Program and Abstracts for the 30th Annual Meeting of the Society of
Population Ecology
第 30 回個体群生態学会大会 プログラム・講演要旨集
10-12 October 2014 University of Tsukuba
2014 年 10 月 10 日(金)∼ 12 日(日)筑波大学
!"#"$%"$&%'#()'*+,-.#"$+('/&+.+01
A76B-6("$%"C
?#16%$#(@
/>$)6("$#.
23435#,67
839#:#:-7#
;323<+7#=$+
!&#.$(0'D7+:'4$D6'E$%"+71'57#$"%
"+'"F6'*+,-.#"$+('<1(#:$&%
#
G3H3I6.%+( J3I3?K+7(%"
$&F$
#) 539#:#(#L 239#:#:
Statistical
significance is
not always
ecological one
Clinics on
Statistics
Organizing Committee of the 30th Annual Meeting of the Society of Population Ecology
第 30 回個体群生態学会大会実行委員会
Program and Abstracts for the 30th Annual Meeting of the Society of
Population Ecology
第 30 回個体群生態学会大会 プログラム・講演要旨集
10-12 October 2014
2014 年 10 月 10 日(金)∼ 12 日(日)
University Hall, University of Tsukuba, Tsukuba City, Ibaraki, JAPAN
筑波大学 大学会館・総合交流会館(茨城県つくば市)
Host: Organizing Committee of the 30th Annual Meeting of the Society of Population
Ecology
主催:第 30 回個体群生態学会大会実行委員会
Co-hosts: The Society of Population Ecology
University of Tsukuba
Colleges of Biological Sciences, University of Tsukuba
共催:個体群生態学会
筑波大学
筑波大学生命環境学群生物学類
Sponsor: Springer Japan
後援:シュプリンガー・ジャパン
Contents
Access 交通アクセス
2
Campus & Restaurant Map キャンパス・飲食店マップ
4
Schedule 大会日程・時間割
5
Venue 会場案内
6
News and notifications 注意事項とお知らせ
7
Population Ecology Young Scientist Award: Award Ceremony and Award
Lecture 個体群生態学会奨励賞 授賞式・受賞講演
12
Autumn School ‘Clinic on Statistics’ 秋の学校「行列のできる統計相談所」
13
Plenary Symposia 基調シンポジウム
14
Plenary Symposium A . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 14
Plenary Symposium B . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 16
Organized Sessions 企画シンポジウム
19
Organized Session 1 . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 19
Organized Session 2 . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 22
Organized Session 3 . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 25
Poster Presentation 一般講演(ポスター)
28
Poster Titles 講演タイトル . . . . . . . . . . . . . . . . . . . . . . . . . . . . 28
Poster Abstracts 講演要旨 . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 31
List of Attendants 大会参加者名簿
48
Bus Timetables バス時刻表
50
Taxi タクシー
54
1
Access 交通アクセス
From Tokyo to Tsukuba by train and bus 電車とバス利用の場合
つくば駅から大学会館(会場)
Tsukuba ~ Daigaku-kaikan (Univ. Hall, Venue)
Nourin-gijutsu-center
農林技術センター
東京からつくば駅
Nijino-hiroba
虹の広場
Tokyo ~ Tsukuba
JR Yamanote Line
JR山手線
Akihabara
秋葉原
Tokyo
東京
Hamamatsucho
浜松町
Tokyo Monorail
東京モノレール
Haneda Airport
羽田空港
Univ. of Tsukuba
筑波大
45min
Daisan-area-mae
第三エリア前
Route Bus
路線バス
10min
Tsukuba Express
つくばエクスプレス
Ichinoya-gakusei
shukusya-mae
一ノ矢学生宿舎前
Daiichi-area-mae
第一エリア前
Daigaku-kaikan-mae
大学会館前
Tsukuba-daigaku-nishi
筑波大学西
Hirasuna-gakusei
shukusya-mae
平砂学生宿舎前
Oikoshi-gakusei
shukusha-mae
Tsukuba追越学生宿舎前
daigaku-junkan
Migi-mawari Tsukubadaigaku
筑波大学循環
byouin-iriguchi
筑波大学病院入口
右回り
Tsukuba
つくば
Highway Bus 高速バス
65min
Narita Express
成田エクスプレス
Highway Bus 高速バス
100min
Narita Airport
成田空港
Tsukuba-medical-center-mae
筑波メディカルセンター前
Tsukubadaigaku-kasuga-area-mae
筑波大学春日エリア前
Azuma-shogakkou-mae
吾妻小学校前
Highway Bus 高速バス
120min
Tsukuba Center
つくばセンター
Tsukuba Sta.
Daigaku-shokubutsu-mihonen
大学植物見本園
Daini-area-mae
第二エリア前
大学会館
Venue
Medical-center-byoin
メディカルセンター病院
TARA-Center-mae
TARAセンター前
Tsukuba-daigaku-chuo
筑波大学中央
Daigaku-kouen
大学公園
Matsumi-ike
松美池
Amakubo-sanchoume
天久保三丁目
Gasshukujo
合宿所
Amakubo-ike
天久保池
Amakubo-nichoume
天久保二丁目
Oikoshi-shukusha-higashi
追越宿舎東
Tsukuba-daigaku-junkan
Hidari-mawari
筑波大学循環
左回り
つくば駅
By Tsukuba Express (TX) train つくばエクスプレス (TX) をご利用の場合
45 min. using rapid service to ‘Tsukuba Sta.’ (terminal) from ‘Akihabara Sta.’ (1,190
yen) ♣
秋葉原駅からつくば行きに乗車、つくば駅(終点)で下車(快速 45 分、1,190 円)。♣
By highway bus from Tokyo Sta. 東京駅から高速バスをご利用の場合
Approx. 65 min. to ‘Daigaku-kaikan’ (i.e., ‘University Hall’) from ‘Tokyo Sta. Yaesu
South Exit’ bus terminal, platform No. 2 (1,180 yen)
東京駅八重洲南口バスターミナル 2 番のりばから、高速バス「筑波大学」行きに乗車、
「大学会館」で下車(約 65 分、1,180 円)、徒歩 0 分。運行間隔は 20-30 分。
From Narita Airport (New Tokyo International Airport) 成田空港から
Approx. 100 min. to the ‘Tsukuba Center’ bus terminal from Narita Airpot by
Limousine Bus (2,600 yen)♣
成田空港第 1 または 第 2 ターミナルから、高速バス「土浦・つくば」行きに乗車、
「つ
♣
くばセンター」で下車(約 100 分、2,600 円)。
From Haneda Airport (Tokyo International Airport) 羽田空港から
Approx. 120 min. to ‘Tsukuba Center’ bus terminal from Haneda Airpot by Limousine
Bus (1,850 yen)♣
羽田空港から、高速バス「つくばセンター」行きに乗車、「つくばセンター」(終点) で
下車(約 120 分、1,850 円)。♣
2
♣ From Tsukuba Sta. to Univ. Hall (Venue) つくば駅から会場まで
About 10 min. to ‘Daigaku-kaikan-mae’ (i.e., ‘University Hall Ent.’) from ‘Tsukuba
Center’ platform No. 6 by Route Bus (200 yen). Route No. 10, 20, 30, 30A (bound for
‘Tsukuba-daigaku-chuo’), and C10 (‘Tsukuba-daigaku-junkan’) are available.
つくばエクスプレス(TX)つくば駅に隣接するバスターミナル「つくばセンター」6
番のりばから、関東鉄道路線バス「筑波大学循環 右回り」または「筑波大学中央」行き
に乗車、「大学会館前」で下車(約 10 分、200 円)、徒歩 0 分。運行間隔は 10-20 分。
なお、「筑波大学循環 左回り」に乗車しても、「大学会館前」で下車できます(約 20
分、200 円)。
Access to Tsukuba by car 自動車利用の場合
Univ. of Tsukuba
筑波大
Daigaku kaikan ( Conference Hall )
大学会館 ( 会場 )
Hiratsuka sen
平塚線
Chuo-guchi
中央口
Hiratsuka-sen
平塚線
Tsukuba Station
つくば駅
Tsutsiurakita I.C
土浦北 I.C
Higashi Odori
学園東大通り
Gakuen sen
Sasagi crossing
大角豆
学園線
Sakura Tsutsiura kita I.C
交差点
Nishi Odori
桜・土浦北 I.C
学園西大通り
Tsukuba Chuo I.C
つくば中央 I.C
Joban Expressway
常磐自動車道
354
Tsukuba
JCT
Yatabe I.C
つくばJCT
谷田部 I.C
Taigeinishi
Gate P
体芸西ゲート
P
大学会館
Hospital
病院
Matsumi-guchi
松見口
Nishi-odiri
西大通り
Higashi-odori
東大通り
Please use FREE parking at the ‘Daigaku-kaikan parking for guests’ (enter through the
Hiratsuka-sen) or at the ‘Taigei-nishi gate parking.’
「大学会館 外来用駐車場」(平塚線沿いから入ることができます)、または「体芸西駐
車場」(大学会館バス停の南へ約 150m に入口ゲートがあります)を無料でご利用いただ
けます。守衛所等での手続きは必要ありません。
3
Campus & Restaurant Map キャンパス・飲食店マップ
University area
University of Tsukuba
11:00-14:00
8:15-14:00
Bus route
Restaurant for Lunch
Cafe
Izakaya (Japanese style bar)
Bento box shop
University Library
JEWEL OF INDIA
XIAN
DALI
BAR
University Hall
200m
Bus route
Pedestrian deck (To Tsukuba station)
4
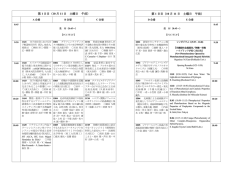
Schedule 大会日程・時間割
Friday, 10 October
Time
Event
Management committee
13:00-16:00
運営委員会
Autumn school ‘Clinic on statistics’ 15:00-18:00
秋の学校「行列のできる統計相談所」
Place
Daisan-kaigishitsu (3F)
3 階第三会議室
Multimedia room (2F)
2 階マルチメディアルーム
Saturday, 11 October
Time
09:00-12:00
12:00-13:00
13:00-14:00
14:00-16:00
16:00-16:40
16:45-18:20
18:30-20:30
Event
Plenary symposium A
基調シンポジウム A
Lunch time
Poster presentation core time
ポスター発表コアタイム
Organized session 1
企画シンポジウム 1
Organized session 3
企画シンポジウム 3
Award ceremony, Award recipient lecture
授賞式・受賞講演
General meeting
総会
Banquet
懇親会
Place
Hall (3F)
3 階ホール
Foyer (2F, 3F)
2 階・3 階ホワイエ
Hall (3F)
3 階ホール
Multimedia room (2F)
2 階マルチメディアルーム
Hall (3F)
3 階ホール
Hall (3F)
3 階ホール
Restaurant ‘Plaza’ (1F)
1 階レストラン「プラザ」
Sunday, 12 October
Time
09:00-12:00
12:00-13:00
13:00-14:00
14:00-16:00
Event
Plenary symposium B
基調シンポジウム B
Lunch time
Poster presentation core time ポスター発表コアタイム
Organized session 2
企画シンポジウム 2
5
Place
Hall (3F)
3 階ホール
Foyer (2F, 3F)
2 階・3 階ホワイエ Hall (3F)
3 階ホール
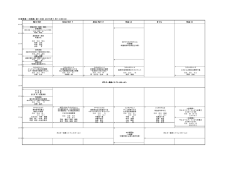
Venue 会場案内
2F
Resting place
休憩室
ラウンジ
Lounge
EV
「大学会館前」バス停
'Daigaku-kaikan-mae'
Bus stop
ホワイエ
Foyer
WC
Reception desk
Poster
マルチメディアルーム
Multimedia room
受付
EV
WC
レストラン
Restaurant
(1F)
統計相談所
企画シンポ3
Clinic on statistics
Organized session 3
Banquet
懇親会 (1F)
WC
3F
Food and drink are allowed
飲食可能なエリア
Management committee
運営委員会
No food or drink allowed in the other areas.
これ以外の場所での飲食はご遠慮ください。
第三会議室
Daisan-kaigishitsu
Springer
Poster
Head office
大会本部
展示
Resting place & cloakroom
&
ホワイエ
Foyer
基調シンポ
企画シンポ1,2
Organized session 1, 2
Award ceremony, Award reception lecture
General meeting
総会
休憩室 クローク
EV
Nursing room
ホール
Hall
Plenary symposium
第二会議室
Daini-kaigishitsu
WC
授賞式、受賞講演
6
授乳室
第一会議室
Daiichi-kaigishitsu
News and notifications 注意事項とお知らせ
Reception 受付
The reception desk (registration counter) will be open at the foyer of University Hall
(2nd floor) during the following time periods.
10th (Fri) 12:00-18:00
11th (Sat) 8:30-18:00
12th (Sun) 8:30-16:00
If you have preregistered online without an advance payment, or if you have not
registered yet, please complete your payment on the day of arrival. The registration fee
is 7,000 JPN for a general PES member, 5,000 JPN for a student member, and 8,000
JPN for a non-member.
Please remember to wear your name tag in the University Hall building.
受付は、以下の時間に、大学会館 2 階ホワイエで行っています。
10 日(金)12:00-18:00
11 日(土)8:30-18:00
12 日(日)8:30-16:00
事前登録をされて参加費・懇親会費が未振込の方、事前登録をされず当日参加される
方は、受付にてお支払いください。参加費は一般会員 7,000 円、学生会員 5,000 円、非会
員 8,000 円です。
会場では名札をおつけください。
Cloak クローク
The cloak will be available at the second meeting room (DAINI-KAIGISHITSU) of
University Hall (3rd floor) during the following time periods.
10th (Fri) 14:00-18:00
11th (Sat) 8:30-18:30 (there will be an alternative cloak at the banquet room
during 18:30-20:30)
12th (Sun) 8:30-16:30
Please keep valuables in your possession at all times. We are not responsible for lost,
stolen, or damaged items. Make sure to pick up all your luggage at the end of the day,
because we cannot keep them overnight!
大学会館 3 階第二会議室にてお荷物をお預かりします。預かり時間は以下のとおり
です。
10 日(金)14:00-18:00
11 日(土)8:30-18:30(18:30-20:30 は懇親会会場に荷物置き場を設けます)
12 日(日)8:30-16:30
貴重品は必ずご自身で管理してください。お預けになった荷物に関しても万一の盗難・
破損の責任は負いかねます。日付をまたいでのお預かりはできません。いずれの日もク
ローク終了時間までに荷物を引き取ってください。
7
Resting & nursing 休憩室・授乳室
Resting room: the second meeting room (DAINI-KAIGISHITSU) of University Hall
(3rd floor) and the lounge of KORYU-KAIKAN (2nd floor)
Nursing room: the first meeting room (DAIICHI-KAIGISHITSU) of University Hall
(3rd floor)
All rooms will be available during the following time periods.
10th (Fri) 14:00-18:00
11th (Sat) 9:00-18:00
12th (Sun) 9:00-16:00
以下の日程で休憩室(大学会館 3 階第二会議室、交流会館 2 階ラウンジ)と授乳室(大
学会館 3 階第一会議室)を開設します。
10 日(金)14:00-18:00
11 日(土)9:00-18:00
12 日(日)9:00-16:00
Symposia シンポジウム
Please start and finish your presentation on time. You will not be allowed to extend the
scheduled time. Liquid-crystal projectors will be provided in all presentation rooms, but
the organizers should be responsible for preparation of computers to which they hook up
the projectors. We will assign a couple of assistants to each presentation room.
設定された時間内に終了するように進行をお願いします。時間延長はできません。シ
ンポジウム の発表には液晶プロジェクターが使用できます。液晶プロジェクターに接続
するコンピューターおよび発表用ファイルは、シンポジウム企画者の責任で準備してく
ださい。開催時間帯の各会場には 1∼2 名のスタッフを配置します。
Poster presentations ポスター発表
Each presenter will be assigned to one poster board indicated by a unique “presentation
number.” Note that the presentation numbers will not always be consecutive between
the adjacent boards, because these are identical to the registration number of the
participants. Please check your registration number on the meeting program and use the
correct board.
Tacks will be provided. You can start putting your poster up on 10th (Fri) 14:00. The
scheduled core time of the presentations is 13:00-14:00 on 11th & 12th. All posters must
be taken down after 16:00 on 12th (Sun).
展示場所は展示ボードに「発表番号」で指示されています。本大会では、ポスターの
発表番号に大会登録番号を転用しているため、発表番号が通し番号になっておりません
のでご注意ください。ポスター発表者は、ご自分の大会登録番号を要旨集等でご確認く
ださい。
ポスターをボードへ貼り付けるための画鋲などは大会事務局で用意しますので、 それ
を用いてください。
ポスターは 10 日(金)14:00 から 12 日(日)16:00 まで掲示することができます。12
日(日) 16:00 以降はすみやかにポスターをお持ち帰りください。
8
ポスター発表のコアタイムは、2 日間設けています。両日とも 13 時∼14 時がコアタイ
ムです。
Awards ポスター賞
We will select one Best Poster Prize and three Excellent Poster Prizes from all poster
presentations, regardless of the career status of the presenters. The winners
(lead/presenting authors) will be announced and awarded during the banquet.
