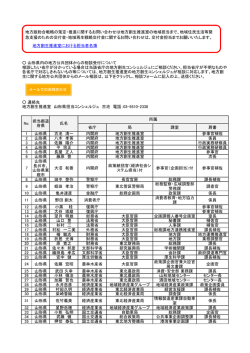
やまがたの 野生動物 山形県立博物館 2013 やまがたの 野生動物 山形県立博物館 2013 2 ︱この冊子について︱ この冊子について この冊子は,全国科学博物館振興財団による平 レッドデータブックカテゴリーとレッドリスト 成25年度科学系博物館活動等促進事業「野生動 環境省レッドデータブックで採用されているカ 物の不思議を知る体験教室」の活動紹介とまとめ テゴリーは1997年に発表され,2006年までに出 の一部をなすものとして刊行した。 版された。哺乳類と鳥類は、「改訂・日本の絶滅 のおそれのある野生生物―レッドデータブック― 「哺乳類」について 1 哺乳類,2 鳥類(環境省 2002a, b)」として 山形県に分布するすべての哺乳類について取り 出版されている。その後,レッドリストはホーム 上げた。それぞれ,形態,生態,分布,分類,絶 ページで公開され、最新版は2012年のものであ 滅危惧などについて概説した。写真は生態写真を る。山形県のレッドデータブックは「レッドデー 中心にしたが,剥製の写真や明治時代の動植物の タブックやまがた 山形県の絶滅のおそれのある 図譜である「両羽博物図譜」からの引用も含まれ 野生動物(山形県 2003)」として発行され,環 る。モグラ類,コウモリ類,ネズミ類については 境省に準ずるカテゴリーである。山形県のレッド 和名と学名だけのものもある。オオカミとカワウ リストは改訂作業中である。 ソなど絶滅種も含んでいる。 国際保護連合(IUCN)レッドリストのカテ 和名,学名等については「日本の哺乳類 改 ゴリーと基準は,2000年2月に、2001年版 訂 第 2 版( 阿 部 ら 2008)」 お よ び「The Wild (Ver.3.1)採択された。最新のレッドリストは Mammals of Japan 第2刷(Ohdachi et al. 2010) 2010年版。軽度懸念を除き,ほぼ環境省の基準 に従い, 「ホームページ Wikipedia」なども参考 と同じである。 にした。種にもとづいた記述を優先し,亜種につ 絶滅(Extinct, EX)既に絶滅したと考えられる種 いては説明中に記載した。 野生絶滅(Extinct in the Wild, EW)飼育・栽培 山形県内の哺乳類に関する話題について5題と 下でのみ存続している種 資料を記述した。講演会で取り上げたものを中心 絶滅危惧I類(Threatened, CR+EN)絶滅の危機に に,それぞれ専門とする分野について研究者から 瀕している種 執筆していただいた。 絶滅危惧IA類(Critically Endangered, CR)ごく 近い将来における野生での絶滅の危険性が極めて 「鳥類」について 高いもの 山形県には、約390種の鳥類が記録されている 絶滅危惧IB類(Endangered, EN)IA類ほどではな が,そのなかから絶滅危惧とされているものを中 いが,近い将来における野生での絶滅の危険性が 心に写真により紹介した。写真は生態写真を主に 高いもの したが,剥製や「両羽博物図譜」からの引用も含 絶滅危惧II類(Vulnerable, VU)絶滅の危険が増 まれる。また,「石沢コレクション」の古い写真 大している種 も一部掲載した。 準絶滅危惧(Near Threatened, NT)存続基盤が 和名,学名等については「日本鳥類目録 第7版 脆弱な種 (日本鳥学会 2012)」に従った。また,「レッ 軽度懸念(Least Concern, LC)IUCNレッドリス ドデータブックやまがた 山形県の絶滅のおそれ トにあるカテゴリー のある野生動物(山形県 2003)」をもとに絶滅 情報不足(Data Deficient, DD)評価するだけの 危惧の高いカテゴリーを掲載した。亜種を基準に 情報が不足している種 取り上げている。 絶滅のおそれのある地域個体群(Threatened 鳥類に関する話題を2題取り上げた。 Local Population, LP)地域的に孤立している個 体群で絶滅のおそれが高いもの 3 写真・図について 環境省(赤),山形県(青)で表記した。環境省 それぞれの写真について,撮影者の氏名または のカテゴリーで山形県と直接関係のない亜種や地 提供団体を掲載した。また,引用がある場合は出 域個体群を含む場合は( )で表した。 典を明記した。撮影者を明記していないものは, 山形県内に近県から侵入しているニホンジカと 山形県立博物館鈴木弘二および編集者による撮影 イノシシについては,県の2003年版カテゴリー や執筆者,協力者の提供写真である。 「絶滅EX」を記載していない。 そのほか、ワシントン条約(CITES)附属書I, 各部の長さについて II, III記載種,国指定特別天然記念物,国指定天 必要に応じ,哺乳類の頭胴長,肩高および尾 然記念物,国際希少野生動植物種,国内希少野生 長,とくにコウモリ類では前腕長,さらに鳥類の 動植物種,県の獣,県の鳥などについても関係の 全長および翼開長を記載した。これらは次の図の 種に明記した。 通りである。 ︱この冊子について︱ レッドリストカテゴリーは国際保護連合(緑), 4 ︱目次・挨拶︱ 目 次 2 この冊子について 4 目次 挨拶 5 哺乳類 6 やまがたの野生動物−哺乳類 伊藤健雄 10 絶滅した仲間 12 サルの仲間 14 吾妻の白猿 縮 文夫 20 イヌ・ネコの仲間 25 ウシの仲間 28 ニホンジカとシノシシの侵入 玉手英利 31 資料 山形県環境エネルギー部みどり自然課 33 ウサギの仲間 34 ネズミの仲間 38 今昔 山形のヤマネ 中村夢奈 42 トガリネズミの仲間 挨 拶 43 コウモリの仲間 やまがたには多くの野生動物が生息していま 44 庄内海岸に漂着するクジラ 長澤一雄 す。カモシカ,ツキノワグマやニホンザルなどは 48 海の哺乳類 近年繁栄し,ひととのかかわりをもつようになっ 49 鳥類 ています。しかし,一般には知られていない野生 50 山形県の野鳥 奥山武夫 動物もたくさんいます。また,ニホンオオカミや 53 鳥海山に舞うイヌワシ 長船裕紀 ニホンカワウソなど絶滅した動物もいます。 59 絶滅危惧種を中心とする鳥類 今年度の「野生動物の不思議を知る体験教室」 73 絶滅した鳥 事業のひとつ,企画展「やまがたの野生動物―け 74 迷鳥 ものととりの多様性―」では県内に生息する哺乳 76 参考文献・協力者および資料提供者 類と鳥類の展示をおこないます。また,野生動物 の不思議を知るための講演会も数回計画していま す。本書はそれらの内容紹介のために発行するこ とになりました。この事業が「やまがたの野生動 物」理解への一助となるよう期待しております。 本書を発行するにあたり,活動助成を受けた全 国科学博物館振興財団をはじめ,資料を提供いた だいた多くの方々に感謝の意を表します。 山形県立博物館長 髙梨博実 表紙 ニホンザル, 裏 表 紙 オ シ ド リ (写真:長船裕紀) 哺乳類 タヌキ (撮影:長船裕紀) 6 ︱ 哺 乳 類 ︱ ︿寄 稿﹀ やまがたの野生動物−哺乳類 伊藤健雄 (山 形 大 学 名 誉 教 授) (1) ツキノワグマ れてきた。従って,7年前の捕獲でその60%近く を失ったことになる。数字だけから見れば,これ は破壊的な痛手と言える。しかし,翌年の目撃件 数はほぼ例年並みで,大きな落ち込みは見られな かった。その後の発表では,本県のクマの数は 1,240頭,最大値で1,500頭と推定されている。 本県では,落葉広葉樹林(ブナ・ミズナラ林) が県土の60%以上を占めているので,クマの生息 環境としては比較的良好な状態にある。クマは広 葉樹の若芽や若葉,ドングリなどの堅果を主な 図1. ツキノワグマ(撮影:東英生) 食物としている。昭和53年の環境庁(当時)の 「クマ異例,出没多発続く県内!」。昨年, 調査では,クマの分布域は県土の55.5%であった 10月31日の山形新聞は,見出しでこう報じた。 が,平成12年には76.8%に拡大している。分布 記事によれば,この年の春以降,異例のペースで 域は,蔵王,吾妻,月山など県内の主要山系に普 クマ(ツキノワグマ)の出没が頻発し,10月15 遍的に広がっている。奥山ではほぼ飽和状態にあ 日現在で目撃は400件を超えたという。だが,県 る一方で,広葉樹の人工林化やナラ枯れによる集 内でのクマの目撃多発はなにもこの年に限ったわ 団枯損などによって,近年平地や集落周辺への出 けではなく,数年前からその傾向は現れていた。 没と定着が起こっている。そのために,突発的な 目撃数が多ければ,当然捕獲数も増える。昭和 接触や農作物被害,さらには人身事故などのクマ 55年以降,県内の年間捕獲数は長い間150頭前 との不適切な関係が増加している。 後で推移してきたが,平成8年から200頭を超え 全国的にみればクマは減少傾向にあり,分布域 るようになり,7年前の平成18年に700頭を回る の孤立化が目立っている。比較的クマが多い本県 という異常な事態に至った。この年は全国的にク でも,いつまでも安泰でいられるかどうかは分か マの捕獲数が多く,本県は僅かの差ながら最多の らない。少なくとも無計画な取り放題だけは避け 長野に次いで2位,1位から10位までに宮城県を なければならない。クマの未来を祈りたい。 除く東北5県がランクされた。 本県にはどのくらいの数のクマが住んでいるの (2) ニホンザル だろうか。全県のクマの数を推定するのは容易な 私たち日本人は,日本にサルが住んでいるのを ことではない。それは,大人のクマが単独生活者 当たり前と思い,何の不思議にも感じないが,世 であること,行動範囲が広く,一日に十数キロ 界的に見ればこれは極めて珍しい現象なのであ メートルも移動すること,出産数,齢別死亡数, る。サル(ニホンザル)が属する霊長類の仲間 寿命などの繁殖指標に関するデータが不足してい は,本来熱帯から亜熱帯地域に住む動物である。 ることなどがその理由であるが,何よりもクマ 霊長類の世界分布を見ると,赤道を挟んだ北緯 の頭数調査法が確立していないのが致命的であ 30度から南緯30度の範囲にそのほとんどが集中 る。それでも本県では,昭和52年以降 巻き狩り している。この範囲より北に位置する日本列島 追い出し法 とでも言うべき方法で毎年調査が継 のサルは,本来の生息地からの はみ出し者 であ 続され,生息数は1,200頭から1,300頭と推定さ る。とりわけ東北地方には「世界最北限のサル生 はいない)。環境省のレッドデータブックでは, 害者としてのサルである。県内での農作物被害は 「東北地方のニホンザル個体群」は「絶滅の恐れ 当初水稲や桑,クリ,ブドウなどにみられたが, のある地域個体群」にランクされている。 その後サクランボ,リンゴ,ラフランスなどの価 格の高い果樹類に集中し,さらに畑作物へ拡大し た。平成13年以降は,被害金額で年間2億円前後 という状態が続いている。県では昭和60年から サルの有害駆除捕獲を実施している。平成8年か ら捕獲数は急増し,近年は年間400∼500頭で推 移しており,全国有数の サル捕獲県 という有り 難くない評価を受けている。被害の甚大な地域で は,加害群のサルに電波発信機を装着し,集落同 士でその動きを追跡しながら農耕地からの追い払 図2. ニホンザル(撮影:長船裕紀) いに役立てている。 日本列島でのサルの主な生息環境は,西日本に 傍若無人に暴れまわるサルがいる一方で,真室 優占する常緑広葉樹林(シイ・カシ林)であり, 川町の及位(のぞき)個体群のように,ひっそり この森林でのサルは広く分布する。一方,東北地 と消滅して行くサルたちがいる。いくつかの地方 方の落葉広葉樹林では,寒冷多雪という厳しい自 では分布域の拡大や被害の増大が認められるもの 然条件が故に分布が制限されており,生息数も多 の,全国的に見れば分布域の縮小や群れの分断に くない(北海道と琉球諸島には分布しない)。分 よる孤立化が起こっており,地域的絶滅が進行し 布が制限された原因は,一つはブナ・ミズナラ林 ている所も少なくない。 の伐採と人工林化による生息地の破壊,もう一つ は高性能の銃の使用による乱獲にあったと言われ (3) カモシカ ている。 一時極端に減少した本県のサルは,狩猟圧から 解放されると徐々に回復し,昭和30 40年代から 増加に転じた。現在,蔵王連峰から吾妻連峰にわ たる奥羽山系に広く分布する個体群,飯豊連峰の 玉川・荒川流域に分布する個体群,朝日連峰の大 鳥川流域,麻耶山地域に分布する個体群など,6 地域個群,100群前後,約3,000頭が生息してい ると推定されているが,調査が十分に及んでいな い地域もあり実態はよく分かっていない。 図3. カモシカ(撮影:長船裕紀) 西吾妻スカイバレー付近では,時折白い体色の カモシカは現在,国の「特別天然記念物」に指 サルが目撃される。俗に言う白子(アルビノ)と 定されており,法律上「国宝」級の扱いを受けて 当初は考えられていたが,よく見ると瞳孔は赤く いる。文化財保護法でいう「特別天然記念物」は ない。遺伝学的な説明では,白い体色は伴性遺伝 「天然記念物のうち世界的にまたは国家的に価値 の一型である限雌性遺伝によるもので,この性質 が高いもの」とされているが,カモシカはなぜ国 はメスだけに現れる。劣性遺伝子をもつ性染色体 宝級の動物なのだろうか。 が対に揃ったときにだけ発現するところから,白 その理由の一つは,指定当時(昭和30年), い個体は早死にするものが多く,成獣にはなりに カモシカは絶滅の危機に瀕しており,法的な強い くいと言われている。 保護を必要としていたこと,二つは生物地理学上 7 ︱ 哺 乳 類 ︱ ︿寄 稿﹀ 最近報道などで話題に上がるのは,農作物の加 息地」という特異性がある (北海道に野生サル 8 ︱ 哺 乳 類 ︱ ︿寄 稿﹀ の遺存種( 生きた化石 )として,有蹄類の系統 直接目撃による観察で10∼30ha,最大でも50ha 進化の中にあって 原型 を留めた価値を有してい と言われていた。しかし山形市西蔵王で行われ ることである。指定後半世紀以上が過ぎた現在, たラジオテレメトリー法による調査では,90∼ 特に前者の状況は大きく様変わりしている。 150haという従来言われていたもののほぼ10倍 カモシカは本県を象徴する動物として,昭和 の広さで,最大500haに達する例もあった。行動 57年「県獣」に指定された。候補の3種(カモシ 圏の中心部には,日常的に極めて高い頻度で利用 カ,ニホンザル,リス)を対象に葉書による県民 されるコアエリア(広さ10ha程度)がある。コ 投票が行われ,投票数1,284票の76.9%をカモシ アエリアはなわばりの占有者によって守られ,隣 カが獲得し当選した。カモシカを県獣に指定して 接個体のそれと重複することはない。 いる県は,山形を含めて栃木,富山,山梨など6 カモシカとシカ(ニホンジカ)が重複して生息 県にのぼる。県民から広く愛され,和と協調に基 する国内の多くの地域では,単独生活者のカモシ づく絆としての県獣だったが,カモシカを取り巻 カと群れ生活者のシカとの間で,生活様式に依存 く環境は山形でも必ずしも安泰ではなかった。 した種間競争が起こり,カモシカは敗退する傾向 カモシカは,クマとともに落葉広葉樹林を代表 が認められている。県内では今のところシカは定 する大型哺乳類である。分布の中心は東北地方か 住していないので,このような現象は知られてい ら中部地方にかけた山岳地帯で,東日本に偏って ないが,今後のシカの動向いかんによっては,近 いる。四国と九州の分布域はごく限られており, い将来同様の現象が起こる可能性はある。 中国地方,北海道,南西諸島には生息しない。か 昭和54年,文化庁,環境庁(当時),林野庁 つて 幻の動物 と呼ばれた時代には高山獣と考え の3庁がカモシカの保護及び被害対策についての られていたが,個体数の増加に伴い現在では山岳 覚書を発表した(いわゆる「3庁合意」)。その 地帯ばかりでなく,海岸部から低山帯や平地に至 骨子は,①カモシカの保護地域を指定し,生息環 るまで普通に生息する身近な動物となった。環境 境とともに保護を図る,②被害防止に努めるが, 庁による昭和58年の調査では,全国の推定個体 保護地域外の地域では状況に応じて個体数調整 数は9,900頭ないし10,200頭であったが,最近の (捕獲)を行う,の2点である。この覚書に基づ 全国規模での調査資料はない。 いて,文化庁は全国のカモシカ生息地に15箇所 県内では,蔵王連峰を含む奥羽山系と朝日・ の保護地域を指定した。そのうち本県にかかるの 飯豊・吾妻の各山系に,連続した広い分布域が は「南奥羽カモシカ保護地域」と「朝日・飯豊カ ある。これらのうち,奥羽山系の標高200mから モシカ保護地域」の2箇所である。各保護地域を 900mにかけた山地帯に,比較的密度の高い地域 対象に,毎年の「通常調査」と5年毎の「特別調 が認められる。かつてこの地帯の生息数はおよそ 査」が実施され,生息状況のモニタリングが行わ 2,000頭,うち山形市域だけで600頭と推定され れている。 たが,生息に関する最近の調査はなく,動向は分 カモシカの個体数が,全国的に回復するに伴っ かっていない。ただ山形市の蔵王山麓周辺地域で て,いくつかの地方では農林業に対する被害問題 は,平成10年頃に比べてここ数年カモシカの目 が深刻化してきた。「3庁合意」に基づく食害防 撃件数が減少している。恐らくこの山系では,低 除のための捕獲は,最初岐阜・長野の両県で林業 密度安定状態に移行しつつあるものと考えられ 被害対策として始まったが,その後愛知,山形, る。 静岡,群馬,岩手へと波及した。山形(山形市) 森林中でカモシカの成獣オスは,なわばりを構 では畑作物・果樹被害への対応として,平成2年 えて生活している。なわばりはオス同士の防衛範 から10年までの9年間に304頭を捕獲し,その後 囲で,そこからメスは必ずしも排除されない。な は休止している。捕獲が始まった昭和50年から わばりを含むカモシカの行動圏は,それぞれ重複 平成22年までの36年間に,全国で捕獲されたカ することなく独立に配置され,その面積は従来の モシカの総数は32,968頭に達している。 は多い。 幻の動物 であった時代,カモシカは日 これに対して,カモシカとクマは西側優勢であ 本の野生動物の中で最も研究資料の少ない,謎に る。毛足の長い密生した冬毛と大きく広がる蹄 満ちた動物であったが,今や物言わぬ死体の発言 が,深い雪上でのカモシカの生活を可能にした。 によって,最も研究の進んだ動物になった。 またクマは,大型獣としては珍しい 冬眠 (正確 には冬季の睡眠)という戦略を採用して,厳しい (4) 哺乳類からみた山形 冬に適応した。ちなみに,県内の哺乳類で冬眠す 森は哺乳類の宝庫 と言われるように,森を住 るものには,クマの他にコウモリ類やネズミ類の 処とする哺乳類は多い。森林が県土の70%を超え 数種,ヤマネ,アナグマなどがある。 る本県は 哺乳類の楽園 と言えそうであるが,現 近年,県内のイノシシ事情が変わってきた。東 実には深刻な問題が少なからず存在する。「山形 北地方では,これまで福島県阿武隈山地だけに限 の自然マップ」によれば,この地に暮らす哺乳類 られていた分布域が西側に広がり,さらに北上し の種類は45種(7目17科37属)。しかし,コウ て宮城県南部に侵入した。10年ほど前,仙台市 モリやネズミの仲間などあまり調査が進んでいな の南西に位置する太白山で,イノシシが目撃され い分類群もあるので,実際には50種を超えるも たという新聞報道があった。恐らくその一部が数 のと思われる。 年後に奥羽山脈を越えて本県に入ってきたのであ 45種という数は,日本全国の哺乳類の種類数 ろう。子どものイノシシを見たという情報や農作 と比べてどうなのだろう。最新の資料では,クジ 物への被害報告もあり,どうやらこの頃に県内で ラ類などの海生哺乳類41種を除く土着哺乳類は の定住化が始まったとみられる。 7目23科57属110種,外来種を加えると31科72 一方シカは,これまで分布がなかった福島県 属134種である。このうち,北海道と南西諸島だ で,群馬県から尾瀬地域へ侵入し,特別天然記念 けに住む種を除くと,本州,四国,九州の陸生種 物「尾瀬」の湿原植物を撹乱していることが確認 は75種で,本県の種類数はその約60%に相当す されており,この 尾瀬のシカ がしだいに県西部 る。これを多いとみるか,少ないとみるかは,見 地域を北上して,本県に接近するという動きを見 解の分かれるところであろう。 せている。また,広大な分布域である岩手県南部 の五葉山地域から,シカが拡大進行中という情報 もある。