ANM176®が認知症の予防と改善に役立つ可能性について
2015 年 6 月 25 日
認知症の予防と改善を推進する会
(目次)
ページ
はじめに ------------------------------------------------------------------------------------------ 1
健忘は認知症の始まりか? ------------------------------------------------------------------ 4
(認知機能とは) ------------------------------------------------------------------------- 4
(認知症について) ---------------------------------------------------------------------- 5
(中枢神経系について) ----------------------------------------------------------------- 6
(脳関門) ---------------------------------------------------------------------------------- 9
体の中で行われている生命維持活動 について ---------------------------------------- 11
(生物は特有な成分で構成された細胞から成り立っている) --------------- 11
(細胞内へのシグナリング) -------------------------------------------------------- 13
(キナーゼによるシグナリング) -------------------------------------------------- 15
(ユビキチン化とプロテアソーム) ----------------------------------------------- 17
(遺伝子の DNA 情報からタンパク質の合成) -------------------------------- 18
生物の恒常性維持について ---------------------------------------------------------------- 22
(細胞ストレスについて) ----------------------------------------------------------- 24
(小胞体ストレス応答) -------------------------------------------------------------- 25
(酸化ストレス応答) ----------------------------------------------------------------- 26
(虚血ストレス応答) ----------------------------------------------------------------- 28
(炎症とは) ----------------------------------------------------------------------------- 29
(細胞毒性)------------------------------------------------------------------------------- 30
(ストレス耐性) ----------------------------------------------------------------------- 32
(細胞ストレスと病気) -------------------------------------------------------------- 34
(神経の中の記憶) -------------------------------------------------------------------- 35
(記憶の分子機構) -------------------------------------------------------------------- 37
アルツハイマー病 ---------------------------------------------------------------------------- 43
(βアミロイドの分泌と APP の役割) ------------------------------------------ 46
(βアミロイドの神経毒性作用) ------------------------------------------------- 48
(βアミロイド神経毒性はどのようにして起こるのか?) ---------------- 51
(βアミロイドの分解) -------------------------------------------------------------- 53
(タンパク質の凝集体とオートファジー) -------------------------------------- 55
(神経変性疾患について) ----------------------------------------------------------- 58
(アルツハイマー病の発症リスク) ----------------------------------------------- 61
(βアミロイドがアルツハイマー病を発症させる仕組み) ----------------- 63
アルツハイマー病は生活習慣病か? ---------------------------------------------------- 67
(糖尿病とは)--------------------------------------------------------------------------- 68
(インスリンシグナリングとグリコーゲン合成)---------------------------- 69
i
(インスリンシグナリングの役割)------------------------------------------------ 70
(インスリン抵抗性)------------------------------------------------------------------ 71
(インスリンは脳でも働いている)------------------------------------------------ 73
現在のアルツハイマー病治療方法 ------------------------------------------------------ 75
アルツハイマー病にどのように備えたらよいか? --------------------------------- 78
(アルツハイマー病の予防は可能か?) ---------------------------------------- 79
(サプリメントはアルツハイマー病に有効か?) ---------------------------- 82
フェルラ酸にはβアミロイド神経毒性を抑制する効果がある ------------------ 86
(フェルラ酸とはどんなもの?) ------------------------------------------------- 89
(フェルラ酸の吸収と排泄) ------------------------------------------------------- 92
(フェルラ酸の生体内での機能) ------------------------------------------------- 93
(フェルラ酸はアルツハイマー病の改善に役立つか?) ------------------- 95
ANM176Ⓡ がアルツハイマー病に役立つ可能性について ----------------------- 97
(ANM176Ⓡ とは) ----------------------------------------------------------------- 97
(ANM176Ⓡの臨床試験の経緯と結果の概要) ------------------------------- 101
(ANM176Ⓡには製造ノウハウがある) --------------------------------------- 104
(ANM176Ⓡの品質管理と安全性) --------------------------------------------- 105
(ANM176Ⓡはアルツハイマー病の予防と改善に役立つ)---------------- 107
最後に ---------------------------------------------------------------------------------------- 113
ii
はじめに
ここで述べる「ANM176」とは、韓国にある翰林大学生薬研究所(Institute of Natural
Medicine, Hallym Universty)の Song 教授の指導の下で Yan 博士らが行った、漢方薬の
生薬の中からアルツハイマー病(以下では Alzheimer’s disease から AD と表記します)の
治療に効果がある成分を探索する研究から開発された製剤の名前です。東洋に古くからあ
る漢方では治療方法の文献が膨大に集積されています。ところが、認知症 ≪以前は痴呆(チ
ホウ)と言っていました≫ に該当する文献は見当たらず、ただ、当帰(以下では「トウキ」
と表記します)と呼ばれる生薬だけが「もの忘れ」に効果があると伝承(デンショウ)されてい
ました。伝承とは、文献に記載できる明確な効果がみられないが、可能性が否定できないと
いう程度のことを言います。
AD を引き起こしていると考えられているベータアミロイド(βamyloid)と呼ばれるタ
ンパク質(以下では「βアミロイド」と表記します)をマウスの脳内に注入すると、記憶
や学習力を低下させます。この記憶や学習力を低下させる作用は、βアミロイドが持って
いる神経毒性作用(以下では「βアミロイド神経毒性」と言います)が原因で起こります
(48 ページ「βアミロイドの神経毒性」)
。Yan 博士らは、βアミロイドがマウスの記憶や学習力
を低下させる作用を定量化する方法を編み出し、βアミロイド神経毒性の程度を計測しま
した。この方法を用いてトウキに含まれているβアミロイド神経毒性抑制成分を探索した
結果、驚いたことに、それは 1 種類だけでなく、13 種類もあったのです(97 ページ
「ANM176Ⓡとは」)。この研究をベースに、トウキと同じ属でガーデンアンゼリカと呼ばれ
るハーブ系の植物に含まれる 13 種のβアミロイド神経毒性抑制成分を含み、βアミロイ
ド神経毒性を最も効果的に抑制するサプリメント用製剤の ANM176®が誕生しました。
この 13 種のβアミロイド神経毒性抑制成分の中で、フェルラ酸(Ferulic acid)は残り
の 12 成分と異なり、どんな植物のどんな部位にも幅広く含まれている成分です。そこで
Yan 博士らは、このフェルラ酸を取り上げ、βアミロイド神経毒性をフェルラ酸が効果的
に抑制する詳細を、マウスを用いた実験で示しました(86 ページ「フェルラ酸はβアミロイド神
経毒性を抑制する効果がある」)。この実験でマウスに投与しβアミロイド神経毒性を抑制した
フェルラ酸の量をヒトに換算すると、本来であれば食事で摂取可能な量でした(89 ページ
「フェルラ酸とはどんなもの?」)。と言うのも、類人猿から進化したヒト(人間を生物として
言う場合は「ヒト」、個人として言う場合は「人」と表記します)は米や麦などの穀物を
食べるようになり、穀物のヌカ部分にあるフェルラ酸を吸収してきたはずだからです。ヌ
カにはヒトでも吸収できるフェルラ酸が含まれていますが、ヌカ以外の植物のどこにでも
あるフェルラ酸のほとんどはヒトでは吸収できない形となっています(92 ページ「フェルラ酸
の吸収と排泄」)。この 100 年ほどで、ヌカ部分を取り除いた白米や精白小麦粉が一般化し
て、今ではヌカが付いたままの玄米(ゲンマイ)や地粉(ジゴナ)は、ほとんど流通しなくな
っています。ヒトに進化してから日常的に吸収してきたフェルラ酸がβアミロイドの神経
毒性を効率よく抑制することを示した Yan 博士らが行った試験結果を改めて考えると、愕
1
ANM176®が認知症の予防と改善に役立つ可能性について
然(ガクゼン)とするものがあります。この試験で神経毒性を引き起こしたβアミロイドの
濃度は、高齢になると誰の脳にもあり得る濃度で、フェルラ酸が吸収できていない現代の
高齢者の脳の中では、日常的にβアミロイド神経毒性が起こっている可能性が考えられる
からです。
3 人に 1 人が 65 歳以上の高齢化社会を迎えようとしている日本において、高齢者の 10
人に 1 人は既に認知症を発症しており、5 年以内に発症する可能性がある予備軍を含めると
4 人に 1 人は認知症になると推計されています(5 ページ「認知症について」)。このような深刻
な課題を抱えている認知症の増加は、単に高齢化だけに原因があるのではなく、生活環境の
変化も関与していると言われています。
最近、AD と生活習慣病の糖尿病
(以下では Diabetes
Mellitus から DM と表記します)が深く関係していると考えられています(67 ページ「アルツ
ハイマー病は生活習慣病か?」)
。AD 患者のほとんどは遺伝とは関係ないが高齢になると発病す
る老年性アルツハイマー型認知症(以下では senile dementia of Alzheimer type から SDAT
と表記します)で、DM と同じ生活習慣病と考えられます(67 ページ「アルツハイマー病は生活
習慣病か?」)。AD も DM も、その患者数は近年急速に増加していますが、これら患者のほ
とんどは、発症の原因に遺伝は関係していません。それにもかかわらず患者が急増している
原因には、高齢化と環境の変化が考えられます。一般に生活習慣病の発症には食事や日常の
運動(活動)が関係していると考えられていますが、さらに、高齢になるとストレス応答の
能力が低下し、内因性の炎症が関与してこれらの生活習慣病を引き起こすと考えられます
(34 ページ「細胞ストレスと病気」)。また、高齢になると低下するストレス耐性も発症の一因と
なっている可能性も考えられます。ストレス耐性とは、一言では「体の抵抗力」です。食べ
物が不足した飢餓状態になった時、病原菌や毒物に遭遇した時、寒さや暑さが続くなどの異
常気象など、異常な事態に遭遇すると体はストレスとして受け止めます。これらの異常事態
に対処するストレス応答と、ストレス応答によって異常事態を乗り越えるストレス耐性の
能力が生まれながらに備わっています(32 ページ「ストレス耐性」)。
このストレス耐性の能力は、このストレス耐性に関連する転写因子 Nrf2 のシグナリン
グとともに高齢になると低下します(79 ページ「アルツハイマー病の予防は可能か?」)。また、
DM の発症に関係するインスリンシグナリング経路も高齢になると低下して、AD の発症
にも関係すると考えられています(107 ページ「ANM176Ⓡはアルツハイマー病の予防と改善に役立
つ」)。
食品に含まれるフェルラ酸など一部の成分は、高齢になって低下するストレス耐性を増
強できる可能性があります。細胞内にある Keap1 と呼ばれるタンパク質は Nrf2 と結合し
て細胞内に引き止め、Nrf2 が細胞核に移行して転写に機能することを抑制しています。フ
ェルラ酸など Nrf2 の転写を活性化する一部の食品に含まれる成分は Keap1 に作用し、
Keap1 から Nrf2 を引き離して核内への移行を促進し、Nrf2 の転写を促進させます(79 ペ
ージ「アルツハイマー病の予防は可能か?」)
。
一方、インスリンシグナリング経路は細胞の生長パターンを抑制しているシグナリング
2
ANM176®が認知症の予防と改善に役立つ可能性について
キナーゼ GSK3βをリン酸化して、GSK3βが抑制していた Nrf2 の転写活性など細胞の生
長に関与する様々な機構を活性化します(70 ページ「インスリンシグナリングの役割」)。
フェルラ酸は、高齢になって低下するインスリンシグナリングを、GSK3βとは無関係
な Keap1 のルートで、インスリンシグナリング経路が十分に活性化できなかった Nrf2 の
転写活性を補強すると考えられます(93 ページ「フェルラ酸の生体内での機能」)。
臨床試験で認知症患者が朝と晩のそれぞれに使った ANM176®の量は 300mg で、
ANM176®の 300mg に配合されたフェルラ酸は 100mg です。このフェルラ酸 100mg は、
本来であればヒトが穀物から 1 日に摂れる量に匹敵します。また、この量をマウスの生理
作用に換算した試験でβアミロイド神経毒性を効果的に抑制したことは、本来であれば食
事で摂(ト)り得るフェルラ酸を補給することによって、AD を予防できる可能性が考えら
れます。またフェルラ酸は、インスリンシグナリング経路を補助できることから、DM に
限らず多くの生活習慣病の予防にも役立つ可能性があります(89 ページ「フェルラ酸とはどん
なもの?」)。
一方、フェルラ酸は AD における記憶力や認知機能の低下の改善に有効である可能性
が示唆されていますが、フェルラ酸に抗炎症作用があるガーデンアンゼリカに含まれる
特定クマリン(coumarin)類を配合することによって AD における神経細胞(ニュー
ロン)死の抑制に有効と考えられます。AD の進行には炎症が関与していると考えられる
からです(95 ページ「フェルラ酸はアルツハイマー病の改善に役立つか?」)。トウキやガーデンア
ンゼリカに含まれるβアミロイド神経毒性を抑制する成分は、フェルラ酸の他に 12 種の
クマリン類です。このクマリン類の一部には抗炎症作用があります(97 ページ「ANM176®と
は」)。AD 用の製剤を開発するにあたり、新たな問題点が分かってきました。それは、βア
ミロイド神経毒性を抑制し抗炎症作用がある 13 種の成分が含まれているトウキやガーデ
ンアンゼリカは稀で、原料を安定的に確保することが難しいことです。
AD と DM には共通したインスリンシグナリング経路の障害が考えられ(73 ページ「イン
スリンは脳でも働いている」)、ANM176®が、どのような作用によって、AD の予防や改善に
有用と考えられるか、また、なぜ単にフェルラ酸とガーデンアンゼリカを配合しただけで
は、ANM176®と同じ効果が期待できないかを、最近の AD 関連の研究成果から、できる
だけ分かりやすく述べるように努めました。
本紙の主題は 67 ページ「アルツハイマー病は生活習慣病か?」から始まりますが、それ
を補うため、前半は、生物学の基礎を述べました。前半部分は飛ばして主題から読みはじめ
る場合は、注に合わせて前半部分を参考にされるとよいでしょう。また、本文に入れると雑
多になり、しかし、理解するに必要な事柄については下部に青字で解説しました。
97 ページ「ANM176®がアルツハイマー病に役立つ可能性について」以降は、AD の予防
と改善に有用と考えられる ANM176®の作用について掘り下げました。
3
ANM176®が認知症の予防と改善に役立つ可能性について
健忘は認知症の始まりか?
50 歳を越えると誰でも「もの覚え」が悪くなり、また、
「もの忘れ」も起こるようになり
ます。しばらく使っていない言葉や映画俳優の名前など「覚えているはず」が、なかなか出
てこなくなります。また、
「どこかにしまったはず」や「置いたはず」の物を、よく探すよ
うになります。別の部屋に何かを取りに行ったとき、何を取りに来たのか思い出せないこと
もしばしば起こります。このように「はず」の名前や物が思い出せないことが頻繁になると、
認知症の前兆ではないかと不安になることは誰にでもあることです。不安になって「もの忘
れ外来」の診察を受けると、認知症の専門医によって認知機能が正常であるか否かが診断さ
れます。認知機能の検査は一種の心理テストで行うもので、血液検査などによる指標では判
りません。認知機能は記憶力とは別もので、こうした専門医による診断の結果は、ほとんど
の場合、正常な年相応の健忘であるようです。高齢になると体の機能の全般が衰えるのは自
然の成り行きで、脳の機能も年齢とともに衰えて、健忘が始まるのです。しかし、認知症の
初期症状も「もの忘れ」で、健忘における良性の「もの忘れ」と混同されることが多いと言
われています 1)。
(認知機能とは)
記憶力は通常よく使われる言葉ですが、認知力という言葉は、あまり馴染みがありません
し、認知機能と言われても、なかなか理解できません。そこで、認知機能を、あえて言えば、
「自分が置かれている状況を無意識のうちに自動的に判断している能力」と言い換えられ
ることができます。
記憶には、意識しなくても自動的に覚える一時的な短期記憶と長い間覚えている長期記
憶があります。短期記憶の例として、よく電話が挙げられます。電話をかける時、少なくと
も 3、4桁の数字を覚えてプッシュするでしょう。何桁かの数字を一時的に覚えられない場
合は電話をかけるのも大変となります。計算にも短期記憶が重要で、一時的に数字を覚えて
計算がスムーズに出来るようになります。このため、短期記憶に問題があると、買い物の支
払いやお釣りの受取りも、うまく出来なくなってしまいます。覚えていても適切に思い出せ
ない場合もあります。思い出したことが適切でないと変なことになります。例えば「グッチ」
というブランドを思い出す時、
「グット」は何となく違うと思うでしょう。
「グッチ」を閃(ヒ
ラメ)いて思い出した時は間違いないと確信するでしょう。
日常生活では、意識しなくても経験したことは自動的に覚えています。この自動的に覚え
た事は適切に引き出され、自分の置かれている状況を常に判断しています。この自動的に覚
える機能に問題があると過去にあったエピソードがなかったかのように振る舞うことにも
なります。覚えていないことは、当然、思い出すことができません。認知機能も同じで、無
意識のうちに正しいか否かを確認していますが、認知機能に異常があると、そのまま言葉に
出したり行動したりします。そのため判断や行動が正常でなくなり、徘徊したり道に迷った
りが起こるのです。
4
ANM176®が認知症の予防と改善に役立つ可能性について
認知症とは、認知機能に問題があることによって、日常生活や社会生活が営なめなくなっ
た状態です。認知症になると食事をしたこと自体を忘れて「嫁がご飯を食べさせてくれない」
と言ったり、
「お金」を自分でしまい込んだことを忘れて「盗られた」と騒いだりするため、
家族や近隣とのトラブルに発展することにもなります。
このような認知症に、どのように対処したらよいのでしょうか。高齢になると早期の診断
が薦められています。
「もの忘れ外来」を訪問した結果、認知症でないと分かれば一安心で
しょう。しかし、認知症ではないと診断されても健忘が激しい場合、軽度認知障害(以下で
は mild cognitive impairmenrt から MCI と表記します)と診断されることがよくありま
す。MCI の半数は認知症の前段階と言われています。しかし、専門医が MCI と診断して
も、認知症の前段階なのかどうかを診分けることは困難と言われています 2,3)。
また、認知症と診断されても、現在の医療では治すことができないという現実もあります。
現在の医薬品では認知症の進行を一時的に抑制できる程度で、進行を止めることはできま
せん。認知症と診断されても、今のところは、医薬品で一時的に進行を抑制するか、患者の
行動の異常が激しい場合は向精神薬を投与するか、患者の行動や生活の改善あるいは家族
の心構えなど、認知症の周囲のことに目を向けるしか方法はありません。
(認知症について)
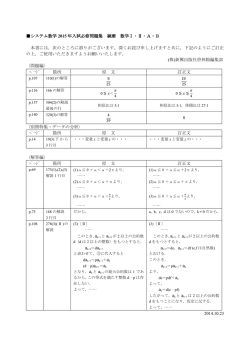
厚生労働省が 2008 年に発表した実態調査では、2002 年の認知症患者数は 228 万人で、
2020 年には 325 万人まで増加すると推察さ
れていました 4)。しかし、そのわずか 5 年後
FTD
PC
の 2013 年の発表では、2012 年の認知症患者
DLB
数は既に 462 万人であったと推計されてい
ます 5)。462 万人という認知症患者数は、65
VD
歳以上の 15%以上に相当します(2012 年の
65 歳以上の推定人口は 3079 万人)。この認
AD
知症患者 462 万人の中でアルツハイマー病
(AD)は 312 万人との推計です。さらに、
最近の推計では、2025 年の認知症患者は 700
万人になるとされています。このように、急
図 1.65 歳以上の日本の認知症患者の割合
2012 年の認知症患者は 462 万人で、その中でアル
ツハイマー病(AD)は 312 万人(67.6%)、血管
性認知症(VD)90 万人(19.5%)、レビー小体型
認知症(DLB)20 万人(4.3%)、パーキンソン病
(PC)20 万人(4.3%)、前頭側頭型認知症
(FTD)4 万人(0.9%)であったと推計されてい
ます 5)。
速に認知症患者が増えているのです。
認知症の内訳では、AD は認知症全体の約
7 割を占め、2 番目に多い脳血管性認知症(以
下では vascular dementia から VD と表記し
ます)は約 2 割です。この 2 つで認知症全体
の約 9 割を占めます(図 1)。
AD は高齢になると誰でも発病する可能性があり、AD になると記憶力や認知機能が段々
5
ANM176®が認知症の予防と改善に役立つ可能性について
と低下し、並行して脳の側頭葉や
脳梁(のうりょう)
大脳皮質
頭頂葉(図 2)が萎縮し、この萎縮
に伴う神経障害が進行していきま
頭頂葉
す。
脳下垂体
(のうかすいたい)
VD は、脳梗塞を繰り返すことが
前頭葉
原因となって起こる認知症です。
日本では、20 世紀末までは VD が
後頭葉
認知症の中で最も多かったのです
側頭葉
が、その後は、食事の低塩化や運動
の推奨など生活習慣病の予防運動
視床下部
によって脳梗塞など脳の血管障害
小脳
(ししょうかぶ)
が減少し、それに伴って VD 患者
海馬(かいば)
脳幹(のうかん)
数も減少しています 6)。
AD と VD 以外の認知症に、レ
ビー小体型認知症
7)
(以下では
dementia with lewy bodies から
DLB と表記します、DLB)
、一部
図 2.脳の側面
Alheimer’s Disease Education & Referral Center(USA)
のパーキンソン病(以下では
Parkinson’s disease から PD と表記します)8)、前頭側頭型認知症(以下では frontotemporal
dementia から FTD と表記します)などがあります。
DLB は認知機能の低下だけでなく、しばしば、ないモノが見える幻視などの異常が起こ
り、それに伴う被害妄想が現れ、鬱(ウツ)にもなります。DLB も PD で見られるようなパ
ーキンソニズム*と呼ばれる症状が、しばしば見られます。また、これらの症状は、日によ
って良くなったり悪くなったりの日内変動があります。
*パーキンソニズムは、小脳の障害に起因する症状で、手足や首、顎、唇、舌に揺れが見られ、体が傾
き、歩行は前かがみで腕の振りが少なく小刻みとなる歩行障害などパーキンソン病に特徴的にみられ
る運動性障害です。
PD は高齢者に発症することが多く、パーキンソニズムが特徴です。また、軽度の認知症
や鬱を伴うことも稀ではありません 8)。
FTD は前頭葉と側頭葉が萎縮する認知症の総称で、性格変化や人格変化に伴う行動異常
が先行し、後に、覚えることが難しくなったり、地理や位置についての異常(空間認知障害)
が現れたりするのが特徴です。ピック病は FTD に分類された代表的な認知症です。
(中枢神経系について)
神経を持っている動物の中には神経が分散している散在型と神経の中心が体の中央部に
ある脊索(セキサク)型があります。脊索型の脊索動物は、受精卵が分裂して体を形成する発生
の過程で表面にある細胞(外胚葉細胞)が溝状に陥没し、溝の両側が閉じて筒のような構造
6
ANM176®が認知症の予防と改善に役立つ可能性について
(神経管)が出来ます。同時に、神経管の先端が膨らんで脳の原型が出来上がります。脊索
動物から進化したヒトのような脊椎(セキツイ)動物では、神経管の周辺に背骨(脊椎骨)が形
成されて、その中は神経が入った脊髄(セキズイ)となります。
脳と脊髄を総称して中枢(チュウスウ)神経系と呼びます。一方、皮膚や目や舌などが感知
した刺激や感覚を中枢神経系に伝えたり、中枢神経系の指令を筋肉に伝えたりする体の神
経を末梢(マッショウ)神経系と呼びます。通常、末梢神経系は脊髄で中枢神経系と繋がって
います。末梢神経系には脊髄と繋がっていない自律神経もあり、自律神経は心臓や腸を動
かしたり、汗腺を開いたりなど無意識で働きます。中枢神経系も末梢神経系も指令伝達の
方式は基本的に同じですが、中枢神経系は記憶や認知機能を担い、末梢神経系は指令や刺
激の伝達に特化していることから、大きく異なるところもあります。例えば、中枢神経系
は、記憶を担っているため基本的には再生できませんが、末梢神経系は再生できます。
神経線維は多くの線維が寄り集まってできていますが、この中の 1 本の線維は、神経伝
達を担う細胞から出ています。この神経伝達を行う細胞をニューロン(neuron)または神
経細胞と呼びます。ヒトは、約 1 兆個の神経系細胞をもって生まれてくると言われていま
すが 9)、この中の約1割がニューロンで、残りは色々な面でニューロンを補助しているグリ
ア細胞と呼ばれる神経系細胞です。まだ生まれる前の胎生期に、神経系細胞からニューロン
とグリア細胞が派生し、次第にニューロンは
神経伝達に、グリア細胞はニューロンの神経
伝達を補助する役割に分化*します。
*1 つの細胞として生まれた後、分裂を繰り返し
ながら親と同じ姿になる過程を個体発生と言
います。個体発生の過程で分裂した細胞は、そ
の後、体の中での役割を運命づけられ、様々な
組織や器官の細胞となります。この様に細胞の
役割が分かれる過程を分化と言います。
神経伝達を補助しているグリア細胞には
ニュ ーロ ンの 周 囲に あるア スト ロサイト
(astrocyte、星状細胞とも呼ばれます)と神
経線維の周りを取り巻いているオリゴデン
ドロサイト(oligodendrocyte)があります。
ニューロンの周囲にあるアストロサイトは
ニューロンに栄養や必要物質を供給したり
ニューロンで要らなくなった物質の浄化に
働いたりなど様々なニューロンの補助活動
をしています。アストロサイトは哺乳動物か
ら出現し、それ以前の動物には見られませ
ん。また、アストロサイトはヒトへの進化過
図 3.ニューロンとシナプスの模式図
程で、その数が急増していることから、言語
7
ANM176®が認知症の予防と改善に役立つ可能性について
や思考などの高次脳機能と強い関係があると考えられています。
ニューロンには軸索と呼ばれる 1 本の長い突起と樹状突起と呼ばれる多数の短い突起が
あり、軸索は他のニューロンの樹状突起と結合します(図 3 上)
。ニューロンの神経伝達に
おいて軸索の表面を流れる電気の電位が重要な役割をしています。この軸索をオリゴデン
ドロサイトがミエリン鞘(ショウ)と呼ばれる電線の被服のようになって包み(図 3 上)
、軸索
表面の電位を増幅したり安定化させたりする役割を果たしています。
体にはマクロファージと呼ばれる細胞外の異物をアメーバ―のように内部に取り込んで
分解処理をする細胞がありますが、脳には存在しません。その代りにマクロファージと同じ
ような働きをするミクログリア(microglia)と呼ばれる細胞が脳に存在します。
軸索と他のニューロンの樹状突起はシナプス(synapse)と呼ばれるキノコのような構
造物によって結合しています(図 3 下)
。軸索側のシナプス先端の膜(シナプス前膜と呼
ばれます)と樹状突起側のシナプス先端の膜(シナプス後膜と呼ばれます)の間にはシナ
プス間隙と呼ばれるわずかな隙間(スキマ)があります(図 3 下)
。上昇した電位が軸索表面
を伝わってシナプス前膜
に達するとシナプス前膜
からグルタミン酸
(Glutamate)など神経
伝達物質と呼ばれるタン
パク質がシナプス間隙に
放出されます。シナプス
後膜にはシナプス間隙に
放出された神経伝達物質
を受け止める受容体があ
図 4.細胞膜受容体
ります。ニューロンに限
1.イオンチャンネル型受容体(図左)
:様々なイオンが結合すると受容体の中
にあるイオンを通すイオンチャンネルが開き、イオンが流入・流出します。
2.酵素連結型の 1 つに G タンパク質受容体があります。様々な刺激によって
G タンパク質に結合した GDP が GPT にリン酸化されます。上図では、活
性化した G タンパク質がイノシトール 2 リン酸(PIP2)をリン酸化するホ
スファチジルイノシトール 3-キナーゼ(PI3K)が活性化してイノシトール
3 リン酸(PIP3)に変換します。活性化した G タンパク質が PIP2 から PIP3
を作り出す酵素には PI3K の他にプロテインキナーゼ C(PKC)がありま
す。
らず様々な細胞の細胞膜
には、特定の物質を通過
させたり、情報の伝達に
関与する特定の物質を特
異的に感知したりする機
能がある受容体が埋め込まれています(図 4)。
神経伝達物質はシナプス後膜にある特定の受容体を刺激して活性化し、次のニューロン
の表面電位が上昇します。その結果、刺激がニューロン間を次々と伝達します。神経伝達物
質はニューロンから次のニューロンへ、いわばスイッチの役割を果たしています。
シナプスによるニューロン同士の結合は、脳内で、網の様に広がっています。この網目構
造をニューロンネットワークと言います 10,11)。
8
ANM176®が認知症の予防と改善に役立つ可能性について
(脳関門)
血管が詰まったり破れたりして起こる脳梗塞で分かるように、血管は脳の奥まで到達し
ています。しかし、血液とニューロンとは接していません。血管とニューロンの間にはア
ストロサイトがあって、アストロサイトはニューロンへの栄養や神経伝達物質の供給・調
整を行い、ニューロンの働きを妨げるような血液中の物質を遮断しています。このアスト
ロサイトの働きのように、血流に乗ってやってくる様々な物質を取り除くシステムを血液
脳関門(以下では Blood Brain
barrier から BBB と表記します)
と呼びます。
細胞は間質液という透明な液に
浸っています。血液により運ばれ
た酸素やタンパク質などは毛細血
管壁から間質液へと拡散し、間質
液から組織の細胞へと拡散しま
す。この間質液は、リンパ液とな
ってリンパ管内を流れて静脈に合
流します。脳にはリンパ管はな
く、その代わりに脳髄液(以下で
は cerebrospinal fluid から CSF
と表記します)があります。CSF
は脳と脊髄の中に限って流れてい
ます。脳の神経系細胞が浸ってい
る脳間質液は CSF 中に拡散しま
す(図 5)。
BBB では、アストロサイトや
ペリサイトと呼ばれる細胞が毛細
血管とニューロンの間にあって、
血管から一部の物を自分自身の膜
図 5.CSF の生産場所と循環
側脳室、第三脳室、第四脳室には血管が豊富な脈絡叢という組
織がある。脳脊髄液(CSF)は主に脈絡叢で産生される(①)。
また、CSF は脳室壁の上衣細胞からも産生されるし、脳内の間
質液も脳室内に入る。脳室内で産生された CSF は、側脳室⇒第
三脳室⇒中脳水道⇒第四脳室へと脳室内を循環(②)する。次
いで第四脳室正中孔および外側孔からくも膜下腔に流れ出る
(③)。そしてテント下からテント上へと膜下腔を流れ(④)、
上矢状洞近傍のくも膜顆粒から吸収され、静脈系に入る(⑤)。
脳室とくも膜下腔にある CSF の量は成人では約 140mL であ
り、1 日の生産量は約 500mL である。
船橋市立医療センター脳神経外科(2002 年 12 月 12 日作成)より
を通過させ、その一部をニューロ
ンに引き渡しています 12)(図 6)。水
など非常に小さい分子や脂溶性あるい
脳毛細血管
血管上皮細胞
ペリサイト
ニューロン
アストロサイト
はプラスにチャージしている分子は、
ある程度、例えば大きさでいえば、小
さいほどニューロンに入ることができ
ます 12)。しかし、血中にある通常の
物質は BBB を透過してニューロンに
図 6.アストロサイトの BBB 機能
9
ANM176®が認知症の予防と改善に役立つ可能性について
入ることはできません 13,14)。
ニューロンにとって重要な物質は、受容体(図 4)やトランスポーター*がアストロサイ
トの細胞膜を透過させます。BBB に関与するトランスポーターにはブドウ糖(以下ではグ
ルコース**と呼びます)など糖類、アミノ酸類、乳酸などモノカルボン酸類等、特定範囲
の物質の透過に関わるものが知られています 15)。
*トランスポーターはチャンネルや受容体と共に細胞膜に存在する膜タンパク質で、受容体とは異
なってエネルギーを使って特定物質の膜透過(輸送)を行います。
**糖には多くの種類がありますが、その中でグルコースは生物に幅広く利用されます。植物の光合
成によって、水(H2O)の水素原子(H)と空気中の炭酸ガス(CO2)からグルコース(C6H12O6)
が作られ、グルコースから様々な糖やタンパク質や脂肪などの様々な生体物質が作られます。また
グルコースはエネルギーの根源でもあります。
グルコースなどの糖類は GLUT1 と GLUT3、アミノ酸類は LAT1 と CAT1 など、フェル
ラ酸や乳酸などのモノカルボン酸類はモノカルボン酸トランスポーター(以下では Mono
Carbonate Transporter から MCT と表記します)が、BBB による透過や小腸からの吸収
にトランスポーターとして働いています。しかし、BBB による透過は小腸からの吸収より
かなり厳密で、特定の分子しか透過させません。
タンパク質の場合は、どんなに小さくても、BBB に機能するタンパク質の助力をなくし
ては脳内から血中へ流出できません
14,16)。また
BBB には、インスリン受容体(以下では
insulin receptor から IR と表記します)***17)、インスリン様成長因子(以下では insulinlike growth factor から IGF と表記します)受容体 18)、レプチン****受容体 19)、血中に含
まれる鉄イオンの輸送を担っているトランスフェリン受容体 20)なども関与しています。
***インスリン受容体は糖尿病と関係があるホルモンのインスリンが結合して細胞内に作用をします。
****レプチンは肥満の抑制や体重増加の制御と関係するホルモンです。
もう1つ触れておきたい BBB に機能する受容体に、悪玉コレステロールと言われている
低比重リポタンパク質(以下では low-density lipoprotein から LDL と表記します)の受容
体があります。LDL が悪玉と言われるのは、動脈硬化を起こして血管が詰まる原因となる
からですが、本当は重要な役割をしています。一方、善玉コレステロールとは高比重リポタ
ンパク質(以下では high-density lipoprotein から HDL と表記します)のことですが、LDL
と HDL ともコレステロールそのものではなく、タンパク質と脂質が結合したリポタンパク
質にコレステロールが包み込まれた粒子です。
体の中で重要な働きをするコレステロールは、体内の様々な細胞内で合成され、リポタン
パク質で包まれた LDL や HDL 粒子にパックされて血液などの循環液中を運ばれます。コ
レステロールは真核生物*****の細胞膜成分ですが、他にも様々な機能があります。体内に
あるコレステロールの 4 分の 1 は脳に存在し、脳ではニューロンの軸索突起を覆うミエリ
ン鞘の膜にあって絶縁体のように機能しています。
*****生物には原核生物と真核生物に大きく分けられます。大腸菌などの原核生物は、遺伝子が
書き込まれた DNA が細胞質に広がって存在しますが、酵母やヒトなどの真核生物の DNA は細胞
質の中にある細胞核にコンパクトに圧縮されて存在します。
またコレステロールは、アンドロゲン(男性ホルモン)
、女性ホルモン(エストロゲン)
、黄
10
ANM176®が認知症の予防と改善に役立つ可能性について
体ホルモン(プロゲステロン)などステロイドホルモン,ビタミン D,胆汁酸などの合成原
料ともなっています。
ヒトを始めとした哺乳類においてコレステロールの大部分は食事に由来するのではなく、
体内で合成されます。コレステロールの生合成は,ホルモンやコレステロールの血中濃度で
調節され、細胞内コレステロール量は厳密に制御されています
21)。すべての細胞はコレス
テロールを合成できますが、一方では、細胞表面の LDL 受容体を通して血液中の LDL を
取り込み、コレステロールを補給することもできます。細胞内のコレステロール量が十分に
満たされない場合は、コレステロールの合成や LDL 受容体の量を増やします。
LDL 受容体の機能を持つタンパク質は数多くあり(これらをまとめて LDL 受容体メン
バーといいます)、ほとんどすべての細胞に存在し、特に肝臓に多くあります。LDL 受容
