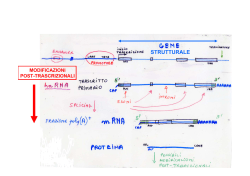
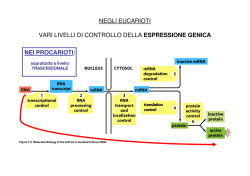
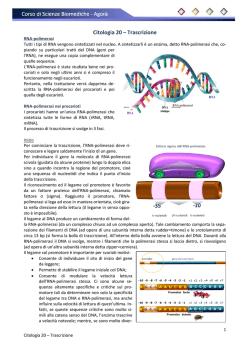
TRASCRIZIONE E TRADUZIONE Due tra gli avvenimenti principali che avvengono all’interno della cellula, si susseguono rispettivamente nel nucleo (la trascrizione) e nel citoplasma (la traduzione). La trascrizione è un fenomeno altamente selettivo: non tutto il DNA viene trascritto, bensì solo alcune parti. Le sequenze trascritte sono chiamate geni. LA TRASCRIZIONE In biologia molecolare, la trascrizione è il processo mediante il quale le informazioni contenute nel DNA vengono trascritte enzimaticamente in una molecola complementare di RNA. Concettualmente, si tratta del trasferimento dell'informazione genetica dal DNA all'RNA. Nel caso in cui il DNA codifichi una proteina, la trascrizione è l'inizio del processo che porta, attraverso la produzione intermedia di un mRNA, alla sintesi di peptidi o proteine funzionali. LA TRASCRIZIONE – CENNI GENERALI La trascrizione avviene attraverso particolari enzimi detti genericamente RNA-polimerasi. Tali proteine sono spesso denominate RNA-polimerasi DNA-dipendenti, dal momento che producono una molecola di RNA a partire da una di DNA. Tali enzimi utilizzano nucleotidi trifosfati (ovvero nucleotidi con tre gruppi fosfato) per formare l'RNA. Durante il processo, dai nucleosidi trifosfati vengono rimossi due gruppi fosfato per formare un legame covalente tra un nucleotide e quello successivo. LA TRASCRIZIONE – CENNI GENERALI La RNA polimerasi si lega al DNA solo presso particolari sequenze, dette promotori, che non sono trascritte. Dal promotore iniziano a inserirsi i nucleosidi trifosfato per formare una sequenza di nucleotidi che sarà complementare al filamento di DNA in via di trascrizione. Dopo l'individuazione del promotore, la RNA polimerasi rende il DNA adatto alla trascrizione. Il filamento di RNA inizia quindi ad allungarsi, attraverso l'aggiunta di un nucleotide per volta. Il primo nucleotide del neofilamento di RNA trattiene i tre gruppi fosfato mentre quelli successivi vengono privati di due gruppi fosfato attraverso una reazione esoergonica. • Quando, durante la trascrizione, nel DNA si incontrano particolari sequenze di basi (poste solitamente alla fine di ogni gene), la trascrizione termina. LA TRASCRIZIONE NEGLI EUCARIOTI Dato l'elevato numero di geni presenti in un organismo eucariote, la trascrizione è un meccanismo molto più complesso perché necessita di un sistema di regolazione che agisca su più livelli. Una prima notevole differenza sta proprio nelle RNA Polimerasi, esse infatti vengono suddivise in tre categorie in funzione dell'ambito genetico in cui lavorano. • L' RNA Polimerasi I presente nel nucleolo è capace di trascrivere i geni per tre dei quattro rRNA ovvero 5.8S, 18S e 28S. • L'RNA Polimerasi II che si trova nel nucleoplasma (parte di nucleo che non contiene il nucleolo) trascrive gli mRNA, i miRNA, i siRNA, alcunisnoRNA e snRNA. • Infine l'RNA Polimerasi III che si trova nel nucleoplasma trascrive l'rRNA 5S, tutti i geni dei tRNA e alcuni snRNA. LA TRASCRIZIONE NEGLI EUCARIOTI Da un solo gene, la trascrizione dà luogo a molti m-RNA ⇒ amplificazione delle informazioni. Come per la replicazione del DNA, la trascrizione procede in direzione 5' → 3'. Più precisamente, il filamento lungo il quale il DNA viene scandito, detto filamento stampo, è percorso dall'enzima in direzione 3' → 5'. Il nuovo filamento di RNA, iden7co al filamento senso viene sintetizzato a partire dal suo 5'. La trascrizione si svolge in varie fasi. LA TRASCRIZIONE – INTRODUZIONE • Il messaggio genetico è frammentato, infatti un gene comprende gli esoni, cioè sequenze contenenti l'informazione ereditaria, che generalmente verranno trascritte e tradotte in proteine, e gli introni, cioè sequenze intercalari, che verranno trascritte ma non tradotte. • Nella fase iniziale vi è la trascrizione integrale degli esoni e degli introni. Gli esoni dunque, a differenza degli introni, in