県内では,4・5年前から内陸盆地や鶴 岡市周辺で,いずれも単独個体の目撃例が10件 ほど得られている。今のところ県内での群れの目 撃例はない。 イノシシやシカの生存を脅かした天敵や熟練の 漁師が存在した往時と異なり,現在の自然環境に は,彼らの勢力拡大をくい止めるパワーはない。 図4. ニホンジカ(撮影:長船裕紀) 一度絶滅した 往時の住人 が戻ってくるのを,私 本県の哺乳類相を東北地方の他の5県と比べて たちは歓迎すべきか,排斥すべきか,選択を求め みると,全体としては大きな違いはない。しかし, られる時期がすぐ目前に迫っている。 奥羽山脈の東側と西側では明らかに違っている。 大方の哺乳類は,本県の豊かな自然林の中で, 福島,宮城,岩手に生息するイノシシとシカは, あるいは改変された環境の中で,健気に生きてい 本県にはいない(いなかった)。両種とも昔から る。彼らをことさら保護もせず,さりとて迫害も いなかったわけではなく,イノシシは明治末期に せず,お互いがさりげなく意のままに暮らしてい シカは大正8年にそれぞれ最後の記録を残して絶 ける,そんな共存関係が成り立つことを望んでい 滅した。日本海側に特有の冬期の深い積雪と強い る。 9 ︱ 哺 乳 類 ︱ ︿寄 稿﹀ 狩猟圧が,絶滅の主な原因と考えられている。 死体は物を言わないが,死体から得られる情報 10 絶滅した仲間 ︱哺乳類︱ (食肉目イヌ科) オオカミ Temminck, 1839とエゾオオカミCanis lupus Canis lupus (Linnaeus, 1758) hattai Kishida, 1931は亜種として区別されてい LC EX EX CITESI る。 図1-1. ニホンオオカミ(国立科学博物館) 図1-2. タイリクオオカミ(ニホンオオカミの別亜種) 形態 亜種のニホンオオカミは,頭胴長95∼ 絶滅危惧等 山形県,環境省ともに絶滅。国際保 114cm,尾長30cm,体重15kg。中型の日本犬 護連合では種として軽度懸念に指定している。ワ くらいの大きさ。体毛は灰褐色から淡黄褐色で背 シントン条約附属書Iに掲載されている。 面の毛には黒色が混じる。頭骨は短小で,吻部は 家畜の害獣としての駆除がおこなわれたことも 短く広い。北海道産亜種のエゾオオカミより小型 影響するが,明治以降にヨーロッパから輸入され である。イヌの頭骨とは,吻部の凹部が小さいこ た洋犬から感染したジステンバーや狂犬病が絶滅 と,裂肉歯が大きいことで区別される。 の直接の原因らしい。 生態 大陸に現存しているオオカミと同様,シカ, そ の 他 「 両 羽 博 物 図 譜 ( 松 森 胤 保 ) 」 に は カモシカ,イノシシ,ノウサギ,ノネズミなどを ヤマイヌとして4つの図が掲載されている。図 集団で捕食して,森林生態系における植食性動物 から判断して一部は野犬と考えられているが, の個体数調整者としての役割を果たしていた。 「明治14年3月8日鶴岡ニ於テ死タル賣物ヲ見ル 分布 【日本】絶滅した。北海道に亜種エゾオオ 所ナリ」との記述がある図(図1-3)などはオ カミ,本州,四国,九州に亜種ニホンオオカミ オカミとみられる。「山形縣の哺乳類(橋本, が分布していた。エゾオオカミは1896年までに 1939)」によれば,山形県ではヤマイヌとして 絶滅したと推定されている。ニホンオオカミは 明治初年頃に鳥海山や吾妻山に相当生息していた 1905年に奈良県鷲家口で捕獲され,アメリカの と記録されている。 動物学者アンダーソンによって収集された若い雄 剥製としては,国内に国立科学博物館,東京大 が最後の個体になったといわれている。1910年 学農学部,和歌山県立博物館にそれぞれ1頭が残 に福井県で捕獲されたものを最後とする見方もあ されているにすぎな る。【世界】基亜種タイリクオオカミ(図1-2) い。そのほか,頭骨 はユーラシア中部,北部,北アメリカに広く分布 や毛皮が関東西部, する。家畜の害獣として駆除されたが,最近はヨ 中国地方の神社に保 ーロッパや北アメリカでは国立公園などに再導入 管されている。 されている。 分類 種としては北半球北部に広く分布する 図1-3. 山犬 が,ニホンオオカミCanis lupus hodophilax (両羽博物図譜より) (食肉目イタチ科) カワウソ 載されている。 ︱哺乳類︱ NT EX EX 特別天然記念物 CITESI 図1-4. カワウソ(斎藤報恩会博物館) 形態 頭胴長65∼82cm,尾長30∼49cm,体重 5∼11kg。全長1mをこえる大型獣。頭部と頸部 は同じ太さで円筒形の胴に移行し,四肢は短く幅 図1-5. ニホンカワウソ絶滅(読売新聞,2012.8.29) が広い。指の間には水かきがある。尾は太く円錐 大正期には全国で年間1,000頭以上の捕獲記録 形をし,先半分は上下に扁平。頭部は平たくて丸 があったが,乱獲と生息地の河川環境の変化によ く,目は頂上にある。背面はややくすんだ褐色 り分布域が縮小した。高知県が最後の生息地とし で,胸から腹側は白っぽい毛色をしている。 て1979年まで確認されているが,その後30年を 生態 河川の中下流部から沿岸部にかけて生息す 経過し絶滅と判断された。標本は四国などの博物 る。薄暮から夜にわたって活動し,水中で魚類や 館に多数残されている。 甲殻類,陸上でノネズミや鳥類などを捕食する。 山形県内での過去の生息記録は極めて少ない。 川岸に複数の巣穴をもち,それらを巡回しながら 「両羽博物図譜(松森胤保)」にカワウソの図が 生活する。巣穴は休憩場所や出産場所として使わ あり(図1-6),これが確実な記録である。「山 れる。 形縣の哺乳類(橋本,1939)」ではすでに絶滅 分布 【日本】かつて北海道,本州,四国,九州 と判断している。西川町大井沢では大正の末ころ などに広く分布していたが,絶滅と判断されてい には見られたという「朝日連峰の狩人(志田・西 る。【世界】イギリス,ヨーロッパから東南アジ 沢,1991)」。山形県で最後の状況は不明であ ア,中国まで,シベリアを除きユーラシアの河川 る。標本等も確かなものは残っていない。 沿いに広く分布する。 分類 種としてはユーラシアに広く分布するが, 日本産のカワウソを亜種ニホンカワウソLutra lutra nippon Imaizumi & Yoshiyuki, 1989と分類 している。また,本州以南にみられるものを種と する考え方もある。種および亜種についての分類 は明確ではない。 絶滅危惧等 山形県は絶滅。環境省はそれまでの 絶滅危惧1A類から新たに絶滅とした。国指定特 別天然記念物とされているが絶滅種として今後の 扱いは不明。国際保護連合では種として準絶滅危 惧に指定している。ワシントン条約附属書Iに掲 11 Lutra lutra (Linnaeus, 1758) 図1-6. 獺(両羽博物図譜より) 12 サルの仲間 (霊長類) ︱哺乳類︱ (オナガザル科) ニホンザル 布する。日本固有種。 Macaca fuscata (Blyth, 1875) LC(LP)CITESⅡ 図1-7. ニホンザル(米沢市小野川,撮影:東英生) 形態 頭胴長雄53∼60cm, 雌47∼55cm,尾長 雄8∼12cm, 雌7∼10cm,体重雄6∼18kg, 雌6∼ 14kg。毛の色は茶褐色または灰褐色で,腹と四 群れ 肢の内側がやや白い。顔と尻は裸出して赤い。 群れ以外 隣接県の群れ 生態 常緑広葉樹林や落葉広葉樹林にすむ。数頭 の雄成体および雌成体とその子どもたちからなる 図1-8. 山形県と近県のニホンザルの分布(第2期山形県ニホンザル保 十数頭から百数十頭までの群れで生活する。群れ 護管理計画より) の行動圏は1km2から80 km2と幅がある。昼行性 で,樹上および地上で活動する。 雑食性で,果実,種子,葉,芽,昆虫その他の 小動物を食べる。植物がおもな食物となってい る。また,いろいろな農作物を食害することがあ り,各地で問題になっている。農作物被害の防止 を目的とした有害駆除がおこなわれている。 交尾期は秋から冬で,出産期は春から夏であ る。雄は6∼8歳で社会的な成熟に達する。さら に,成長するにしたがい自分の生まれた群れを離 れ,10∼11歳までに完全にいなくなる。雌は基 本的に生まれた群れにとどまり,5∼7歳で初産 を経験する。通常2∼3年に1回の割合で,一度に 普通1頭の子を産む。大部分の個体は25歳までに 死亡する。 群れ 群れ以外 分布 【山形県】奥羽山系,吾妻山系,朝日山 系,飯豊連峰などに分布するが南部に多い(図 1-8)。【日本】本州,四国,九州,金華山,淡 図1-9. 1978年の山形県内ニホンザルの分布(第2期山形県ニホンザル 路島,小豆島,屋久島。東北地方では断続的に分 保護管理計画より) その他 吾妻山系には毛が白色となった白猿が確 た。 図1-10. ニホンザルの群数と推定個体数(第2期山形県ニホンザル保 護管理計画より) 絶滅危惧等 山形県のレッドリストの指定はな い。環境省では種としての指定はないが,北奥 羽,北上山系,金華山のものは絶滅のおそれのあ 図1-12. 吾妻の白猿(撮影:遠藤亨) る個体群に指定されている。 30年前ころまでは山形県内の分布が限られて 国際保護連合では軽度懸念に指定している。ま いた(図1-9)。近年は村山地方などで分布域を た,ワシントン条約附属書IIに指定され国際間で 広げている。農作物に被害をおよぼす動物として の取引が規制されている。 問題になっている。有害獣として捕獲される個体 もある(表1-1)。 平成25年3月末には子ザル3頭が山形市内の小 学校にあらわれ,そのうち2頭が捕獲され山に帰 された(図1-13)。 図1-11. ニホンザル雄(小国町大滝,撮影:東英生) 図1-13. 学校にあらわれた子ザル(山形市立南山形小学校) ⾲┴ෆ䛻䛚䛡䜛䝙䝩䞁䝄䝹ᤕ⋓ᩘ䛾᥎⛣䠄➨2ᮇᒣᙧ┴䝙䝩䞁䝄䝹ಖㆤ⟶⌮ィ⏬䜘䜚䠅 ᖺᗘ 㡯┠ ᭷ᐖᤕ⋓ H12 321 H13 564 H14 423 H15 576 H16 572 H17 470 H18 704 ಶయᩘㄪᩚ ᤕ⋓ィ 321 564 423 576 572 470 704 H19 H20 H21 H22 ༢䠖㢌 H23 12᭶ᮎ 290 181 108 67 2 33 180 261 462 357 323 361 369 529 359 ︱哺乳類︱ (図1-12)。吾妻の白猿については別に記述し 13 認され,米沢市の天然記念物に指定されている 14 吾妻の白猿 ︱ 哺 乳 類 ︱ ︿寄 稿﹀ 縮 文夫 (白 猿 会) 吾妻山での白猿発見 昭和13年3月米沢市南原在住の二人のハンター (藤田三郎氏・小林安寿氏)により,西吾妻山中 で白猿を発見捕獲,南原で1か月半ほど飼育した 後,上野動物園に引き取られ大変な話題になりま した。噂を聞いた作家の戸川幸夫の名作「吾妻 の白猿神」のモデルにもなったが,上野動物園 では環境に順応出来なかったせいか一年程で亡 くなりました。可哀そうな事をしたと藤田三郎 氏。・・・ その後白猿の話題が途切れたが,昭和49年 図2. まいちゃん 秋,西吾妻山にきのこ採りに入った人が偶然にも 白猿に遭遇。その話を聞いた,市内の歯科医故遠 藤亨氏の耳に入り,白猿の探索が始まりました。 昭和50年に3歳位のメス猿「はつこ」から52年 に3歳位の「ふうこ」,52年に生まれたばかりの 「みつちゃん」,54年に生まれたばかりの「よ つちゃん」まで4頭の白猿を発見しています。こ の4頭の白猿は同じ群れの中に生まれました。 図3. はくてん 図1. 上野動物園に行った白猿と捕獲した藤田三郎氏 平成の白猿誕生 その後白猿の発見は途切れたが,平成3年7月 に西吾妻スカイバレーでアマチュアカメラマンの 図4. ゆいちゃん 高橋勉氏が,白猿よつちゃんの群れに遭遇,その 群れの中に生まれたばかりの白の子猿を発見。そ そのメンバーで白猿会を結成し白猿の観察を続 の話を聞いた仲間のスカイバレー通いが始まる。 けています。吾妻の白猿は昭和58年に米沢市の 物園に行った白猿を除き,23頭の白猿が西吾妻山 白猿は大きな猿の群れの中で一緒に行動してお 一帯に生まれています。西吾妻山には全部で13群 り,米沢で白猿が確認されるのは,関根・南原・ ぐらい居る中の7群れの中に1∼2頭の白猿が入って 大平集落・白布温泉・天元台・スカイバレー・綱 います(表1,図5)。ただ3,4年たつと白猿が見 木・簗沢・小野川・八谷等の群れの中に1∼2頭 えなくなります。長生き出来ないのかもしれませ ずつ入っています。群れには行動範囲があり,通 ん。でも,白猿「よつちゃん」は子供を5匹ほど生 常それ以外には行かないが,たまに群れと群れが み16年も生きていたが,見えなくなりました。猿 重なる場所もあります。また,吾妻山の曲がり竹 の年齢は人間の約4倍と言われています。4,5歳で のシーズン,6月頃には各方面の殆どの群れが竹 成猿,亡くなるまで妊娠可能と言われています。交 の子を食べに,山に上がるようです(スカイバレ 尾期は10月から12月位まで。出産期は4月頃から7 ーで出会える)。冬期間は,標高1,000m以上に 月の始め位まで,吾妻の白猿は平成3年から平成24 は上がっていないようです。 年まで立て続けに生まれ発見されています。 平成25年5月現在確認出来ている白猿は(白天, 初めて白猿に巡り合う チッチ,結)の3頭のみです。 私が初めて白猿に出会ったのは昭和54年3月 頃,今の綱木川ダムが出来る前の烏川地区でし 猿の習性・生態 た。日曜日の晴れた日,雪景色の撮影に出かけた 白猿は毛が白いのは勿論,顔・手・足と肌の見え ところ,田んぼの雪原を移動している2匹の大き る所はピンク色で目はブルーです。白猿の寿命は普 な白猿(ふうこ,みつちゃん)に遭遇しました。 通の猿に比べて短い様な気がします。ただ昭和54 大変感激した事を覚えています。その姿の虜にな 年に発見された,よつちゃん(メス)だけは16年 り白猿を追いかけ始めることになりました。 も生きて5頭の子猿を生んでいます。猿の年齢は人 間の4倍と言われている(よつちゃんの場合人間で 白猿発見と名称の由来 言うと64歳ぐらいまで生存)。 これまで確認された白猿は24頭だが,昭和13 猿は4,5歳から出産でき,上手くいって2年ごと 年に上野動物園に引き取られた白猿には名前がな で,4月∼7月初めぐらいまでが出産期です。たま く,昭和50年以降に発見された白猿23頭に名前 に,双子の子ザルを生むことがあります。 を付けられる様になりました。 10月∼12月の交尾期以外,オス猿とメス猿は別 ○昭和3年(1928年)から平成24年(2012年) 行動をとっており,通常群れ(30頭から120頭) までの白猿誕生記録一覧(表2) として見かけるのは,メス猿の集団です。本当のボ 白猿「はつこ」から「よつちゃん」までは,遠 ス猿はメス猿で,群れの中に居るオスのボス猿と言 藤亨氏が名付けた名前。 われています(オッポをピンと上げているのですぐ 「ふくちゃん」から「はくてん」までは白猿会 分かる)。猿は一時任されているだけで,餌場の確 で付けた名前。「チッチ,まゆ,のぞみ,ゆいち 認や群れの秩序維持が出来なくなると交代させられ ゃん」の4頭の呼名は市民からの公募により選ば る。だから必死になり群れを引っ張っています。次 れた名前です。 の餌場に行く為には,場所を確認してから移動する 名前の由来として,最初の4頭は,「ひい,ふ 為,1∼2日前に偵察役の猿を出しています。 う,みい,よつ」と数の数え方。次の「ふくちゃ あの広い山の中で何処の栗の実が食べ頃か等,殆 ん,すけちゃん」は,高橋氏の床屋さん福助の名 ど分かって居るようで,毎年同じ時期に現れるよう 前から取り,次の「むつちゃん」は6番目に生ま です。 れて来たから。「あいちゃん,たかちゃん,なお ちゃん」は,上杉家の武将にちなんで名付けら れ,「ホクロちゃん」は顔と肩に3つのホクロが 15 ︱ 哺 乳 類 ︱ ︿寄 稿﹀ 生息している場所(範囲) 「天然記念物」に指定されました。現在まで上野動 16 ︱ 哺 乳 類 ︱ ︿寄 稿﹀ 図 5. 吾 妻 山 系 に お け る ニ ホ ン ザ ル の 分 布 (赤 字 は 白 猿) 17 ︱ 哺 乳 類 ︱ ︿寄 稿﹀ 有り,「れいちゃん」は山の峰(嶺)で発見され 員会文化課に報告しています。 たことから,「まいちゃん」は「れいちゃん」と <白猿観察活動の展望> 姉妹だから,「さとみちゃん」は里でも見られる 長年の観察により,各群れの行動範囲が分かっ 事から名付けられ,「かぶとちゃん」は兜山の麓 てきたので観察がしやすくなったが,なにせ野生 で,「いくちゃん」は平成19年に発見されたた の猿達なので裏をかかれる事ばかり。それでも, め,「あきちゃん」は発見者の名前で,「はくて 白猿の群れに逢えた時の感動は変わりません。 ん」は白布温泉の「白」と天元台の「天」から取 りました。 観察を通して気づいたこと 観察活動を始めた頃は,猿に会う事自体が難し 白猿会について く,猿による農作物の被害等は殆ど無かったが, <会発足の経緯> 最近7,8年前頃から人里近くまで下りて来て, 平成3年10月に設立されました。平成3年7月 農作物に悪さをする様になりました。 に,新たに生まれた白い子猿が発見され,毎朝ス 農家の方達は大変困って,電気柵や,花火等で カイバレーに上がって白猿を観察していた仲間で 追い払っていたが,あまり効果がない状態のよう 情報交換する目的で作った会です。 です。最近では犬(モンキードッグ)を導入して <活動内容> 追い出しが行われるようになり,これが最も効果 吾妻の白猿は昭和58年に米沢市の天然記念物 的な良い方法のようです。(昔は犬が放し飼いだ に指定されたため,行動範囲,生息頭数等の現状 った事を思い出してみてください。)猿に対して について,写真や動画に記録して米沢市の教育委 ここは怖い処,危険な処と学習させる事が出来れ 18 ︱ 哺 乳 類 ︱ ︿寄 稿﹀ ば,効き目が大きいと思う。 ―夏― <猿の行動について> シロザ,アカザ,オオバコ,イヌビュウ,ツユク •親猿から子猿に手渡しでエサを与えず,全て自 サ,クローバー,桑の実,モミジイチゴ,草イチ 分で探して自分で食べている。 ゴ,ヘビイチゴ。 •猿の行動,移動は,ボスの合図により動いてい ―秋― る。川等を渡る時は同じ所を渡らせる。安全のた おもに木の実(クリ,ブドウ,アケビ,ブナの め。 実,サルナシ,マタタビ,ヤマボウシ)等。 •食事の後には休憩時間有り。子猿が騒げば,親 ―冬― 猿が手で押さえ休ませる。後々の移動のため。 木の実の残り,樹皮(とくにクワ,柳),新芽 •寒い夜寝る時等は,5,6匹枝に一列になり休 等,クローバー等は一年中食べているようだ。湯 み,夜中に内側と外側が交代する。その繰り返 花,セメント,土等,冬期間は道路に散布した融 し。 雪剤の塩分。 •次に行く場所の下見に,様子見の猿を1,2日前 <猿に威嚇され囲まれた時> に出している様子。食べ物が有るか,安全か。 •こんな時は,逆らわず,しらんふりをして,ご •食べ物がたくさんあっても全部食べていかな く普通に歩く様にして離れる方が良い。 い。次回に回ってきた時に食べる。 •逆らえば,大勢の猿に囲まれる。とくに長い棒 <一般に知られていない猿の行動> 等は絶対に持たない事。 •猿の本当のボスは,メス猿である。オス猿は一 •目と目を合わせない。大声を出さない。とくに 時任されているだけ。役に立たなくなると交代。 子猿等にはいたずらをしない事。 •春に,オタマジャクシ等を食べていた。川原の 石下にいる虫等も食べる。蛋白源か。 西吾妻山の猿には餌付けをしない。今後も絶対 •夏に,毒草のトリカブトの花になる前のツボ に餌付けをしないつもりです。餌付けをすると, ミのところを食べていた。1∼2頭。猿の漢方薬 生態系が崩れ,奇形,野性味が無くなり人間に悪 か。 さをする様になる。自然に任せておいた方が良 •子猿が餌等を食べる時は,親猿から手渡しには い。 貰えない。全て自分で拾わなければならない。自 今思っていることは,猿が農作物の畑に入り込 立。 んで作物を食い荒らしている事。悪さをしような •猿の交尾期は,10月から12月頃まで。この時期 どと思って来る訳ではないが,結果として悪い事 に離れていたオス猿の群れが各群れに近づく。 をしています。それで,猿害の名のもとに駆除さ •猿のお産は,4月から7月の初め頃まで。お腹に れている状態です。やはり山中の食べ物より, 居るのは6か月ぐらい。 里の食べ物,とくに畑の作物はたやすく取れ美 •猿の群れは,メス猿が主体である。子供のオス 味しいので,山に帰る事を忘れてしまっていま 猿と。交尾期を除いては。 す。