個体群生態学会大会では、優秀なポスター発表を選んでポスター賞を授与します。
対象者:本大会では例年と異なり、一般講演を行う方全員がポスター賞の対象となり
ます。
授賞の件数と表彰方法:得票数順に『最優秀賞』(1 件)および『優秀賞』(約 3 件)を表
彰します。懇親会のなかで表彰式を行ないます。
Photography 会場内での撮影・記録
Please be advised that you are responsible for the copyrights or portrait rights of photos
and videos that you take during the meeting. Regardless of the presentation type, all
phototakers should be meticulously careful not to cause any inconvenience to scientific
activity of the presenters.
著作権、 肖像権を侵害しない範囲で行ってください。シンポジウム講演、ポスターな
ど、すべての形態の研究発表について、 発表者に不利益が生じるかたちでの記録を決し
て行わないようにお願いいたします。
Steering committee 運営委員会
The committee will be held during 13:00-16:00 on 10th (Fri) at the third meeting room
(DAISAN-KAIGISHITSU) of University Hall (3rd floor). Please make sure to be there if
you are a member of the committee.
10 日(金)13:00 から 16:00 まで、大学会館 3 階第三会議室で開催します。運営委員の
方はご参加ください。
PES general meeting 総会
The general meeting will be held during 16:45-18:20 on 11th (Sat) at the hall of
University Hall (3rd floor). Note that PES has just re-started as an incorporated
non-profit organization from this April (for details, see
http://www.population-ecology.jp), and we need to discuss important matters such
as the dissolution of the former PES as a private organization, and the transition of
members to the incorporated NPO. All PES members are highly encouraged to attend
the meeting!
11 日(土)の 16:45 から 18:20 まで、大学会館 3 階ホールで行われます。
今年4月に NPO 法人としての個体群生態学会がスタートしました。今回の総会ではい
よいよ、任意団体としての学会の解散や会員の皆様の NPO 法人への移行手続きなどに関
9
する重要な議題があります。会員の皆様におかれましては、ぜひご参加いただきますよ
うお願いいたします。
Banquet 懇親会
The banquet will be held during 18:30-20:30 on 11th (Sat) at the plaza of University
Hall (1st floor). Prior registration is required for attendance. We cannot accept
additional participants in principle; however, if you are eager to attend, please consult
with the reception staff on arrival. Our staff will be checking your name tag for the
“paid” stamp at the entrance, so please make sure to bring it. We cannot allow anyone
to join the banquet unless you have the stamp on your name tag.
11 日(土)18:30-20:30 に、大学会館 1 階プラザで行ないます。参加者は事前に申込み
をされた方とさせていただきます。当日参加は基本的にお受けできませんが、希望され
る方は受付にてご相談ください。懇親会会場入り口にて、 大会スタッフが名札の懇親会
参加印の確認をさせていただきますので、名札を必ず持参ください。名札の参加印がな
い場合は、ご入場いただけません。
Smoking キャンパス内での喫煙
Smoking is prohibited on all campus property at the University of Tsukuba. Thank you
for your understanding and cooperation.
大学構内は、全面禁煙です。ご協力をお願い致します。
ATM/post office ATM・郵便局
We have a post office and ATM in the Annex University Hall, which is located at the
east side of the University Hall, near the foot of the TOYO bridge (桐葉橋).
大学会館別館(大学会館の東側の建物)の桐葉橋わきに、郵便局と常陽銀行の ATM が
ありますのでご利用ください。また、10 日(金)17 時までは、大学会館別館 1 階の筑波
大学内郵便局をご利用いただけます。
Parking 駐車場
TAIGEI-NISHI parking lot will be available throughout the meeting period. The visitor
parking of the University Hall will also be available. The car-stop fence gate will be
unlocked, so you can directly drive into the parking lot from the Hiratsuka line.
「大学会館 外来用駐車場」(平塚線沿いから入ることができます)、または「体芸西駐
車場」(大学会館バス停の南へ約 150m に入口ゲートがあります)を無料でご利用いただ
けます。守衛所等での手続きは必要ありません。
Lunch 昼食
Many of the eating facilities within campus will be open during 10th (Fri). Part of them
will still be open on 11th (Sat), but few on 12th (Sun). Please bring your own lunch or
try off-campus eateries in the neighborhood.
10
Participants can also make reservations for 700-JPN lunch boxes (incl. tea) prior to the
meeting through online reservation form
(http://www.pe.ska.life.tsukuba.ac.jp/PES30lb); deadline: September 19th, Fri.
The payment should be made at the reception desk on arrival.
学内の食堂は、10 日(金)は多数営業しています。11 日(土)も一部営業しています
が、12 日(日)はほとんど営業していません。会場周辺の飲食店をご利用いただくか、
ご自身でご用意ください。
また、弁当 (お茶付き) を 700 円で予約限定販売します。 希望する方は、お弁当申込
みフォーム(http://www.pe.ska.life.tsukuba.ac.jp/PES30lb)から、9 月 19 日
(金)までに申込みをしてください。弁当代金は、学会受付にてお支払いください。
Misccellaneous その他
The University Hall has wireless LAN Internet access throughout the building for those
who have their own computer. To access the internet, you will need 1) access point’s
password and 2) login name & password to the network authentication. They will be
provided on request at the reception desk.
For this meeting, we will not have the program in printed format. All information will
be posted on our PES30 website. Please keep your eyes on it!
当日、会場では無線 LAN を使用できます。接続するには、1: アクセスポイントへのパ
スワード、2: 認証ネットワークのログイン名・パスワード の 2 ステップが必要です。ご
希望の方は、受付にてお申し出ください。
本大会では、冊子体でのプログラム・要旨集の配布をいたしません。大会に関わる情
報はすべて、大会 HP へ掲載しますので、皆様には HP を閲覧くださいますよう、お願い
いたします。
11
Population Ecology Young Scientist Award: Award
Ceremony and Award Lecture
個体群生態学会奨励賞 授賞式・受賞講演
Saturday, 11 October 16:00-16:40 Hall (3F)
Language: English
Mathematical models for effective management of populations subject to
uncertainty
Hiroyuki Yokomizo (Center for Environmental Risk Research, National Institute for
Environmental Studies)
We need to mitigate various anthropogenic threats to species as efficiently as possible
because both time and budgets available for management are limited. However, finding
optimal solutions is complicated by the uncertainty caused by both stochasticity and
imperfect knowledge of targeted populations. One important question is how much we
should reduce those uncertainties by monitoring, which also incurs a cost. Different
degrees of uncertainty need to be modeled differently. First, I derive optimal
conservation and monitoring effort levels under uncertainty on population size of an
endangered population. Second, I demonstrate the importance of knowing the
relationships between density of invasive plants and its cost of impact for effective weed
managements. Third, I propose a mathematical model to spatially allocate conservation
efforts based on imperfect knowledge of the rate of change in population size for
conservation of Japanese endangered vascular plants. Mathematical models enable us to
derive rational and objective management strategies. We can show assumptions clearly
in the models, which may contribute to consensus building in decision-making. I show
how mathematical models can give us useful insights and provide tools for effective
management of populations.
12
Autumn School ‘Clinic on Statistics’
秋の学校「行列のできる統計相談所」
Friday, 10 October 15:00-18:00 Multimedia room (2F)
Language: English and Japanese
Consultants are Dr. Eiichi Kasuya (Kyushyu Univ.) and Dr. Nobuhiro Minaka
(NIAES). Anyone attending to PES30 can visit the clinic with his/her tough statistical
problems and ask the experts for solutions. Discussion on the problem will start and
may end with some fruitful advices or resignation.
相談員として、粕谷英一氏(九州大学)と三中信宏氏(農業環境技術研究所)がご参
加くださいます。PES30 に参加する人は誰でも、自分の抱える統計学的難問を持ち込み
相談できます。相談はおそらく議論を呼び、最終的に有意義な解法か諦めの、どちらか
を持ち帰ることになります。
How to consult 相談の手順
Your registration for your statistical problems through the PES30 website is
recommended, but anyone can ask the experts for their advice without submitting
registration beforehand.
At the day of clinic on statistics, please give us a brief presentation concerning your
consultation. Please prepare a few slides (Keynote, PowerPoint or pdf) for the
presentation. The order of presentation is determined until the day of clinic.
You can resister two or more consultations, but the second and following
consultations are allowed only when we have enough time. Please note that we cannot
complete all consultation due to the limitation of time. 相談は大会 HP を通じて随時受け付けております。当日飛び入り参加も可能です。
当日は、相談者の方に、相談内容を説明する簡単なスライドを説明していただきます。
議論の共有をはかるため、可能であれば、データや対象の図などの表示をお願いします。
相談者の方の希望によっては、用意していただいたスライドを、実行委員が代理で紹介
することも可能です。それをもとに、統計の達人方に解法の指南を乞い、会場の皆様と
ともに、議論を深めます。
なお、質問は複数でもかまいませんが、なるべく多くの皆様の相談が可能となるよう、
2 つ目以降のご質問は時間が余った場合にのみ対応する、ということにさせてください。
また、もし応募者多数の場合は、時間内に相談ができなくなる可能性がありますが、ど
うかご了承くださいますようお願い申し上げます。
13
Prenary Symposia 基調シンポジウム
Prenary Symposium A:
[SpA] Statistics and Population Ecology
Saturday, 11 October 9:00-12:00 Hall (3F)
Language: English
Organizer: Yukihiko Toquenaga (Univ. of Tsukuba)
E-mail: toque[at]biol.tsukuba.ac.jp
The following three speakers will discuss population ecology from three statistical perspectives:
error statistic, evidential, and Bayesian. The names of the three camps are based on “The
nature of scientific evidence” (2004) eds. M.L. Taper and S.R. Lele. Error statisticians and
evidentialists are often (mis-) labelled as frequentists and information statisticians,
respectively. After a briefly introduction of the three statistical perspectives by the organizer,
the three speakers will give us talks with the following titles;
[SpA-1] Evidential statistics: a path between the paradigms
Mark L. Taper (Department of Ecology, Montana State University / Department of Biology,
University of Florida, USA)
E-mail: markltaper[at]gmail.com
For two and a half centuries scientists wishing to make statistical inferences have been faced
with a choice between two very different paradigms: Error statistics (often called classical
statistics or frequentism) and Bayesian statistics. Evidential statistics has arisen as a response
to this tension, and has been constructed, more or less consciously, from both paradigms by
appropriating good features and jettisoning problematic features. In evidential statistics, the
evidence for one model over another is a data based estimate of the relative distance of each
model to truth. There is a broad family of evidence functions including the familiar ones such
as the likelihood ratio and the difference of information criteria. In this talk, I will define the
design characteristics of evidential statistics more closely. I will also discuss a number of axes
along which error and Bayesian statistics are commonly thought to be differentiated and how
evidential statistics relates to each. I will then indicate how several difficulties with the two
paradigms are easily addressed using the evidential approach. Several examples of the use of
the evidential approach in scientific analyses will be briefly introduce. I will close by discussing
my personal views of the central problem facing scientists and how evidential statistics is well
suited to solve that problem.
[SpA-2] Bayes approach as an approximation to maximum likelihood
estimation
Kohji Yamamura (National Institute for Agro-Environmental Sciences)
E-mail: yamamura[at]niaes.affrc.go.jp
14
Ronald A. Fisher (1922), who is the founder of maximum likelihood estimation, criticized the
Bayes approach (1763) of using a uniform prior distribution, simply because we can invent
estimates arbitrarily in the Bayes approach by changing the transformation before the
analysis. Thus, the Bayes approach lacks the scientific objectivity, especially when the amount
of data is small. However, we can utilize the Bayes approach as an approximation to the
objective maximum likelihood estimation if we use an appropriate transformation that makes
the posterior distribution close to a normal distribution (Yamamura et al. 2008). The Bayes
approach generally makes the calculation of maximum likelihood estimates very easy because
no complicated optimization technique is required. We illustrate it by the example of
maximum likelihood estimation of sika deer populations. We show that we can create various
estimates in the Bayes approach by changing the transformation arbitrarily, but that only one
transformation is appropriate as an approximation to the maximum likelihood estimation.
One-to-one correspondence exists between a uniform prior distribution under a transformed
scale and a non-uniform prior distribution under the untransformed scale. For this reason, the
Bayes approach of maximum likelihood estimation is nearly identical to the Bayes estimation
using non-informative prior of Jeffreys (1946).
[SpA-3] Bayesian data analysis in ecology: motivations, methods, and benefits
Robert M. Dorazio (Southeast Ecological Science Center, U.S. Geological Survey, USA)
E-mail: bdorazio[at]usgs.gov
The goal of this presentation is to provide guidance to ecologists who would like to decide
whether Bayesian methods can be used to improve their conclusions and predictions. I begin
by providing a concise summary of Bayesian methods of analysis, including a comparison of
differences between Bayesian and frequentist approaches to inference when using hierarchi- cal
models. Next I provide a list of problems or circumstances where Bayesian methods of analysis
may arguably be preferred over frequentist methods. These problems and circum- stances are
usually encountered in analyses based on hierarchical models of data. I briefly describe the
essentials required for applying modern methods of Bayesian computation (i.e., MCMC), and I
use real-world examples to illustrate these methods. I conclude by summa- rizing what I
perceive to be the main strengths and weaknesses of using Bayesian methods to solve
ecological inference problems.
15
Plenary Symposium B:
[SpB] Scaling from Life History Traits to the Population Dynamics
Sunday, 12 October 9:00-12:00 Hall (3F)
Language: English
Organizers: William A. Nelson (Queen’s University, Canada), Takehiko Yamanaka (National
Institute for Agro-Environmental Sciences)
From single-celled algae to vertebrate carnivores, populations can have pronounced
stage-structure owing to the life-history of organisms. Juveniles, for example, often have lower
ingestion rates than adults and less per-capita impact on resources. When vital rates are
resource-dependent, this creates an interaction between life-histories and resource densities
that can generate unexpected ecological dynamics. These individual stage-structues result in a
rich variety of population dynamics and furthermore they shape unique community dynamics.
The following talks showcase recent examples where accounting for life-histories help
disentangle ecological dynamics.
[SpB-1] Recurrent insect outbreaks caused by temperature-driven changes in
system stability
William A. Nelson (Queen’s University, Canada)
E-mail: nelsonw[at]queensu.ca
Insects often undergo regular outbreaks in population density that can have devastating
impacts on agricultural crops. For some pests, identifying the causal mechanism for outbreaks
has proven difficult. Here we show that outbreak cycles in the tea tortrix Adoxophyes honmai
can be explained by temperature driven changes in system stability. Wavelet analysis of a 51yr
time series spanning over 200 outbreaks reveals a distinct threshold in outbreak amplitude
each spring when temperature exceeds 15◦ C, and a secession of outbreaks each fall as
temperature decreases. To better understand temperature-dependence in these outbreaks, we
developed a mathematical model of insect population dynamics that explicitly incorporates the
temperature-dependent response of A. honmai life history as observed in the laboratory. The
independently parameterized model predicts that insect populations will cross a Hopf
bifurcation from stability to sustained cycles as temperature increases, which is in close
quantitative agreement with the observed outbreaks. Our results suggest that temperature can
alter system stability and provide an explanation for generation cycles in multivoltine insects.
[SpB-2] Multi-generation population dynamics in seasonal environments:
generation separation versus smearing
Ottar N. Bjornstad (Pennsylvania State University, USA)
E-mail: onb1[at]psu.edu
Many insects have multiple generations each year. They are found across biomes that vary in
their strength of seasonality and, depending on location and species, can display a wide range
of population dynamics. Some species exhibit cycles with distinct generations (generation
16
separation), some exhibit generation overlap with multiple life stages present simultaneously
(generation smearing), while others have intermediate dynamics with early season separation
followed by late season smearing. There are two main hypotheses to explain these dynamics.
The first is the ’seasonal disturbanc’ hypothesis where winter synchronizes the developmental
clock among individuals, which causes transient generation separation early in the season that
erodes through the summer. The second is the ’temperature destabilization’ hypothesis where
warm temperatures during the summer cause population dynamics to become unstable giving
rise to single generation cycles. Both hypotheses are supported by detailed mathematical
theory incorporating mechanisms that are likely to drive dynamics in nature. I will discuss the
theory and propose a conceptual framework that allows for an intuitive description of the
relative importance of each mechanism.
[SpB-3] Moth outbreak dynamics across Japan suggest evolution at the
northern range limit
∗
Takehiko Yamanaka (National Institute for Agro-Environmental Sciences), Yasushi Sato
(National Institute of Vegetable and Tea Science)
E-mail: apple[at]affrc.go.jp
We analyzed the long-term data of the Japanese smaller tea tortrix, Adoxphyes honmai at
several stations from Saitama to Kagoshima. We found that populations at all the stations
showed strong evidence of generation cycles, a landmark of internal population regulation.