seguito al processo di splicing del trascritto primario, detto hnRNA, si ritrovano negli mRNA maturi. Intervengono le RNA-polimerasi I, II, III: i 2 filamenti di DNA si scostano per rottura dei ponti-H, al passaggio dell'enzima, e ha luogo la trascrizione di un filamento di DNA. Subito dopo si riformano i pontiH. L’enzima riconosce il punto d’inizio grazie a un promotore, che comprende anche il TATA-box. Il segnale di fine gene è una sequenza AATAAA, detta poliadenilazione. LA TRASCRIZIONE – L’INIZIO Convenzionalmente la trascrizione genica inizia quando a livello del promotore si assembla una classe di proteine nota come fattori generali di trascrizione della Polimerasi II (TFII). Il compito di queste proteine è di segnalare alla RNA Polimerasi II la collocazione del promotore su un dato gene e le sue sequenze consenso, stabilizzarla in loco, aprire la doppia elica di DNA, attivare la RNA Polimerasi II e svolgere tutte quelle funzioni che garantiscono un corretto inizio del processo di trascrizione. Ci sono cinque fattori generali di trascrizione essenziali: TFIIB, TFIID, TFIIE, TFIIF e TFIIH. TFIID è descritto come il primo ad entrare in gioco, si tratta di un grosso complesso proteico formato da 12 subunità, tra queste particolarmente importante è la subunità TBP (TATA binding protein). LA TRASCRIZIONE – L’INIZIO La TBP si lega a livello del TATA-box, una sequenza consenso presente a monte di molti geni generalmente ad una distanza di 25-30 nucleotidi dal punto di inizio della trascrizione. Questo forte angolo non sarà la causa dell’apertura del promotore (o melting) ma ha la funzione di allargare i solchi maggiori al fine di favorire una migliore interazione delle altre proteine del complesso di attivazione basale. LA TRASCRIZIONE – SECONDA FASE In realtà la TBP intercetta la sequenza TATA spesso legata ad altri fattori chiamati TAF's, e in questa forma viene chiamata TFIID (“TBP+TAF's” = TFIID) per formare il complesso di inizio preliminare. A questo punto la topologia del DNA è tale da poter reclutare altri fattori di trascrizione ovvero il TFIIA e il TFIIB. Tale complesso trimerico TBP TFIIA TFIIB e chiamato anche DAB. Il complesso DAB così assemblato è capace di legare un quarto fattore di trascrizione che porta con sé la RNA Polimerasi II che è TFIIF. LA TRASCRIZIONE – TERZA FASE Queste prime fasi, solitamente di stallo, diventano processive e progressivamente più rapide con una serie di reazioni che causano l’allargamento conformazionale alla RNA polimerasi II, che se opportunamente fosforilata nella regione C-Terminale supera la fase di stallo. • Nonostante ciò nelle prime fasi la velocità di trascrizione non può essere elevata a causa delle ancora forti interazioni con tutti gli altri fattori ancora legati alla RNA Polimerasi II. La fase di pulizia del promotore avviene grazie al rilascio del DAB e di TFIIF che avevano inizialmente il solo ruolo di stabilizzare il complesso di inizio. Tale rilascio avviene grazie a opportune fosforilazioni/defosforilazioni della RNA Polimerasi II a livello del dominio C-Terminale che potremmo definire una sorta di pannello di controllo su cui accedono molti altri regolatori della trascrizioni sia in fasi precoci che tardive. LA TRASCRIZIONE - TERMINAZIONE l terminale 3’ dell’RNA eucariotico è definito da modifiche posttrascrizionali, in pratica l’RNA trascritto viene accorciato in seguito al taglio a livello di una specifica sequenza che si trova 20-30 nucleotidi a valle del sito di poliadenilazione (AAUAAA); immediatamente dopo il taglio una specifica polimerasi aggiunge al 3’ del nuovo trascritto una coda di poli-A (ca. 200 nucleotidi). Funzioni della coda poli-A: • esportare l’mRNA maturo dal nucleo; • influenzare la stabilità degli mRNA nel citoplasma LA TRASCRIZIONE INVERSA Per retrotrascrizione (o trascrizione inversa) si intende una trascrizione che non segue il cosiddetto dogma centrale della biologia molecolare, secondo il quale l'informazione genetica fluisce dal DNA, all'RNA, alle