・・・原因は色々あると思うが,いろんな動 <季節ごとの猿の食べ物> 物(クマ,カモシカ,ハクビシン,タヌキ,イタ ―春― チ,2,3年前からイノシシ)達が里の人間社会 ふきのとう,ツクシンボ,スイパ,イタドリ,ミ に近づいてきています。 ツバ,シシウド,クコの若芽,草ソテツ,イヌド ウナ,ウワバミソウ,モミジガサ,アケビの新 芽,ウド,コシアブラの芽,オオバギボウシ,根 曲り竹の子,ギョウジャニンニク,シオデ,ワサ ビ等。 こんな姿をいつまでも,次に写真を掲載します。 19 ︱ 哺 乳 類 ︱ ︿寄 稿﹀ 図6. れい ち ゃ ん 図10. ま い ち ゃ ん 図7. れい ち ゃ ん 図11. は く て ん 図8. さ つ き ち ゃ ん 図12. ゆ い ち ゃ ん 図9. さ つ き ち ゃ ん 図13. ゆ い ち ゃ ん 20 イヌ・ネコの仲間 (食肉目) ︱哺乳類︱ (イヌ科) キツネ (イヌ科) タヌキ Vulpes vulpes Linnaeus, 1758 LC Nyctereutes procyonoides Gray, 1834 LC 図1-14. キツネ(撮影:鳥海隼夫) 形態 頭胴長60∼75cm,尾長40cm,体重4∼ 図1-15. タヌキ(撮影:鳥海隼夫) 7kg。背面は赤褐色で顎の下から腹部は白色。四 形態 頭胴長50∼60cm,尾長15cm,体重3∼ 肢の足先前面には黒毛が少し入る。尾は背面と同 5kg。全身白毛が少しまだらに入った灰黒色で, じ赤褐色で先端だけに白毛がある個体がいる。ほ 長いさし毛がある。目の周囲はやや濃い黒毛。尾 かのイヌ科動物とは,体色,頭胴長に対して尾が はふさふさする。ほかのイヌ科の種に比べて四肢 長いこと,吻部が細くとがることで区別できる。 が短い。接地する後足の指は4本で,5本指のイ 生態 おもに森林と畑が混在する田園地帯を好む タチ科と足跡で区別できる。 が,都市郊外から山岳部までさまざまな環境に生 生態 郊外の住宅地周辺から山地まで広く生息す 息する。ノネズミ類,鳥類,大型のコガネムシ類 る。鳥類,ノネズミ類などの小動物,昆虫,野生 などおもに小動物を捕食する。果実類を食べるこ 果実類などを食べる。甲虫の幼虫やミミズなど土 ともある。また,畑のトウモロコシやニワトリ, 壌動物の採食も多い。親子あるいは家族が近い距 家畜死体,人家のゴミを食べることもある。春先 離に集まり生活,行動をする。排泄物を特定の場 3∼4月に平均4頭の子を産み,夏まで家族群で生 所に集中するタメ糞を行う。タメ糞は個体または 活する。 家族集団間のナワバリ識別の役割があると考えら 分布 【山形県】広く分布する。【日本】北海 れている。春に3∼5頭を出産する。秋まで家族 道,本州,四国,九州,淡路島,小豆島。【世 群で行動する。 界】ユーラシアの大部分と北アフリカに分布。 分布 【山形県】広く分布する。【日本】北海 オーストラリアや北アメリカでもヨーロッパから 道,本州,四国,九州,佐渡,瀬戸内諸島など広 の移入個体が分布を広げた。 く分布する。一部に移入個体が分布。【世界】自 分類 ヨーロッパ産と区別され,本州以南の個 然分布域は中国東部からロシア南東部,朝鮮,日 体をホンドキツネVulpes vulpes japonica Gray, 本など。毛皮獣として導入されたものが東ヨー 1868,北海道産をキタキツネと,亜種とするこ ロッパを中心に野生化している。 とがある。 分類 日本産は大陸産と区別され,Nyctereutes 絶滅危惧等 山形県、環境省のレッドリストの指 procyonoides viverrinus Beard, 1904という亜種 定はない。国際保護連合では軽度懸念に指定して の扱いをされる場合がある。 いる。 絶滅危惧等 山形県、環境省のレッドリストの指 定はない。国際保護連合では軽度懸念。 る。冬眠中に2∼3年間隔で1∼2頭の子を出産す Canis familiaris (Linnaeus, 1758) 野生化種 る。 21 (イヌ科) イヌ ︱哺乳類︱ ペットとして世界中に分布する。放棄されたペ ットや狩猟犬がノイヌとなる。日本でも野生化し たノイヌが各地でみられる。集落近くや林道沿い などに生息することが多い。野外に適応できる中 型犬以上が多い。 ネコ (ネコ科) Felis catus (Linnaeus, 1758) 野生化種 イエネコが野生化しノネコとなったものがい る。日本では冬季の生活条件から,温暖な西日本 に多く寒冷地では少ない。また,森林や原野では 少なく,都市部に多い。人からの給餌と野外のエ サの両方を利用する半野生化ネコが多い。 図1-17. ツキノワグマ(Wikipediaより) 分布 【山形県】広く分布する(図1-18)。 【日本】本州,四国に分布。九州では絶滅した。 【世界】ヒマラヤの南側山麓から東南アジア北 部,中国東北部,ロシア南東部,台湾,海南島に 分布する。 図1-16. ネコ(霞城公園,撮影:鈴木弘二) ツキノワグマ (クマ科) Ursus thibetanus (G. Cuvier, 1823) LC(LP) 国際希少野生動植物種 CITESI 形態 頭胴長120∼145cm,体重70∼120kg, 尾は短い。全身黒色で胸に白い三日月模様があ る。まれに,三日月模様がない個体もある。本 1978年のみ 2003年のみ 州,四国では最大哺乳類。 両方で確認 生態 春はブナの若芽や草本類,夏はアリ,ハチ などの昆虫類,秋はクリ,ミズナラ,コナラ,サ 図1-18. 山形県内ツキノワグマの分布(第2期山形県ツキノワグマ保 ワグルミなどの木の実を採食する。カモシカなど 護管理計画より) の死体を捕食することもあるが食糧全体に占める 分類 大陸産はAsian Black Bearアジアクロ 割合は小さい。食糧を求めて人家の近くでも見ら ク マ と 呼 ば れ る が , 日 本 産 は そ の 亜 種U r s u s れることがあり,有害獣として駆除されることが thibetanus japonicus Schlegel, 1857とされてい ある。母子をのぞき単独で行動する。12月∼4月 る。 までは冬眠する。越冬場所としてはブナ,スギな 絶滅危惧等 山形県のレッドリストの指定はな どの大木の樹洞,あるいは岩穴や土穴を利用す い。環境省では下北半島の個体群を絶滅のおそれ 22 ︱哺乳類︱ があるとしている。国際保護連合では軽度懸念に があり,納屋に巣をつくることもある。齧歯類, 指定している。国際希少野生動植物種およびワシ 鳥類,両生類,爬虫類などの小型脊椎動物,昆虫 ントン条約附属書Iに指定され国際間の取引が規 類,ムカデなどの土壌動物,ヤマグワ,マタタビ 制されている。 などの果実類と多様なものを採食する。交尾は夏 におこなわれるが,出産は翌春になる。4∼5月 に2∼4頭の子を樹洞など比較的簡単な巣の中で 出産する。 分布 【山形県】広く分布する。【日本】本州, 四国,九州,淡路島,対馬に分布。北海道南部お よび佐渡には人為的に導入された。【世界】朝鮮 半島にも分布するという報告がある。 絶滅危惧等 山形県のレッドリストの指定はな い。国際保護連合では軽度懸念に指定している。 ツシマテンは,環境省のレッドリストでは準絶滅 危惧種とし,国の天然記念物にも指定されてい 図1-19. 捕獲後放獣前のツキノワグマ(提供:東英生) る。 その他 近年人里にあらわれ,農作物に被害を与 えたり,人に危害を加えたりすることで問題にな っている。捕獲数は資料(P. 32)に記載した。 テン (イタチ科) Martes melampus (Wagner, 1841) LC(NT) 図1-21. 貂(夏毛,両羽博物図譜より) イタチ (イタチ科) Mustela itatsi (Temminck, 1844) LC 図1-20. テン(撮影:長船裕紀) 形態 頭胴長45cm,尾長19cm,体重1.1∼ 1.5kg。毛色には,胸にオレンジ斑をもち,あざ やかな黄色をした個体(キテン)から,褐色をし たもの(スステン)まで変異が大きい。耳介がは っきりと外に出ること,前臼歯が4/4であること でほかのイタチ類と区別できる。 生態 樹上空間を多く利用するため森林を生息地 とする。樹木があれば人家周辺にも見られること 図1-22. イタチ(Wikipediaより) 雄12∼16cm, 雌7∼9cm。全身が茶から山吹色, 分布 【山形県】山岳地帯に分布する。【日本】 額中央部から鼻頭部にかけて濃い褐色の斑紋があ 北海道と本州中部の白山,北アルプス,中央アル る。雌は雄より小型。シベリアイタチは本種より プス以北の山岳地帯。【世界】ユーラシア北部と 大型で尾が長いことで区別できる。 北アメリカに広く分布する。 生態 雌は一定の行動圏をもち,土穴などを巣と 分 類 本 州 産 は 亜 種 ホ ン ド オ コ ジ ョM u s t e l a する。雄はいくつかの雌の行動圏に重なる。カエ erminea nippon Cabrera, 1913,北海道産は亜種 ル,ネズミ類,鳥類,昆虫類など陸上小動物,ザ エゾオコジョとされている。 リガニなどの甲殻類や魚を捕食する。平均3∼5 頭の子を産む。 分布 【山形県】広く分布する。【日本】本州, 四国,九州,佐渡,淡路島,屋久島などに分布。 北海道,一部の南西諸島などにネズミの駆除のた め導入され定着した。日本固有種。 分類 屋久島に生息する個体を亜種コイタチと いう。本種をシベリアイタチMustela sibirica 図1-24. オコジョ(冬毛,剥製) (Pallas, 1773) の亜種とする見方もある。 絶滅危惧等 山形県,環境省のレッドリストとも 絶滅危惧等 山形県,環境省のレッドリストの指 に準絶滅危惧に指定されている。国際保護連合で 定はない。国際保護連合では軽度懸念に指定して は軽度懸念に指定している。また,ワシントン条 いる。 約附属書IIIに掲載されている。 オコジョ (イタチ科) アナグマ (イタチ科) Mustela erminea Linnaeus, 1758 Meles meles (Linnaeus, 1758) LC LC NT NT CITESIII 図1-25. アナグマ(撮影:長船裕紀) 形態 頭胴52cm,尾長14cm,体重12kg程度。 図1-23. オコジョ(夏毛,提供:みどり自然課) 全体にくすんだ褐色で,四肢と胸部はやや濃い褐 形態 頭胴長雄18cm,雌16cm,尾長雄6cm, 雌 色をしている。両眼部は黒っぽい褐色。その間の 5cm,体重約100g。夏毛は背面が濃い褐色で, 鼻頭部中央は白色。ずんぐりしたからだつきで耳 腹面は白い。冬毛は全身白色で,尾の先端だけは 介は短い。四肢の爪は長く湾曲している。色彩か 黒毛が残る。 らほかの種と容易に区別できる。 生態 山岳地帯より高い地域に生息する。夏は高 生態 山地帯下部から丘陵部の森林,潅木林に生 山帯の岩場などでも活動する。高山に生息する鳥 息する。トンネルを掘り,雌を中心とする家族集 類,ノネズミ類,昆虫など小動物を捕食する。春 団で生活する。雄の行動圏は重ならない。土壌動 23 ︱哺乳類︱ に5頭前後の子を出産する。 形態 頭胴長雄27∼37cm,雌16∼25cm,尾長 24 ︱哺乳類︱ 物や小動物をおもに採食する。交尾は4∼7月で 生息記録がないことから外来種とみられる。【世 翌年春に出産する。狩猟獣であり生息数は年々減 界】東南アジア大陸部から中国南部,海南島,ス 少している。丘陵部の果樹園周辺などでは,タヌ マトラ,ボルネオ,ジャワ,台湾に分布する。 キやハクビシンに混じってワナで捕獲されること 分類 数亜種に分けられるが,どこから外来した もある。 か明確ではないため,亜種名は使用していない。 分布 【山形県】広く分布する。【日本】本州, 絶滅危惧等 山形県,環境省のレッドリストの指 四国,九州,小豆島。【世界】ヨーロッパから極 定はない。国際保護連合では軽度懸念に指定して 東までユーラシア北部に広く分布する。 いる。外来種であるが,渡来時期が明確でないた 分類 日本産は亜種ニホンアナグマMeles meles め,外来生物法での指定はない。 anakuma Temminck, 1844とされる。 絶滅危惧等 山形県,環境省のレッドリストの指 アライグマ (アライグマ科) 定はない。国際保護連合では軽度懸念に指定して Procyon lotor (Linnaeus, 1758) LC 外来種 いる。 頭胴長42∼60cm,尾長20∼41cm。灰褐色の 体毛をもち,眼のまわりから頬にかけて黒い斑紋 ハクビシン (ジャコウネコ科) がある。タヌキと誤認されることが多いが,尾に Paguma larbata (Smith, 1827) LC 外来種 黒い横縞があるのが大きな特徴である。また,足 は白っぽく耳には白い縁取りがある。 原産地は北アメリカ南部。水辺近くの森林に生 息するが,湿地,農耕地,海岸や都市などにも定 着できる。山形県ではこれまでも目撃情報がよせ られていたが,2013年6月10日に国道112号線 鶴岡市熊出付近で交通事故死と思われる個体が発 見された(図1-27)。 狩猟獣,特定外来生物に指定されている。 図1-26. 街中のハクビシン(山形市,撮影:原田裕行) 形態 頭胴長61(雌)∼66(雄)cm,尾長 40cm,体重3kg程度。体色は灰褐色で,顔面と 四肢の下部は黒褐色,額下部から鼻鏡部中央に白 線が入り他種と容易に区別できる。 生態 山地帯下部から集落周辺に生息する。最近 は町の中でも見られることがある。木登りが得意 で樹上をよく利用する。人家の屋根裏をねぐらと 図1-27. アライグマ(提供:鶴岡市自然学習交流館「ほとりあ」上山 することもある。鳥類とその卵,昆虫その他の小 剛司) 動物から果実類まで食性の幅が広い。ブドウ,ト ウモロコシなど果樹や畑作物を食害することもあ る。出産は初夏から秋までと考えられている。 分布 【山形県】広く分布し,市街地でも確認さ れている。【日本】北海道,本州,四国,九州の 各地で確認されているが,とくに北海道と西日本 では断続的な分布。江戸・明治期における確実な 部からユーラシアに広く分布する。 Sus scrofa Linnaeus, 1758 LC 国内外来種 分類 本州,四国,九州では亜種ニホンイノシ シSus scrofa leucomystax Temminck & Schlegel, 1842,南西諸島では亜種リュウキュウイノシシ と区別されている。 絶滅危惧等 山形県では明治末期に絶滅したとい われ,絶滅に指定されている。しかし,最近イノ シシは分布域を広げ,宮城県では定着し,さらに 山形県に侵入が報告されている。本書では絶滅を 除外した。環境省の指定はない。国際保護連合で は軽度懸念に指定している。 図1-28. イノシシ 形態 本州産では,頭胴長雄110∼160cm,体 重雄50∼150kg。雌では雄の60∼80%程度。南 西諸島産は小型。全身褐色または暗黒色の剛毛で おおわれ,鼻の先端は円盤状となる。耳介は小さ い。指の数は前後とも4本で,2個の蹄をもつ。 犬歯はよく発達し終生成長をつづける。とくに雄 では牙となる。 生態 常緑広葉樹林,落葉広葉樹林,里山の二次 図1-29. 山豕(両羽博物図譜より) 林,低山帯と隣接する水田,農耕地,平野部に広 その他 両羽博物図譜に記録がある(図 く分布する。雑食性で,地表から地中にかけての 1-29)。1907年,豚コレラで日本中のイノシシ 各種の植物と動物を掘りかえして採食する。植物 が激減,東北地方では壊滅したといわれている。 ではクズ,ヤマイモ,ススキの根茎や各種の葉, 「東北地方には明治末期すでにオオカミはいなく 果実など,動物では昆虫類,ミミズ,タニシ,カ なっており,豚コレラにかかったイノシシの死骸 エル,ヘビなどを食べる。出産期は春∼秋で,通 をオオカミの代わりにイノシシが共食いして一気 常1年に1回出産する。野外では4∼5頭の子をう に蔓延したのではないか。」という説もある。 む。初産は1∼2歳で毎年繰り返し繁殖をおこな 現在,県内でも有害捕獲がおこなわれている。 う。雄の成熟は2歳。平均寿命は1歳以下で,若 齢での生存率が低い。最長でも10歳以下。地域 により狩猟が盛んにおこなわれている。群れで生 活するが,通常,雄と雌は別々に行動する。雌は 母親とともに母系的な群れをつくる。雄は1∼2 歳で母親のもとを離れ,小さな群れをつくるか単 独生活をする。 分布 【山形県】太平洋側の宮城県などから侵入 がある(図1-30,資料P. 32)。【日本】本州, 四国,九州,南西諸島。【世界】北アフリカの一 図1-30. 狩猟捕獲イノシシ(2009.1.23) ︱哺乳類︱ (イノシシ科) イノシシ 25 ウシの仲間 (鯨偶蹄目) 26 ︱哺乳類︱ (シカ科) ニホンジカ 季節的変化は少なく,木の葉を採食する。出産期 Cervus nippon Temminck, 1838 は5月下旬∼7月上旬で通常1子を出産する。交尾 LC 国内外来種 期は9月下旬∼11月で,妊娠期間は約230日であ る。初産齢は2歳,最長寿命は雄15歳,雌20歳 前後である。雌は母親と母系的な群れをつくる が,雄は1∼2歳で母親のもとを離れる。雄の一 部は交尾期にナワバリをもち,ハレムをつくる。 雄は「フィー」という鳴き声を発し求愛をおこな う。マーキングや攻撃行動をおこなう。 図1-31. ニホンジカ(雄,冬毛) 図1-33. 雪の中のニホンジカ(2011.1.29,大江町,撮影:庄司光幸) 分布 【山形県】太平洋側の宮城県などから侵入 が見られる(図1-33,資料P. 31)。【日本】北 海道,本州,四国,九州,瀬戸内諸島,馬毛島, 図1-32. ニホンジカ(撮影:長船裕紀) 屋久島,種子島,対馬など。【世界】ベトナムか 形態 北海道産のエゾジカが最大で,屋久島産の ら中国東部,ロシア沿海地方,台湾,朝鮮半島, ヤクジカが最小。成体で,頭胴長雄90∼190cm, 日本に広く分布する。さらに,ヨーロッパや北ア 雌90∼150cm,肩高雄70∼130cm, 雌60∼ メリカに移入分布する。 110cm,体重雄50∼130kg, 雌25∼80kg。夏毛 分類 本州に分布する亜種をホンシュウジカ は茶色で白斑があり,冬毛は灰褐色である。黒い Cervus nippon centralis (Kishida, 1936),北海道 毛でふちどられた大きな白い尾斑がある。新生 産亜種をエゾジカ,四国,九州産亜種(基亜種) 子には細かい白斑がある。換毛期は5∼6月,9∼ をキュウシュウジカ,馬毛島産亜種をマゲジカ, 10月である。顕著な性的二型を示す。雄は角を 屋久島産亜種をヤクジカ,慶良間列島産亜種をケ もち1歳は1ポイント,2∼3歳は2∼3ポイント, ラマジカに分ける分類がある。しかし,ミトコン 4歳以上は4ポイント。角は毎年生え変わり,成 ドリアDNAから日本産のニホンジカは,北海道 体では30∼80cm。 から兵庫県までのグループとそれ以西の2つに分 生態 常緑広葉樹林,落葉広葉樹林,寒帯草原な けられるという報告がある。エゾジカは東日本の ど多様であるが,草地が入り組んだ森林に多く生 ホンシュウジカと同じ亜種となり,これまでの亜 息する。積雪地域の個体は雪を避け小規模な季節 種の分類は正確なものではないことになる。最近 的移動を行う。落葉広葉樹林に生息するエゾジカ の分類にはDNAを用いたものが重要視される傾 やホンシュウジカは,イネ科草本,木の葉,果 向があるが,種としてのニホンジカは変わりな 実,ササ類などを季節に応じて採食する。常緑広 い。 葉樹林に生息するキュウシュウジカなどは食性の 形態 成体では,頭胴長70∼85cm,肩高70∼ の長い体毛におおわれる。体色には個体変異があ る。両性とも黒い円錐形の角をもつ。角は角質の 真性角で,加齢とともに伸長し生えかわらない。 性的二型はほとんどない。体の大きさは3歳まで に成長が止まる。 生態 低山帯から亜高山帯にかけてのブナ,ミズ ナラなどが優占する落葉広葉樹林,針葉広葉混交 林に多く生息し,各種木本類の葉,広葉草本,サ サ類などを食べる。反芻胃を持つ。タメ糞をする 習性がある。山地に近い田畑ではカモシカによる 図1-34. 鹿(両羽博物図譜より) 食害が発生している。カモシカ用のネットで侵入 絶滅危惧等 山形県では両羽博物図譜(図 を防いだり,有害獣として駆除がおこなわれた 1-34)に記録があるように,明治時代には生息 りしている。出産期は5∼6月,交尾期は10∼11 していることがわかる。しかし,大正期に絶滅し 月,妊娠期間は215日で,通常1子を出産。平均 たといわれる。山形県のレッドデータブックでは 寿命は5歳前後で,最長で20歳を超える。単独生 絶滅として扱われているが,最近宮城県などから 活をすることが多く,4頭以上の群れをつくるこ 山形県に侵入が報告されている。