That is, larvae compete for food resources and the reproductive period is shortened. This
internal competition mechanism has been shown to create regular generation cycles, a
phenomenon that was observed in the data that we analyzed. We also evaluated the effect of
temperature increase over several decades on the generation cycles at each station. The
northern populations, such as in Saitama, were found to prolong their generation periods to
adjust to the onset of winter. Tracking the change in growth speed of the individuals in the
northern populations based on the temperature increase in these decades would provide the
evidence for microevolution. We suspect that the harshness of the winter drastically
accelerates local adaptation to the environment.
[SpB-4] Timing and propagule size of invasion determine its success by a
time-varying threshold of demographic regime shift
Masato Yamamichi (Program-Specific Assistant Professor of Hakubi Center for Advanced
Research / Center for Ecological Research, Kyoto University)
E-mail: yamamichi[at]ecology.kyoto-u.ac.jp
Theory of invasion ecology indicates that the number of invading individuals (propagule size)
and the timing of invasion are important for invasion success. Propagule size affects
establishment success due to an Allee effect and the effect of demographic stochasticity,
whereas the timing of invasion does so via niche opportunity produced by fluctuating
predation pressure and resource abundance. We propose a synthesis of these two mechanisms
by a time-varying dose-response curve where the dose is propagule size and the response is
establishment probability. We show an example of the synthesis in a simple predator-prey
model where successful invasion occurs as a demographic regime shift because of the bistability
17
of the system. The two mechanisms are not independent, but simultaneously determine
invasion success in our model. We suggest the difficulty of understanding invasion process
based on a dose-response curve of propagule size as no unique curve can be determined due to
the effects of invasion timing (i.e., the threshold of demographic regime shift is time varied).
The results of our model analysis also have an implication on eco-evolutionary feedbacks and
the phase relationship between population cycles of predators and prey.
18
Organized Sessions 企画シンポジウム
Organized Session 1:
[So1] 相互作用系の多面的理解から明らかにする個体群生態学
Comprehensive Understanding of Population Ecology by
Multidisciplinary System Research of the West Indian Sweet Potato
Weevil
Saturday, 11 October 14:00-16:00 Hall (3F)
Organizer: 熊野了州 1,2,3 (1 沖縄県病害虫防除技術センター、2 琉球大学、3 琉球産経株式
会社)
Norikuni Kumano1,2,3 (1 Okinawa Prefectural Plant Protection Center, 2 Ryukyu-Sankei Co.,
Ltd., 3 University of the Ryukyus)
E-mail: nrkumano[at]gmail.com
自然界には競争、捕食、寄生、共生といった様々なメカニズムの生物間相互作用が存在している。
個々の相互作用を見てみると、作用するレベルやスケールは個体間から宿主と体内微生物の関係
に至るまで、極めて多様である。ある一種を中心に据え、これらの相互作用を俯瞰すると、レベ
ルやスケールの異なる相互作用同士の関連性が見えてくる可能性がある。こうした相互作用系の
解明は、群集生態学や生物多様性の理解を進めると同時に、害虫管理においてはは天敵昆虫の利
用で直接的な利益をもたらす事が可能なため極めて有用になる。本シンポジウムでは近年、野外
と増殖環境で興味深い種間相互作用が明らかになりつつある、熱帯∼亜熱帯に広く分布するサツ
マイモ害虫であるイモゾウムシの種間相互作用に焦点を当てる。そして、本種と競争、捕食-被捕
食、寄生、相利関係にある、4つの異なるレベルで作用する種間相互作用系(競争者、寄生蜂、
病原虫、微生物)の話題を演者の方々に紹介していただき、対象とする生物(イモゾウムシ)の
相互作用系の網羅的理解から個体群生態学の新たな視点を考えたい。
In nature, a wide variety of biological interactions are observed, such as competition,
predation, parasitism, or mutualism. Some biological interactions are conducted in the body or
the cell. If we study the variety of biological interactions with a focus on a specific species, we
may able to find some additional information. This viewpoint allows us to cultivate a better
understanding of community ecology and biodiversity. Furthermore, we may be able to
develop techniques to control pest insect populations. In this symposium, we focus on the
biology of the West Indian sweet potato weevil (Euscepes postfasciatus), a major pest insect
species for sweet potato in tropical and subtropical regions. This species is now being
intensively studied, and some curious biological interactions have been found in the laboratory
and field. In this session, four speakers would elucidate four different levels of biological
interactions for this weevil, such as resource competition, host-parasitic interaction (parasitic
wasp), host-parasite interaction (protozoan pathogen), and mutualistic interaction. We will
also discuss a new perspective for population ecology through a comprehensive understanding
of the biological interactions of a specific species.
[So1-1] 同一資源を利用するイモゾウムシとアリモドキゾウムシの種間
競争
19
Inter-specific competition between two sweetpotato weevils
栗和田 隆 (鹿児島大・教育・生物)
Takashi Kuriwada (Kagoshima University)
E-mail: t.kuriwada[at]gmail.com
イモゾウムシとアリモドキゾウムシは両種ともにサツマイモを寄主としてその内部で幼虫期を過
ごし、羽化後イモから脱出してきた成虫が新たな寄主を求めて分散していく。両種は時間的にも
空間的にも同所的に生息しており餌資源も共有しているため、強い種間競争が生じていると予測
される。しかし、両種の種間関係については全く調べられてこなかった。そこで、様々な密度・
割合で両種を飼育し、次世代の羽化個体数や体サイズ、発育期間を調査した (Kuriwada et al.
2013)。その結果、イモゾウムシはアリモドキゾウムシと比べ種間競争によってより強い負の影響
を受けていることがわかった。一方で、イモゾウムシの方が同種の高密度条件に対してはかなり
強い耐性を持つことも明らかになった。講演では両種の生態の違いからこの結果を考察する。
Kuriwada et al. (2013) Journal of Applied Entomology. 137: 310-316
[So1-2] イモゾウムシの対寄生蜂戦略: 植食性ジェネラリストにおける天
敵不在食草の選好性
Defense against parasitic wasps for enemy-free plants in the West Indian
sweet potato weevil Euscepes postfasciatus
◦ 鶴井香織 1,2,3 、熊野了州 1,2,3 、豊里哲也 1,2 、松山隆志 1 、立田晴記 3
(1 沖縄県病害虫防除技術センター、2 琉球産経株式会社、3 琉球大・農)
∗
Kaori Tsurui1,2,3 , Norikuni Kumano1,2,3 , Tetsuya Toyozato1,2 , Takashi Matsuyama1 , Haruki
Tatsuta3
(1 Okinawa Prefectural Plant Protection Center, 2 Ryukyu-Sankei Co., Ltd., 3 University of the
Ryukyus)
E-mail: tsuruikaori[at]gmail.com
ヒルガオ科を広く食害するイモゾウムシは卵・幼虫・蛹を茎内で過ごす。茎内は捕食者と物理的に
隔離され比較的安全だが、外部単寄生蜂イリムサーキバラコマユバチが幼虫期の天敵として知ら
れている。寄生蜂は野外で局所的・一時的に高密度となり、イモゾウムシ個体群に壊滅的な打撃を
与える一方で、寄生が全く見られないパッチも多くある。食草と寄生蜂密度に関する調査の結果、
enemy-free-space を形成するグンバイヒルガオは、幼虫と成虫にとって栄養的に劣る餌であった
一方、栄養的に優れたノアサガオにおける寄生蜂密度は高かった。母親はグンバイヒルガオへ好
んで産卵したことから、イモゾウムシの食草選択において、栄養的な質よりも子の幼虫期におけ
る死亡リスクが重要だと考えられた。近年の沖縄県久米島において、グンバイヒルガオでのイモ
ゾウムシ密度はノアサガオの約 10 倍に達している。しかし、食草間に見られるイモゾウムシ密度
の違いは、イモゾウムシ・寄生蜂・寄主植物の3者の拮抗的共進化により変動するかもしれない。
[So1-3] イモゾウムシ大量増殖における病原性原虫(Farinocystis sp.)の
発生
Occurrence of Farinocystis sp. on the mass production of West Indian Sweet
potatoweevil
◦ 大石 毅 1 、照屋清仁 1 、鶴井香織 1,2,3
(1 沖縄県病害虫防除技術センター、2 琉球産経株式会社、3 琉球大・農)
∗
Tsuyoshi Oishi1 , Kiyohito Teruya1 , Kaori Tsurui1,2,3
20
(1 Okinawa Prefectural Plant Protection Center, 2 Ryukyu-Sankei Co., Ltd., 3 University of the
Ryukyus)
E-mail: ooishits[at]pref.okinawa.lg.jp
沖縄県ではイモゾウムシ(以下 イモゾウ)根絶事業の一環として、人工飼料飼育と生イモ塊根
飼育を組み合わせた大量増殖による不妊虫の生産を行っている。これまで、イモゾウに感染する
病原性原虫(Farinocystis sp.)は、生イモ塊根飼育の成虫で蔓延することが知られていたが、人
工飼料飼育における感染は知られていなかった。しかし、2013 年 6 月にはじめて、人工飼料飼育
において成虫から原虫が検出された。原虫によるイモゾウの生産低下が懸念されたため、人工飼
料飼育および生イモ塊根飼育におけるイモゾウの原虫の感染状況と生産への影響について調査し
た。人工飼料飼育においては、成虫だけでなく幼虫、蛹においても感染が確認され、卵(糞栓)
も原虫に汚染されていた。それにもかかわらず、人工飼料飼育において、原虫感染が生産に及ぼ
す影響はほとんど確認されなかった。一方、生イモ塊根飼育においては、原虫感染は成虫に限ら
れているにもかかわらず、生産頭数は著しく低下した。これらの結果をもとに、餌の種類が原虫
感染したイモゾウの適応度に及ぼす影響の可能性について議論する。
[So1-4] イモゾウムシの内部共生細菌が宿主に与える影響
Effects of bacterial endosymbionts on a sweet potato weevil host
細川貴弘 (九大・理・生物)
Takahiro Hosokawa (Kyushu University)
E-mail: th-hosokawa[at]aist.go.jp
多くの昆虫類において体内共生微生物の存在が知られており、宿主昆虫の正常な成長や繁殖に共
生微生物が必須となっている。演者らはイモゾウムシの共生微生物について調べたところ、中腸
の前端部に菌細胞塊が発達しており、その内部に共生細菌が存在していることを発見した。分子
系統解析の結果、この共生細菌は一部のゾウムシ類で報告されている共生細菌 Nardonella の一
種であることが明らかとなった(Hosokawa et al. 2010)。抗生物質の投与によってイモゾウムシ
の幼虫から共生細菌を除去すると、生存率には異常が見られなかったが、羽化時の外骨格の黒化・
硬化がともに弱くなった。したがって共生細菌はゾウムシ類の堅牢な外骨格形成に重要な役割を
担っていることが示唆された(Kuriwada et al. 2010)。この共生細菌がイモゾウムシの食草選択
や寄生蜂抵抗性に影響を与える可能性についても議論する。
Hosokawa et al. (2010) Appl. Entomol. Zool. 45: 115-120.
Kuriwada et al. (2010) PLoS One 5: e13101.
Commentator: 鈴木紀之(立正大学・地球環境科学部・環境システム学科)
Noriyuki Suzuki (Rissho University)
21
Organized Session 2:
[So2] To save our dining tables with a variety of foods:
popuelation ecology of crop pollination and pest control
バラエティに富んだ食卓を守るために∼農業をめぐる送粉者と害虫防除の
個体群生態学∼
Sunday, 12 October 14:00-16:00 Hall (3F)
Language: Japanese with English slides
Organizers: ∗ Tomoyuki Yokoi (Univ. of Tsukuba / Pollination Services Society of Japan),
Kazuharu Ohashi (Univ. of Tsukuba)
◦ 横井智之(筑波大学生命環境系/日本送粉サービス研究会)
・大橋一晴(筑波大学生命環
境系)
E-mail: tomoyoko[at]envr.tsukuba.ac.jp
“Washoku”, traditional Japanese dietary culture, was added to UNESCO’s Intangible Cultural
Heritage list in 2013. One of the reasons for listing was “the consumption of various natural,
locally sourced ingredients such as rice, fish, vegetables and edible wild plants”. Various foods
not only sustain our dietary culture, but also play an important role in the maintenance of our
good health through balanced nutrients. In the front line of agricultural production, beneficial
organisms such as pollinators for crop pollination and natural enemies for biological pest
control contribute to the labor-saving and stable crop production. Efficient and sustainable use
of these organisms is essential for our diet. However, these organisms are facing/causing
problems in agroecosystem; intensive use of agrochemicals, invasion of alien species, and
biodiversity loss in agroecosystem. Both basic and applied research approaches based on
population ecology are necessary to solve these problems. In this symposium, we invite
researchers working on crop pollination, biological pest control, and the effect of agrochemicals
on biodiversity in paddy fields. We hope to deepen our discussions.
2013 年に「和食」がユネスコ無形文化遺産に登録された。登録の最大の理由は「多様で新鮮な食
材とその持ち味の尊重」と言われている。多様な食材は私たちの文化を支えるだけでなく、バラ
ンスの良い栄養摂取を通じた健康維持にも大きな役割を果たしている。食材の生産のなかで最も
産業規模の大きな農作物生産の現場では、授粉を行う送粉者(ハナバチ)や害虫の生物的防除を
行う天敵といった「生物資材」と呼ばれる生物種が、農作業の省力化と安定した作物生産に貢献
している。これら生物資材の効率的・持続的な利用によって、私たちの食生活は維持されている。
しかし近年、生物多様性の減少や外来生物の侵入、農薬利用に関わる問題が取りざたされている。
これからも私たちの食生活を守っていくためには、農業の現場で利用される生き物を対象に、個
体群生態学による基礎と応用の両側面からの研究が不可欠である。企画者らによる日本送粉サー
ビス研究会も、機関・分野を超えて新たな研究課題に取り組む場と位置づけている。本シンポジ
ウムでは、農業をめぐる送粉者、天敵による生物的防除等を研究されている研究者の方をお招き
してお話を伺い、じっくりと議論を行いたい。
[So2-1] Two serious problems in agroecosystems; invasive alien species and
chemical pesticides
Atsushi Kasai (NIES)
22
笠井 敦 (国環研)
E-mail: kasai.atsushi[at]nies.go.jp
Farmlands play important roles in human society. While the maintenance of crop quality and
the sustainability of yields are the first principles in agricultural production, the environment
of farmlands is a system that can sustain by depending on the biodiversity and endemism.
There are often trade-offs between agricultural production and agricultural environment
because farmlands themselves are the environments that enhance biodiversity and endemism.
Especially, disturbances such as chemical pesticides used for convenience and invasion of alien
species due to globalization may reduce the sustainability of agroecosystems. Here I introduce
Camellia spiny whitefly (Aleurocanthus camelliae) that invaded Japan in 2004 and its
parasitoid wasp, Encarsia smithi, as an example that alien species invading farmlands are not
only agricultural pests but also their natural enemies. I also introduce the efforts of evaluating
the impacts of three neonicotinoid insecticides on biodiversity in Japan. These insecticides are
widely used as nursery-box-applied insecticides for rice production in this country, whereas EU
banned their use from December 2013 due to concerns about their risks to bees. I hope to
provide an opportunity to discuss what environment of farmlands we should expect by
reviewing the details of the disturbances observed in these case studies.
[So2-2] Biological control: use of natural enemies that serve as agents of
agro-ecosystem service
生物的防除:農地生態系サービスのエージェントである天敵生物の活用
Takatoshi Ueno (Kyushu University)
上野高敏 (九大)
E-mail: ueno[at]grt.kyushu-u.ac.jp
Any organism suffers from biological and non-biological factors that cause its mortality, which
results in the reduction of its population. The process, i.e., natural control, can be used to
suppress agricultural pests. Natural enemies are organisms that cause the biological mortality
of other organisms and can be beneficial to us if they are enemies of target pests. Indeed,
biological control is the use of natural enemies, such as predators and parasitoids, to combat
pests and has been often successful in pest management. In my talk, I will give successful
examples of biological control against invasive insect pests, showing how biological control can
be an effective approach to manage invasive alien organisms and to attain sustainable or
eco-friendly agriculture. Also, evidence is given that natural enemies are ideal as “biological
indicators” to assess agro-biodiversity and environmental soundness of agricultural practices in
rice paddies. I will show that we can link among farmers, consumers, sustainable rice
production, and biodiversity, by using natural enemies as bio-indicators in rice paddies. Thus,
my talk will address usefulness of natural enemies, agents of ecosystem service, in agriculture.
[So2-3] The importance of pollination service to crop production in Japan
Akihiro Konuma (NIAES)
小沼明弘 (農環研)
E-mail: akihirok[at]niaes.affrc.go.jp
23
Pollination by animal vector has attracted many scientists as main target of their study for
long time. Many of those studies have focused on relationship between wild plants and their
pollinators in evolutionary/conservation context because animal pollinators are significant
evolutionary factors of flowering plants. Recently, world pollinator declining comes into
spotlight together with sudden disappearing of honeybee such as CCD (Colony Collapse
Disorder). Wild insects, apparently, contribute to productivity of fruits and fruit vegetables
around the world. For example, TEEB (The Economics of Ecosystems and Biodiversity)
reported the total economic value of insect pollination was estimated at 153 billion Euros in
2005. This is 9.5% of world agricultural output for human food. Because of the importance of
pollination service to crop production, Intergovernmental Platform on Biodiversity and
Ecosystem Services (IPBES) decided to set the “ Pollination and food production ” as a work
program in fast track thematic assessment. However, knowledge regarding pollination by wild
insects as an ecosystem service, especially in crop production, is still limited.In this talk, I may
discuss the importance of pollination ecology in crop production, study of insect pollination as
an ecosystem service to agriculture and food production in Japan.
