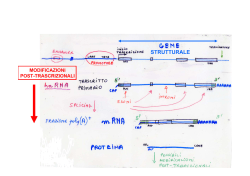
Citologia 19 – RNA (parte 2) Transfer RNA (tRNA) È la molecola che permette di associare ad ogni codon dell’mRNA il corrispondente amminoacido. I tRNA hanno una struttura chimica bidimensionale che ricorda un trifoglio, con 4 tratti a doppia elica, 3 tratti lineari formanti un loop (+ eventuale loop accessorio tra sito di attacco dell’enzima e sito di attacco all’mRNA) ed un tratto terminale contenente l’estremità 3’-OH. La sua forma tridimensionale ricorda una L rovesciata, dove sono ben visibili ed accessibili 4 zone strategiche della molecola: 1. Sito di attacco dell’amminoacido (non specifico) 2. Sito di riconoscimento dell’mRNA (specifico ed ha una struttura complementare al codone riportato sull’mRNA), 3. Sito di riconoscimento dell’enzima che attiva l’amminoacido (specifico), 4. Sito di riconoscimento del ribosoma. I tRNA sono più di 20 (45-51 negli eucarioti), per cui ad ogni amminoacido corrisponde più di un tRNA. Invece, gli enzimi che legano l’amminoacido al rispettivo tRNA (chiamati aminoacil-tRNA-sintetasi) sono 20, esattamente come il numero degli amminoacidi. Ciascun enzima trasferisce un determinato tipo di amminoacido al rispettivo tRNA. Legame tra tRNA e amminoacido È un processo che avviene secondo una specifica sequenza di eventi. In particolare: 1. Un aminoacil-tRNA-sintetasi si lega ad un ATP. 2. L’enzima si attiva e, in maniera specifica, si lega all’aminoacido. 3. Il gruppo enzima + amminoacido + ATP si lega al braccio destro del tRNA (dove c’è il sito di attacco per l’enzima). 4. Grazie all’energia fornita dall’ATP, l’amminoacido si stacca dall’enzima e si va a legare sull’estremità 3’ del tRNA (sito di attacco dell’amminoacido). 5. L’aminoacil tRNA sintetasi effettua una verifica dell’amminoacido legato (mediante un suo componente, detto proof-reading). Dopo la verifica, se l’amminoacido è corretto, il complesso tRNAamminoacido si stacca e si rende disponibile per la sintesi proteica. Se l’aminoacido è sbagliato, il proof-reading lo stacca dal tRNA ed amminoacido e tRNA ritornano liberi nel citoplasma. Ribosomal RNA (rRNA) Le molecole di rRNA servono a formare i ribosomi. Ribosomi 1 Citologia 19 – RNA (parte 2) I ribosomi sono organuli formati da RNA-ribosomiale (rRNA) e proteine, presenti nel citoplasma, sulla membrana nucleare e sul RER. Il ribosoma (80S; S è il coefficiente di sedimentazione) è formato da 2 subunità diverse, tenute assieme da magnesio. La subunità maggiore (60S) è formata da: rRNA 28 S. L’rRNA 28S ha 3 importanti funzioni: Strutturale: si piega in conformazioni tridimensionali che formano l’impalcatura sulla quale si legano le proteine ribosomiali. Enzimatica: lega assieme gli amminoacidi (attività enzimatica peptidiltransferasi) durante la sintesi proteica. Per questo motivo l’rRNA 28S è considerato un ribozima, cioè un enzima a RNA (o RNA catalitico), una molecola di RNA in grado di svolgere una reazione chimica1. Legame con il tRNA, in quanto contiene i siti di attacco per l’RNA-trasportatore. rRNA 5.8 S rRNA 5 S 50 proteine (distribuite prevalentemente in superficie). La subunità minore (40 S) è formata da: rRNA 18 S. Ha 3 funzioni: Strutturale Riconoscimento del tRNA È coinvolto nel posizionamento corretto dell’mRNA. 33 proteine Essa contiene il sito di attacco per l’RNA-messaggero; quest’ultimo viene accolto in una specie di tunnel determinato dal ripiegamento della molecola dell’rRNA 18S. 1 Sebbene la maggior parte dei ribozimi abbia una concentrazione cellulare irrilevante, il loro ruolo è essenziale per la vita. L'RNA può anche essere considerato una molecola che viene ereditata tra cellula madre e figlia (come avviene per il DNA). Ciò ha indotto Walter Gilbert a proporre che nel passato le cellule primordiali si servissero di RNA sia come materiale genetico che come molecola strutturale e catalitica: solo successivamente questi ruoli vennero affidati a DNA e proteine. Questa ipotesi è nota come ipotesi del mondo a RNA. Queste considerazioni implicano che i ribozimi presenti oggi nelle cellule, così come i ribosomi stessi, siano da considerare fossili viventi di una vita ancestrale basata esclusivamente sugli acidi nucleici. 