今後見直しが必 とはほとんどない。積雪に強く,季節による移動 要である。本書では絶滅を除外した。 はあまりない。土地への定着が高く,雌雄ともナ 環境省の指定はない。国際保護連合では軽度懸 ワバリをつくる。同性間のナワバリでマーキング 念に指定している。 行動を盛んにおこなう。面積は10∼15haで,地 その他 最上町,遊佐町や河北町などで,飼育さ 域や生息環境により異なるが,雄の方がやや大き れている。飼育個体が逃げた報告はない。 い。ナワバリの重なり合う雄と雌が配偶行動をお 最近のニホンジカとイノシシの侵入については こなう。子は母親と約1年間ともに生活する。 別に報告した。 分布 【山形県】山地を中心に広く分布する。 【日本】本州,四国,九州。東北地方や中部地方 カモシカ (ウシ科) を中心に分布域が拡大している。日本固有種。 Capricornis crispus (Temminck, 1844) 分類 亜種に分けられていない。 LC(LP)要注目 絶滅危惧等 山形県の獣に指定されている。国の 国指定特別天然記念物 県の獣 特別天然記念物に1955年指定された。環境省で は九州の個体群を絶滅のおそれがあるとしてい る。国際保護連合では軽度懸念に指定している。 その他 かつては生息数が減少し,1934年に国 の天然記念物,1955年に特別天然記念物に指定 された。しかし,現在は増加し林業や農業の食害 が問題になっている。1975年ころから有害駆除 がはじまった。山形県でも駆除がおこなわれてい る。 図1-35. カモシカ ︱哺乳類︱ 蹄が発達している。全身白色または灰色,灰褐色 27 75cm,体重30∼45kg。四肢は太くて短く,側 28 ニホンジカとイノシシの侵入 ︱ 哺 乳 類 ︱ ︿寄 稿﹀ 玉手英利 (山 形 大 学 理 学 部 教 授) 山形のシカとイノシシの歴史 カがいなかった青森や秋田でも,2009年以降,目 ニホンジカ(Cervus nippon ,以下,シカ)と 撃例が増えており,従来からシカが生息していた宮 イノシシ(Sus scrofa )は古くから日本人の貴重 城,岩手では分布域がさらに拡大しています。 な狩猟資源として,その皮革,肉,骨角などが利 用されてきました。山形県内の縄文遺跡からはシ カの角で作った道具やイノシシの骨などが出土し ています。近世以降もシカとイノシシは利用され ており,徳川吉宗の時代に幕府の役人が各地の産 物を調べた「諸国産物帳」には,羽前でイノシシ を産したことが記載されています 1。シカについ ても,近世中期の歴史資料に,山形の「鹿」や「 シカ」に関する記述があることが報告されていま す2。 山形県のシカとイノシシは江戸末期から明治 期になると数を減らしました。1919年(大正8 年)を最後に,県内でのシカの捕獲は途絶え(山 形県 2003)3,昭和50年代までの約60年間,目 撃例もわずか数件となっています。このようなシ カとイノシシの局所的絶滅が起こった原因は,明 治維新により藩の猟場が開放されたことと,猟銃 が普及したことで,乱獲が進んだためと考えられ ています。さらに,シカやイノシシは積雪が深い 場合は容易に捕獲されるので,豪雪地域が多い山 形では特に個体数が減少したと考えられます。こ のような歴史により,現在の山形県はシカとイノ シシの分布空白域となっています(図1)4。 ところが, 2009年以降,山形県内では毎年、 シカが散発的に目撃されるようになりました。 2012年までの目撃例は17件にのぼり,交通事 故死した個体も6件確認されています。出没場所 は、最上,村山,置賜,庄内の県全域にわたりま す5。イノシシも2004年に県内で3頭が捕獲され たのをはじめとして,捕獲数が増えています(図 2)。明治期の絶滅からおよそ100年ぶりに,山 形県にシカとイノシシが戻ってきたのです。 図1. ニホンジカとイノシシの分布域 このようなシカやイノシシの分布拡大は,山形 生息が確認された地域を赤色で表示。環境省自然環境保全基礎調査動 以外の東北各県でも起こっています。これまでシ 物分布調査「日本の動物分布図集」(2010) より引用。 4 イノシシは福島から宮城,そして山形へと分布 29 域が広がりました。図2でみられるように,福島 ︱ 哺 乳 類 ︱ ︿寄 稿﹀ では平成16年頃から捕獲数が増加し,その数年 後に宮城でも同様のパターンで捕獲数が増加し始 めました。今後,山形でも福島・宮城と同じよう に個体数が推移するならば,数年後には県内のイ ノシシは急速に増えると予想されます。イノシシ は現在,おもに置賜,村山に生息していることが 確認されていますが,生息空白地であった秋田県 の南部でも新たな目撃が報告されていることか ら,山形県の全域がすでにイノシシの分布域と なっている可能性も考えられます。 図3. 各地のシカのミトコンドリアDNA配列のネットワーク図 ◇は山形のシカ,○は他地域のシカの遺伝子配列(ハプロタ イプ)を示します。ネットワーク上で近い位置にあるほど,遺 伝的に近縁と考えられます。山形で捕獲された個体のうち,山 形1は岩手のシカと同一でしたが,山形2は尾瀬のシカ,山形 3と山形4はいずれも栃木(日光)のシカと遺伝的に近縁でし 6 た。佐藤他(2013) の図を一部改変して引用。 図2. 山形県および近隣2県でのイノシシ捕獲数の推移 左側の縦軸は福島県と宮城県の年間捕獲数(狩猟と有害駆除の合 計数),右側の縦軸は山形県の年間捕獲数を示します。 山形のシカとイノシシはどこから来たのか? 山形県に進出しつつあるシカとイノシシの出所 を明らかにする目的で,山形大学理学部がDNA 分析を利用した調査を行いました 6。シカの調査 では,村山市,鶴岡市,小国町で交通事故により 死亡した4個体から組織を採取してミトコンドリ アDNAの塩基配列を調べました。エゾシカから ヤクシカまで日本各地のシカが持っているミトコ ンドリアDNAの配列と照合したところ,1個体の 遺伝子型(ハプロタイプ)が岩手県北上山地のシ 図4. 山形・宮城・福島3県におけるイノシシの集団構造 カでみられる遺伝子型と一致しました。一方,他 DNA分析の結果,東北南部のイノシシは大きく3つのグルー の3個体は,尾瀬や日光など北関東以西のシカと プ(青,緑,黄色で示す)に分かれていました。それぞれのグ 系統的に近いことがわかりました(図3)。この ループに属する個体の分布密度が高い地域を各色の線で囲んで 結果から,山形県のニホンジカは,少なくとも南 表示しています。 北2つの地域から,別々に進出している可能性が 示されました。山形県森林環境研究センターによ る出没状況の調査でも,複数のルートでシカが県 内に進出していることが示されています5。 30 ︱ 哺 乳 類 ︱ ︿寄 稿﹀ イノシシについては分布拡大の状況を把握する ニホンジカとカモシカの違い ために,山形県,宮城県,福島県で有害捕獲され シカ(ニホンジカ)とカモシカ(ニホンカモシ た個体のDNA分析を行いました。人間の親子判 カ)はよく混同されますが,ニホンジカはシカ 定や犯罪捜査で使われるマイクロサテライト遺伝 科,ニホンカモシカはウシ科に属し,形態や生態 子を用いた分析により,東北南部のイノシシは大 は大きく異なっています。シカ科とウシ科では角 きく3グループに分かれることが明らかになりま の形が違い,シカ科の多くの種は,雄のみが枝分 した(図4の緑,青,黄で示した地域)。このう かれして毎年生え変わる枝角(antler)を持つの ち黄色で示したグループは上山から宮城県南部, に対して,ウシ科は雌雄ともに枝分かれせず生え 青色で示したグループは天童から宮城県中部にか 変わらない角(horn)を持っています。ニホン けて連続しています。これから,宮城県からは複 ジカは群れで生活することが多いのに対して,ニ 数のルートで山形にイノシシが入ってきたと考え ホンカモシカは通常は単独で暮らしています。 られます。また,県南部の緑色で示したグループ については福島から入って来た可能性も考えられ イノシシとブタの違い ます。これらの調査から,シカとイノシシのどち イノシシとブタは同じ種(Sus scrofa )に属す らも県外の複数の地域から山形に侵入し,分布を る動物で,野生のイノシシを家畜化したものがブ 拡大している状況が明らかになってきました。 タです。ブタの家畜化は人間が定住生活を始める シカやイノシシが東北南部で急速に分布を拡大 とともにヨーロッパや西アジア,中国などで並行 している原因としては,里山環境の変化や狩猟者 して起こったことが知られています。現在の日本 の減少,気候の温暖化などが考えられており,生 で飼育されているブタの品種の多くは,ヨーロッ 息状況の調査・研究が続けられています。 パイノシシから家畜化された系統に属していま す。家畜化されたブタはイノシシに比べて繁殖性 シカとイノシシが増えることによる問題 や産肉性などが向上しており,気性もより扱いや 全国の野生鳥獣による農作物被害は金額にして すいものに変化しています。 約239億円(平成22年度)に上りますが,その 約5割がシカとイノシシによるものです。ニホン 文献 ジカは草食性で,シダ類や有毒植物を除くほとん 1 日本野生生物研究センター 1987. 過去における鳥獣分布 どの種類の植物を旺盛に食べます。そのため,シ 情報調査報告書. カの増加が著しい地域では,森林の更新が妨げら 2 伊藤愛・箕口秀夫・三浦慎悟 2009. 文献資料に基づいた れて草地化が進行したり,樹皮食いによって樹木 東北地方におけるニホンジカの過去の生息状況. 第56回日 が枯死したりするなど深刻な農林被害が生じてい 本生態学会大会発表. ます。また,高山植物や希少種の植物への食害も 3 山形県 2003. レッドデータブックやまがた. 山形の絶滅 問題となっています。イノシシは雑食性ですが主 のおそれのある野生動物. な餌は植物質で,根や地下茎なども掘り起こして 4 環境省自然環境局生物多様性センター 2010. 自然環境保 食べます。被害作物は稲,野菜から飼料作物まで 全基礎調査動物分布調査. 日本の動物分布図集. 多様で,田畑や植林地を掘り返すことによる被害 5 齊藤正一・古澤優佳 2012. 生物多様性の森作り実証モデ も生じています。シカやイノシシの食害が農業生 ル事業実施報告書―シカの目撃情報とその立地環境の整理 産のコストに及ぼす影響は大きいため,両種が共 を中心に. 存する西日本の被害地域では,有害駆除だけでな 6 佐藤真ほか 2013. 山形県で確認されたニホンジカの出自 く集落単位で囲う電気柵や緩衝帯を設置するなど ―ミトコンドリアDNA多型に基づく推定. 哺乳類科学, 53 様々な被害防止の取組が行われています。 (1): 印刷中. ᖹᡂ ᖺ ᭶ ᪥⌧ᅾ ᒣᙧ┴⎔ቃ࢚ࢿࣝࢠ࣮㒊ࡳࡾ⮬↛ㄢ 㐟బ⏫ ┿ᐊᕝ⏫ 㓇⏣ᕷ ┠ᧁ㸸H24.9 ┿ᐊᕝ⏫ᕝࣀෆ㏆ 㔠ᒣ⏫ ┠ᧁ㸸H24.72 㩬ᕝᮧ ┠ᧁ㸸H24.8 ┠ᧁ㸸H24.85 ᪂ᗉᕷ㔠ἑ㏆ ┠ᧁ㸸H24.7 ୕ᕝ⏫ ┠ᧁ㸸H24.82 㭯ᒸᕷῧᕝ㏆ ᗉෆ⏫ ᪂ᗉᕷ㔠ἑ㏆᪂ᗉᕷ ᭱ୖ⏫ ᭱ୖ⏫ᚿⱱ㏆ 㭯ᒸᕷ⩚㯮㏆ ┠ᧁ㸸H24.8 ᡞἑᮧ ㏻ᨾ㸸H21.9.27 ⯚ᙧ⏫ ᪂ᗉᕷ㫽㉺㏆ ᒣᙧ⮬ື㌴㐨᷸ᘬ㹑㸿㏆ ┠ᧁ㸸H21.6 ┠ᧁ㸸H24.82 ㏻ᨾ㸸H23.10.15 ▼⏣⏫ᷓἑ㏆ ᡞἑᮧゅᕝ㏆ 㹐㸵ሀⱏἑ㏆ 㭯ᒸᕷ ⶶᮧ ᑿⰼἑᕷ ▼⏣⏫ ┠ᧁ㸸H22,11.10 㭯ᒸᕷᮌ㔝ಛ㏆ ᐮἙỤᕷ Ἑ⏫ すᕝ⏫ ┠ᧁ㸸H23,1 Ụ⏫ᰗᕝ㏆ ㏻ᨾ㸸H21.10.23 ᮧᒣᕷᅵ⏕⏣ᆅෆࡢᅜ㐨 13 ྕ⥺ ᮧᒣᕷ ㏻ᨾ㸸H22.10.17 ᮧᒣᕷ㔠㇂ᆅෆࡢዟ⩚ᮏ⥺ ᮾ᰿ᕷ Ụ⏫ ኳ❺ᕷ ୰ᒣ⏫ ┠ᧁ㸸H23,1 ᮅ᪥⏫ⓑ㏆ ᮅ᪥⏫ ᒣ㎶⏫ ᒣᙧᕷ ⓑ㮚⏫ 㛗ᕷ ┠ᧁ㸸H21.10 ᒣᙧᕷᕝ㏆ ୖᒣᕷ ༡㝧ᕷ ┠ᧁ㸸H21.10 ᮾ㑥○▼ओ ᅵሙ ᑠᅜ⏫ 㧗␊⏫ ㏻ᨾ㸸H24.1.12 ᕝす⏫ ┠ᧁ㸸H24.04 ᑠᅜ⏫ᒾἑᆅෆࡢᅜ㐨 113 ྕ⥺ ᕝす⏫ᇼ㔠㏆ 㣤㇏⏫ ⡿ἑᕷ ┠ᧁ㸸H23,3 ⡿ἑᕷ⥘ᮌ㏆ ۑ㸸㏻ᨾࠊە㸸┠ᧁሗ 㸸࢜ࢫ࣭࣓ࢫࡢࡘࡀ࠸ ︱ 哺 乳 類 ︱ ︿資 料﹀ 31 㸺㈨ᩱ㸼ࢽ࣍ࣥࢪ࢝ࡢ┠ᧁሗ➼ ︱ 哺 乳 類 ︱ ︿資 料﹀ 32 㸺㈨ᩱ㸼┴ෆ࠾ࡅࡿࣀࢩࢩᤕ⋓ᩘࡢ᥎⛣㸦H16㹼H24㸧 ᒣᙧ┴⎔ቃ࢚ࢿࣝࢠ࣮㒊ࡳࡾ⮬↛ㄢ 㻡㻜 㻠㻜 㻟㻜 ᤕ ⋓ ᩘ ᭷ᐖᤕ⋓ ⊁ ⊟ 㻞㻜 㻝㻜 㻜 㻴㻝㻢 㻴㻝㻣 㻴㻝㻤 㻴㻝㻥 㻴㻞㻜 㻴㻞㻝 㻴㻞㻞 㻴㻞㻟 㻴㻞㻠 ᖺᗘ ༢䠖㢌 㻴㻝㻢 㻴㻝㻣 㻴㻝㻤 㻴㻝㻥 㻴㻞㻜 㻴㻞㻝 㻴㻞㻞 㻴㻞㻟 㻴㻞㻠 䠄 䠤㻝 㻢 䡚䠤㻞 㻠 䠅 㻥 ᖺᖹᆒ ィ ⊁䚷䚷⊟ 㻜 㻜 㻝 㻣 㻝㻝 㻝㻤 㻟㻠 㻝㻢 㻝㻠 㻝㻜㻝 ᭷ᐖᤕ⋓ 㻟 㻜 㻜 㻜 㻡 㻜 㻟 㻜 㻟 㻝㻠 㻝㻝㻚㻞 㻝㻚㻢 ᤕ㻌⋓㻌ィ 㻟 㻜 㻝 㻣 㻝㻢 㻝㻤 㻟㻣 㻝㻢 㻝㻣 㻝㻝㻡 㻝㻞㻚㻤 ᖹᡂ㻞㻡ᖺ㻟᭶㻟㻝᪥⌧ᅾ 㸺㈨ᩱ㸼┴ෆ࠾ࡅࡿࢶ࢟ࣀ࣡ࢢ࣐ᤕ⋓ᩘࡢ᥎⛣㸦H12㹼H24㸧 ᒣᙧ┴⎔ቃ࢚ࢿࣝࢠ࣮㒊ࡳࡾ⮬↛ㄢ 㻣㻜㻜 㻢㻜㻜 㻡㻜㻜 ᤕ ⋓ ᩘ 㻠㻜㻜 ᭷ᐖᤕ⋓ ⊁ 㻟㻜㻜 ⊟ 㻞㻜㻜 㻝㻜㻜 㻜 㻴㻝㻞 㻴㻝㻟 㻴㻝㻠 㻴㻝㻡 㻴㻝㻢 㻴㻝㻣 㻴㻝㻤 㻴㻝㻥 㻴㻞㻜 㻴㻞㻝 㻴㻞㻞 㻴㻞㻟 㻴㻞㻠 ᖺᗘ ༢䠖㢌 㻴㻝㻞 㻴㻝㻟 㻴㻝㻠 㻴㻝㻡 㻴㻝㻢 㻴㻝㻣 㻴㻝㻤 㻴㻝㻥 㻴㻞㻜 㻴㻞㻝 㻴㻞㻞 㻴㻞㻟 䠄 䠤㻝 㻟 䡚䠤㻞 㻞 䠅 㻝 㻜 ᖺᖹᆒ 㻴㻞㻠 㻞㻜㻡 㻞㻝㻤 㻞㻞㻥 ⊁䚷䚷⊟ 㻠㻠 㻝㻟 㻠㻤 㻟㻢 㻞㻣 㻡㻞 㻞 㻠㻝 㻞㻟 㻞㻤 㻝㻞 㻟㻠 㻠 㻞㻤㻚㻞 ᭷ᐖᤕ⋓ 㻝㻞㻡 㻞㻜㻟 㻝㻜㻜 㻝㻤㻢 㻞㻞㻣 㻝㻠㻣 㻢㻥㻜 㻤㻡 㻝㻠㻠 㻝㻜㻡 㻞㻞㻝 㻝㻜㻥 㻞㻤㻞 㻞㻝㻜㻚㻤 㻡㻥 㻞㻥 㻡㻜 㻢㻥 㻢㻥 㻣㻢 㻢㻥 㻡㻝㻚㻢 㻠 㻣 㻟 㻝㻟 㻜 㻠 㻢 㻚㻤 㻝 㻝 㻝 㻜 㻝 㻝 㻚㻜 㻞㻟㻟 㻝㻠㻟 㻞㻤㻢 ᤕ⋓ᩘୖ㝈ᩘ Ꮨᤕ⋓ 㻡㻟 㻠㻤 㻠㻠 㻟㻥 㻡㻞 㻡㻣 ᨺ⋇ᩘ 㻙 㻙 㻙 㻙 㻙 㻙 㻙 㘒ㄗᤕ⋓ 㻙 㻙 㻙 㻙 㻙 㻙 㻙 㻙 ᤕ㻌⋓㻌ィ 㻝㻢㻥 㻞㻝㻢 㻝㻠㻤 㻞㻞㻞 㻞㻡㻠 㻝㻥㻥 㻢㻥㻞 㻝㻞㻢 㻝㻢㻣 㻝㻟㻟 㻙 㻙 㻙 ⊁⊟⮬⢔ 䕿 䕿 㻙 㻞㻟㻜 䕿 䈜Ꮨᤕ⋓䛿䚸᭷ᐖᤕ⋓䛾ෆᩘ䛷䛒䜛䚹䜎䛯䚸ᖹᡂ䠎䠍ᖺᗘ䛛䜙ᒣᙧ┴䝒䜻䝜䝽䜾䝬ಖㆤ⟶⌮ィ⏬䛻䛚䛡䜛ಖㆤ⟶⌮䛻క䛔䚸 䚷㻌ணᐹᤕ⋓䛿Ꮨᤕ⋓䛻⛣⾜䛧䛶䛔䜛䚹 䈜㻴㻝㻢ᖺ㡭䛛䜙䛾䛂䜲䝜䝅䝅䛃䛾ὶධ䛻䜘䜚䚸㏆ᖺ䚸䛂㘒ㄗᤕ⋓䛃䛾䛜ᩓぢ䛥䜜䛶䛝䛶䛔䜛䚹 䈜⊁⊟⮬⢔䛻䛴䛔䛶䛿䚸⮬⢔䜢⾜䛳䛯ᖺ䛻䕿䜢グ㍕䚹 㻞㻟㻥 Hollister, 1912,佐渡産を亜種サドノウサギ,隠岐 Lepus brachyurus Temminck, 1845 LC (NT) 産を亜種オキノウサギ,それ以外を基亜種キュウシ ュウノウサギとして区分する場合がある。 絶滅危惧等 山形県のレッドリストの指定はない。 環境省ではサドノウサギを準絶滅危惧種としてい る。国際保護連合では軽度懸念に指定している。 狩猟獣。かつて90万頭前後が捕獲されていた が,現在は十数万頭に減少している。 カイウサギ(アナウサギ) (ウサギ科) Oryctolagus cuniculus (Linnaeus, 1758) 図1-36.ニホンノウサギ(冬毛,撮影:鳥海隼夫) 野生化種(県内での野生化はない) 図1-37. ニホンノウサギ(夏毛,撮影:長船裕紀) 形態 頭胴長43∼54cm,尾長2∼5cm,耳長6∼ 8cm,後足長13∼15cm,体重1.3∼2.5kg。腹が 図1-38. カイウサギ 白色である以外はほぼ全身茶褐色で,耳の先端が 日本では,カイウサギとして家畜化している。小 黒い。本州の東北部や日本海側の積雪地帯および 学校などで飼育されているのは本種である。野生化 佐渡の個体は,冬になると耳の先端を除き全身白 している場所の多くは外敵のいない無人島である。 色になる。 地下でトンネルを掘り群れで生活する。山形県内で 生態 低地から亜高山帯までの森林や草原など様 も飼育個体が逃げ出すことがあるが,野生化は確認 々な環境にみられるが,低山から山地帯に多い。 されていない。 植物食性で,葉,芽,枝,樹皮を食べる。夜行性 原産地はイベリア半島とアフリカ北部であるが, で巣はつくらない。春から夏まで連続して3∼5 ヨーロッパ,オーストラリア,ニュージーランド, 回の出産を繰り返す。妊娠期間は42∼47日。子 チリなど世界各地に移入され野生化している。 は1∼4頭,ふつう2頭である。寿命は4年未満。 分布 【山形県】広く分布する。【日本】本州, 四国,九州,佐渡,隠岐諸島,淡路島,小豆島, 五島列島,天草諸島など。日本固有種。 分類 本州で冬季に白色の毛になるものを亜種 トウホクノウサギLepus brachyurus angustidens ︱哺乳類︱ (ウサギ科) ニホンノウサギ 33 ウサギの仲間 (兎目) 34 ネズミの仲間 (齧歯目) ︱哺乳類︱ (ヤマネ科) ヤマネ レッドリストの指定は最近除外されている。国際 Glirulus japonicus (Schinz, 1845) 保護連合では軽度懸念に指定している。 LC VU 国指定天然記念物 国の天然記念物になっている。 ヤチネズミ (ネズミ科) Eothenomys andersoni (Thomas, 1905) NT 形態 頭胴長79∼118mm,尾長40∼63mm,体 重11∼40g。中部山地や紀伊半島ではやや大型。 分布 【山形県】広く分布する。【日本】本州の 中部・北陸以北と紀伊半島。日本固有種。 スミスネズミ (ネズミ科) Eothenomys smithii (Thomas, 1905) 形態 頭胴長70∼115mm,尾長30∼50mm,体 重20∼35g。 分布 【山形県】長井ダム,新庄市升形から記録 がある。日本では北限にあたる。【日本】本州の 新潟県・山形県以南,四国,九州。日本固有種。 ハタネズミ (ネズミ科) Microtus montebelli (Milne-Edwards, 1872) LC 図1-39∼40. ヤマネ(撮影:やまがたヤマネ研究会) 形態 頭胴長90∼136mm,尾長29∼50mm,体 形態 頭胴長68∼84mm,尾長44∼54mm,耳 重22∼62g。 長6∼9mm,後足長15∼17mm,体重は夏14∼ 分布 【山形県】広く分布する。【日本】本州, 20g,冬34∼40g。背面は淡褐色で,腹面はより 九州,佐渡。日本固有種。 淡色。背中に黒褐色の縦線が1本あり,目のまわ りも黒褐色。ネズミに似るが,尾には約20mmの カヤネズミ (ネズミ科) 長い毛がある。 Micromys minutus (Pallas, 1771) DD 生態 低山帯から亜高山帯の成熟した森林に生息 する。夜行性でおもに樹上で活動する。からだの 大きさの割には広い行動圏をもつ。樹洞や木の枝 の間に樹皮やコケを集めて球形の巣をつくる。果 実,昆虫そのたの小動物,小鳥の卵などを食べ る。寒冷期に冬眠する。雌は年2回,3∼5子を出 産する。寿命は約3年。 分布 【山形県】西川町や鶴岡市(旧朝日村)な ど山地から知られている。【日本】本州,四国, 九州。日本固有種。 図1-41. カヤネズミ(Wikipediaより) 絶滅危惧等 山形県では絶滅危惧II類。環境省の 形態 頭胴長 50∼80mm,尾長 60∼90mm,体 【山形県】白川ダムで記録がある。日本で 分布 後足長18∼21mm,体重10∼20g。頭胴長より は北限にあたる。 【日本】本州では山形県・宮城 尾長が大きいことによりアカネズミと区別でき 県以南,四国,九州。【世界】ヨーロッパからシ る。 ベリア,中国,台湾,朝鮮半島,日本などに分布。 分布 【山形県】山地を中心に分布する。【日 本】北海道,本州,四国,九州,周辺の島々。日 アカネズミ (ネズミ科) 本固有種。 Apodenus speciosus (Temminck, 1844) LC ドブネズミ (ネズミ科) Rattus norvegicus (Berkenhout, 1760) 外来種 形態 頭胴長186∼280mm,尾長149∼ 220mm,後足長27∼46mm,体重150∼500g。 分布 【山形県】広く分布する。【日本】広く分 布する。【世界】世界的に分布する。 その他 河川,海岸,湖畔,市街地では下水,台 所の流し,ゴミ捨て場,地下街,食品倉庫など水 図1-42. アカネズミ(撮影:鳥海隼夫) が十分に摂取できる湿った場所を好む。植え込み の地中,耕作地の堆肥の中などに巣を作る。 ドブネズミ,クマネズミ,ハツカネズミの「家ネ ズミ」はほぼ世界中に分布する。いずれも人間と ともに大陸から渡ってきた史前帰化動物と考えら れる。世界に広がる前の野生動物としての原産地 は,メソポタミア地方の河川流域や沼沢地帯と考 えられている。 