proteine. La retrotrascrizione è infatti mediata dall'enzima trascrittasi inversa, che catalizza la sintesi di DNA complementare ad un RNA. Tale evento è frequente nei virus a RNA (come l'HIV ed il virus del sarcoma di Rous, il primo ad essere stato scoperto), che sono in grado di replicarsi solo attraverso la conversione del loro genoma da RNA a DNA. LA TRADUZIONE La traduzione è il processo biochimico attraverso il quale l'informazione genetica contenuta nel mRNA (RNA messaggero), viene convertita in proteine che svolgono nella cellula un'ampia gamma di funzioni. La sintesi proteica inizia da un filamento di mRNA, prodotto a partire da un gene sul DNA attraverso il processo di trascrizione. Questo filamento è usato come stampo per la produzione di una specifica proteina. mRNA L'mRNA è il trascritto di un gene nel linguaggio dell'RNA, che prevede quattro basi, identiche al DNA fatta eccezione per l'uracile che sostituisce la timina. Un mRNA maturo appena esportato dalla nucleoplasma attraverso la membrana nucleare è stato inoltre modificato mediante aggiunta di una 5'-metilguanina all'estremità 5' (cappuccio dell'mRNA), mediante una coda poliadenilata (AAAAA...) all'estremità 3' e associato a diverse tipologie di proteine utili a far riconoscere al resto della cellula l'avvenuta maturazione ed esportazione di questo fuori dal nucleo. mRNA Il ribosoma legge le basi dell'mRNA a triplette (dette codoni) e a ciascuna tripletta (per un totale di 64, che si ottiene elevando il numero delle basi dell'mRNA, cioè 4, al numero di cifre di una tripletta, 3) fa corrispondere un amminoacido rispettando il codice genetico. In realtà, essendo il codice ridondante, alcune triplette codificano per lo stesso amminoacido e solo due amminoacidi sono specificati da una sola tripletta (AUG per la metionina e UGG per il triptofano), esistono inoltre triplette che specificano la fine della traduzione, dette codoni di stop, sono UAA, UAG e UGA, esse non codificano per nessun amminoacido. mRNA Il ribosoma legge le basi dell'mRNA a triplette (dette codoni) e a ciascuna tripletta (per un totale di 64, che si ottiene elevando il numero delle basi dell'mRNA, cioè 4, al numero di cifre di una tripletta, 3) fa corrispondere un amminoacido rispettando il codice genetico. In realtà, essendo il codice ridondante, alcune triplette codificano per lo stesso amminoacido e solo due amminoacidi sono specificati da una sola tripletta (AUG per la metionina e UGG per il triptofano), esistono inoltre triplette che specificano la fine della traduzione, dette codoni di stop, sono UAA, UAG e UGA, esse non codificano per nessun amminoacido. I tRNA Il tRNA è una piccola molecola di RNA composta mediamente da 80nucleotidi. Ha una caratteristica forma simile a quella di un quadrifoglio (la forma è data dal fatto che i nucleotidi che lo compongono sono appaiati in modo tale da originare una molecola con quattro sporgenze; due di queste sono più importanti: la prima è costituita da due nucleotidi non appaiati che sono sempre gli stessi per tutte le molecole di tRNA, cioè A C C; la seconda molecola è opposta alla prima, è costituita da una tripletta di basi anch' esse non appaiate che sono diverse per ciascuno dei 61 diversi tipi di tRNA) tRNA Vi sono quattro aree di particolare interesse in ciascuna molecola di tRNA, in particolare due anse, dette ansa D e ansa T (spesso qui si concentrano i nucleotidi modificati), l'anticodone, specifico per ciascun tRNA e complementare ad un codone del mRNA e l'attacco dell'amminoacido corrispondente al codone all'estremità 3'. Esistono più tRNA per uno stesso amminoacido (infatti vi sono solo 48 anticodoni diversi codificati da circa 500 geni) e nel contempo un tRNA può associarsi a più di un codone. Questo accade perché ciascun anticodone di tRNA si associa saldamente solo alle prime due basi di un codone, mentre è poco specifico e più tollerante per la terza base, tanto che non raramente si verificano appaiamenti sbagliati, causando il