Commentator: Jun Nakamura (Tamagawa Univ.)
中村 純(玉川大)
24
Organized Session 3:
[So3] Rice, Bugs, and Egrets
田んぼ、蟲、鷺
Saturday, 11 October 14:00-16:00 Multimedia room (2F)
Language: English
Organizer: Miyuki Mashiko (NIAES)
益子美由希(農業環境技術研究所)
The venue of the PES30 is located in the center of the Tsukuba Science City in Ibaraki
Prefecture, which is a planned city developed in the 1960s. The city has more than 300
research institutes that are surrounded by rice and lotus fields supporting the largest breeding
populations of colonial Ardeidae species in Japan. Herons and egrets often forage in rice fields,
and Japanese people have traditionally worshiped them as gods protecting crops from pest
bugs and locusts. But the population sizes of herons and egrets in Japan had declined after
the World War II, and now their breeding colonies are often hated by neighbors because of
their feces and noisy sounds. To build a future in which we live in harmony with animals living
in a human-influenced landscape, we can learn more from studies on population ecology. This
session will bring together researchers that study different aspects of colonial herons and egrets
such as long-term population dynamics, mechanisms of colony site selection, and ecological
functions of insects living in the ground of egret colonies. The statistical methods we introduce
would also come up for discussion and can be applied in various practice in population ecology.
本大会の会場・筑波大は、研究学園都市という、人的に計画された土地にあります。しかし一歩
外は、田舎の田園風景。水田面積国内 2 位の茨城県は、準絶滅危惧種のチュウサギも多い、サギ
たちの楽園です。サギ類は、人が創出した水田環境を主な餌場とする高次捕食者で、日本では古
くから、害虫を食べる益鳥として祀られてきました。しかし、戦後の開発期には個体数を減らし、
近縁の複数種・数十∼数千羽からなる彼らのコロニー(集団繁殖地)は、時に悪臭でけむたがら
れています。私たちのそばに生きる彼らとうまく付き合っていくためにも、野外データによる個
体群生態学から、ヒントが得られるはずです。本企画では、茨城県とイタリア北部の 2 地域での
長期研究を比較しながら、サギ類の個体数動態に関わる要因を探ります。また、毎年同所にコロ
ニーを形成したがるサギ類の習性と、周辺の土地利用情報とを天秤にかけた、コロニー場所選択
の解析から、群れの意思決定メカニズムを議論します。さらに、サギ類の羽毛や死骸がコロニー
の林床に生息する昆虫によって食物連鎖へ組み込まれる様を垣間みます。各演者の用いたユニー
クな統計手法も、議論を呼ぶことを期待しています。
[So3-1] Colony dynamics of heron and egret communities in Japan:
ecological and historical perspectives
日本におけるサギ類群集の繁殖コロニーの動態についての生態学的・歴史的研究
∗
Miyuki Mashiko (NIAES), Yukihiko Toquenaga (Univ. of Tsukuba)
◦ 益子美由希(農業環境技術研究所)、徳永幸彦(筑波大)
E-mail: mashiko3yuki[at]affrc.go.jp
For colonial breeding birds, the presence and persistence of suitable colony sites are important
factors for the maintenance of bird populations. Long-term studies of colonial herons and
egrets are well conducted in Europe and the USA, but not in Asia. Here we investigated
25
colony dynamics based on 51-years data around Ibaraki Prefecture, Japan. First, 10-year
population dynamics from 2002 to 2011 were addressed. The population dynamics differed
among the six constituent species, and the marked increase of the Ardea cinerea contributed to
the increasing temporal variation in colony size and in species composition ratio. Second, we
examined whether colonies exhibit site fidelity by applying the correlated random walk
method. Two-third of 61 colonies during 1963-2013 exhibited site fidelity despite egrets being
facultative reusers of previously occupied colony locations. Collective philopatry may originate
from a few faithful pioneer individuals with many others following them. Third,
pioneer/follower relationship among the constituent species of colonies were analyzed by
considering A. cinerea as a pioneer. After the increase of A. cinerea in 2005, colonies with A.
cinerea had a greater propensity to persist. A. cinerea began to take the initiative in
establishing colonies, and other species began to follow them in colony site selection.
[So3-2] Long-term trends of breeding herons and egrets, and their foraging
ecology in ricefields of Italy
イタリアにおけるサギ類の長期動態と水田における採餌生態
Mauro Fasola (Pavia Univ., Italy)
E-mail: fasola[at]unipv.it
Breeding herons and egret were monitored since 1972 in Northwestern Italy, an area of 57,591
km2 with large surfaces of rice cultivation (2,000 km2 ). The heronries increased from 40 to
130, and the nests peaked in 2000, up to 23 times the initial number for Grey Herons. This
spectacular increase was due to lower human-induced mortality, to climatic changes, and to
changes in rice cultivation practice. But since 2000, a decreasing trend has become evident. In
order to check the influence of rice cultivation practices on population trends, we compared the
results obtained in 2013-2014 with those 1977-2000, about the following topics. 1) Changes in
chicks diet; some staple prey (amphibians) have diminished, while new prey of recent
colonization (the invasive Procambarus clarkii) have increased. 2) Changes in the submerged
rice, studied using satellite imagery; compared to 100% submersion until 1990, the submerged
surface in 2013 were <50%. 3) Foraging success of the breeders in agricultural versus
seminatural foraging habitats. A modeling is under way of the influence of these changes in
prey availability, of the climatic changes, and of other factors, on these declining heron
populations.
[So3-3] Arthropods and their ecological function in heron breeding colonies
サギ類コロニー下における節足動物の生態的機能
Shinji Sugiura (Kobe Univ.)
杉浦真治(神戸大)
E-mail: ssugiura[at]people.kobe-u.ac.jp
Herons frequently breed in inland forests. In the forest floor, organic materials are provided as
carcasses of prey (that they drop) and chicks (that die). Such resource subsidies of organic
materials potentially increase arthropod abundance and diversity in forests. Furthermore,
increased arthropods can enhance the decomposition of organic materials. Here I investigated
26
arthropod communities and their ecological function at heron colony sites in the southern area
of Ibaraki Prefecture, central Japan. I used pitfall and baited traps to compare arthropod
densities and diversity between heron breeding colonies and areas outside of colonies. I found
that scavenging arthropods such as carrion beetles and earwigs increased in heron breeding
colonies. Furthermore, I investigated the decomposition rate of heron carcasses in the colonies
to clarify the ecological function by scavenging arthropods in the heron colonies. I
experimentally placed heron feathers (a part of carcasses) in the forest floor and thereafter
investigated the weight decrease of the feathers. Four months after the feather placement,
86-99% and 21-68% of the feathers were consumed in colony and non-colony sites, respectively.
These results suggest that the population increase of scavenging arthropods could enhance the
rate of feather decomposition in heron breeding colonies.
[So3-4] Changes in colony site selection strategies of herons and egrets over
the years: new statistical methods for studying habitat selection
サギ類におけるコロニー場所選択の戦略の年次変化:生息場所選択を研究するための新
たな研究手法
∗
Luis Carrasco, Yukihiko Toquenaga (Univ. of Tsukuba)
E-mail: luis[at]pe.ska.life.tsukuba.ac.jp
Conventional habitat selection models using log-linear or more sophisticated regressions often
fail to predict distribution of species, due to the complexity of the data available or to their
lack of generalization power. Statistical tools that assume no linearity and capable to
extrapolate the predictive models across landscapes are needed to deal with the difficult
relationships between habitat variables and species distribution data. We created colony site
selection models for herons and egrets using a random forest algorithm: a classification
algorithm using multitude of decision trees as training time and outputting the mode of classes
output by each tree. To obtain the data used for constructing the models, we used land use
maps and long-term data of colony locations in Ibaraki, Japan. We also analyzed the collective
site fidelity by analyzing the distance from each colony site to previous year colonies ’
locations. This methodology allowed us to observe changes of the habitat selection strategy
over the 30 years of study. The most striking change was that colony site fidelity increased
drastically its importance in relation to the land use preferences in recent years. This could
reflect the changes in species composition of the colonies of the last few years.
Commentator: Yukihiko Toquenaga (Univ. of Tsukuba)
徳永幸彦(筑波大)
27
Poster Presentations 一般講演(ポスター)
Poster Titles 講演タイトル
Note that poster ids are not in serial, and hence there are missing numbers.
ポスターは連番ではなく、番号が飛んでいます
[P01] 佐渡市における環境と経済の好循環のための数理的研究:生態系動態とヒトの選択
動態の結合モデル – 横溝裕行(国環研)
[P02] アブラムシに化学擬態する捕食者のアリを騙すメカニズム – ○林正幸、野村昌史
(千葉大院・園芸・応用昆虫)
[P04] 少数個体からなる群れからみる群れの起源 – ○新里高行 (筑波大)、三具和希 (筑波
大)、村上久 (神戸大)、都丸武宣 (神戸大)、園田耕平 (滋賀大)、西山雄大 (大阪大)、
郡司ペギオ幸夫 (早稲田大)
[P05] Male-killer prevalence in a sympatric population of specialist and generalist
Harmonia ladybirds – *Suzuki Noriyuki (Tohoku Univ.), Naoya Osawa (Kyoto
Univ.)
[P06] The form of an evolutionary tradeoff affects eco-evolutionary dynamics in a
predator-prey system – *Minoru Kasada (Univ. of Tokyo), Masato Yamamichi
(Univ. of Kyoto), Yoshida Takehito (Univ. of Tokyo)
[P08] 鳥のさえずりの地理的変異と種の認知:方言に対する反応の非対称性 – 濱尾章二
(国立科学博物館・動物研究部)
[P10] 徒花の意味 – 江副日出夫(大阪府大・院・理)
[P12] 生態学的個体群制御と群集ネットワークの複雑性 – ○近藤倫生(龍谷大・理工)、
舞木昭彦(島根大・生物資源)
[P16] シロアリ女王の秘策:卵門を閉じて王と同居しながら単為生殖 – ○矢代敏久(京
大院・農・昆虫生態)、松浦健二(京大院・農・昆虫生態)
[P18] 生息環境の細分化の程度と種間競争の結果 – 穴澤正宏(東北工大・工・環境)
[P19] Accounting for the spatial structure of communities in the estimation of transition
probabilities subject to observation errors – Keiichi Fukaya (ISM), J. Andrew Royle
(USGS), Takehiro Okuda (NRIFSF), Masahiro Nakaoka, Takashi Noda (Hokkaido
Univ.)
[P20] 性的対立の波及効果:より発達した交尾器の棘をもつオスほど他種メスにコスト
を与える – ○京極大助、曽田貞滋(京都大・理・動物生態)
[P21] セイタカアワダチソウの侵入地とその起源集団におけるアワダチソウグンバイの
密度の比較 – ○坂田ゆず (京大・生態研), Timothy Craig (University of Minnesota,
Duluth), Joanne Itami (University of Minnesota, Duluth), 大串隆之 (京大・生態研)
28
[P22] Natural selection, genetic variation, and plasticity in eco-evolutionary dynamics:
a case study of Plantago lanceolata – Richard P. Shefferson (University of Tokyo),
Deborah A. Roach (University of Virginia)
[P26] キアゲハの季節多型(春型・夏型)の適応的意義:体温調節か保護色か? – ○正
木奈穂、西口泰平、石原道博 (大阪府大・院・理)
[P27] 小標本データに対するブートストラップ信頼区間の構成 – ○岡村 寛・市野川 桃子(中央水研)
[P28] 性の存在が群集動態に与える影響:ギルド内捕食系を用いた理論的検証 – 川津一隆
[P29] ダムによる魚の移動阻害の影響―メタ生態系モデルを用いたメコン川ダム開発の
影響評価― – ○吉田勝彦(国立環境研・生物)、広木幹也(国立環境研・生物)、 冨
岡典子(国立環境研・地域)、村田智吉(国立環境研・地域)、福島路生(国立環境
研・生物)
[P30] ハダニの交尾前ガード行動と捕食リスク:気にする♀、しない♂ – ○奥 圭子 1,2,3 ,
Erik H. Poelman2 , Peter W. de Jong2 , Marcel Dicke2 (1JSPS 海外学振, 2Wageningen
Univ., 3 中央農研)
[P31] Does group size influence aggressive behavior of Formica japonica in group fighting?
– *Suguru Sasaki and Gaku Takimoto (Toho university)
[P32] 生活史雑食による捕食−被食関係の逆転とギルド内捕食系の共存 – ○川田尚平、
瀧本岳(東邦大・生物)
[P33] 個体群成長率とプロセスの時空間変動性:岩礁潮間帯の固着生物群集における推
移行列モデルを用いた解析 – ○金森由妃(北大・院・環境科学),深谷肇一(統数
研),岩崎藍子(北大・院・環境科学),野田隆史(北大・地球環境)
[P36] 針葉樹林業地域における広葉樹林がカスミザクラの結実と交配に与える効果 – ○
永光輝義、滝久智、菊地賢、加藤珠理(森林総研)
[P38] A facultative mutualist increases the persistence of obligate mutualism – Gaku
Takimoto (Toho University)
[P39] 状態空間モデルによるアライグマの個体数推定と空間明示的な作物被害の許容密
度の推定 – 〇栗山武夫、長田 穣(東大・院・生物多様性)、浅田正彦(AMAC)、
宮下 直(東大・院・生物多様性)
[P40] 同所的に生息する二種の同属ダンゴムシに感染する繁殖操作の異なるボルバキア –
角拓人(岡山大)、澤谷祐輝(岡山大)、三浦一芸(NARO)、○宮竹貴久(岡山大)
[P41] Temporal chain reactions of indirect interactions initiated by intraspecific variation
in an early-season herbivore – Shunsuke Utsumi (FSC, Hokkaido Univ.)