日本の家ネズミのうち,最も個 体数が多いのはドブネズミである。 図1-43. アカネズミ(撮影:やまがたヤマネ研究会) これらのネズミ類は,貯食性,貯脂肪性,冬眠 形態 頭胴長80∼140mm,尾長70∼130mm, など,冬を越すために必要な習性を欠く。そのた 後足長22∼26mm,体重20∼60g。 め,冬期の食料を人間に頼るために人間の生活の 分布 【山形県】広く分布する。【日本】北海道, 場に入り込んだと考えられる。その結果として, 本州,四国,九州,周辺の島々。日本固有種。 人類とともに世界中に分布を広げ,他のネズミ類 以上に繁栄することとなった。 ヒメネズミ (ネズミ科) Apodenus argenteus (Temminck, 1844) LC クマネズミ (ネズミ科) Rattus rattus (Linnaeus, 1758) 外来種 形態 頭胴長146∼240mm,尾長150∼ 260mm,後足長22∼40mm,体重150∼200g。 分布 【山形県】広く分布する。【日本】広く分 布する。【世界】世界的に分布する。比較的乾燥 した場所でも生息できる。 ハツカネズミ (ネズミ科) 図1-44. ヒメネズミ(撮影:やまがたヤマネ研究会) Mus musculus Linnaeus, 1758 外来種 35 ︱哺乳類︱ 形態 頭胴長65∼100mm,尾長70∼110mm, 重 7∼14g,日本では一番小さなネズミ。 36 ︱哺乳類︱ 形態 頭胴長57∼91mm,尾長42∼80mm,体 形態 頭胴長16∼22cm,尾長13∼17cm,耳 重10∼25g。体色は変異にとみ,白色,灰色, 長22∼31cm,後足長48∼58cm,体重250∼ 褐色,黒色など。短毛で腹側は淡い。両羽博物図 310g。背面は夏毛では赤褐色,冬毛では灰褐 譜の南京鼠(図1-45)は本種である。 色。腹は白色。尾の先が白い。耳の先にふさ毛が 分布 【山形県】広く分布する。【日本】広く分 生じる。 布する。【世界】世界的に分布する。 生態 平野部から亜高山帯までの森林に生息する が,低山帯のマツ林に多い。昼行性でおもに樹上 で活動する。ほぼ植物食で花,芽,種子,果実, キノコのほか,昆虫その他の節足動物も食べる。 春から秋に食物を地面に浅く埋めたり,木の枝に 挟んだりして貯蔵する。小枝,樹皮などで球状 の巣を樹上の枝の間などにつくる。行動圏は約 10ha。初春から夏までに1∼2回,産子数は2∼6 頭。寿命は約5年。 分布 【山形県】広く分布する。【日本】本州, 四国,九州,淡路島。中国地方には少なく,九州 と淡路島からは最近確実な生息記録がない。日本 固有種。北海道にはキタリスとシマリスが分布す る。また,クリハラリスが外来している。 図1-45. 南京鼠(両羽博物図譜より) 絶滅危惧等 山形県のレッドリストでは準絶滅危 惧,国際保護連合では軽度懸念に指定している。 ニホンリス (リス科) Sciurus lis Temminck, 1844 環境省のレッドリストでは中国地方および九州地 LC(LP)NT 方のものは絶滅のおそれのある個体群とされた。 ムササビ (リス科) Petaurista leucogenys (Temminck, 1827)LC NT 図1-46. ニホンリス(撮影:鳥海隼夫) 図1-47. ニホンリス 図1-48. ムササビ(剥製) 長35∼43cm,後足長61∼71cm,体重700∼ 色。胸部は白色。目のまわりが黒褐色。 1000g。飛膜が首から前肢,前後肢の間,後肢 生態 山地帯から亜高山帯までの森林に生息す と尾の間に発達する。背面は褐色系であるが色彩 る。夜行性で,樹上で活動し,飛膜を使って木々 変異が大きく,北方のものの方が淡い色をしてい の間を滑空する。おもに樹洞を巣にする。ほぼ完 る。腹面は白い。目と耳の間から頬にかけて帯状 全な植物食で花,芽,葉,種子,果実,キノコ類 に淡色の部分がある。体長および飛膜が首と前 を食べる。年に2回,3∼5子を産む。 肢,後肢と尾の間にあることによりニホンモモン ガと区別できる。 生態 自然林,発達した二次林や針葉樹植林など に生息する。低地から亜高山帯まで見られるが低 地に多い。夜行性で樹上活動する。ほぼ完全な植 物食で花,芽,葉,種子,果実を食べる。雌は1 ∼1.5ha,雄は2∼3haの行動圏で互いに重ならな い。繁殖は年2回で,冬と初夏に交尾し,春と秋 の2回,通常2子を産む。寿命は約10年。 図1-50. ニホンモモンガ 図1-49. 野衾(両羽博物図譜より) 図1-51. モモンガ(両羽博物図譜より) 分布 【山形県】広く分布する。【日本】本州, 分布 【山形県】広く分布する。【日本】本州, 四国,九州。日本固有種。 四国,九州。日本固有種。 絶滅危惧等 山形県のレッドリストでは準絶滅危 絶滅危惧等 山形県のレッドリストでは絶滅危 惧,国際保護連合では軽度懸念に指定している。 惧II類,国際保護連合では軽度懸念に指定してい 環境省の指定はない。 る。環境省の指定はない。 その他 両羽博物図譜に図が掲載されている(図 その他 両羽博物図譜に図が掲載されている(図 1-49)。 1-51)。 ニホンモモンガ (リス科) Pteromys momonga Temminck, 1844 LC VU 形態 頭胴長14∼20cm,尾長10∼14cm,耳 長15∼25cm,後足長32∼39cm,体重150∼ 220g。目が大きく,前肢と後肢の間に飛膜があ 37 ︱哺乳類︱ る。尾は偏平。背面は夏毛が褐色,冬毛が灰褐 形態 頭胴長34∼48cm,尾長28∼41cm,耳 38 今昔 山形のヤマネ ︱ 哺 乳 類 ︱ ︿寄 稿﹀ 中村夢奈 (や ま が た ヤ マ ネ 研 究 会) 1 ヤマネとは 生息環境である森林の減少による絶滅の危険性 ニホンヤマネGlirulus japonicus (以下ヤマネ) から,山形県では絶滅危惧種Ⅱ類に指定されてい は日本の森林に生息するネズミ目の一種である。 る。世界にヤマネ科動物は8属26種が現存してい 体長は約7cm,尾の長さは約5cmである。形態的 るが,ニホンヤマネは極東に現存する唯一の種で 特徴は,目の周りの黒い縁取り,茶褐色の体毛の あり,日本固有種かつ貴重種として,昭和60年6 背面中央に見られる黒い筋(黒条線),ふさふさ 月26日に天然記念物に指定された。 とした尾である(図1)。夜行性で木の実や昆虫 2 さまざまな呼称 を食べる。 昔の日本人は山中で見かけたヤマネに対してそ の不思議な生態や見かけから,地方独特の名前で 呼んでいた。とくに,触ると死んだように冷たい 状態で冬眠する生態に由来するものが多い(表 1)。その名前から推察するに,当時の人はヤマ ネに対して親しみよりも驚きや不気味さを感じて いたことが伺える。山形西川町では「木鼠(キネ ズミ)」と呼ばれていたようである。これは,木 の上を生活するネズミの意味として,全国的に広 図1. ニホンヤマネGlirulus japonicus く使われていた呼称であったようだ。 表1. 各地域のヤマネの呼称(中島 2001参考) 図2. 冬眠中のヤマネ 冬には飲まず食わずで,樹洞や土の中で玉のよ 3 昔のヤマネ うに丸まって冬眠する(図2)。ヤマネは冬眠中 ヤマネの生態や生息記録が初めて記録されたの に体温を外気温に同調させながら0℃近くまで下 は1700年代と考えられている。1757年1月26日 げることもできる上に,呼吸数や心拍数も大幅に に捕獲されたヤマネについて肥後(現在の熊本 減少させて仮死状態で冬を越すという生理システ 県)六代藩主細川重賢が「毛介綺煥(もうかいき ムを持っている。そのため,冬眠中のヤマネに触 かん)」に挿絵と観察から得られた生態を記述し れるとひんやり冷たい。夜行性,樹上性,特異な ている。ここではヤマネと断定されておらず,後 冬眠行動が故に山中で出会うことが難しい動物で に朱書きで「関東ニテヤマ子ト云日光山中ニ多 ある。 有」と追記させている。そのため,ヤマネの呼称 が初めて出てくる文献は1780年代で,仙台藩主 39 伊達家に生まれ,後に江戸幕府の若年寄を務めた ︱ 哺 乳 類 ︱ ︿寄 稿﹀ 堀田正衡が記述した「観文獣譜(かんぶんじゅう ふ)」であると考えられている。ここではヤマネ の挿絵はなく文章のみで,日光産のヤマネを譲り 受けて飼育観察した内容が記述されている。 図3. 「日本動物誌(Fauna Japonica)」のヤマネ 江戸時代に医師としてオランダから来日してい 図4. 博物館獣譜のヤマネ(画像提供:東京国立博物館) たシーボルトは1833∼1850年に刊行された「日 この「動物訓蒙」を読んだ松森胤保は「両羽博 本動物誌(Fauna Japonica)」においてヤマネの 物図譜」(庄内地方の動植物が記録されている) 全図と頭骨図を記載している(図3)。シーボル に初めて山形でヤマネの記述を行った人物であ トはこの時ヤマネを新種として考えており,特に る。ヤマネ自体を松森胤保は見たことがなく「動 美しい毛並みを称賛している。ここで,初めて海 物訓蒙」のヤマネを見た感想として『北海道産の 外のヤマネ(ヨーロッパヤマネ)と比較記述がな シマリスに似ており,尾はニホンリスのようで された。 ムササビとは異なるようだ』と記している(図 同時期である明治初期に,官吏として後の東京 5)。 国立博物館や東京都恩賜上野動物園の基礎を築い しかし,ニホンリスの章を見ると『月山山麓の た田中芳男が編纂した「博物館獣譜」はヤマネの 志津のあたりの人々はリス(栗鼠)と木に登る鼠 挿絵を最も多く記載した文献である(図4)。何 (木鼠)は別物で,木鼠は小さいものであると述 種類かの図譜を切り張りしたもので,それぞれに べている。木鼠は体色がテンの様に淡赤茶で,尾 「竹鼠」や「阿蘭陀鼠」などと呼称が書かれてい の毛が扁平で細長い形をしている。リスとは異な るが,どれも黒条線などの特徴を記しており,ヤ り,大きさはネズミのサイズだ。』と記述してい マネであることが判断される。残念なことに,当 る。この文章を見るに,志津の人が言っている 時の政府の目指す西欧近代生物学とは相いれない 「木鼠」はヤマネの可能性がある。その第1の理 ものと判断され,集大成されることなく終焉を迎 由として,毛色である。個体によってヤマネは赤 えた。しかしこの後,田中芳男は現代の動物図鑑 みがかった茶色に見えることもある。第2に,尾 ともいえる「動物訓蒙 哺乳類初編」を1875年 である。ヤマネは扁平の尾をもっている。さら に出版し,ヤマネの生態を記述している。 に,現代においてもヤマネはリスの子と間違われ ることや,ネズミにリスの尾を生やしたものと形 容されることから,当時の志津の人々が混同して 40 ︱ 哺 乳 類 ︱ ︿寄 稿﹀ いる可能性は容易に想像できる。第3に,現在の 大井沢小中学校(現廃校)では,学校教育の一環 山形県西川町近辺では,昭和初期には既にヤマネ として地域の自然研究がおこなわれており,その を「木鼠」と呼んでいたと言われている。ヤマネ 成果を1954年に「峠の小道」として出版してい の大きな特徴である黒条線について記載がされて る。この中に,ヤマネの冬眠に関する研究を行っ いないことからヤマネとは断定はできないが,当 た中学校1年の生徒がいる。ここでは,飼育下に 時リスやネズミとは異なるヤマネのような「ネズ おいて冬眠前の摂食種類の記述,外気温7℃で冬 ミサイズのリスの様な尾をもつ生物」が確認され 眠を行い13℃で活動を始めること,呼吸数と脈 ていたことは確かである。 拍数もカウントして冬眠に伴う回数減少について も言及している(図6)。また,山形大学名誉教 授の大津高は山形県産のヤマネを用いた冬眠に関 する学術研究論文を1993年に発表している。ヤ マネの冬眠生理機構に関する学術研究は1930∼ 1950年代に下泉重吉が数例行って以来ほとんど 進んでいない中,山形県産のヤマネを使用した研 究が2例も存在している。 図6. 「峠の小道」内のヤマネの写真 4 今のヤマネ 過去の文献・記録や学術研究から,山形のみな らず全国的にヤマネが珍しい生き物として扱われ ているのがわかる。それは,めったに出会うこと ができないためである。筆者は,2008年より山 図5. 「両羽博物図譜」におけるヤマネの記述部(画像提供:酒田市 形県でもヤマネの記録数や発見数の多い西川町大 立光丘文庫) 井沢地区においてヤマネの生態調査を行ってき 20世紀に入ると山形県でも野生動物の生息調 た。調査結果から,なぜ山形でヤマネに出会える 査が行われるようになり,現在の庄内町,鶴岡 きっかけが少ないのか見えてきた。 市,西川町,金山町,天童市(面白山),東根市 (船形山),米沢市(吾妻山)などの山麓部(ブ 原因1 冬眠期間 ナの原生林や広葉樹林内)でヤマネは発見されて 森林内にヤマネの巣箱を設置して定期的に観察 いる。 したところ,ヤマネの巣箱利用は5月中旬から10 昭和60年に天然記念物に指定される以前は、 月下旬に観察された(図7)。一般的にヤマネの 野山で発見されたヤマネを持ち帰り,飼育観察し 研究では,巣箱の利用が見られない月の期間を冬 た報告例を目にすることがある。山形県西川町立 眠期間(生態的冬眠期間)と考えられている。こ のことより,山形のヤマネは約6か月半∼7か月 41 の間冬眠していることと考えられる。夜行性で日 ︱ 哺 乳 類 ︱ ︿寄 稿﹀ 中は寝ている上に,1年の半分以上樹洞や土のな かで冬眠しているのでは,出会える期間は非常に 短い。 図8. 背中に専用接着剤で小型発信機を付けられたヤマネ 5 これからのヤマネ ヤマネの調査をしているなかで,興味深い結果 が得られた。それは,ヤマネがスギやカラマツな 図7. 2010∼2012年の間に各月に巣箱で捕獲された合計個体数 どの植林を自然林と同様に行動圏として頻繁に利 と各月の平均・最高・最低気温(大井沢測候所観測データ参 用し,巣材としても好んで利用するということで 考)。灰色点線は野外で冬眠を誘導する境界温度(平均気温 ある。さらに,養蚕のために林縁部に植樹された 9℃)を示す(Shimoizumi 1940)。 ヤマグワに対して高い選好食性があり,初夏の重 要な食料源であることも分かっている。山形県に 原因2 日中の睡眠場所(日内休眠場所) はブナやナラなどの豊かな原生林が残されてお ヤマネは巣を持たない動物で,毎日転々と寝る り,ヤマネの生息域はこのような自然林であると 場所を変える。ヤマネに超小型の電波発信機を取 考えられてきた。しかし,山形のヤマネは植林な り付けて追跡してみると,ヤマネは2m以上の高 どの人的活動による環境変化にも上手に適応しな 所を日内休眠場所として利用していた(表2,図 がら生き残ってきたと考えられる。現在,ヤマネ 8)。日中の休眠場所も高所にあるのでは,人の は生息域である森林の減少から全国的に減少傾向 目につくことも少ない。 にあることが指摘されている。今後は,ヤマネを 表2. 日内休眠場所の特徴(ヤマネの報告の多い長野県との比較) 含む野生動物たちの適応が追いつかないほど速い 環境の変化のないような森林管理が求められるだ ろう。 参考文献 中島福男 2001. 日本のヤマネ. 信濃毎日新聞社. Shimoizumi, J. 1940. Studies on the hibernation もしも,昔のヤマネも同じような冬眠期間や休 of Japanese dormouse Glirulus japonicus (Schinz). 眠場所の高さであったならば,自然をよく観察し 1. On the hibernation period. Bunrika Daigaku ていたであろう松森胤保さえもヤマネを見たこと (Science Reports of Tokyo), Sect. B, 4: 51-61. がなかったとしても納得がいく。 Shibata, F., Kawamichi, T. & Nishibayashi, K. 2004. Daily rest-site selection and use by the Japanese dormouse. Journal of Mammalogy, 85: 30-37. 44 ︱ 哺 乳 類 ︱ ︿寄 稿﹀ 庄内海岸に漂着するクジラ 長澤一雄 (山 形 県 立 山 形 中 央 高 等 学 校) クジラという動物 にラクダ亜目が分かれ,次にイノシシ亜目,ウシ 水中に高度に適応した哺乳類で,現在世界の海 亜目が分かれた。最後にクジラはカバ亜目との共 洋には80余種が棲息している。分類学上「クジ 通祖先から分かれて進化した。この見解に基づい ラ目」に属する動物で,そのうち歯を有するグ て新しい分類名である「クジラ偶蹄目」が提唱さ ループを「ハクジラ亜目」,歯がなくヒゲ板を備 れた。 えるグループを「ヒゲクジラ亜目」と呼ぶ。 古いクジラとしては,始新世約5,000年前のパ ハクジラ亜目は,体長数mの小型の種が多く, キケタスやアンブロケタスなどが有名である。こ これらはイルカと呼ばれる。一般に多数の歯を有 れらは四肢を有するが,陸上と水中を行き来して するが,アカボウクジラ科などは,進化とともに いた。こうした古いクジラの仲間を「ムカシクジ 歯を極端に減少させた。ハクジラは小魚やイカな ラ亜目」と呼ぶ。その後クジラは急速に水中適応 どを餌としており,群れをなす場合が多い。そ を進めた。漸新世3,000万年前になると,歯を消 の種類は70種ほどでクジラ目の種の大半を占め 失してヒゲ板を発達させた新しいクジラが分かれ る。最大の種はマッコウクジラである。ヒゲクジ て進化した。ヒゲクジラである。こうしてクジラ ラ亜目は,体長10mを超えるナガスクジラなど の2大グループであるハクジラ亜目とヒゲクジラ の大型の種が多い。歯がなく代わりに上顎だけに 亜目が成立した。中新世2,000万年前になると, 硬く弾力のあるヒゲ板をもつ。これはケラチン質 現在見ることができるすべての科が出現し,現代 の長い三角形状をした薄い板で,上顎の両側に数 型クジラの時代になった。 百枚が重なる。餌は小魚やオキアミ類で,ヒゲ板 世界の各地から多くのクジラ化石が産出してお によって餌が口内で濾しとられる。夏は餌場の り,海の繁栄者としての進化をたどることができ 極圏へ,冬は繁殖場の暖海域へと大規模に回遊 る。日本の最古の化石は漸新世3,000万年前の化 する。その種類は10数種と少ないが,古来より 石で,北九州地域や北海道足寄地域から産出して 捕鯨の対象とされてきた。なお,イルカとクジラ いる。足寄地域からは「歯をもつヒゲクジラ」つ は正式な分類名ではなく慣習的な呼び名である。 まりハクジラとヒゲクジラの中間的な形質をもつ 体長が概ね4m以下のものがイルカで,それより 化石が産出している。中新世以降は,日本各地か 大きいものがクジラと呼ばれてきた。英語名の ら化石が産出する。大型脊椎動物の化石として dolphin とwhaleの使い分けもほぼ同様である。 は,クジラの産出頻度が最も高い。ただし,全身 が揃った化石は少なく,断片骨として産する。こ クジラの進化と系統 クジラは新生代の初期に陸から海へ進出した哺 乳類で,中生代の末期に絶滅した大型海棲爬虫類 のニッチを占有した。クジラの系統については, 1990年代に遺伝学と形態学によるクジラの新た な研究によって,クジラは偶蹄目(ウシ目)のカ バ亜目に最も近縁であるという新設が提案され た。偶蹄目は現在多様な種を含む大きなグループ を形成するが,それによると,クジラの祖先は新 生代の初期に偶蹄目に属しており,そこから最初 図1. オウギハクジラの漂着(鶴岡市鈴,1993年) 漂着するクジラ んだ個体または生きた個体が海岸に乗り上げるこ と。②漂流:死んだ個体が海を漂流しているこ と。③混獲:死んだ個体または生きた個体が意図 せずに漁網等で捕獲されること。④迷入:生きた 個体が漁港などに迷い込むこと。これらのうち, 死後の漂着個体が最も多いため,単に漂着(スト ランディング)という場合は,この意味で使われ 図2. オウギハクジラの頭部(鶴岡市米子,1999年) ることが多い。生存個体が漂着した場合は,区別 れは,日本が常に変動帯であったことに関係す するため,生存漂着(ライブ・ストランディン る。地殻変動は,海底の骨格を移動させ分散させ グ)または座礁と呼ぶ。イルカなど小型のハクジ た。 ラが集団で座礁する場合があり,それを集団座礁 山形県では,中新世以降の地層から化石が産出 (マス・ストランディング)と呼ぶ。 する。最も化石記録が多い地域は新庄盆地であ クジラの漂着は日本各地であり,その報告は年 る。ここは中新世後半から鮮新世にかけて,深く 間200∼400件にのぼる。