per l’azione fusogena delle proteine virali di membrana espresse dalla cellula ospite. 3 Proteine HN ed F La proteina HN è bifunzionale: la neuroaminidasi rimuove acido sialico dal recettore cellulare per il virus (CD46 o SLAM); poi l’emagglutinina si lega al recettore. Questo evento attiva la proteina F (fusion) che è responsabile direttamente della fusione tra pericapside virale e membrana cellulare. 4 SLAM • Glicoproteina di membrana espressa da linfociti B timociti immaturi linfociti T • Ha funzioni costimolatorie per i linfociti 5 SLAM è il recettore cellulare per MV Lo dimostrano esperimenti di transfezione con plasmidi recanti il cDNA per SLAM Cellule umane resistenti all’infezione di MV diventano suscettibili se transfettate con il gene che codifica SLAM L’effetto immunosoppressore di MV si esplica in gran parte per il solo legame delle proteine virali a SLAM, anche senza effetti citopatici diretti, per il blocco dell’attività costimolatoria 6 Emagglutinina ingegnerizzata • Flag-tag per purificazione ed immunoblotting (DYDDDK) • TM: dominio transmembrana • Fxa: sito di taglio riconosciuto dal fattore Xa • scFv: single-chain Ab (c-ter) anti- CD38 (marker di mieloma) 7 • Mutazioni puntiformi nel sito di riconoscimento per CD46 ed in quello per SLAM aboliscono la capacità della proteina di legarvisi, mentre scFv conferisce nuova specificità recettoriale. • La fusione può così essere ristretta solo ad un particolare tipo cellulare, che esprime la proteina antigenica riconosciuta da scFv. 8 Tre diverse linee cellulari esprimenti diversi recettori per H o per Hmutata sono state cotransfettate con un plasmide recante il gene F ed un plasmide con H o H-modificata. 9 L’attività della proteina H dipende dall’efficienza del suo trasporto alla superficie cellulare. Perciò è stata rilevata la quantità totale di H e quella presente in membrana mediante FACS ed immunoblotting 10 Sono possibili altri bersagli? Sono state prodotte emagglutinine chimeriche doppio-mutanti anti-CEA o anti-EGFR. Tranfettando le cellule con F e con H o H anti-CEA o H anti-EGFR o H antiCD38 si osservano sincizi solo se la linea cellulare esprime l’antigene riconosciuto da scFv 11 12 Applicazioni Per determinare l’efficacia della fusione cellulare nella terapia tumorale citoriduttiva sono stati prodotti vettori adenovirali esprimenti la proteina F ed una proteina tra: H, H anti-CD38, H anti-EGFR. Cellule SKOV3ip.1(cellule di tumore ovarico umano) sono state transfettate con questi vettori. 13 La linea SKOV3ip.1 esprime bassi livelli di CAR (recettore primario per coxsackievirus-adenovirus): ha una certa resistenza ai vettori adenovirali, perciò si utilizza una MOI (multiplicity of infection) elevata (300p/cellula). 14 La fusione e l’uccisione più efficiente delle cellule tumorali si ha con il vettore esprimente H anti-EGFR. Il processo mediato da H non mutata è meno efficiente, probabilmente perchè la cellule tumorali hanno una densità di EGFR superiore a CD46. 15 Effetti della fusione cellulare su tumori in vivo Cellule SCOV3ip.1 trattate con vettori: • AdH/F • AdH-CD38/F • AdH-EGFR/F sono state impiantate in topi privi di timo in modo da evitare il rigetto. 16 Il vettore AdH-EGFR/F è più efficiente nel limitare la crescita tumorale data la sua capacità di formare sincizi. 17 La fusione cellulare può servire anche per formare ibridi selettivi tra cellule tumorali e APCs, indirizzando e stimolando la risposta immunitaria. 18 Le cellule K562 transfettate con H-EGFR o con HCD38 non si fondono tra loro, ma con cellule epiteliali tumorali (A431 ) o con cellule T Jurkat CD38+ . Quindi è possibile la fusione eterologa purchè la cellula transfettata non possieda il recettore di H. 19 Un anticorpo monoclonale anti-EGFR blocca la fusione tra K562 esprimente H anti-EGFR/F e A431 esprimente EGFR. 20 MV oncolitici • Il virus del morbillo puo’ essere utilizzato allo scopo di uccidere cellule tumorali, come quelle del mieloma multiplo, costituendo quindi un nuovo agente terapeutico 21 Allo scopo sono necessari almeno 3 passaggi: • identificare un recettore specifico del tumore contro cui indirizzare selettivamente il virus • ottenere un’ emagglutinina mutata fusa con un scFv anti-recettore (le mutazioni eliminano il riconoscimento dei recettori naturali) • produrre virioni infettanti dotati della proteina H cosi’ modificata 22 • Le cellule di mieloma multiplo esprimono quantita’ elevate di CD38 • E’ stato ottenuto il virus ricombinante MV-Edm anti-CD38 • Cellule CHO (CD38-) sono resistenti all’infezione di