体メンバーは共通して、細胞膜を1回貫通したタンパク質で、場合によっては同じ物質を違
ったメンバーが感受し
22,23)、それらの物質を細胞内に取り込んだり、細胞内に情報伝達し
たり(以下では「シグナリング」と言います)など、その機能も多様です 24,25)。LDL 受容
体メンバーの LRP1 (LDL-related protein receptortype 1)は、脳内のリポタンパク質な
ど巨大分子の輸送や
26)、ウイルスやアルツハイマー病の原因タンパク質と考えられている
βアミロイドなど様々な毒性物やタンパク質の脳からの排泄に働いています 27–29)。
体の中で行われている生命維持活動について
生物には様々な機構が備わり、この機構によって生命が維持されています。生命が誕生し
たのは海の中だったと言われています。そのため、最初に誕生した原始の海のような環境が
保たれた細胞の内部(細胞質と呼ばれる)で行われている生命の維持活動は巧妙に制御され
ていますが、何らかの要因でこの制御された維持活動が正常に機能しなくなると、様々な障
害や病気をもたらします。
(生物は特有な成分で構成された細胞から成り立っている)
バクテリアなど細胞が 1 つから成る単細胞生物が進化して多細胞生物が誕生したのです
が、多細胞生物も子孫を残す過程で一旦は単細胞生物と同じように一つの細胞となり、次世
代では細胞分裂によって再び多細胞生物になります。
細胞は、タンパク質、核酸、脂質、糖類といった生物に特有な成分で、できています。
これらの成分の間には、生物に特有な方式の相互関係があります。例えば、親から子へ引
き継がれる遺伝子の情報は DNA と呼ばれる核酸の配列に組み込まれており、この情報に
基づいてタンパク質を構成するアミノ酸の配列が決まります。このようにして出来上がっ
たタンパク質には、細胞の骨格形成、酵素反応、シグナル伝達など多くの役割がありま
す。細胞の骨格を担うタンパク質は単に物理的な構造を決めているだけでなく、細胞の運
動や細胞内での物質輸送にも関与しています。
酵素の役割を果たすタンパク質は、様々な生体物質を作ることや物質を分解することに
11
ANM176®が認知症の予防と改善に役立つ可能性について
関与しています。酵素の役割を果たすタンパク質は、生物内の低温条件においても、物質
の合成や分解(反応)を行うことができます。日常経験している自然界の化学反応の典型
は火で、例えば家庭で使うガスは 200℃以上になると空気中の酸素による酸化反応が起こ
り、さらに温度が上昇して反応が周辺に広がり、炭素成分が赤く光って炎に見えます。ま
た、自然界で行われる穏やかな反応の例として、空気中の酸素が鉄を酸化して錆となる反
応が挙げられます。この場合は、紫外線などの放射線が鉄の原子に作用して酸素と反応さ
せるため、ゆっくりとした変化となります。こうした自然界の化学反応と比べると生体内
で行われる酵素による反応は厳密で一定の反応しか行わず、また、反応させる相手を酵素
の活性部位の構造にはめ込み、1 分子から正確に行うことができる特殊な反応です。
シグナリングの役割を担うタンパク
質の役割も基本的には酵素反応と似て
A
おり、タンパク質の特定な構造に別の
タンパク質がはめ込まれることによっ
てタンパク質の構造が変化し、その結
B
果、タンパク質の機能が変化します。
この変化はスイッチのような役割を果
たし、生命活動で欠くことができない
様々な情報伝達を調整したり制御した
りします。
ヒトの体は約 40 兆個以上の細胞があ
ると言われており、皮膚、内臓、脳、骨
など組織も細胞で成り立っています。微
図 7.脂質二重層のリン脂質分子の代表的な構造
A.リン脂質のうち細胞膜で多数を占めるグリセロリン脂
質の基本構造。(R)の部分は分子によって異なり、コリ
ンが付くとホスファチジルコリンになります。
B.細胞ではリン脂質分子は自発的に集合し二重層を作りま
す。リン脂質の中で、量が最も多いホスファチジルコリ
ンは主に細胞膜の外側にあり、ホスファチジルセリン、
ホスファチジルエタノールアミンは内側にあります。
WIKIPEDIA より
生物など細胞が1つだけの単細胞生物
に限らず、多くの細胞で成り立っているヒト
も、膜(細胞膜)によって隔てられた細胞に
よって成り立っています。
外界と細胞内を隔てる細胞膜は、主に水溶
性の分子が油溶性の脂質の端に結合したリ
ン脂質(図 7A)で成り立っています。細胞膜
は石鹸の泡やシャボン玉に例えられます。石
鹸の成分は脂肪酸(脂肪酸も脂質の類)(図
7A)で、泡やシャボン玉では水溶性部分が水
側(シャボン玉は中側)に並んで膜を作りま
す。同じように細胞膜でも様々なリン脂質が
膜の形成に寄与しています。シャボン玉は脂
肪酸が一列の一重ですが、細胞膜の場合はリ
PIP2
PIP3
図 8.ホスファチジルイノシトール(PI)
図中の PO はリン酸を示す。
伊集院 壮,竹縄 忠臣 脳科学辞典
12
ANM176®が認知症の予防と改善に役立つ可能性について
ン脂質の油溶性部分同士が内側、水溶性部分が外側になった 2 重膜構造(脂質二重層)です
(図 7B)。
細胞膜の受容体から細胞内に行う細胞内シグナリングにホスファチジルイノシトール 3
リン酸(以下では Phosphatidyl-inositol 3,4,5-trisphosphate から PIP3 と表記します)が
重要な役割を果たしています。PIP3 が切り出されるイノシトールリン脂質のホスファチジ
ルイノシトール(phosphatidyl-inositol;PI)
(図 8)も細胞膜の内側にあります。さらに、
多くの場合、細胞膜には膜で様々な機能を果たしている膜タンパク質が脂質の中に埋め込
まれるか脂質に結合しており、この脂質や膜タンパク質には、糖鎖やリン酸が結合し、コレ
ステロールも入り込んでいます。したがって細胞表層は全体として複雑な構造となり、細胞
の種類ごとに特徴が異なっています。細胞膜にあるコレステロールは、膜を固くする役割が
ありますが、細胞内で重要なコレステロールの保存場所でもあります。
細胞膜の脂質二重層は電気的に中性で極めて小さな分子、例えば酸素分子や二酸化炭素
分子は通しますが、イオンあるいはアミノ酸、糖類、核酸と言った生体物質の大部分は通る
ことができません。細胞膜に埋め込まれている受容体(図 4)が、特定の物質を通過させた
り、情報の伝達に関与する特定物質を特異的に感知したりする機能を果たしています。細胞
の外には、マトリックスと呼ばれる様々な成分が含まれる不定形の構造があり、マトリック
スと細胞は様々な相互作用をしています。
このように細胞膜は、外界からの刺激や毒物あるいは体内からのシグナルや状況を感知
する場所として、また、物質の細胞内外の交換、細胞間のコミュニケーション、細胞同士の
接合など多くの機能に関与しています。
(細胞内へのシグナリング)
多くの細胞から構成される生物体の中で一定の役割を分担する細胞は、周りや全体の細
胞に統率されて機能します。この統率に働くシグナリングに関与するタンパク質にサイト
カイン*とホルモンがあります。近隣の細胞へのシグナリングにはサイトカインが担ってい
ます。ホルモン類は血液やリンパ液など循環液に乗って移動し、組織間でシグナリングしま
す。
*サイトカインは数百種類が発見されています。炎症と関係する炎症性サイトカインには、白血球が
分泌し免疫系の調節に機能するインターロイキン(以下では Interleukin から IL と表記します)、
白血球の遊走を誘導するケモカイン、ウイルス増殖阻止や細胞増殖抑制の機能があり免疫系で重要
な役割があるインターフェロン、さまざまな細胞に増殖や分化を誘導したりアポトーシスを誘発し
たりする腫瘍壊死因子(以下では tumor necrosis factor から TNF と表記します)などがありま
す。IL は現在 30 種以上が知られています。TNF ファミリーには構造的にも機能的にも類似してい
る TNFαと TNFβがあります。
特定の物質を感知する受容体には、細胞膜に埋め込まれた膜結合型と細胞内にある細胞
質型があります。
膜結合型受容体は、その機能によってイオンチャンネル型と酵素連結型に分けられます
(図 4)。イオンチャンネル型は刺激物が結合することによって受容体の構造が変化し、ナ
13
ANM176®が認知症の予防と改善に役立つ可能性について
トリウム、カルシウム、カリウムに代表されるイオンを通過させるか否かの開閉を行いま
す。酵素結合型にはサイトカインあるいはホルモンなど特定の刺激物を認識する単純伝達
型、ホスファチジルグアニンと結合している G タンパク質結合型、微生物由来の糖、脂
質、タンパク質あるいは核酸などによる分子構造(病原体関連分子パターン)を検出する
パターン認識受容体(pattern recognition receptor;PRR)があり、これらの酵素結合型
受容体は、イオンを通過させるものと刺激物を認識して膜の機能や細胞内のシグナル伝達
に関わる酵素を活性化するものがあります。
単純伝達型受容体は、タンパク質中のチロシンと呼ばれるアミノ酸をリン酸化する活性
(リン酸化活性を持っている酵素は「キナーゼ」と呼ばれます)を持っているためチロシン
キナーゼ型受容体(以下では receptor tyrosine kinase から RTK と表記します)とも呼ば
れます。RTK の中にあるアミノ酸のチロシンを特異的にリン酸化するチロシンキナーゼ活
性は、特定のタンパク質中のチロシンをリン酸化して活性化し、様々なシグナリングを発し
ます。RTK は、主にサイトカインやホルモンの受容体で、細胞の分裂、増殖、形態形成、
分化に重要な役割を果たしています。糖尿病と関係があるインスリン*や成長因子**の受容
体も RTK 型受容体です。
*インスリンは糖尿病と関係が深いホルモンです(詳細は 68 ページ「糖尿病とは」の項を参照してくだ
さい)。
**成長因子とは、特定の細胞の増殖や変化の調節や促進に関与する内因性タンパク質の総称で、標的
細胞の表面の受容体タンパク質に特異的に結合することにより、細胞間の信号物質として働きます。
代表的な成長因子には上皮成長因子(Epidermal growth factor;EGF)、IGF、トランスフォーミン
グ成長因子(transforming growth factor;TGF)、神経成長因子(nerve growth factor;NGF)、脳
由来神経栄養因子(brain-derived neurotrophic factor;BDNF)などがあります。サイトカイン、ホ
ルモン、ビタミンは成長因子には含まれません。
G タンパク質結合型の G とはグ
アニンのことで、グアニンは DNA
を構成する 4 つの核酸の 1 つです
(G タンパク質中のグアニンは
DNA の遺伝情報とは関係ありませ
ん)
。DNA を構成する 4 種の核酸
の中のグアニンとアデニンは(他
GTP
GDP
グアニン
にはシトシンとチミンがありま
す)
、容易に電子を受けたり離した
りする性質があり、DNA 以外にも
図 9.GTP と GDP 間のリン酸化・脱リン酸化
GDP がリン酸化され GTP となります。一方、GTP はホスファ
ターゼによって脱リン酸化され GDP に戻ります。
細胞内で様々な役割を果たしてい
ます。G タンパク質にはグアニンにリン酸 2 個が付いた GDP かリン酸が 3 つ付いた GTP
のどちらかが結合しています(図 9)。 平常は、G タンパク質には GDP が結合してお
り、不活性型です。GDP がリン酸化され GTP になると G タンパク質が活性化します(図
10)。 G タンパク質自身には GTP を脱リン酸化する作用があり、活性型の GTP 結合型は
刺激がなくなると自動的に GDP 結合型に戻って不活性となります。さらに GTP アーゼ活
14
ANM176®が認知症の予防と改善に役立つ可能性について
性化タンパク質(GTPase
activating proteins ;GAP)によ
って GTP から GDP への脱リン酸
化が加速されます 30)。このように
刺激によって ON になり刺激が無
くなると自動的に OFF になる G タ
ンパク質はスイッチのような働きを
します 31)。また、細胞内にシグナ
ルを発する時に重要な制御をするグ
アニンヌクレオチド交換因子(以下
図 10.G タンパク質の活性化
では guanine nucleotide exchange
・GDP が結合した不活性型 G タンパク質はグアニンヌクレオチ
ド交換因子(GEF)により GDP 型から GTP 型へ変換され活性型
となります。
・G タンパク質は GTP 末端リン酸基を加水分解して脱リン酸化
し、GDP にする GTP アーゼ活性をもつので、GTP が結合し
た活性型 G タンパク質は自然に GDP 型(不活化型)にもどり
ます。
・G タンパク質自身のもつ GTP アーゼ活性は一般的に低いが、
GTP アーゼ活性化タンパク質(GAP)と結合することにより酵
素活性が亢進し不活性型に戻り易くなります。
(WIKIPEDIA より改変)
factor から GEF と表記します)は
GDP を GTP に変換します 32,33)。
RTK 型受容体のあるものは GEF
を活性化して G タンパク質に結合
した GDP を GTP に変換し、G タ
ンパク質を活性型にします。GEF
の機能を持つ SOS と言うタンパク質は、チロシンキナーゼによるアロステリック制御に
よって GEF の機能が活性化されます 34)。アロステリック制御とは、酵素反応ではなく、
相手のタンパク質に結合することによって、その構造を変化させ、その結果、相手のタン
パク質の機能を変化させてシグナリングの調整や分岐に機能する方法です。
G タンパク質結合型受容体には、3 量体(1 つのタンパク質を 1 量体といいます)となっ
て活性化する 3 量体 G タンパク質に連携して働く G タンパク質共役受容体(以下では G
protein-coupled receptor から GPCR と表記します)があります。GPCR には神経に作用
する物質の受容体など多数の種類があり、様々な刺激を感知する役割を果たしています。
微生物の外殻にある病原体関連分子パターンを認識するパターン認識受容体(PRR)に
はスカベンジャー受容体やインテグリン*受容体のように他の受容体の協力がないと細胞
内シグナリングをしないタイプと、微生物を感知する Toll 用受容体(以下では Toll like
receptor から TLR と表記します)などのように単独でシグナリングをするタイプがあり
ます。LDL 受容体(11 ページ)はスカベンジャー受容体の仲間です。
*インテグリンは細胞膜にあって細胞同士や細胞の外にある物質と細胞の接着に機能し、体の形態形成、
細胞の移動、傷の修復、炎症反応、生体防御、止血、血管新生など非常に広い範囲の生体現象に重要
な役割を果たしています 35)。インテグリンには 24 種の仲間があり、共通してαβサブユニットの 2
量体を形成して受容体として機能します。
(キナーゼによるシグナリング)
特定のアミノ酸にリン酸を付けてシグナリングに関与するキナーゼはリン酸化を受ける
と自身のタンパク質の構造が変化して、自身の中にあるリン酸化酵素活性が活性化し、活性
15
ANM176®が認知症の予防と改善に役立つ可能性について
化したキナーゼは別のキナーゼを活性
化してシグナルが次から次へと伝わり
ます。このシグナリングに関わるキナ
ーゼには、相手のタンパク質中のリン
酸化するアミノ酸の種類によって、チ
ロシンをリン酸化するチロシンキナー
ゼとセリンあるいはスレオニンをリン
酸化するセリン・スレオニンキナーゼ
の 2 通りがあります。
チロシンキナーゼは主に受容体の活
性化と細胞膜の機能に関与し、チロシ
ンがリン酸化されることによってアダ
プタータンパク質とよばれるタンパク
図 11.MAPK 経路による DNA の転写活性化
PP2A、PTPase、MKP-3 は脱リン酸化酵素で、MPAK シ
グナリングを抑制します。
質が結合し、次のキナーゼが活性化さ
れます。チロシンキナーゼ型受容体(RTK)
(14 ページ)もチロシンキナーゼの 1 つです。チ
ロシンキナーゼからセリン・スレオニンキナーゼへのシグナリングは、主にアロステリック
制御(15 ページ)によって行われます。また、その後のセリン・スレオニンキナーゼのシグナ
リングもアロステリック制御によって、途中で分岐したり、強められたり弱められたり、複
雑に制御されて最終目標の標的タンパク質の活性化や合成が起こり、様々な事態に適切に
対応します。
セリン・スレオニンキナーゼの細胞内シグナリングに、よく知られた分裂促進因子活性化
タンパク質キナーゼ(以下では Mitogen-activated Protein Kinase から MAPK と表記しま
す)があります。代表的な MAPK はサイトカインなどが活性化した RTK 型受容体によっ
て G タンパク質の Ras が活性化し、Ras によって活性化した MAPKK キナーゼ(MAPKKK)
は MAPK キナーゼ(MAPKK)を、MAPKK は MAPK をリン酸化して活性化して NF-κB
などの転写因子を活性化し、細胞の対応に重要なタンパク質の転写を行います 36)(図 11)。
G タンパク質を活性化する GEF の機能を持つ SOS(15 ページ)は MAPK 経路を活性化しま
37)。MAPK
す
は微生物を感知する TLR によっても活性化されます。p38(p38 mitogen-
activated protein kinase)は代表的な MAPK で、p38 の仲間には ERK1/2、JNK(C-JunN-terminal kinase)、ERK5 などがあり、熱、紫外線、重金属や微生物など様々なストレス
*
によって活性化され、場合によってはアポトーシス(apoptosis)**と呼ばれる細胞の自己
死を引き起こすなど、細胞の種類により、様々なストレス応答*に関与しています 38,39)。
*一般にストレスというと精神的な負担や圧迫といった意味ですが、生物的なストレスというと、外部
環境や傷害によって起こる負担や対応(応答とも言います)のことです。
**アポトーシスとは遺伝子に組み込まれ制御された細胞の自己死のことです。細胞は、ストレス応答
や発生・分化などの必要に応じて、細胞死による影響を他の細胞に及ぼさないように制御されたアポ
トーシスを引き起こして消滅する仕組みが備わっています。ヒトのように多数の細胞から成り立って
いる生物(多細胞生物)の場合は、1 つの細胞が死んでも個体は死にませんし、また、アポトーシスに
よって古い細胞を新しい細胞に置き換えて個体の生命が維持されます。
16
ANM176®が認知症の予防と改善に役立つ可能性について
アポトーシスに対してネクローシス(necrosis)は、遺伝子による対応に間に合わないような緊急
な細胞死のことです。最近では、一部のネクローシスは遺伝子が関係すると報告されています。
p38 と JNK は酸化ストレスやヒートショックなど環境ストレスによって活性化され、細
胞質から核内に移動して、さまざまなストレスに応答する
40)ことからストレスキナーゼ
(Stress-activated protein kinase;SAPK)とも呼ばれています 38)。
G タンパク質はシグナリングの起点となる重要な役割を果たしています。その起点の 1
つにホスファチジルイノシトール 3-キナーゼ(以下では Phosphatidyl-inositol 3-kinase
から PI3K と表記します)があります。G タンパク質によって活性化された PI3K は 41)、
細胞膜にあるイノシトールリン脂質に結合しているホスファチジルイノシトール 2 リン酸
(PIP2)から PIP3(図 8)(13 ページ)を作り出します 42,43)。
PIP3 はアロステリックな制御によって多くのタンパク質を活性化します。その中でも、
細胞の形態変化、増殖、分化などに関連するセリン・スレオニンキナーゼの Akt(PBK と
もいいます)はストレスキナーゼ経路も制御し、生命の維持に重要です。PIP3 が Akt を活
性化する経路には、PIP3 が Akt を直接活性化する直接的経路 44)と PIP3 に結合して活性
化したセリン・スレオニンキナーゼの PDK1 が Akt を活性化する間接的経路があります
45,46)。
(ユビキチン化とプロテアソーム)
細胞の中で行われているタンパク質の分解は新陳代謝に寄与するだけでなく、シグナリン
グに関係するタン
パク質を調整・制
御します。
ユビキチン化と
は、76 個のアミノ
酸から成るユビキ
チン(Ubiquitin)
と呼ばれる目印の
タンパク質を標的
のタンパク質に付
図 12.ユビキチン化とプロテアソーム分解
ユビキチン化には 3 つのカテゴリーのタンパク質、E1、E2、E3 が関係します。ユ
ビキチン活性化酵素 E1 は ATP のエネルギーを利用して、E1 自身のシステイン残
基にユビキチン(Ub)を結合させ、E1-Ub となります。ユビキチン結合酵素 E2 は
E1-Ub を自身のシステイン残基に結合させ E2-Ub を生成します。ユビキチンリガー
ゼ E3 は、標的タンパク質を識別し、標的タンパク質のリジン残基にユビキチンを結
合させます。Web.「ユビキチン化を介したタンパク質分解のしくみ」小林麻己人より
けることです。ユ
ビキチン化されたタンパク質の分解システムには、プロテアソームという細胞内の器官で分
解するユビキチン・プロテアソーム系(図 12)とリソソームという細胞内の器官で分解す
るオートファジー・リソソーム系があります。ユビキチン・プロテアソーム系では、ユビキ
チン化によって特定のシグナリングに関するタンパク質をプロテアソーム分解に送り込んで
制御することによって、細胞の重要なプロセスに関わり細胞機能を調節します 47,48)。
ユビキチンを標的タンパク質のアミノ酸リジンに結合させるユビキチンリガーゼ E3 は哺
17
ANM176®が認知症の予防と改善に役立つ可能性について
乳類で約 1,000 種が見出されており、また、E3 はユビキチン化する標的タンパク質のタイ
プによって、幾種類かに分類されています。
(遺伝子の DNA 情報からタンパク質の合成)
遺伝子は細胞核にある染色体(図 13A)の中にあります。染色体は両親から 1 本ずつ受
け継いだ同じ形の染色体(これを相同染色体と呼びます)の 2 本が一組になっています。
ヒトには 23 組の相同染色体があります。ただし、性染色体と呼ばれる 1 組は、女性は XX
の組合せで相同ですが、男性は XY で相同ではありません。性染色体以外は常(ジョウ)染
色体といわれ、ヒトの常染色体は 1 から 22 までの番号が付けられています。相同染色体
には両親から引き継いだ同じ遺伝子があり、この 2 つの遺伝子を対立遺伝子と言います。
対立遺伝子に書き込まれたタンパク質のほとんどは同じですが、場合によってはアミ
ノ酸の一部が異なっている場合があり、この中で形態や機能に寄与する遺伝子を優
勢、機能していない遺伝子を劣勢と言います。
染色体には、ヒストン(図 13A)と呼ばれるタンパク質に DNA が巻き付いています。
DNA は核酸が連なった構造体で(図 13B)
、遺伝子は DNA の中に情報として書き込まれ
ています。DNA の配列を構成している核酸は塩基と呼ばれ、塩基にはシトシン(C)
、チ
ミン(T)、アデニン(A)、グアニン(G1)の 4 種類があります(図 13B,C)。この 4
つの塩基の中の 3 つの組合せ(アミノ酸に対応する 3 個の塩基配列はコドンと呼ばれま
す)が 1 つのアミノ酸を指定します。「あ、い、う、え、お、・・」と同じように DNA の
コドンは文字の役割があり、DNA 中の一定の長さの塩基配列は特定のタンパク質を表わ
しています。タンパク質を構成するアミノ酸は 20 種類があり、例えば、コドン TTT はア
ミノ酸のフェニルアラニン、CTT はロイシン、CAA はグルタミンを指定します。ヒトの
場合、親から引き継いだ DNA には 3 万から 4 万種のタンパク質の情報が書き込まれて
います 49)。
ヒトには約 40 兆個の細胞があるといわれていますが、それぞれの細胞には必ず DNA が
あり、その中には体を作る全ての情報が書き込まれています。DNA 情報の一部しか使わ
ななくても、全ての情報を持つことは無駄にも思えますが、多数の細胞を狂いなく制御す
るには、この方が良いのでしょう。その代りに細胞によって DNA からのタンパク質の読
み取り、すなわち転写が、転写因子によって制限されて細胞の多様性が生じます。DNA
の塩基配列は転写因子によってメセンジャーRNA(mRNA)と呼ばれる塩基配列に転写さ
れます。細胞分裂では DNA が複製されますが、DNA から mRNA の転写も DNA の複製
と同じような方法で行われます。
ヒトの転写因子は約 2,000 種類があると言われています。DNA に書き込まれたタンパ
ク質(遺伝子)は 3 万種以上あるということは、一つの転写因子は幾つかのタンパク質の
読み取りを担当しているということになります。このため DNA に書き込まれたタンパク
質情報には特定の転写因子が結合する転写エレメント*があります。
18
ANM176®が認知症の予防と改善に役立つ可能性について
* 転写因子が認識して結合する DNA 領域
の特異的塩基配列を転写エレメントと
呼びます。転写エレメントには様々な系
列があり、その系列に関連するタンパク
質が連動して DNA から読み取られま
す。
A
個体の各細胞にある DNA の塩基配
列は同一ですが、同じ種であっても別
の個体とは、その配列が少しずつ異な
ります。DNA に書き込まれたタンパク
質のアミノ酸が他のアミノ酸に置き換
B
わっていることが集団の中で、ある程
度の頻度で見られ、それに伴って形質
の変化をもたらす場合(例えば髪の毛
の色や血液型など)、その遺伝子は遺伝
的に多型であると言います。また、遺
伝子 DNA 中で形質変化を起こさない
で、塩基 1 つだけ異なっている場合を
一塩基多型(以下では single-
C
nucleotide polymorphisms から SNP
と表記します)と言います。ヒトの
DNA にある SNP は約 300 万ヵ所以上
あると言われています。SNP は劣勢遺
伝子の中にあって、次の変異のベース
となります。ヒトの DNA に書き込ま
れたタンパク質を決定する塩基配列が
複数見られる遺伝子コピーが約 3,000
個あり、これを遺伝子コピー数多型と
いいいます。また、タンパク質を決定
する DNA の塩基配列の中に同じ配列
が重複している場合があり、10 数から
数 10 個の塩基が繰り返している場合を
配列多型と言います。これらの遺伝子
多型の他に 、DNA のメチル化なども
個人ごとに異なり、その結果、個性が
現れ、病気に対する感受性や体質・素
質が個人ごとに異なります。
図 13.細胞核の概要
A.DNA は細胞核内にある染色体中に含まれ、ヌクレオソ
ームと呼ばれる核酸が連なった長い分子 2 本の DNA
が二重となって螺旋構造を形成しています。
B.DNA の二重螺旋は、アデニン/チミン、グアニン/シト
シンという核酸の組み合で 2 本の DNA が結合していま
す。
C.核酸結合の分子式
A:MYCODE,
B:「理解しやすい新生物」水野丈夫編 文英堂, C:Z 会高校
講座のホームページより改変
19
ANM176®が認知症の予防と改善に役立つ可能性について
細菌など原始的な単細胞生物の多く
の細胞は、DNA が細胞質の中にある原
核細胞ですが、ヒトなど高度に進化した
多細胞生物の細胞は DNA が細胞の中心
部分の細胞核の中にある真核細胞です。
真核細胞の場合には細胞質内に 小胞体
(以下では endoplasmic reticulum の頭文
字から ER と表記します)と呼ばれる袋状
の構造が張り巡らされており、核の周りの
核膜と繋がっています(図 14)。
タンパク質の合成は、まず DNA から転
写され核の外に輸送された mRNA が、ER
の膜に埋め込まれたリボソームと呼ばれ
る粒のような構造物(図 15)に結合しま
す。真核生物のリボソームは、大サブユニ
図 14.細胞核の概要
(細胞の中心部分だけを示している)
(1) 核膜 (2) リボソーム (3) 核膜孔 (4) 核小体
(5) 染色体 (6) 細胞核 (7) 小胞体 (8) 核質
WIKIPEDIA より改変
ット 60S と少サブユニット 40S(S は沈降
速度を表す Svedberg の略で値が大きい
ほど沈降速度は速く、大きいこ
とを示している)がダルマのよ
うにくっ付いた形をしていま
す(図 16)。リボソーム上で
mRNA からタンパク質に転換
されることを翻訳といいます。
mRNA のアミノ酸配列情報に
基づいて順番にアミノ酸が結
合し、ER の表面で DNA に書
き込まれた配列が 翻訳されて
タンパク質が合成されます
図 15.リボソーム上でのタンパク質の翻訳
真核生物のリボソームは大サブユニット 60S と少サブユニット 40S か
ら成っており、小サブユニットに mRNA が結合します。その後に大サ
ブユニットが結合して、開始コドンを持った開始複合体が結合し、続い
て mRNA のコドンに該当するアミノ酸をもった tRNA が結合します。
Ajamaica(分子生物学)Net より
(図 17)。
リボソーム上の mRNA からタンパク質の翻訳は翻訳制御因子で調整されます。真核生
物でのタンパク質合成の開始には、少なくとも 10 種類の真核生物開始因子(以下では
eukaryotic initiation factor から eIF と表記します)が必要で、eIF には、それらを構成
するサブユニットや関連因子を含めると 30 種類以上が知られています。翻訳の開始は、
開始コドン(AUG)が付いた転移 RNA(transfer RNA;tRNA)と eIF2 などが結合した
翻訳開始複合体が形成されるところから始まります(図 16)。この翻訳開始複合体には、
さらに eIF3 などいくつかの翻訳開始因子とリボソームの小サブユニット 40S が結合し、
20
ANM176®が認知症の予防と改善に役立つ可能性について
同時に eIF4A、eIF4E および eIF4G は複合体
eIF4F を形成して mRNA 分子の 5’末端(読み取り
がスタートする側、読み取りの最終側は 3’末端)に
あるグアニンがメチル化されたキャップ構造に結合
します。キャップ構造は翻訳の開始に関係するだけ
でなく、mRNA の保護にも寄与しています。この
eIF4F を目指して翻訳開始複合体が mRNA に引き
寄せられ、開始コドン*(AUG)に行き当たるとリ
ボソームの大サブユニット 60S が結合してタンパク
図 16.tRNA の構造と翻訳
質合成が開始されます。
*開始コドンはホルミル化(-COH 基が結合)されたメチ
オニンが結合しており、タンパク質の最初のアミノ酸が
リボソームに結合する時に外れます。
その後、個々のアミノ酸と結合した tRNA は該当
する塩基配列(コドン)を認識して、開始コドンの
mRNA 上のコドンに該当するアミノ酸を
3’-末端に結合した tRNA 中には、mRNA の
コドン(上図の例では AUG)を認識するア
ンチコドン(UAC)があり、リボソームで
翻訳中のタンパク質の末端に該当するアミ
ノ酸を引き渡します。AUG はメチオニン
(Met)の塩基配列。
(図は「薬の情報~専
門薬学ウェブサイト」より加工)
後のアミノ酸配列に従って
リボソームに運ばれ、タン
転写因子
パク質が合成されます(図
15)。
elF2 はα、β、ɤ のサブ
ユニットから成っており、
このαサブユニットが
eIF2αキナーゼによってリ
ン酸化されると開始複合体
が形成されず、翻訳が抑制
されます。eIF2αキナーゼ
には何通りかの系列があっ
て、タンパク質の抑制に関
与して恒常性維持(ホメオ
スタシス)に寄与していま
す。もう一つの翻訳開始複
合体が形成を阻害してタン
図 17.DNA の読み取りとタンパク質への翻訳
転写因子が関与して DNA のエキソンから転写された mRNA は核膜孔か
ら核外に運ばれ小胞体膜に結合したリボソーム(小サブユニット 40S と大
サブユニット 60S がダルマ状に結合している)でタンパク質に翻訳されま
す。合成されたタンパク質(ポリペプチド)は小胞体内で自動的に折りた
たまれます(フォールディング)。
(永田和宏他「細胞生物学」放送大学教育振興会から改変)
パク質翻訳を抑制する機構
は、複合体 eIF4F を構成するユニットの 1 つである eIF4E に eIF4E 結合タンパク質
(以下では eIF4E binding protein から 4EBP と表記します)が結合することによって
eIF4F 複合体形成を抑制するメカニズムです。
21
ANM176®が認知症の予防と改善に役立つ可能性について
生物の恒常性維持について
生物としてのヒトは 38 億年という壮大な時間をかけて進化してきました。生物の進化の
過程には、過激な自然環境の変化や病原菌や肉食動物など他の生物による厳しい攻撃があ
りました*。
* 現在の地球の大気は窒素(78%)、酸素(21%)が主で、二酸化炭素はごくわずか(約 0.03%)です。
しかし、約 45 億年前に誕生した原始地球の大気は、ほと
んどが二酸化炭素で、酸素はなかったそうです。
そのため、38 億年前に誕生した生物は、酸素のない環境
でしか生きられない嫌気性の生物でした。
32 億年前に現れた光合成生物によって酸素が排気され、
次第に海洋中の酸素濃度が上昇してきました。しかし、そ
れは生命にとっては危険でした。酸素は細胞膜や遺伝子を
傷つけ生命に害を及ぼすからです。この危機を乗り越える
ことができたのは「酸化酵素」の獲得です。ヘムと呼ばれ
る酸素分子を活性型に変換する補欠分子(タンパク質の機
能を補う分子)(図 18)を進化で持てるようになって酸素
分子を物質の分解に利用できるようなりました。この結
果、生物は、酸素を無毒化し、酸素が遺伝子を傷つけるこ
とから自分を守ることが可能になったのです。また、この
酸素を利用し、有害物質を分解することができるようにも
なりました。さらに、ミトコンドリアを獲得し、酸素を利
図 18.ヘムの構造と役割
用して大きなエネルギーを作り出すことができるように
ヘムはポルフィリン骨格に鉄イオンがキ
なったのです。
レート結合によって挿入された構造をし
約 27 億年前の地球冷却化にともなって生じた地磁気と
たヘモグロビン、ミオグロビン、チトクロ
ームなどヘムタンパク質に結合する補欠
地球の周りの磁気圏は、生命に有害なα、β線などの太陽
分子です。ヘムタンパク質は酸素分子と結
風を遮断するようになりました。
合でき生体内の酸素の利用、運搬に機能し
約 20 億年前に真核生物が誕生しました。真核生物が原
ています。
核生物と異なるのは、原核生物は細胞の中に広がっていた
DNA を真核生物では細胞の中心にある核の中にコンパクトに圧縮し大量の情報を担うことができるよ
うになったのです(10 ページ注****)。
約 19 億年前から本格的な大陸が出現しました。
7.5~6 億年前には地球は大きな氷河期に突入し、地球の冷却と凍結は生命に危機を及ぼしました。そ
れが過ぎると大気と気温は次第に安定し、生物が繁栄するための環境が整ってきました。
約 5 億 5 千年前、それまで数十種しかいなかった生物が、突如 1 万種にも爆発的に増加しました(カ
ンブリア爆発)。こうして本格的な生物同士間の生存競争が始まりました。この時期に、生物に体の中
に脊索を持つ脊索動物が生まれ、後に、この脊索動物から脊椎動物が誕生します。
大気中に増えて成層圏まで達した酸素によって、5 億年前にオゾン層が形成され、生命に有害な紫外
線が防がれて、地上で生命が生き延びられる環境が整ってきました。
植物が本格的に地上に進出したのは約 5 億年前です。重力に耐えられる強靱さなど植物は自分の体の
構造を適応させながら陸地に進出していきました。植物の上陸に続き、約 4 億年前頃から昆虫類も地上
に進出します。
約 3 億 6000 万年前に両生類が誕生し、後の爬虫類、鳥類、ほ乳類へ進化しました。
約 6,500 万年前の新生代に入り、地球の気温は一旦暖かくなりましたが、約 4,000 万年前頃から気温
が低下してきました。特に 180 万年前から始まる第 4 期は本格的な氷河期の時代でした。現在は第 4 氷
河期の比較的温度が落ち着いている間氷期です。
ヒトの祖先はアフリカ大陸で誕生したと言われています。大昔にアフリカ大陸の森林で暮らしていた
サルは豊富な食料に囲まれて暮らしていました。そこにプレートの移動によるアフリカ大陸を南北分断
する亀裂(グレートリフトバレー)が起こり、環境が激変しました。亀裂の西側は昔通りの森林に覆わ
れていましたが、東側では降水量が減り、森林は草原地帯へと姿を変えました。草原地帯での生活を強
いられたヒトの祖先は、生き残りのために、自らを変化させていく必要に迫られました。
類人猿とヒトが分かれたのは約 500 万年前のことと言われています。知能が発達した現代人の直接の
祖先であるクロマニヨン人は 15 万年前に誕生しました。
ヒトは誕生してから 15 万年にすぎませんが、この誕生は 38 億年にわたる過酷な生物進
22
ANM176®が認知症の予防と改善に役立つ可能性について
化の結果であり、ヒトの DNA には 38 億年という進化の歴史が書き込まれていると言える
でしょう。ヒトの DNA に書き込まれている情報は、文字数にすれば 30 億字以上で、本に
すると数万冊分です。ヒトの DNA に書き込まれた約 3 万種のタンパク質は、DNA が読み
取られる時に選択的スプライシング*と呼ばれる方法によって、更に 10 倍のタンパク質が
作られると言われています。
* 真核生物の細胞では DNA がそのまま mRNA に転写されることは稀で、ある部分(イントロン)は
除去されてエキソン(図 17)のみが再結合してmRNA が形成されます。この機構を選択的スプライ
シングといいいます。 選択的スプライシングによって 1 つの遺伝子から異なる構造のタンパク質が
複数でき上がります。
進化の過程が如何に過酷であったかの 1 例を挙げましょう。植物は食べられないように
様々な毒物を新たに作り出し、それに対して動物は新たな毒物を無毒化できるように進化
してきました。その結果、動物は、毒物を分解するチトクローム p450(以下では p450 と
表記します)という酸化酵素の遺伝子を約 700 種類(ヒトでは約 50 種類)以上もっていま
す。更に、1 種類の p450 には、微妙に異なる遺伝子が 100 種類以上あります。つまり、植
物の毒を分解するために、7 万種もの p450 が進化したと考えられ、植物とそれを食べる動
物の攻めぎ合いの激しさが窺えます。こうして進化した p450 遺伝子の種類と数は人種や個
人によって異なることから、医薬品の分解速度に個人差が現れ、個々の人によって医薬品の
効果が異なる原因の1つにもなっています。
一方、必要でなくなったタンパク質は、進化の過程で DNA から読み取られなくなります。
例えばビタミン C は必須ビタミンですが、食べ物からビタミン C を摂取できるようになっ
たヒトには、DNA にビタミン C 遺伝子の痕跡があっても実際には役に立っていません。生
物は結構横着で、外部から吸収できるものを優先して使う傾向があるようです。長いこと外
部から吸収し、その遺伝子が機能しない進化が起こっても差しさわりはないでしょう。しか
し大航海時代になると、長い間経験しなかったビタミン C 欠乏症に陥ったのです。このビ
タミン C の例のように、生物はいつ訪れるか予測がつかない環境の変化や攻撃に対して常
に備えるだけの余裕はなく、生命を脅かす様々な生物や物質あるいは環境の変化に対応す
る場合も、その事態になってから対応する免疫という仕組みを備えるようになったと考え
られます。
進化で獲得したもう一つの柱は、エネルギーの利用方法でした。生物にとってエネルギー
は必要不可欠です。一方、自然界のエネルギーは豊富な時と不足する時が気ままに訪れます。
様々な事態を乗り越えて進化してきた生物には、この不安定なエネルギーを有効に利用す
る方法が備わっています。エネルギーが豊富な時は、植物では澱粉として、動物ではグリコ
ーゲンとして細胞内に蓄積し、不足時や異常事態には、蓄積した澱粉やグリコーゲンからエ
ネルギーを作り出すことができます。細胞が分裂するためにはエネルギーを十分に蓄えて
生長する必要があります。エネルギーが満ち足り細胞が生長*すると 1 つの細胞が分裂し、
分裂した細胞が生長を繰り返します。
*ここでは、形態が発達して大きくなることを成長、太ったり熟成したり栄養的に増加することを生長
といいます。
23
ANM176®が認知症の予防と改善に役立つ可能性について
エネルギーが長期間不足すると、細胞分裂などあらゆる活動をなげうって、最低限の生命生
命維持だけに集中します。
どんな状態においても、細胞内は、生物が誕生した時の海のように、一定の安定した環境
に保つ仕組みがあり、この安定した状態を恒常性(ホメオスタシス)と呼びます。エネルギ
ー供給の元である太陽の動きに対する生物内の日内変動も、生物に備わっているホメオス
タシスの仕組みの一環と言えます。危機に遭遇した時には生存を優先したストレス応答(16
ページ注*)のパターンに切り替えられ、蓄えたエネルギーを総動員して、体の一部を犠牲に