これは,日本のどこか 静かな内湾であったが,そうした環境がクジラに で毎日のように漂着があることを示している。こ 適したのだろう。新庄盆地の化石は,大型のヒゲ れらの記録は,㈶日本鯨類研究所や国立科学博物 クジラが多い。1990年代に山形県立博物館で発 館動物研究部で収集しており,データベースとし 掘したクジラ化石の中には,体長推定15mのナ てHPでも公開されている。漂着個体の生物学的 ガスクジラ科の化石が含まれていた。こうした化 研究は,国立科学博物館,大学,水族館,博物館 石は現在の日本海のクジラの祖先である。 等によって,現地での計測,解剖,内臓サンプル の採取・分析などが実施されている。死因につい 日本海のクジラ は特定されない場合が多いが,地道な調査の蓄積 世界の80余種のクジラのうち,日本列島周辺 は今後も必要である。 で確認されている種はその半分の約40種であ 漂着クジラの調査は,絶えず人的および予算不 る。このうち日本海では,その半分の約20種が 足の問題を抱える。同時に,漂着クジラが国内法 確認されている。種の確認は,洋上での目視や漂 ではゴミと同様の一般漂着物としか扱われないと 着個体の調査によって行われている。 いう根本的な問題がある。日本列島周辺海域の環 日本海のハクジラ亜目としては,シワハイル 境と生物の実態を正しく認識する上でも,漂着鯨 カ,マダライルカ,マイルカ,ハンドウイルカ, カマイルカ,オキゴンドウ,ハナゴンドウ,マゴ ンドウ,ネズミイルカ,イシイルカ,スナメリ, シロイルカ,コマッコウ,ツチクジラ,オウギハ クジラ,アカボウクジラなど,ヒゲクジラ亜目と しては,ナガスクジラ,ミンククジラ,ザトウク ジラ,セミクジラなどが確認されている。このう ち,日本海で漂着が多い種は,マイルカ,オキゴ ンドウ,カマイルカ,イシイルカ,オウギハクジ ラ,ミンククジラなどである。 図3. ミンククジラの漂着(遊佐町湯ノ田,1993年) ︱ 哺 乳 類 ︱ ︿寄 稿﹀ う。広義には次の4つの場合がある,①座礁:死 45 漂着はストランディング(stranding)ともい 46 類の法的整備を進め,自治体と研究機関による調 査研究のネットワークづくりが望まれる。ちなみ ︱ 哺 乳 類 ︱ ︿寄 稿﹀ に,広い国土と長い海岸線を有するアメリカ合衆 国では,年間数千件のクジラなどの海棲哺乳類の ストランディングがあるという。このためストラ ンディングの対応を明確にするために,MMPA (Marine Mammal Protection Act:海棲哺乳類保 護法)が1972年に制定されて,生きた個体や死 んだ個体の保護と調査研究が法律の定めによって 実施されている。 図4. オウギハクジラの漂着(鶴岡市釜谷坂,1995年) ハクジラ亜目としては,ツチクジラ,オウギハ 庄内海岸の漂着クジラ クジラ,カマイルカ,ハナゴンドウ,イシイル 最近の山形の漂着クジラを表に示す。ほとんど カ,ヒゲクジラ亜目としては,ミンクジクラ,ナ が死後漂着で冬季に集中する。これは他県でも同 ガスクジラが記録されている。多い種はオウギハ 様の傾向で,冬季は個体が衰弱しやすく,また強 クジラ,カマイルカ,ミンククジラで,特にオウ い季節風の影響で流れつきやすいためと考えられ ギハクジラがこの20年で10個体と最も多い。山 る。ただし,その数は他県に比較すると少ない。 形県立博物館では,国立科学博物館の協力のも 海岸線が短いことが一因であろうが,他の要因に とで1990年代に漂着クジラの調査を実施してお ついても検討していく必要はある。隣県の新潟県 り,現地解剖を実施するとともに,オウギハクジ では,佐渡島を含めて長い海岸線を有するために ラやミンククジラの骨格標本を収集して日本海の 漂着クジラが多く,食糧資源としても利用されて クジラの標本として保管している。 きた。それを「寄り鯨」と呼び,また供養のため の「鯨塚」なども点在する。鯨塚は山形に存在し 謎のオウギハクジラ ないことからみても,昔から漂着クジラが少な 庄内海岸での最多記録は,オウギハクジラであ かったようである。 る。その漂着は,1990年代に集中していたが, 2000年代に入ると漂着が無くなる。これは日本 海の他県でも同様で,1980年代から急にオウギ ハクジラの漂着が相次いだが,2000年に入ると 漂着が激減した。原因は明らかでなく,研究者も 首をかしげる。このクジラは,以前は世界的にも 漂着がほとんどなかったため,研究が進んでいな かった「謎のクジラ」だった。しかし,近年の日 本海での多数の漂着によって研究が一気に進展し た。 オウギハクジラは,体長5m,体重1t程度で,ア カボウクジラ科のオウギハクジラ属に属し,学名 をMesoplodon stejnegeri という。オウギハクジ ラ属の種は,現在13種が認識されている。クジ ラにおいて,1つの属がこのように多数の種に分 化している例はほかにない。骨格や外部形態の差 とDNAの違いなどで種が分けられるが,相互の 外見は類似性が高い。このことは,比較的新しい ディスプレイと繁殖行動用であると考えられる。 る。なぜこのような磨耗が起こるのかは明らかで ない。歯の磨耗と主食のイカとの関係が薄いため 大きな謎となっており,研究者の頭を悩ませてい る。 骨格では,頭骨に最も特徴が表れる。前方に鳥 のように突き出す吻部は,摂餌方法の特殊化に 伴って変化した。上顎骨には歯は全くなく,歯槽 図5. オウギハクジラ頭骨 左:背面観 右:側面観 が浅い筋状の痕跡になっている。このことは,古 年代に種分化が進んだことを示唆している。 い時代に歯を消失したことを示唆している。吻部 日本列島周辺海域には,オウギハクジラ,イ の後方に亜円形の前頭部があり,鼻孔周辺が浅い チョウハクジラ,コブハクジラ,ハッブスオウギ 凹状をなしている。ハクジラは音波を増幅するた ハクジラの4種が棲息している。いずれの種も, めに鼻孔の上にメロンと呼ばれる脂肪塊をもつ 漂着や目撃例が極端に少ない謎のクジラとされて が,前頭部の形態はメロンの受け皿の形をしてい きた。このうち日本海には,オウギハクジラ1種 る。中央部の鼻孔から後方の鼻骨にかけては,頭 のみが棲息する。日本海では,漂着個体に妊娠例 頂部は急激に上昇して鶏冠のようにオーバーハン があり,また新生児の漂着があることなどから, グする。ここには鼻骨を中心として多種の頭蓋骨 この種が日本海で繁殖していることが確実とされ が集合している。ここでは,後退する前頭骨と前 る。つまり,オウギハクジラは日本海を古里とす 進する後頭骨が頭頂骨を押し潰すように複雑に重 るクジラである。 なり合っている。こうした複雑な頭頂部の特徴 は,クジラの頭蓋に共通して見られ,「頭蓋の短 オウギハクジラの形態 縮」またはテレスコーピング(telescoping)と その名前の由来となっている大きな特徴は,下 呼ばれる。 顎の扇形をなす1対の大型の歯である。歯の数は ハクジラ亜目で最も少ない。オスの歯は大きく萌 漂着クジラのこれから 出するが,メスの歯は歯肉に埋もれたまま外部に クジラは数千万年かけて進化してきた哺乳類で 姿を現すことがない。イカ類を主食とするが,漂 ある。しかし,シロナガスクジラなど一部のクジ 着個体の胃内容物の調査からも確かめられた。吻 ラは絶滅に瀕しているし,ヨウスコウカワイルカ 部は小さく突出し,口は大きく開かない。これは は,21世紀になって絶滅した。クジラをどのよ イカ類を吸い込みために適している。体表に無数 うに保護し,また活用するかについては,クジラ の傷を有する。これは繁殖期にオスの歯によって 自体をさらによく知る必要があるだろう。そのこ できたものと推定される。オスの歯は個体誇示の とは,人類という生物学上の種を理解し,また将 来を考える手がかりにもなるだろう。 かつて,解剖学者でありクジラ研究の先駆者で あった小川鼎三は,生物界には2つの大きな山が あると例えた。それはヒト山とクジラ山であり, クジラ山を極めることによって,ヒトの深い理解 が得られるのだと説いた。生物の種の多様性の理 解は,DNAだけでは進まない。実物の形態比較 図6. オウギハクジラの歯 左から右へ成長・加齢変化 によってより明確化される。商業捕鯨は現在禁止 ︱ 哺 乳 類 ︱ ︿寄 稿﹀ 同時に,歯の基部の前方で半円状の磨耗が進行す 47 オスの歯は,加齢とともに急速に大型化が進む。 48 海の哺乳類 (食肉目) ︱哺乳類︱ 庄内海岸に漂着した哺乳類を記録する。山形県 のレッドリスト掲載はない。下記以外にも記録さ れている可能性があるが詳細は不明。 図1-59.アシカ(両羽博物図譜より) オットセイ (アシカ科) Callorhinus ursinus (Linnaeus, 1758) VU 一匹のオスが複数のメスを独占し海岸に近い場 所にハーレムを形成する。ハーレムを持たないオ 図7. 保存されたミンククジラ骨格標本(山形県立博物館蔵,1993年 スは群れをつくって生活する。 遊佐町湯ノ田) されており,クジラを研究する上での漂着個体の ニホンアシカ (アシカ科) 価値は計り知れない。これを調査研究すること Zalophus japonicus (Peter, 1866) EX CR は,自然科学の一つの大きな仕事である。 明治期には庄内でも多く確認されている。両 アメリカのクジラ研究者ジェームズ・ミード 羽博物図譜のアシカ(図1-59)は貴重な記録で は,漂着クジラについて次のように述べた。「漂 ある。1974年に礼文島で幼獣1頭が捕獲され, 着は発見されなければならない。次にそれは報告 1975年に竹島で目撃した記録が現在における最 されなければならない。そして,報告は記録され 後のニホンアシカ生息事例となっている。以後は なければならない」と。さらに一言加えよう。 生息の情報は得られておらず,絶滅したものとみ 「標本は保存されなければならない」と。 られている。国際保護連合では絶滅だが,環境 省のレッドリストでは絶滅危惧Ia類とまだ絶滅に 文献 なっていない。 国立科学博物館 2013. 海棲哺乳類ストランディングデータべ ース.http://www.kahaku.go.jp/ 長澤一雄・山田格 1997. 山形県温海町漂着オウギハクジラ属 Mesoplodon の頭蓋にもとづく分類の再検討.日本海セトロ トド (アシカ科) Eumetopias jubatus (Schreber, 1776) EN NT 両羽博物図譜に記録がある。 ジー研究, 7: 27-36. Shimano, M., Yasue, H., Oshima K., Abe, H., Kato, H., Kishiro, ゴマフアザラシ (アザラシ科) T., Munechika, I. & Okada, N. 1997. Molecular evidence Phoca largha Pallas, 1811 from retroposons that whales form a clade within even- 繁殖は一夫一妻で4月に交尾する。約一年後の toed ungulates. Nature, 388: 666-670. 3∼4月に流氷上で一子を出産。このほかのアザ ラシも日本海でみられる場合がある。 鳥類 オオハクチョウ (撮影:長船裕紀) 山形県の野鳥 奥山武夫 (山 形 県 立 博 物 館) 50 ︱ 鳥 類 ︱ ︿寄 稿﹀ 1. 戦後,野鳥の世界に何が起こったのか 30年を境にして日本の動植物の生態系は大きく 日本の明治時代以降,動物の種類によっては生 転換していった。水田やその周辺からトンボ類, 息環境の減少や乱獲などからニホンオオカミ,コ ホタル,ガムシ類などの水生昆虫が姿を消し,イ ウノトリ,トキなどのように数を減らしたり絶滅 ナゴ,タニシ,ドジョウ,フナ,クモ類,カエル したりしたものも多くいたといえる。 類,ヘビなども激減していった。 (1) 昭和20年代 そのため,水生昆虫などを餌にするツバメ,ヒ クイナ,セグロセキレイ,魚を餌にするカワセ ミ,カイツブリなどが,田園地帯では激減して いった。 反対に,汚水に強い種類のユスリカの仲間が大 発生したため,それを餌にするイワツバメがツバ メに変わって激増していった。それまで,市街地 では繁殖することがなかったイワツバメが,ビル や橋の下,駅舎などで集団営巣するのが見られ, その害が問題になる程であった。その後,イワツ 図1. イワツバメ(撮影:長船裕紀) バメは徐々に減少していったが,ツバメは現在も 先の大戦中や以後も食糧難のため,野生動物も 回復していない。 採集され食料とされていたが,生態系がかわるよ うな変化は認められなかった。しかし,20年代 後半になると強力な農薬や殺虫剤などが散布され るようになって動物の世界では大きな変化のきざ しが見られるようになってきた。 その1つの例として,その時まで平地の人家付 近では見られなかったイワツバメの営巣が昭和 28年に初めて山形市内で見られた。 (2) 昭和30年代 図3. ツバメ(撮影:やまがたヤマネ研究会) (3) 昭和40年代 日本人の生活が豊かになり,生活排水の増加, 工場排水による河川の汚染,山林の伐採,各種の 開発などが進んで,日本の自然は最悪の時代であ った。 また,一方で自然保護思想の啓蒙も進み,これ も自然に大きな影響を与えた。 オオヨシキリ,コチドリ,イソシギ,コアジサ 図2. ツバメ(提供:鈴木かおり) シなどの水辺の鳥も減少していった。 タルなどが回復のきざしが見られるようになっ た。 越冬するハクチョウ類,留鳥のカラス,キジな どが増加してきて目立つようになった。 地で越冬し,あとは南へ移動してゆくのが普通で あったが,平地や市街地でも繁殖するように変化 してきた。 (5) 昭和60年代,平成時代 図4. ムクドリ(撮影:長船裕紀) 増加してゆく鳥と減少してゆく鳥との差が,さ 山形県内に生息するムクドリは40年代初めま らに大きくなってきている。 では夏鳥で冬期間は雪のあまり降らない地方に渡 河川では魚類が以前のような多様性はないが, っていくのが普通であった。それが,この時より 限られた種類のウグイをはじめオイカワ,アブラ 一部が県内に残り越冬するようになった。温暖化 ハヤなどの増加が見られ,更に,放流によるアユ などにより小雪になったということもあるが,当 の増加が重なってゴイサギ,カワセミなどが回復 時は食品類の残飯の廃棄や果樹類の取り残しなど し,本来本県では数が少なかったコサギ,アオサ が見られるようになったのも,越冬を可能にした ギも増加していった。 原因の1つであったと思われる。 山地の鳥ではヤマドリ,サンショウクイ,ヨタ 一方,日本人全般の意識も変化してきており, カ,チゴモズ,トラツグミ,センダイムシクイな 今までほとんど人を近づけなかったカルガモが市 どが50年代に続いて減少していった。蔵王山頂 街地でも繁殖するようになり,渡ってきたハクチ や,雨雲の日などに平地でもたくさん見られたア ョウ,カモ類などが身近な所で越冬するようにな マツバメもあまり見られなくなっている。 った。 哺乳類のニホンカモシカも生息地の拡大がはじ 2. 新しく県内で繁殖が確認された鳥 まり,里山でも生息が見られるようになった。 (4) 昭和50年代 図6. オナガ(剥製,村山市大淀産) アマサギ,チゴハヤブサ,オナガなどがいる が,オナガが県内で初めて確認されたのは,昭和 図5. ヒヨドリ(撮影:長船裕紀) 36年(1961年)6月12日に村山市大淀で採集さ 自然保護思想がさらに進み,農薬,工場排水な れてからである。その後,徐々に県内に広がって どの規制により河川の水質が徐々に良くなってい いった。 くが,開発行為は大規模化する。 また,以前県内でのカワウは最上川河口など 水田,水辺の一部ではイナゴ,アカトンボ,ホ で,たまに観察される程度で繁殖はしていなかっ ︱ 鳥 類 ︱ ︿寄 稿﹀ 夏に山地で繁殖し秋になれば山を降り,一部は平 51 山形県内に生息するヒヨドリは,この時まで春 たが,平成4年頃から飯豊町で繁殖が確認され 蔵王山,飯豊山,月山,鳥海山などの山頂部に た。その後,県内を北上して繁殖地を広げ,アユ はイワヒバリ,ホシガラス,カヤクグリ,ビンズ などに被害が出はじめ,河川関係の漁業組合など イなどが繁殖し,ブナ,ミズナラ帯の中腹部には で問題になっている。 イヌワシ,クマタカ,キビタキ,トラツグミなど 52 が繁殖している。中腹部から里山地帯にはアオゲ ︱ 鳥 類 ︱ ︿寄 稿﹀ 3. 山形の野鳥について ラ,コガラ,センダイムシクイ,オオルリ,ヤマ 日本列島は南北に連なる多くの島々からなって ガラ,メジロ,カケス,ホオジロなど多くの鳥類 おり,海岸線も長く,北は亜寒帯から南は亜熱帯 が繁殖する。 までの様々な気候が見られ,面積のわりには多様 平野部には身近な所でキジ,カルガモ,ツバ な環境に恵まれているといわれている。 メ,モズ,スズメ,ムクドリなどが繁殖し,河川 今まで日本では,600種ほどの鳥類が確認され 敷などのヨシ原ではオオヨシキリが営巣し,河川 ているといわれていたが,「日本鳥類目録 改訂 敷林ではゴイサギ,アオサギ,コサギ,カワウな 7判(2012)」によると,633種が掲載されてい どが集団で営巣し,コロニーをつくっている。 る。この種類数は,世界の鳥類の6%ほどをしめ ており,日本の面積からすると種類数は多いとい われている。しかし,日本で繁殖する鳥類はお およそ240種で,相対的には少ないといわれてい る。 また,日本で見られる鳥類のうち,おおよそ 30%は迷鳥と呼ばれる種類の渡り鳥である。 山形県は本州東北地方の日本海側に位置し, 冷温帯,広葉樹林帯に属し,多雪地帯である。 2,000m級の山地が連なり蔵王,吾妻,飯豊,朝 図8. コハクチョウ(撮影:長船裕紀) 日の各連峰,月山,鳥海山など日本百名山といわ 冬期にはハクチョウ類,カモ類,サギ類,冬鳥 れている山々の中で6峰に接している県である。 の小鳥類など多くの種類が越冬したり,県内を通 そこにはブナ,ミズナラ帯,スギ林などが広が 過したりする。とくに,最上川河口をはじめ庄内 り,平地には水田,果樹畑などがしめている。 地方では多く見られる。 また,流域面積が県土の76%をしめている最上川 庄内海岸にはウミネコ,イソヒヨドリなど,内 があり,庄内地方には日本海に接する海岸線が約 陸部ではほとんど見られない種類も生息してい 80kmの長さにわたっている。更に,離島である る。春秋の渡りのシギ,チドリ類も旅鳥として通 飛島もあり,山形県は多様な自然に恵まれた環境 過してゆくが,太平洋側に多くある干潟などがな にあるといえる。 いため個体数,種類数はあまり多くない。 酒田市の飛島は,ウミネコの繁殖地として国の 天然記念物として指定されている(昭和25年8月 29日指定)。また,飛島は春秋の渡りの時期の 中継地として,全国的にも有名で多くの種類が記 録されている。日本での初記録種や,本土ではめ ったに見られない迷鳥,普通種も含めて300種に 近いといわれている。 県内で現在までに確認された種類数は390種以 上とされている。 図7. クマタカ(撮影:長船裕紀) 鳥海山に舞うイヌワシ 長船裕紀 (環 境 省 猛 禽 類 保 護 セ ン タ ー) 53 も,近年の巣立 ① イヌワシの基本データ ちする雛数が減 少している為, 地域単位での繁 殖番の絶滅が心 配されている。 図1. 日本におけるイヌワシの生息メッシュ図(環境省) ③ イヌワシの生活サイクル ② イヌワシの分布と生態 イヌワシは世界に6亜種いると言われており, 日本には日本と朝鮮半島付近に生息する亜種ニホ ンイヌワシApuila chrysaetos japonica が生息して いる。ニホンイヌワシは6亜種のなかで最も小さ く,生息域が狭い。以降イヌワシは,日本におけ る亜種ニホンイヌワシについて記す。 ニホンイヌワシは,本州以南に分布する留鳥の 図2. イヌワシの生活サイクル 猛禽類の中では最も大きく,最小の亜種とはいえ 繁殖シーズンは11月頃から始まり,1月にかけ 翼開長は最大2mに達する。猛禽類全般に言える て雄と雌が並んで飛行する横並び飛行や波状飛 ことで,体重は雄が3.0∼3.5kg,雌が3.5∼5.5kg 行,足と足を絡ませるなどの行動が求愛期に見ら で雌の方が大きい。北海道から九州までの低山地 れるようになる(求愛期)。1月∼2月は岸壁の 帯から高山帯にかけて生息し,個体数は全国で 岩棚に樹木の枝や葉,ススキなどを運び,巣を 650羽,番数は260番程度(環境省2004)と推 造る(造巣期)。2月上旬∼中旬頃に産卵し,約 定されている。しかし最近の情報では,西日本で 45日抱卵する(抱卵期)。巣内での育雛期間は のイヌワシの生息状況が悪化しており,特に四国 70∼94日で,5月下旬∼6月中旬頃に巣立ちする や九州での確認例は激減し,中国地方において (巣内育雛期)。