2 Citologia 19 – RNA (parte 2) Tasche ribosomiali Nei ribosomi ci sono 3 tasche, dette siti, che accolgono i tRNA durante la sintesi delle proteine, ed un tunnel: Sito A (acceptor = accettore), dove si attaccano i tRNA con l’amminoacido attivato da inserire nella catena polipeptidica. Sito P (peptidyl = peptidico), dove viene trattenuta la catena polipeptidica in fase di allungamento, ancora attaccata all’ultimo tRNA. Sito E (exit=uscita), detto anche sito di uscita; è il sito in cui attendono i tRNA che hanno fornito l’amminoacido alla catena prima di uscire dal ribosoma. Tunnel, contenuto nella sub-unità maggiore; da esso fuoriesce la catena polipetidica nascente. Ribosomi mitocondriali Nelle cellule eucariotiche ci sono, oltre ai ribosomi citoplasmatici, anche quelli mitocondriali. Questi assomigliano molto, sia come struttura che come funzione, ai ribosomi procariotici (60S con subunità di 50S e 30S). Sintesi dei ribosomi I ribosomi si formano dentro al nucleo. Specificatamente, l’rRNA viene sintetizzato nel nucleo, mentre la componente proteica è sintetizzata nel citoplasma e successivamente trasportata nel nucleo, ove viene unita agli rRNA. I ribosomi così formati, abbandonano quindi il nucleo (attraverso i pori) e migrano nel citoplasma. Il nucleolo è formato da particolari regioni cromosomiche, dette NORs (nucleolus organizer regions) che sono presenti sul braccio corto dei cromosomi acrocentrici (cromosomi 13, 14, 15, 21 e 22). Nelle NORs sono localizzati i geni per gli rRNA 5.8S, 18S e 28S. I geni per il 5S sono invece localizzati al di fuori dei NORs sul cromosoma 1. I geni per gli rRNA presenti nelle NORs sono disposti in successione, separati tra loro da brevi sequenze "spaziatrici" non tradotte. Pertanto, dalla trascrizione di questi geni si forma dapprima un filamento immaturo di rRNA (pre-rRNA), contenente sia gli rRNA 28S, 18S e 5.8S che le sequenze non tradotte. Poi, questo filamento subisce dapprima un processo di pseudouridilazione e metilazione e poi viene sottoposto a taglio ed eliminazione delle sequenze non utili, fino ad arrivare agli rRNA 28S, 18S, 5.8S. 3 Citologia 19 – RNA (parte 2) Small Cytoplasmic RNA (scRNA) Sono piccole molecole di RNA presenti nel citoplasma. Il loro ruolo funzionale non è stato ancora del tutto chiarito. Uno di questi (scRNA 7S) è coinvolto, assieme ad alcune proteine, nel processo di importazione delle proteine destinate alla secrezione, nel reticolo endoplasmatico. Small Interfering RNA (siRNA) Sono piccole molecole (19-21 nucleotidi) a doppio filamento, sfasati di un paio di nucleotidi. Il precursore del siRNA è una molecola di RNA a doppio filamento (dsRNA = double strand RNA), generalmente di origine esogena (dsRNA virali o artificiali), e raramente endogeni (derivanti da unità ripetitive, pseudogeni e trasposoni). I siRNA fanno parte del sistema RNAi (RNA interference), un processo che blocca l’espressione di singoli geni. Un uso particolare del sistema RNAi è il knock-out artificiale dei geni, ovvero il silenziamento dell’espressione di uno più geni per studiarne gli effetti e le conseguenze e quindi le funzioni svolte. Il processo di maturazione che seguono i siRNA è il seguente: I doppi filamenti di RNA dei precursori dei siRNA vengono tagliati da un enzima (chiamato Dicer), producendo piccole sequenze di dsRNA (19-21 paia di basi), chiamati ds-siRNA (double strand-siRNA = siRNA a doppio filamento). I ds-siRNA si associano al complesso enzimatico RISC (dall'inglese RNA Induced Silencing Complex = complesso silenziatore indotto dall’ RNA). L'RNA a doppio filamento viene aperto (probabilmente da una elicasi) e uno dei due filamenti (quello senso) viene degradato, mentre l’altro (quello antisenso) rimane associato a RISC, formando il complesso siRISC. Il siRISC effettua una scansione degli mRNA presenti nel citoplasma fino a trovarne uno complementare al frammento di RNA antisenso associato al complesso stesso. Generalmente, l'appaiamento tra siRNA e mRNA è perfetto. Dopo l’appaiamento delle basi, un componente della RISC effettua un taglio dell'mRNA; i due frammenti di mRNA risultanti, privi di cappuccio al 5' uno e di coda di poliA al 3' l'altro, vengono rapidamente degradati. Micro RNA (miRNA) 4 Citologia 19 – RNA (parte 2) Sono piccole molecole (20-22 nucleotidi) monofilamento. Di origine endogena, derivano da precursori a forma di forcina (con tratti circolari a filamento unico e tratti a doppio filamento), trascritti da geni non codificanti proteine, ad opera della RNA-polimerasi II. I miRNA individuati nell’uomo sono circa 500 (dati del 2011) ma si stima che ne siano almeno 1000. Sono considerati i più importanti regolatori dell’espressione