してでもホメオスタシスを維持し、生命を保ちます
50)。異常事態に遭遇した時のストレス
応答の相(パターン)切り替えのスイッチは、高度に進化した様々な仕組みによって巧妙か
つ厳密に実行されます。ストレス応答時にエネルギーが不足すると細胞は自己死のアポト
ーシスにスイッチを入れることもあります。
(細胞ストレスについて)
「天災は忘れた頃にやって来る」の譬(タトエ)のように、生物は、いつ異常事態に遭遇す
るか分かりません。しかし、前項で述べたように、生物の細胞も常に目一杯に、あらゆるス
トレスに備えるだけの余裕はなく、ストレスが来た時に対処します。
よく風邪のウイルスに免疫があるとかワクチンによって免疫を付けるとか言いますが、
免疫とは、文字通り、病気にならない、あるいは病気を軽くする仕組みで、微生物からヒト
に至るまで、生物に幅広く備わっています。よく言われるウイルスに対する免疫やワクチン
による免疫は獲得免疫のことです。獲得免疫は、出会ったことのある病原菌や物質を記憶し
て、次回に同じものに出会ったときは直ぐに対応できる仕組みで、背骨の構造のような脊椎
を持った脊椎動物にしか備わっていません。脊椎動物以前の生物には、遭遇した事態に、そ
の都度ごとに対処する自然免疫の仕組みしかありません。脊椎動物が高度に進化できたの
は、獲得免疫システムを備えたことによると言われています。獲得免疫のおかげで生存率が
格段に高まり、遺伝子に多くの情報を蓄えることができるようになったのです。しかし皮肉
なことに、ヒトは、獲得免疫系の仕組みを備えたために、花粉症など様々なアレルギー疾患
を引き起こすリスクが生じます。また高齢になると生活習慣病になるのは、生存率が高くな
って寿命が延びたことが関係しています。
自然免疫は、英語でinnate imunity(内因性免疫)と言い、まさに体に備わった内因性
の免疫の意味です。脊椎動物でも獲得免疫で異常事態を感知した後は、生物に共通した自
然免疫システムで対処します。異常事態に遭遇した時、細胞膜にある受容体は異常事態を
感知して細胞内にシグナリングし、このシグナルによって様々なタンパク質が細胞の内外
に現れ、細胞は生命維持に向けたストレス応答をします51)。このストレス応答には一連の
パターンがあります。例えば風邪をひいた時、咳、鼻水、体温の上昇などの炎症の症状が
現れます。風邪で起こる炎症や飢餓に対する応答は分かり易いですが、細胞内でのタンパ
ク質合成を制御する小胞体ストレス応答、異常物質の分解に関与する酸化ストレス応答、
24
ANM176®が認知症の予防と改善に役立つ可能性について
低酸素状態に対処する低酸素応答などは分かり難いですが、これらも細胞で行われている
重要なストレス応答です。さらに、これらのストレス応答には細胞が死滅するアポトーシ
スの仕組みも繋がっています52)。例えば、化膿した皮膚が瘡蓋(カサブタ)となって、そのう
ちに、はがれ落ちるのはアポトーシスによって傷んだ部位が切り捨てられるからです。ア
ポトーシスは見えないところでも常に起こっており、体を構成する細胞の新陳代謝も関与
しています。ストレス応答によって、当面必要のない細胞がアポトーシスによって切り捨
てられます。ストレス応答は個体の生命を守ることが最終的な目的で、そのために生命と
は関係しない日常の活動は差し置かれます。考えてみれば当然のことです。
ストレス応答には、風邪をひいた時のように即座に対処しなければならない場合と、そ
の後の回復期のように長時間かけて対処する場合があり、同じストレスに対しても時期や
状況によって仕組みが異なります。ストレス応答が長引いて、いつまでたっても対処でき
ない場合は慢性化して、場合によっては慢性病や個体の死につながることにもなります
53)。細胞は病原菌だけでなく花粉など様々な物質を感知し、場合によってはアレルギーを
起こします。アレルギーは異物に対するストレス応答が慢性化した状態です。
これらのストレス応答の仕組みは、状況によって相互に関係します。たとえば、風邪を
ひいた訳ではないのに、急に寒くなったりすると風邪と同じような症状が起こるのは、事
態は異なっていても、ストレス応答の一部が共有されているからです。
(小胞体ストレス応答)
真核生物で mRNA からタンパク質を翻訳する現場の小胞体(ER)(図 14、17)は、タ
ンパク質合成に限らず、糖などをタンパク質に付けるなどのようなタンパク質 2 次加工や
タンパク質の構造を管理するなどのタンパク質品質管理に重要な場所です。タンパク質は
ER で合成された後、自動的に立体構造を取ります。タンパク質の立体構造は酵素の反応
やシグナリングのアロステリック制御(15 ページ)に関係しますので、正確であることが必
須です。そのため ER では、合成したタンパク質が一定の構造を取っているか否かの品質
管理を行います。正確な構造が取れなかったタンパク質は再び組み立て直されるか、ER
の外に排出されて分解されます。
ストレスによって撹乱された ER の状態を小胞体ストレスといいます 54)。タンパク質の
合成場所である ER は内外のストレスを受けやすいため、ER にはタンパク質合成を抑制
するなど小胞体ストレスを回避する仕組みが備っており、これを小胞体ストレス応答(以
下では unfold protein response から UPR と表記します)と言います 55,56)。UPR は文字
通り、タンパク質の折り畳み(fold)がうまくいかなかった時の応答です。外部ストレス
によって立体構造が異常なタンパク質が増えたり、ER の働きが限界に達したりした時に
UPR が起こり、一時的にタンパク質の合成が抑制されます。こうした UPR によって ER
の働きが減速され、また、ER に送り込まれるタンパク質量が抑制されて、ER の負荷が低
減されます 55,56)。
25
ANM176®が認知症の予防と改善に役立つ可能性について
UPR には IRE1、PERK、ATF6 と呼ばれるタンパク質がストレスセンサーとして機能
しています。平常時は、これらストレスセンサーに GRP78 という小さなタンパク質(シ
ャペロン分子と言います)が結合しています 57)。UPR によって GRP78 が外れると
PERK 、IRE1、ATF6 が活性化し、活性化した PERK はタンパク質合成の全般を停止さ
せます 55,56)。また、活性化した IRE1 と ATF6 は、ER の品質管理に関連する様々なタン
パク質の DNA からの読み取りを促進します 58)。UPR に必須の因子が欠如したり、ER の
機能低下が起こったりするとアポトーシスを誘導し、その細胞が死ぬことにもなります。
(酸化ストレス応答)
細胞は、大量の異物や微量であっても毒物や病原性微生物など異常な物質(これらを外来
異物といいます)に出会った場合、酸化ストレス応答を引き起こします。酸化ストレス応答
と呼ばれるのは、外来異物を細胞内に取り込むと酸化分解に係る様々なタンパク質が細胞
内に現れるからです。外来異物は血液やリンパ液あるいは組織中にあるマクロファージな
どの食細胞*内に取り込まれ 59,60)、その細胞内で酸化分解されます。
*外来異物をアメーバ―のように取り込み分解する食細胞にはマクロファージ、単球、好中球、破骨細
胞、樹状細胞などや脳内だけに存在するミクログリアがあります 59,60)。
外来異物の分解には数百種類の酸化酵素が関与し
A
ます。これら酸化酵素には酸素分子を活性型に変換
する補欠分子ヘム(図 18)が結合しています。酸素
は生体内のエネルギー代謝にも関係しますが、生体
物質を酸化して生命を障害する可能性もありますの
で、ヘムの制御は重要です。血中で酸素を運ぶヘモグ
ロビンは、酸素分子と結合しても酸化反応ができな
くなったヘム酵素の変異型で、酸素の多いところで
は酸素分子に結合して、酸素の少ないところで結合
していた酸素を放します。参考までに、ヘムは分子の
中に鉄原子を取り込んでいますが、ヘムと似た分子
B
の骨格の中にマグネシウム原子を取り込んだクロロ
フィルは植物中で太陽の光を利用して水を活性化
し、それを分解して酸素分子を放出します。
酸化分解酵素は、様々な物質を酸化分解できます
が、一方、酸化分解を受けた副生物は何にでも反応す
るため即座に処理されなければなりません。そのた
め、生物内では分解に係る酵素群(第Ⅰ相系酵素群と
いいます)と中和に係る酵素群(第Ⅱ相酵素群といい
ます)が連動して働きます。生物が進化で酸素を活用
図 19.NADP の分子構造
生物で利用するエネルギーは活性化した
電子と考えられます。電子は原子の電子
軌道にあるりますが、光や他の分子の影
響を受けて不安定となり活性化します。
NADP(図 A)のアデニンジヌクレオチ
ドリン酸に結合したニコチンアミド(図
B)の水素(H)の活性化した電子(e-)
は不安定で、生物内で行われる酸化反応
に利用されます。
26
ANM176®が認知症の予防と改善に役立つ可能性について
できるようになったのは(22 ページ注)、この第Ⅰ相系酵素群と第Ⅱ相系酵素群の連動システ
ムを獲得できたからといっても過言ではありません。
第Ⅰ相系酵素群は酸化分解に酸素分子の電子を使う p450 類と過酸化水素の電子を使う
ペルオキシダーゼ類が代表的です。この酸化分解で、還元型のニコチンアミドアデニンジヌ
クレオチドリン酸
(以下では nicotinamide adenine dinucleotide phosphate H から NADPH
と表記します)が酸化されて NADP となります(図 19)。NADPH は生物界に幅広く存在
し、光合成経路あるいは解糖系経路などで用いられている基本的な電子担体で、NADPH ⇌
NADP の変換によって細胞内内外の電子伝達に機能しています。
第Ⅰ相系酵素群による分解物は、即座に、第Ⅱ相系の酵素群によってグルタチオン(以
下では glutathione から GSH と表記します)やグルクロン酸などの抱合物質(酸化分解
物と結合して毒性を中和することを抱合といいます)に結合して無毒化され、体外に排泄
されます。GSH の場合はグルタ
チオン S -転移酵素(以下では
Glutathione S-transferase から
GST と表記します)が抱合に関
与します(図 20)。
GSH は 3 つのアミノ酸が特殊
な結合をした物質で(図 21)、通
常のタンパク質分解酵素では分
図 20.グルタチオンの代謝
解されません。健康診断でよく
1
耳にするγ-GTP(γ-glutamyl
2
transpeptidase、γ-グルタミル
トランスフェラーゼ)はグルタ
チオンを直接分解できる稀な酵
素です。肝癌やアルコール性肝
障害など肝・胆道系に異常があ
るとγ-GTP が肝臓で増えて血中
に流出します。このため血中の
γ-GTP 活性は肝機能の指標とし
3
4
5
γ-グルタミルシステインシンセターゼ
グルタチオン合成酵素
γ-グルタミルトランスフェラーゼ(γ-GTP)
ジペプチダーゼ
グルタチオンS転位酵素(GST)
CysGly:シスチルグリシン
Csy:システイン
Gly:グリシン
Glu:グルタミン酸 γGluCys:γグリシルシステン
GSH:グルタチオン
X:γ‐グルタミル残基のアクセプター Y:外来異物
分解物と結合したグルタチオン(GS-Y)は排泄されます。
このサイクルで生成されたシスチルグリシン(CysGly)は
グルタチオン合成に再利用されます 787)。
て利用されます。γ-GTP の数値
が高いことは、肝臓での酸化分解活動が活発か、病
的な状態であることを示しています。
食細胞の仲間の好中球や単球の細胞内では、外来
異物に即座に対応して急速に酵素群が増加します。
この急速な反応を呼吸バーストといいます
61)。この
呼吸バーストによって、p450 類、ペルオキシダーゼ
図 21.グルタチオン
グルタミン酸側鎖のカルボキシル基と結合
したシステインがグリシンとアミノ酸結合
しています。
27
ANM176®が認知症の予防と改善に役立つ可能性について
類など酸化分解酵素類の遺伝子の転写が促進され、異物が酸化分解されます
62)。この分解
と中和・排泄の調和は、特定のタンパク質の転写に関わる転写エレメント(19 ページ注)で制
御されます。
第Ⅰ相系酵素群の転写には転写因子 AhR が関与し、AhR は補助因子と複合体になって
DNA の転写エレメント XRE に結合して p450 やペルオキシダーゼなど第Ⅰ相系酵素群の
転写を促進します 63)。XRE は異物応答エレメント(xenobiotic response element)の頭
文字です。第Ⅱ相系酵素群には転写因子 Nrf2(Nuclear factor erythroid 2-related factor
2)が DNA の転写エレメント ARE に結合し 64–66)、ARE を配列の中に含む GST、キノン
リダクターゼ、γ-GTP など第Ⅱ相系タンパク質が揃って読み取られます 67–69)。 ARE は
抗酸化応答エレメント(antioxidant response element)の頭文字です。Nrf2 を不活性化
した細胞では、弱い酸化ストレスにさえ耐えきれずに死を起こすと報告されています 70)。
細胞内の環境における第Ⅰ相系と第Ⅱ相系のバランス、すなわちレドックス制御 (redox
regulation) の機構は生体のホメオスタシスの維持において重要な役割を担っており、こ
の機構は多くの外来性の因子、すなわち薬剤、放射線、紫外線、環境物質である農薬、ダ
イオキシンなど、また、高熱、低温、低酸素状態に適応するように機能します。レドック
ス制御は酸化ストレスを契機とするストレス応答反応とも捉えることができるため、レド
ックス制御と酸化ストレス応答は表裏一体の関係にあると考えられています 71)。
(虚血ストレス応答)
脳梗塞などにより一時的に血液が届かない事態を脳虚血と言います。脳虚血を起こした
組織は著しく障害され細胞が死ぬことにもなります
72)。低酸素によっても虚血と同じ現象
が起こります。高山病も虚血障害の一種と考えられます。一酸化酸素中毒によって知的障害
が起こるのは、脳のニューロンが酸素欠乏状態に脆弱(ゼイジャク)なところに、酸素を運ぶ
血中のヘモグロビンに一酸化炭素が結合してニューロンに酸素が届かなくなったことによ
って脳虚血と同じ事態になり、起こる障害です。また、一時的に心機能が停止した後も、脳
への酸素運搬量が減少して同じ事態となります。一時的な脳虚血の後、再び酸素が戻っても、
ニューロン死のプロセスは止まりません。このように、細胞が低酸素状態で起こす一連の応
答を虚血ストレス応答(または低酸素応答)と言います。
動物による脳虚血では、脳梗塞が数時間から数日かけて虚血中心部から周辺に広がって
いきます 73)。脳虚血によって、脳中の異物掃除をしているミクログリアが活性化して(6 ペ
ージ「中枢神経系について」)
、IL-1β74)など炎症に関係する様々なサイトカイン(13 ページ「細胞
内へのシグナリング」)などが多数活性化され、また、虚血における細胞内シグナリングには
MAPK 経路の p38(15 ページ「キナーゼによるシグナリング」)が関与していると報告されていま
す 75,76)。また、脳虚血ストレスで細胞のサバイバルに関与する Akt の活性が低下し(15 ペー
ジ「キナーゼによるシグナリング」)
、その結果、アポトーシス(細胞の自己死)を引き起こすと
報告されています 77,78)。
28
ANM176®が認知症の予防と改善に役立つ可能性について
ニューロンを補助しているアストロサイトは(6 ページ「中枢神経系について」)、脳梗塞などの
虚血性の血管疾患状況においてグリア線維性酸性タンパク質(以下では glial fibllial acids
protein から GFAP と表記します)*を活性化して炎症に抑制的に作用し、ニューロンの防
御に重要な役割を果たしていると報告されています 73,79)。
*GFAP はアストロサイト中で見られる線維で、しばしば、アストロサイトの活発な形状変化と GFAP
が同時に観察され、GFAP はアストロサイトの細胞の骨格を形成する役割があると考えられています
80)。アストロサイトの活性化と加齢によって GFAP が増強されます 81)。
(炎症とは)
一般には、患部が赤くなったり(紅斑)
、痒くなったり、腫れたり、熱を帯びたり、ある
いは喉が痛くなったりすることを「炎症」と呼んでいます。この症状は炎症メディエーター
と呼ばれるヒスタミンやブラジキニンが分泌されることによって起こります。ヒスタミン
は血管を拡張させ、紅斑や発熱を発することに関与します。ブラジキニンは皮膚などにある
感覚受容器を刺激して痛みや痒みを引き起こします。
細胞における炎症は、もっと広範囲で、様々なストレスに対して緊急に対処するストレス
応答につながった仕組みです。異物の種類や環境の変化などストレスの内容に応じて、前項
で述べたような小胞体ストレス応答や酸化ストレス応答などが起こります。その応答後の
処理、例えば、問題を起こした物質の除去、問題を局部で解決し他の部位に波及させない、
問題が起こった患部の修復、などがあります。これらの処理は、様々なストレス応答に共通
しており、炎症として様々なタンパク質の活性化や DNA から新規なタンパク質の転写が行
われて、元の状態(ホメオスタシス)に戻すことを目指します。
感覚受容器が受け止めた刺激を脳で感じる痛みや痒みは、炎症患部を休ませ保護する等、
動物にとって意味がありますが、感覚受容器がない体中では炎症が起こっているかどうか、
自分の感覚では分らず、とかく見過ごしがちです。
細胞内の炎症は免疫機構(24 ページ「細胞ストレスについて」)の一部で、様々なストレス応答
に連動した自然免疫の現象です。病原菌に感染した時、組織が損傷した時、毒物や異物に遭
遇した時など異常事態において、炎症性サイトカイン(13 ページ注)が細胞外に分泌され 82,83)、
自身や他の細胞にシグナリングし、リンパ球の活性化や動員が行われ、様々なタンパク質が
分泌され、自然免疫系の応答(24 ページ「細胞ストレスについて」)を促進します 84)。炎症は細胞
の死、すなわちアポトーシス(16 ページ注**)にも繋がっていますが、問題が局部で解決さ
れないで全体に及ぶと重大な病気や場合によっては個体の死をもたらすことにもなります。
また、何時までも問題が解決されないで炎症が続く場合は、慢性の病気になることもありま
す。
脳内でも自然免疫のシステムがあり炎症が生じます。ただし、脳には獲得免疫系はあり
ません。その理由は、免疫応答は神経作用を誤作動させる可能性があり、BBB(9 ページ
「脳関門」)が血液やリンパ液に乗ってやって来る物質を厳密に選別し、獲得免疫系でター
ゲットとされる細菌やウイルスあるいは異物は脳には入れないからと考えられます。しか
29
ANM176®が認知症の予防と改善に役立つ可能性について
し、脳内においても、トラウマ、梗塞、毒素などによる様々な刺激によって、即時的で一
時的な内因性炎症システムの活性化が誘導されます 85,86)。このことから、脳内においても
免疫や炎症のダイナミックな応答があり得ると考えられています 87,88)。脳内にしかないミ
クログリアは(7 ページ)、活性化すると同時に大量の炎症性因子を放出することから 89)、脳
内でも炎症が起こっていると考えられています 90)。マウスなどを使って脳梗塞を実験的に
起こした研究では、梗塞進展に先立ってミクログリアが活性化され 91)、小胞体ストレス応
答(UPR)が起こり 92)、IL-1β、IL-6、INFγなど様々な炎症性サイトカインあるいは炎
症に関連するプロスタグランジン合成酵素(以下では cyclooxygenese から COX と表記し
ます)やアラキドン酸代謝経路の活性化(図 22)*などを誘導し、炎症反応の証拠が観察
されています 74)。
*細胞膜のホスファチジルイノシ
トールに結合しているアラキド
ン酸からプロスタグランジン
(prostaglandin )H2(PGH2)
が合成されます。膜結合タンパク
質の COX はヘム酵素で、このア
ラキドン酸から PGH2 合成の反
応に関わっています。PGH2 は最
終的に PGE2、PGD2、PGI2 に変
換されます 93)。
最近、PGE2 はマスト細胞か
らヒスタミンを分泌させること
が示されました 94)。また、血液
タンパク質ブラジキニンはホス
ホリパーゼ A2 を活性化させて
図 22.アラキドン酸から各種プロスタグランジンの合成
PG 合成を促進させ、血圧を低
下させる方向に働きます。
COX には、COX1 と COX2 の 2 つのアイソマーがあり、COX1 は刺激により誘導されることはな
く常時細胞内に一定量が存在しています。一方、COX2 は脳や腎臓などでは恒常的に存在していま
すが、その他の組織では、種々の刺激により炎症組織において誘導されます 95)。COX2 は主に核膜
に存在し、IL-1α、TNF-α、細菌外殻のリポ多糖類(以下では lipopolysaccharide から LPS と表
記します)など炎症刺激物質や多くの発癌物質による刺激により誘導されます。一方、COX2 の活
性はステロイド類によって強く阻害されます。非ステロイド抗炎症剤( non-steroidal antiinflammatory drugs;NSAID)によって活性が抑制するのも COX2 です。
アスピリンは例外で、COX1、2 両方をアセチル化することで不可逆的に阻害します。副腎皮質ス
テロイドホルモンは、COXs の合成を阻害して抗炎症作用や鎮痛作用などを現します。
(細胞毒性)
細胞毒性(cytotoxicity)は、細胞に対して死、もしくは機能障害や増殖阻害の影響を与
える物質や作用などを言います。生物は細胞で成り立っているため、細胞に障害を引き起こ
す物質や作用の細胞毒性の判定は、生物の生命維持活動を解明する上で重要です。細胞毒性
の要因としては、細胞を形作る物質・構造の破壊、細胞の生存に必須な呼吸、基本的代謝、
DNA の複製や転写あるいは翻訳等の阻害、細胞周期や細胞内シグナル伝達への影響など、
様々な現象が考えられます。
外来異物に遭遇して細胞にストレスが加わると、平常のパターンが停止してストレス応
30
ANM176®が認知症の予防と改善に役立つ可能性について
答のパターンに切り替わり、酸化分解や分解物の中和などに関連するタンパク質が新規に
現われ、また、それらのタンパク質の活
性化が起こります。
細胞毒性を判定する様々な方法があ
りますが、その中で MTT 法
A
96)*と呼ば
れる方法は、細胞内の酸化分解に関連す
るタンパク質が活性化されているか否
かを観察し、細胞毒性の有無を判定しま
す 97)。
* MTT はテトラゾリウム塩の一種である
dimethylthiazolyl-diphenyltetrazolium Bromide のことで、その水
溶性は黄色ですが、酸化酵素によって
MTT はホルマザンに変化します。このホ
ルマザンは青色の非水溶性の結晶で生成
後は沈殿します。これを有機溶媒で溶解
させると赤紫色の溶液となり、細胞毒性
が観察できます。
B
MTT 法による発色は、そこに活発な
電子があることを示しており、そこでは
ミトコンドリア*のコハク酸脱水素酵素
の活性化 98)や小胞体 99,100)、細胞質 100)、
細胞膜
101,102) 等にある電子伝達系が活
発に活動しています
98,102)。細胞内の電
子伝達系は分解中の物質から取り除か
れた電子を受け止めて、その電子を、エ
ネルギーを生み出す機構や物質を組み
立てる機構あるいは解毒する機構に運
ぶ役割を果たしています。
*ヒトの細胞には 1 個あたり平均 2,000 個
のミトコンドリアがあります。20 億年以
上前に、酸素を利用する好気性バクテリ
アが他の生物の体内に侵入した物がミト
コンドリアの始原ですが(22 ページ注)、
ミトコンドリアを細胞内に取り込んだ真
核生物の DNA は細胞核の中にあるた
め、自身のミトコンドリアが作り出す活
性電子による DNA の損傷から免れるこ
とができました。
ミトコンドリアでは ATP が合成され
ます。ミトコンドリアの内膜に埋め込ま
れた電子伝達系で、電子は、複合体 I
(NADH デヒドロゲナーゼ/ユビキノ
ン)→ 複合体 II(コハク酸デヒドロゲ
図 23.ミトコンドリアとその電子伝達系
A.ミトコンドリアの構造
ミトコンドリアは外膜と内膜の脂質膜に囲まれており、内
膜に囲まれた内側をマトリックス、内膜と外膜に挟まれた
空間を膜間腔と呼びます。内膜はマトリックスに向かって
陥入してクリステと呼ばれる特徴的な構造となっていま
す。内膜はミトコンドリアの機能的な特徴を担っており、
酸化的リン酸化に係る呼吸鎖複合体などの酵素群が規則的
に配列しています。内膜は外膜とは対照的に基本的には不
透性で、内外で物質を輸送するためには、それぞれの物質
に特異的な輸送体が必要となります。
マトリックスでは、TCA 回路やβ酸化などミトコンドリ
アの代謝機能に関わる酵素群が数多く存在しています。こ
こには DNA も含まれており、ミトコンドリア独自の遺伝
情報が保持されています。
B.ミトコンドリアでの ATP 合成
ミトコンドリア内側の TCA 回路によって ATP が生産され
ます。解糖系によって 6 個の炭素があるブドウ糖を 3 個の炭
素のピルビン酸まで分解し、このプロセスで ATP を 2 個、
NADH を 2 個が生まれます。ピルビン酸はミトコンドリア
の中に入り、炭素が 2 個のアセチル CoA になります。この
時、炭酸ガス CO2 と NADH を放出します。アセチル CoA
は回路からやってきたオキサロ酢酸と化合し、クエン酸にな
り(この時、CoA を放出し)
、さらに TCA 回路の例の連続的
な酸化反応が進みます。
31
ANM176®が認知症の予防と改善に役立つ可能性について
ナーゼ/ユビキノン)→複合体 III (コエンザイム Q/シトクロム c レダクターゼ) →複合体 IV(シ
トクロム c オキシダーゼ)の順番に、一連の酸化還元反応を通して最終的な電子受容体である酸素
分子に移動します。電子の伝達によって得られたエネルギーは、ミトコンドリアマトリックスから
膜間空間に活性電子を移動するのに用いられ、このとき輸送された活性電子によりミトコンドリア
内膜の内外に電気化学的なエネルギーが生じます。これが原動力となり、ATP 合成酵素が ADP と
無機リン酸から ATP を合成します(図 23)。
この電子の活動が細胞毒性と関係する理由は、ミトコンドリアの活動と好中球と呼ばれ
る免疫系細胞の働きから推測することができます。生きた細胞では、ミトコンドリアにあ
る脱水素酵素の活性によって MTT 法で発色しますが、死んだ細胞では発色しません。好
中球は、細菌など外来異物と遭遇して活性化し、これら外来異物を細胞内に取り込んで分
解しますが、この時に、呼吸バースト(27 ページ)とよばれる急速な酸化反応が起こり
41,103)、電子伝達系が活発に活動して外来異物の分解に関与します。外来異物に出会った好
中球が活発に活動している状態は、まさに細胞が毒物に対処していることを示唆していま
す。異常事態によって活性化した G タンパク質は、細胞内にシグナリングすると自動的に
OFF になりますが(15 ページ)、このことからも異常事態の刺激は細胞にとっては危機的な
状況であることが窺えます。G タンパク質によって活性化された PI3K は膜にあるイノシ
トールリン脂質から PIP3 を作り出します(17 ページ)。PIP3 は小胞体 99,100)や細胞質 100)に
存在する数多くの酸化分解に関連する第Ⅰ、Ⅱ相系酵素群(27 ページ)を活性化します。
NADPH を NADP(図 19)に変換する NADPH オキシダーゼの活性化によって細胞質か
ら細胞膜に電子が運ばれます 41,104)。NADPH オキシダーゼは多くのユニットが集まって
機能する複合酵素です。好中球で呼吸バーストが起こると、PIP3 は NADPH オキシダー
ゼを構成しているユニットの1つ p47phox に結合して NADPH オキシダーゼを活性化しま
す 41,43)。PIP3 は NADPH オキシダーゼによって生成された NADPH が細胞膜の運動や細
胞骨格の形成に関与する線維であるアクチン 105)や細胞の運動 106,107)にも関与します。アク
チンは好中球やマクロファージなど食細胞による異物の取り込み時の細胞膜の動きにも関
係しています 108,109)。PI3K の活性化も MTT 法による細胞毒性を示します 41)。
(ストレス耐性)
獲得免疫系がない原核生物でも(10 ページ注*****)、以前と同じ事態に遭遇した時には、
以前よりうまく対処できる仕組みを備えています。それは、一度経験したことのあるストレ
スによって次のストレスに対する耐性が備わる仕組みで、この仕組みをストレス耐性と言
います。このストレス耐性は生物界に幅広く行き渡っている防御メカニズムで
66)、物質を
記憶する獲得免疫とは異なり、経験した環境や状況に対する馴れとも言い換えることがで
きます。
よくストレス耐性の例として引き合いに出されるのは虚血耐性です。脳のニューロンは
虚血に脆弱ですが(28 ページ「虚血ストレス応答」)、一度、弱い虚血状態を経験したニューロ
ンは、次に遭遇した虚血が強くても容易に死ぬことはありません。実際、過去に 1 回だけ
脳梗塞を起こしたことがある患者は、初めて脳梗塞を起こした患者より梗塞予後が良好で
32
ANM176®が認知症の予防と改善に役立つ可能性について
あったと報告されています 110)。
ストレス耐性は虚血耐性だけでなく、熱中症 111)、低体温 112)、薬物 113)、物理的障害に
対する耐性が知られています。虚血耐性は低酸素耐性の 1 種です。ヒマラヤなど高山に登
る時に、中間地点にしばらく滞在して体を慣らすのも、マラソン選手が高地でトレーニン
グするのも、低酸素耐性を強化するためです。これらのストレス耐性には同じようなメカ
ニズムが関与しているのではないかと考えられています 114)。
中枢神経系においては、ニューロンは虚血(低酸素)に脆弱ですが、一方、アストロサ
イトは耐性的です(29 ページ)。この虚血に対するアストロサイトの応答が小胞体(ER)の
機能と関係していると考えられています 115) 。ラットの神経系細胞を低酸素に曝すと、小
胞体ストレス応答(UPR)関連の分子(25 ページ「小胞体ストレス応答」)が観察されるように
なります 116)。これらのタンパク質の中には、UPR 関連の ATF6 に類似した OASIS、
BBF2H7、ATF4 などがあります 117)。OASIS はアストロサイトで、BBF2H7 はニューロ
ンで観察されます。アストロサイトでは、まず ATF6 が関係した UPR が起こり(初期応
答)
、その後 OASIS による 2 段階目のサバイバルシグナルが活性化されると報告されてい
ます(後期応答)118)。この OASIS によるシグナルが、アストロサイトの小胞体ストレス
耐性を生み出す根源であると考えられています 118)。UPR で転写される遺伝子には小胞体
ストレス応答エレメント(ER-stress response element;ERSE)あるいは cAMP 応答エ
レメント(cAMP response element;CRE)の転写エレメント(19 ページ注)あります
58,119)。ATF6
は主に ERSE に作用するのに対して、OASIS は CRE に優勢的に作用する
とのことです 118)。
転写因子 Nrf2 の活性化は神経系細胞の中ではアストロサイトにだけ観察されますが
120)、Nrf2
に欠陥がある変異マウスの神経細胞は、酸化ストレスに対して過敏であったと
報告されています 121)。Nrf2 は酸化ストレス、小胞体ストレスなどによる親電子性物質、
活性酸素、血流によるずり応力など物理的ストレスなど様々な環境ストレスによって細胞
質から核内に移行して転写が活性され、抗酸化や解毒酵素群の遺伝子発現を誘導し
69,122)、ストレス応答を統一的に制御します。また、Nrf2
66,123)。Nrf2
は虚血耐性にも関係します
はストレス耐性とも関係があるといわれています 124)。Nrf2 が関与する様々
なタンパク質の転写によってストレスに対する防御が強化され 124)、致死的ストレスに対
しても防御的に働きます 125)。
ラットの培養アストロサイトを低酸素に曝すと、ヘム(図 18)を分解するヘムオキシゲ
ナーゼ1(以下では Heme oxygenase-1 から HO-1 と表記します)*が観察されます 116)。
UPR が起こると、ATF4 は Nrf2 の転写活性化に関与して HO-1 遺伝子の転写を開始させ
ます 126)。虚血状態でアストロサイトに現われる HO-1 が、虚血耐性に寄与するとも考え
られています 127,128)。
* ヘムを分解する HO-1 は NADPH および酸素を使ってヘムを段階的に分解し、線形テトラピロール
ビリベルジンとビリルビン(図 24)に変換されます 127)。ビリベルジンとビリルビンには抗酸化活
性と一酸化炭素に対する細胞防御作用があります。
33
ANM176®が認知症の予防と改善に役立つ可能性について
一方、特定のストレス耐性獲得時に別のストレ
A
ス耐性が生じることがあり、この現象をクロス・
トレランスと言います。クロス・トレランスの例
として、吸入麻酔薬による脳の虚血耐性 131)、低
体温治療による脳保護作用 72,112)など多数が知ら
れ、医療でも多方面に応用されています。カロリ
ー制限による外科的ストレスや熱ストレスの耐性
増強 132,133)あるいは鍼灸治療もクロス・トレラン
スの例です。また、植物の毒性成分のアルカロイ
B
ドが含まれているトリカブトなどの漢方生薬を治
療に用いるのも、毒を以て毒を制するの諺(コトワ
ザ)にあるように、ストレス耐性の作用によるも
のと考えられます。
クロス・トレランスは様々なストレスの応答や
耐性が繋がっていることによって生じると考えら
図 24.線形テトラピロールビリベルジン
(A)とピルビン(B)の構造
WIKIPEDIA より
れます。高レベルの活性酸素種(reactive oxygen species;ROS)は 細胞に有害で加齢や
老年性関連の様々な病気と関係すると言われています 134,135)。一方、通常レベルの活性酸素、
スーパーオキシド、過酸化水素などの電子供与体は、外来性および内因性の様々な物質を酸
化するためや細胞膜の運動などに必要不可欠で 136,137)、傷害に直接的に関与していないと言
われています 138)。また、細胞内のスーパーオキシドの濃度が上昇するほど細胞自己死のア
ポトーシス(16 ページ注**)を抑制することができ
139)、あるケースでは無毒の範囲内で活
性酸素に晒すと細胞死を促進するよりむしろ生長を刺激する効果を示す証拠がいくつも提
出されています
140,141)。これら細胞の生長に寄与する理由の1つには、ROS
を産生する環
境は酸化ストレスを引き起こして酸化ストレス耐性を誘起することが背景にあること考え
られます。毒性学では少量のストレスが細胞の防御機構を誘導し引き続く重篤なストレス
に耐性が備わる現象を“ホルミシス(hormesis)”と呼んでいますが、転写因子 Nrf2 は酸
化ストレスに対するホルミシスを仲介し、様々な疾患の予防・治療に有効であると考えられ
ています 66)。
(細胞ストレスと病気)
シグナリングに関与するキナーゼは、ヒトの DNA 中に 518 種が書き込まれていると言
われています 142)。このキナーゼによるシグナリングに更に他のタンパク質がアロステリッ
ク制御(15 ページ)に関与して微妙に調整され、様々な受容体や転写因子などの制御因子を刺
激します。このように膨大なタンパク質が複雑に関係して微妙に調整されている現象を解
明することは容易ではありません。AD と同じように多くの病気でも原因物質による発症の
方法が解明されていないのは、生命現象が複雑なことも関係しています。
34
ANM176®が認知症の予防と改善に役立つ可能性について
しかし、38 億年にわたって進化した微妙な制御方法には、病気にならない免疫の仕組み
も備わっています(24 ページ「細胞ストレスについて」)。それでも成人病や認知症あるいは様々
なアレルギー疾患が急増しているのは、今までに生物が経験しなかった高齢化や環境変化
などの事態に対応できないからでしょう。この事態には、食事や生活環境の変化もあるのか
も知れません。
小胞体(ER)ストレスは幾つかの病気と関係すると言われています。例えば、ER ストレ
スは肥満症を誘導し、インスリンが十分に機能しないインスリン抵抗性を生じさせます 143)
(詳細は 71 ページ「インスリン抵抗性」)。インスリン抵抗性は 2 型糖尿病を発症させます
144)。
さらに肝臓における ER ストレスは炎症を引き起こし 117)、脂肪代謝、グルコース、イオン
のホメオスタシス(恒常性維持)
(22 ページ「生物の恒常性維持について」)に影響し、幾つかの病
理につながるストレス応答を引き起こすと言われています 145)。
セリン・スレオニンキナーゼであるアポトーシスシグナリングキナーゼ(以下では
Apotosis signaling kinase から ASK と表記します)の活性化によって細胞の自己死であ
るアポトーシス(16 ページ注**)を起こすことが知られています。平常 ASK は活性化され
ずに静かにしていますが、様々な環境ストレス、活性酸素種(ROS)、小胞体(ER)スト
レス、炎症に関連する IL-1βや亜酸化窒素(N2O)など内因性因子 146)に応答して活性化
し、ストレスキナーゼ SAPK 経路(16 ページ)を活性化します 147,148)。ASK はガン、糖尿
病、血管性疾患に関連し 149)、アルツハイマー病などの神経変性疾患にも関係しているこ
とが示唆されています 146,150)。各種神経変性疾患では小胞体ストレス応答(UPR)の防御
機構(25 ページ「小胞体ストレス応答」)が破綻して変性タンパク質の蓄積を招き、アポトーシ
スによるニューロン死に繋がることが示されています 151,152)。
(神経の中の記憶)
脳内で神経作用に関与するニューロンのシナプスから分泌される神経伝達物質は 100 種
類以上あり、その中でアセチルコリン(以下では acetyl choline から ACh と表記します)
は記憶や学習などに関係し、セロトニンは眠りや脳を鎮めた時に分泌されます。また、これ
らの神経伝達物質は、どのニューロンにも存在するのではなく、ACh を主に分泌するコリ
ン作動性ニューロンやノルアドレナリンを主に分泌するアドレナリン作動性ニューロンな
ど、ニューロンは特定の神経伝達物質を分泌するように特化しています。ですから、シナプ
スの結合によって形成されるニューロンネットワーク(8 ページ)には、特定な神経伝達物質
によって作動する様々な種類のニューロンが複雑に関係しています。
ヒトは、生まれた時は約 1,000 億個のニューロンがあるが、成人になると1日に 10 万個
単位のニューロンが死んで行くと言われています。それでも記憶が失われないのは、記憶は、
新しいニューロンが生まれて生じるのではなく、ニューロンネットワーク内に新たな結合
が形成されたり、今まである結合が強化されたり 153)、あるいは様々な要因によるニューロ
ンの信号伝達能力が向上したりすることによって、記憶が生じると考えられているからで
35
ANM176®が認知症の予防と改善に役立つ可能性について
す 154)。
本能も記憶の一種で、ニュ
a
ーロンネットワークに刻ま
れて生まれてきます。よく、
幼少期を外国で過ごした子
供は、ほぼネイティブスピー
カーと同様に外国語の発音
や聞き取りができるように
なりますが、大人になってか
ら外国に数年滞在しても、ネ
イティブスピーカーに近づ
c
b
くことは難しいことが知ら
れています。このように脳に
は最も適応できる時期があ
り、その時期までは、ニュー
ロンネットワークは骨格だ
けで、その後の刺激によって
細部ができあがるのではな
図 25.ラット海馬(a)およびテタヌス刺激(b)と LTP(c)
いかと考えられています。赤
(a) ラット海馬のシェファー側枝 (Schaffer collateral;SC)、苔状線維
(mossy fiber;MF) 、中隔海馬経路(septohippocampal pathway;
SHP)
(b) ニューロン表面の電位が長い時間にわたり上昇する長期増強
(LTP)の現象(c)は、電気刺激を貫通線維、苔状線維、シャッ
ファー側枝などのニューロンに連続して与えることで誘導されます
827)。
a 図は Schultz S, Panzeri S, Rolls E, Treves A [Quantitative analysis of
a Schaffer collateral model] In Information Theory and the Brain,
ちゃんは、よく両手を顔の前
で大きく揺すりますが、これ
も人によって異なる両眼の
焦点を調節するためで
155)、
幼児期に両手が自由に揺す
られない状況が長く続くと
斜視の原因になると言われ
R.Baddeley, P. Hancock, P. Foldiak eds., Cambridge University Press,
UK (1998)を改変
b,c 図 は The Synaptic Organization of the Brain, Ed: Gordon M.