巣立ち後もしばらく巣外での育 ︱ 鳥 類 ︱ ︿寄 稿﹀ (1)イヌワシとは 雛期間があり,親が幼鳥に餌を与える(巣外育雛 動が頻繁に観察 期)。 されることが多 い。 ④ イヌワシの行動圏と巣について 54 ︱ 鳥 類 ︱ ︿寄 稿﹀ 特定の動物の個体(つがいや集団)が,生活環 図4. イヌワシの狩り のある時期に通常の生活のために動きまわる地域 のイメージ のことを行動圏という。イヌワシの場合,この行 環境省作製 動圏の中には巣やねぐら,休息場所,狩り場,巣 などが存在し,広さは平均60㎢にも及ぶ(日本 (2)鳥海山とイヌワシ イヌワシ研究会)。 鳥海山は山形 営巣環境は,岸壁のオーバーハングした岩棚に 県と秋田県の県 巣を構えるが,まれに樹木に造ることがある。行 境に位置し, 標 動圏の中の最低標高地点から最高標高地点の中間 高は2,236mにも 標高地点付近か,それよりも低標高地に巣がある 及ぶ。標高は山 ことが多い。巣の標高が狩り場の標高と比較し, 形県の最高峰, より低いほど巣への餌の運搬労力が軽減されるこ 東北地方の中で とや,高度が高いほど気象条件が厳しいことが関 も2番目に高く 係していると言われている。 日本百名山・日 鳥海山 → 本百景で知られ 図5. 山形県におけるイヌワシ ⑤ イヌワシの食性と狩り ている。 生息メッシュと鳥海山位置 食性は主にノウサギ,ヤマドリなどのキジ類, アオダイショウなど大型ヘビ類などだが,タヌキ やキツネ,テンなどの小型哺乳類の他,カモシカ やシカなどの幼獣も狩ると言われている。 図6. 酒田市八幡地区から望む鳥海山 鳥海山には亜高山帯の針葉樹林帯がほとんど見 られず,ブナ帯が森林限界の上限となり,それよ り上は偽高山帯の低木や,風衝草原,高山ハイデ が分布する。標高1,100m付近上部にはミヤマナ ラやナナカマド,ダケカンバ,チシマザサ群落, ショウジョウスゲ群落が発達する。1,100m以下 では自然植生のブナ群落が優占し,ヒメヤシャブ 図3. イヌワシの餌資源(左上:ノウサギ,右上:アオダイショウ, シやタニウツギ,キタゴヨウが散在する。標高が 左下:ホンドギツネ,右下:ヤマドリ) 低くなるにつれ,ブナ−ミズナラ群落,スギやカ イヌワシが狩りを行う環境は,自然裸地や自然 ラマツなどの植林,クリ−ミズナラ群落がまと 草地、伐採跡地など開けた環境で行う。林縁部の まって分布する。鳥海山の四季折々の景色は壮観 木や見通しの良い岩に止まり,待ち伏せでの狩り で,観光客や登山者のみならず多くのイヌワシ愛 や,上空から獲物を探し急降下して狩りをする。 好家,バードウォッチャーを惹きつけてやまな なお雨の日に行われることは少なく,主に晴天や い。現在,鳥海山には北麓と南麓に一番ずつ, 曇天に行われる。したがって,長雨の後は探餌行 計4羽が生息し,観察情報ではそれ以外の個体も 時々観察されることから,4+α生息しているこ ことで,北麓イヌワシの番とは別の番が南麓に存 とになる。 在することが判明した。1993年から,本格的に 個体の追跡調査が行われ,遂に1995年に巣が発 見された。その他の事項は図10を参照。 鳥海山にイヌ 現在まで,環境省や山形県,東北森林管理局, 55 ワシが生息して 研究者,地元のイヌワシ関連団体などが調査を行 いることは今で い,内部構造解析の結果,行動圏や高利用域が少 は広く知られて しずつ解明されていった。植生調査やウサギの糞 いることだが, 量調査,餌資源回復 イヌワシはかつ や狩り場環境の創出 ︱ 鳥 類 ︱ ︿寄 稿﹀ て生態のみなら を目的に,列状伐採 図7. 保護された渓谷 などの取り組みも実 えも謎に包まれていた。鳥海北麓で暮らす人々に 施された。繁殖状況 は,古くから「鳥海ワシ」と言い伝えられ,畏怖 調査も行われ,1993 の念を抱き伝説的存在でもあった。その鳥海ワシ 年から現在に至るま が我々の前に姿を現したのは1970年のことであ で,年ごとの繁殖の る。6月のある日,北麓の渓谷沿いの林道で2羽 成否が把握されてい のイヌワシが保護された。当時の様子の詳しい記 る(図11)。 録は多くないが,幼綿羽が残っており,2羽は林 図9. 南麓のイヌワシ調査に向かう調査員 ず,その存在さ 道のさらに奥の断崖の巣棚で育った兄弟であっ た。巣立ちする時期であり,何らかの要因で地上 へ落下し,飛び立てなくなった状態で保護され た。そのまま現在の秋田市大森山動物園に収容さ れ,それぞれ「鳥海」と「白滝」と名付けられ, 長期的に飼育が開 始された。その後, 白滝は1989年に死 亡したが,鳥海は 現在も40歳を過ぎ 図10. 鳥海南麓におけるイヌワシと人々の歴史 ても存命で,国内 最高齢を更新中で ある。 図 8. 秋 田 市 大 森 山 動 物 園 で 飼 育 さ れ て い る イ ヌ ワ シ の 鳥 海 (2012 年 撮 影) ② 鳥海南麓イヌワシ 一方で,山形県側の鳥海南麓におけるイヌワシ の歴史は北麓のイヌワシより浅い。麓で暮らす 人々には,北麓同様伝説として知られていたかも しれないが,広くイヌワシが知られるようになっ たのは,1989年に旧八幡町の鳥海南麓にスキー 図11. 鳥海南麓におけるイヌワシの繁殖状況(1993年∼2012年) 場開発の計画が持ち上がったことがきっかけであ <巣の補修> る。大規模な開発行為を危惧した地域の日本山岳 図10. に記したように,1996年及び1998年, 会の有志が,山岳研究の一環として調査を始めた 2回にわたって巣の崩落が生じた。1回目の1996 ① 鳥海北麓イヌワシ 年は繁殖活動終了後に崩落したため,繁殖に影 響はなかったが,1997年の繁殖には失敗した。 1998年は繁殖に成功したが,繁殖活動終了後2 回目の崩落が生じ,壊滅的状態となった。それま 56 での調査から,他に営巣出来そうな場所が少ない ︱ 鳥 類 ︱ ︿寄 稿﹀ と判断され,日本山岳会及び,継承するかたちで 発足した鳥海山ワシタカ研究会,猛禽類専門家ら によって崩落した巣の補修が行われ,人工巣が設 置された。1999年以降は,3年連続繁殖が行わ 図13. 全国及び東北地方,山形県におけるイヌワシの近年の繁殖成功 れなかったが,2002年∼2007年までの間,計4 率推移(環境省,日本イヌワシ研究会) 回人工巣で繁殖が行われた。 現在の日本の山地植生は,自然植生では林冠が 発達し,人工林においても手入れが行き届かず荒 れたまま放置され,鬱閉した環境も少なくない。 前述のように,開けた環境で狩りを行うイヌワシ にとって,狩り場環境の善し悪しは繁殖成功の鍵 となっている。 鳥海山は多雪地であり,冬季の強風は凄まじ く,急峻な地形は頻繁に雪崩を引き起こす。よっ て鳥海南麓のイヌワシの行動圏には,自然裸地や 自然草地が散在し,多雪風衝地には低木群落が成 図12. 鳥海南麓の補修後の人工巣(提供:鳥海山ワシタカ研究会) 立し,牧場として利用されていた場所などの開け <繁殖成功率> た環境も多く, イヌワシの現状を語る上で,「繁殖成功率」 それらが狩り場 (繁殖に成功した巣の数÷巣立ちの成否を調査 となっている可 した巣の数×100)という重要なキーワードがあ 能性がある。ま る。現在,全国的に繁殖成功率の数値が著しく低 た猛禽類の餌場 下しおり,日本イヌワシ研究会及び環境省の報 環境の創出を目 告から整理すると,1981年∼1985年までは平均 的とした列状伐 図14. 探餌する鳥海南麓のイヌワシ 47.2%の繁殖成功率であったが,2010年までの 採が実施されていることが,繁殖成績の良好な理 10年間では,23.9%と半減している。 由かもしれない。 鳥海南麓のイヌワシでは観察開始後,2013年 ところが,鳥海南麓のイヌワシを脅かしかねな で20年になる。繁殖の成否は判明している1993 い不安要素も少なくない。イヌワシの行動圏の流 年∼2012年の20年で,9回繁殖に成功した。繁 域において,支流では土砂崩れや油の流出が発生 殖成功率にして45%を誇る。2012年までの過去 し,本流では河川工事が行われている。治山治水 10年では40%とやや低下するが,全国的に見て に伴う調査は,航空からも行われ,観光や山菜採 も良好な数字と言える。 り以外の人の出入りも多くなっている。さらに, 全国的な繁殖率の低下の要因は,採餌環境の減 冬季から春季にかけて,イヌワシの行動圏におけ 少が大きいと言われている(由井ほか)。餌の獲 る山岳地域へ,スノーモービルの乗り入れが確認 得が難しくなることで,繁殖に必要な親鳥の体力 されていることや,オスプレイの低空飛行訓練の の維持,育雛期に必要な十分な餌の供給が出来な 飛行ポイントにも近い(ピンクルート)。これら くなったと考えられる。 の人間活動が,必ずしもイヌワシへ影響をもたす とは限らないが,影響は未知数であり今後も追跡 調査が望まれる。 主な参考文献 山岸哲・森岡弘之・樋口広芳 2004. 鳥類学辞典. 57 環境省 2012. 猛禽類保護の進め方(改訂版)−特にイヌワ 山形県文化環境部 1997. 鳥海山南麓イヌワシ生息調査報告. 日本山岳会 1997. 鳥海山南麓イヌワシ調査報告書. 鳥海山ワシタカ研究会 2000. 鳥海山南麓のイヌワシ. 日本イヌワシ研究会オフィシャルサイト. 図18. 冬の鳥海南麓 狩場となる積雪した草地にウサギの足跡 図15. 風衝草原で探餌するイヌワシ 図19. 春の鳥海北麓 図16. 秋の鳥海南麓 図20. 田植えを終えたばかりの水田と鳥海山(酒田市八幡地区) 図17. 秋の鳥海南麓 鶴間池 図21. 夏の鳥海南麓 鶴間池 ︱ 鳥 類 ︱ ︿寄 稿﹀ シ,クマタカ,オオタカについて−. 絶滅危惧種を中心とする鳥類 58 山形県の絶滅危惧種から重要なものを取り上 (タカ目タカ科) 図2 A∼C クマタカ げ,生態写真を中心に掲載した。 Spizaetus nipalensis (Hodgson, 1836) LC EN EN ︱鳥類︱ 国内希少野生動植物種 CITESII イヌワシ (タカ目タカ科) 図1 A∼C 全長雄72cm, 雌80cm,翼開長140∼165cm。 Aquila chrysaetos (Linnaeus, 1758) 鳥海山から高坂ダムにかけての北部の山地,神室 LC EN CR 国指定天然記念物 山系,御所山,蔵王山,葉山,月山,朝日連峰, 国内希少野生動植物種 CITESII 酒田市の鳥 飯豊連峰などの山系に生息するが多くはない。 全長雄81cm, 雌89cm,翼開長170∼213cm。 成鳥は全身が黒褐色で,後頭部は金褐色。県内で オオタカ (タカ目タカ科) 図2 D∼F は鳥海山∼丁岳,弁慶山地,神室山地,御所山, Accipiter gentilis (Linnaeus, 1758) LC EN VU 蔵王山,豪士山,吾妻山,摩耶山,月山,葉山, 国内希少野生動植物種 CITESII 朝日連峰,飯豊連峰で生息が確認されている。 全長雄50cm, 雌58cm,翼開長105∼130cm。 上面は灰色で胸部から腹部にかけて黄斑がある。 オオワシ (タカ目タカ科) 図1 D∼F 県内全域の低山の林を中心に比較的多くみられ Haliaeetus pelagicus (Pallas, 1811) る。冬季は餌となるカモの集まる河川,湖沼など VU NT VU 国指定天然記念物 で観察される。生息数は回復傾向にある。 国内希少野生動植物種 CITESII 全長90cm∼100cm,翼開長220∼245cm。く ハイタカ (タカ目タカ科) 図4 C∼D ちばしは大きく,黄色で下に湾曲している。飛翔 Accipiter nisus (Linnaeus, 1758) 時には翼後縁部がふくらみ,尾は長く白色のくさ LC NT EN CITESII び形をしている。北海道北部や東部,東北地方, 全長雄32cm, 雌39cm,翼開長60∼79cm。雄 中部地方まで飛来する。 成鳥は胸から腹にかけて橙褐色。雌は褐色の横斑 があり上面は灰褐色。本州以北の山地で繁殖し, オジロワシ (タカ目タカ科) 図1 G∼I 冬は大陸から飛来することもある。 Haliaeetus albicilla (Linnaeus, 1758) LC VU VU 国指定天然記念物 ツミ (タカ目タカ科) 国内希少野生動植物種 CITESI Accipiter gularis gularis (Temminck & Schlegel, 全長80∼90cm,翼開長180∼230cm。くちば 1844) しと虹彩は黄色でオオワシより小型。飛翔時の翼 全長雄27cm, 雌30cm,翼開長51∼63cm。雄 は長方形に近く,尾は白色の緩やかなくさび形。 成鳥は胸から脇は淡橙色。渡りの時期に山地や飛 北海道北部や東部で少数繁殖するが,東北・中部 島で比較的多く観察される。 LC EN NT 地方まで飛来することがある。 ハチクマ (タカ目タカ科) 図3 C∼D クロハゲワシ (タカ目タカ科) 図3F Pernis ptilorhyncus (Temminck, 1821) Aegypius monachus (Linnaeus, 1766) NT LC NT VU CITESII 全長雄57cm, 雌61cm,翼開長121∼135cm。 全長雄100∼110 cm,翼開長250∼290cm。 淡色型と暗色型の2型がある。雄成体は尾羽に太 日本で記録されたタカ科の鳥の中で一番大きい。 い2本の黒色帯,雌成体は2∼3本の黒色帯がある。 全身が黒褐色の羽毛で覆われ,頸部は襟巻き状の 低山から平地の森林で繁殖する。小動物や地バチ 羽毛で覆われる。頭部には羽毛がなく灰色の皮膚 等を地上でとらえる。県内の山地に生息する。 が露出している。嘴は太く,鉤状になり先端部は 黒い。 チュウヒ (タカ目タカ科) 図4A∼B Circus spilonotus Kaup, 1847 LC EN NT 全長雄48cm, 雌58cm,翼開長113∼137cm。 の縦斑や細かい黄斑がある。赤色型は全体に赤褐 羽色や模様に個体差がある。雄成体は上面が暗褐 色がかっている。短い羽角があり,虹彩は黄色。 色,下面は淡褐色で暗褐色の縦斑がある個体と, 九州以北の山地に夏鳥として飛来する。 上面が暗色で羽縁が白く,下面は淡色で胸に黒色 オオコノハズク(フクロウ目フクロウ科)図5D から暗褐色。県内では,南下北上の渡りの途中で Otus lempiji semitorques Te m m i n c k & 通過していく場合に見られることが多い。 Schlegel, 1844 (VU) DD サシバ (タカ目タカ科) 図4 E∼F に黒褐色,灰黒色,薄橙色,白色に斑が複雑には Butastur indicus (Gmelin, 1788) LC VU NT いる。くちばしは黒く,足指には羽毛がはえる。 全長雄47cm, 雌51cm,翼開長105∼115cm。 虹彩は橙色。県内では留鳥または漂鳥であるが渡 成鳥の上面は茶褐色,下面に太い茶褐色の横斑が りのとき飛島で見られる。 ある。県内各地の低山帯の林に夏鳥として渡来 し,その付近の田畑等で小動物をとらえる。 トラフズク(フクロウ目フクロウ科)図5 E∼F Asio otus otus (Linnaeus, 1758) NT ノスリ (タカ目タカ科) 図4G∼H 全長38cm,翼開長35cm。長い羽角をもつ。 Buteo buteo japonicas Temminck & Schlegel, 上面は灰褐色で,黒褐色や黄褐色,白色などの斑 1844 がある。下面は淡黄褐色の地に,黒い縦斑と交わ る横斑がある。県内全域の平野部に生息する。 トビ (タカ目タカ科) 図3E Milvus migrans lineatus (Gray, 1831) LC ハヤブサ (ハヤブサ目ハヤブサ科)図6 A∼C Falco peregrinus Tunstall, 1771 LC VU VU ミサゴ (タカ目ミサゴ科) 図3 A∼B 国内希少野生動植物種 CITESI Pandion haliaetus haliaetus (Linnaeus, 1758) 全長雄42cm, 雌49cm,翼開長84∼120cm。 LC EN NT 頭部,背,翼の上面は青黒色,下面は白地に細か 全長雄54cm, 雌64cm,翼開長155∼175cm。 い横縞がある。九州以北に生息し,おもに海岸の 上面は黒褐色で下面は白い後頭に短い冠羽があ 崖の岩棚で繁殖する。鳥をねらって急降下してと る。一部のダムで繁殖の記録がある。渡りの時期 らえることが多い。冬季は北方から南下する個体 に最上川,赤川その他の河川,湖沼で確認されて もあり,内陸の平野部の水田や湖沼でも見られ いる。 る。 フクロウ (フクロウ目フクロウ科)図5 A∼B チゴハヤブサ (ハヤブサ目ハヤブサ科) 図6D Strix uralensis Pallas, 1771 (EN) CITESII Falco subbutteo subbutteo Linnaeus, 1758 全長50cm,翼開長100cm。体上面は灰白色。 LC NT CITESII 胸は白く褐色の縦斑がある。本州北部からは, 全長雄33cm, 雌35cm,翼開長72∼84cm。上 亜種フクロウStrix uralensis hondoensis (Clark, 面は灰黒色で,胸から腹にかけて白色に黒色の縦 1907), 亜種モミヤマフクロウStrix uralensis 斑がならぶ。下面は赤褐色。頬には黒いひげ状の momiyamae Taka-Tsukasa, 1931が記録されてい 斑がある。夏鳥として飛来する。春と秋の渡りの るが亜種の分布が明確ではない。山形県ではモミ 時期には各地でみられることがある。平地の社寺 ヤマフクロウを絶滅危惧IB類としている。ここで の大木や鉄塔に営巣し,周辺を飛びまわり小鳥や は,種としてのフクロウとした。 昆虫をとらえる。冬季は南へわたる。 コノハズク (フクロウ目フクロウ科)図5C チョウゲンボウ (ハヤブサ目ハヤブサ科) Otus sunia japonicus Temminck & Schlegel, Falco tinnunculus Linnaeus, 1758 LC 図6 E∼F 1844 LC VU CITESII 全長 30∼35 cm,翼開長70∼80 cm。羽毛は 全長20cm,翼開長40cm。体色は灰色型で 赤褐色で黒斑がある。雄の頭と尾は青灰色,雌は は,上面が灰褐色で黒褐色や黄褐色の斑があり, 褐色で翼の先が尖る。農耕地で身近に見られる。 肩には黄白色の斑がある。下面は汚白色で黒褐色 樹洞で繁殖するが,人工物で営巣が増えている。 ︱鳥類︱ 全長24cm,翼開長57cm。雌がやや大。全体 59 の縦斑がある個体がある。雌成体は全身が淡褐色 60 ︱鳥類︱ 図 2-1. A ∼ C, イ ヌ ワ シ ; D ∼ F, オ オ ワ シ ; G ∼ I, オ ジ ロ ワ シ. 撮 影 : 今 井 正 (A); 長 船 裕 紀 (B ∼ C, G); 高 橋 雄 成 (D, F); 角 田 分 (E, H ∼ I). 61 ︱鳥類︱ 図 2-2. A ∼ C, ク マ タ カ ; D ∼ F, オ オ タ カ. 撮 影 : 今 井 正 (A); 長 船 裕 紀 (B ∼ C, E); 鳥 海 隼 夫 (D); 角 田 分 (F). 62 ︱鳥類︱ 図 2-3. A ∼ B, ミサ ゴ ; C ∼ D, ハ チ ク マ ; E, ト ビ ; F, ク ロハ ゲ ワ シ (剥 製). 撮 影 : 長 船 裕 紀 (A ∼ B, E); 鳥 海 隼 夫 (C); 高 橋 雄 成 (D). 63 ︱鳥類︱ 図 2-4. A ∼ B, チ ュ ウ ヒ ; C ∼ D, ハ イ タ カ ; E ∼ F, サ シ バ ; G ∼ H, ノ ス リ. 撮 影 : 長 船 裕 紀 (A ∼ C, E ∼ H); 角 田 分 (D). 64 ︱鳥類︱ 図 2-5. A ∼ B, フ ク ロ ウ ; C, コ ノ ハ ズ ク ; D, オ オ コ ノ ハ ズ ク ; E ∼ F, ト ラ フ ズ ク. 撮 影 : 長 船 裕 紀 (A ∼ B); 角 田 分 (C); 高 橋 雄 成 (D);鳥海隼夫 (E ∼ F). 65 ︱鳥類︱ 図 2-6. A ∼ C, ハ ヤ ブ サ ; D, チ ゴ ハ ヤ ブ サ ; E ∼ F, チ ョ ウ ゲ ン ボ ウ. 撮 影 : 長 船 裕 紀 (A ∼ C); 後 藤 拓 弥 (D); 鳥 海 隼 夫 (E ∼ F). (カモ目カモ科) 図7A コクガン 肩羽に毛状の長い飾り羽を生じる。嘴と脚は黒色 。 Branta bernicla (Linnaeus, 1758) LC VU VU 全長61cm。