genica (il 33% dei geni umani è controllato, nell’espressione, dai miRNA). Inoltre, i miRNA sono coinvolti nei tumori e in molte altre malattie. I miRNA intervengono, come i siRNA, nel processo di RNAi; essi però sono più numerosi e seguono un processo di attivazione diverso: I precursori dei miRNA, piuttosto grandi, vengono ridotti, attraverso diversi passaggi (dagli enzimi drosha e dicer), a piccoli frammenti di 20-21 nucleotidi (ds-miRNA). Ciascun ds-miRNA si lega al complesso proteico RISC (miRISC) che prima distrugge uno dei due filamenti e poi guida l’altro verso l'mRNA complementare. Una volta raggiunto, l’mRNA subisce essenzialmente un blocco funzionale. Una degradazione dell’mRNA è stata supposta nei casi in cui la corrispondenza tra sequenza del miRNA e mRNA sia alta. In generale, però, il legame tra miRNA e mRNA non è perfetto. Quest’apparente difetto, li rende meno specifici ma, al tempo stesso, attivi verso un maggior numero di mRNA. RNA nucleari Gli RNA nucleari sono: gli small nuclear RNA (snRNA), gli small nucleolar RNA (snoRNA) e gli heterogeneous nuclear RNA (hnRNA). Small Nuclear RNA (snRNA) Sono molecole di RNA, formate da circa 150 nucleotidi, localizzate nel nucleo e coinvolti nelle prime fasi di maturazione degli mRNA. In associazione con alcune proteine, formano gli snRPN (small nuclear ribo-nucleoproteins, dette snurps) che unendosi costituiscono lo spliceosoma, responsabile del processo dello splicing. Sono state individuate almeno 6 classi di snRNA: U1 snRNA, U2 snRNA, U3 snRNA, U4 snRNA, U5 snRNA, e U6 snRNA, che danno origine a 6 corrispondenti classi di snurps (U1, U2, U3, U4, U5, U6). 5 Citologia 19 – RNA (parte 2) Di questi, tutti, ad eccezione dell’U3, fanno parte dello spliceosoma. Small Nucleolear RNA (snoRNA) Sono molecole di RNA formate da 60-300 nucleotidi, localizzati nel nucleolo e coinvolti nella maturazione degli RNA pre-ribosomiali. Infatti, gli RNA ribosomiali maturi che vanno a costituire il ribosoma, per poter funzionare correttamente, devono subire modificazioni chimiche. Queste avvengono proprio grazie agli snoRNA. Negli eucarioti, quasi tutti gli snoRNA fino ad oggi individuati possono essere suddivisi in due grandi famiglie, a seconda delle sequenze che hanno conservato nel corso dell’evoluzione: snoRNA di tipo C/D, che hanno conservato i box C (5’RUGAUGA3’) e D (5’CUGA3’). Talvolta, sono presenti copie addizionali dei due box. In questo caso, si parla di box C’ e box D’. Gli snoRNA C/D hanno una forma a singola forcina. La loro funzione è metilare (aggiungere gruppi CH3) il pre-rRNA (aggiungere gruppi CH3). Il sito operativo che svolge la metilazione è compreso tra i box C e D. Un sottogruppo degli snoRNA C/D (U3, U8, U14 e U22), svolge una funzione di taglio sui pre-rRNA. snoRNA di tipo H/ACA, che hanno conservato chiamato box H (ANANNA, dove N sta per qualsiasi nucleotide) e la box ACA Gli snoRNA H/ACA presentano una forma a doppia forcina (hairpin), con un filamento singolo (hinge) che unisce le due forcine. Il box H si trova sul filamento hinge, mentre il box ACA si trova a tre nucleotidi di distanza dall’estremità 3’. Gli snoRNA della classe H/ACA sono coinvolti nella conversione delle uridine in pseudo-uridine. La parte operativa (quella che esegue la funzione di conversione) è contenuta all’interno della grande bolla delle forcine. L’operazione di taglio viene effettuata da un particolare sottogruppo di snoRNA appartenenti alla classe C/D. E’ importante ricordare che entrambe le classi di snoRNA interagiscono con diverse e specifiche proteine per espletare le proprie funzioni. L’associazione con queste proteine è fondamentale per le operazioni di taglio, conversione e metilazione. Heterogeneus Nuclear RNA (hnRNA) 6 Citologia 19 – RNA (parte 2) Si tratta di un gruppo assai indefinito di molecole che comprende tutte le forme di pre-RNA (cioè tutte le molecole di RNA che devono ancora maturare. A volte, gli hnRNA si congiungono ad alcune proteine, formando gli hnRNP, assumendo funzioni non ancora definite. Nota. Le cellule eucariotiche necessitano di grandi quantità di rRNA (perchè hanno bisogno di molte proteine). Per questo motivo, esse sono dotate di molte copie dei geni dell’rRNA, disposte testa-coda. La figura mostra molecole di RNA-polimerasi I mentre trascrivono geni di rRNA adiacenti (TU = transcription unit, NTS = nontranscribed spacer). 7 Citologia 19 – RNA (parte 2)
© Copyright 2026 Paperzz