Shepherd, Oxford University Press (2003)から改変
ています 156)。
記憶や学習において、両耳の奥にある海馬(図 2)が重要な働きをしており、一時的に覚
える短期記憶(4 ページ)は、刺激によって一時的に海馬のニューロンが活発になる現象と考
えられています 154)。実験的に前後のニューロンを刺激した時にニューロンの表面電位が長
い時間上昇する長期増強(以下では Long Term Potentiation の頭文字から LTP と表記し
ます)という現象(図 25)が確かめられ 157)。短期記憶は、この LTP に保持されると考え
られています 158,159)。
ニューロンのシナプス(図 3)はシナプスで分泌される主な神経伝達物質による違いだけ
でなく、興奮の仕方によっても種類が異なり、後シナプスの膜電位を上昇させるシナプスは
興奮性シナプスと呼び、上昇した電位を抑制するシナプスは抑制性シナプスと呼びます。興
36
ANM176®が認知症の予防と改善に役立つ可能性について
奮性シナプスが記憶の形成に関係します。興奮性シナプスは信号を受け取るとシナプス後
膜(図 3)で興奮性シナプス後電位(以下では excitatory postsynaptic potential から EPSP
と表記します)と呼ばれる活動電位を発し、EPSP によって後シナプスの「脱分極」と呼ば
れる電位上昇が起こります。中枢神経系ではグルタミン酸が主な興奮性シナプスの神経伝
達物質です。
LTP によって樹状突起の先端部にスパインと呼ばれる微小な突起が現れ、スパイン上に
形成されたシナプスが次々とつながってニューロンネットワークが形成されます。このニ
ューロンネットワークに長期記憶が組み込まれると考えられています 160,161)。実際に、ニュ
ーロンネットワークの中に地理記憶などがあることが、21 世紀になって実証されました
162,163)。2014
年にノーベル生理学・医学賞を受賞した研究は、一定の場所を走り抜ける時に
海馬で活発に活動するニューロンの発見でした
164,165)。同じように、特定の景色を見た時
166,167)や特定の場所に来た時 168)あるいは特定の時 169)に活動するニューロンが次々と報告
されてきています 170)。
このようにニューロンネットワークにある特定のニューロンの集団が突然活発になる現
象を発火(フラッシュ:flash)と言います。思い出す時には、スキャニング中に特定の記憶
に当てはまるニューロンネットワークで電位の上昇が起こり、フラッシュのようにパッと
思い当ります。英語でも突然思い出すことをフラッシュ・バック(flash back)といいます
が、我々も思い出す時に、
「あ、これだ」と突然に思い出します。記憶に関わった海馬のニ
ューロンが優先的に活動しやすくなることで記憶が再生されると考えられています 171)。
(記憶の分子機構)
記憶は長期増強(LTP)から始まり
AMPA 型
グルタミン酸
受容体
ます。短期記憶は、LTP によって上昇
するニューロンの表面電位である興
奮性シナプス後電位(EPSP)によっ
て担われています。EPSP を引き起こ
す主な神経伝達物質グルタミン酸(以
下では Glutamate から Glu と表記し
ます)の受容体にはイオンチャンネル
共役型と代謝調節型があります。イオ
ンチャンネル共役型受容体には
AMPA 型、NMDA 型、カイニン酸型
があり、これらの受容体はニューロン
の膜を通過するイオンの種類と量を
調節します(図 26)。
EPSP の分子機構は大よそ以下の
図 26.LTP の分子機構
シナプスには、通常のシナプス伝達を主に担う AMPA 型受容
体と、特に細胞が脱分極したときに活性化する NMDA 型受容
体があります。NMDA 受容体は Ca2+を透過させますが、平常
では Mg イオンがブロックしているため活性化していません。
高頻度 刺激 等によ りシ ナプス 後細 胞が 強く脱 分極 す ると、
NMDA 受容体の Mg イオンのブロックがはずれて Ca2+が細胞
内に流入します。
Ca2+はカルモジュリン(Calmodulin)を活性化することによ
って、キナーゼの CAMKII を活性化し、これが AMPA 受容体
をリン酸化すると、シナプス後電流の振幅が増大します 828,829)。
また、CAMKII は核への情報伝達系を活性化し、新規タンパク
質合成を誘導します。830)を改変
37
ANM176®が認知症の予防と改善に役立つ可能性について
通りです。シナプス前膜(図 3)からシナプス間隙に放出された Glu が次のニューロンのシ
ナプス後膜に埋め込まれた AMPA 型受容体に結合するとニューロン内にナトリウム(Na)
イオンが流れ込み、シナプス電位が上昇します。このシナプス電位の上昇が一定のレベルに
達すると、NMDA 型受容体をブロックしていたマグネシウム(Mg)イオンが外れてカルシ
ウムイオン(Ca2+)がニューロン内に流入します
172)。後シナプスにおいて
NMAD 受容体
を通して流入した Ca2+により活性化したカルシウム/カルモジュリン依存性プロテインキナ
ーゼⅡ(calcium-calmodulin- dependent protein kinaseⅡ;CaMKⅡ)によって細胞質か
らシナプス後膜に AMPA 受容体が挿入され AMPA 受容体が増強されます。同時に NMAD
受容体の自己リン酸化が促進され、LTP に必要なキナーゼ活性が持続して 172)、シナプス結
合が強化・安定化します 173)(図 26)。
シ ナ プス 後膜 には 後
後シナプス肥厚部(PSD)
シナプス肥厚部(以下で
は postsynaptic density
から PSD と表記します)
と呼ばれる膜が厚くな
った複雑な構造が発達
しています 174)(図 27)
。
樹 状 突 起 の PSD に は
PSD-95 ( postsynaptic
density-95)などの足場
タンパク質*があり、グ
ルタミン酸受容体の
図 27.マッシュルーム型スパインの構造
スパインの構造において、頭部には PSD が顕著に見られ、頭部と頸部には繊
維状アクチンの細胞骨格が発達しています。 191)
NMDA 型や AMPA 型など多くのタンパク質を繋げています 175,176)。
*足場タンパク質とは、タンパク質複合体形成の足場となるタンパク質のことで、PDZ や SH3**など
タンパク質同士の結合に関わる複数のドメインで構成され、シグナル伝達に関わる分子を結びつける
機能があます。
通常、タンパク質は一定の構造をしており、その構造には、数個から十数個のアミノ酸からなる
モチーフと呼ばれる二次構造と、さらに二次構造が集まり局所的にコンパクトな半独立のユニット
を作ったドメインと呼ばれる三次元構造が含まれます 177)。ドメインは、タンパク質同士の相互作用
を行います。
特にシナプスには受容体、接着分子、シグナル分子など多様なタンパク質が PDZ ドメ
イン**を足場として、シナプス構造や適切なシグナル伝達に重要な役割を果たしています
178)。シナプス後膜にある
PSD-95 は 3 つの PDZ と SH3***などのドメインから成る興奮
性シナプスの主要な足場タンパク質の一つで、グルタミン酸受容体の NMDA 型や AMPA
型 179,180)など多くのタンパク質の足場となり 175,176)、膜タンパク質と細胞内シグナル伝達
系を繋げる役割を担っていると考えられています 181)。
**タンパク質の中にある PDZ ドメインは種を超えて幅広く豊富に存在します 182)。PDZ ドメインの
多重性により、様々な機能性タンパク質が一ヵ所に集積され、それらの相互作用によってシグナリン
グの調節が行われると考えられています。PDZ ドメインはターゲットタンパク質のβシート構造
38
ANM176®が認知症の予防と改善に役立つ可能性について
図 28.TARP の AMPA 受容体補助サブユニットとしての機能
(A)TARP は AMPA 受容体のチャネル活性を修飾しシグナル強度を増強します。(B)TARP は AMPA 受容体の
発現を安定化し細胞表面の AMPA 受容体量を増大します。
(C)TARP と PSD-95 の結合は AMPA 受容体複合体を
シナプスに係留しシナプスの AMPA 受容体活性を維持します。187)
183,184)を結合サイトとして認識します。
***SH3 ドメインは、アミノ酸のプロリンが豊富にある領域をもち、様々なターゲットタンパク質
と幅広く結合してリン酸化に関与します。
TARP と呼ばれるタンパク質は AMPA 受容体に結合してチャンネル機能(図 4)を変化
させる AMPA 受容体補助サブユニットです 185)(図 28)。TARP の C 末端部位は PDZ ドメ
インによって PSD
(図 27)にある PSD-95 と結合し、この TARP と PSD-95 の結合が AMPA
受容体をシナプスにつなぎ留め、シナプスの AMPA 受容体活性を維持します。こうして
AMPA 受容体の数が増加し、安定化して 186)、シグナル強度が増強します 187)。この PDZ ド
メインによるタンパク質の相互作用が LTP(36 ページ)の原理で、短期記憶形成に機能して
188,189)、短期記憶が形成されると考えられています 158,159)。
では、長い間保持される長期記憶はどのようにして形成されるのでしょうか?長期記憶は
シナプスで繋がったニューロンネットワーク(8 ページ)に新たなシナプスが形成されること
によって形成されると考えられます。この新たなシナプス形成の方法には諸説があります
が、最も有力と考えられているのは、最初に運動性の高い細長い突起が樹状突起に現れ、こ
れに軸索が接着してスパイン(37 ページおよび図 27)ができ、そのスパイン上にシナプスが形
成されるという説です 190,191)。
長期記憶に関与するタンパク質に線維を構成するアクチンなど細胞膜の形成に関与する
タンパク質があります 159,192)。LTP に伴ってシナプス内のアクチンが大きく変化します
193,194)。LTP
によって PSD-95 の PDZ ドメインに A MPA 型と NMDA 型の Glu 受容体あ
るいは TARP など多くのタンパク質が結合してシナプスへ挿入され、この挿入に伴い後シ
ナプスにあるアクチンの重合が亢進し、その結果、樹状突起のスパインが拡大して後シナ
プスタンパク質の結合量が亢進すると考えられています 195)。アクチンは、「G アクチン」
と呼ばれる球状タンパク質が連なってできた線維状タンパク質で微小線維とも呼ばれます
39
ANM176®が認知症の予防と改善に役立つ可能性について
(図 29)。筋肉細胞が収縮するのはアク
チンの働きです。細胞中にあるアクチン
は、マクロファージなど食細胞がアメー
バ―のように細胞外の異物を取り込む時
の細胞膜の運動も関係します 108,109)。ニ
ューロン軸索や樹状突起の伸長にもアク
チンが働いています。
このアクチンの制御には G タンパク質
の Rho の仲間分子(ファミリー)が関与
し 196)、ニューロン突起の形態形成や制御
に不可欠な役割を果たしています 197,198)。
G タンパク質は、その働き方から Ras、
Rab、Rho ファミリーに分けられていま
す。Ras ファミリーは、主に細胞の増殖や
分化に関与し、ストレスに応答して炎症
図 29.アクチン(微小繊維)の構造
(A)アクチンの電子顕微鏡写真 (B) アクチンを構成する
G アクチン分子は 2 量体となって ATP と結合し、螺旋状
に連なって、線維になります。(C) G アクチンを色分けし
て隣同士密着を示しています。
などの様々な生理現象と関係します。Rab
ファミリーは、主に膜の形成と関係があり、小胞体機能やエンドサイトーシスとよばれる細
胞外物質の細胞内への取り込み、神経伝達物質などの分泌などに関与しています。Rho フ
ァミリーは主に細胞膜の動きと関係し、食細胞による異物の取り込みやニューロンの突起
の形成や制御に関係しています。
細胞内シグナリングのスイッチのような役割を果たしている G タンパク質は(14 ページ)、
ニューロンの受容体にも多数あり、例えば、代謝調節型受容体のムスカリン性アセチルコリ
ン受容体(muscarinic acetylcholine receptor;mAChR)、アドレナリン受容体、ドーパミ
ン受容体、γアミノ酪酸(γ-aminobutyric acid;GABA)受容体 B 型、セロトニン受容体、
嗅覚受容体、ロドプシン(網膜にある光受容体)、アヘン成分と結合するカンナビノイド受
容体とオピオイド受容体など数多くの受容体は G タンパク質と連携して働く G タンパク質
共役受容体(GPCR)です(15 ページ)。これらの GPCR は長期記憶の形成に関係するニュー
ロンの軸索および樹状突起の形成や伸展などシナプス形成過程において、細胞の構造を支
えるアクチンと微小管(以下では microtubules から MT と表記します)による細胞骨格の
動的役割に不可欠です 199)。
シナプス形成の過程が終了すると MT が軸索に挿入され、その後、持続的な軸索伸展と
強化が起こると考えられています 197)。MT はアクチンより 3 倍ほど太いチューブ状の線維
でαチュービュリンとβチュービュリンと呼ばれる球状タンパク質が中空の周りを交互に
結合しています(図 30)。MT は骨格形成の役割だけでなく、小胞体輸送と呼ばれる分泌タ
ンパク質の細胞質内輸送にも関与しています。ニューロンの小胞体(ER)で合成された神
経伝達物質は小胞と呼ばれる粒子内に包まれ、MT のレールに沿ってシナプスまで送り込ま
40
ANM176®が認知症の予防と改善に役立つ可能性について
れ、シナプス間隙に放出されます。この
輸送は、種々の微小管結合タンパク質
( 以 下 で は microtubule-associated
protein から MAP と表記します)によ
って調節されています。
微小管結合タンパク質 1(以下では
microtubule associated protein 1 か
ら MAP1 と表記します)とコラプシン
反応媒介タンパク質 2(以下では
Collapsin response mediating protein
2 から CRMP2 と表記します)は
MAP の仲間です。MAP1 には
MAP1A、MAP1B 、MAP1S のサブタ
イプがあり、MAP1A と MAP1B は主
にニューロン、MAP1S は多くの組織
で見られます。MAP1B は発達期のニ
ューロンの特に軸索に見られ、軸索の
形成を誘導し維持します 200,201)で。
MAP1A は成熟した脳のニューロンの
図 30.微小管の構造
α:αチューブリン β:βチューブリン
微小管は、互いによく似た球状タンパク質αチューブリンと
βチューブリンが交互に会合し一回り 13 個のチューブリン
分子からなる中空の繊維です。
東京医科歯科大学教養部生物和田勝 HP より
樹状突起にあります。MAP1B はチュ
ーブリンの重合・脱重合による MT
の形成と維持に関与し、神経細胞の
形態形成や機能を制御しています
202)。
MAP1 と異なり CRMP2 にはキナ
ーゼ活性があり、MT の構成分子であ
るチューブリンと結合して MT の重
合を促進して安定化させます。この
MT の重合がニューロンの極性を決定
し軸索伸長に必要であることが明ら
かになっています 203)。さらに
CRMP2 は、軸索形成に必要なタンパ
ク質の輸送や接着分子のリサイクリ
ングにも関与しています 204)。G タン
パク質 Rho ファミリーによるシグナ
図 31.CRMP2 を介したシグナル伝達機構
久保祐亮、稲垣直之 web site「脳科学辞典-CRMP」
リングによって CRMP2 がリン酸化
41
ANM176®が認知症の予防と改善に役立つ可能性について
します 201,205)(図 31)。リン酸化された CRMP2 はチューブリンへの結合性が弱くなって
軸索の退縮が促進されます
205)。
短期記憶は既に細胞中にあ
るタンパク質の活性化によっ
て保持されるのに対して、長期
記憶には新たなタンパク質の
合成が必要です。外部からの刺
激によって活性化した G タン
パク質によるシグナリングは、
シナプスの形成や熟成に関係
して長期記憶が保持されます。
一般に細胞の成長を制御す
る G タンパク質 Ras ファミリ
ーはアポトーシスを引き起こ
図 32.Ras 経路(東北大学大学院医学系研究科遺伝病学分野 HP)
図 33.SRF 細胞内制御機構
転写因子 SRF は DNA の CArG ボックス(CC(A/T)6GG 配列)に 2 量体で結合し、c-fos などの最初期遺伝子
(immediate early gene;IEG)やβアクチンなど細胞骨格系遺伝子の発現を制御します 207)。
転写因子には Ternary complex factor(TCF)831)など様々なコファクター(転写共役(調節)因子)が DNA 転写
の制御に関与しています。TCF は、MAPK によりリン酸化されて活性調節され 831)、CArG ボックス近傍の DNA
配列(GGAA/T)と SRF に結合して 3 量体を形成し、下流遺伝子の発現を制御します 831)。MKL/MRTF も SRF
のコファクターで 832)、MKL/MRTF は Rho シグナル活性化によるアクチン細胞骨格の再編成によって G-アクチ
ンから解離し、核移行して下流遺伝子の発現を制御すると考えられています 833)。
田渕 明子 [血清
応答因子] 脳科学辞典 2012 年 5 月 1 日 より
42
ANM176®が認知症の予防と改善に役立つ可能性について
す Akt やストレス応答に関連する ERK などの MAPK 経路、あるいは様々な応答に関連
しますが(図 32)、転写因子の血清応答因子(serum response factor;SRF)*の活性化
にも関与しています。
*転写因子 SRF は 206)、c-fos などの最初期遺伝子(immediate early gene;IEG)やβアクチンな
ど細胞骨格系遺伝子の発現を制御し 207)、中枢神経系においては、海馬の神経回路形成 208)、樹状
突起や軸索形態 208,209)、シナプス機能 207,209)などに関与しています。
SRF 制御のための細胞内情報伝達は、MAPK 経路と低分子量 G タンパク質 Rho 伝達経
路の大きく 2 つがよく知られています 210)(図 33)。
アルツハイマー病
アルツハイマー病(AD)は高齢になって発症す
る代表的な認知症です。
AD はドイツの精神科医アロイス・アルツハイマ
ー(Alois Alzheimer)によって 1906 年に初めて報
告された認知症で、その特徴は以下のようです。
(1) 成人期を過ぎてから発病します。
(2) 記憶力の低下や異常行動が見られ、これらは
段々と悪化します。
(3) 死後解剖の脳で以下のことが確認されます。
① 脳神経細胞の死滅による大脳の萎縮(図
34)。死後解剖で通常の脳の重さは 1200~
図 34.正常者と AD 病患者の脳 213)
1400g ですが、AD では 1000g 以
下となっています。
②顕微鏡観察で以下のものが数多く
観察されます。
ⅰ) 脳組織に老人斑と呼ばれる斑点
老人斑には主にβアミロイド
タンパク質が含まれており
211,212)、このβアミロイドは電
子顕微鏡で観察すると線維状
になっています 213)(図 35)。
ⅱ) 中枢神経系のニューロン内に
図 35.老人斑内βアミロイドの電子顕微鏡写真 213)
神経原線維変化(以下では neurofibrillary tangle から NFT と表記します)と呼
ばれる線維状の構造物があり
(図 36)、NFT 中には主にリン酸化されたタウ
(tau)
と呼ばれるタンパク質がよく見られます
214,215) 。タウは微小管結合タンパク質
43
ANM176®が認知症の予防と改善に役立つ可能性について
(MAP)(41 ページ)の 1 種で、
タウのリン酸化や脱リン酸化
は微小管(MT)による細胞骨
格形成や細胞内の物質輸送に
関係します 216,217)。
AD になると記憶力が著しく低下する
だけでなく人格が失われたような言動が
見られます。昨日の出来事が全くなかっ
たかのように言ったり行動したり、作り
事を言ったり、いままでマメだった買い
図 36.神経原線維変化(NFT)の顕微鏡写真 213)
物をおっくうがったり、季節外れの着物を着たり等の明らかにおかしい言動が、症状が進む
にしたがって段々と目立つようになります。また、引きこもったり、家族と話したがらなく
なったり、趣味だったことをしなくなったり、物事に関心を示さなくなったり等の鬱が見ら
れるようになります。初期では昔のことは良く覚えています。また、無関心や意欲の低下が
見られ、ささいなことで感情を爆発させたりします。症状が進むと、あたかもあったことの
ように言う妄想、死んだ人がいるかのような幻覚、話し声が聞こえるような幻聴、徘徊ある
いはしてはいけない場所での排泄をする等の異常な事態がしばしば起こるようになります。
AD を発病する年齢は、ほとんどは 65 歳以上になってからで、このことから老年性アル
ツハイマー型認知症(SDAT)とも呼ばれます。まれに 50 代からでも発病することがあり、
これを若年の早期発症型アルツハイマー病(以下では early-onset AD から EOAD と表記し
ます)と呼ばれます。EOAD 患者数は全 AD 患者数の 5-15%を占める 218)と言われていま
す。EOAD では、患者の親族に AD 発病の頻度が高くみられることから、家族性アルツハ
イマー病(以下では familial Alzheimer's disease から FAD と表記します)とも呼ばれま
す。SDAT と FAD は発症時期と進行速度に違いがあるだけで症状は同じです。そのため、
以下では断りがない限り、SDAT と FAD を区分けせず AD と言います。AD 診断後の余命
は 6.1 年(血管性認知症の場合は 5.5 年)と報告されています 219)。診断を受けるまでの期
間を入れると、個々人による差はありますが、AD は発病から約 10 年で進行すると考えら
れます。
AD になぜなるのか?の問いには、なかなか答えられないのが現状のようです。というの
は AD の原因タンパク質と言われているβアミロイドが、どのようにして AD 発症に関与
しているのか、まだ、定説がないからです。
βアミロイドは以下の理由から AD 発症の原因タンパク質と考えられています。
① AD の死後脳の組織中
220–222)、血清中 223,224)に見られるβアミロイド濃度は、健常者
に比較して明らかに高いこと。さらに、AD モデルマウスの脳内でもβアミロイドの増
加が見られること 225)。
② AD の脳内に特徴的に見られる老人斑中に、主にβアミロイドが含まれること 211,212)。
44
ANM176®が認知症の予防と改善に役立つ可能性について
③ FAD 患者で見つかっている変異した遺伝子は共通して、βアミロイドの分泌と関係し
ていること 226,227)。また、ダウン症候群(Down's syndrome)はβアミロイドの遺伝子
がある 21 番常染色体(18 ページ)が 3 本になる遺伝病で(性染色体以外の常染色体は常
に 2 本が一組です)
、AD 患者と同じように、血清でβアミロイドの濃度が上昇し 228)、
成人期以降に認知症を発症すること 229–231)。
④ βアミロイドはニューロンの LTP(36 ページ)を抑制し
232)、βアミロイドが多量に見
られるニューロンでシナプス機能の不全や変性が観察され 233)、また動物試験で記憶や
学習力を低下させる 234)など記憶に関連する様々な機能を障害するβアミロイドの作用
が示されていること。さらに、βアミロイドが大量に見られる AD モデルマウスのシナ
プスが正常に機能しないこと 235)。
AD 患者の脳にはβアミロイドを含む老人斑だけでなく、リン酸化されたタウを含む NFT
(図 36)が見られるため、βアミロイドとタウのどちらが AD の原因タンパク質なのか、
長い間、論議されてきました。また、AD 患者の脳髄液(CSF)中で、βアミロイド濃度が
低下するが 236)、タウの濃度は増加するため 237)、一時は、AD の原因タンパク質はβアミロ
イドでなくタウではないかと言われました 238)。
しかし、βアミロイドはタウの過リン酸化とニューロンの機能不全や変性を導き
239,240)、
さらに、タウのリン酸化の前にβアミロイドによってタウの異常と微小管の崩壊が観察さ
れています 241)。また現在では、AD 患者の CSF 中のタウ濃度が増加する理由は、AD 患者
の脳関門(BBB)に関与する受容体の 1 つである LRP1(11 ページ)が減少し 242,243)、また、
BBB の能力も低下し
られています
244,245)、βアミロイドの脳外への排出量が減少することによると考え
28,244,245)。これらのことから、AD
の CSF 中のβアミロイド濃度が低下する
理由も AD の発症によって LRP1 の能力が低下することに原因があることが分かり、現在
では、βアミロイドが AD 発病の原因ということが確定的です。しかし、βアミロイドと
AD との関係について、例えば、以下の問題点に十分に答えらません。
ⓐ βアミロイドは、どのようにして AD におけるに認知機能の障害に関与しているのか?
ⓑ 健常者であっても壮年期になると脳内や CSF 中のβアミロイドが増加し 246,247)、また、
老人斑は多くてあっても AD でない人が多数いる 214,248)。
ⓒ 遺伝とは関係しない SDAT の患者で老人斑が多く見られない症例がある 249,250)。
ⓓ 老人斑があまり見られない FAD が見つかっている 251,252)。
ⓔ AD モデルマウスではβアミロイドの沈着が現れる何ヵ月も前から脳の機能障害が現れ
る 225)などβアミロイドの存在と AD が必ずしも相関していない。
以上のように、AD とβアミロイドの存在が相関しない場合があるため、βアミロイドが
AD の原因タンパク質だとしても、AD 発症の機構がうまく説明できないのです。しかし、
最近の多くの研究成果から、βアミロイドによる AD 発症の機序が推察されています。以降
では、その内容を述べます。
45
ANM176®が認知症の予防と改善に役立つ可能性について
(βアミロイドの分泌と APP の役割)
βアミロイドはアミロイド前駆タンパク
質(以下では amyloid precursor protein か
ら APP と表記します)と呼ばれるタンパク
APP 分子
細胞膜
質から、幾つかの酵素によって切り出されま
す 253,254)。APP は1回だけ細胞膜を貫通した
タンパク質*で、βアミロイドは APP の細胞
細胞内
膜の外側から数 10 分子離れた場所と細胞膜
貫通部の中が切られた部分です(図 37,38)。
*細胞膜を貫通するタンパク質には、1回膜を
貫通するものや、4、7 回など複数回貫通する
ものがあります。
βアミロイド
APP はどんな動物にも共通してみられ、し
かも種による変化が少ないタンパク質です。
通常タンパク質は、進化によって種による違
酵素
いがあります。種による変化が少ないという
ことは、種に共通した重要な機能があること
老人斑
を示唆しています。また APP は、様々な種
類の細胞に幅広く見られ、脳では、特にニュ
ー ロン のシ ナプ スに 高 濃度 で存 在 します
255,256)。
21 番常染色体にある APP の遺伝子が
DNA から読み取られる時に、DNA の一部が
読まれない選択的スプライシング(23 ページ
注)で、長さが異なる 3 種
図 37.β-アミロイドと老人斑の生成
(Alzheimer Disease Education & Referral Center)
類の APP ができあがりま
す。最長の APP は 770 個
のアミノ酸からできてお
り、APP 770 と呼びます。
APP 770 から 19 個のアミ
ノ酸が抜け落ちて APP751
が読み取られ、更に 56 個
のアミノ酸が抜けた
APP695 が読み取られます
257)(図
38)。 ニューロン
に は APP695 、 APP751 、
APP770 の全てが見られま
図 38.アミロイド前駆タンパク質(APP)の切断
アミノ酸数が 770 個からなる APP770、24 個からなる P3 と 42 個からなる
βアミロイドを示した。
46
ANM176®が認知症の予防と改善に役立つ可能性について
すが 255,258)、ニューロンを補助するアストロサイトやオリゴデンドロサイトのグリア細胞(7
ページ)では APP695 は見られません
259)。
APP が切断される場合、3 つの切断ヵ所があります。それは、①P3(図 38)と呼ばれる
毒性のない小さなタンパク質の切断に関与するα部位、②毒性のあるβアミロイドの分泌
に関与するβ部位、③α、β部位で切断された後 APP の細胞膜に結合した部分から APP の
細胞膜貫通部分が切断され、P3 とβアミロイドの細胞膜貫通部分が切り出されるγ部位で
す(図 38)。α、β、γ部位を切断する酵素を、それぞれαセクレターゼ、βセクレターゼ、
γセクレターゼと呼びます。
ほとんどの APP は切断されずに細胞膜に残っています
260)。βアミロイドが分泌される
場合、βセクレターゼは APP の細胞の外側に出た細胞膜から 29 番目と 30 番目のアミノ酸
の間を切断します。βセクレターゼは BACE1 と呼ばれる酵素です
261)。BACE1
はタンパ
ク質のアミノ酸配列中のアスパラギン酸がある部分を切断するため、APP の特定な位置を
切断します。一方、APP の膜貫通部分はγセクレターゼと呼ばれる複合酵素によって膜表
面から 11 から 14 個目のアミノ酸と次のアミノ酸の間が切断されます。このため、39 から
43 個のアミノ酸で構成されたβアミロイドが切り出されます 227,255)。ヒトの脳には 40 個の
アミノ酸から成るβアミロイド(以下では Aβ40 と表記します)がβアミロイド全体の約 8
割、次いで Aβ42 が 2 割程度を占め、Aβ43 は若干量です。AD に限らず高齢者の脳にある
老人斑中に占める主なβアミロイドは Aβ42 です 262)。
APP が切断される場合、通常は、αセクレターゼ(ADAM と呼ばれる酵素と言われて
います)によって細胞外の細胞膜から 12 個目と 13 個目のアミノ酸間が切られるため(図
38)、βアミロイドは分泌されません 263,264)。ADAM は細胞の外のマトリックス(13 ペー
ジ)にある酵素です。各細胞がマトリックス成分と、どのように関係するかによって、細
胞内へのシグナリングや細胞の移動が行われ、体組織が維持・形成されます。このため
ADAM などの細胞外タンパク分解酵素はシナプスの LTP(36 ページ)やニューロンネット
ワークの形成(35 ページ)にも重要です 265)。ADAM は細胞の外部にあり、一方、BACE1
は膜に結合しており、両者は存在場所が異なることからも ADAM と BACE1 による APP
の切断には異なる意味があると考えられます。また、APP が BACE1 によって切断を受け
る領域は種によって異なることから、脳におけるβアミロイドの分泌は APP の通常の役
割とは関係していないと考えられています 266)。
APP の機能はβアミロイドの分泌に限定されていません。APP は胎児の段階で神経細
胞が成熟してニューロンに分化する時に増強され 267,268)、また、ヒトでもショウジョウバ
エでも強いストレスを受けた脳で増強されます 269–271)。最近では、APP にはニューロンの
保護、細胞移動、軸索輸送、シナプス形成などの機能があるといわれています 272)。
以前から、βアミロイドは神経作用に特定の役割を果たしているのではないかといわれ
ていました。その理由は、
1) 50 歳ごろから脳内のβアミロイドの量は増え出し、人によって濃度がバラつくよう
47
ANM176®が認知症の予防と改善に役立つ可能性について
になりますが、実は、若いころから一定量のβアミロイドが脳にある 273)。
2) APP の膜貫通部分を切断するγセクレターゼは 4 つのタンパク質が寄り集まった複
合タンパク質で、その構成タンパク質の 1 つである PS1 は、他の重要な神経作用に
関係するタンパク質の切断にも関与している 274)。
3) APP からβアミロイドの切り出しに関与するβセクレターゼとγセクレターゼの両
方とも、正規のプロセスで細胞内を移動し 275)、通常のタンパク質分泌を行う個所に
存在している 276)。
4) 培養したニューロンからβアミロイドを極端に減らすとニューロンが死ぬという報
告がある 277)。
ニューロンの興奮が適切に抑制されることによって、正常な神経活動ができると考えら
れています 278,279)。また、低濃度の Aβ40 は培養ニューロンを活発にし 261,277)、マウスの
試験で内因性のβアミロイドを抑制すると LTP や記憶力が低下したと報告されています
280)。シナプスの活動はβアミロイドの分泌を促進することから 281–283)、βアミロイドは過
剰なシナプス活動を自動的に抑制する役割を果たしているかもしれないとも考えられてい
ます 279)。βアミロイドは高濃度でも低濃度でもシナプス伝達を障害するため 284)、適正な
濃度が必要であると報告されています 278)。神経作用で興奮が適切に抑制されることが重
要なことは、興奮性の神経伝達物質であるアドレナリンとアドレナリンの作用を抑制する
ノンアドレナリンの例からも窺えます。
(βアミロイドの神経毒性作用)
A
βアミロイドは、動物を用いた試験で記憶や学習
力を低下させ、細胞試験ではニューロンの LTP(36
ページ)やシナプス形成など正常な神経活動を障害し
232,285)、通常の細胞においても機能を障害する 286)
B
作用があります。
動物を使った記憶・学習試験は多数ありますが、
その中の受動的回避試験では、明るい部屋(明
室)と暗い部屋(暗室)の間に仕切りがある箱を
C
使います。マウスを明室に入れると本来の性格か
ら直ぐに暗室に移動します。マウスが暗室に移動
してから明室の間にある仕切りをしめ、床に微弱
な電流を流しますとマウスはショックを受けます
(図 39)。このショックを受けたマウスを明室
に入れると、今度は暗室にはなかなか入ろうとし
ません。ところが、βアミロイドを脳の奥にある第
三脳室(図 5、マウスの第三脳室もヒトの脳と同じ
図 39.マウスによる受動的回避試験
A.マウスを明るい部屋(明室)に入れる
と暗い部屋(暗室)に入る習性がある。
B.全てのマウスが暗室に入ったら床に微
弱な電流を流してショックを与える。
C.ショックを与えた後明室に入れると、
正常なマウスは暗室に入ろうとしない。
暗室に入るまでの時間が長いほど正常に
近いことを示している。
48
ANM176®が認知症の予防と改善に役立つ可能性について
ような場所にある)に注入したマウスは、ショックを受けた記憶がよく定着していないた
め、直ぐに暗室に入ってしまいます。このβアミロイドの注入は極細のガラス管を、脳を
傷つけないように、耳の下から第三脳室内に差し込んで行います。第三脳室内にある脳髄
液はニューロンの周辺にある間質液に浸透することができるため、第三脳室に注入したβ
アミロイドはニューロンに作用します。
このように、学習力や記憶力を抑制するβアミロイドの作用は以前から知られており、
このβアミロイドが神経活動を害する作用を「βアミロイド神経毒性」と言います。実際
に脳に存在するほとんどのβアミロイドは構成するアミノ酸の数が 40、42、43 個のそれ
ぞれ Aβ40、Aβ42、Aβ43 ですが、構成しているアミノ酸集が多いほど、つまり、Aβ40<
Aβ42<Aβ43 の順で神経毒性が強いと言われています 287)。
ところが、1 分子のβアミロイド 288)(以下では「βアミロイド・モノマー」と言いま
す)や完全に線維状になったβアミロイド(以下では「線維状βアミロイド」と言いま
す)あるいはβアミロイドとアミノ酸配列が逆のタンパク質 234,289)には神経毒性はありま
せん。このことは何を意味しているのでしょうか?
この疑問を解く鍵は、βアミロイド分子の立体構造にあります。タンパク質の立体構造は
アミノ酸の配列によって決まります。βアミロイドとアミノ酸が逆配列のタンパク質は、重
さも成分もβアミロイドと全く同じですが、立体構造だけが異なっています。では、βアミ
ロイド・モノマーと線維状βアミロイドには、なぜ神経毒性がないのでしょうか? タンパ
ク質の立体構造はα螺旋
(らせん)構造とβシー
ト構造(図 40)および一
定の構造を取っていない
ランダム構造から成り立
っています。α螺旋構造
α螺旋構造
β シート構造
線維状
図 40.βアミロイドタンパク質の構造
アミノ酸が連なったタンパク質を青い線で示しています。
は一定方向に折れ曲がり螺旋のような構造です。
アミノ酸が椅子(シート)のように左右に折れ曲
A
B
がり、この折れ曲がりが幾つも連なって階段状と
なった構造をβシート構造と言います。βシート
構造は、他のタンパク質のβシート構造と密着す
る性質があり、βアミロイドの分子同士がβシー
図 41.βアミロイドの凝集体(A)と
線維(B)288)より
ト構造同士でピッタリくっ付くと水に溶けない線
維状βアミロイドになります。脳内の老人斑中に蓄積しているのは線維状βアミロイドで
す(図 41)。
21 世紀になってから、βアミロイドの神経毒性の元は、数個のβアミロイド分子が寄せ
集まったβアミロイド・オリゴマーにあると考えられるようになってきました 232,285)。βア
ミロイド・オリゴマーは、βアミロイド同士がβシート構造によって緩く密着したもので、
49
ANM176®が認知症の予防と改善に役立つ可能性について
水溶性です。βアミロイド・オリゴマーは、短期記憶と関係があるニューロンの LTP やシ
ナプスでの神経活動に悪影響を及ぼし 232,285)、記憶力の低下を招きます 285,290)。
βアミロイドの「アミロイド」とは、ヨウ素試薬で紫に発色する物質の総称です。小学校
の理科で、ヨード液をのり状の澱粉に垂らすと紫色に発色しますが、澱粉を口の中でよく噛
んだ後は発色しなかった実験を思い出します。のり状澱粉は、長く連なったグルコースが螺
旋構造になっています。その螺旋構造中の空間にヨード液中のヨウ素分子が入って紫色に
発色します。ヨウ素を発色させるのは澱粉に限りません。キトサンなどの多糖類やタンパク
質もヨウ素で紫色に発色することがあります。澱粉の場合とは違って、タンパク質の螺旋構
造にはヨウ素分子は入れず発色しません。一方、幾つかのβシート構造が形成したクロスβ
シート構造と呼ばれるタンパク質の立体構造にはヨウ素分子が入って発色します。このよ
うにヨウ素で発色する構造を総称してアミロイド構造と言います。
DNA の情報からタンパク質が出来上がると、通常は、タンパク質は自動的に適正な立
体構造になります。正常な立体構造はタンパク質の機能に必須のため、タンパク質を合成
する現場である小胞体(ER)では、タンパク質品質管理機構によってタンパク質の立体構
造を高度に管理しています 291,292)(25 ページ)。この機構によって、正常な高次構造を形成し
たタンパク質だけが分泌経路に乗って、それぞれのタンパク質が働く現場に出ることがで
きます。立体構造が正常でないタンパク質は、再度、組み立てられ直し、それでも正常に
ならなかったタンパク質は ER から細胞質に輸送されて、タンパク質を分解する細胞内器
官のプロテアソーム(図 12)で分解されます 291)。このタンパク質品質管理機構に問題が
起きると、細胞の内外に変性タンパクが凝集して、細胞の機能不全や細胞死が起こると考
えられています 55)。
細胞内にはタンパク質の立体構造の形成を補助するシャペロンと呼ばれる小さなタンパ
ク質が多数あります。シャペロンには、細胞が高熱に曝されるなどの熱ショックによって
細胞内に現れるものがあり、これを熱ショックタンパク質(以下では heat shock protein
から Hsp と表記します)と言います。タンパク質は熱によって変性しやすく、Hsp はタン
パク質立体構造の正しい形成に働いているだけでなく、立体構造が正しくないタンパク質
の分解プロセスにも関係します。Hsp ファミリーの Hsp22 や Hsp27 の突然変異は、パ
ーキンソン病、アルツハイマー病、アレキサンダー病*、多発性硬化症**などの神経変性
疾患の発症を促進する可能性が報告されています 293)。
*アレキサンダー病(Alexander Disease)は幼児期に発症し、大頭、痙攣、運動異常、延髄性症状が緩
慢に進行し、通常は 10 歳未満で死に至ると言われています。この患者の脳内アストロサイトの細胞
質にはローゼンタール線維と呼ばれる封入体があり、その中には Hsp ファミリーの hsp27 やαB ク
リスタリンあるいは変質した GFAP(29 ページ注)の凝集体が見られ観察されます 294)。
**多発性硬化症(multiple sclerosis)は脳の神経に広範囲にわたって神経伝達の絶縁体のような役
割があるミエリン鞘(図 3)の脱落と硬化が多発的に起こり、さらにグリア細胞(7 ページ)の炎症
状態によって、運動麻痺、感覚障害、深部反射亢進、視力障害、病的反射など多彩な症状が現れる
病気です。特定の症状が決まって起こるわけではありません。長期にわたって症状が良くなったり
悪くなったりを繰り返します。原因はよく分かっていませんが、毒物、一部の薬剤、過度の飲酒な
ども発病の引き金となることがあると言われています。日本では、約 80%が 15~50 歳で発症し、
患者数は、人口 10 万人あたり 3~5 人と言われています。
50
ANM176®が認知症の予防と改善に役立つ可能性について
Hsp のファミリーには、βアミロイド・オリゴマーの毒性を低下させるものがあります
295,296)。Hsp
のαB クリスタリン(Hsp16 あるいは CRYAB とも呼ぶ)は、βアミロイド
の線維形成を減少させ、βアミロイド神経毒性を低減させると報告されています 296)。ま
た、Hsp のプレフォールディン(PFD)は、異常や変質したタンパク質を安定させ再び正
しい構造にすることが示されています 297–299)。PFD はヒトの脳内にもあって 300)、βアミ
ロイド・モノマーと PFD を一緒にして 48 時間経ってもβアミロイドの線維形成を抑制し
ており、PFD がβアミロイド・オリゴマー形成に関与している可能性を示唆しています
301)。Hsp
の Omi302)はβアミロイド・オリゴマーにある PDZ ドメインで結合して濃度依
存的にβアミロイド・オリゴマーの毒性を著しく弱めることができます(303,304)。一方、
Hsp のアポリポタンパク J(Clusterin とも呼びます、以下では apolipoprotein J から
ApoJ と表記します)はβアミロイド・オリゴマーの形成を促進し 305,306)、細胞死とも関係
すると報告されています 307)。
(βアミロイド神経毒性はどのようにして起こるのか?)
βアミロイドの形状によって MTT 法で示される毒性が異なります。数分子から成るβ
アミロイド・オリゴマー308–310)と数十分子から成るβアミロイド凝集体 309–311)は MTT 法
による細胞毒性を示し、βアミロイドの凝集状態と細胞毒性はよく相関すると報告されて
います 286)。また、42 個のアミノ酸から成る Aβ42 は Aβ40 に比してすばやく凝集体を形
成し 308,312)、MTT 法で観察する細胞毒性も Aβ40 と比較し Aβ42 の方がはるかに強いと報
告されています 310,313)。一方、βアミロイド・モノマーやβアミロイドとアミノ酸が逆配
列のタンパク質 314)には MTT 法による細胞毒性はありません 288,315)。アミノ酸の構成は同
じであっても、アミノ酸の配列が異なると立体構造は全く異なります。つまり、βアミロ
イドが寄り集まったものは、MTT 法で感知される刺激を細胞に及ぼしたということにな
ります。MTT 法による発色は、細胞質 100)、細胞膜 101,102)等にある電子伝達系に活発な電
子があることを示しています(31 ページ)。
細胞に障害をもたらす異物を細胞膜が感知したシグナルは G タンパク質の活性化を経て
PI3K を活性化し、膜にあるイノシトールリン脂質から PIP3 が増産されます(17 ペーシ)。
PIP3 は NADPH オキシダーゼを構成しているユニットの1つ p47phox に結合して NADPH
オキシダーゼを活性化します 41,43) (32 ページ)。アクチン重合や細胞膜の変化が関与する細
胞運動には PIP3 が活性化した NADPH オキシダーゼによって生成された NADPH が関与
します。PIP3 は小胞体 99,100)や細胞質 100)に存在する数多くの酸化分解に関連する第Ⅰ、Ⅱ
相系酵素群を活性化します(32 ページ)。
病原菌などの細菌を感知する TLR(15 ページ)は大阪大学の審良教授らが発見した有名な
受容体です 316)。TLR は、攻撃してきた細菌の外壁物質を感知して細胞内に伝達し、細胞の
様々な防御システムを稼働させます。この細菌の外壁物質に、よく毒性実験で使用されるリ
ポ多糖類(LPS)があります。TLR の相同体 TLR4 は LPS を感知する代表的な受容体です
51
ANM176®が認知症の予防と改善に役立つ可能性について
が 316)、一般に細胞が自然界の異物に対処する時には、免疫に関係する受容体、異物の構造
認識に関係する受容体、異物を細胞膜が包み込むことに関係する受容体など様々な機能を
持った多くの受容体が協調して行われます 317–319)。
インテグリン(15 ページ注)ファミリーの MAC1(またはインテグリンαMβ2)は好中
球の呼吸バーストや細胞表面での電子伝達系の活性化に関与しています 108,318)。インテグ
リンは 2 量体となることによって受容体としても機能します。MAC1 の受容体 MAC1L*
は NADPH オキシダーゼを活性化します 103)。
*MAC1 の膜表面受容体 MAC1L には細胞間の接着と細菌の外殻分子構造を認識するパターン認識受
容体(14 ページ参照)の両方の機能があり、接着、遊走、ファゴサイトーシス 318,320)、呼吸バースト
108)、走化性の機能に関与しています 108,321)。
細菌類外壁の LPS が
MAC1L を含む複合受
容体によって感知され
ると G タンパク質の活
性化を経て PIP3 が細
胞膜で作られます 41)。
好中球やマクロファー
ジにおいては、取り込
んだ異物を包み込んで
形成されるファゴソー
ムの成熟は表面膜から
分離・シール後、直ぐ
に開始され、ファゴソ
ームは早期エンドソー
ム、後期エンドソー
ム、リソソーム
図 42.リソソームへの経路と機能
細胞内外成分を取り込んだファゴソームは、初期ファゴソームから後期ファゴソ
ームを経るエンドサイトーシス経路で、細胞内に蓄積された物質はオートファジ
ー経路でリソソームへ輸送される。リソソーム内には各種の加水分解酵素が存在
し、基質をアミノ酸、脂質、糖などにまで分解する。リソソームは細胞外に放出
される(エキソサイト―シス)こともある。
(lysosomes)と融合
します 322)(図 42)。このプロセスに PIP3 が関係します 323,324) 。
βアミロイド凝集体は、ミクログリアの細胞膜にある MAC1L・インテグリンα6β1・
CD36・CD47 複合受容体(以下ではβアミロイド複合受容体といいます)を刺激して G
タンパク質を活性化します 103,325)。βアミロイド複合受容体を構成している CD36 と
CD47 はスカベンジャー受容体と呼ばれる仲間で、CD36 は他の受容体と一緒になって
LPS など細菌が作った様々な物質の構造を認識します。ミクログリアでβシート構造を持
ったβアミロイドによるチロシンキナーゼシグナリング(16 ページ)の活性化が観察されて
います 325,326)。CD47 は細胞内へのチロシンシグナリングの開始にも関与します 327)。βア
ミロイドはβアミロイド複合受容体を通して呼吸バーストや代表的な炎症性サイトカイン
である IL-1βの分泌を促進します 103,325,328)。
52
ANM176®が認知症の予防と改善に役立つ可能性について
βアミロイドはミクログリアとアストロサイト(7 ページ)を活性化し 234,329)、炎症性サイ
トカイン IL-1β234)の発現や IL-6 と IL-8 の分泌を促進します
330)。この
IL-1βによるサイ
トカイン分泌はβシート構造を持ったβアミロイドに見られたが、βシート構造を持たな
いβアミロイドの部分では観察されなかったと報告されています 330)。また、IL-1βの存在
下では、βアミロイド凝集体は著しく(3~8 倍)サイトカインを分泌させました 330)。
βアミロイドと細菌類外壁の LPS による神経毒性作用は似ており、LPS によって細胞膜
受容体 TLR4(51 ページ)が活性化するシグナリング経路とβアミロイドが活性化するシグナ
リング経路とは区分けがつかないと報告されています(3325,331)。βアミロイドと LPS の両方
とも IL-1βの分泌を促進し
41,332,333)、βアミロイド 232,334)と
LPS147,335)の両方とも LTP を
抑制するとともに海馬が関係する学習や記憶を害します 336,337)。また、βアミロイドと LPS
の両方とも MTT 法(31 ページ)による細胞毒性があります 98,309,310)。LPS は LRP4 を活性化
して MAPK 経路の p38(16 ページ)を活性化します
NF-κB を活性化し
します
は炎症と関係する転写因子
339,340)、その結果、炎症を引き起こします 40,341)。βアミロイド凝集体
も、ミクログリアにおいて p38 を活性化し
344–346)。LPS
338)。p38
342,343)、転写因子の
NF-κB と c-Jun を活性化
や線維状βアミロイド処理による IL-1β濃度上昇とシナプス機能の傷
害は、抗炎症剤を投与すると抑制されることから
333,337,347)、LPS
とβアミロイドによる細
胞毒性や神経毒性は炎症と関係すると考えられます。
以上から、細胞外にあるβシート構造をもったβアミロイド凝集体がミクログリアに取
り込まれる時に、ミクログリア表面の電子伝達系が活発になり、PI3K が活性化して PIP3
が作られ細胞内にシグナリングし、酸化ストレス応答が起こります。この酸化ストレス応答
や活発化した電子伝達系に繋がる内因性の炎症がニューロンの LTP などの神経作用を抑制
し、記憶や学習力を阻害する神経毒性を示すと考えられます。
(βアミロイドの分解)
βアミロイドは、その形状や存在する場所に応じて、様々なタンパク分解酵素やリソソー
ムやオートファジーなど細胞内器官(図 42)の関与によって分解され解消されます。βア
ミロイドを酵素で分解する場合は、
膜上ではネプリライシン
(以下では neprilysin から NEP
と表記します)348,349)、細胞外ではプラスミン 350)などのマトリックスプロテアーゼ*、細胞
内ではリソソーム内のインスリン分解酵素**(以下では insulin degrading enzyme から
IDE と表記します)351,352)やカテプシン D 353,354)などがあります 255,353)。
*細胞外のマトリックスにあるタンパク質分解酵素のマトリックスプロテアーゼはマトリックスや細胞
膜にあるタンパク質を切断して、細胞の接着や細胞内シグナリングに重要な役割を果たしています 355)。
βアミロイドの前駆タンパク質 APP のα部位を切断して無毒の P3 の分泌に関わる ADAM もマトリ
ックスプロテアーゼです。
**インスリン分解酵素(IDE)は脳、肝臓、筋肉に多く発現し 356,357)、細胞内に普遍的に見られるが、
一部は細胞膜やペルオキシソーム、エンドソーム、ミトコンドリア中にも見られ、βアミロイド、グ
ルカゴン(glucagon)、アミリン(amylin)、ソマトスタチン(somatostatin)、心房性ナトリウム利尿
ペプチド(natriuretic peptide)などを分解します 358–361)。
インスリン分解酵素 IDE の活性を阻害するインスリンあるいはカテプシン D の阻害剤
53
ANM176®が認知症の予防と改善に役立つ可能性について
pepstatin A を投与した 72 時間後のミクログリアでは、βアミロイド・オリゴマーの減少
が抑制されたが、膜でβアミロイドを分解する NEP の阻害剤には、この効果がなかったと
報告されています 362)。このことは、ミクログリアが取り込んで分解する場合と細胞膜での
NEP によるβアミロイドの切断は、本質的に異なっていることを示唆しています。NEP は
体中に広く見られますが 363)、脳の神経系細胞では、主に海馬における貫通線維束、Schaffer
側枝、反回側枝など(図 25)のニューロンで 364)、特に前シナプスに存在し 365)、βアミロ
イドの分解はそこで行われると報告されています 348,366)。
細胞内シグナリングや神経伝達に働いているタンパク質の分解は生理作用に影響し、生
命にとって重要な意味があります。そのためタンパク質を分解する細胞内システムは基本
的に何らかの制御を受けていると考えられます。NEP は膜に結合した酵素で、膜に結合し
た別のタンパク質の配列中を切断します。NEP はβアミロイドだけでなく、エンケファリ
ン、ソマトスタチン、サブスタンス P などの神経作用に関係するタンパク質や膵臓ランゲ
ルハンス島β細胞から分泌されるアミリン(IAPP とも呼ばれます)あるいは血中やリンパ
液中を循環しているエンドセリン、ブラジキニンなど多くのタンパク質を分解して様々な
生理機能に関係しています
367)。NEP
は分泌されたβアミロイドのレベル調整を果たして
いる有力な酵素ですが 368)(図 43)、細胞膜に局在しています。
図 43.ネプリライシン(neprilysin;NEP)によるβアミロイド分解における pH の影響
神経細胞内の分泌顆粒内は pH 5~の酸性であるのに対して、細胞間隙(細胞外)は pH 7 を超える
中性です。Aβ40 はいずれの pH でもネプリライシンに分解されますが Aβ42 は中性でのみ効率的に
分解されます 369)。理化学研究所 HP
一方、エンドサイトーシス(図 42)によって細胞内に取り込まれた細胞外のβアミロイ
ドを分解するリソソーム中は pH5 以下の酸性状態です。42 個のアミノ酸から成る Aβ42(47
ページ)は pH 7.2 に比して pH 5 の条件下では 1/10 程度しか分解されないと報告されてい
ます 369)。これらのことは、既に細胞内外に蓄積した Aβ42 は分解され難いことを示唆して
います。
54
ANM176®が認知症の予防と改善に役立つ可能性について
(タンパク質の凝集体とオートファジー)
細胞内には様々な分解機構がありますが、その代表的なリソソームには 70 種類もの分
解酵素が含まれており、様々なタンパク質を分解できます。リソソーム中は酸性でしかも
強力な酸化分解酵素が含まれているため、リソソームに分解物が細胞質に漏れ出さないよ
うに 2 重膜で厳重に包まれています。このため、分解するものを袋に包んで、その包みを
リソソームに融合するという特殊な方法が取られます。分解物を包み込みリソソームに運
び込んで分解する方法は大きく 2 通りあります。その 1 つはファゴサイト・リソソーム系
で、もう一つはオートファジー・リソソーム系です。ファゴサイト・リソソーム系では、
細胞外の物質を細胞膜が陥没して包み込み、この包みをエンドソーム(図 44)と呼びま
す。このエンドソームを細胞内に輸送してリソソームと融合してファゴサイトリソソーム
中の内容物を分解する方法です。
オートファジー(autophagy)は細胞内にある不要物や古くなった細胞内構造物を包み
込むシステムで、オートファジー・リソソーム系では、エンドソームと同じようにオート
ファジーにリソソームが融合してオートリソソーム中のタンパク質や構造物を分解します
370,371)(図
44)。オートファジーによって浄化された細胞内には構造物が新たに作られま
す。細胞分裂しない神経細胞や 1 年以上入れ替わらない肝細胞などではオートファジーに
よる細胞質内の分解・浄化機構が重要です。
図 44.オートファジーの過程
飢餓などによりオートファジーが誘導されると、細胞質に出現した隔離膜が伸張し、細胞質のタンパク質やミトコ
ンドリアなどの細胞内構造物を含みながら閉鎖することでオートファゴソームが形成されます。オートファゴソー
ムの外膜とリソソームが融合し、オートファゴソームの内膜と取り囲まれた細胞質成分が分解されます。オートフ
ァジーが生じると、主要な産物としてアミノ酸が供給されます。(西村多喜、水島昇 [オートファジー] 脳科学辞典
2012 年 4 月 17 日)
オートファジーには大きく 2 通りがあります。その 1 つは、栄養が足りなかったり、外か
ら供給される栄養が不足したり、細胞に危機が訪れたりすると盛んになる選択的オートファ
ジーで 372)、もう1つは、常に一定レベルで行われている基底オートファジー(非選択的オ
ートファジーとも呼びます)です 370,371)。
LC3 はオートファジーの膜にあるタンパク質で、オートファジーの開始に関与していま
す。選択的オートファジーに最も特徴的なタンパク質である p62 と呼ばれるタンパク質が
オートファジーの取り込みに関与しています 373,374)。p62 は LC3 と結合し、一方、p62 の C
末端側にはユビキチンと結合するユビキチン鎖結合ドメイン(UBA)があるため(図
55
ANM176®が認知症の予防と改善に役立つ可能性について
45A)、ユビキチン化され
たタンパク質凝集体やダメ
ージミトコンドリアなどの
脂肪内器官などと UBA ド
メインで結合し、これらを
オートファジーに送り込み
ます 375,376)。すなわち、オ
ートファジーによってユビ
キチン化タンパク質の選択
的な分解が行われます(図
45B)。
オートファジーを抑制し
ているセリン・スレオニン
キナーゼの mTOR
(mammalian Target of
Rapamycin)は、
mTORC1(mTOR
Complex1)と mTORC2
の 2 種類の複合体を形成
し、この複合体はそれぞれ
独立したネットワークで機
能しています 377,378)。オー
トファジー開始に関与する
タンパク質複合体は
379)、
冨栄養状態では細胞質に拡
散して mTOR1 によって不
活性化されています 380)。
または、成長因子、ホルモ
ン、ストレスなどによって
mTOR1 複合体内の結合が
弱くなり、その結果
mTOR が自己リン酸化し
て活性化し、オートファジ
図 45.P62 と LC3 との相互作用を介した選択的オートファジー
A.N 末端の PB1 ドメインを介して、他の PB ドメインを持つタンパク質と
オリゴマー構造を作ります。UBA ドメインではユビキチン鎖と相互作用し、
ジンクフィンガードメインを介して RIPI や TRAF6 と相互作用します。LC3
認識配列(LRS)に LC3 を結合してオートファジーへ局在化されます。
B.上記のドメインを介してユビキチン化タンパク質が凝集体を形成した後に
オートファゴソームに至り分解されます。
C.肝臓における Nrf2 の Keap1 による制御とオートファジーの関わり.