[P42] 果実の摘果が生物的防除の効率に与える影響についての数理的考察 – ○池川雄亮、
江副日出夫、難波利幸 (大阪府大・院・理)
29
[P43] 遺伝子と個体数分布に基づいたサドガエル個体群のネットワーク構造の推定 – ○
山中美優 (東大・院・農), 小林頼太 (東環工専), 関谷國男 (新潟大), 宮下 直 (東大・
院・農)
[P44] A stranger is tastier than a neighbor: Cannibalism in Mediterranean and
desert populations of pit-building antlions – *Ofer Ovadia and Erez David Barkae
(Ben-Gurion University of the Negev)
[P46] サンショウウオの共食いがトップダウン効果に及ぼす影響は年変異する – 〇高津
邦夫(北大・院・環境科学)、岸田治(北大・北方圏 FSC)
[P47] Multinomial mixture model によるオニヒトデ捕獲除去データからの駆除達成率の
推定 – ⃝熊谷直喜(国環研)、山川英治・白木一太朗(沖縄県環境科学センター)、
比嘉義視(恩納村漁協)、岡地賢(コーラルクエスト)
[P48] 人為的撹乱に対する山菜の応答∼収穫することで翌年のタケノコの生産は増す –
⃝片山昇、岸田治、坂井励、早柏慎太郎、伊藤欣也、実吉智香子、浪花愛子、高橋
廣行、高木健太郎(北大・FSC)
[P49] 気候と景観構造の地域差がもたらす高次捕食者サシバの繁殖適地の違い – ○藤田
剛(東大・農)、東 淳樹(岩手大・農)、野中 純(オオタカ保護基金)、堺 義昭、堺 初美(印西サシバの会)、伊関文隆(希少生物研)、深澤圭太(国環研)、
宮下 直(東大・農)
[P50] 農法と周辺景観が水田のアシナガグモ‐餌昆虫の季節動態に及ぼす影響 – ⃝筒井
優(東大・農)、田中幸一(農環研)、馬場友希(農環研)、宮下直(東大・農)
[P51] 共生する植物-バクテリアでみられる野外集団間による変異 – ○鍵谷進乃介(北大・
環境科学院)、内海俊介(北大・北方生物圏 FSC)
[P52] ハムシの餌選好性はどの形質と遺伝的に相関するか – ○小野寺裕乃 (北大・環境科
学院)、内海俊介 (北大・北方生物圏 FSC)
[P54] 大きなオタマがいるとき、小さなオタマが捕食されやすいのはなぜか? – ○山口
彩(北大・院・環境科学), 岸田治(北大・北方生物圏 FSC)
[P55] 異質環境下におけるニホンジカの将来分布の予測:セルベースの状態空間モデル
による解析 – ○長田穣(東大・院・生物多様性)、浅田正彦(AMAC)、栗山武夫
(東大・院・生物多様性)、横溝裕行(国環研)、宮下直(東大・院・生物 多様性)
[P56] 寄主植物を共有するホソオチョウとジャコウアゲハの出現タイミングが両種間相
互作用に与える影響 – ○橋本洸哉、大串隆之(京大・生態研センター)
30
Poster Abstracts 講演要旨
[P01] 佐渡市における環境と経済の好循環のための数理的研究:生態系動態とヒトの選択動
態の結合モデル
横溝裕行(国環研)
水田生物の多様性は、農業者や米の消費者の行動と直接的、または間接的に関わっ
ていると考えられる。本研究は、佐渡市における環境と経済の好循環を目標として、
生物多様性の動態と農家や米の消費者の選択動態を結合した数理モデルを構築した。
水田の生物数、農家の環境保全型農業への取り組み、米の販売量を増加させるため
の対策の効果を明らかにするために、数理モデルによる解析を行った。水田におけ
る生物数を増加させる取り組みや、環境保全型農業への取り組みに対する助成制度
だけでは十分でなく、認証米の販売戦略や米の品質の向上など、消費者の認証米の
購入量を増加させるための対策が重要である事が明らかになった。また、対策が打
ち切られた場合に、生物数、環境保全型農家の割合、認証米の消費者の割合がどの
ように推移して行くのかを解析した。水田における生物数を増加させる取り組みが
逆に生物数を減少させてしまうことや、補助金を打ち切った場合でも環境保全型農
家の割合が高い状態で保たれる可能性が示唆された。さらに、環境保全型農業によ
り、生物数が増加した事を消費者に伝えることにより、認証米の魅力を高める事が
重要である事がわかった。
[P02] アブラムシに化学擬態する捕食者のアリを騙すメカニズム
○林正幸、野村昌史(千葉大院・園芸・応用昆虫)
アブラムシとアリは共生関係を結ぶことが知られ、アブラムシはアリに甘露を提供
する一方、アリはアブラムシの天敵を攻撃し排除する。しかし一部の天敵は、アブ
ラムシに化学擬態することでアリからの攻撃を免れ、アブラムシを捕食する。これ
はアリのアブラムシ認識が体表化学成分に依存するためである。しかし、アリがア
ブラムシの体表化学成分を先天的に識別するのか、それとも学習によって認識する
のかはわかっていない。もしアリがアブラムシを学習し認知するならば、アブラム
シに化学擬態する天敵に対するアリの行動は、アブラムシ随伴経験によって変わる
かもしれない。そこで我々は、アリのアブラムシ認識に学習が関与しているか、ア
リのアブラムシ随伴経験が天敵に対する行動を変化させるか検証した。その結果、
トビイロシワアリは、マメアブラムシに随伴した経験により同種への攻撃性を低下
させた。更に、アリはアブラムシ随伴経験により化学擬態する天敵2種(カオマダ
ラクサカゲロウ、コクロヒメテントウ)に対する攻撃性を低下させた。これらの結
果から、アリはアブラムシを学習すること、アブラムシ天敵の化学擬態の機能はア
リの学習に依存することが示唆された。
[P04] 少数個体からなる群れからみる群れの起源
○新里高行 (筑波大)、三具和希 (筑波大)、村上久 (神戸大)、都丸武宣 (神戸大)、園田耕平
(滋賀大)、西山雄大 (大阪大)、郡司ペギオ幸夫 (早稲田大)
近年になって,画像解析の発達により,魚の群れや鳥の群れの内部でそれぞれの個
31
体がどのような運動をしているのか,徐々に解析されるようになってきた.たとえ
ば、Ballerini et al. (2008) や Cavagna et al.(2010) における鳥の群れの解析は,従来
考えられてきたような群れのモデルが必ずしも正しくはないという結果や,Couzin
らに代表されるように,魚の群れ (Golden Shiner) から集合知の創発を問うたりし
ている.このような流れにありながら,
「群れ」とはどのような現象をさすのか,
「群
れ」が所謂「群れ性」を獲得するのはいかなる条件下において出現するのか,といっ
たことは不問のままである.本研究では,一匹から5匹までの個体群 (アユの群れ)
を長時間観察することにより,その振る舞いに本質的な差異はどこにあるのか,と
いった事を議論する.
[P05] Male-killer prevalence in a sympatric population of specialist and
generalist Harmonia ladybirds
*Suzuki Noriyuki (Tohoku Univ.), Naoya Osawa (Kyoto Univ.)
We compared male-killer bacterial prevalence in two sibling species of the ladybird
that use different host ranges. Harmonia yedoensis feeds mostly on the highly elusive
aphid and its hatchlings can greatly enhance their prey capture ability by consuming
unhatched eggs in the clutch. In contrast, Harmonia axyridis feeds on a wide range
of prey and consumption of unhatched eggs by the hatchlings does not necessarily
increase their foraging success. In the study area, the prevalence of male-killer,
based on low percentage of hatched eggs, female-biased sex ratios, and Spiroplasma
infection, was significantly higher in H. yedoensis than H. axyridis. These results
suggest that the difference in the male-killer prevalence may reflect differences in
the quality of their prey and their foraging behaviour.
[P06] The form of an evolutionary tradeoff affects eco-evolutionary dynamics
in a predator-prey system
*Minoru Kasada (Univ. of Tokyo), Masato Yamamichi (Univ. of Kyoto), Yoshida Takehito
(Univ. of Tokyo)
Rapid evolution on an ecological time scale has been increasingly recognized.
Ecological and evolutionary dynamics can be tightly linked and important to predict
future dynamics, but there is a significant gap between theoretical predictions
and empirical tests, especially on the effects of the nature of genetic variation
such as the form of a fitness tradeoff. Using a predator-prey (rotifer-algal)
experimental system in laboratory microcosms, we studied how different forms of
a tradeoff between prey defense and growth affect eco-evolutionary dynamics. Our
experimental results showed for the first time that different forms of the tradeoff
produced remarkably divergent eco-evolutionary dynamics, including near fixation,
near extinction, and coexistence of algal genotypes, with quantitatively different
population dynamics. A mathematical model, parameterized from completely
independent experiments, explains the observed dynamics. The results suggest that
knowing the details of heritable trait variation and co-variation within a population
32
is essential for understanding how evolution and ecology will interact, and what
form of eco-evolutionary dynamics will result.
[P08] 鳥のさえずりの地理的変異と種の認知:方言に対する反応の非対称性
濱尾章二(国立科学博物館・動物研究部)
鳥のさえずりは種の認知において重要なはたらきをもつ。また、さえずりには種内
でも地理的変異(方言)がある。鳥は異なる地域の方言を同種のさえずりと認知す
るであろうか。この疑問に答えようとして過去に行われた音声再生実験の一部は、
非対称な結果を得ている。つまり、二つの地域のうち一方の個体は他方の方言にも
よく反応するが、逆の実験では異なる方言には反応が弱かった。個々の結果の解釈
は試みられているが、理由は明らかではない。私は、
「さえずりの似た他種が同所的
に分布しているとさえずりを区別する能力が養われ、その結果同種の異なる方言を
も区別するようになる」という仮説を立て、南西諸島のヤマガラとシジュウカラを
用いて検証を行った。複数の島の間で実験を行った結果、ヤマガラでは、シジュウ
カラが同所的に分布している島の個体は、分布していない島の同種さえずりへの反
応が弱い一方、シジュウカラが分布していない島の個体はいずれのさえずりにも同
程度反応した。シジュウカラでも結果は同様であった。このことは、同所的に分布
しているさえずりの似た種の存在が種の認知に影響を与え、生殖行動や遺伝子流動
の方向に影響する可能性を示唆する。
[P10] 徒花の意味
江副日出夫(大阪府大・院・理)
植物の過剰な花生産の適応的意義については、今まで複数の仮説が提案されている。
この研究では、過剰な花生産は、種子捕食者による種子の捕食率を下げることで繁
殖効率を改善しうるという新しい仮説を簡単な数理モデルを用いて検討する。植物
個体は一定量の資源をもち、花生産と果実(種子)生産に配分すると仮定する。こ
の植物の花に無作為に産卵する種子捕食者昆虫が存在し、1 株の生産花数が多いほ
ど 1 花あたりの捕食者密度が低下するとする。1)植物は、全ての花を捕食者産卵
の有無に関わらず成熟させる、2)花のうち一定の割合を捕食者産卵の有無に関わ
らず無作為に遺棄し残りを成熟させる、3)捕食者に産卵された花だけを選択的に
遺棄し残りを成熟させる、という 3 つの戦略を考える。1 と 2 の場合、産卵された
が遺棄されない花は最終的に全部消費されるとする。解析の結果、植物の資源量が
少なく捕食者密度が高い場合、花を遺棄しない戦略は生産する花数が少ないためほ
ぼ全部の花が捕食されるのに対し、遺棄する戦略は花数を多くできるため捕食率が
低くなり、無作為な遺棄であっても繁殖効率を大きく改善しうることがわかった。
[P12] 生態学的個体群制御と群集ネットワークの複雑性
○近藤倫生(龍谷大・理工)、舞木昭彦(島根大・生物資源)
生物群集は互いに直接・間接の効果を及ぼし合う個体群のネットワークとみなせる。
では、この種間効果を利用して特定の個体群を制御する事はできるだろうか?そし
33
て、その制御可能性は群集ネットワークの複雑性とどう関係するだろうか?私たち
は、簡単な群集動態モデルを利用し、捕食者・餌資源の個体群密度を変化させるこ
とによる標的個体群の制御(生態学的制御)の困難さについての理論研究を行った。
単純な群集ネットワークでは、標的種と強い関わりを持つ捕食者・餌生物種の個体
群密度を変化させる事で標的個体群により大きな影響を与える事ができる。しかし、
その効果には大きなばらつきがあり、
「思いもよらない反応」が生じやすいという問
題があることがわかった。それに対して、複雑な群集ネットワークでは、標的種と
の直接の関わりの強弱に関わらず、捕食者・餌生物種の個体群密度の変化が標的個
体群に及ぼす影響は非常に小さかった。このことは、標的個体群に望ましい効果を
及ぼすのには、より多くの努力が必要になる事を意味している。これらの結果は、
生態学的制御に伴う困難さは、群集ネットワークの複雑さによって異なる可能性を
示唆している。
[P16] シロアリ女王の秘策:卵門を閉じて王と同居しながら単為生殖
○矢代敏久(京大院・農・昆虫生態)、松浦健二(京大院・農・昆虫生態)
オスは有性生殖でしか次世代に遺伝子を残すことが出来ない。それゆえ、メスによ
る単為生殖の進化はオスにとっては適応度上の大きな脅威であり、オスはメスに有
性生殖を強いるように対抗進化すべきである。しかし、ヤマトシロアリの女王は同
居している王と継続的に交尾を行い有性生殖で職蟻や有翅虫等を生産する一方で、
同時に二次女王(巣内で創設女王の生殖を引き継ぐ)を単為生殖で生産している。
一体、シロアリ女王はどのようにして王と同居しながら「単為生殖の子」を生産し
ているのだろうか?本研究によって、女王は卵門(精子が卵に侵入する穴)の無い
卵を産むことによって、王の存在下であっても受精を拒否し、単為生殖の子を生産
していることが明らかになった。さらに、女王は継時的(齢・季節)に卵門数を調
節することによって、生産される単為生殖卵の割合を制御していることも明らかに
なった。この「卵門を閉じる」というシロアリ女王の戦術は、二倍体生物における
メスによる卵の受精制御メカニズムの初めての解明であり、オスからの干渉の有無
に関わらずメスの戦略として単為生殖が進化し得ることを示している。
[P18] 生息環境の細分化の程度と種間競争の結果
穴澤正宏(東北工大・工・環境)
一般に、個体群や群集の動態は、生息環境の空間構造により大きな影響を受けると
考えられる。パッチ状の環境において共通資源をめぐり競争している生物の場合、
生息地の細分化の程度が変わると、種間競争の結果(共存するか、どちらが勝つか)
はどのように変化するだろうか?全体の資源量が一定のままパッチ数が増加すると、
パッチあたりの資源量が小さくなるため、少ない資源量で多くの子供を産める種が
競争に有利になっていくかもしれない。このように、生息環境がより細分化したと
きの影響は、各個体が繁殖するのに必要な資源量や資源競争の様式(スクランブル
かコンテストか)など、個体レベルの資源競争の詳細に依存するはずである。本発
表では、個体間の資源競争に基づいて第一原理的から導かれた動態モデル(方程式)
を使って、生息地の細分化の程度が変化したときの、種間競争の結果や個体群動態
の変化について、考察した結果を報告する。特に、細分化の程度が大きくなるにし
34
たがい種間競争の勝者が入れ替わることがあることや、このような勝者の入れ替わ
りの起きやすさは競争の様式(スクランブルかコンテストか)により異なることな
どを紹介する。
[P19] Accounting for the spatial structure of communities in the estimation of
transition probabilities subject to observation errors
Keiichi Fukaya (ISM), J. Andrew Royle (USGS), Takehiro Okuda (NRIFSF), Masahiro
Nakaoka, Takashi Noda (Hokkaido Univ.)
Markov models have been applied to sessile assemblage data to infer transition
probabilities, although estimates may be biased when the observation error of the
occupancy state exists. New methods have recently been proposed to estimate
transition probabilities while accounting for the observation error. In these methods,
however, any spatial structure of the community is not considered explicitly. Since
sessile organisms typically represent a spatially aggregated pattern, it seems likely
that the local structure of the community composition affects observation, state
transitions, or both. Hence, accounting for such spatial dependence may be a key
to capture underlying community dynamics and observation processes accurately.
In this study, we consider a hierarchical model to estimate Markov community
dynamics of sessile species which explicitly considers the spatial variation in species
frequency and its effect on observation processes. By using a nonparametric
kernel regression estimator, the model estimates the relative frequency of each
species (state) ’near’ each observation point which is related to the probability
distribution of observed data. We apply this model to a long-term intertidal sessile
community data set and show that the new model provides corrected estimates of
transition probabilities, even when the spatial dependence of community structure
is anisotropic.
[P20] 性的対立の波及効果:より発達した交尾器の棘をもつオスほど他種メスにコストを
与える
○京極大助、曽田貞滋(京都大・理・動物生態)
オスとメスの進化的な利害の対立(性的対立)は有性生殖種に広く見られる。性的
対立の典型的な帰結のひとつは、オスにとっては有利だがメスにとってはコストと
なるようなオス形質の進化である(e.g. 交尾器の棘)。オスはしばしば近縁種のメス
とも配偶を試みる。このため、性的対立の結果発達してきたオスの加害形質が、種間
交尾などを介して他種のメスに対してもコストを与える可能性が考えられる。我々
はアズキゾウムシ・ヨツモンマメゾウムシを用いてこの仮説を検証した。アズキゾ
ウムシのオス交尾器には棘があり、またアズキゾウムシのオスとの種間交尾によっ
てヨツモンマメゾウムシの産卵数が下がることが先行研究から分かっている。実験
の結果、棘の長いアズキゾウムシのオスほどヨツモンマメゾウムシのメス生殖器を
傷つけやすいこと、また生殖器の傷ついたヨツモンマメゾウムシのメスの産卵数が
低下することが明らかとなった。これらの結果は種内の性的対立によって発達して
きたオス形質が他種メスにとってもコストとなることを示唆している。このような
35
性的対立の種間関係への波及効果が、近縁種間の共存を妨げる要因の一つとなって
いるのかもしれない。
[P21] セイタカアワダチソウの侵入地とその起源集団におけるアワダチソウグンバイの密
度の比較
○坂田ゆず (京大・生態研), Timothy Craig (University of Minnesota, Duluth), Joanne
Itami (University of Minnesota, Duluth), 大串隆之 (京大・生態研)
植物の防御形質は、生物・物理的環境に応じて変化するため、種内で地理的変異が
見られる。そのため、外来植物の侵入地での防御形質の進化を明らかにするために
は、侵入集団の起源となる原産地の集団との比較が必要である。外来植物セイタカ
アワダチソウは 2000 年に侵入した外来の植食性昆虫アワダチソウグンバイと日本
で再会し、グンバイの侵入が防御形質に急速な適応進化を生じさせていることが分
かってきた。本研究では、中立マーカーによる遺伝解析から明らかになった日本の
セイタカと最も近縁な北米南部の集団において、グンバイの個体数と食害率の調査
を行った。グンバイは北米南部地域の広域にわたって高い密度で分布しており、食
害率も高かった。気候要因を考慮して比較したところ、北米に比べて日本ではグン
バイの密度が高いことが示された。これらの結果から、グンバイの密度には気候要
因が大きく関わっているが、日米の違いには、それに加えて日本でのセイタカの防
御形質の低下が関わっている可能性が考えられる。今後、北米南部集団のグンバイ
への抵抗性を調べることで、日本のセイタカの侵入プロセスにおける防御形質の進
化が明らかになることが期待される。
[P22] Natural selection, genetic variation, and plasticity in eco-evolutionary
dynamics: a case study of Plantago lanceolata
Richard P. Shefferson (University of Tokyo), Deborah A. Roach (University of Virginia)
The demography of wild populations is often relatively stable over many years.