MVa-CD38 • Cellule CHO tranfettate con plasmidi recanti il cDNA per CD38 sono uccise da MVa-CD38 ma non da MV 23 Esperimenti in vivo su roditori dimostrano che il virus ricombinante causa la formazione di cellule sinciziali multinucleate con danno esteso alle cellule tumorali ma molto ridotto su cellule normali 24 Conclusioni • E’ possibile fondere tra loro selettivamente alcuni tipi cellulari (contro cui sia stato prodotto un Ab specifico) mediante proteine fusogene virali ingegnerizzate • Questa tecnica puo’ essere impiegata nella terapia antitumorale (fusione omologa/ eterologa) • Mediante simili tecniche di ingegneria proteica si possono reindirizzare viurus umani contro particolari tipi 25 cellulari, come i tumori Un’altra categoria di proteine fusogene è la classe II. La proteina “E” è una proteina di fusione di classe II, strutturalmente diversa da quelle di classe I Classe I myxo- paramyxovirus retrovirus (HIV) filovirus (Ebola) Classe II yellow fever West Nile virus dengue virus alphavirus 26 La proteina “E” è costituita da tre domini,I,II e III,ciascuno dotato di una specifica funzione,e si presenta sul pericapside sotto forma di dimero. La struttura secondaria prevalente è il β-foglietto 27 • I virus dotati di proteine di classe II entrano nelle cellule per endocitosi (endosomi) • L’acidificazione del pH endosomale determina dei significativi cambiamenti strutturali nella proteina caratterizzati dal ripiegamento e dal ri-orientamento dei tre domini strutturali l’uno rispetto all’altro • In particolare tra i domini I e II è stata riscontrata una regione“cerniera” pHsensibile(Kl loop, residui 270-279) • Mutazioni a questo livello alterano il valore soglia del pH di fusione 28 29 • Dominio III della proteina virale: tramite interazioni glicano-mediate lega il recettore cellulare • Dominio II: presenta un loop con una sequenza idrofobica conservata responsabile del legame alla membrana bersaglio • A pH acido il dimero si dissocia per formare trimeri 30 Il trimero mantiene le trutture secondarie del dimero, ma con differenti orientazioni reciproche • Il dominio II ruota di 30° rispetto al D. I • Il dominio III ruota di 70° verso il D. II, portandosi quindi vicino al “fusion loop” del dominio II La formazione del trimero procede dal dominio II verso il dominio III, mediante interazioni sia polari che apolari 31 • Il riarrangiamento molecolare porta all’esposizione del dominio II alla cui estremità si trova il “loop” di fusione • Il loop di fusione è responsabile dell’ancoraggio alla membrana bersaglio; essenziali per questa sua attività sono i residui idrofobici Trp101, Leu-107,Phe-108 32 Figure 4 The dengue sE trimer. a, Ribbon diagram coloured as in Fig. 1b. Hydrophobic residues in the fusion loop (orange) are exposed. The expected position of the hydrocarbon layer of the fused membrane is shown in green. Representative lipids are shown to scale. A chloride ion (black sphere) binds near the fusion loop. b, Surface representation of the trimer. The dashed grey arrow indicates the most likely location for the stem (see text). An extended cavity is visible near the tip of the trimer; access to this cavity will probably be occluded by the stem. The glycan on Asn 67 and representative 33 lipids are shown in space-filling representation. Meccanismo della fusione mediata da proteine di classe II • E si associa ad un recettore della superficie cellulare mediante il dominio III • Questo legame permette l’endocitosi del virus 34 • Il ridotto pH dell’endosoma causa la dissociazione dei dimeri sul virione • Esposizione dei ”fusion loops” 35 Trimerizzazione • Si ipotizza che la membrana dell’endosoma catalizzi la trimerizzazione promuovendo la formazione di un intermedio 36 • La trimerizzazione causa il ripiegamento del dominio III • L’energia libera sprigionata dal ripiegamento spinge le membrane l’una verso l’altra 37 • Nel trimero si creano contatti ulteriori tra la regione “stem” (vicino alla porzione transmembranaria) ed il dominio II • Questo permette l’emifusione 38 • Le membrane si fondono formando un poro 39 • Tatsuo,H.,Ono,N., Tanaka,K. & Yanagi, Y. “SLAM (CDw150) is a cellular receptor for measles virus” Nature 406, 893-897 (2000) • Yargo Modis, Steven Okata, David Clements, and Stephen C. Harrison “Structure of the dengue virus envelope protein after membrane fusion” Nature 427, 313-319 (2004) • Peng,K.W. et al. “Oncolytic measles viruses diplaying a single-domain antibody against CD38, a myeloma marker” Blood, Blood 2557-2562 (2003) 40
© Copyright 2026 Paperzz