通常 Nrf2 は Keap1 によってユビキチン鎖の付加を受けプロテアソームで
分解されます。オートファジーによって増加した P62 や酸化ストレスによっ
て Keap1 は Nrf2 のユビキチン化ができなくなり、分解を免れた Nrf2 は核
に至り酸化ストレス防御タンパク質の翻訳を開始させます 371)。
ーが抑制されます 381)。
一方、mTORC1 のキナーゼ活性は獲得免疫に関わる T 細胞の増殖活性化を阻害する作
用を持つ免疫抑制剤ラパマイシン(Rapamycin)*により阻害されてオートファジーが誘
56
ANM176®が認知症の予防と改善に役立つ可能性について
導されます 382,383)。
*ラパマイシンはマクロライド系抗生物質で、移植における免疫抑制剤や細胞分裂を抑制する作用を応
用して血管を拡張する「薬剤溶出ステント」として利用されています。
ラパマイシンは、AD モデルマウスでβアミロイドとタウの産生経路を抑制し
384)、また、
培養細胞でβアミロイドの解消を促進したと報告されています 385)。
細胞の生長に関わるシグナリングキナーゼの Akt は mTOR1 を活性化してオートファジ
ー誘導を負に制御しています 386,387)。Akt は PDK1 からリン酸化されて活性化しますが
(17 ページ)、もう 1 つの mTOR の複合体 mTORC2 は、Akt が PDKⅠによってリン酸化さ
れるアミノ酸とは異なるアミノ酸をリン酸化して Akt の活性を抑制すると報告されていま
す 388)。このことから、Akt は mTORC によるポジティブとネガティブのフィードバック
の両方を通じて制御していると考えられています。
翻訳開始に関係する複合体 eIF4F を構成する eIF4E に 4EBP が結合することによって
eIF4F 複合体形成が阻害され、翻訳が抑制されます(21 ページ)。一方、mTOR はアミノ酸や
グルコースなどの栄養源状態を感知し、また、増殖因子、低酸素、ATP 枯渇などからのシ
グナルを統合して、4EBP の相同分子 4EBP1 をリン酸化し eIF4E 結合能が抑制され、
mRNA の翻訳とタンパク質合成を亢進させ 389,390)、細胞の成長、増殖、生存など広範な機
能を調節します 377,391)。mTOR は低酸素、ATP 枯渇などからのシグナルを統合して、4EBP
の相同分子 4EBP1 をリン酸化し eIF4E 結合能が抑制され、mRNA の翻訳とタンパク質合
成を亢進させ 389,390)、細胞の成長、増殖、生存など広範な機能を調節します 377,391)。
選択的オートファジーに取り込まれたタンパク質は、アミノ酸やペプチドに分解され
392)、細胞内にアミノ酸が供給されます 393)。一般的に飢餓などの危機的な状態では細胞内の
アミノ酸合成やタンパク質合成は低下しますが、ストレス応答に関連するなど特定のタンパ
ク質合成はむしろ増加します 394,395)。この時に、オートファジーによるアミノ酸供給が重要
で、飢餓時にオートファジーが誘導されない実験では、細胞死を引き起こします 395)。この
ため、選択オートファジーの開始に関係している P62 が細胞内タンパク質の品質管理とし
て機能し、また、細胞内の浄化機構によって生体の恒常性維持に関与しているだけでなく、
炎症やアポトーシスなど様々なシグナル伝達経路の中継地点としても機能しています
371,374)。
タンパク質や老廃物が細胞内に溜まると細胞に悪影響を及ぼします。さらに、過剰なタン
パク質はしばしば凝集して様々な毒性を引き起こすことにもなります。細胞の中で過剰な
タンパク質が寄り集まって形成された凝集体は、オートファジーや不等細胞分裂*の際に処
理されます 396)。
*細胞が 2 つに分裂する時に、細胞の中央で均等に分裂する等分裂と、偏って分裂する不等分裂があり
ます。
三次構造に問題があるタンパク質を修復したり分解したりする細胞内浄化機構の能力が限
界になった時、細胞質でつくられる微小な凝集物は微小管(MT)に沿って中心体に能動的
に輸送され、凝集体として蓄積します 396)。そのような凝集体はオートファジーや不等細胞
57
ANM176®が認知症の予防と改善に役立つ可能性について
分裂の際に処理されます。オートファジーの開始に関与している LC3 の名は microtubuleassociated protein light chain 3 を短縮したものです。ニューロンの軸索や樹状突起にある
微小管結合タンパク質(MAP)の MAP1(41 ペーシ)は全長が翻訳された後、C 末端側の微小
管結合部分である LC3 が切断され、残された N 末端側の重鎖と複合体を形成して MT に結
合し MT を安定化します。MAP1 はアクチンにも結合できます。つまり、LC3 はオートフ
ァジーの開始に関与するだけでなく、MT の安定化やアクチンの形成にも関与しています。
LC3 に結合する p62 は常にオートファジーによって分解され、細胞内の p62 は適切な
濃度に調整されています 397)。過剰な p62 は、様々なストレスや炎症によって Keap 1 に
作用して Nrf2 を核内に移行させ、一群の第Ⅱ相系の抗酸化タンパク質などの Nrf2 ターゲ
ット遺伝子の転写を活性化します 374,398)(図 45C)。p62 は炎症やアポトーシスなど様々
なシグナル伝達経路の中継地点として機能します 374)。
p62 は、ユビキチンに結合する UBA ドメインとは別に、N 末端側に多量体化および凝
集体形成を行う PB1(Phox and Bem1p)ドメインを持っています(図 45A)。このドメイ
ンにより p62 は、ポリユビキチン化タンパク質に結合すると同時に自身が多量体となり、
不溶化ポリユビキチンタンパク凝集体を形成します。
オートファジーはアルツハイマー病(AD)
、レビー小体型認知症、パーキンソン病、ハン
チントン病*、筋萎縮性側索硬化症(以下では amyotrophic lateral sclerosis から ALS と表
記します)といった神経変性疾患(次の項を参照)だけでなく、炎症性腸疾患のクローン病
399)やアルコール性肝炎、
脂肪性肝炎の発症や肝臓ガン 400)の抑制にも関与していることが知
られています。また、オートファジーは AD 患者で抑制されている傾向があると報告されて
います 401,402)。
*ハンチントン病(Huntington's disease;HD)は遺伝性で、ハンチンチン(huntingtin)と呼ばれる
原因タンパク質が分かっています。症状としては歩行障害など小脳症状の不随意運動(このため舞踏
病とも呼ばれる)があります。HD 患者の 80%が発症後 10~15 年以内に何らかの精神障害をきたし
ます 403)。欧米人では 1 万人に 1 人ぐらいの発症率ですが、日本人などアジア人種では、その 10 分
の 1 と言われています。
(神経変性疾患について)
アルツハイマー病(AD)の特徴である大脳の神経原線維変化(NFT)やβアミロイドの
集積より、炎症に関連するマーカーの方が遥かに AD と関連し 404)、多くの免疫応答は数多
くの神経疾患、特に AD で明白であるといわれています 405,406)。脳、軟骨、骨格筋の分化や
制御・修復、あるいは癌化や脳内炎症に関係する S100B は、脳障害やニューロン死のプロ
セスで活性化したアストロサイトで観察されています
407)。そのため
S100B は、脳炎症の
マーカーとしても知られています。また、血清中 S100B 濃度上昇は、炎症や BBB 機能に
ダメージがあることを示しています
408)。
S100B が脳内で結合する相手は、アストロサイ
ト内の GFAP409)(29 ページ注)、ニューロン内の微小管構成分子のチューブリン、微小管関連
タンパク質のタウ、アクチン結合タンパク質など細胞骨格に関連するタンパク質
407,410)や
58
ANM176®が認知症の予防と改善に役立つ可能性について
G タンパク質の Rac1 など多数の細胞内シグナルや神経伝達に関連するタンパク質が報告
されています 407,410)。
神経系細胞の変質が進行する病気を神経変性疾患と言います。神経変性疾患には AD 以
外にも、レビー小体型認知症(DLB)、パーキンソン病(PC)など多数が知られています。
これらの神経変性疾患には脳内の炎症が深く関わっていると言われています 411)。ニューロ
ンの軸索を取り囲んでいるミエリン鞘(図 3)の炎症が長期にわたって進行する多発性硬化
症(50 ページ注**)では脳内の広範囲に炎症の証拠がみられ、脳梗塞でも細胞の崩壊を引き
起こす炎症状態が示されています 411)。
これらの神経変性疾患の原因タンパク質は、AD ではβアミロイド、DLB ではαシヌク
レインのように、それぞれ病気で異なります。PC 病の場合はαシヌクレインの場合と他の
タンパク質の場合があります。ポリグルタミン病(polygglutamine disease)は、アミノ酸
のグルタミン酸が幾つも連なったポリグルタミン(polygglutamine、以下では polyQ と言
います)の凝集が脳内に見られる神経変性疾患の総称で、遺伝子にアミノ酸のグルタミンを
コードする CAG(シトシン-アデニン-グアニン)配列の繰り返し(リピート)があるこ
とから CAG リピート病とも呼ばれます。ポリグルタミン病に分類されるハンチントン病、
脊 髄 小 脳 失 調 症 ( spinocerebellar ataxia ) * 、 歯 状 核 赤 核 淡 蒼 球 ル イ 体 萎 縮 症
(dentatorubropallidoluysial atrophy;DRPLA)**は原因タンパク質が異なりますが、共
通して原因タンパク質の DNA 中に CAG リピートがあります。
*脊髄小脳変性症は運動失調を主症状とする原因不明の神経変性疾患の総称です。体が不安定で起立の
維持が困難であったり、歩行時にふらついたりや手足の動きがチグハグだったりなどの運動失調症状
や筋固縮や無動などのパーキンソニズムが徐々に出現し、良くなったり悪くなったりしながら進行し
ます。運動失調だけの場合もありますが、起立性低血圧や排尿障害を中心とする自律神経症状、末梢
神経症状などを示す場合も少なくありません。さらに、ロレツが回らない、あるいは音と音がつなが
ってしまうなどの言葉の障害も見られます。脊髄小脳変性症の推定有病率は、10 万人当たり約 18 人
程度であり、そのうち約 70%は遺伝性でない孤発性で 30%が遺伝性です。
**歯状核赤核淡蒼球ルイ体萎縮症(DRPLA)の名前は脳基底核である歯状核、赤核、淡蒼球、ルイ体
に異常があることに由来します。DRPLA の症状は、ミオクローヌス発作(突然倒れたりする発作)
、
認知症、協調運動の障害、不随意運動です。12 番染色体上の DRPLA 遺伝子に CAG リピートが多い
遺伝病です。
polyQ を含む封入体は神経系に限らず多くの臓器に広く見られますが、神経系だけが選
択的に障害されます。polyQ は核内と細胞質の両方に見られ、核内の polyQ は転写調節機
構に影響を及ぼし、また、ニューロンの細胞質にある polyQ はタンパク質品質管理機構に
影響して神経機能障害や神経細胞死を引き起こすと考えられています。ポリグルタミン病
は共通して、原因タンパクの発現量は同じでも CAG リピートの数が多いことと発病が相関
しています。例えば、CAG リピートが長いほど発症年齢が若年化し重症化傾向があります。
CAG リピートはβシート構造(図 40)をとりやすく 412)、βシート構造に富んだアミロイ
ド線維を形成し 413,414)、βシート構造が相互に結合して凝集することが示されています 415)。
これらのことから、伸長したポリグルタミン鎖を持ったタンパク質中のβシート構造がポ
リグルタミン病の発病に関連しているのではないかと考えられるようになってきました。
ク ロ イ ツ フ ェ ル ト ヤ コ ブ 病 { 牛 の 場 合 は ウ シ 海 綿 状 脳 症 ( Bovine Spongiform
59
ANM176®が認知症の予防と改善に役立つ可能性について
Encephalopathy;BSE)}はプリオンが原因タンパク質のプリオン病の 1 種です。プリオン
病も認知機能が障害されます。
*クロイツフェルトヤコブ病(CJD)は、日本におけるプリオン病のうち 1 割程度を占め、伝染性が
あります。多くの場合は 40~50 歳代で発症し男女差はありません。認知症は徐々に進行し数年後に
無言無動状態になります。
認知症ではない神経変性疾患もあり、宇宙論で有名なホーキング博士がかかった筋萎縮
性側索硬化症(ALS)は筋肉を動かす神経に異常が生じる神経変性疾患の1つですが、認知
機能は正常です。ピック病と ALS は TDP-43 と呼ばれるタンパク質が原因と言われます
416)。神経変性疾患は原因タンパク質が異なっても、それが脳内や神経で凝集することが共
通しています。原因タンパク質が凝集する脳の部位が異なることによって、様々な症状が起
きるのです。例えば AD では記憶に関係する海馬の周辺、DLB や PC の場合は運動に関係
する小脳に異常が集中して見られます。
また、神経機能とは関係ない変性疾患もあります。透析患者の手の付け根にある手根管
417)と呼ばれる血管の内壁、糖尿病患者の腎にあるメサンギウム領域 418)、家族性アミロイド
ポリニューロパチーで体中、などで蓄積したアミロイドタンパク質は、それぞれの局部に特
有な症状を引き起こします。これらの原因タンパク質は、透析ではβ2 ミクログロブリン
417)、糖尿病患者の腎メサンギウム領域 418,419)、粥状動脈硬化性血管病変 420)では終末糖化産
物(AGE)
、家族性アミロイドポリニューロパチーではトランスサイレチンです
421)。凝集
したタンパク質が原因で起こる病気を総称してアミロイドーシスと言い 421)、現在までに 28
種類のアミロイドーシスが知られています 422)。アミロイドーシスはタンパク質のアミロイ
ド構造(49 ページ)が引き起こす病気という意味で、AD もβアミロイドが原因タンパク質の
アミロイドーシスの一種です。
多発性硬化症(50 ページ注**)、脳梗塞、AD などの多くの病気は、細胞の機能不全や崩壊
を引き起こす炎症状態を示しています 411)。
アポトーシスに関連しているセリン・スレオニンキナーゼの ASK(35 ページ)は、ポリグ
ルタミン病や AD において認められる神経細胞死を仲介していることが示唆されています
146,150)。ASKl
欠損細胞ではポリグルタミン(polyQ)による細胞死が抑制されることか
ら、polyQ の過剰発現による神経細胞死には ER ストレスから IRE1-ASK-JNK 経路が
関与すると考えられています 423)。線虫において、老化による熱ショックタンパク質
(Hsp)の発現量の低下は polyQ 凝集体の形成を早めることが示されています 424)。ま
た、ハンチントン病において Hsp27 は防護的な影響を持つことが報告されています 425)。
神経変性疾患の発症には、それぞれの疾患の原因となるタンパク質にアミノ酸配列の類
似性はありません。しかし、これらタンパク質の構造に共通した原因があり、いずれもβシ
ート構造に富んだアミロイド線維を形成し 413,414)、これらが細胞内に蓄積して細胞傷害を引
き起こすと考えられています 426)。これらの立体構造がどのようにして神経変性疾患を引き
起こしているのか分かっていないことも、それぞれの原因タンパク質に共通しています。
60
ANM176®が認知症の予防と改善に役立つ可能性について
(アルツハイマー病の発症リスク)
遺伝性の家族性アルツハイマー病(FAD)の遺伝子にはβアミロイド分泌に直接関係して
いる変異があります 226,227,286)。一方、アルツハイマー病(AD)患者の 95%以上を占める老
年性アルツハイマー型認知症(SDAT)ではβアミロイド分泌に関する遺伝子は正常です。
FAD でも SDAT でも、死後解剖で脳内に多数見られる老人斑には、主にβアミロイドが
凝集していますが、AD ではなかった健常者の死後脳内にも老人斑が見られます。今でも保
存されているアインシュタインの脳には多数の老人斑があるとのことです。このように、脳
内にβアミロイドが多いことは AD 発症の必要条件ですが、十分条件ではないようです。
SDAT は高齢になってから発症することや進行が遅いだけで、FAD とほとんど区別でき
ない症状を示します。このことは、両者に共通したメカニズムが存在することを示唆してい
ますが
427)、このメカニズムはまだよく分かっていません。遺伝性の
FAD においても、発
症率や重症度は家系により異なることが多く、βアミロイドの分泌に関連する遺伝子の変
異も 30%から 50%しか該当しないとのことです 428)。一方、βアミロイドの分泌に異常がな
い SDAT でも脳内βアミロイドの濃度が上昇することは、βアミロイド関連以外の遺伝子
や環境などの要因が AD の発病に関係していることを示唆しています 279,429)。つまり、βア
ミロイドの分泌とは直接的に関係しないが AD の発症とは関係する遺伝子変異が FAD にも
SDAT にもあり、これらの変異も AD 発症のリスクと考えられるのです。βアミロイドの分
泌とは直接関係しない AD 発症リスクで、アポリポタンパク質(以下では Apo lipoprotein
から Apo と表記します)E がよく知られています。リポタンパク質の LDL と HDL(10 ペ
ージ)には標識となる Apo がくっ付いています。Apo は標識の役割だけでなく、細胞膜への
コレステロールなど脂質の輸送や脂質代謝の活性化にも関与し、ニューロンの神経突起の
伸長やシナプスの形成中の末端にコレステロールを供給する役割があります 430,431)。Apo は、
その働き方や働く場所によって ApoA から ApoE まで 5 種類あり 432)、個々人は両親から受
け継いだ ApoE2、ApoE3、ApoE4 の中の 2 種類を持っていいます。つまり、血液型に A 型
(A/A、A/O)
、B 型(B/B、B/O)、AB 型(A/B)
、O 型(O/O)があるように、ApoE にも
ApoE2/ApoE2 、 ApoE2/ApoE3 、 ApoE3/ApoE3 、 ApoE3/ApoE4 、 ApoE2/ApoE4 、
ApoE4/ApoE4 の組合せがあるのです。この組み合わせで、ApoE4 が多いほど、つまり、
ApoE4/ApoE4、ApoE4/ApoE3(あるいは ApoE4/ApoE2)、Apo4 がない組合せの順でβア
ミロイドの脳内集積を増加させると報告されています
433) 434)。また疫学調査でも、健常人
で ApoE4 遺伝子を持っている人は 10% 程度であるのに対し、AD 患者では 30% 以上で
あったとのことです 435,436)。ApoE4 は SDAT においてもリスク因子の1つと考えられてい
ます 433,437,438)(表 1)。しかし ApoE4 が多いか少ないかは、あくまでリスクの話で、約 50%
の AD 患者は ApoE4 を持っておらず、FAD の 20%と SDAT の 10%以下しか ApoE4 と関
連付けられなかったとのことです 439,440)。
LDL 受容体メンバー(11 ページ)は、LDL や DHL などのリポタンパク質に結合した ApoE
も感受します。また、LDL 受容体メンバーは脳関門(BBB)(9 ページ)に機能し、脂質異常
61
ANM176®が認知症の予防と改善に役立つ可能性について
表 1.アポリポタンパク質遺伝子の組み合わせと頻度(人)
石浦正一「タンパク質の反乱」講談社
対立遺伝子組合せ
白人
日本人
E2 /E2
E2 /E3 E3 /E3
1998
E2 /E4
E3 /E4
E4 /E4
症例数
通常人
0.4
14.3
61.5
2.8
19.3
1.8
834
AD 患者
0.2
2.6
36.8
2.9
45.4
12.1
546
通常人
0.1
7.6
74.5
1.5
15.7
0.6
866
AD 患者
0.4
4.8
48.1
0
37.7
9.1
231
症(高脂血症)441)や AD23)など様々な病気にも関係しています。βアミロイドやプリオン
442,443)の排泄には、LDL
受容体メンバーの LRP1 と ApoE2 あるいは ApoE3 が複合体を形
成することが必要です 29,444)。また、様々な物質と結合した LRP1 は、結合した物質に応じ
たシグナルを細胞内に伝えます
445) 。リポタンパク質-βアミロイド複合体と結合した
ApoE2 と ApoE3 は、LRP1 によって BBB を通過し、脳髄液(CSF)中に流出しますが 444)、
ApoE4 は ApoE2 と ApoE3 と比較してβアミロイドの BBB 通過能力が低いことが示され
ています 440)。このことが、ApoE4 が AD のリスクファクターとなっていると考えられてい
ます 446)。
ApoE の遺伝子は DNA の塩基の 1 つが異なる一塩基多型(SNP)(19 ページ)です。ヒト
の SNP が、インスリンが活性に関与している Akt と FoxO の遺伝子(DNA)や 447,448)ある
いはインスリン抵抗性を低下させる脂肪細胞由来ホルモンのアディポネクチン*遺伝子
449)
でも見つかっています。このアディポネクチン遺伝子の多型(19 ページ)がインスリン抵抗性
と糖尿病のリスクと関係すると報告されています 449,450)。
*アディポネクチン(adiponectin)によって脂肪酸の燃焼が増加し組織内中性脂肪含量が低下しま
す 450)。
また、βアミロイドの分解に関与する酵素の NEP451)、ACE 452,453)、エンドセリン-1
454)、カルパイン(CAPN10)455)、インスリン分解酵素(IDE)456)、カテプシン
D 457)も
遺伝子多型であることが知られています。これらβアミロイドの分解に関連する遺伝子多
型の存在が、高齢化に伴うβアミロイド濃度の上昇と個人による濃度のバラつき 247)に関
係していると考えられています。βアミロイド濃度は、高齢になると上昇するだけでな
く、個人によって極端にバラつくようになり、特に Aβ42 は低い人と高い人の間に 3 倍以
上のバラつきが見られるようになると報告されており 223,247)、個人によるβアミロイド分
解酵素や解消の遺伝子多型による差が影響するのでしょう。
17 番染色体に遺伝子がある微小管関連タンパク質(MAP)のタウ、DNMBP もリス
ク遺伝子として報告されています 429)。さらに、Nrf2 にある 1 ヵ所の SNP は Nrf2 タン
パク質のレベルに著しく影響し、ヒトで酸化剤が急性肺障害(acute lung injury)を引き
起こすリスクと関係していると考えられています 458)。以上の他にも、βアミロイドの分
泌に遺伝的原因のない SDAT の発症リスクに関係している遺伝子多型があり、様々な個
性の差と外的な状況が関与していると考えられていることから AD は老年性の多因子疾患
であるとも言われています。
62
ANM176®が認知症の予防と改善に役立つ可能性について
(βアミロイドがアルツハイマー病を発症させる仕組み)
分泌直後のβアミロイド・モノマーには神経毒性はなく 286)、高齢でなくても、アルツハ
イマー病(AD)でなくても、脳内には一定量のβアミロイドが存在し 247)、適正濃度のβア
ミロイドは正常なニューロンの働きに役割があると言われています 278,279)(46 ページ「βアミ
ロイドの分泌と APP の役割」)
。βアミロイドの分泌に関連する遺伝子に変異がなくても、高齢
者の脳内には主にβアミロイドが沈着した老人斑
222,459,460)や目のレンズタンパク質には終
末糖化産物(AGE)の蓄積 419)などタンパク質をコアとした沈着物が増加します。これら沈
着物の増加は、一般に、老化現象の 1 つと捉えられています。
βアミロイドを分解する代表的な酵素と考えられている NEP は、主にニューロンの 364)、
前シナプス膜上で 365)、βアミロイド・モノマーを分解すると考えられます。NEP は神経伝
達を行うニューロンの前シナプスで Aβ40 と Aβ42(47 ページ)の両方を分解しますが
461)、
Aβ42 より Aβ40 の方を効率よく分解します 462) (54 ページ)。NEP は、加齢によって低下す
ると考えられていますが 368,463)、NEP の活性を増強するホルモンの 1 種であるソマトスタ
チン 369)の脳内量も、年齢によって減少すると報告されています 369,464,465)。
一方、脳から循環液へのβアミロイドの排泄を促進する LRP1(11 ページ)は高齢者と AD
患者の脳で減少しています 242,243)。また、循環液から脳内へのβアミロイド流入に寄与する
受容体 RAGE*466,467)は高齢や AD 患者で増加します 466,468)。
*RAGE はスカベンジャー受容体(scavenger receptor;SR)の仲間で、SR は様々な変性蛋白、細菌
から放出される毒性物質などとも結合し、BBB の通過に関与しています。
脳髄液(CSF)中に検出されるβアミロイドの大部分は 40 個のアミノ酸から成る Aβ40
です 469,470)。健常者と AD 患者の Aβ40 と Aβ42 の比較で、AD では脳内の Aβ42 濃度が健
常者より高いにもかかわらず、CSF 中の Aβ42 が低下し 236,471)、Aβ42 に対する Aβ40 の比
率が健常者より上昇します 236,471)。この理由は、AD 患者では BBB における LRP1 量が同
年代のヒトに比べて減少し、BBB 能力も低下している
244,245)からと考えられています(47
ページ)。また、LRP1 との親和性が Aβ40 より Aβ42 の方が低いため
472)、Aβ40 が
Aβ42 よ
り効率よく BBB を透過し、CSF 中の Aβ42 が低下します 472)。神経系細胞が直接的に接し
ている間質液(9 ページ)から BBB を介した循環血液中への Aβ40 輸送は、LRP1 のリガンド
である ApoE の遺伝子を欠損させたマウスにおいて低下することが示されています 243,444)。
ApoE が AD のリスクであることについても、ApoE の相同体によって LRP1 の透過能力
に差があることによると考えられています。
熱ショックタンパク質(Hsp)の ApoJ は膜組織の再生やリン脂質の輸送等、多くの生
理的機能と関係していますが、βアミロイド・オリゴマー形成の促進作用もあります(51
ページ)。 BBB を経由したβアミロイドの脳外排出に関係している LRP1243)の相同体
LRP2*は、脳髄液(CSF)から循環血液への Aβ40 の輸送に関与しています 473)。
*LRP2 の機能は LRP1 と似ており、甲状腺ホルモン・チロキシンのトランスポーターであるトランス
サイレチン 474)、インスリン様成長因子 1(IGF1)475)あるいはレプチン 476)などを細胞内に取り込む
働きがあります。
63
ANM176®が認知症の予防と改善に役立つ可能性について
LRP2 は ApoJ と複合体を形成した Aβ40 の細胞内への取り込みに関与し 473)、βアミロイ
ドの BBB を経由した脳内からの解消は ApoJ によって制御されます 477,478)。このことによ
り、βアミロイドが BBB を通過するにはβアミロイドの構造が関係し、解消されなかっ
た Aβ42 がβアミロイド・オリゴマーの形成に関係している可能性が考えられます。ま
た、ApoJ が関与して形成されたβアミロイド・オリゴマーは MTT 法による毒性(31 ペー
ジ)も示します 305)。
Aβ42 の NEP 分解能は Aβ40 より低く 462)(54 ページ)、上で述べたように、LRP1 の BBB
通過能力は Aβ40 に比較して Aβ42 が低いこと 440)、および、Aβ40 の BBB 透過に関与する
ApoJ と LRP2 複合体は、Aβ42 の透過には関与していないこと、の理由で、AD 脳におけ
る Aβ40 に対する Aβ42 の比率が上昇すること、が説明できます。また、分泌されたばかり
のβアミロイドを分解する NEP だけでなく、インスリン分解酵素(IDE)456)、カテプシン
D
457)などβアミロイドの分解に関わっている多くの酵素(53
ページ)や BBB 透過によるβ
アミロイドの排出に関わっている LRP1 そのものが遺伝子多型である
242,479)こと、によっ
ても個人によるβアミロイド濃度のバラつきが説明できます。ニューロンのシナプスで
APP から切り出されたβアミロイドは NEP によって適正な濃度に制御されていると考え
られます。一方、遺伝と関係がある家族性アルツハイマー病(FAD)の一部では、βアミロ
イドに対する NEP の酵素分解能に抵抗性があることが示されています 480,481)。
シナプス後膜にある足場タンパク質 PSD-95 中の PDZ ドメイン(38 ページ)は様々なタン
パク質にあるβシート構造に結合します 183,184)。PSD-95 中の PDZ ドメインは、NMDA 型
と AMPA 型のグルタミン酸受容体や多くの神経作用に関連するタンパク質 175,176)にあるβ
シート構造が結合する場 183,184)として機能しますが 482,483)(38 ページ注**)、AD 型変異マウ
スやβアミロイド・オリゴマーによるシナプス障害を起こした大脳ニューロンで PSD-95 の
レベルが減少したと報告されています 484,485)。また、NMDA 型受容体の働きを抑制すると、
βアミロイドが誘導したスパイン減少を抑制し 486)、βアミロイド神経毒性に対してニュー
ロンを保護できると報告されています 487)。一方、βアミロイド・オリゴマーにある PDZ ド
メインで結合して濃度依存的にβアミロイド・オリゴマーの毒性を著しく弱めることがで
きる Hsp の Omi は
303,488)(51
ページ)、βアミロイド・オリゴマーとは結合しますが、βア
ミロイド・モノマーやβアミロイドとアミノ酸が逆配列のタンパク質には殆ど結合しませ
488)。これらのことから、βアミロイド・オリゴマーは、NMDA
ん
型受容体と PSD-95 の
PDZ ドメインに影響して NMDA 型受容体の働きに悪影響を及ぼし、神経作用を障害する
と考えられています 279)。老化細胞では、タンパク質の品質管理に関連するシャペロン分子
の著しい減少が見られ
424,489–491)、これらシャペロン分子の
DNA からの読み取りに関与す
る転写因子 HSF1 の発現減少や転写能力の減退が起こっていると報告されています 492,493)。
βアミロイドの分泌に直接関係する遺伝子の変異がなくても、高齢による NEP 分解能
の低下と個人による NEP 活性のバラつきによってβアミロイドの増加を招き、シナプス
後膜のβアミロイド、その中でも Aβ42 が増加し、ApoJ など Hsp 関連のタンパク質品質
64
ANM176®が認知症の予防と改善に役立つ可能性について
管理などが加わって、増加したβアミロイドはβアミロイド・オリゴマーを形成し、βア
ミロイド・オリゴマー中のβシート構造が樹状突起の後シナプス肥厚部(PSD)(図 27)
にある足場タンパク質 PSD-95 の PDZ ドメインに結合して NMDA 型受容体などに影響し
(48 ページ「βアミロイドの神経毒性作用」)
、シナプスの活動を抑制し
278,494)、健忘や
MCI ある
いは AD の認知機能障害の一因となっているのかも知れません。
上に述べたようにβシート構造を含むβアミロイド・オリゴマーはシナプス後膜で直接、
神経伝達に関与する受容体を障害すると考えられますが、線維状βアミロイドは膜受容体
が感知します。βアミロイド・オリゴマーと線維状βアミロイドによる神経毒性は細胞内シ
グナリング経路が異なると言われています 495)。βアミロイド・オリゴマーは AD 患者の海
馬のシナプスで観察されましたが 496,497)、アミロイドを含む老人斑が見られた認知機能は正
常であった高齢者のシナプスにはβアミロイド・オリゴマーは観察されなかったと報告さ
れています 498)。
細胞外に分泌されたβアミロイドは、ミクログリアに取り込まれて分解されるか、アスト
ロサイトが関与する BBB によって脳外に排出されます。老化細胞では、プロテアソーム(図
12)におけるタンパク質分解経路の変調など細胞のタンパク質品質管理システム全般の低
下が起こっていると報告されています 499–501)。また、βアミロイドを取り込むミクログリア
の活力も高齢になると低下します 502)。さらに、脳からのβアミロイド排出のシステム能力
が高齢者や AD 患者で低下し 23,503)、脳内にβアミロイド、特に Aβ42 が蓄積します。
βアミロイド凝集体は、ミクログリアの細胞膜にある MAC1L などインテグリン(15 ペー
ジ注)が関与するβアミロイド複合受容体(52 ページ)を経て G タンパク質を活性化します
103,325)。ニューロンネットワークに長期記憶が組み込まれていると考えられていますが(8
ページ)、このニューロンネットワークの形成には G タンパク質が関係していると考えられ
ます。膜機能に関与するインテグリンによる G タンパク質 Ras ファミリー(40 ページ)の
活性化は 34)、膜の突起形成プロセスと関係すると考えられています 162)。また G タンパク
質 Rab ファミリーの一員である Rab11 が軸索先端へのインテグリンのβ1 サブユニット
の輸送を制御することで、海馬ニューロンの軸索伸長を制御すると報告されています
504)。様々な刺激を感知する
G タンパク質共役受容体(GPCR)(15 ページ)であるムスカリ
ン性アセチルコリン受容体(mAChR)、アドレナリン受容体、ドーパミン受容体などの
神経伝達に関与する受容体はアクチンや微小管(MT)による細胞骨格の動きに寄与して
います 199)。
タウ 216,217)や CRMP2200)(図 29)など微小管結合タンパク質(MAP)はニューロンに
ある MT の動きを調節します(41 ページ)。AD 脳に見られるタウのリン酸化の前に、βアミ
ロイド・オリゴマーによって、ニューロンでタウの誤局在化と樹状突起で MT の崩壊が観
察され、スパインの脱落、ミトコンドリアと神経線維の変異が伴ったとのことです 241)。
βアミロイドが神経作用を害すると考えられるもう一つのルートは、炎症による影響で
す。βアミロイドが増えただけでは AD は発症しないと考えられています。事実、マウスの
65
ANM176®が認知症の予防と改善に役立つ可能性について
脳内に注入したβアミロイドが学習力を低下させる試験では、実際の高齢者の脳内にある
レベル濃度のβアミロイドが動物の記憶や学習力を低下させましたが、AD に見られるよう
な神経細胞の死を示してはいませんでした 234)。また、動物の脳内に注入したβアミロイド
が、例えオリゴマーであっても、βアミロイドの影響は数日間で回復することが可能です
234,288,505)。つまり、動物実験で記憶や学習力を低下させるβアミロイドの作用は
AD の全て
を現してはいません。健忘と AD は異なり、実際の AD 患者では、まず認知機能の低下が先
行しますが、その後は徐々に神経細胞が死んで脳の萎縮が進行します。脳内に注入したβア
ミロイドよって、海馬のアストロサイトで炎症性サイトカイン IL-1βが分泌されることが
示されています 234)。脳の神経炎症と深く関係する IL1α、IL1β、IL1R antagonist protein
(Il-1Ra)の全てには ApoE と同じように著しい多型(19 ページ)があります 506–509)。
また、活性化したインテグリン MAC1 が関係するβアミロイド複合受容体が細胞内にシ
グナリングし、IL-1βなどの分泌やストレス応答が導かれることが報告されています 41)。
βアミロイドが脳内で引き起こした炎症が神経作用に影響すると考えると、動物の脳内に
注入したβアミロイドは 30 分以内に NEP によって分解されましたが 348)、その後もβア
ミロイドの影響が数日間も残っていたこと 234,288,505)を説明できます。動物試験ではβアミ
ロイドはニューロンの死が起こらないのに、なぜ AD ではニューロンの死が進行するのか
も、炎症の慢性化がアポトーシスにつながっていることから説明できます。
小胞体ストレス応答(UPR)は AD の脳内でも起こっていると報告されています 510)。ま
た、UPR センサーの GRP78(26 ページ)を過発現させるとβアミロイドの分泌を抑制するこ
とが示されています
えられています
511,512)。UPR
513)。AD
が増強し長引くと、遂にはアポトーシスを誘発すると考
脳では p38 のリン酸化(活性化)が増加し、Akt の発現が抑制さ
れていることが報告されています 201,514)。ニューロンネットワークはストレス応答ともつな
がっていると考えられています 162,163)。
年を取ると、個人によって、外部からの細菌や毒物などの攻撃や内因性のストレスに対し
て感受性が高くなり
491)、UPR
が起こりやすくなり
515)、熱ストレス応答 516)や酸化ストレ
ス応答 491)などのストレス応答が全般にわたって低下します。酸化ストレス応答に導くシグ
ナリング経路をβアミロイド・オリゴマーが活性化させ 517,518)、AD 脳の障害に関係してい
ると考えられています 519)。
非ステロイド抗炎症剤(NSAID)は AD の発病や進行を遅らせる可能性を示す報告が多
数あり、AD の進行抑制に NSAID が有効ではないかと考えられています 520)。NSAID は細
胞膜のホスファチジルイノシトールに結合しているアラキドン酸からプロスタグランジン、
トロンボキサンあるいはロイコトリエンなどの炎症メディエーターを合成する COX
(図 22)
の酵素活性を抑制します。NSAID は炎症の抑制だけでなく、アポトーシス抑制や癌の抑制
など多くの生理作用に関与しています 93,521)。NSAID には低分子量 G タンパク質 Rho とそ
のエフェクターである ROCK
(Rho-associated kinase)の活性を阻害する作用もあり 522,523)、
COX 活性に非依存的なメカニズムで AD に有効性があるとも考えられています 524)。
66
ANM176®が認知症の予防と改善に役立つ可能性について
ミクログリアでβアミロイド複合受容体に関連する MAC1L は
41)、βアミロイドだけで
なく、パーキンソン病の原因タンパク質α-シヌクレインによる神経毒性の誘導にも重要な
役割を果たしていると報告されています 525,526)。AD に限らず神経変性疾患では、原因タン
パク質が異なり、損傷される脳の部位が異なっていても、クロスβシート構造によるタンパ
ク質同士の集合体が細胞のストレス応答をひき起こす引き金となり、脳内炎症を引き起こ
し、場合によっては損傷された部位のニューロン死を招くと考えられます。脳内炎症の慢性
化がニューロン死を引き起こし、AD と関係していると考えられています
89,527)。また、脳
内炎症は AD に限りません。ハンチントン病や脊髄小脳失調症などのポリグルタミン病の
原因となるポリグルタミン酸は ER ストレスから IRE1-ASK-JNK 経路が関与する可能
性が考えられています 423)。また、ニューロン死は AD だけでなく全ての神経変性疾患に同
じことが言えます。神経変性疾患で共通している現象は脳内炎症と考えられています。
アルツハイマー病は生活習慣病か?