Many have interpreted this to suggest that natural selection is unimportant in
determining demographic patterns. An alternative hypothesis is that plants deal
with strong, temporally variable selection via adaptively plastic demographic
responses. We explore this hypothesis with a long-term, experiment study of
Plantago lanceolata. Approximately 8,000 plants in four cohorts of equivalent
genetic background were planted at different times between Nov 2000 and Nov
2002, and their fates, sizes, and flowering patterns were monitored from then until
the present. We used these data to build density-dependent simulations of Plantago
population dynamics in which genotypes had varying levels of plasticity in their
vital rates. We found that in the deterministic simulations, and in simulations with
fluctuating environments in which plants were not allowed to respond plastically,
natural selection led to consistently increasing population growth rate, reflective
of Fisher’s fundamental theorem. However, this tendency was severely slowed in
simulations allowing adaptive plasticity in vital rates. We suggest that demographic
buffering most likely stabilizes populations because of natural selection, rather than
36
in spite of it, since our simulations suggest that adaptive plasticity in vital rates
should be selected for.
[P26] キアゲハの季節多型(春型・夏型)の適応的意義:体温調節か保護色か?
○正木奈穂、西口泰平、石原道博 (大阪府大・院・理)
変温動物である昆虫は外気温の影響を直接受けるため、体温を適切に制御できなけ
れば飛翔や繁殖行動に負の影響が及ぶ。キアゲハの成虫は、春型と夏型という 2 つ
の季節多型を示す。夏型は気温の高い夏に出現し、春型よりも体サイズが大きく、
翅の色彩は黒色が強い。一方で、春型は気温の低い春に出現する。そのため春型と
夏型の間に体温調節という点で違いがあると考えられる。そこでキアゲハの卵を短
日条件と長日条件に分けて飼育し、人為的に春型と夏型を生じさせた。各季節型の
成虫は 5 ℃に置かれ、体温を十分に下げられた後、25 ℃の恒温器に移して加温され
た。加温開始から 10 分間、サーモグラフィーを用いて熱画像を撮影し、胸部体温
の経時変化を解析した。その結果、春型は夏型よりも速く体温を上昇させた。しか
し、白熱電球で加熱する同様の実験では、夏型の方が春型よりも体温が速く上昇し
た。このため、両者に生理的な違いはあるものの、適応的意義という点では翅の色
彩の体温調節への関与は弱いことが示唆された。本講演では、季節型間で見られる
翅の色彩の違いが保護色として機能している可能性についても検討する。
[P27] 小標本データに対するブートストラップ信頼区間の構成
○岡村 寛・市野川 桃子(中央水研)
個体群生態学では,小標本のデータに対して統計的推測を行うことも少なくない.小
標本のとき,興味の対象のパラメータに対する信頼区間に t 分布を使用することがよ
く行われるが,t 分布に基づく方法がいつもうまく働くわけではない.我々は,小標
本データに対する統計的推測のための新しいブートストラップ法,Double residual
bootstrap(DRB)法,を開発した.DRB では,残差を独立に二度リサンプリング
する.平均の推定に関して,DRB によるブートストラップ分布は t 分布に一致する.
シミュレーションによって,より一般的な問題に対してもうまく働くことが確認さ
れた.
[P28] 性の存在が群集動態に与える影響:ギルド内捕食系を用いた理論的検証
川津一隆
生物群集の多数派を占める有性生物は,オスによる子への投資が少ないという共通
の特徴を持つ.また,子への投資の性差は性選択や性的対立を生むため,結果とし
て捕食行動や防衛行動でもオス・メス間で違いがみられる例が少なくない.これら
の性差は相互作用強度や栄養転換効率などの食物網におけるエネルギーの流れ方に
構造をもたらし,その結果,有性群集は無性的な群集と比べると異なる動態をみせ
ると予想される.しかしながら,群集生態学の多くは食物網ネットワークの形など
の群集レベルで現れる大きな構造に着目するのみで,性差のような行動レベルで現
れる構造が群集動態に与える影響についてはわかっていないことが多い.そこで,
37
本研究では被食者・捕食者側それぞれの性構造が系の動態と安定性に与える影響を
調べるために,3 種からなるギルド内捕食系という最小限の生物群集を用いて解析
を行った.また,環境生産力を一定に保つことで2つの階層の性構造が群集サイズ
に与える影響についても同時に解析を行った.発表では得られた結果にもとづき性
構造が現実の生物群集においてどのような機能を果たしているかについての考察を
行う.
[P29] ダムによる魚の移動阻害の影響―メタ生態系モデルを用いたメコン川ダム開発の影
響評価―
○吉田勝彦(国立環境研・生物)、広木幹也(国立環境研・生物)、 冨岡典子(国立環境
研・地域)、村田智吉(国立環境研・地域)、福島路生(国立環境研・生物)
メコン川は淡水魚の多様性が高く、世界最大の漁獲量を持ち、流域住民の食糧供
給に重要な役割を果たしている。しかし、近年の著しい経済発展に伴う電力需要の
増大などを受けて、複数のダム建設計画が進められている。そこで本研究では、これ
まで開発を進めてきた川のメタ生態系モデルを用いてコンピュータシミュレーショ
ンを行い、ダム建設に伴う魚類の移動阻害が魚類生態系に与える影響を解析した。
その結果、ダムの直上、直下で魚種の絶滅率が有意に高くなった。季節移動を行う
魚よりも、行わない魚の方がダム建設の影響を強く受け、絶滅率が大きくなった。
しかし本研究シミュレーションでは、1 回ごとのシミュレーションの結果のばらつ
きが非常に大きかった。その原因は、メコン川の生態系には、多くの撹乱要因が内
包されていることにあると考えられる(短期間で大規模に環境が変化すること(雨
季と乾季)、生物相の入れ替わりが起こること(魚の季節移動)など)。本研究の結
果は、メコン川でのダム建設のコストは大きめに見積もる必要があることを示唆し
ている。
[P30] ハダニの交尾前ガード行動と捕食リスク:気にする♀、しない♂
○奥 圭子 1,2,3 , Erik H. Poelman2 , Peter W. de Jong2 , Marcel Dicke2 (1JSPS 海外学振,
2Wageningen Univ., 3 中央農研)
雌雄間において、必ずしも損得が一致するとは限らない。ハダニの♀は初回交尾の
み受容するため、♂成虫が成虫化直前の♀を交尾前ガードする。交尾前ガード時の、
雌雄それぞれの捕食リスクに対する反応と捕食され易さの関係を調べた。捕食リス
クがない時、♂にガードされた♀(ガード♀)と単独♀が居ると、多くの♂は前者
の♀を選ぶ。しかし、♀が捕食リスクを経験していると、♂は♀に対する選好性を
示さなかった。その一方、♂は捕食リスクの有無に関わらず、ガード♀への選好性
を示し、♀をガードした。以上のことから、♀だけが捕食リスクに反応を示すこと
が示唆された。ハダニのガードペアは、単独♀よりも捕食者カブリダニに見つかり
易かった。カブリダニはガードペアの♀と♂を同程度に攻撃したが、最終的には♂
よりも♀を多く捕食した。ハダニ♂成虫は自由に動き回ることができるが、成虫化
直前の♀はまったく動くことができない。♀が捕食され易いのは、この行動制限の
ためであり、♀が捕食リスクに反応を示すのは、交尾ガード時の捕食を避けるため
であると考えられた。
38
[P31] Does group size influence aggressive behavior of Formica japonica in group
fighting?
*Suguru Sasaki and Gaku Takimoto (Toho university)
In fights between groups of social animals, individuals often use information
on group size to decide whether to engage in struggles, because group size can
be an important factor that influences battle outcomes. Ants are social insects,
and fight strongly with intraspecific different colonies or interspecific colonies to
defend their territories, nests and food resources. It may thus be possible that
group size influences decision-making of ant individuals when fighting. In this study,
we conducted a series of laboratory experiments to test whether group size affects
individual’s decision of Formica japonica about whether to fight with Camponotus
japonicus. Individuals of F. japonica became more aggressive against C. japonicus
when they belonged to larger groups prior to fighting. In addition, the probability
of joining a fight increased when a F. japonica individual was in larger groups during
fighting. These results demonstrate that group size influences aggressive behavior
of F. japonica individuals, and parallel previous similar findings from vertebrates,
such as canids and primates. We argue that group size can be an important cue
for making a decision about joining a fight not only in vertebrates but also in
invertebrates.
[P32] 生活史雑食による捕食−被食関係の逆転とギルド内捕食系の共存
○川田尚平、瀧本岳(東邦大・生物)
ギルド内捕食系(IGP 系)は基底種、基底種を捕食する消費者、基底種と消費者両
方を捕食する雑食者から構成される単純な食物連鎖である。初期の理論研究では、
IGP 系において基底種の生産性が高い時、雑食者と消費者が共存できないとされて
いる。しかし、実際の IGP 系では基底種の生産性が高くても共存できる場合があり、
必ずしも理論の予測と一致するとは限らない。この問題を解決するため、本研究で
は、雑食者の生活史雑食を組み込んだ IGP 系のモデルを構築した。雑食者の幼体は
消費者に捕食されるが、成体になると消費者を捕食するようになり、捕食−被食関
係が逆転する状況を想定した。このような捕食−被食関係の逆転は、例えばプラン
クトン群集などで知られている。解析の結果、基底種の生産性が高い時でも、消費
者と雑食者が共存する可能性があることが分かった。消費者と雑食者の共存には、
消費者が雑食者の幼体を捕食する率が、雑食者の成体が消費者を捕食する率と同等
以上であることが必要であった。この結果は、IGP 系における理論的予測と実証的
な結果の不一致の解消に貢献するものである。
[P33] 個体群成長率とプロセスの時空間変動性:岩礁潮間帯の固着生物群集における推移
行列モデルを用いた解析
○金森由妃(北大・院・環境科学),深谷肇一(統数研),岩崎藍子(北大・院・環境科
学),野田隆史(北大・地球環境)
39
環境の異質性は生物の数と分布を決める主要因とされ,環境の異質性と群集構造
の対応関係については多くの知見が蓄積されている.一方で環境の時空間変異に対
応して群集動態がどのように変化するかについての研究例は極めて少ない.この隔
たりが群集パターンの生成プロセスや多種共存機構の理解不足の一因かもしれない.
固着生物群集は全ての種が空間を巡る競合関係にあるため,古くから群集動態や
多種共存機構を研究するモデル系として注目されてきた.推移行列モデルは固着生
物群集の動態,つまり各種の空間の占有状態の時間変化,を表す一つの方法である.
このモデルを用いることで群集構成種の個体群動態とそれに影響するプロセスの時
空間変異性を明らかにすることが可能である. 岩礁潮間帯の固着生物群集では群
集構造が潮位や季節によって変化することが知られている.そこで本研究では岩礁
潮間帯の固着生物群集を対象に推移行列モデルを適用し,各種の個体群成長率とそ
の背後にあるプロセス(加入,存続,置換,死亡)について,潮位や季節による変
動性を明らかにする.
[P36] 針葉樹林業地域における広葉樹林がカスミザクラの結実と交配に与える効果
○永光輝義、滝久智、菊地賢、加藤珠理(森林総研)
林業地域の生物多様性の保全には、残存する広葉樹林の生態系機能を明らかにする
必要がある。そこで、ハナバチの送粉機能に注目し、広葉樹林の面積と形状で表さ
れる景観構造がハナバチ媒であるカスミザクラの結実と交配に与える影響を調べた。
茨城県北部の 15 km 四方の針葉樹林業地域に残された広葉樹林に、互いに 1 km 以
上離して 12 サイトを設けた。各サイトで 2012 年と 2013 年に、カスミザクラの開花
木を記録し、約 12 母樹のサイズと葉緑素濃度(SPAD 値)を測り結実した枝を採取
した。そして、サイトごとに、カスミザクラの結実数、果実重、充実率(胚の発育
率)、他殖率、二親性近交係数(両親の血縁度)、母樹内父性相関(交配の単純性)
を求めた。その結果、送粉機能の景観構造依存性が示された。つまり、広葉樹林の
面積が広いと、カスミザクラの胚の発育と交配の多様化が促進された。一方、カス
ミザクラの結実は母樹の資源(サイズと SPAD 値)に、胚の発育は両親の血縁度に
依存していた。このように、景観構造でない要因の重要性も明らかになった。
[P38] A facultative mutualist increases the persistence of obligate mutualism
Gaku Takimoto (Toho University)
Mutualism is widespread and important biological interactions, but mathematical
theory on mutualism has been less developed compared to other types of
interactions. A basic Lotka-Volterra type model of two-species mutualism predicts
unbounded population growth, which is circumvented by additional assumptions
such as strong negative density-dependence and saturating benefits from mutualistic
interactions. Existing models of two-species obligate mutualism typically predict
bistability, in which interdependent mutualists inevitably go extinct when their
abundance fall below certain thresholds. Here I consider a case in which one of
the obligate mutualists interacts with a facultative mutualist. A mathematical
model shows that the facultative mutualist increases the persistence not only of
its interacting obligate mutualist, but also of another obligate mutualist with
40
which the facultative mutalist does not interact directly. This result highlights the
importance of multi-species context in understanding the persistence of mutualistic
communities.
[P39] 状態空間モデルによるアライグマの個体数推定と空間明示的な作物被害の許容密度
の推定
〇栗山武夫、長田 穣(東大・院・生物多様性)、浅田正彦(AMAC)、宮下 直(東大・
院・生物多様性)
アライグマは北米原産の外来中型哺乳類で、在来生物への捕食、人家や寺社への侵
入、人獣共通感染症の媒介や農作物被害など様々な問題を起こし、特定外来生物と
して捕獲事業が進められている。アライグマの農作物被害を軽減させるためには、
各地域の許容生息密度を推定し、管理目標を設定することが重要である。さらに、
農地周辺の景観構造が地域によって均一ではないため、生息密度と被害程度の関係
が局所的に異なることが予想される。そこで本研究では、千葉県を対象地とし、ア
ライグマの被害許容密度を、農地周辺の景観構造から2四方で空間明示的に予測す
ることを目的に、まず個体群動態を明示的に扱った状態空間モデルを用いて個体数
推定を行い、推定された個体数と景観構造を独立変数としたロジスティック回帰を
行うことで被害程度との関係を明らかにした。の状態空間モデルは、捕獲事業によ
る捕獲効率(CPUE)をもとに構築し、市町村単位で生息密度を算出した。の被害
予測モデルは、応答変数に農作物の被害アンケートを使用し、被害地域周辺の景観
構造は、森林・耕作地・耕作放棄・市街地・河川面積等を用いた。
[P40] 同所的に生息する二種の同属ダンゴムシに感染する繁殖操作の異なるボルバキア
角拓人(岡山大)、澤谷祐輝(岡山大)、三浦一芸(NARO)、○宮竹貴久(岡山大)
同所的に生息する 2 つの同属の種類において異なる繁殖操作を受けていると考えら
れるダンゴムシが見つかった。古くから日本の常緑樹林に生息するコシビロダンゴ
ムシは、多くの種に分化しているとされているが、岡山市内の半田山には 2 種のみ
が分布していた。両種は、ともにコナラ属、常緑広葉樹の林床で同所的に生息し、
異なるボルバキアに感染していた。Spherillo dorsalis sensu Nunomura (1990, 2003)
は、性比が大きくメスに偏っており(90%以上)、メスの8割がボルバキアに感染
し、オスの感染率は1割程度であった。これに対して、Spherillo sp. shi-1 sensu 唐
沢・川野(2014)は、メスバイアス(65%)ではあるものの、両性ともに 9 割がボ
ルバキアに感染していた。この結果は、前者のボルバキアがメス化もしくはオス殺
しであるのに対し、後者のそれは繁殖を操作していないか、もしくは細胞質不和合
であると考えられた。また両種とも、感染率の季節変動は認められなかった。
[P41] Temporal chain reactions of indirect interactions initiated by intraspecific
variation in an early-season herbivore
Shunsuke Utsumi (FSC, Hokkaido Univ.)
Plant-insect interactions represent intricate networks at all levels of biological
41
complexity. Although systems approach to linking these complex networks at
different levels of biological integration will be a major challenge, analyzing
chain reactions of indirect interaction units may provide an insight into the
complexity. The hypothesis in the present study is that feeding evolution of the leaf
beetle Plagiodera versicolora long influences community structure through indirect
interactions. To test this hypothesis, a field experiment was conducted. The
gourmet type beetle exclusively fed on new leaves of willows. In contrast, the
no-pref type displayed non-preferential feeding between new and mature leaves.
Different proportions of gourmet and no-pref type beetles were introduced onto
mesh-covered willow shoots in spring. After six days, the covers and beetles
were removed, and the community assembly dynamics of the naturally colonized
herbivore species was investigated. Then, community assembly dynamics were
long influenced by the interaction between initial beetle treatment and subsequent
colonizer species identities. This process was driven by temporal chain reactions of
indirect interactions. As a result, species diversity in local herbivore communities
significantly differed between initial treatments.