fenomeno dello wobblingcioè il tentennamento della terza posizione. tRNA I tRNA che intervengono nella sintesi proteica sono il prodotto di pre-tRNA più lunghi che vengono modificati nel nucleo grazie ad uno speciale splicing che segue un meccanismo "taglia e cuci", diverso da quello comune che prevede la formazione di una sorta di cappio per rimuovere le sequenze introniche. Successivamente unamminoacil-tRNA-sintetasi specifica per ciascun amminoacido (circa 20 tipi diversi) accoppia covalentemente l'amminoacido corrispondente all'estremità 3' di ciascun tRNA. I RIBOSOMI Un ribosoma è un grande ribozima e rappresenta la macchina esecutrice della sintesi proteica. Possiede due grandi subunità, la maggiore (60 S) e la minore (40 S); ciascuna ribosoma, oltre agli rRNA, contiene anche proteine, con una composizione complessiva dei 2/3 in RNA e per 1/3 in proteine. La subunità minore funge da sostegno e da sito di ingresso dell'mRNA da tradurre, mentre la subunità maggiore è la principale macchina catalizzatrice del complesso. Quando sono unite le due subunità possiedono quattro siti d'attacco, uno per l'mRNA e i siti E, P, A per i tRNA in arrivo o in uscita. • Un ribosoma umano aggiunge circa 2 amminoacidi al secondo TRADUZIONE NEGLI EUCARIOTI La sintesi proteica inizia sempre dall'Nterminale di una proteina in formazione verso il suo C-terminale. Il primo amminoacil-tRNA ad essere aggiunto è invariabilmente quello con legata metionina, detto tRNA iniziatore, poiché la sequenza d'inizio di ciascun mRNA è AUG. Generalmente la metionina è sempre rimossa alla fine della traduzione da una proteasi specifica. La subunità minore del ribosoma si attacca all'estremità 5' dell'mRNA che viene riconosciuta poiché possiede un cappuccio di 7metilguanosina e i fattori di inizio eIF4G e eIF4E, poi, una volta trovata, vi si attacca anche la subunità maggiore e quindi il ribozima cerca la prima tripletta AUG sull'mRNA che è il suo segnale d'inizio della sintesi proteica. TRADUZIONE NEGLI EUCARIOTI Il movimento della subunità minore è ulteriormente facilitato dall'idrolisi di ATP in ADP+P. Il tRNA si lega al sito P del ribosoma, mentre il tRNA con l'anticodone corrispondente al codone successivo nell'mRNA legato al fattore d'allungamento EF1 si attacca nel sito A. EF1 unisce il tRNA al sito A ed effettua un controllo di qualità per non venire aggiunto se la corrispondenza codone-anticodone non è corretta. EF1 inoltre lega il tRNA sull'mRNA in una conformazione curva che impedisce l'immediato legame dell'amminoacido legato al tRNA cui è attaccato al resto del polipeptide in crescita. Una volta riconosciuto un appaiamento corretto, l'rRNA sulla subunità minore idrolizza il GTP legato a EF1 a GDP e si stacca dal tRNA. TRADUZIONE NEGLI EUCARIOTI A questo punto la peptidil transferasi, contenuta nella subunità maggiore del ribosoma, catalizza lo spostamento del legame che unisce l'amminoacido al suo tRNA nel sito P formando un legame peptidico tra l'amminoacido del tRNA presso il sito P e quello presso il sito A. La subunità maggiore si sposta quindi di tre nucleotidi verso l'estremità 3' dell'mRNA e così fa anche la subunità minore così che, alla fine degli spostamenti, il primo tRNA aggiunto si trova nel sito E e il secondo nel sito P mentre nel sito A si lega il fattore di allungamento EF2 con legato GTP. Il primo tRNA ormai privo di amminoacido si stacca dal sito E ed esce dal ribosoma, mentre un nuovo amminoacil-tRNA si attacca al sito A, il GTP di EF2 è idrolizzato a GDP e EF2 si stacca dal sito A. Quando il codone di stop è raggiunto nella fase di terminazione il ribosoma cattura una molecola d'acqua che idrolizza il polipeptide ormai completo il quale si distacca dal ribosoma. Il processo è coadiuvato da fattori di rilascio (RF1, RF2, RF3 nei batteri, RF1 ed RF3 negli eucarioti), proteine che simulano l'azione del tRNA, si legano al sito A e liberano la proteina nel citoplasma (fenomeno conosciuto come mimetismo molecolare).
© Copyright 2026 Paperzz