糖尿病(DM)、高血圧、肥満、高脂血症などは「成人病」と言われています。しかし、こ
れらの病気は若年者でも多発し、その発病に生活や環境の変化あるいは遺伝要因も関係し
ていると考えられるようになったことから、近年では「生活習慣病」とも呼ばれるようにな
りました。これらの生活習慣病は急増しています。例えば、DM が強く疑われる人は 740 万
人、DM の可能性を否定できない予備軍は 880 万人と推定されており(厚生労働省が 2002
年実態調査)
、最近の 5 年間だけでも予備軍を含めた DM 関連者は 250 万人も増加してい
ます
144)。花粉症やアトピー性皮膚炎などアレルギー疾患も、1970
たのが、2000 年代には 4 人に 3 人に増加したと言われています
年代は 4 人に 1 人だっ
528)。これらのアレルギー
疾患も、アレルギーを引き起こす原因物質だけでなく、遺伝的要因や環境や食べ物や生活の
変化も発症に関係していると考えられています。5 ページで述べたようにアルツハイマー病
(AD)の推定患者も毎年急増しており、2015 年の厚生労働省による報告では、2012 年の
312 万人から、2025 年には倍以上の 700 万人になると推計されています。
一方、疾患の患者患者が減少している生活習慣病もあります。その 1 つは高血圧症(血圧
の収縮期 140 mmHg 以上もしくは拡張期 90mmHg 以上)です。厚生労働省による 2000 年
の循環器疾患基礎調査によれば,30 歳以上の日本人のうち男性 51.7%,女性 39.7%が高血
圧ですが、40~74 歳では 1960 年代前半がピークで、その後は低下しています。この理由
は、食事減塩や降圧剤の使用が普及したことによると言われています。高血圧症のように原
因の 1 つに食事の習慣があって、その習慣を変えることによって発症率を低下させること
ができた生活習慣病は稀で、生活習慣病と言われながら、生活のどこを改善したら良いのか
分かっていない病気か多いのです。
最近、高齢化率を上回って増加している AD と DM には何らかの共通した背景があるの
ではないかと考えられています。AD 患者の 80%は DM かその前兆が見られ 529)、高齢者の
DM の一部には認知機能の低下が高頻度に見られると報告されているからです
530,531) 。
67
ANM176®が認知症の予防と改善に役立つ可能性について
イギリスで行われた平均年齢 56 歳の男女 5,647 人を調査した研究では、DM の疑いがあっ
て、かつ、認知症が強く疑われる人は、健常者 1 人に対し、男では 2.45 人、女では 1.83 人
であったとのことです 532)。また、DM 患者は健常者の 2 倍の頻度で AD に進行するといわ
れています 533)。日本においても福岡県久山町で行われた調査で、DM と AD の疑いがある
人には相関があることが示されています 531)。
AD と DM には、近年患者が増加していること以外にも以下のことが共通しています。
1.成人や高齢になってから急増する
2.発症機構に不明なところが多く、根本的な治療方法がない
3.発症に主因以外の要因も無視できない
4. DM も AD も約 95%は遺伝とは関係しない
最近、DM に関係するインスリンは、AD の発症にも関係していると考えられています
519,534)。以下では、まず
DM の概要について述べ、その後は、AD と DM の細胞内の生理作
用について掘り下げてみたいと思います。
(糖尿病とは)
尿に一定量以上の糖が常に検出されるために糖尿病(DM)と言われますが、通常 DM と
は、血液中のグルコース濃度が病的に高い糖尿病型高血糖症のことです。糖とはグルコース
のことで、血液中のグルコースは電解質や水分と共に、いったんは腎臓で濾過され原尿中に
出ますが、そのほとんどが再吸収され血液中へ戻ります。しかし、原尿中のグルコースが
200 mg/dL(100cc に 20g の濃度で、コーヒーカップに小さじ 2 つ分の砂糖を入れたぐらい
の甘さです)を越すと再吸収能を超えたグルコースが尿の中に溢れ出ることになります。食
事直後は、健常人でも尿に糖が出ることがありますが、一過性です。また、腎臓機能障害に
よりグルコースの再吸収が十分で無い腎性糖尿病では血糖値は正常です。
検査前日の夕食後から検査当日の朝まで食事を摂らない状態で測定した血糖値(空腹時
血糖値)が 126 mg/dL 以上もしくは食後の時間経過にこだわらずに測定した血糖値(随時
血糖値)が 200 mg/dL 以上であれば、
「糖尿病型」高血糖症と診断されます。空腹時血糖値
が 110 mg/dL 未満、随時血糖値が 140 mg/dL 未満であれば「正常型」
、
「糖尿病型」にも「正
常型」にもあてはまらないものが「境界型」です。 DM が長引くと色々な臓器に障害が生
じます。
動物は食べ物から様々な物質を吸収しますが、澱粉などの糖類の一部は腸内で分解され
て主にグルコースとして吸収されて血流に乗ります。血液中のグルコース濃度(血糖値)が
高くなると肝臓や骨格筋などの細胞内にグルコースが取り込まれます。細胞内に取り込ま
れたグルコースは、生体成分の原料やエネルギーとして利用されるかグルコースが連なっ
たグリコーゲンとなって細胞内に蓄えられて血糖値が低下します 144)。
DM には、膵臓から分泌されるインスリンの障害が深く関係しています。膵臓は肝臓の
下、十二指腸の横にあって(図 46)、食べ物の吸収や吸収した後の元栓のように、最も根源
68
ANM176®が認知症の予防と改善に役立つ可能性について
のグルコースの制御に関わっていま
す。膵臓の中には腸内で食物の消化に
関与する酵素類を分泌する外分泌細胞
と、外分泌細胞に取り囲まれて、あたか
も島のように分散している内分泌細胞
があります。この内分泌細胞は発見者
パウル・ランゲルハンスの名前にちな
んでランゲルハンス島(以下では膵ラ
島といいます)と呼ばれています。膵ラ
島にはα、β、δと呼ばれる 3 種類の
細胞があります(図 47)
。この各細胞か
ら分泌されるホルモンは異なってお
り、β細胞からは血糖値を下げる作用
があるインスリンとアミリン、α細胞
からはインスリンと反対に血糖値を上
図 46.膵臓の位置
[中学生の知識で分かる私たちの体の仕組み] 秦順一ホームペ
ージより
げる作用があるグルカゴン、
δ細胞からはインスリンの
働きを増強する作用がある
ソマトスタチンが分泌され
ます
144)。ソマトスタチンは
視床下部や消化管からも分
泌され、中枢神経系では神経
外分泌部の腺房細胞
ペプチドの一種として成長
ホルモンの分泌量を抑制し、
赤血球
消化管では胃酸や胃液の分
泌量を調整しています。
図 47.膵臓断面
(インスリンシグナリングとグリコーゲン合成)
膵ラ島β細胞から分泌されたインスリンは血糖値、すなわちグルコース濃度を低下させ
ます。血中に分泌されたインスリンは、まず、様々な細胞の細胞膜にあるインスリン受容体
(IR)に結合します。IR はチロシンキナーゼ型受容体(14 ページ)の一種で、インスリンが
結合すると構造が変化して、IR 内のチロシンキナーゼが活性化し 535)、インスリン受容体基
質(以下では insulin receptor substrates から IRS と表記します)中のアミノ酸であるチ
ロシンをリン酸化して活性化します 536)。IR によって活性化した IRS は PI3K を活性化し、
活性化した PI3K は細胞膜にあるイノシトールリン脂質に結合した PIP2 をリン酸化して
PIP3 に変換してイノシトールリン脂質から切り離します 537,538)(図 8)
(17 ページ)。PI3K は
69
ANM176®が認知症の予防と改善に役立つ可能性について
G タンパク質によっても活
性化されます。このように
して生成した PIP3 は、直接
もしくは PDK1 の活性化を
経て Akt を活性化します
(17 ページ)。また Akt は、
インスリンシグナル経路や
G タンパク質によって活性
化された PDK1 経由だけで
なく、Akt の自己リン酸化
によっても活性化されます
45,46)。活性化した
Akt はグ
図 48.肝臓におけるグルコースの取り込み
144)
リコーゲン合成酵素キナーゼ 3β(以下では glycogen synthase kinase 3βから GSK3βと
表記します)をリン酸化します(図 48)。
血中から細胞に取り込まれたグルコースは、平常ではグリコーゲン合成酵素(以下では
Glycogen synthetase から GS と表記します)によってグリコーゲンに合成されて細胞内に
蓄えられます(図 48)。一方、GSK3βは GS をリン酸化して不活性にしてグリコーゲン合
成を抑制します 539,540)。Akt によって GSK3βがリン酸化されると GSK3βの活性が抑制さ
れ、GSK3βの GS に対するリン酸が抑制されて GS が活性化し 539,540)、グリコーゲンの合
成が始まります。
(インスリンシグナリングの役割)
インスリンを分泌するのは膵ラ島のβ細胞だけですが、インスリンと似た作用をするイ
ンスリン様成長因子(IGF)は様々な細胞が分泌します。このインスリンもしくは IGF に
よって、インスリン受容体→IRS→PI3K→PIP3→PDK1 経路で活性化した Akt が様々な作
用をする経路をインスリンシグナリング経路と呼びます 541)。
平常の細胞内では、グリコーゲン合成酵素キナーゼ 3β(GSK3β)がグリコーゲン合
成酵素(GS)の活性を抑制し、グルコースからのグリコーゲン合成を抑制していますが、
血中のグルコース濃度が高くなると、インスリンシグナル経路によって活性化した Akt
は、GSK3βを抑制して GS を活性化し、グルコースからグリコーゲンへの合成を促進し
ます。このように、栄養状態になるとインスリンシグナリング経路でグリコーゲンが細胞
内に蓄積されるようになり、他の細胞内機構も制御されて、細胞は生長のパターン(24 ペー
ジ)に切り替わります。GSK3βは哺乳類組織の細胞に幅広く分布し 542,543)、GS 以外に
も、タンパク質合成に関わる転写因子の Nrf2 や C/EBFαあるいは真核生物翻訳開始因子
2B(elF2B)、細胞やシナプスの骨格に関与するβカテニン(catenin)(図 49)あるいは
微小管(MT)の重合を促進する微小管結合タンパク質(MAP)の CRMP2205) (図 31)
70
ANM176®が認知症の予防と改善に役立つ可能性について
や MAP1B などのリン酸化に関与し
201,205)、細胞分裂サイクル、遺伝子転写、細
胞内骨格体の集積、神経軸索形成などを抑
制的に制御します 544,545)。一方、インスリ
ンシグナリング経路は、グルコース代謝に
関係するだけでなく、GSK3βの全般的な
生長抑制作用を停止させます。また、イン
スリンシグナリング経路は、様々なストレ
ス応答的な機能やアポトーシスを抑制し、
細胞分裂サイクル、細胞内骨格体の集積な
ど数多くの細胞の生長の機能を促進して、
細胞生長を促進する役割を果たしています
545)。
図 49.接着結合と細胞膜裏打ちタンパク質
インスリンシグナリングはストレス応答
にも関係します
接着結合(アドヘレンス・ジャンクション)は、細胞細胞間接着および細胞-基質間接着の接着装置の 1 つで
す。細胞接着を担う膜タンパク質には、接着結合の形
成と維持に関与するカドヘリン、細胞基質接着に関わ
るインテグリンが代表的な細胞接着分子で、その細胞
内ドメインで細胞骨格と結合しており、アクチンなど
を動員した接着装置を構築しています。カドヘリンは
まずβ-カテニンと相互作用します。ビンキュリンはカ
テニンの結合分子です。(図は WIKIPEDIA より)
546) 。GSK3βは転写因子
Nrf2 を直接リン酸化することによって Nrf2
の細胞核内への移行を抑制しています
547)。
インスリンシグナリング経路によって活性
化した Akt は Nrf2 による転写を不活性にし
ていた GSK3βをリン酸化して Nrf2 が細胞核に移動し、ARE エレメントの関連する酸化
分解や分解物の解毒に関連する第Ⅰ、Ⅱ相系のタンパク質の DNA からの転写が始まります
548)(28
ページ)
。Nrf2 は酸化ストレスに限らず様々なストレスによって核内に移行し、第Ⅰ、
Ⅱ相系のタンパク質を誘導してストレス応答を統一的に制御します 69,122)。
インスリンシグナリング経路によって活性化した Akt は mTOR の複合体 mTORC1 を
活性化してオートファジーを抑制します(57 ページ)。このオートファジーに対する Akt の
作用は、グルコースからグリコーゲンを作り出すグリコーゲン合成の場合と似ています。
オートファジーは細胞のエネルギーが不足した異常な事態でアミノ酸の供給に機能してお
り、Akt がオートファジーを抑制する作用とグリコーゲン合成を促進する作用が連動して
いるのは当然のことと考えられます。しかし、Akt によるオートファジーの制御は単純で
はなく、mTOR の複合体には mTORC1 と mTORC2 があり、Akt は mTORC1 を活性化
する一方
mTORC2 は Akt の活性を抑制しており、Akt によるオートファジーの抑制は
微妙に制御されていると考えられます 46)。
(インスリン抵抗性)
糖尿病(DM)には 1 型と 2 型があることは良く知られています。1 型 DM は膵ラ島のβ
細胞に異常があるなどによって、正常にインスリンが分泌されないことが原因です。2 型
71
ANM176®が認知症の予防と改善に役立つ可能性について
DM は、インスリン分泌能力に異常はありま
せんがインスリンの分泌量が減少する低イ
ンスリン症と、インスリンの分泌量はさほど
減少していないにもかかわらずインスリン
の効果が発揮されていない高インスリン症
があり
144)、どちらにせよ、インスリンの効
果が発揮されないことが原因です。日本人で
740 万人いると言われている DM 患者の約
95%は 2 型で、しかも低インスリン症が過半
図 50.日本人と欧米人のインスリン抵抗性
日本人は空腹時血糖値 100mg/dL を越える頃から徐々
に糖負荷後の平均インスリン分泌量が低下していきま
すが、米国白人ではこのピークが 120mg/dL 前後にシ
フトしており、全体的な分泌量も日本人の倍以上です。
清野裕 [糖尿病の新しい概念] 最新医学 (1995)
50:639—45
ですが、欧米人では高インスリン症が多いと
のことです 549)(図 50)。
2 型 DM において、膵ラ島β細胞が正常
であっても血中の グルコース 濃度が上昇
する理由はインスリンが正常に働いていないからで、この状態はインスリン抵抗性と呼
ばれています。インスリン抵抗性は高インスリン症と低インスリン症の両方に関係しま
す。高インスリン症では、インスリンの効果が発揮されないため血糖値が下がらず、その結
果、ますますインスリンが分泌されます。一方、低インスリン症では、ストレスなどが膵ラ
島β細胞に影響してインスリンの分泌量が低下し、その結果、血糖値が上昇します 144)。
血糖値が上昇すると膵ラ島β細胞から分泌されるのはインスリンだけでなく、インスリ
ンと共に血糖値を下げる作用があるホルモンのアミリンも分泌されます 550,551)。インスリ
ン抵抗性によってインスリンだけでなく過剰なアミリンも分泌させることになり 552)、過
剰に分泌されたアミリンは線維状アミリンになります。線維アミリンは膵ラ島β細胞を消
滅させ、2 型 DM を進行させると言われています 553)。1 型 DM ではインスリンもアミリ
ンも不足するため線維状アミリンは形成されません。そのため、線維状アミリンの存在は
2 型 DM の判定に利用されています。
膵ラ島β細胞は小胞体(ER)のストレスセンサーである IREl や PERK(26 ペーシ)を豊
富に発現しています。膵ラ島β細胞で ER ストレス応答(UPR)が起こると PERK が
RNA からのタンパク質の翻訳を抑制しますが、この抑制されるタンパク質にはインスリ
ンも含まれています。また、IRE1 は UPR に関連する様々なタンパク質の DNA からの転
写を促進します 58)。こうして、タンパク質の翻訳を抑制する PERK はブレーキの役割
を、タンパク質の転写を促進する IRE1 はアクセルの役割を果たし、インスリン合成が調
節されます 55,56,554)。膵ラ島β細胞はインスリンを分泌するために特化された細胞で、多量
のインスリンを合成しているため、ER におけるタンパク質合成の負荷、すなわち UPR
が、通常から高い状態に置かれています 554)。そのため膵ラ島β細胞は様々なストレスに
脆弱で 555,556)、ブレーキ役の PERK とアクセル役の IRE1 とのバランスが崩れるとβ細胞
の死に繋がることもあり得ると考えられています 557)。このように UPR はインスリン合成
72
ANM176®が認知症の予防と改善に役立つ可能性について
や膵ラ島β細胞の生存に深く関わり 554)、2 型 DM の発症に重要な役割を演じていると言
われています 556)。
ストレスキナーゼの JNK は(16 ページ)、インスリン抵抗性に関与していると考えられて
います。JNK を削除したマウスの実験で、TNF-α、IL-6 など炎症性サイトカイン(13 ペー
ジ注)の発現が抑制され、インスリン抵抗性や 2 型 DM を防げました
558,559)。このことは、
JNK が炎症を引き起こし、炎症がインスリン抵抗性と関連することを示唆しています。
アルツハイマー病でのβアミロイドと 2 型 DM でのアミリンは異なるタンパク質ですが、
過剰になると両者とも線維状になり、また、その結果起こる酸化ストレスがそれぞれの発病
に関係していると考えられています 560)。
(インスリンは脳でも働いている)
脳の重量は体重の 2.5%程度ですが体全体の約 3 分の 2 のグルコースを消費すると言われ
ています 561)。しかし、脳にはグリコーゲンのようなグルコース貯蔵機構がなくグルコース
の供給は血流に頼っているため、血液の約 15%が脳に供給されます。脳には脳関門(BBB)
と呼ばれる血液関門があって、血流に乗ってくる物質が脳内に入ることを制限しています
13)。 BBB
では細胞膜上のグルコーストランスポーターである GLUT1 と GLUT3 が関与し
て(10 ページ)、大量のグルコースが脳に供給されています 562,563)。
グリコーゲン貯蔵機構がない脳ではグリコーゲン合成酵素キナーゼ 3β(GSK3β)を抑
制してグリコーゲン合成酵素(GS)を活性化する必要はありませんが、インスリンシグナ
リング経路は、それ以外の重要な役割を脳で果たしています。体のインスリンは BBB を通
って脳に入ることができますが
564)、脳内でもインスリン 565,566) やインスリン様成長因子
(IGF)565,566)が作られています。脳内のインスリンシグナリング経路は、摂食調節作用 541)、
神経細胞の分化や増殖作用 567,568)、ニューロンの保護 569,570)などの役割を果たしています。
空腹時に空腹感を感じることを摂食欲望と言います。グルコース、インスリン、レプチ
ン(10 ページ注****)など細胞のエネルギー状態を反映する分子によって脳の奥にあ
る視床下部が刺激され、その結果、視床下部から発するシグナリングが脳全体のニューロ
ンネットワークに伝達され摂食欲望が生じます。レプチン受容体やインスリン受容体
(IR)が高密度に分布している弓状核は、視床下部の内側基底部にあって、血中の物質濃
度の変化を検出し(図 51)、摂食欲望を抑制します。このように、インスリンなどが脳で
調整する摂食欲望や体のグルコース状態は神経作用からもバランスが取れるようになって
いると考えられます。摂食欲望が病的な場合は、過食し肥満を引き起こします。
小胞体ストレス応答(UPR)は膵ラ島β細胞の生存に深く関わっていると考えられてい
ますが 571,572)、神経変性疾患においても UPR の防御機構が破綻して神経細胞死に繋がる
と考えられています 151,152)。UPR に関与する PERK の活性化は、タンパク質の翻訳を抑
制するだけでなく 573)、GSK3βを通して 574,575)、ストレス応答に関係する転写因子 NF-κ
B を活性化し炎症を増強しますが 145)、脳においてもインスリンシグナリング経路で
73
ANM176®が認知症の予防と改善に役立つ可能性について
GSK3βを非活性化することがアポト
ーシスの重要な制御要因であることが
小脳顆粒ニューロン 576)や PC12 細胞
および線維芽細胞 577)で示されていま
す。
最近のインスリンシグナリング経路
の研究成果から、βアミロイド神経毒
性とアルツハイマー病(AD)の発症
にインスリンシグナリング経路が関係
している可能性を考察することができ
ます。インスリン受容体(IR)はニュ
ーロンやニューロンの周りにあるグリ
ア細胞(7 ページ)を問わず脳内に幅広
く見られ、特に、記憶・学習や認知機
能に関与している大脳皮質、海馬、嗅
球 578)や摂食欲望を制御している視床
下部に多く見られます 564,579)。海馬ニ
ューロンに結合した Aβオリゴマーが
IR を細胞膜から細胞質に移動さるこ
図 51.弓状核のニューロン活動の制御
レプチンは全身の脂肪細胞で作られ、食欲と代謝の調整のた
めに体脂肪量を脳へ伝えます。弓状核には NPY/AgRP/GAB
と POMC/CART の 2 種類のニューロンがあり、レプチンお
よびインスリンの受容体は、この 2 種類のニューロンにあ
ます。NPY/AgRP/GAB は摂食促進性で、POMC/CART は
抑制性です。NPY/AgRP/GAB は POMC/CART を抑制しま
す。満腹時ではレプチンやインスリンの刺激によって、
POMC/CAR が摂食を抑制性するアルファ・メラノコルチン
刺激ホルモン (α-MSH)を分泌します。
浦野明央 [肥満の神経内分泌学] 本能と煩悩(全 12 回)
Web TOKAI より
とが示されています 580,581)。IR のチロシンキナーゼ活性が阻害されるとβアミロイド・オ
リゴマーがニューロンに結合することをブロックするインスリンの働きが失われることか
ら 581)、βアミロイド・オリゴマーに対するインスリンの防御メカニズムは、βアミロイ
ド・オリゴマーが結合する場所の抑制とそれに引き続く IR の細胞質内への移動、シナプ
スの減少や可塑性の障害の防御に関係しているのではないかと考えられています 519,581)。
AD のモデル動物 582,583)や実験的に膵臓ランゲルハンス島を破壊した動物 584)で、脳のイ
ンスリンシグナリング経路に問題が生じていることが示されています。またインスリンと
同じ働きをする IGF1 は、AD のモデル動物で AD の特徴の 1 つである認知機能障害を弱
めることができます 585)。インスリンシグナリング経路に問題が生じると記憶力が低下
し、ニューロンの LTP(36 ページ)が障害されます 586,587)。
インスリンが NMDA 型受容体によるシナプス伝達を増強することが示されています
588,589)。イノシトールリン脂質から
PIP3 を作る PI3K は(17 ページ)、インスリンや G タン
パク質によって脳内でも増強し 590)、PI3K によって生じた PIP3 がニューロンの樹状突起
や軸索形成の初期に働き 591)、細胞極性、神経細胞の軸索形成 591)などに広く関わっている
と考えられています。このことは、インスリンシグナリング経路がシナプスの樹状突起形
成の起点となり 580,581)、記憶・学習を強化することを示唆しています 583,592)。
インスリンシグナリング経路によって GSK3βが抑制され 574,575)、AD に特徴的なタウ
74
ANM176®が認知症の予防と改善に役立つ可能性について
の過剰リン酸化が阻止されることが示されています 593,594)。G タンパク質 Rho ファミリー
の活性化よって GSK3βが活性化され、活性化した GSK3βは微小管(MT)の重合と安
定化を促進する微小管結合タンパク質(MAP)のタウ 595,596)、CRMP2(図 31)、MAP1B
をリン酸化し 201,205)、軸索伸長を抑制します 203,597)。一方、インスリンシグナリング経路
によって活性化した Akt は GSK3βの活性を抑制して CRMP2、MAP1B のリン酸化を抑
制し、MT の重合・脱重合を調節して軸索伸長を促進させる方向に寄与します 598,599)。以
前から、インスリンシグナリング経路は神経成長因子(NGF)
、ニューロトロフィン 3
(NT-3)、BDNF などの神経栄養因子と同じような働きがあると考えられます。GSK3β
は、神経栄養因子の NT-3 や脳由来神経成長因子(BDNF)によっても阻害され、CRMP2
のリン酸化が抑制されることで軸索伸長が促進されます 205)。
インスリンシグナリング経路によって活性化した Akt は、βアミロイド神経毒性(48 ペ
ージ「βアミロイド神経毒性作用」)に対しても神経細胞を保護し
600,601)、神経細胞の生存を維
持します 576,602)。
インスリンの働きを増強するソマトスタチン(69 ページ)は NEP の活性を増強しますが
369)、NEP
はソマトスタチンを分解できます
367)。一方、膵臓ラ島β細胞からはインスリン
と同じように血糖値を下げる作用があるアミリンが分泌されますが、NEP はアミリンの分
解にも関係しています 367)。これらのことは、NEP の働きは微妙に制御されていることを示
唆しています。事実、NEP の mRNA とタンパク質の量は高齢者や AD 患者で減少してい
ますが 603,604)、一方では NEP の活性は加齢や AD で増加していると報告されています 605)。
このような活性の上昇は、AD に関する他のタンパク質でも観察されており、AD における
ホメオスタシスの乱れや炎症応答が影響しているのではないかと考えられています 606)。
インスリンシグナリング経路によって AD に特徴的な脳でのβアミロイドの沈着が抑制
されることが示されています 593,594)。DM と AD を併発するモデルマウスでは、老年期前の
時点で、すでに認知機能障害が明らかに現れ 582)、さらに、このマウスでは脳内インスリン
量の減少や神経細胞におけるインスリン感受性の低下が見られたと報告されています
582)。
以上のことなどからインスリンシグナリングの低下が AD 発症に関係しているのではない
かと考えられています 607)。2 型 DM で、膵β細胞がインスリンを増産して血中インスリン
濃度が高くなると BBB は輸送するインスリンを減少させ、脳内のインスリンが不足し、神
経細胞が正常に働かなくなり、細胞死へと向うと考えられています 534,608)。
以上のようにインスリンシグナリングは、DM だけでなく、タウのリン酸化やβアミロイ
ドの集積を防御することから、早期あるいは進行が中等度の AD の治療や AD の予防に重
要と考えられています 519,609)。
現在のアルツハイマー病治療方法
アルツハイマー病(AD)の進行抑制に使用されている医薬品のドネぺジル(アリセプト
®/エーザイ)は、記憶や認知機能に関係している神経伝達物質のアセチルコリン(ACh)
(35
75
ANM176®が認知症の予防と改善に役立つ可能性について
ページ)を分解する酵素であるアセチルコリンエステラーゼ(以下では Acetylcholinesterase
から AChE と表記します)の働きを抑制する作用があります。AD610,611)やレビー小体型認
知症(DLB)612,613)の患者の脳では ACh の濃度が減少しており、ACh を分解する AChE の
働きを医薬品によって抑えて Ach の量を補うことによって、AD や DLB 患者における記憶
や認知機能の低下の進行を一時的に抑制できます。ドネぺジル以外に、日本で販売されてい
る AChE 抑制医薬品にはガランタミン(レミニールⓇ/武田薬品)、リバスチグミン(イクセ
ロンⓇパッチ/ノバルティス ファーマ・小野薬品共同販売)があります。
記憶や認知機能に関係している ACh のシナプスでの適切な濃度は重要で、ACh の合成と
分解に係る酵素のバランスによって常に一定の濃度になるよう厳密に管理されています。
ACh は、コリンアセチルトランスフェラーゼ(choline acetyltransferase;ChAT)と言う
酵素によってアセチルコエンザイム A(アセチル
CoA )とコリンから作られます。また ACh は
AChE によってコリンと酢酸に分解されます(図
コリン
52)。ACh の神経伝達作用は適正な濃度が必要で
様々な経路で制御されていますが、一方、βアミ
ロイドは ACh の量を抑制することが知られてい
ます
614,615)。AD
患者の脳ではβアミロイド・オ
アセチルCoA
ChAT
AChE
酢酸
ACh
リゴマーが ChAT610)および AChE616,617)の活性
低下、ACh 量の低下 618)を引き起こしていると報
告されています。また、ACh の受容体の 1 つで
あるニコチン性アセチルコリン受容体(nicotinic
図 52.アセチルコリンの合成経路
ACh: アセチルコリン
ChAT:コリンアセチルトランスフェラーゼ
AchE:アセチルコリンエステラーゼ
acetylcholine receptor; nAChR)がβアミロイ
ドによって障害されることも報告されています 619,620)。これらのことから AD で記憶・認知
機能が低下するのは ACh が関与する脳のコリン作動性ニューロン(35 ページ)が障害を受け
ることによると考えられています
621)。最近では、βアミロイドによる
ACh の低下は副次
的な作用で、βアミロイドは最初に神経細胞に直接的に働いて記憶や認知機能を障害する
ことが示されています 622,623)。
神経作用は微妙にコントロールされているため、医薬品によってコリン作動性神経に働
く AChE を阻害して脳内の ACh 量を増加させても、再び AChE が増加するようになり、
AChE を阻害する作用がある医薬品の記憶や認知機能に対する効果は一時的となるわけで
す。高血圧や糖尿病など他の生活習慣病の治療も対処療法ですが、AChE 阻害剤による ACh
量を増加せる AD の治療と異なるのは、脳内の ACh 量は一定になるように制御されている
からです。ドネぺジルを使用した臨床試験では、健常者では記憶力や注意力などの脳機能が
低下し 624)、3 年以上継続して使った AD 患者では却って記憶力や認知機能の悪化が進行す
る 625)と報告されていることも、AChE 阻害剤が AD の長期的な治療に向いていないことを
示唆しています。
さらに、ACh は脳の中だけでなく腸や心臓の運動にも関係しており、AChE
76
ANM176®が認知症の予防と改善に役立つ可能性について
阻害剤は体にも影響する可能性があり、吐き気などの副作用の原因となっている可能性が
あります。
NMDA 受容体拮抗剤*はグルタミン酸による損傷からニューロンを保護する作用があり、
AChE を阻害する作用とは異なります。
*拮抗剤とは受容体を活性化する物質と競合して受容体の働きを抑制する物質です
AD 用の医薬品としての NMDA 受容体拮抗剤には
626)、メマンチン(メマリー®/第一三
共)があります。直接神経に作用するメマンチンは、AChE 阻害剤と同じように、NMDA
の量が一定になるように制御されていると考えられます。そのため、AD の進行を抑制する
メマンチンの有効期間は、AChE 阻害医薬品よりも短いことが示されています 627)。
このよに、AChE や NMDA 受容体拮抗剤のようにコリン作動性ニューロンをターゲット
する医薬品は、一時的に AD の進行を抑制するだけで 625,628)、果たして患者や医療費抑制に
貢献しているのか疑問もあります。
インスリンを鼻腔内へ投与した結果、AD 患者の言語想起が改善したことが示されてい
ます 629)。このインスリンの作用は、AD で脳内のインスリンが低下したことに起因する神
経作用に対する悪影響をインスリンが抑制したのかもしれません。しかし、外部から投与
したインスリンも実際に体で働く場合はインスリンを分解するインスリン分解酵素
(IDE)や脳関門(BBB)透過の機構などによって適切な濃度に制御されます。また、イ
ンスリンが細胞に届けられると、その多くは、直ちに IDE によって分解されると言われて
おり 630)、鼻腔内に投与したインスリンが認知症の治療に役立つか疑問です。
炎症抑制にインスリン抵抗性を改善する非ステロイド抗炎症剤(NSAID)(30,66 ページ)
のチアゾリジン(以下では thiazolidinedione から TZD と表記します)薬が AD に有効で
ある可能性が示唆されています 631)。TZD 薬は、主に脂肪細胞に存在するペルオキシゾー
ム増殖剤応答性受容体γ(proliferator-activated receptor γ;PPARγ)*に結合して抗
炎症作用を発揮します 631)。
*PPAR にはα,γ,δ(β)のサブタイプがあり、ほとんどの脊椎動物にある核内受容体です
632)。いずれの PPAR サブタイプも脂溶性低分子化合物や NF-κB を活性化するシグナル伝達分子
によって転写活性が生じ、DNA のペルオキシソーム増殖剤応答エレメント(PPRE)に結合し、
STATs、NFAT、C/EBP、AP-1 などの転写因子との相互作用によって抗炎症作用を示します
633)。PPAR は細胞内のペルオキシゾームを増やし、また、炭水化物、脂質、タンパク質等の代謝
や細胞の分化にも密接に関連しています 632)。
TZD 薬は抗炎症作用によって神経を保護します 346)。しかし、TDZ 系医薬品を AD 用医
薬品に適応する試みは、ことごとく失敗しています。また、TDZ 系医薬品には肝臓機能を
低下させるなどの副作用があることを考えると、長期間にわたり使用する AD には向いて
いないと考えられます。
今までにインスリンも含め様々な既存の医薬品が AD の治療用医薬品として試されまし
たが効果が確認されていません。花粉症を含め様々な慢性炎症疾患にも有効な医薬品がな
い場合が多く、AD が脳内神経の慢性炎症ともいわれていることからも、AD に有効な医
薬品がなかなか見つからない現状が頷けます。
77
ANM176®が認知症の予防と改善に役立つ可能性について
βアミロイドは AD の原因タンパク質であるという説が有力ですが、しかし、βアミロ
イドをターゲットにした AD の治療開発の難しさについて、米国スタンフォード大学
Longo とカルフォルニア大学サンフランシスコ校 Massa の両博士は次のように述べてい
ます 634)。
① βアミロイドレベルを減少させることが、AD の発病や進行を遅らせることになるとは
限らない。
② βアミロイドの生理的機能はまだ解明されていない。
③ AD の症状が現れる何年か前のβアミロイドの集積段階からβアミロイドを減らす薬
を投与するというような考えは、治療表明が始まる何年か前に極端に低毒性で信頼性
の高い薬が必要となり、不可能だろう。
④ βアミロイドをベースにした治療だけでは神経細胞を保護するだけで、その機能や可
塑性を回復させることはできない。
⑤ 遺伝とは関係しない老年性アルツハイマー病(SDAT)の明確な原因因子はβアミロイ
ドの集積であることは、最近の膨大な報告により示されているが、他の多数のメカニズ
ムも重要な因子と考えられる。
βアミロイドは脳の中で定常的に分泌されており 635)、前に述べたようにニューロンの
神経機能に関与している可能性もあります 278,279)(51 ページ「ベータアミロイドの神経毒性はど
のようにして起こるのか?」)。また、AD に限らずレビー小体型認知症、パーキンソン病、前
頭側頭型認知症、筋萎縮性側索硬化症など他の神経変性疾患や糖尿病などの生活習慣病の
全般に言えることですが、環境の変化、高齢化、体質など様々な要因が発症に関与してい
る可能性があります。これらの病気のほとんどは遺伝とは関係ありませんが、原因タンパ
ク質は推定できていても、その原因タンパク質が、どのようにして、それぞれの病気を引
き起こすのか分かっていません。そのため、一元的な作用を標的とした治療方法には限界
があり、個体差や環境も考慮することが必要です。また、医薬品のほとんどは合成した化
学物質ですが、これら合成物の長期使用は、副作用があるため 636,637)、その副作用を上回
る効果の意義を勘案する必要があります。また、インスリン製剤 143)、ニトログリセリン
638)、L-ドーパ 639)等のように医薬品に薬物耐性が数多く見られ 640,641)、長期使用の医薬品
においては薬物耐性も考慮に入れる必要があります。
アルツハイマー病にどのように備えたらよいか?
現在の医療では、アルツハイマー病(AD)は不治の病と考えられています。しかし、AD
に限らず、糖尿病や高血圧など、いわゆる生活習慣病の多くも不治の病で、多くの場合、元
の正常な状態に戻すことは困難です。ただし、AD と他の生活習慣病が異なることは、多く
の生活習慣病は、対症療法によって正常に近い状態にして通常の生活を送ることができま
すが、AD では、わずかに 1 年そこそこの進行抑制しかできない医薬品しかないので症状は
どんどん悪化していきます。AD では人格の根源である認知機能に障害が起こるため、それ
78
ANM176®が認知症の予防と改善に役立つ可能性について
に伴う家族や周りの社会の負担は他の生活習慣病よりはるかに大きく、また、人格を喪失し
た悲しみは家族に大きな心の傷を残します。そこで、記憶や認知機能低下の進行は止むを得
ないとして、AD の周辺症状を緩和する医療や介護が行われています。
AD の発症に個人の形質や生活環境の変化が影響していると考えられていることから、運
動プログラムを AD の治療や介護に取り入れる方法が一部で実践されています。環境要因
も発症に関係していると考えられる AD に、運動による活動性だけでなく、食事も重要と考
えられますが、現状では定説はありません。
本来食事で摂り得るレベルのフェルラ酸がβアミロイドの神経毒性を抑制し、トウキに
含まれるフェルラ酸と特定のクマリン類を安定的に摂取すると AD の主症状の改善に役立
つ可能性があります。
(アルツハイマー病の予防は可能か?)
体の個々人の違い(個体差)
、例えば身長などの形質の違いは遺伝的要因に環境要因が加
わって現れます。免疫系の誤作動による炎症の慢性化が関与していると考えられている花
粉症やアトピー性皮膚炎、リウマチなどの慢性炎症疾患にも個体差を決定する多くの遺伝
子と環境要因が関係しています。同じように生活習慣病も、遺伝的な個体差と様々な環境因
子が発病に関与していると考えられます。基本的に生物は、生まれてから生殖期までが進化
による淘汰を経てきましたが、高齢化は進化に反映し難い因子です。また、高齢化はヒトに
おいて近年起こった現象で、高齢期に関係する機能の淘汰は行われていないと推察されま
す。一般的に、淘汰が行われていない形質の個体差は大きくなります。遺伝子が異なっても
成人期までは、体に備わったホメオスタシス機構によって個体差があまり現れませんが、高
齢になってホルモンバランス、代謝、運動機能などが変化し、ホメオスタシス(24 ページ)の
機構が十分に機能しなくなると個体差が大きくなると考えられます。そのため高齢期にな
ると様々な形質に個体差が現れ、人によって、虚血性心疾患、糖尿病(DM)
、高脂血症、ガ
ンなど様々な生活習慣病を発症することになります。
生活習慣病にならないようにするためにはホメオスタシスが重要です。しかし、ホメオス
タシスの仕組みは複雑である上に微妙なバランスの上に成り立っているため、どのように
したらホメオスタシスが維持できるのか、よく分かっていません。タンパク質だけでもヒト
で、約 30 万種があるといわれており、多数の分子が織りなす複雑な仕組みがバランスして
いるホメオスタシスを制御して実際の臨床や予防に役立てることは困難です。例えば、イン
スリンシグナリング経路の伝達に重要な PI3K は、インスリン以外の刺激によっても活性化
します。慢性神経疾患の発症に深く関係していると言われている G タンパク質
642,643)は微
妙に制御されていることが知られています 644,645)。活性酸素、スーパーオキシド、過酸化水
素などは有害であるといわれていますが 138)、あるケースではむしろ生長を刺激する効果を
示す証拠がいくつも提示されています
139,141)。ほとんどの
DM は、遺伝とは直接関係しな
い 2 型で、インスリンの分泌や作用の低下が DM の発症に関係していますが(71 ページ「イ
79
ANM176®が認知症の予防と改善に役立つ可能性について
ンスリン抵抗性」)
、なぜインスリンの分泌や作用が低下するのか、よく分かっていません。花
粉があっても花粉症にならない人がいたり、人によって DM になったりならなかったり、
多くの生活習慣病は、原因物質が分かっていても、それがどのようにして病気を引き起こし
ているか分っていないことや、なぜ発症する人としない人がいるのか分からないことが共
通しています。このように、誰がどの生活習慣病になり易いかは、事前には分からないため、
生活習慣病を防ぐには基本的な予防対策が必要と考えられます。
血糖値を上げることは様々なストレス応答に必要ですが、一方、平常時でのホメオス
タシスにはインスリンの作用が重要です。アドレナリン、成長ホルモン、ステロイドホ
ルモン、甲状腺ホルモンなど多くのホルモンは血糖値を上げる作用がありますが、血糖
値を下げられるホルモンはインスリンとアミリンだけです 550,551)。1 型 DM ではインスリ
ンもアミリンも不足しますが 2 型 DM ではアミリン分泌は正常です。ほとんどの DM 患者
は 2 型ですので、グルコースの調整に問題を起すホルモンはインスリンだけということに
なります。生物は長い間、飢餓に苦しみ、食べ物が少ない環境下においても生きていけるよ
うに進化してきたのでしょう。飢餓に限らず、生物の生存を脅かす環境変化に対処できるよ
う様々なストレス応答の仕組み(24 ページ「細胞ストレスについて」)を進化の中で備えてきまし
たが、飽食や高齢に対する事態は 38 億年の進化の過程ではなかったことでしょう。初めて
遭遇した飽食と高齢化において、血糖値を下げる事が困難となってしまい、ホメオスタシス
に破綻を来し、様々な生活習慣病の爆発的な増加につながっていると考えられます。
インスリンシグナリング経路によって活性化した Akt は、アルツハイマー病(AD)の原
因タンパク質と考えられているβアミロイドが引起すβアミロイド神経毒性に対してニュ
ーロンを保護する作用があります
600,601)。
一方、ラットの海馬ニューロンで、βシート構
造をもったβアミロイドが PI3K の阻害を通して Akt の活性を抑制し、GSK3βを活性化す
ることが示されています 646)。PI3K はアロステリック制御(15 ページ)を受ける酵素で 647)、
βアミロイドのβシート構造が PI3K の活性に作用してインスリンシグナリング経路に直
接影響する可能性が考えられます。このように、脳におけるβアミロイド・オリゴマーの影
響はインスリンシグナリング機能不全と密接に関係すると考えられています 519,592)。インス
リンシグナリング経路によって活性化した Akt は、GSK3βを中継して糖代謝(70 ページ)と
βアミロイド神経毒性の抑制(75 ページ)の両方に関係します。GSK3βは Nrf2 を直接リン
酸化することによって Nrf2 の細胞核外への移行を促進して Nrf2 の転写活性を抑制してい
るため、GSK3βをリン酸化して不活性にするインスリンシグナリング経路による Akt の
活性化は、Nrf2 の核内移行と転写活性を促進します 548)。
転写因子 Nrf2 の核外移行は、酸化ストレスのセンサーである Keap1 のユビキチン化も
関与しています。平常では、Nrf2 は Keap1 と結合して細胞質内にあって核内の Nrf2 レ
ベルは低く抑えられています 66)。また、Keap1 は Nrf2 をユビキチン化し、ユビキチン化
された Nrf2 はプロテアソーム内で分解されます 648)。特定の求電子剤は、Keap1 と結合し
て Nrf2 を遊離させて核へと移動させ
649,650)、酸化ストレス応答やストレス耐性に関与す
80
ANM176®が認知症の予防と改善に役立つ可能性について
るタンパク質群の転写を促進します 651)。すなわち Keap1 は、酸化ストレスセンサーとユ
ビキチン化という 2 つの機能を持ち、酸化ストレスに対する防御因子の発現スイッチとし
ての役割を果たしています 66)。また、インスリンシグナリング経路で、GSK3βを経由せ
ず、Akt が直接 mTOR を経由して Nrf2 を活性化させるルートも報告されています。この
ようにして、Nrf2 は酸化ストレス等に対するホメオスタシス機構として働き 66)、様々な
環境ストレスに対する防御が強化され 124)、致死的ストレスに対する防御機構として広く
関与していると考えられています 125)。ヒトに限らず Nrf2 シグナリングは、加齢と病気に
相関して低下していることが示されています。長寿や寿命に関係する遺伝子については、
ハエなどの昆虫や寄生虫など線虫で研究が進んでいます。遺伝子の中には著しく進化した
ものと、進化の上でかなりかけ離れている生物でもかなりの部分が共通しているものとが
あります。Nrf2 の遺伝子は昆虫や線虫でもヒトとあまり変わっていません 652,653)。Nrf2
転写を活性化するシグナリングは寿命を延長することができることがハエ、線虫で示さ
れ、また、カロリー制限が Nrf2 による寿命延長の効果に
必要なことが示されています 653,654)。
一般に加齢した生
物では、Nrf2 標的遺伝子のストレス応答性は低下し
655,656)、また、加齢に伴うストレス応答の低下はヒートシ
ョックタンパク質関連の転写因子 HSF1 や p38 などのシ
グナリングでも起こっていると報告されています 657,658)。
図 53.フェルラ酸の分子構造
植物の中には Keap1 と結合して Nrf2 による転写を活
性化できるフェルラ酸(図 53)やスルフォラファン(図
54)などの成分があります。フェルラ酸については、動
物は進化の過程で常に摂取してきたと考えられますが、
今日のヒトの食事ではほとんど摂れていないと考えられ
ます。ホメオスタシスにはストレス耐性が重要な役割を
果たしており、ストレス耐性の破綻がホメオスタシスに
影響を及ぼし、老化や生活習慣病に関係していると言わ
図 54.スルフォラファンの構造
スルフォラファンはブロッコリの芽
中にみつけられた物質で、カリフラ
ワー、キャベツなどにも含まれ、抗ガ
ン作用があります。
れています。
遺伝子に直接的な原因がない老年性アルツハイマー型認知症(SDAT)は、βアミロイド
が炎症、活性酸素、タウ、ミトコンドリア、オートファジーなど様々な分子や機構に関与し
ていると報告されていますが、決定的な経路は明らかとなっていません。一方、SDAT を引
き起こす経路には DM など他の生活習慣病と発症機構が似ており、高齢になると脳内で増
加したβアミロイドに他の何かの条件が加わることで発病すると考えられます。
食事のコントロールや運動などは DM の予防に有用と言われています。今まで述べたよ
うに、DM と AD の発症には共有した経路があり、DM を予防することは AD の予防にも繋
がるでしょう。βアミロイド中のβシート構造が細胞のストレス応答と内因性の炎症を引
き起こします。この炎症が何時までも収束しないで慢性的に継続することが AD の発症に
81
ANM176®が認知症の予防と改善に役立つ可能性について
関与していると考えられます。花粉症の症状を発するとなかなか治らないと同じように、慢
性炎症疾患を治療するのは大変です。ですから、AD の予防は非常に重要です。しかし、β
アミロイドは神経作用に役割を果たしている可能性があり、一方では毒性も発揮するβア
ミロイドを制御することは厄介です。
(サプリメントはアルツハイマー病に有効か?)