[P42] 果実の摘果が生物的防除の効率に与える影響についての数理的考察
○池川雄亮、江副日出夫、難波利幸 (大阪府大・院・理)
農業害虫に対する生物的防除の有効性を検討する場合、害虫と天敵の生活史、行動、
個体数変化のみが注目するだけでなく、人為的な栽培管理が害虫と天敵の個体群動
態や生物的防除の効率に及ぼす影響を考慮する必要がある。本研究では、栽培管理
の一例として質の良い果実を得るために一部の幼果を間引く摘果処理に注目し、摘
果が生物的防除の効率に及ぼす影響を、作物-害虫-天敵の系に関する数理モデルを
用いて調べた。作物の果実には 3 つの生長段階(花、幼果、果実)があり、幼果期
にすべての作物個体に対して同程度の摘果が行われると仮定した。作物個体は 2 次
元格子空間上に分布し、害虫及び天敵は近傍作物のみに分散することができると仮
定した。計算機シミュレーションによる解析の結果、摘果を行わない場合は、天敵
個体群の増加によって、害虫はたとえ一時的に増加しても速やかに減少した。一方、
摘果量が増加すると、天敵個体群は存続しているにもかかわらず、摘果なしの場合
に比べて害虫個体群の減少が鈍くなった。この傾向は、天敵の分散率が小さい場合
により顕著であった。以上より、栽培管理が生物的防除の効率に与える影響を天敵
の分散率と関連付けて考察する。
[P43] 遺伝子と個体数分布に基づいたサドガエル個体群のネットワーク構造の推定
○山中美優 (東大・院・農), 小林頼太 (東環工専), 関谷國男 (新潟大), 宮下 直 (東大・院・
農)
希少種の有効な保全策を考える上では、複数の空間スケールから個体群構造を把握
し、その成因を探る必要がある。本研究では、佐渡島固有種のサドガエル (Rugosa
sussura) を対象に、以下の項目の解明を目的とした。(1) 個体の遺伝子型による地域
個体群の特定;(2) 地域個体群内の生息地ネットワークの推定;(3) 個体数情報を用
いた生息適地の条件となる環境要因の特定;(4) 遺伝子情報と個体数情報に基づいた
42
保全や再生適地の探索。遺伝子解析では、24 集団 373 個体についてマイクロサテラ
イト 7 座位を用いた。(1)STRUCTURE 解析により、サドガエルは 3 つの地域個体
群に分けられた。(2) 生息地間の遺伝的共分散を用いた Population Graph から、地
域個体群内では一部の生息地が環状構造を形成していたことがわかった。(3) 佐渡島
の水田 42 か所において GLM を用いて個体数に影響する環境要因を探索した結果、
水田周辺の水路が浅く、湛水面積が大きい水田群で個体数が多かった。(4) 以上の
結果をもとに、サドガエル保全に重要な生息地を、生息地の遺伝的多様性とネット
ワークの中心性から推測する。
[P44] A stranger is tastier than a neighbor: Cannibalism in Mediterranean and
desert populations of pit-building antlions
*Ofer Ovadia and Erez David Barkae (Ben-Gurion University of the Negev)
Cannibalism is a widespread trophic interaction, which can be enhanced by various
environmental stresses. Individuals originating from Mediterranean habitats are
typically better adapted for coping with competition an adaptation that can
manifest itself in increased cannibalism tendency. Conversely, dessert environments
are characterized by long periods of food shortage, also selecting for increased
cannibalism tendency. We tested for differences in the frequency of cannibalism
between Mediterranean and desert populations of the pit-building antlions. We also
quantify the frequency of cannibalism within and between antlion populations. We
could not detect a significant geographic variation in the frequency of cannibalism
neither within nor between climate regions. We suggest that plastic responses to
environmental stresses buffer against divergent selection, and thus no significant
differentiation in cannibalism tendency could occur. Notably, the frequency of
cannibalism was higher between than within populations, however, cannibal identity
was almost exclusively determined by its initial body mass advantage, rather than
by its population-of-origin. We suggest that this intra-specific discrimination ability
is mediated by behavioral and or chemical cues, associated with co-occurring in a
common environment. Future research should focus on exploring these cues, part
of which could be the basis for a kin recognition system to evolve.
[P46] サンショウウオの共食いがトップダウン効果に及ぼす影響は年変異する
〇高津邦夫(北大・院・環境科学)、岸田治(北大・北方圏 FSC)
相互作用は、生物種の特徴(個体数や構成個体の表現型)を変えることで、その種
が関わる別の相互作用に影響することがある。その影響の方向や強さは時間ととも
にどのように変わるのだろうか。本発表では、「捕食者種の種内相互作用(共食い)
は、捕食者種から餌種への捕食圧に影響を与えるが、その方向が餌種のコホートに
よって異なる」ことを示した研究を報告する。私たちは、孵化直後のエゾサンショ
ウウオ幼生(サンショウウオ)の共食いを操作して、その年に孵化したエゾアカガ
エル幼生のコホート(当年オタマ)と翌年孵化したコホート(翌年オタマ)に対す
る捕食圧の大きさを調べた。サンショウウオが共食いしなかった場合に比べ、共食
いした場合には、当年オタマに対する捕食圧は 1.5 倍だった(共食いが捕食圧を強め
43
た)が、翌年オタマに対する捕食圧は半分以下だった(共食いが捕食圧を弱めた)。
これは、サンショウウオの共食いがある場合には、共食いに成功したサンショウウ
オが巨大化して当年オタマに強い捕食圧をかけたが、翌年まで池に残るサンショウ
ウオが非常に少ないため、翌年加入してきたオタマに対して捕食圧がほとんどかか
らなかったためである。
[P47] Multinomial mixture model によるオニヒトデ捕獲除去データからの駆除達成
率の推定
⃝熊谷直喜(国環研)、山川英治・白木一太朗(沖縄県環境科学センター)、比嘉義視(恩
納村漁協)、岡地賢(コーラルクエスト)
個体数を直接的に調査できない動物は多いが、近年の統計モデリングの発展によって、
間接的データからでも個体数推定が可能になった。本研究は Multinomial mixture
model による、食害生物の反復捕獲除去データからの生息数推定結果を報告する。
研究対象は、不定期に大発生し造礁サンゴ群集を食害するオニヒトデとし、沖縄県
恩納村漁協で実施の駆除作業のデータを使用した。駆除は潜水作業が主要手段であ
り、駆除の成功には時間的・人的資源を効率的に配分する必要がある。このため生
息数推定に加え、駆除達成の目安作業量の推定も行った。1 回の作業による推定駆
除率は 0.2 となった。また個体数を 5% まで減少させるのに必要な駆除回数は 15
回となり、これは恩納村で重点駆除を実施している海域の作業回数と同等であった。
元の生息数は 2003500 個体/km2、推定駆除達成率は平均 60% となり、多くの地点
では駆除後は許容限度個体数以下に抑制されたと見なせる。今後さらに推定誤差を
抑えるため、駆除作業範囲の空間的重なり等を考慮した拡張モデルによる推定も行
う予定である。
[P48] 人為的撹乱に対する山菜の応答∼収穫することで翌年のタケノコの生産は増す
⃝片山昇、岸田治、坂井励、早柏慎太郎、伊藤欣也、実吉智香子、浪花愛子、高橋廣行、
高木健太郎(北大・FSC)
「山菜」は、森林から享受される「生態系サービス」として古くから人に愛されて
きた存在である。人は山菜を収穫することで山菜に撹乱を与えるが、そのような人
為的撹乱に対して、山菜はどのように応答するのか?このような問題の解決は「い
つどこで山菜がたくさん採れるのか?」という一般の人の素朴な疑問に科学的に答
えることであり、社会的な意義は小さくない。本研究では、高級山菜として知られ
るチシマザサのタケノコを対象とした大規模な操作実験を実施し、タケノコの生産
に対する収穫の影響について調べた。北大天塩研究林内に 10 x10 m の実験区を 21ヵ
所設置し、そのうちの半数を収穫区として、一般的なタケノコ採取の基準と強度で
タケノコを収穫した。もう半数は対照区としタケノコの採取を行わなかった。翌年
に各実験区でタケノコ密度を調べた結果、収穫区では対照区の3倍のタケノコが生
産されていた。多くの植物種では、損傷に応答して損傷で失った以上に新しい組織
を生産する(補償反応)。このような補償反応によって、「収穫後にタケノコの生産
は増える」と考えられる。本研究は、基礎生態学的な知見を実学的な課題へと応用
する試みといえる。
44
[P49] 気候と景観構造の地域差がもたらす高次捕食者サシバの繁殖適地の違い
○藤田 剛(東大・農)、東 淳樹(岩手大・農)、野中 純(オオタカ保護基金)、堺 義昭、堺 初美(印西サシバの会)、伊関文隆(希少生物研)、深澤圭太(国環研)、
宮下 直(東大・農)
生物分布と景観構造の関係を統計的に推定する生息適地モデルは、保全優先度の評
価や保護区策定、開発の影響予測など、景観スケールでの保全活動で重要な役割を
果たしてきた。データ収集などに労力がかかるため、保全活動では他地域で作成し
たモデルをそのまま対象地域に適用する例が多いが、こうしたモデルの転用は、過っ
た生息地評価を導く可能性がある。そこで演者らは、里山景観の代表的な高次捕食
者サシバを対象に、複数地域 (分布中心3地域、周縁2地域) にわたって生息適地を
評価し、その地理的差異と共通点を調べた。サシバは、農地と森に生息するカエル
や昆虫などを採食するため、両方を利用できる林縁が地域を通して重要な景観要素
だと考えられる。一方、著しく分断化の進んだ地域では、農地や森などの個々の要
素の面積の重要度が高まっている可能性がある。また、気候や農事暦の変化によっ
て餌密度等の生息地の質に違いが生じ、それが農地や森の重要度を変化させる可能
性も考えられる。発表では、解析結果にもとづき、生息適地の地理的差異が生じる
機構を議論するとともに、生息地保全上の意義も議論する予定である。
[P50] 農法と周辺景観が水田のアシナガグモ‐餌昆虫の季節動態に及ぼす影響
⃝筒井優(東大・農)、田中幸一(農環研)、馬場友希(農環研)、宮下直(東大・農)
水田は湿地棲生物の代替生息地として知られており、田面と用排水路が組み合わさっ
た構造をしているため、農事歴による田面環境の変化に対して多くの生物が個体群
を維持していることが知られている。しかしながら、農業の集約化による、水路の
暗渠化や農薬・化学肥料の散布が多くの生物を減少させつつある。これに対処する
ため、生物多様性や生態系サービスの向上を目的の一つとした環境保全型農業が広
まりつつある。環境保全型農業が生物多様性に及ぼす効果は、周囲の景観要因や生
物間相互作用で異なり、慣行栽培と差が生じない場合も知られている。本研究では、
害虫密度と負の相関があり、水田で優占する捕食者として知られるアシナガグモ類
とその餌昆虫としてユスリカを主とした双翅目に着目した。これらは、水環境に敏
感に応答するため、農事歴により大きく個体数が変動すると推測される。そこで田
面環境の異なる 3 時期で環境保全型農業と森林からの距離が田面と水路のアシナガ
グモ類と小型飛翔昆虫に及ぼす影響を調査した。本発表では各時期で森林からの距
離と環境保全型農業がアシナガグモ類と餌昆虫に影響を及ぼしたプロセスについて
議論する。
[P51] 共生する植物-バクテリアでみられる野外集団間による変異
○鍵谷進乃介(北大・環境科学院)、内海俊介(北大・北方生物圏 FSC)
植物-バクテリアの共生関係として、大気中窒素を固定し宿主植物に供給する根粒菌
とマメ科植物との関係が知られ、広く研究されている。その根粒菌とは系統的に遠
縁である放線菌にも、ハンノキ属やヤマモモ属などの根に根粒を形成し、窒素固定
45
産物を供給するという根粒菌と類似した共生関係を築く土壌細菌、フランキアがい
る。しかし、根粒菌-マメ科共生系に比べ、フランキア共生系に関する研究は進んで
おらず、野外での実態も解明されていない。近年、様々な共生系において、共生関
係の帰結に大きく影響する共生者両種の種内遺伝変異が広く存在すること、遺伝変
異の効果は生物・非生物的環境要因によっても変化することがわかってきている。
そこで、我々はフランキアとケヤマハンノキを使い、各々の野外集団間によって共
生関係に変異があるという仮説の検証実験を行った。北大雨龍研究林に生育するケ
ヤマハンノキ 3 集団の合計 14 個体から種子を採取し、その発芽実生に同ハンノキ 3
集団から採取したフランキアを摂取した。摂取後のハンノキの生存率、野外環境下
での成長率や成長速度を比較した結果、ハンノキとフランキアそれぞれの集団間で
特性が異なることが示唆された。
[P52] ハムシの餌選好性はどの形質と遺伝的に相関するか
○小野寺裕乃 (北大・環境科学院)、内海俊介 (北大・北方生物圏 FSC)
近年、生物群集と適応進化を統合的に理解する試みがなされている。遺伝相関は、
ある形質が進化する際に他の形質がどう進化するかをあらわすので、形質進化が生
物間相互作用へどう波及するか示唆を与える。同時に、遺伝相関は生物群集の構造
が形質進化に影響を与える主要なメカニズムのひとつである。したがって、遺伝相
関を明らかにすることは、進化が生物群集に波及するプロセスと、生物が形質進化
に影響するプロセスについて洞察が得られる。本研究では、ヤナギルリハムシ (以
下、ハムシ) を対象に、餌選好性とその他形質の遺伝相関を明らかにすることを目
的として研究を行った。一般に植食者の餌選好性は、植物の質や子の適応度から理
解されることが多いが、自身の捕食者回避や移動分散などの形質と遺伝相関を明ら
かにした研究はほとんどない。まず私たちは、北海道北部の半径 40km 圏内の 4 つ
のハムシ集団間に餌選好性の遺伝的な変異が存在することを発見した。次にこれら
4 集団を用い、全きょうだい家系を造成し、餌選好性、捕食者回避行動、顎・翅サ
イズ等の外部形態の解析を行った。それらの形質間の遺伝相関について報告し議論
する。
[P54] 大きなオタマがいるとき、小さなオタマが捕食されやすいのはなぜか?
○山口彩(北大・院・環境科学), 岸田治(北大・北方生物圏 FSC)
個体群内の個体のサイズのばらつきは、食う‐食われる関係の強さを決める要因に
なる。このことは捕食者種のサイズのばらつきに注目することで明らかにされてき
たが、昨年私達は、被食者種のサイズのばらつきもまた重要であることを発見した。
捕食者となるエゾサンショウウオ幼生(サンショ)がいるとき、エゾアカガエルの
オタマジャクシ(オタマ)は小さな個体だけで集団が構成される場合よりも、大き
な個体が集団の中に含まれる場合のほうが、小さな個体がより多く捕食された。今
回私たちは、この「サンショを介した大きなオタマから小さなオタマへの負の間接
効果」が生じるメカニズムについて研究した。私達は、「大きなオタマが活発に動
くため、小さなオタマが刺激されて動き、サンショとの遭遇が増えることで捕食さ
れやすくなる」と考えた。この仮説を検証するために2つの実験を行った。野外の
密度を反映した条件で、サンショと大小オタマの有無を操作し、彼らの活動量と小
46
オタマの生存率を調べた。さらにメカニズムの詳細を明らかにするため、小さいス
ケールで、サンショ1個体と大小オタマ各1個体の有無を操作し、彼らの行動と捕
食の過程の関係を調べた。
[P55] 異質環境下におけるニホンジカの将来分布の予測:セルベースの状態空間モデルに
よる解析
○長田穣(東大・院・生物多様性)、浅田正彦(AMAC)、栗山武夫(東大・院・生物多様
性)、横溝裕行(国環研)、宮下直(東大・院・生物 多様性)
野生動物の効果的な管理を行うためには、将来の個体数分布を予測することが不可
欠である。しかし、今までの研究では異質環境下での移動を十分 に考慮してこな
かったため、将来どのように野生動物が分布拡大していくか予測することは困難で
あった。本発表では、セルベースの状態空間モデ ルを用いることで、個体数の増加
しやすさだけでなく、移動のしやすさが周辺環境によって変化する、生物が周辺環
境の移動のしやすさに従っ て複数の経路を移動することを明示的に考慮した。本発
表では、最新の MCMC 手法を組み込んで、千葉県房総半島におけるニホンジカの
糞粒調 査・区画法調査のデータに考案した統計モデルを適用し、ニホンジカの個体
群増加や移動に関わる環境要因を明らかにした。また、その推定したパ ラメータを
用いて将来の個体数分布予測を行った。
[P56] 寄主植物を共有するホソオチョウとジャコウアゲハの出現タイミングが両種間相互
作用に与える影響
○橋本洸哉、大串隆之(京大・生態研センター)
寄主植物の量すなわち植食者にとっての資源量は、植食者間の相互作用の結果を左
右する重要な要因である。資源量は過去の食害量だけでなく植物の成長にも影響さ
れることから、過去の食害のタイミングが資源量の決定に重要であると予測される。
しかし、食害タイミング・植物の成長・資源量の関係を具体的に調べた研究はほと
んどない。我々は、ウマノスズクサを寄主植物とするホソオチョウとジャコウアゲ
ハの相互作用を調べた。両種幼虫はおおむね交互に発生し、出現の時間間隔は種内
よりも種間で短い傾向にあった。両種の出現時期を模して食害タイミングを操作し
た実験および野外調査により、食害タイミングに関わらずジャコウによる食害を経
験した株では地上部の量が有意に少なくなること、この影響は次世代の幼虫出現時
まで持続することがわかった。これに対して、ホソオの影響は操作実験と野外調査
のいずれでも認められなかった。以上から、資源量の減少によるジャコウの負の効
果は、後から現れるホソオとジャコウ双方にもたらされると考えられた。これらの
結果は、食害タイミングとその影響の強さによっては食害の影響が自種の次世代ま
で残り得ることを示している。
47
List of Attendants 大会参加者名簿
(P??: Poster, Sp? and So?: Plenary and Organized Sympo, StCl: Stat. Clinic, BQ: banquet)
Name
Anazawa, Masahiro
Bjornstad, Ottar
Carrasco, Luis
Cohen, Joel E.