雌雄同色。体色は頭部から頸,胸 ヒクイナ (ツル目クイナ科) 図8 G∼H 部は黒色。喉に白い輪状の斑紋がある。背上面は Porzana fusca erythrothorax (Temminck & 黒褐色で細い淡色の羽縁がある。頸は短く太い。 Schlegel, 1849) LC NT CR 全長23cm。後頸,背,翼は暗褐色で,前頭か 66 ︱鳥類︱ トモエガモ (カモ目カモ科) 図7 C∼D ら胸,腹は赤褐色。下尾筒には白い線があり,脚 Anas formosa Georgi, 1775 は赤褐色,嘴は青黒色。 LC VU NT 全長約40cm。雄は頭頂から後頭まで黒褐色。 顔には黄白色の地に緑黒色のともえ形の模様があ ケリ (チドリ目チドリ科) 図9B る。雌は全体的に褐色。 Vanellus cinereus (Blyth, 1842) オシドリ (カモ目カモ科) 図7 E∼G ヘラシギ (チドリ目シギ科) Aix galericulata (Linnaeus, 1758) Eurynorhynchus pygmeus (Linnaeus, 1758) LC DD NT 県の鳥 CR CR CR 全長約45cm。雄は紫赤褐色も冠羽や参列風切 全長15cm。くちばしの先がへら状になってい 羽のイチョウ羽などが目立つ。雌は地味で目立た る。雌雄同色。夏羽は頭から胸にかけて赤褐色, ない。繁殖期をすぎると雄も地味な色になる。 冬羽は上面が灰色で白い羽縁があり下面は白い。 オオハクチョウ (カモ目カモ科) 図7B コアジサシ (チドリ目カモメ科) 図9C∼D Cygnus cygnus (Linnaeus, 1758) Sterna albifrons sinensis Gmelin, 1789 LC EN LC LC NT 全長140∼165cm。全身が白い。くちばしは 全長28cm。背と翼の上面は淡青灰色で,体と 先端が黒く,基部は黄色。黄色の部分が大きく, 尾は白い。頭と過眼線は黒色。額は白い。夏羽で その先端は三角形にとがって黒色部に食い込む。 は嘴が黄色で先端が黒い。脚は橙黄色。 幼鳥は全身が淡い淡褐色。 ウミガラス (チドリ目ウミスズメ科) 図9E オオハム (アビ目アビ科) 図11H Uria aalge inornata (Salomonsen, 1932) Gavia arctica viridigularis Dwight, 1918 LC VU LC CR CR 国内希少野生動植物種 全長70cm。雌雄同色。成鳥冬羽は胸から腹は 全長40cm。雌雄同色。夏羽では頭,胸,背が 白色で背中は光沢のある黒色。雨覆に小さな白斑 黒褐色で,胸から腹にかけての体下面,下尾筒は がある。 白色。嘴と脚は黒色。冬羽は顔の下半分と側頸, 体下面が白色で眼の後方に細い黒い線がある。 ヨシゴイ (ペリカン目サギ科) 図8 A∼B Ixobrychus sinensis sinensis (Gmelin, 1789) ハシブトウミガラス(チドリ目ウミスズメ科 ) LC NT VU Uria lomvia arra (Pallas, 1811) LC CR 図9F 全長36cm。全身が黄褐色。雄は頭上が黒く, 全長43cm。雌雄同色。夏羽では頭,胸,背が 水切羽は黒色。雌は下面に不明瞭な褐色の縦しま 黒褐色で,胸から喉にかけて三角形の白色部があ がある。幼鳥は上下面に縦斑がある。 る。腹から体下面は白色。冬羽の顔は黒く,喉と 前頸,体下面が白色。 クロサギ (ペリカン目サギ科) 図8 C∼D Egretta sacra sacra (Gmelin, 1789) LC VU ウミスズメ (チドリ目ウミスズメ科) 図9G 全長約62cm。全身が黒色型と白色型がある。 Synthliboramphus antiquus (Gmelin, 1789) 嘴や足の色は黄褐色から黒色など個体変異がある 。 LC CR CR 全長23cm。頭部は大きい。夏羽の上面は灰黒 チュウサギ (ペリカン目サギ科) 図8 E∼F 色,冬羽は黒色。下面は白い。 Egretta intermedia intermedia (Wagler, 1829) LC NT NT カンムリウミスズメ(チドリ目ウミスズメ科 ) 全長約70cm。全身白色で繁殖期には胸および Synthliboramphus wumizusume (Temminck, 1836) VU VU DD 図9H Schegel, 1845 全長24cm。雌雄同色。夏羽は頭頂部に黒い冠 全長19cm。雄は額,目先,過眼線は黒い。頭 羽がある。上面は黒色,胸から体下面は白い。 は青灰色。背面は赤褐色で,黒くて細い黄斑があ LC CR CR る。 ケイマフリ (チドリ目ウミスズメ科) 図9A Cepphus carbo Pallas, 1811 VU DD アカモズ (スズメ目モズ科) 図11B 眼のまわりが白い。上下の嘴基部に1つずつの白 LC EN EN 点がある。嘴は黒く,脚は赤い。冬羽は喉から 全長20cm。雌雄ほぼ同色。上面赤褐色,下面 頬,側頸から体下面が白くなる。 は白く胸から脇にかけて淡い橙色。 ヨタカ (ヨタカ目ヨタカ科) オオセッカ (スズメ目センニュウ科) 図 Caprimulgus indicus Lahtam, 1790 LC NT EN 11D Locustella pryeri pryeri (Seebohm, 1884) 全長29cm。全体が黒褐色で,白色,褐色,黒 NT EN DD 国内希少野生動植物種 色などの複雑な枯れ葉模様がある。雄は頸線が白 全長14cm。頭から背,翼にかけてやや赤みの く,喉や風切,尾羽外側先端に白斑がある。 ある褐色で,黒褐色の縦斑がある。下面は白く尾 はくさび形。 ブッポウソウ (ブッポウソウ目ブッポウソウ科) 図10A セッカ (スズメ目セッカ科) Eurystomus orientalis calonyx Sharpe, 1890 Cisticola juncidis brunniceps (Temminck & LC EN CR Schlegel, 1850) 全長30cm。雌雄同色。全身光沢のある青黒色 。 全長13cm。雌雄同色。頭部から背,腰にかけ LC CR て黄褐色で,頭頂部に黒の縦斑がある。喉から体 ヤマセミ (ブッポウソウ目ブッポウソウ科) 下面は白色で,腹側面は黄褐色。尾は長く暗褐色。 Megaceryle lugubris pallida (Momiyama, 1927) LC VU 図10B キバシリ (スズメ目キバシリ科) 図11E 全長38cm。上面は白と黒のまだら模様で,頭 Certhia gamiliaris japonica Harten, 1897 の羽毛は長く冠羽になる。雄は胸に黄褐色と黒 LC EN 色,雌は黒色の斑点がある。腹は白色。 全長14cm。雌雄同色。上面の羽色は樹皮に似 ている。くちばしは細長く,下に湾曲している。 オオアカゲラ (キツツキ目キツツキ科) 図 10C Dendrocopos leucotos stejnegeri (Kuroda, コジュリン (スズメ目ホオジロ科) 図11F 1921) LC NT Emberiza yessoensis yessoensis (Swinhoe, 1874) NT VU EN クマゲラ (キツツキ目キツツキ科) 図10D 全長15cm。雄成鳥の夏羽は,頭部が黒く,背 Dryocopus martius martius (Linnaeus, 1758) は淡褐色と黒色の縦縞がある。頸から腹部下面は LC VU DD 汚白色で腰は赤褐色。雌成鳥の夏羽は,頭部が黒 褐色で淡褐色の頭央線があり,眉斑と頬線は汚白 サンショウクイ(スズメ目サンショウクイ科) 色。 Pericrocotus divaricatus divaricatus (Raffles, 1822) LC VU VU 図11C オオジュリン (スズメ目ホオジロ科) 図 全長20cm。体は細く尾が長い。雄の頭部は白 11G Emberiza schoeniclus pyrrhulina (Swinhoe, いが後頭部や過眼線は黒い。上面は青灰色で翼は 1876) LC EN 黒く,下面全体が白色。雌は後頭から後頸は灰 全長16cm。雄成鳥の夏羽は,頭部から顔,上 色。嘴と脚は黒色。 腹部まで黒い。頬線から後頸,胸,腹部は白い。 背や翼は赤褐色で黒褐色の縦縞がある。雌成鳥は チゴモズ (スズメ目モズ科) 図11A 頭部の黒色はなく,眉斑と頬線は白い。 Lanius bucephalus bucephalus Temminck & 67 ︱鳥類︱ Lanius cristatus superciliosus Latham, 1802 全長37cm。雌雄同色。夏羽では全身が黒色で 68 ︱鳥類︱ 図 2-7. A, コクガン ; B, オ オ ハ ク チ ョ ウ ; C ∼ D, ト モ エ ガ モ ; E ∼ G, オ シ ド リ. 撮 影:高 橋 雄 成 (A);長 船 裕 紀 (B, G);角 田 分 (C ∼ F). 69 ︱鳥類︱ 図 2-8. A ∼ B, ヨシ ゴ イ ; C ∼ D, ク ロ サ ギ ; E ∼ F, チ ュ ウ サ ギ ; G ∼ H, ヒ ク イ ナ. 撮 影 : 角 田 分 (A, D ∼ F); 高 橋 雄 成 (B ∼ C, G ∼ H). 70 ︱鳥類︱ 図 2-9. A, ケイマフ リ ; B, ケ リ ; C ∼ D, コ ア ジ サ シ ; E, ウ ミ ガ ラ ス ; F, ハ シ ブ ト ウ ミ ガ ラ ス ; G, ウ ミ ス ズ メ (剥 製); H, カ ン ム リ ウ ミ ス ズメ (剥製). 撮影 : 角 田 分 (A, C); 鳥 海 隼 夫 (B, D); 高 橋 雄 成 (E ∼ F). 71 ︱鳥類︱ 図 2-10. A, ブッポ ウ ソ ウ ; B, ヤ マ セ ミ ; C, オ オ ア カ ゲ ラ (剥 製); D, ク マ ゲ ラ (剥 製, 山 形 大 学 理 学 部). 撮 影 : 高 橋 雄 成 (A); 長 船 裕 紀 (B). 72 ︱鳥類︱ 図 2-11. A, チゴモ ズ ; B, ア カ モ ズ ; C, サ ン シ ョ ウ ク イ ; D, オ オ セ ッ カ ; E, キ バ シ リ ; F, コ ジ ュ リ ン ; G, オ オ ジ ュ リ ン ; H, オ オ ハ ム. 撮 影 :鳥海隼夫 (A); 角 田 分 (B ∼ C,F ∼ G); 鈴 木 崇 文 (D ∼ E); 高 橋 雄 成 (H). 絶滅した鳥 かつては北海道・本州・伊豆諸島・佐渡島・隠 Nipponia nippon (Temminck, 1835) 岐諸島・四国・九州・琉球諸島といった日本各地 73 EN EW 国指定特別天然記念物 のほか,ロシア極東,朝鮮半島,台湾,中国と東 国内希少野生動植物種 アジアの広い範囲に生息していた。日本では東北 ︱鳥類︱ 地方や日本海側に多く見られた。 図2-14. 朱鷺(両羽博物図譜より) しかし,乱獲や開発によって19世紀から20世 紀にかけて激減し,朝鮮半島では1978年の板門 図2-12. トキ(斎藤報恩会博物館) 店,ロシアでは1981年のウスリー川で観察され 全長約76cm。翼開長約130cm。体重雄1,800 たのが最後である。日本でも2003年に最後の日 ∼2,000g,雌1,450∼1,600g。顔は濃いピンク 本産トキ「キン」が死亡した。野生では中国(陝 色(トキ色)の皮膚が露出する。くちばしは下方 西省など)に約1,000羽が生息している。日本 に湾曲。後頭部にやや長い冠羽がある。全身は白 の佐渡島において2008年秋から2012年秋まで 色,春から夏にかけての繁殖期には首すじから黒 に人工繁殖のトキ計107羽が放鳥されている。 い分泌物が出てからだに塗りつけるため,頭から 山形県でも最初の放鳥後に観察されている(図 背のあたりが灰黒色になる。翼の下面はトキ色を 2-15)。飼育下では中国に620羽,日本に180 している。脚もトキ色。 羽,韓国に13羽がおり人工繁殖が進められてい る。 図2-13. 野生のトキ(佐渡,撮影:佐藤春雄,1966.11.23) 図2-15. 放鳥トキの飛来(米沢市,撮影:鳥海隼夫) (ペリカン目トキ科) トキ 迷 鳥 74 ︱鳥類︱ (コウノトリ目コウノトリ科) コウノトリ 太平洋戦争前後の食料不足の中で食用にされたこと Ciconia boyciana Swinhoe, 1873 もあり,1956年には20羽にまで減少した。そのた EN CR 国指定特別天然記念物 め,コウノトリは同年に国の特別天然記念物に指定 国内希少野生動植物種 された。 1966年に福井県小浜市の国内最後の野生繁殖地 の個体が姿を見せなくなり,1971年5月25日には 豊岡市に残った国内最後の一羽である野生個体を保 護するがその後死亡。このため人工飼育以外のコ ウノトリは国内には皆無となり,さらには1986年 2月28日に飼育していた最後の個体が死亡し,国内 の野生個体群は絶滅した。しかし,これ以降も不定 期に渡来する複数のコウノトリが観察され続けてい る。 図2-16. コウノトリ(山形県,撮影:角田分) 形態 全長110∼115cm。翼開長160∼ 200cm。体重雄4∼6kg。羽は白と金属光沢のあ る黒。くちばしは黒みがかった濃い褐色。脚は赤 く,目の周囲も赤い。 生態 成鳥になると鳴かなくなり,かわりに「ク ラッタリング」と呼ばれる行為が見受けられる。 くちばしを叩き合わせるように激しく開閉して音 を出す行動で,ディスプレイや仲間との合図に用 いられる。樹上に雌雄で造巣する。3∼5個の卵 図2-17. 飼育されるコウノトリ(2013.5, 多摩動物公園) を産み,抱卵期間は30∼34日である。抱卵,育 多摩動物公園では,中国から譲り受けて人工飼育 雛は雌雄共同でおこなう。雛は約58∼64日で巣 を続けていた結果,1988年4月6日に国内初の人工 立つ。 繁殖に成功した。豊岡市のコウノトリ飼育場でも, おもにザリガニなどの甲殻類やカエル,魚類を この国内繁殖野生個体群の絶滅の約7か月前である 捕食する。ネズミなどの小型哺乳類を捕食するこ 1985年7月27日に当時のソ連から幼鳥6羽をもらい ともある。 受けており,1989年5月16日に人工繁殖に成功し 分布 東アジアに限られる。総数も推定2,000∼ ている。これ以後,毎年の繁殖に成功している。 3,000羽と少なく,絶滅の危機にある。中国東北 現在,豊岡市コウノトリの郷公園に約100羽,多摩 部地域やアムール・ウスリー地方で繁殖し,中国 動物公園に約50羽,その他約50羽が飼育されてい 南部で越冬する。渡りの途中に少数が日本を通過 る。 することもある。山形県でもまれにみられること 2005年までに豊岡市のコウノトリの郷公園周辺 がある。 地域にコウノトリの生息可能な環境が整備されつつ その他 日本列島にはかつて留鳥としてコウノト あり,周辺の農家も農薬の散布を控え,無農薬栽培 リが普通に生息していたが,明治期以後の乱獲や に切り替える等の協力をしている。2005年9月24 巣を架ける木の伐採などにより生息環境が悪化, 日には世界初の放鳥が行われ,34年ぶりにコウノ トリが大空に羽ばたくこととなった。 きるようになる。 食性は雑食で,昆虫,甲殻類,貝類,魚類,カ Grus japonensis (Müller, 1766) 分布 日本(北海道東部),ロシア南東部,中華 EN VU 国指定特別天然記念物 人民共和国,大韓民国北部,朝鮮民主主義人民共 国内希少野生動植物種 CITESI 和国。夏季に中華人民共和国北東部,アムール川 やウスリー川中流域で繁殖し,冬季になると朝鮮 半島,長江下流域に南下し越冬する。 日本では周年生息する。おもに北海道に生息す るが,山形県にも飛来することがある。慶応から 明治初年の記録として両羽博物図譜にマナヅル, ナベヅルとともに記述がある。 図2-18∼19. タンチョウ(撮影:角田分) 形態 全長125∼152cm。翼開長約240cm。体 重6.3∼9kg。全身の羽は白色。眼先から喉,頸 部にかけての羽は黒い。次列風切や三列風切は黒 い。頭頂には羽毛がなく赤い皮膚が露出する。虹 彩は黒や暗褐色。くちばしは長く,黄色や黄褐 色。脚は黒い。 図2-20. 丹頂(甲),真ナ鶴(乙),牛鶴(丙)(両羽博物図譜よ 生態 湿原,湖沼,河川などに生息する。冬季に り) は家族群もしくは家族群が合流した群れを形成す その他 日本では1935年に繁殖地も含めて国の る。オスが長く1回鳴いたあとに,メスが短く2 天然記念物,1952年に「釧路のタンチョウ」 ∼3回鳴くことを繰り返し,これにより縄張りを として特別天然記念物,1967年に地域を定めず 主張したり,ペアを維持したりするのに役立つと に種として特別天然記念物に指定されている。 考えられている。繁殖形態は卵生。繁殖期に1∼ 1993年に種の保存法施行に伴い国内希少野生 7km2の縄張りを形成する。湿原や浅瀬に草や木 動植物種に指定されている。北海道での1952 の枝などを積み上げた直径150cm,高さ30 cm ∼1953年における生息数は33羽,1962年にお に達する皿状の巣を作り,日本では3∼5月に1∼ ける生息数は172羽,1988年における生息数は 2個の卵を産む。雌雄交代で抱卵し,抱卵期間は 424羽,2000年における生息数は740羽。 31∼36日。雛は孵化してから約100日で飛翔で 75 ︱鳥類︱ エル,植物の茎,種子などを食べる。 タンチョウ (ツル目ツル科) 参考文献 (おもなもの) 76 ︱参考文献・協力者および資料提供者︱ 阿部 永・石井信夫・伊藤徹魯・金子之史・前田 T. 2010 . The Wild Mammals of Japan 2nd 喜四雄・三浦慎悟・米田政明 2008. 日本の哺 print. Shoukadoh, Kyoto, 544 pp., 4 maps. 乳類 改訂2版. 東海大学出版会, 神奈川, 207 pp. 大津 高(編) 2004. 山形県陸産淡水産動物目 橋本賢助 1939. 山形縣の哺乳類. 山形縣教育, 録改訂版. 山形県動物環境調査会, 山形, 309 pp. 585: 12-18. 山形県(編) 2000. 第54回愛鳥週間「全国野鳥 環境省自然環境局野生生物課(編) 2002a, b. 改 保護のつどい」記念誌 やまがたの野鳥. 山形県 訂・日本の絶滅のおそれのある野生生物―レッ 文化環境部環境保護課, 山形, 192 pp. ドデータブック― 1 哺乳類, 179 pp., 2 鳥類, 山形県希少野生生物検討委員会動物部会(編) 279 pp., ㈶自然環境研究センター, 東京. 2003. レッドデータブックやまがた 山形県の 日本鳥学会(編) 2012. 日本鳥類目録改訂 第7 絶滅のおそれのある野生動物. 山形県文化環境 版. 日本鳥学会, 兵庫, 439 pp. 部環境政策推進室環境保護課, 山形, 303 pp. Ohdachi, S. D., Ishibashi, Y., Iwasa, M. A. & Saitoh, 協力者 および 資料提供者 (敬称省略) 相澤周一,伊藤健雄,今井正,遠藤亨,岡部夏 環境省猛禽類保護センター,国立科学博物館,斎 雄,奥山武夫,桶舎富士子,長船裕紀,押切智 藤報恩会博物館,酒田市立光丘文庫,世界淡水魚 紀,角田分,加藤雄祐,小城伸晃,後藤拓弥,後 園水族館,全国科学博物館振興財団,仙台市科学 藤亮,庄司光幸,鈴木かおり,鈴木弘二,鈴木崇 館,東京国立博物館,西川町大井沢自然博物館, 文,高橋雄成,鐸木能光,玉手英利,縮文夫,鳥 山形県環境エネルギー部みどり自然課,山形大学 海隼夫,長澤一雄,中村夢奈,羽根田裕,原田裕 理学部,山形大学附属博物館,やまがたヤマネ研 行,半澤直人,東英生,本間憲一,水落朋子,村 究会,山形の野生動物を考える会 上愛,由井正敏,横倉明 やまがたの野生動物 平成 25 年 7 月 31 日 発行 発 行 山形県立博物館 〒 990-0826 山形市霞城町 1 番 8 号 編 集 吉田 哉 印 刷 寒河江印刷株式会社 本書は,一般財団法人全国科学博物館振興財団の活動助成を受け発行した。 © 山形県立博物館 Wild Animals of Yamagata, Yoshida, H. (ed.) 2013
© Copyright 2026 Paperzz


![日刊[華鐘通信] - 上海中经互联网络有限公司](http://s3.paperzz.com/store/data/005492934_1-a7ac04d9eb457d84c42bf7d87127b81a-250x500.png)