動物は、通常、食べ物からエネルギーと体を構成する物質だけでなく特定の生理活性成
分を摂取しています。このように他の生物が作った生理活性成分の中には、本来、自分の
中で合成できるものもあったのでしょうが、外部から安定的に摂取できるようになると、
往々にして自分から合成する能力を失うことがあります(23 ページ)。環境が変わって、こ
れらの成分が摂れなくなると、しばしば重い病気にかかります。ビタミン C 欠乏症の壊血
病やビタミン B1(チアミン)欠乏症の脚気が、この例として有名です。大航海時代の壊
血病や白米を食べる食文化は、進化の過程では経験したことのない事態だったのです。ビ
タミンに限らず、カルシウム、ヨードなどのミネラルの欠乏症もあります。これらの欠乏
症になると欠乏した成分を補給すれば治るという単純ではないようです。
生活が変わり、食事の内容が変化すると知らず知らずの間に、様々な欠病症に陥ること
があります。高齢化も進化の過程で経験したことのない事態で、様々な機能低下に必須成
分の欠乏が加わり、一部の生活習慣病を引き起こしている可能性が考えられます。
一方、過剰症もあります。ナトリウムなど塩分の過剰摂取は高血圧や血管性疾患のリス
クファクターであることは有名ですが、必須ビタミンも過剰に摂取すると様々な過剰症が
現れます。例えば、ビタミン C の大量療法を実践すると腎臓や尿管結石を生じる危険性が
あります。一般にビタミン類は必要以上に摂取しても効果はありません。また、ヒトが食
べたことのない高濃度や物質を長期間使うリスクは計り知れません。
体のホメオスタシスには様々な生理機能因子などが複雑に関係しており、その全容は分
かっていません。そうはいっても病気にかかってしまった時にはワラにもすがる思いで
色々と試してみようということになるでしょう。しかし、病気は刻々と進行し、意味のない
商品や使い方によっては却って悪くなるものもあります。
認知症に役立つ食品として、ブドウ種子抽出物 659)、ワイン 660,661)、ザクロ 662)、ブラ
ックベリー
663)、ワイルドベリー664)、緑茶 665,666)、エクストラバージンオリーブオイル
667)が、またサプリメントではイチョウ葉エキスやポリフェノール 668,669)などの植物抽出成
分あるいはビタミン E670)などのビタミン類 671)など多数が挙げられています。しかし、欧
米で消費量が多いビタミン E とイチョウ葉エキスは、国の費用で効果があるか否かの臨床
試験が行われ、両方とも効果を認めることができませんでした 672–675)。
動物試験でアルツハイマー病(AD)に効果があるといわれている天然物質が数多くあ
りますが、ヒトに効果があるとは必ずしも言えません。特に知的な機能が障害される AD
の場合では、動物とヒトの試験の結果は、かなりの違いがあります。ヒトの精神活動が障
82
ANM176®が認知症の予防と改善に役立つ可能性について
害される認知症用のサプリメントの場合は、生理的なバックグランドとヒトによる臨床試
験が重要で、動物試験だけでは不十分です。また、AD の臨床試験では、心理的な要因が
完全に排除できず、臨床試験の結果も確実に信頼できるとはいい難いのが現状です。
よく、ワインを飲用している国民は長寿で AD や生活習慣病も少ないというフレンチパ
ラドックスが引き合いに出されますが、このような疫学的な推計は、安心感をもたらす効用
はありますが、病気に直面している人にとってはあまり意味がありません。
一般に、サプリメントで気をつけなければならない要点として、以下のことが挙げられ
ます。
1.充分な学術的背景があるか?
2.必要以上に大量に摂取することにならないか?(濃縮された生理作用のある成分
は危険性があることに注意する必要があります)
3.吸収や標的臓器に到達して作用しているか?
4.排泄は充分行われ蓄積することはないか?
5.長年にわたりヒトは食べていた経験があるか?
6.体のホメオスタシス(恒常性)を崩すことにならないか?
7.混入する副産物はどうなっているのか?
そこで以下では、比較的多く使用されている認知症用のサプリメントについてまとめま
した。
[イチョウ葉エキス]
イチョウ葉エキスは、特に脳血管循環の改善効果を有するという報告 676,677)があることか
ら世界中で認知症用のサプリメントとしても注目されています。イチョウ種子の外皮に接
触皮膚炎やアレルギーを起こすギンコール酸は葉にも 0.1~1.0%含まれています。そのた
め、イチョウ葉エキスからできるだけギンコール酸を除去する必要があります。。ドイツの
植物由来の薬品を評価する委員会「Commission E」に承認されたイチョウ葉エキス標準品
(EGb 761)の効能・効果は、脳機能障害の症状の軽減などですが認知症は該当しません。
2000-2008 年にかけ米国で行われた二重盲検の臨床試験では、軽度認知障害(MCI)(5
ページ)がある者と軽度のアルツハイマー病(AD)の約 3,000 人に 120mg の EGb761 を 1
日 2 回に分けて使用した結果、AD と MCI の改善や進行抑制の効果がなかったと示されて
います 678)。
[DHA、EPA]
分子中に二重結合を多く含む多価不飽和脂肪酸はメチル末端から数えた二重結合の位置
によって、ω3 系列とω6 系列に分けられています(図 55)。代表的なω3 系列多価不飽和
脂肪酸(以下ではω3 系列脂肪酸と表記します)には、α-リノレン酸、ドコサヘキサエン
酸(以下では docosahexaenoic acid から DHA と表記します)およびエイコサペンタエン
酸(以下では eicosapentaenoic acid から EPA と表記します)などがあります。魚油に豊
富に含まれるω3 系列脂肪酸は DHA と EPA です。生物体中にある脂肪酸は主にリン脂質
83
ANM176®が認知症の予防と改善に役立つ可能性について
の構成脂肪酸として細胞膜に含まれています。生体の総脂肪酸の中で DHA は約 11%を占
めますが、α-リノレン酸は約 0.2%,EPA は 0.1% 以下と極めて少量しか含まれていませ
ん 679)。神経系細胞には DHA を合成する酵素がないため、脳内の DHA の大半は体から血
液脳関門(BBB)に機能するアストロサイトを経由し 680)、ニューロンに供給されます
681)。
ラットの試験で DHA は、空間認知機能の向上とともに海馬・大脳皮質の BDNF 産生が
亢進したが、EPA では、これらの効果が認められなかったと報告されています 682)。ま
た、DHA がβアミロイド・オリゴマーの分泌とβアミロイドによるニューロン死、βア
ミロイドの脳内沈着などを抑制することが示されています 683,684)。
現在の欧米型の食事では DHA は相対的に欠乏する可能性があり、ω3 系脂肪酸の摂取
不足によって脳の発達障害、認知症、鬱(ウツ)病などの様々な精神神経疾患の発症要因の
一つとなっていると
考えられています
681)。
ω3 系脂肪酸から
合成されるレゾルビ
ンとプロテクチン
は、炎症収束を促進
する作用があります
53)(図
55)。ω6 系
列のアラキドン酸か
ら合成される炎症メ
ディエーターのプロ
スタグランジンとロ
イコトリエンは炎症
図 55.ω3 PUFA の抗炎症作用機序
ω3 系列の EPA や DHA はω6 系列のアラキドン酸代謝系と拮抗的に働くことで起
炎性のエイコサノイドの産生および作用を抑制しますが、ω3PUFA 由来のレゾルビ
ンとプロテクチンは炎症収束を促進すます 685)。
反応の初期過程で中
心的役割を果たしています(30 ページ)。一方、遊離型の DHA や EPA がアラキドン酸カス
ケードを阻害し、炎症誘発物質のトロンボキサン A2 やロイコトリエンの産生を拮抗的に
阻害することで抗炎症作用を発揮します。最近では、ω3 系脂肪酸はプロテクチン D1
(PD1)やレゾルビンなど炎症収束を促進する抗炎症因子を作り出すと報告されており
685)、DHA
や EPA はこれらの因子を介して AD や脳血管性認知症の発症・進行を防御する
可能性も推察されています 686)。また、多くの臨床や調査の研究で AD に対する DHA や
EPA の有用性が示されています 687)。
魚に含まれる DHA と EPA はバラつきがあるものの、生の 100 当り 1g 前後で、ヒト
によって食べる量も異なることから、成人期以降はサプリメントで補給することに意義が
あると考えられます。
84
ANM176®が認知症の予防と改善に役立つ可能性について
[レスベラトロール]
レスベラトロールは赤ワインに含まれる有効成分で、βアミロイド誘導性の細胞死を大
幅に低下させ 688)、AD モデルマウスで Sirt1*を活性化して、
海馬での神経変性を抑制し 689)、
認知機能の改善が示されています
689,690)。しかし、その逆の可能性も報告されています 691)。
*サーチュインと呼ばれる一連のタンパク質は、酵母、ショウジョウバエ、線虫などの動物試験で
寿命を延長させると報告されています 692,693)。しかし、マウスなど哺乳類での寿命延長の効果は
ハッキリしていません 694,695)。サーチュインの活性は、通常、年齢に伴って低下し 696)、また、カ
ロリー制限によってカロリーの消費を低下させ 697,698)、ストレス耐性を強化したと報告されてい
ます 699,700)。マウスのサーチュインである Sirt1(ヒトの場合は SIRT1)は脱アセチル化を行う酵
素で、DNA が巻き付いているヒストン(18 ページ、図 13)を脱アセチル化してタンパク質の読
み取りに関与しています 701,702)。また SIRT1 は、膵ラβ細胞におけるインスリン分泌の調整や
703)、糖生成 704,705)、メタボリックシンドローム 706,707)、炎症 708)に関係していると報告されていま
す。AD 患者の脳でサーチュインが健常者もしくは MCI 症例者よりも低いと報告されていま
す 709)。
[クルクミン]
クルクミン(curcumin)はショウガ(生姜)の一種であるウコンに含まれる色素で、カ
レーやインスタントラーメンの黄色の元になっています。クルクミンは、培養した免疫系細
胞でβアミロイドの解消を促進し 710,711)、AD モデルマウスの脳でβアミロイドの解消を増
強し
712)、老人斑を減少させる 713)ことが示されています。また、統計解析からアジアにお
けるクルクミンを含む食事と AD 予防との関係が示されています 714)。しかしクルクミンは
腸管からわずかしか吸収されないため、食事からはほとんど体内に吸収されません 715,716)。
サプリメントとしてクルクミンに期待される作用にはガンのリスク低減などがありますが、
逆に発ガン性があるとも報告されています 717)。
[フペルジン A]
フペルジン A(またの呼び方はヒューペルジン A、Huperzine A;Hup A) は、ヒカゲ
ノカズラ科に属するヒカゲノカズラ(日陰鬘、日陰蔓、学名 Lycopodium clavatum)と同
種のトウゲシバ(またの名は千層塔、学名 H uperzia serrata)に含まれるセスキテルペン
アルカロイドで 718)、乾燥したトウゲシバから 0.011%が抽出されます。厚生労働省の「医薬
品の効果を標榜しない限り医薬品と判断しない成分(本質)リスト」には「センソウトウ、
全草」と記載があります。
千層塔は、漢方において古くから腫れ、発熱、血液疾患の治療に用いられてきた生薬で、
止血、消炎、解毒の効果があります。可能性のある副作用は、呼吸障害、喉あるいは胸の圧
迫感、胸痛、蕁麻疹、発疹、肌のかゆみあるいはむくみ、胃のむかつき、下痢、吐き気、行
動過剰、不眠症などがあります。
中国における臨床試験で、1 日当たり 400μg の Hup A がアルツハイマー病の治療に効
果があることが示されています 719)。Hup A はアセチルコリンエステラーゼ(AChE)を阻
害する作用があります 720)。AChE 阻害力は医薬品のドネペジル(75 ページ「現在のアルツハイ
マー病治療方法」)より低いですが 721)、他の AChE 阻害剤と違うことは、NMDA 受容体拮抗
作用を併せ持つとのことです 719)。
85
ANM176®が認知症の予防と改善に役立つ可能性について
Hup A は、アメリカ合衆国では記憶支援のためのサプリメントとして販売されおり、現
在、アメリカ国立老化研究所はアルツハイマー病治療薬として、Hup A の安全性および有
効性を評価する第 II 相試験としてランダム化比較試験を行っています。以上のことから
Hup A は医薬品に属すると考えられます。
[ポリアミン]
ポリアミンは第 1 級アミノ基
(NH2-)を 2 つ以上もつ脂肪族炭
化水素の総称で,体内には 20 種類
以上のポリアミンが存在します。そ
の中でも代表的なポリアミンはスペ
ルミジン、スペルミン、プトレスシ
ンです(図 56)。これらはヒトを
含めたすべての生物に含まれます
が、胎児や新生児の細胞ではポリア
ミンの細胞の増殖能が高く、合成も
図 56.ヒトの体にある代表的なポリアミン
活発になっています。
ポリアミンは細胞内でアミノ酸であるアルギニンから合成されます。アルギニンはアル
ギナーゼの作用でオルニチンになり、さらに何段階かを経てスペルミンに合成されます。
加齢に伴って一連のポリアミン合成能は低下します 722)。高ポリアミン食を摂取すると血
中のポリアミン濃度が上がる事がヒトの臨床試験により明らかになっています 723)。
フェルラ酸にはβアミロイド神経毒性を抑制する効果がある
翰林大学生薬研究所の Yan(Ji-Jing Yan)博士らの研究は、漢方薬で「もの忘れ」に効
果があるといわれていた生薬のトウキ*の中から、βアミロイド神経毒性を抑制できる成分
を探求することでした。
300
Step-Through Latency (sec)
*トウキ(当帰、学名 Angelica acutiloba)
は、漢方薬として用いられるセリ科シシ
#
200
#
ウド属の多年草です。原産地は中国で、
Control
日本では Angelica acutiloba Kitagawa
Ab1-42
*
FA 0.002% + Ab1-42
が大和当帰(大深当帰)として、Angelica
FA 0.004% + Ab1-42
100
acutiloba Sugiyamae Hikino が北海当
FA 0.006% + Ab1-42
帰として栽培・生産されており(第一二
改正日本薬局方解説書)、市場では前者
0
の方が良品と評価されています 724)。ト
Passive Avoidance Test for FA (I)
ウキの全草に強いセロリに似た芳香が
図 57.フェルラ酸(FA)濃度による受動的回避テスト
あり、漢方薬ではトウキの乾燥根が使用
異なる濃度の FA を給水中に溶かし、マウスに 4 週間与えた
され、下部先端は最も強く血流を促し、
後の受動的回避テストの結果.
最上部(頭部)は強壮作用が強いといわれ
410pmol/ℓの Aβ1-42 を第三脳室に注入した翌日に暗室床に
微弱電流(1mA、5 秒間)のショックを与えました。更にその
ています。貧血症、月経不順、更年期障
翌日に明室から暗室に入るまでの時間(Step Through
害など、婦人科に広く使用します。
βアミロイドがマウスの記憶や学習
Latency)で記憶の定着度を判定しました 234)。
対照(Control)は Aβ42 の逆配列タンパク質
86
ANM176®が認知症の予防と改善に役立つ可能性について
を低下させることは既に分かっていま
が画期的だったのはβアミロイドの神
経毒性を数値化したことでした(図
39)。48 ページの「βアミロイドの
神経毒性作用」で述べたように、暗室
内でショックを受けたマウスは暗室に
Step-Through Latency (sec)
したが 725)、Yan 博士らが行った研究
250
200
#
Control
Ab1-42
150
*
FA (2 week) + Ab42
FA (3 week) + Ab42
FA (4 week) + Ab42
50
0
は入ろうとしませんが、βアミロイド
を脳内に注入したマウスは直ぐに暗室
に入ってしまいます。一方、βアミロ
イド神経毒性を抑制する物質を事前に
FA (1 week) + Ab42
100
Passive Avoidance Test for FA (II)
図 58.異なる期間による受動的回避テスト
マウスに 0.006%の FA をさまざまな期間与え、図 53 と同じ
テストを行いました。0.006%は 14~19mg/Kg/day に相当し
ます 234)。
マウスに与えておくと、暗室へ入るま
600
用いて、トウキ(使用されたのはトウ
キの仲間で「韓国トウキ、Angelica
gigas Nakai」でした)の根に含まれ
るβアミロイド神経毒性を抑制する成
分を調べました。その結果、ポリフェ
Acetylcholine (pg/mg protein)
での時間が長くなります。この方法を
ノールの仲間でフェノール酸と呼ばれ
るグループに属するフェルラ酸(図
53)と 12 種のクマリン類がβアミロ
イド神経毒性を抑制することが分かっ
500
400
300
*
Control(Ab42注入)
Ab42注入
FA + Ab42注入
200
100
0
Ab42を脳室注射後5日目
図 59.アセチルコリンに対するフェルラ酸の効果
マウスに 0.006%濃度のフェルラ酸(FA)を含む給水を 4
週間行った後、410 ピコモル/ℓ濃度の Aβ42 を第三脳室に注入
し、その 5 日後の大脳アセチルコリン量を測定した 234)。
たのです 726)。
βアミロイド神経毒性を抑
制する効果が、どのようにし
て生じるのかを詳細に研究す
るため、トウキに含まれる 13
種のβアミロイド神経毒性抑
制成分の中から、フェルラ酸
が選ばれました。この研究結
果、すなわち、事前に投与した
フェルラ酸がβアミロイド神
経毒性を抑制する作用の詳細
は、英国薬理学会雑誌に掲載
図 60.マウス海馬 GFAP 免疫染色(a-c) / IL-1β免疫染色(d-f)
水だけ(a,b,d,e)とフェルラ酸を 0.006%含んだ水を 4 週間与えた(c,f)
後、第三脳室内に Aβ42 の逆配列タンパク質(a,d)、Aβ42(b,c,e,f)を注入し
ました。フェルラ酸は明らかにアストロサイトでの GFAP(glial
fibrillary acidic protein)と IL-1βの発現を抑制しました 234)。
されました 234)。
まず、この研究結果を紹介しましょう。フェルラ酸は、マウスに与えた濃度が高いほど
87
ANM176®が認知症の予防と改善に役立つ可能性について
(図 57)、また、与えた期間が長いほど
(図 58)、βアミロイド神経毒性を抑制
しました。またフェルラ酸は、βアミロ
イドがアセチルコリン(図 52)のレベル
を低下させる作用を抑制しました(図
59)。
フェルラ酸は、給水中に溶かしてマウス
に与えられました。給水中に溶かしたフェ
ルラ酸の濃度は 0.002~0.006%でした。マ
ウスの体重のばらつきから、マウスの体重
を 1Kg に換算した 1 日の量は、0.14~
0.57mg でした。マウスに投与した 0.006%
のフェルラ酸は、0.42~0.57mg/30g/day
で、ヒト換算では 100~135 mg*に相当し
ます。
*マウスに対する神経作用の有効量をヒト
に換算にあたり、体重(マウス 30g:ヒト
60kg でマウスの 2,000 倍)より体表面積
(マウス 0.007m2:ヒト 1.66m2 でマウス
の 237 倍)の方が適切です。
さらに、βアミロイドによる神経毒性
が細胞内にどんな変化を起こしているの
かを追求し、海馬のアストロサイトで炎
症性サイトカイン IL-1β(28 ページ)とア
ストロサイト(7 ページ)が活性化させてい
ることを発見しました(図 60 の b,e)。
一方、βアミロイドを脳内に注入する前
にフェルラ酸を投与することによって、
図 61.フェルラ酸による GFAP、IL-1βの発現
水だけ(a,f)とフェルラ酸を 0.006%含んだ水を 1(b,g)、
5(c,h)、14(d,i)、28(e,f)日間にわたり飲んだマウス海馬の
GFAP(a-e)、IL-1β(f-j)の免疫染色.
OML(outer molecular layer/分子層表層部)、
234)
DG(dentate gurus/歯状回)
βアミロイドによるアストロサイトの活性化と IL-1βの分泌が抑制されました 234)(図 60
の c,f)。この時に、フェルラ酸自身によって一時的にアストロサイトが微弱に活性化され
ることを見出しました。この活性化したアストロサイトはフェルラ酸を与えた 5 日後には
観察でき、14 日後にはピークとなり、28 日後には元のベースレベルに戻っていました
(図 61)。マウスの第三脳室にβアミロイドを注入して、βアミロイドによる記憶・学習
の障害作用を観察したのは、フェルラ酸を与えて 14 日間が経過した後でしたので、フェ
ルラ酸によるアストロサイトの活性化がピークを過ぎてからでした。さらにフェルラ酸を
与えた期間が 14 日間より長いほど、βアミロイドによる記憶・学習の障害作用をよく抑
制しました(図 58)。マウスにβアミロイドを注入した後に与えたフェルラ酸は何ら効果
88
ANM176®が認知症の予防と改善に役立つ可能性について
がありませんでした。この現象について、Yan 博士らは次のように述べています。
「脳をマイルドなストレスに事前に慣らしておくことにより、その後の障害に対する
耐性が増強することは、すでによく知られています。この例として脳虚血 727)(28 ペー
ジ)、熱中症 111,728)、細菌外殻成分のリポ多糖類(LPS)の連続投与 729)があります。ま
た、事前処置刺激によるアストロサイトの活性化によって虚血耐性(32 ページ)が誘導
されることが報告されています 730,731)。本来、LPS は炎症を引き起こし細胞や神経に
対して毒性ですが、事前に投与した LPS によって虚血耐性が生じることは 729)、他の
ストレス応答が別のストレス耐性を生じさせるクロス・トレランス現象(34 ページ)と
考えることができます。これらの現象は事前投与したフェルラ酸がβアミロイド神経
毒性を抑制する効果とよく似ており、フェルラ酸による海馬におけるアストロサイト
の一時的な活性化とその後の平常化により、βアミロイドが脳に与える障害を回復さ
せたと考えられます。
」
以上のように、フェルラ酸がβアミロイド神経毒性を抑制したことにストレス耐性(32
ページ「ストレス耐性」)が関与している可能性があると Yan 博士らが報告しました。その
後、虚血耐性や酸化ストレス耐性には Nrf2 の転写活性化が関与している、と考えられる
ようになりました 66)(32 ページ「ストレス耐性」)。最近では、βアミロイド神経毒性をフェル
ラ酸が Nrf2 を活性化して抑制することが示されています 732)。
(フェルラ酸とはどんなもの?)
フェルラ酸は植物に含まれるフェノール酸と呼ばれる仲間の 1 つです。フェノール酸に
はフェルラ酸の他に p-クマリン酸(p-coumaric acid)、カフェイン酸(caffeic acid)、シ
ナピン酸(sinapic acid)、シリング酸(syringic acid)があります 733)。
フェルラ酸は、植物の細胞中で、アミノ酸のフェニルア
ラニンから何段階かを経て合成されます。また、フェノー
ル酸からは植物の細胞壁を形成するリグニンが合成されま
す。 植物細胞には動物界にはない細胞壁(図 62)が細胞
膜(植物の場合は「原形質膜」とも呼ばれます)の外側に
あって、植物の強度、細胞の保護などに役立っています。
細胞壁は、主にグルコースが長く連なったセルロース(動
物のグリコ―ゲンとはグルコースの結合方法が異なりま
す)およびリグニンで構成されています。リグニンはセル
ロース繊維の間を埋め、細胞壁の強化と耐水性に寄与して
図 62.細胞壁(小原圭介)
います。
多数のセルロースが束になってセルロース線維となります。フェルラ酸がアラビノース
と呼ばれる糖と結合したフェルラ酸アラビノース(feruloylate arabinose)が細胞壁に分
泌され、分泌されたフェルラ酸アラビノースはセルロース繊維に枝のように結合します。
89
ANM176®が認知症の予防と改善に役立つ可能性について
この枝の突端にあるフェルラ酸は、別のセルロース繊維の枝にあるフェルラ酸と結合しま
す。こうしてフェルラ酸同士が結合したフェルラ酸ダイマーが橋渡しをして多数のセルロ
ース線維が網の目のように結合してセルロースのネットワークが形成されます(図 62)。
また、フェルラ酸ダイマーは隣接するリグニンとも結合し、細胞壁がさらに強化されます
734)。このように細胞壁でセルロース同士の間やセルロースとリグニンの間の橋渡しをして
いるのは、フェノール酸の中でもフェルラ酸だけです。
フェルラ酸ダイマーになるためのフェルラ酸同士の結合は、植物中の酵素だけでなく、
紫外線によっても行われるため、様々な形のフェルラ酸ダイマーができあがります 735)。
酵素反応による合成物は一定の形をしているため自然界にある酵素によって分解されます
が、酵素反応以外の反応による合成物は様々で、この様々な化合物を分解できる酵素は、
なかなか見当たりません。例えば、石油製品のプラスチックが分解されないのも、それを
分解する酵素がないからです。紫外線によってフェルラ酸同士が結合したフェルラ酸ダイ
マーは菌類(カビや酵母など)が持っている酵素に対しても抵抗性があるため 736)、木材
はなかなか分解されないのです。それでも少しずつ木材が腐るのは、フェルラ酸ダイマー
を分解できる特殊な酵素をもっているカビやキノコなどの菌類が、わずかながらいるから
です。フェルラ酸が植物界に広く存在する理由は進化と関係していると考えられます。約
5 億年前頃(22 ページ注)に地表へ進出した植物は、地面に固定して光合成を行うために上
へ上へと背丈が伸びました。背丈が伸びるためには、植物体に物理的な強度が必要で、ま
た、根から吸収した水や養分を一番上にも届けなければなりません。さらに、雨や風、強
い紫外線、細菌や動物の攻撃にも耐えられなければなりません。この課題に大きな役割を
果たしてきたのが植物の細胞壁にあるフェルラ酸でした。フェルラ酸同士が結合したフェ
ルラ酸ダイマーが、セルロース間やセルロースとリグニン間をつなぎ、細胞壁に強度と耐
水性が備わったのです。
植物は動物に食べられないように様々な忌避物(いやな匂いなどを発する物質)や毒物
を細胞壁外に分泌します。これら分泌物は、植物自身にとっても好ましい物でなく、細胞
膜の外で合成する必要がありました。そのため、細胞壁に分泌されたフェノール酸が必然
的に忌避物や毒物の合成原料となります。一方、動物の神経伝達機構のような仕組みを持
たない植物は、他の何らかの方法で自分が攻撃されことを察知し、攻撃された箇所を補修
しなければなりません。 菌類などによる攻撃を受けセルロースやリグニンから切り離され
たフェルラ酸ダイマーは、細胞核に移動し細胞の修復や事後対処に必要な遺伝子の転写を
開始させているのではないかと考えられています 737)。もし、フェルラ酸が忌避物や毒物の
出発原料となれば細胞内への伝達物質とはなり得なかったでしょう。なぜなら、忌避物や
毒物は植物の種類やその組織によって含量が著しく異なるからです。 例えばクロロゲン酸
はコーヒー1杯の中に 70-350mg も含まれています。 また、リンゴ、プラム、チェリーな
どの果物に含まれるコーヒー酸も種や組織によって含量が著しく異なります 738)。 フェノ
ール酸の中でフェルラ酸だけが細胞壁の強化に関係し、侵襲情報を細胞核に伝達する役割
90
ANM176®が認知症の予防と改善に役立つ可能性について
をもったが故に忌避物や毒物の出発原料の役割が免除され、一定量が、どの植物体にも存
在するようになったのではないでしょうか。
このように、どんな植物にも一定量が含まれるフェルラ酸は、それを食べる動物も利用
するようになったと考えられます。上記のように、フェルラ酸は植物の細胞壁の必須成分
であるが故に自然界に幅広く存在しますが、細胞壁にあるフェルラ酸化合物を分解(消
化)できる酵素は動物の消化管にありません。草食動物は細胞壁の成分も吸収できます
が、その理由は、牛は胃の中で、マウスやウサギは後腸(ヒトでは退化して虫垂となって
いる)と呼ばれる大きな袋状の腸の中で、微生物による発酵によって植物細胞壁を分解で
きるからです。このように腸内細菌で分解する消化システムを進化で失ったヒトは、野菜
や果物を食べても、その中のセルロース繊維の大部分は分解されずに排泄されます。
しかし一部の植物の部位、たとえばトウキが属するセリ科シシウド属などの根 739)、穀
類の種皮と胚乳の間に存在する糊粉層といわれるヌカ部分 738)、一部の果物 740)などには、
何にも結合していないフリーかヒトの腸にある消化酵素でも分解できるアルコール結合物
として、ヒトでも吸収できる形態のフェルラ酸が含まれています 738,741,742)。穀物のヌカに
含まれているフェルラ酸の多くはトリテルペンアルコール類や各種植物ステロール、ステ
リルなどのアルコール類と結合して油脂中に溶け込んでいます 743)。総フェルラ酸が穀粒
1Kg あたり、小麦粒では 200~400mg 744,745)、米では約 100mg746)が含まれており、その中
の約 10%がフリーのフェルラ酸です 747,748)。しかし、ヌカとして糊粉層を取り除いた白米
や精白小麦粉の中には、ヒトでも吸収できるフェルラ酸は、ほとんど含まれていません
749,750)。
植物の種子に含まれるフェルラ酸は、様々なストレスから種子を守る役割や種子の発芽
や芽の生長の調節に関与していると考えられています 751–753)。穀類のヌカの中でフェルラ
酸がアルコール類と結合していますが、気温が暖かくなってフェルラ酸とアルコール類の
結合を切断する酵素が活性化するとフェルラ酸とアルコール類の結合を切断します。その
結果、フリーとなったフェルラ酸が発芽のシグナリングに関与すると考えられています。
麦の中でもオート麦やライ麦に含まれるフェルラ酸の含量が小麦と比較すると少ないのは
750,754)、発芽条件が小麦と異なることからもうなずけます。
米ヌカから油(米油)を抽出した後に残ったフェルラ酸とアルコールの結合物を分離し
て精製されたフェルラ酸が食品用の抗酸化剤として利用されています。
また、フェルラ酸およびそのアルコール類との化合物は、
① 糖尿病モデルマウスで著しく血清インスリンを増加させ血糖を低下させる効果 755)
② 糖尿病モデルマウスで尿タンパク質レベルを低下させる効果 756)
③ 癌の抑制効果 757,758)
④ 抗炎症作用 748,759)
⑤ 炎症性サイトカインの IL-1βと TNF-αのレベルを低下させ免疫系細胞の数を減少さ
せてリウマチ性関節炎の症状を抑制する作用 760)
91
ANM176®が認知症の予防と改善に役立つ可能性について
⑥ 活性酸素種(ROS)の解消
⑦ コレステロールや LDL などの脂質過酸化に対する防御 761,762)
⑧ 肝臓防御 763)
⑨ 坑血栓症や虚血に対する防御 764,765)
⑩ 網膜ニューロンの増殖作用 766)
⑪ 脳下垂体からプロラクチン(黄体刺激ホルモン)分泌 767)、ソマトトロピン(成長ホ
ルモン)合成 768)の促進作用
⑫ 血圧降下作用 769)と高脂質症、両方に対する改善効果の可能性 770)
など様々な生活習慣病や慢性炎症疾患の予防や改善に役立つ可能性が報告されています。
(フェルラ酸の吸収と排泄)
フェルラ酸の吸収と排泄についての詳細は Zhao(Zhao Zhaohui、趙 朝輝)博士らによ
って報告されています 771–773)。
ラットを用いた試験で、フェルラ酸と糖のアラビノースが結合したフェルラ酸アラビノ
ースは、約 40%が小腸までに、57%が後腸(ヒトの盲腸部分)までに吸収され、残りは主
に後腸でエステル結合が切り離されて吸収されます 771)。一方、何にも結合していないフ
リーのフェルラ酸は胃で 75%も吸収され、後腸(小腸と大腸の間にあってヒトでは盲腸に
当たる部位)に至るまでに、ほぼ完全に吸収されると報告されています 772)。フェルラ酸
アラビノースなどフェルラ酸と糖類の結合物やフェルラ酸とアルコール類の結合物である
フェルラ酸アルコールエステルは胃の中では分解されません。
ヒトの場合、フェルラ酸アルコールエステルは小腸上皮細胞*のカルボキシエステラー
ゼによって加水分解されフリーのフェルラ酸となり、能動的に吸収されます 745)。その吸
収にはモノカルボン酸トランスポーター(MCT)774) が関与する能動的な膜透過機構によ
って行われると報告されています 773,775)。
*小腸は腸管内側から、粘膜層、粘膜下組
織、固有筋層、漿膜下層、漿膜の層となっ
ています。さらに粘膜層は上皮、粘膜固有
層、粘膜筋板に分けられます。上皮は平坦
でなく、突出する部分と落ち込んだ部分が
あり、突出する部分が栄養吸収を司る絨
毛、落ち込んだ部分が分泌を司る腸腺です
(図 63)。吸収の働きを行う小腸の上皮細
胞は、同時に栄養素の消化の最終段階を行
うところでもあり、消化と吸収が同時に行
われていると考えられています。
脳関門(BBB)にも MCT が関与して
おり、フェルラ酸が BBB を直接通過する
図 63.小腸の繊毛
愛-madoka.com HP より
ことも確認されています。
胃や腸で吸収されたフェルラ酸は一旦フリーで血中を循環して肝臓に入り、そこで一部
92
ANM176®が認知症の予防と改善に役立つ可能性について
は再び糖類や硫酸残基が結合し 771,773)、次の標的細胞に到着するまでは、その結合物のまま
体内を循環します 745,773)。この循環中のフェルラ酸結合物の一部は、ペルオキシダーゼの活
性補助剤として分解されてグルクロン酸に中和され、尿中に排泄されます 773,776)。ラットで
はフェルラ酸の尿中への排泄は非常に速く投与後 1.5 時間でピークとなります 777)。ヒトで
のフェルラ酸の尿中への排泄ははもう少しゆっくりで、投与後 7-9 時間にピークとなりま
778)。修飾されていないフェルラ酸の尿中からの回収は摂取量のたった
す
ヒトでもネズミでも同程度であったとのことです
4-5%にすぎず、
777,778)。このことは、フェルラ酸のほとん
どは分解されるか抱合体として排出されることを示しています。
重要なことは、マウスやラットは後腸の中に住んでいるバクテリアがセルロースに結合
しているフェルラ酸アラビノースを分解し、細胞壁に組み込まれたフェルラ酸を吸収でき
ますが、ヒトではほとんど利用できません 737,771)。
(フェルラ酸の体内での機能)
フェルラ酸の機能は、3 つが挙げられます。
1つ目は、動物が異常事態に遭遇した時に役立つフェルラ酸の機能です。そもそもフェ
ルラ酸はペルオキシダーゼによる反応を受けやすい物質で、植物の細胞壁を構成している
フェルラ酸ダイマー(90 ページ)は太陽光の中にある紫外線によって形成されますが、細胞
壁にあるペルオキシダーゼによってもフェルラ酸同士が結合して形成されます。フェルラ
酸にはペルオキシダーゼなどのヘム(図 18)という補欠分子を活性部位にもったヘム酵素
(26 ページ)の活性を著しく増強する作用があります
779–781)。Henriksen
らは、ペルオキシ
ダーゼの活性部位に結合したフェルラ酸が、ペルオキシダーゼの活性部位にあるヘムの鉄
イオンから分解する物質へ、電子を受け渡す架け橋の役割をしている、というモデルを提
唱しています 779,780)。ペルオキシダーゼ類は多数あり、その活性部位は基本的に同じで、
フェルラ酸はペルオキシダーゼ類全般の活性力を強化している可能性が考えられます。ペ
ルオキシダーゼ類は、第Ⅰ相系の酸化反応(27 ページ)による異物の分解にも関与します。
様々な異物を分解する細胞内器官のペルオキシソームは、エネルギー生産に関わっていた
器官が進化して分解に特化したと考えられています。ペルオキシソームではペルオキシダ
ーゼなどの酸化分解酵素によって、内部に取り込んだ異物を分解します。ペルオキシソー
ムでの異物の分解は急ぐ必要があり、体の中にあるフェルラ酸の力を借りて分解を素早く
行えるようになったのではないでしょうか。一方、フェルラ酸以外の他のフェノール酸に
は、ペルオキシダーゼ活性を増強する効果はありません 782,783)。
フェルラ酸は転写因子 Nrf2 を活性化して第Ⅱ相系の酵素群を増加させることが示され
ています 732,784)。細胞質内にある転写因子 Nrf2 が核内に移行すると、DNA の ARE エレ
メントに結合し 66)、ARE エレメントを配列の中に含む第Ⅱ相系に関与する様々なタンパ
ク質がそろって読み取られます 69)(27 ページ)。細胞質内にある GSK3βは Nrf2 を直接リン
酸化して Nrf2 の核内移行を抑制しています。インスリンシグナリング経路によって活性
93
ANM176®が認知症の予防と改善に役立つ可能性について
化した Akt は GSK3βをリン酸化し、その結果、Nrf2 が細胞核内に移動して ARE エレメ
ントが含まれる DNA の転写が始まります 548)。また、Nrf2 は Keap1 によっても細胞質内
につなぎ留められ(80 ページ)、一部は Keap1 によってユビキチン化されて(17 ページ「ユビ
キチン化とプロテアソーム」)、プロテアソーム内で分解され
648)、DNA
の転写が抑制されてい
ます。特定の求電子剤は Keap1 機能の調節に重要なシステイン残基と結合し 649,650)、Nrf2
を遊離して核へと移動させます 651)(80 ページ)。フェルラ酸は Keap1 と結合できる求電子
剤の機能があり、インスリンシグナリング経路とは無関係に Keap1 の機能に影響して
Nrf2 による転写を活性化します。生物の生理的な要因が全く関与しない実験で酸化反応速
度を測定した結果では、フェルラ酸の酸化防止能力はアスコルビン酸より若干劣っていま
す 785)。一方、生理的な実験系、例えば細胞内での LDL を過酸化する実験では、フェルラ
酸の酸化防止効果はアスコルビン酸よりはるかに高いことが示されています 782,783,786)。こ
の理由はフェルラン酸が転写因子 Nrf2 の転写を活性化させ酸化還元酵素群の遺伝子から
の転写を増加させたことによります。酸化分解物は第Ⅱ相系のグルタチオン(GSH)
(図
21)やグルクロン酸などの抱合剤が結合して即座に中和されます 787)(27 ページ)。フェルラ
酸は GSH を増加させることが示されています 788)。脳内でも第Ⅱ相系のタンパク質に存在
するエレメント ARE が関与する転写が活性化され 18,789)、フェルラ酸 732)、tBHQ*、DMF
等のある種の抗酸化剤はアストロサイトと ARE の活性化が同じ細胞で同時に観察されま
*
すが 790)、抗酸化剤でも beta-naphthoflavone*や tert-butylhydroxide*には同じ効果はな
かったと報告されています 791)。
*tBHQ、DMF、beta-naphthoflavone、tert-butylhydroxide は全て化学的合成物です。
毒物や細菌・ウイルスなどの攻撃性生物の撃退も、最終的には、その生物体を分解する
ことで行われます。フェルラ酸は Nrf2 の転写を活性させることによって、高齢になると
低下する自然免疫(24 ページ)の応答レベルを上昇させ、正常なストレス応答を維持するこ
とによって、炎症や細胞アポトーシスなど二次応答に陥ることを防御していると考えられ
ます。毒物や侵襲生物の分解物は反応性が高いため、第Ⅰ相系の分解と第Ⅱ相系の中和・
解毒は連動している必要があり、第Ⅰ、Ⅱ相系両方を増強するフェルラ酸の働きは理にか
なっています。
2 つ目は、フェルラ酸がストレス耐性を強化できることです。フェルラ酸は脳虚血ダメ
ージを抑制し 765,792,793)、虚血耐性や酸化ストレス耐性などストレス耐性を増強することが
示されています 732,784)。Nrf2 はストレス耐性の強化にも関係します。よく体について抵抗
力があるとかないとか言われますが、この抵抗力の根拠はストレス耐性にあると考えられ
ます。ストレスの強さに合わせて、応答のレベルを調整する方法、つまりストレス耐性を
備えた方が淘汰にかなっているのでしょう。2010 年に北京放射線医学研空所の Ma 博士ら
は、照射による活性酸素種(ROS)の生成はフェルラ酸によって濃度依存的に抑制される
ことを示しました。さらに、放射線照射によって細胞内の GSH と NADPH のレベルは著
しく減少するが、照射 16 時間前に投与したフェルラ酸の濃度に依存して GSH と NADPH
94
ANM176®が認知症の予防と改善に役立つ可能性について
の減少が抑制され、フェルラ酸による Nrf2 の核内移動と GSH および NADPH レベルの
上昇に ERK1/2 と PI3K が関与していることを示しました 732)。Nrf2 は虚血耐性や酸化ス
トレス耐性にも関係していますので 66)、この Ma らの報告によって Yan 博士らが 2001
年に予言したフェルラ酸のβアミロイド神経毒性抑制作用を、Nrf2 の転写を活性化するこ
とによってストレス耐性が高まり、βアミロイドが引き起こす細胞ストレスや神経伝達に
対する悪影響を抑制したことによる、とストレス分子レベルで説明できるようになりまし
た。
最近、細胞系の実験で、冨栄養条件であっても、フェルラ酸がラパマイシンと同程度に
mTOR(56 ページ)のシグナリング経路を抑制し、オートファジーを活性化して長寿命タン
パク質のオートファジー分解を著しく増強することが示されました 794)。この結果から、フ
ェルラ酸には高齢になって低下すると言われている基底オートファジー371)を増強する作用
があると考えられています 794)。
フェルラ酸の機能 3 番目は、消化器官からの吸収性が抜群に高いことです。フェルラ酸
は胃からも吸収されます 772)。フェルラ酸に限らず一部のポリフェノールなど胃から吸収で
きる天然物も僅かにありますが 795,796)、その生理作用はまちまちです。
Yan 博士らは、本来、食事で補給できるレベルのフェルラ酸が、ストレス耐性を強化す
ることによってβアミロイド神経毒性を抑制することをマウスで示しましたが 234)、現代の
食事では、このレベルのフェルラ酸が補給できていません。βアミロイドの分解が遅れる
と炎症をひき起こすラットの試験で、フェルラ酸として吸収されるフェルラ酸ソーダ 771)は
濃度依存的にβアミロイドが誘導するアストロサイトでの IL-1βの分泌を抑制し、Akt を
活性してニューロンのアポトーシスを抑制すると報告されています 797,798)。
βアミロイドが引き起こす神経毒性、コリン作動性ニューロンの障害、ミクログリアと
アストロサイトの活性化、アポトーシスを抑制するインスリンシグナリング経路の阻害を
フェルラ酸は抑制できますが、これらフェルラ酸の作用にも Nrf2 が関係するストレス耐
性の強化によると考えられます。フェルラ酸は、MTT 法で観察されるβアミロイドによる
細胞毒性(51 ページ)を抑制します 732)。フェルラ酸によって事前にストレス耐性を増強して
おけば、繊維状βアミロイドを取り込んだミクログリアは周囲に炎症を蔓延させることな
く、繊維状βアミロイドを抱えたまま静かに眠りにつきます。 事実、老人斑ではペルオキ
シダーゼ活性がみられ 799)、培養細胞ではペルオキシダーゼがβアミロイドの影響から細胞
を守っていると報告されています 800–802)。
(フェルラ酸はアルツハイマー病の改善に役立つか?)