Dorazio, Robert
Ezoe, Hideo
Fasola, Mauro
Fujii, Koichi
Fujita, Go
Fukaya, Keiichi
Hamao, Shoji
Hashimoto, Koya
Hayashi, Masayuki
Hosokawa, Takahiro
Ikegawa, Yusuke
Ishihara, Michihiro
Kagiya, Shinnosuke
Kanamori, Yuki
Kasada, Minoru
Kasai, Atsushi
Kasuya, Eiiti
Katayama, Noboru
Kawakami, Yasuko
Kawata, Shohei
Kawatsu, Kazutaka
Kiritani, Keizi
Kishida, Osamu
Kondoh, Michio
Konuma, Akihiro
Kumagai, Naoki
Kumano, Norikuni
Kuriwada, Takashi
Kuriyama, Takeo
Kyogoku, Daisuke
Masaki, Nao
Mashiko, Miyuki
Minaka, Nobuhiro
Miyashita, Tadashi
Miyatake, Takahisa
Moriya, Seiichi
Nagamitsu, Teruyoshi
Jpn.Name
穴澤 正宏
Affiliation
東北工大・工・環境
PennState
Univ. of Tsukuba
Rockefeller University
SESC
大阪府大・院・理
Pavia Univ., Italy
筑波大
東大・農
ISM
国立科学博物館・動物研究部
京大・生態研センター
千葉大院・園芸・応用昆虫
九州大・理・生物
大阪府大・院・理
大阪府大・院・理
北大・環境科学院
北大・院・環境科学
Univ. of Tokyo
NIES
九大理学動態生物学
北大・FSC
大阪市博・外来研
東邦大学・生物
龍谷大・理工
伊東市
北大・北方圏 FSC
龍谷大・理工
NIAES
国環研
沖縄害虫防セ・琉大農・琉球産経
鹿児島大・教育・生物
東大・院・生物多様性
京都大・理・動物生態
大阪府大・院・理
NIAES
NIAES
東大農生物多様性科学
岡山大
中央農研・病害虫
森林総研・森林遺伝
江副 日出夫
藤井 宏一
藤田 剛
濱尾 章二
橋本 洸哉
林 正幸
細川 貴弘
池川 雄亮
石原 道博
鍵谷 進乃介
金森 由妃
粕谷 英一
片山 昇
河上 康子
川田 尚平
川津 一隆
桐谷 圭治
岸田 治
近藤 倫生
熊谷 直喜
熊野 了州
栗和田 隆
栗山 武夫
京極 大助
正木 奈穂
三中 信宏
宮下 直
宮竹 貴久
守屋 成一
永光 輝義
48
Talk
P18
SpB
So3
SpA
P10
So3
P49
P19
P08
P56
P02
So1
P42
P51
P33
P06
So2
StCl
P48
P32
P28
P12
So2
P47
So1
So1
P39
P20
P26
So3
StCl
P40
P36
BQ
⃝
⃝
⃝
⃝
⃝
⃝
⃝
⃝
⃝
⃝
×
⃝
⃝
⃝
⃝
⃝
⃝
⃝
⃝
⃝
⃝
⃝
×
×
×
⃝
⃝
⃝
×
⃝
⃝
⃝
⃝
⃝
×
⃝
×
⃝
⃝
×
×
Name
Nakamura, Jun
Nelson, William
Niizato, Takayuki
Nishida, Takayoshi
Ohashi, Kazuharu
Ohgushi, Takayuki
Oishi, Tsuyoshi
Okamura, Hiroshi
Oku, Keiko
Onodera, Hirono
Osada, Yutaka
Ovadia, Ofer
Saitoh, Takashi
Sakata, Yuzu
Sasaki, Suguru
Sato, Kazunori
Shefferson, Richard
Shimada, Masakazu
Sugiura, Shinji
Sumi, Takuto
Suzuki, Noriyuki
Takada, Takenori
Takatsu, Kunio
Takimoto, Gaku
Tanaka, Koichi
Tanigawa, Mayuko
Taper, Mark
Toquenaga, Yukihiko
Tsubaki, Yoshitaka
Tsurui, Kaori
Tsutsui, Masaru
Ueno, Yusuke
Ueno, Takatoshi
Utsumi, Shunsuke
Yamaguchi, Aya
Yamamichi, Masato
Yamamura, Kohji
Yamanaka, Miyu
Yamanaka, Takehiko
Yashiro, Toshihisa
Yokoi, Tomoyuki
Yokomizo, Hiroyuki
Yoshida, Katsuhiko
Jpn.Name
中村 純
Affiliation
玉川大
Queen’s University
筑波大・システム情報系
滋賀県立大・環境生態
筑波大・院・生物
京大・生態研
沖縄県病害虫防除技術センター
中央水研
農研機構・中央農研
北大・環境科学院
東大・院・生物多様性
Ben-Gurion University of the Negev
北海道大学フィールド科学センタ
京大・生態研センター
Toho University
静岡大・院・数理システム
東京大学
東大・総合文化・広域科学
Kobe Univ.
岡大・院・環境生命
Tohoku Univ.
北大・地球環境
北大・院・環境科学
Toho University
農環研
PE Editorial Coordinator
Montana State Univ.
Univ. Tsukuba
京大生態研センター
沖縄防技セ琉球産経琉大農
東京大学・院・農
国交省・国総研・緑化生態
Kyushu University
FSC, Hokkaido Univ.
北大・院・環境科学
京大・白眉/生態研
農業環境技術研究所
東大・院・農
NIAES
京大院・農・昆虫生態
筑波大・生命環境系
国立環境研究所
国立環境研・生物
新里 高行
西田 隆義
大橋 一晴
大串 隆之
大石 毅
岡村 寛
奥 圭子
小野寺 裕乃
長田 穣
齊藤 隆
坂田 ゆず
佐藤 一憲
嶋田 正和
角 拓人
高田 壮則
高津 邦夫
田中 幸一
椿 宜高
鶴井 香織
筒井 優
上野 裕介
山口 彩
山道 真人
山村 光司
山中 美優
山中武彦
矢代 敏久
横井 智之
横溝 裕行
吉田 勝彦
49
Talk
So2
SpB
P04
So1
P27
P30
P52
P55
P44
P21
P31
P22
So3
P05
700
P46
P38
SpA
SpA
So1
P50
So2
P41
P54
SpB
SpA
P43
SpB
P16
So2
P01
P29
BQ
×
⃝
×
⃝
×
⃝
×
×
⃝
×
×
×
⃝
×
×
⃝
⃝
⃝
×
×
⃝
⃝
⃝
⃝
×
⃝
⃝
⃝
⃝
⃝
⃝
×
⃝
⃝
⃝
⃝
⃝
⃝
⃝
⃝
⃝
⃝
⃝
Bus Timetables バス時刻表
つくばセンター 6 番のりば Platform No. 6 at Tsukuba Center
行先
Destination
筑波大学中央
Tsukuba-daigaku-chuo
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
30
08
04
08
08
08
08
08
08
03
03
03
08
33
08
08
08
48
22
12
12
23
23
23
23
23
23
23
18
23
25
20
14
43
43
43
43
43
33
33
38
43
47
27 28 35 45 55
23 38 45 53 56
53
53
53
58
平日 Weekday
筑波大学循環(右回り)
Tsukuba-daigaku-junkan
(Migi-mawari)
Loop-line Clockwise
筑波大学循環(左回り)
Tsukuba-daigaku-junkan
(Hidari-mawari)
Loop-line Counterclockwise
00
20
00
00
00
00
00
00
00
00
00
00
00
00
00
00
20
20
00
10
10
10
10
10
10
10
10
10
10
10
10
10
20
00
40
50
20
20
20
20
20
20
20
20
20
20
20
20
20
40
40
40
40
40
40
40
40
40
40
40
40
40
40
40
30
30
30
30
30
30
30
30
30
30
30
30
30
50
50
50
50
50
50
50
50
50
50
50
50
50
40
筑波大学病院行きは、大学会館前まで行きません。Do not ride bus bound for Tsukuba-daigaku-byoin (Univ. Hospital).
行先
Destination
筑波大学中央
Tsukuba-daigaku-chuo
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
33
23
02
07
03
03
03
03
03
03
03
03
03
13
33
38
48
43
08
08
23
23
23
23
23
23
23
43
38
53
23 28 38 40 47 53
23 40 43
43
43
43
43
43
43
43
土日祝日 Weekend
筑波大学循環(右回り)
Tsukuba-daigaku-junkan
(Migi-mawari)
Loop-line Clockwise
筑波大学循環(左回り)
Tsukuba-daigaku-junkan
(Hidari-mawari)
Loop-line Counterclockwise
00
20
00
20
00
20
00
20
00
20
00
20
00
20
00
20
00
20
00
20
00
20
00
20
00
20
00
20
00
20
00
20
00
20
40
40
40
40
40
40
40
40
40
40
40
40
40
40
40
40
40
筑波大学中央行きか、筑波大学循環(右回り)に乗車し、大学会館前で下車ください(約 10 分、200 円)。
筑波大学循環(左回り)でも大学会館前で下車できますが、約 20 分です(200 円)。
Approx. 10 min. (200 JPY) to Daigaku-kaikan-mae by route bus bound for Tsukuba-daigaku-chuo or loop-line
Tsukuba-daigaku-junkan clockwise. Counterclockwise also goes to Daigaku-kaikan-mae but it takes 20 min. (200 JPY).
50
大学会館前バス停 ‘Daigaku-kaikan-mae’
(Univ. Hall Ent.)
Bus Stop
1. 路線バス 左回り方向 Route Bus Hidari-mawari Counterclockwise
行先
Destination
土浦駅西口
Tsuchiura Sta.
経由
Via
つくばセンター
Tsukuba Center
平日 Weekday
筑波大学循環(左回り) ひたち野うしく駅
Tsukuba-daigaku-junkan Hitachinoushiku Sta.
(Hidari-mawari)
筑波大学病院入口
つくばセンター
Univ. Hospital Ent.
Tsukuba Center
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
52
17
17
07
12
12
12
12
12
12
02
07
12
32
37
32
32
38
18
28
08
08
08
08
08
08
08
08
08
08
08
08
08
18
37
32
22
32
32
32
32
32
32
22
27
27
57
52
47
37
52
52
52
52
52
52
37
37
37
52
52
52
52
58
48
28
28
28
28
28
28
28
28
28
28
28
28
38
58
48
48
48
48
48
48
48
48
48
48
48
48
10
20
荒川沖駅西口
Arakawaoki Sta.
つくばセンター
Tsukuba Center
05
15
06
10
筑波大学循環(左回り)
Tsukuba-daigaku-junkan (Hidari-mawari)
Loop-line (Counterclockwise)
Daigaku-kaikan-mae Venue
大学会館前
Tsukuba Center つくばセンター
Tsukuba Sta.
つくば駅
経由
Via
土日祝日 Weekend
土浦駅西口
筑波大学循環(左回り)
Tsuchiura Sta.
Tsukuba-daigaku-junkan
(Hidari-mawari)
つくばセンター
筑波大学病院入口
Tsukuba Center Univ. Hospital Ent.
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
57
17
07
02
12
12
12
12
12
12
12
12
22
42
22
02
02
行先
Destination
47
27
17
32
32
32
32
32
32
32
47
52
47
32 52
52
52
52
52
52
52
52
38
18
38
18
38
18
38
18
38
18
38
18
38
18
38
18
38
58
58
58
58
58
58
58
58
いずれの行先も、つくばセンターで下車できます(約 10 分、200 円)。つくばセンターへは、こ
ちらが早く着きます。
All buses above go to Tsukuba Center (approx. 10 min, 200 JPY).
51
2. 路線バス 右回り方向 Route Bus Migi-mawari Clockwise
行先
Destination
経由
Via
平日 Weekday
筑波大学循環(右回り)
Tsukuba-daigaku-junkan
一ノ矢学生宿舎前
Ichinoya-gakusei-syukusya-mae
土日祝日 Weekend
筑波大学循環(右回り)
Tsukuba-daigaku-junkan
一ノ矢学生宿舎前
Ichinoya-gakusei-syukusya-mae
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
07
27
07
07
07
07
07
07
07
07
07
07
07
07
07
07
27
07
27
07
27
07
27
07
27
07
27
07
27
07
27
07
27
07
47
57
27
27
27
27
27
27
27
27
27
27
27
27
27
47
47
47
47
47
47
47
47
47
47
47
47
47
47
47
47
47
47
47
47
47
47
47
筑波大学循環(右回り)
Tsukuba-daigaku-junkan (Migi-mawari)
Loop-line (Clockwise)
Daigaku-kaikan-mae
大学会館前
Venue
Tsukuba Center つくばセンター
Tsukuba Sta.
つくば駅
右回りでもつくばセンターへ着きますが、左回りよりも時間がかかります(約 20 分、200 円)。
なお、筑波大学中央行きは、つくばセンターまで行きませんので、ご注意ください。
Approx. 20 min. (200 JPY) to Tsukuba Center by clockwise loop-line. Do not ride bus bound
for Tsukuba-daigaku-chuo because it does not go to Tsukuba Center.
52
3. 東京駅行き高速バス Higiway Bus bound for Tokyo Sta.
行先
Destination
経由
平日 Weekday
東京駅(日本橋口)
Tokyo Sta.
都営浅草駅・上野駅
Via
Toei Asakusa Sta, Ueno Sta.
4
5
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
47
17
17
17
17
47
07
17
17
17
17
07
07
07
07
07
07
27
47
47
47
47
47
47
47
47
47
27
47
27
27
27
27
47
土日祝日 Weekend
東京駅(日本橋口)
Tokyo Sta.
土曜:都営浅草駅・上野駅
日曜・祝日:東京駅直行
Sat.: Toei Asakusa Sta, Ueno Sta.
Sun. & Holoday: nonstop
17
17
07
07
07
07
17
17
17
17
17
17
17
17
17
17
17
47
47
47
47
47
47
47
27
27
27
27
47
47
47
47
47
47
47
47
47
47
47
47
47
47
47
Daigaku-kaikan Venue
大学会館
Tsukuba Center つくばセンター
Tsukuba Sta.
53
つくば駅
Taxi タクシー
大曽根タクシー(株)
(有)松見タクシー
フジ急(株)
土浦タクシー(株)
(有)中央タクシー
Ozone Taxi
Matsumi Taxi
Fujikyu
Tsuchiura Taxi
Chuo Taxi
Tel:
Tel:
Tel:
Tel:
Tel:
54
029-864-0301
029-851-1432
0120-0229-03
029-821-5324
0120-39-0184
24H
24H
7:00 - 2:00 (26:00)
7:00 - 2:00 (26:00)
7:00 - 23:30
Program and Abstracts for the 30th Annual Meeting of the Society of Population Ecology
第 30 回個体群生態学会大会 プログラム講演要旨集
Printed on 2014-10-04
Published by the Organizing Committee of the 30th Annual Meeting of
the Society of Population Ecology
Chair: Kohji Yamamura (NIAES) and Yukihiko Toquenaga (Univ. of Tsukuba)
Secretary General: Yukihiko Toquenaga (Univ. of Tsukuba)
Members: Takehiko Yamanaka (NIAES)
Tomoyuki Yokoi (Univ. of Tsukuba)
Katsuhiko Yoshida (NIES)
Kazuharu Ohashi (Univ. of Tsukuba)
Masako Katsuki (Univ. of Tsukuba, PD)
Miyuki Mashiko (NIAES, PD)
Contact: (c/o) Yukihiko Toquenaga
Faculty of Life and Environmental Sciences, University of Tsukuba
1-1-1 Tennoudai, Tsukuba City, Ibaraki 305-8572, JAPAN
E-mail: toque[at]biol.tsukuba.ac.jp
Illustration: Yukihiko Toquenaga (Univ. of Tsukuba)
Website: http://www.pe.ska.life.tsukuba.ac.jp/PES30/
30
th
© Copyright 2026 Paperzz