βアミロイドはアルツハイマー病(AD)の原因タンパク質であると考えられています
が、Yan 博士らが示したβアミロイド神経毒性を抑制する効果があるフェルラ酸は、AD
の治療や改善に役立つのでしょうか?
そもそも、Yan 博士らがフェルラ酸によるβアミロイド毒性抑制効果を研究した理由
95
ANM176®が認知症の予防と改善に役立つ可能性について
は、生薬のトウキ根が AD に有効であるか否かを確認するためでした。トウキ根に含まれ
るβアミロイド神経毒性抑制成分は、フェルラ酸の他に 12 種のクマリン化合物がありまし
た。βアミロイド神経毒性の抑制機構を解明するために、これらの成分の中から、たまた
まフェルラ酸が取り上げられました。現在では世界の多くの研究者が、Yan 博士らが発表
したフェルラ酸のβアミロイド神経毒性機構を参照していますが、Yan 博士ら見出したβ
アミロイド神経毒性を抑制する 12 種のクマリン化合物 726)の作用の解明は、なおざりにな
っています。
動物は植物に一定量含まれているフェルラ酸を吸収し、ヒトも主に穀類からフェルラ酸
を補給してきました。ヒトが米や小麦などから補給してきたフェルラ酸は1日当たり 85~
340mg と推定されますが、果物や葉や茎からヒトが吸収できるフェルラ酸の量は多く見積
もっても数 mg です。 Yan 博士らは、米や小麦からヒトが補給できるレベル量のフェルラ
酸がβアミロイド神経毒性を抑制できることを示しました。現代では、ほとんどの人は、
お米は胚芽を取り除いた白米を食べていますが、小麦粉も、小麦の表面にある糊粉層を取
り除いた精白小麦粉を食べています。玄米や精白していない小麦粉(地粉または全粒粉と
言います)を使った食事をしていればフェルラ酸は十分に補給できるはずですが、フェル
ラ酸がほとんど含まれていない白米や精白小麦粉で作られたパンや麺類を食べている現代
人は、1日に数 mg のフェルラ酸しか吸収できていないのではないでしょうか。このよう
な例は他にもあって、食用油は脱臭工程で酸化防止効果のあるビタミンEを取り除いてし
まっているため、わざわざ窒素を封入しています。 ただし、ビタミンEは必須ビタミン
ではありますが、現代の食事で豆類などから十分に補給できています。 また、壊血病を
引き起こすビタミン C や脚気を引き起こすビタミン B1 は、現代の食事で不足することは
ありません。βアミロイドの神経毒性を著しく抑制するフェルラ酸の働きを考えると、近
年の老年性アルツハイマー病(ADAT)の急増が、食事の変化に原因があると考えてもお
かしくありません。
フェルラ酸は転写因子 Nrf2 を活性化できる唯一のフェノール酸です。 白米や精白小麦
粉を使った食事をしていると、少なくとも何億年も前からフェルラ酸に接していた細胞
は、初めてフェルラ酸の欠乏に直面しているのではないでしょうか。高齢化を含め近年の
生活環境の変化は、生物としてのヒトにとって経験した事がない大事件かもしれません。
βアミロイド神経毒性を抑制する 12 種のクマリン化合物の中には本来であればセロリ
―やニンジンなどの野菜から摂れる成分もあります。これらのクマリン化合物は、植物が
虫や細菌の攻撃を防御するために合成するのですが、近年では、品種改良によって本来含
まれるクマリン化合物をほどんど含まない野菜を食べています。
Yan 博士らがフェルラ酸で行った試験で、βアミロイドによって低下した記憶や学習力
は何日か経つと少しずつ回復し、また、AD に見られるような神経細胞の死は起こってい
ませんでした。その後、Yan 博士らは、AD モデルマウスに6ヵ月間投与したフェルラ酸
がβアミロイドの脳内濃度を低下させ、βアミロイド神経毒性を抑制した試験結果を報告
96
ANM176®が認知症の予防と改善に役立つ可能性について
しましたが 803)、そもそも、AD モデルマウスでは、ヒトの AD に見られる神経細胞の死は
起こらないことが知られています。つまり、AD モデルの動物試験は、βアミロイドによ
る記憶力や認知機能の低下は示しているものの AD における神経細胞(ニューロン)の
死を現してはおらず、AD に効果があるか否かはヒトによる臨床試験で確認する必要があ
ると言われています。
ANM176Ⓡ がアルツハイマー病に役立つ可能性について
本来であれば、ヒトが食事で補給できる成分を配合した ANM176Ⓡは、アルツハイマー病
(AD)用の医薬品やトウキに含まれるβアミロイド神経毒性抑制成分あるいはそれらの成
分の組合せの中で、βアミロイド神経毒性を抑制する作用が最も高く(データ非公開)
、AD
の予防や改善に役立つ可能性があります。ANM176Ⓡに配合されたフェルラ酸とガーデンア
ンゼリカ根抽出物は、長い間、ヒトが食事で摂ってきた成分です。フェルラ酸は、お米や麦
のヌカ部分に含まれる成分で、ANM176Ⓡで補給するフェルラ酸の量は、玄米や地粉を食べ
ていたら摂れる量です。ガーデンアンゼリカはヨーロッパで古くから食べられていた植物
で、その根はハーブとして民間薬に利用されていました。ANM176Ⓡに配合されたガーデン
アンゼリカ根抽出物はハーブとしての利用量より少ない量です。
こんな常識的な成分が配合された ANM176Ⓡが AD の予防や改善に役立つ可能性が示さ
れています。そこで、以下ではこの可能性について詳しく述べます。
(ANM176Ⓡとは)
ANM176Ⓡという名前は、INMⓇ176 という名前に由来し、INMⓇ176 と同等の効果があ
るアルツハイマー病(AD)の予防と改善に使用するサプリメント用の配合剤であることを
表わしています。では INM Ⓡ 176 とは何でしょうか? INM Ⓡ は Institute of Natural
Medicine(生薬研究所)の頭文字で、翰林大学生薬研究所(Institute of Natural Medicine
of Hallym University)のことです。INMⓇには 1~307 まであり、INMⓇ176 は翰林大学生
薬研究所が研究したβアミロイド神経毒性を抑制する 176 番目の配合剤という意味です。
この物語は 1998 年まで遡ります。翰林大学生薬研究所において、Song 教授の指導の下
で行われた Yan 博士らの研究目的は、漢方薬で「もの忘れ」に効果があると言われていた
生薬のトウキの中から、βアミロイド神経毒性を抑制できる成分探求することでした。そこ
で、まず、βアミロイドによる神経毒性を抑制できるトウキ根のアルコール(エタノール)
抽出物を多数に区分けし(この区分けを分画と呼びます)、各分画をマウスに与えて、受動
的回避試験(48 ページ)でβアミロイドによる学習・記憶力の低下、すなわちβアミロイド神
経毒性を抑制する程度を調べました。その結果、1 つだけでなく多くの分画がβアミロイド
神経毒性を抑制することが分かりました。また、これらの分画を組合せて混ぜると、単独の
各分画に比較して、βアミロイド神経毒性を相乗的に抑制することが判明しました 726)。そ
こで、βアミロイド神経毒性抑制する分画を組み合わせた INMⓇ001 から INMⓇ307 まで
97
ANM176®が認知症の予防と改善に役立つ可能性について
の配合剤のβアミロイド神経毒性抑制効果を比較した結果、試験番号 INMⓇ176 が、βアミ
ロイド神経毒性抑制力が最も高かったのです。その後、INMⓇ176 に含まれるβアミロイド
神経毒性を抑制する成分の分析が行われて、フェルラ酸と 12 種のクマリン類が見出されま
した 726,804)。
マウスの受動的回避試験でβアミロイド神経毒性抑制力が最も高かった INMⓇ176 の濃
度をヒトに換算した濃度で、実際の AD 患者を対象とした 2 重盲検試験*が行われました。
*2 重盲検試験とは試験する物質を配合した A 製剤(実薬)と試験する物質を全く配合していない B 製
剤(偽薬)を用意し、この試験に参加した患者と医療関係者の両方には、A と B の製剤の何れが実薬
か分からないようにして、投薬されます。このような 2 重盲検試験は、医薬品開発に用いられます。
2 重盲検試験によって、医薬品を使ったことによる心理的な効果(いわゆる「プラセボ効果」)が除外
できます。認知症用医薬品の試験では一種の心理テストを用いて効果を評価するため、プラセボ効果
が出やすいといわれています。
この、2001 年に三星ソウル病院で 80 名の軽度から中等度の AD 患者を対象に行なわれ
た臨床試験で、INMⓇ176 が、ADAS-cog (Alzheimer's Disease Assessment ScaleCognitive subscale)と呼ばれるアルツハイマー病用心理テストで AD の改善に有効なこ
とが初めて示されました 805)(図
Administration of INM176® and Changes in ADAS-Cognition
64)。この試験は 3 ヵ月間という短
INM176
いものでしたが、同じく三星ソウル
Placebo
るドネペジル(アリセプトⓇ)
、ガ
20
ADAS-Cognition
病院で行われた AD 用の医薬品であ
25
ランタミン(レミニールⓇ)、リバ
Ⓡ
スチグミン(イクセロン )や当時
15
10
の開発中の医薬品であるフェンセリ
5
ン(Phenserine) の試験結果と比較
0
Post-trial
(12wk. After)
Pre-trial
すると興味ある結果でした。 つま
り、これら全ての医薬品による
図 64.軽度から中度の AD 患者に INMⓇ176 と偽薬を使
った 3 ヵ月間の ADAS-cog 得点比較 805)
Changes of ADAS-Cog Scale
ADAS-cog の平均
値は、投与開始後
-6
2 ヵ月目には改善
-4
のピークが過ぎ悪
-2
っていましたが、
Scale
化のトレンドに入
0
0
6
12
18
24
Weeks
®
2
INM 176 の臨床試験結果
INM 176 は 3 ヵ
Ⓡ
月目でも悪化する
4
ことはありません
6
でした(図 65)
805)。
このようにし
Aricept
Reminyl
INM176
A-Placebo
R-Placebo
I-Placebo
図 65.
Phenserine
Exelon
P-Placebo
E-Placebo
(既存医薬品との比較)
Dr. DK Kim ; Samsung Medical Center 80 subjects having mild & moderate cognitive
impairment 12 weeks, Double-blind & placebo-controlled clinical trial
Aricept:アリセプト®, Reminyl:レミニールⓇ, Phenserine:開発中断, Excelon:イクセロンⓇ
、INM176 は韓国 8 病院で行った臨床予備試験のトレンド
98
ANM176®が認知症の予防と改善に役立つ可能性について
て、INMⓇ176 が AD に有効なことが臨床試験で確認され
A.
ました。しかし、INMⓇ176 を商品化するにあたり大きな
問題が生じました。と言うのは、実際に流通している韓国
トウキ根の中に含まれるβアミロイド神経毒性抑制成分の
中で、一部の成分の含量が極端にバラついていることが判
B
.
ったのです。フェルラ酸の含量は安定しているのですが、
クマリン類のフロクマリン(またはフラノクマリン)(図
66)と呼ばれる一連の含量が原料によって極端にバラつい
ていたのです(図 67)。
この理由は日本の研究者によって明らかとなりました
806)。
図 66.クマリン骨格(A)とフ
ロクマリン骨格(B)
北海道衛生研
究所の姉帯博士ら
は、いろいろなト
ウキ類の根に含ま
れるフロクマリン
化合物を調べ、気
候、土地などの変
化により 2.3~
74.6mg/100g の幅
でバラついていた
と報告しています
806)(図
67)。この
図 67.北海当帰のフロクマリン含量
806)
バラつきの理由
は、植物がストレスに対応しているためと考えられています 806)。つまり、トウキ類の根
には、そもそも一連のフロクマリンは含まれておらず、収穫した後の処理によって、初め
て根の中で生まれる成分であることが明らかとなったのです。トウキ類は冬になると茎や
葉などの地上部は枯れますが、条件が良ければ根は越年し、春になると、この根から新し
い芽が出てきます。このためには、根が腐ったりモグラなどによって食べられたりしない
ように、地上部が枯れた後に、根の中で新たに合成されるようになったストレス応答物質
が一連のフロクマリンはではないかと考えられています 806)。古くから高級品として取引
されてきた大和当帰や深川当帰には一連のフロクマリン類が確保されていたと報告されて
います 806)。姉帯博士らは、この理由について、これら伝統的なトウキ産地で古くから行
っている収穫後の特殊な後処理が、収穫後の根にフロクマリン類が生成することになった
ものと推察しています。
このようにして、重要なβアミロイド神経毒性抑制成分であるフロクマリン類含量のバ
ラつき理由が判ったのです。フェルラ酸もクマリン類も細胞の外に分泌されるポリフェノ
99
ANM176®が認知症の予防と改善に役立つ可能性について
ールの仲間です。フェルラ酸は植物の種類によって含量は異なっても同種の植物の特定な
場所には安定的に含まれる物質です(89 ページ「フェルラ酸とはどんなもの?」)。
翰林大学生薬研究所の Yan 博士らによるβアミロイド神経毒性抑制作用の研究を支援
した韓国のサイジェニック社(Scigenic Co.Ltd.,Seoul)は、この Yan 博士らの研究成果を
ベースに、韓国トウキ根を原料としてβアミロイド神経毒性を抑制する成分の含量を規格
化した AD 用の医薬製剤 INMⓇ176 を開発し、さらに、日本向けの認知症の予防・改善を
目的としたサプリメント用製剤 ANM176Ⓡを開発しました。
INM Ⓡ 176 に使用されている韓
国当帰は、日本では「専ら医薬品とし
て使用される原材料リスト」(表 2)
に収載されているトウキに該当
するため医薬品用としてしか販
売できません。
表 2.専ら医薬品として使用される原材料リスト(平成 14
年 12 月 25 日付食基発第 1225001 号)
名称
他名等
オニノダケ / カラトウキ
トウキ
根
表 3.医薬品的効能効果を標ぼうしない限り食品と認められる
原材料リスト(平成 13 年 6 月 28 日付食基発第 20 号)
名称
セ リ 科 シ シ ウ ド 属
薬用部位等
他名等
アンゼリカ
ガーデンアンゼリカ
薬用部位等
全草
(Umbellifeae Angelica)は、洋
の東西を問わず古くから幅広い効果がある薬草として使
われてきました。シシウド属には 40 種ほどあり、その中
で、漢方薬でトウキと呼ばれているものは 9 種です。
そこで、「医薬品的効能効果を標ぼうしない限り食品
と認められる原材料リスト」
(表 3)に収載されている唯
一のシシウド属で、ヨーロッパでは古くから食品やハー
ブとして使われているガーデンアンゼリカ( Angelica
archangelica)(図 68)に着目しました。
漢方薬で使用されているトウキとガーデンアンゼリカ
以外に幅広い人種のヒトが長い間に亘って摂ってきたシ
シウド属の植物はありません。ヨーロッパでは、ガーデ
ンアンゼリカの根は婦人用万能薬、鎮静、強壮、抗腫瘍、
風邪治療の目的で使われました 807)。ガーデンアンゼリカ
図 68.ガーデンアンゼリカ
表 4.既存添加物名簿収載品目リスト 平成 8 年 5 月 23 日 衛化第 56 号 厚生省生活衛生局長通知
「食品衛生法に基づく添加物の表示等について」別添1
番号
名 称
基原・製法・本質
264
フェルラ酸
イネ科イネ(Oryza sativa LINNE)の糠より得られた米糠油を、室温時弱アル
カル性下で含水エタノール及びヘキサンで分配した後、含水エタノール画分に
得られたγ-オリザノールを、加圧下熱時硫酸で加水分解し、精製して得られた
もの、又は細菌(Pseudomonas)を、フトモモ科チョウジノキ(Syzygium
aromaticum MERRILL et PERRY)のつぼみ及び葉より水蒸気蒸留で得られた
丁子油、又は丁子油から精製して得られたオイゲノールを含む培養液で培養
し、その培養液を、分離、精製して得られたものである。成分はフェルラ酸で
ある。
用 途
酸化防止剤
100
ANM176®が認知症の予防と改善に役立つ可能性について
の地上部は厳冬期に肉料理に使われ、その茎は砂糖漬けにしてケーキの飾りに使われてお
図33.Aβ神経毒性の抑制比較
り、その名もまさにアンゼリカ(アンゼリカはエンゼルのこと)
と呼ばれています。しかし、
Evolution is going on with Scigenic products.
ANM176Ⓡを設計するにあたり、ガーデンアンゼリカ根にはヒトでも吸収できるフェルラ酸
がほとんど含まれていなかったた
ANM176 showed superior performance even to INM-176.
め、食品用のフェルラ酸(図 53、
200
神経毒性を抑制する特定のクマリ
ン類が一定量含まれるように規格
化された ANM176 が開発されま
Ⓡ
した。ANM176Ⓡ は、マウスを用
いた確認試験でβアミロイド神経
毒性を抑制する効果が INM 176
Ⓡ
にも勝るとも劣らないことが確認
されました(図 69)。そこで、
ANM176Ⓡが実際の認知症患者に
•Step-Through Latency (S)
表 4)が配合され、βアミロイド
150
100
50
0
Saline
Ab
FA
INM®176
ANM176®
図 69.βアミロイド神経毒性抑制力の比較
Mouse, 30.mg/kg/day, 4 weeks p.o., Aß1-42 i.c.v.
生理食塩水(Saline)、βアミロイド(Aβ)、フェルラ酸
(FA)、INM®176、ANM176®の試験物質を1日に1回、4 週間に
わたりマウスの胃に直接投入し、対照(Saline)群を除いた各群の
第三脳室に 410 pmol/ℓのAβを注入し、受動的回避テストの入室
遅延時間(step through latency)を比較しました。
役立つか否かを確認しました。
以下では ANM176Ⓡで行った臨床試験の経緯と試験結果を述べます。
(ANM176Ⓡの臨床試験の経緯と結果の概要)
通常、医薬品開発のために臨床試験が行われますが、サプリメントなど食品の効果を確
認する臨床試験は行われません。この理由は、一般的にサプリメントなどの食品は病気の
治療に用いられるものではないと考えられているからでしょう。しかし、一律に食品と医
薬品を分けると問題も生じます。例えば、ビタミン C、B1 という食品成分の欠乏症であ
る壊血病や脚気は、本来は、こうした欠乏病が起こらないように食品から吸収できるもの
です。しかし、これら必須成分が吸収できない何らかの事情が起こった場合は、その原因
を取り除くことも重要な医療と考えられます。予防を含めて総合的に対処するのが本来の
医療でしょうが、今日の医療は予防の観点が欠落気味と思われます。
実際に、ANM176Ⓡのアルツハイマー病(AD)に対する確認試験も、財団法人ぼけ予
防協会(現在、認知症予防財団)の佐藤哲朗常任理事(現在、ふれあい福祉協会理事長)
に候補の医療機関をご紹介いただきましたが、なかなか引受先が見つかりませんでした。
その中で、岡山県笠岡市にある認知症専門病院「きのこエスポアール病院」の佐々木健院
長は、有効な医療方法がない認知症医療の現状においては、少しでも可能性のある方法に
は真摯に対応すべきである、との本来あるべき医療精神を持たれていました。
ANM176Ⓡ関連のデータを検討した佐々木先生は、まず、少人数で可能性があるか否か
を確認しようと、2005 年の秋から了承をいただいた 9 名の AD 患者に 6 ヵ月間にわたり
朝と晩に 300mg の ANM176Ⓡ を使った予備試験が行われ、期待以上の可能性が確認され
101
ANM176®が認知症の予防と改善に役立つ可能性について
ました。 そこで、2007 年の春から 8 医療機関、9 名の認知症専門医の協力を得て、軽度
から重度の AD 患者 143 名を対象に 9 ヵ月間の試験を行うことになりました。きのこエス
ポアール病院で行った予備試験にはサイジェニック社から輸入したカプセル商品「アルチ
ーマ-J」(1カプセルに ANM176Ⓡが 150mg 配合)を使用しました。
2007 年から行った本格試験の目的の1つは、どんな進行程度の患者に、つまり軽度と
重度のどちらに、より効果があるか、を観察することにありました。そのため、試験に参
加する AD 患者の進行程度に制限を加えないことにしました。そうすると、場合によって
は嚥下障害があってカプセル商品は利用できない事態が想定されたため、ANM176Ⓡの原
料をサイジェニック社から提供を受け、日本国内で顆粒にしてスティックに詰めた商品で
試験をすることになりました。この製品に協力いただいたのが G 社で、その商品名がフ
ェルガード®でした。しかし、試験開始直後に G 社は無償で提供した ANM176Ⓡで作った
フェルガード®を、試験用に提供することを拒否したため、改めて輸入した ANM176Ⓡの
原料を用いて製造した「ANM176 顆粒」で試験を行いました。現在のフェルガード®には
ANM176Ⓡは使われていません。この結果、統計的にも ANM176Ⓡ が AD の改善や進行
抑制に有用であることが確認され Geriatric Medicine 誌に発表されました 808)。
この試験の目的は、ANM176Ⓡ が以下の効果を示すかを確認することでした。
1.AD の進行度に対して、どのような効果があるか? つまり、進行程度が軽度と重度
を比較して、どちらに効果が高いか?
2.アリセプト®を 1 年以上使用して進行抑制の効果がなくなった後、アリセプト®と併
用した場合、効果があるか(このころは AD 用の医薬品はアリセプト®しか認可され
ていませんでした)
そのため、アリセプト®を使用していないか、アリセプト®を使用している場合は使用後
1年以上が経過して、効果が感じられなくなった AD 患者を対象としました。 すでにア
リセプト®を使用している AD 患者に関して このような制限をした理由は、アリセプト®は
使用開始から1年間を経過すると AD の進行抑制効果が失われていると考えられており
625)、かといって使用を中断すると進行が早まるとの報告もあった 809)からです。また予備
試験でも、アリセプト®と併用した場合でも ANM176Ⓡ の効果がありました。偽薬を使用
する二重盲検検査は、以下の理由で行わないことにしました。
1.偽薬を使うことにヘルシンキ宣言に盛られているような倫理問題がある
2.1 人の患者における ANM176Ⓡ 使用前と使用後を比較することによっても、改善効
果を評価できる
3.9 ヵ月間の評価をすることによってプラセボ効果を無視できる
試験に参加した AD 患者は、試験開始と同時に朝と晩に ANM176Ⓡ を 300mg 服用し、
開始時、3 ヵ月後、6 ヵ月後、9 ヵ月後に認知症用の心理テスト MMSE(Mini Mental
State Examination)および/あるいは日本語版 ADAS-jcog(Alzheimer's Disease
Assessment Scale-Cognitive subscale Japanese edition)で記憶力や認知機能が評価さ
102
ANM176®が認知症の予防と改善に役立つ可能性について
れ、同時に一般健康診断と炎症に関する血液検査が実施されました。
以上の方法で行った本試験の結果は、動物で行われた試験や INMⓇ176 の臨床試験 805)
あるいは事前に行われた確認試験から想定された内容より良い結果で、詳細は以下の通り
でした。
1.MCI を含め軽度から重度までの AD 患者の4割弱が 9 ヵ月前に比較して、ADASjcog 得点で維持か低下していました(ADAS-jcog 評価では得点が低いほど正常に近
い)(表 5)。
2.軽度の患者ほど効果が高い傾向でした(図 70)。
3.65 歳前より 65 歳以上で発病した場合の方が、効果が高い内容でした(表 5)。
4.アリセプト®を使って1年以上経った人より使っていない人の方が、効果が高い内容
でした(図 71)。
5.アリセプト®を使い始めてから1年以上経過すると ADAS-jcog 得点が改善すること
はありませんが、ANM176Ⓡ を併用した場合の4割弱の 9 ヵ月後は開始時と同じか
改善していました(表 5、図 71)。
6.どのケースにおいても「米国アルツハイマー病における健康と経済の評議委員
会」が採用している AD 患者仮説モデル(図 70,71 中の*810))やアリセプト®使用1
年後の進行推移(図 70,71 中の**811))より進行が抑制されていました。
表 5.ANM176Ⓡ 試用9ヵ月後の ADAS-jcog が試験開始時と同等以下(改善)であった比率
開始時
ADAS
65 歳前
65 歳後
抑制率
総数
改善数
総数
改善数
3
3
100%
1
1
100%
2
2
100%
<20
20-30
≧30
9
3
6
4
0
0
44%
0%
0%
1
1
0
1
0
100%
0%
-
8
2
6
3
0
0
38%
0%
0%
総員
18
4
22%
2
1
50%
16
3
19%
<20
20-30
≧30
34
22
21
16
13
8
50%
59%
38%
7
6
3
5
4
0
71%
67%
0%
27
15
18
12
9
8
44%
60%
44%
総員
77
38
49%
16
9
56%
60
29
48%
98
45
45%
19
11
58%
78
33
42%
全体
ADAS-jcog評価平均の変化±標準誤差
-3.00
-3.00
-2.00
-2.00
-1.00
-1.00
0.00
3ヵ月目
6ヵ月目
9ヵ月目
0.00
1.00
1.00
2.00
2.00
3.00
4.00
5.00
6.00
ANM176Ⓡ +アリセプト®
改善数
MCI
発
症
年
齢
ANM176Ⓡ
総数
ADAS-cog<20 (n=26)
ADAS-cog/20-30 (n=21)
ADAS-cog>30 (n=20)
Caro et al*
Rogers et al**
図 70.ANM176®を使用した AD 患者の ADAS-jcog 推移
抑制率
抑制率
ADAS-jcog評価総合得点の平均±標準誤差
3ヵ月目
6ヵ月目
9ヵ月目
3.00
4.00
5.00
6.00
ANM176®
ANM176®+アリセプト®
Caro et al*
Rogers et al**
図71.ANM176Ⓡ単独とアリセプトⓇ併用のADAS-jcog推移
103
ANM176®が認知症の予防と改善に役立つ可能性について
(ANM176Ⓡには製造ノウハウがある)
INMⓇ176 は、トウキ根に含まれるβアミロイド神経毒性を抑制する分画を組み合わせ
た 307 通りから選ばれた最も有効にβアミロイド神経毒性を抑制する製剤です。また、
INMⓇ176 を元として開発された ANM176Ⓡは、フェルラ酸よりβアミロイド神経毒性を
抑制する効果が高く、INMⓇ176 と同等以上であることが確認されました(図 69)。その
後、INMⓇ176 および ANM176Ⓡは、実際にアルツハイマー病(AD)の治療に有効である
ことが確認されたことは 805,808)、前に述べた通りです。
翰林大学生薬研究所の Song 教授らのグループと、この試験を支援したサイジェニック
社は、フェルラ酸ならびに一部のクマリン化合物がβアミロイド神経毒性を抑制すること
によって AD の予防や治療に使用できる特許を共同で出願し、2009 年には日本の特許庁か
ら承認されています。
前に述べたように、漢方では、古くから生薬のトウキに「もの忘れ」を予防する効果が
あると伝承されています。近年でも、トウキが処方されている当帰芍薬散(トウキシャクヤクサン)
が AD の改善に有効なことを示す臨床試験結果が報告されています 812)。また、この効果
がアセチルコリン受容体の増加に由来する可能性が示されています 813,814)。それにもかか
わらず、認知症医療現場では、当帰芍薬散は使われていないのが現状です。この理由が、
Yan 博士らのβアミロイド毒性抑制成分と姉帯博士らのトウキに含まれる生理活性物質の
研究、ならびに、サイジェニック社が INMⓇ176 を開発する過程で分かってきました(98
~99 ペーシ)。すなわち、βアミロイド神経毒性を抑制する成分の中で、フロクマリン類の含
量が極端にバラついていたのです。
慢性系の病気に対して即効作用のある薬はありません。このような慢性系の病気に効果
があったとしても、それは長期間に亘って少しずつ改善する方法です。慢性炎症疾患の 1
種と言われている AD の場合も同じで、原料のトウキに含まれるβアミロイド神経毒性を
抑制する成分の含量のバラつきは、AD 用としては、決定的な致命傷です。
クマリン類含量のバラつきの理由が判っただけでは INMⓇ176 の商品化の問題が解決で
きません。というのも、当時でもβアミロイド神経毒性を抑制するクマリン類は 12 種も
発見されており、これらの全成分を規格化したトウキ抽出物を作るのは不可能であるから
です。これを解決したのは Yan 博士らのβアミロイド毒性抑制成分と姉帯博士らの研究で
した。両博士らは、何ら接点がないにもかかわらず、同じ結論に達していたのです。クマ
リン類には抗炎症、抗菌、抗ガンなど様々な生理作用があることで知られています 815)。
植物に含まれるクマリン類は 30 グループ、150 種に分類され、2 次代謝物まで含めると約
1300 種があるといわれています 815,816)。このようにクマリン類には多数の種類があり、ま
た、植物や部位によって含量は大きく異なるものの、同一の植物部位では、フロクマリン
類以外は、比較的に安定して含まれています。βアミロイド神経毒性を抑制するフロクマ
リンには 3 種類があり(図 67)、これら 3 種類のフロクマリンの含量には相関関係がある
ことが判明しました。つまり、フロクマリン類は含量のばらつきが多いのですが、βアミ
104
ANM176®が認知症の予防と改善に役立つ可能性について
ロイド神経毒性を抑制する 3 種類のフロクマリンは常に一定の比率を保っていることが分
かったのです。この事は実際に製剤を生産する上で非常に重要なことで、13 種のβアミロ
イド神経毒性抑制が安定的に含まれるように規格化した INMⓇ176 と ANM176Ⓡが誕生し
ました。
実際に流通しているトウキやガーデンアンゼリカのほとんどにはβアミロイド神経毒性
を抑制するクマリン類が含まれていません。市販のフェルガードⓇを分析した結果、βア
ミロイド神経毒性を抑制できるクマリン類が全く含まれていませんでした。
「きのこエスポアール病院」での確認試験では、サイジェニック社から提供された「ア
ルチーマーJ」
(1カプセルに ANM176Ⓡが 150mg 充填されている商品)を朝晩 2 カプセ
ルずつ用いました。 2007 年から 143 名の AD 患者で行われた試験では、嚥下(エンゲ)障
害のある重症患者も含まれる可能性があったため、ANM176Ⓡをサイジェニック社から供
給を受けて顆粒商品を開発し、日本タブレット株式会社(宇治市)の GMP 適合工場で製
造した 60 スティック入り「ANM176 顆粒」を朝晩 1 スティックずつ用いました。
「きのこエスポアール病院」で行った確認試験は、本来公表してはいけないものです。
しかし、この試験がフェルガードⓇで行ったように改ざんしてネットで公開されていま
す。また、木村武美著の「BPSD 症状から学ぶ治療戦略」フジメディカル出版の中では、
ANM176Ⓡで行った試験をフェルガードⓇで行ったと改ざんされています。フェルガードⓇ
には ANM176Ⓡが使われていません。また、βアミロイド神経毒性を抑制する 13 成分を
規格化した ANM176Ⓡは製造ノウハウがあり、このノウハウは一切公開していません。フ
ェルラ酸とガーデンアンゼリカを配合した他社品は ANM176Ⓡとは、効果が明らかに異な
ることは前述した通りです。ANM176Ⓡと同じ効果があるように思わせて、実際に病気が
進行している AD 患者にフェルラ酸とガーデンアンゼリカを配合しただけの商品を販売し
ていることは倫理上の問題があります。また、認知症医療に役立てるために、ANM176Ⓡ
の臨床試験にボランティアで参画して頂いた多くの先生方に却ってご迷惑をおかけしてい
る結果となっているのは残念でなりません。
(ANM176Ⓡの品質管理と安全性)
現在では、サイジェニック社から特許ライセンスと製造・品質管理のノウハウを受け、
ANM176Ⓡ を日本国内で製造しています。 その製造方法は、βアミロイド神経毒性を抑
制できる 12 種のクマリン化合物が規定量以上含まれるガーデンアンゼリカ根を選定して
ヨーロッパから輸入し、その抽出物にβ
アミロイド神経毒性抑制成分が製造規格
表 6.フェルラ酸の自主規格
表 6)のフェルラ酸を配合して粉末にしま
重金属
本品を乾燥したものは、フェルラ
酸(C10H10O4) 98.0%以上を含む
Pb として 20μg/g 以下
す。ANM176Ⓡ は配合製剤であり、実際
ヒ素
乾燥減量
強熱残分
As2O3 として 4.0μg/g 以下
0.50%以下(105℃、3時間)
0.10%以下
値に適合するよう食品添加物規格(表 4、
に ANM176Ⓡ が使用されている商品は、
含量
105
ANM176®が認知症の予防と改善に役立つ可能性について
表 7 に記載された 4 商品だけです。
表 7.ANM176®が配合されている商品
商品名
1 カプセル(スティック)の含量
剤形と形体
ANM176 顆粒
60 スティック入り箱
ANM176 顆粒 180
180 スティック入り箱
ANM176 カプセル 60
60 カプセル入りボトル
ANM176 カプセル
360 カプセル入りボトル
180
ANM176Ⓡ
総内容量
顆粒 1,800mg
粉剤 300mg
300-301mg
150-151mg
臨床試験に使われた ANM176Ⓡ は朝と晩にそれぞれ 300mg で、そのなかに処方されて
いる有用成分のフェルラ酸とガーデンアンゼリカ根抽出物はそれぞれ 100mg です。フェ
ルラ酸 100mg は、米に換算すると約1Kg、小麦では 250~500g の穀粒に含まれている量
に、ほぼ相当します。今日のように精米技術や製粉工業が発達していない頃に、通常の食
事から摂れたフェルラ酸の量は、ANM176Ⓡの標準使用量と、そんなにかけ離れていなか
ったと考えられます。
92 ページ「フェルラ酸の吸収と排泄」の項で述べたようにフェルラ酸の排泄は早く、ヒ
トの場合は投与 7~9 時間でピークとなり、24 時間以内に完全に分解もしくは硫化かグル
コシル化されて排泄されます 773)。フェルラ酸は、かつては、天然食品添加物として販売
されていましたが、現在では既存食品添加物リストに収載されており(表 4)、抗酸化剤と
して食品に利用されています。また表 6 の規格で管理されているフェルラ酸は、通常の使
用量である限り安全であることが確認されております 817)。
サイジェニック社が行った ANM176Ⓡ の急性毒性試験は以下内容でした。
実験用マウスをオスとメスおのおの 5 匹、計 10 匹で構成する 5 群に分け、1 群に
は対照として 0.5%濃度の CMC-Na(カルボキシメチルセルロース:生理活性がな
い物質)溶液を残りの 4 群それぞれにマウスの体重1Kg につき ANM176Ⓡ
250mg、500mg、1000mg、5000mg を 15mℓの溶液にし、2週間にわたり各群に毎
日胃に直接投与し、医学的変化を毎日観察しました。
2 週間の観察が終わったあと全てのマウスは解剖され、全ての臓器についての解
剖学的観察の結果は各群と対照群との差はなく、全てのマウスになんら医学的な異
常はありませんでした。
143 名の AD 患者が 9 ヵ月間にわたり ANM176Ⓡを朝晩 300mg ずつ使用した今回の臨
床試験で全ての参加者が 3 ヵ月ごとに行った血液検査において、血糖値、肝臓機能指標
GOT(AST)、GPT(ALT) 、ALP 及びγ-GTP、腎臓指標 BUN、心臓指標 CPK、炎症指標
CRP の全ての項目で、ANM176Ⓡを使用したことによる異常は観察されませんでした。
昔からヨーロッパにおいて、ガーデンアンゼリカは、民間薬として主に根と種が使われ
106
ANM176®が認知症の予防と改善に役立つ可能性について
807)、強壮や便秘、差込み、咳、皮膚病、痔、炎症、潰瘍、月経不順、リューマチ、排尿障
害、胃腸内にガスがたまる鼓腸などを治療する民間薬として利用されてきました。食品とし
ては蜂蜜や嗜好飲料などの香料あるいは野菜として、また、お菓子などの装飾材、健康に役
立つハーブとして使われてきました。ガーデンアンゼリカの葉や茎や根など全ての部分は食
品として使用できると厚生労働省のリスト(表 3)に収載されています。ところが、ガーデ
ンアンゼリカの抽出物を配合した化粧品によって皮膚炎症が起こったことから、ガーデンア
ンゼリカの安全性が欧米で問題になりました。
そもそも植物に幅広く含まれるフェノール類は(ポリフェノールもフェノール類の仲間で
す)
、光の感受性があり、その結果、皮膚炎症を引き起こす様々な副産物が生じます 818)。ま
た、フロクマリン類のソラレンは、紫外線が併用される発癌性が報告されています 819)。こ
れらのことはガーデンアンゼリカに限らず、レモンなどのかんきつ類やセロリ―などセリ科
の野菜にも共通しており、食事で通常取る量の何十倍の量を食べたり皮膚につけたりすると
思わぬ副作用が生じることがあり注意が必要です。さらにフロクマリン類のキサントトキシ
ンには異物の分解に関与する p450 の活性を阻害する作用があります 820)。フロクマリン類
のキサントトキシンはパセリやセロリなどにも含まれています。
欧州医薬品庁(EMEA)はガーデンアンゼリカ製剤のリスクファクターには光変異性と光
発癌性の両方があるとしています。そのため EMEA は、フロクマリン類の 1 日の平均摂取
基準を 1.45mg、最高摂取量を 14 mg と表明しています。この EMEA の指標に基づいた英
国食品基準庁(UK FSA: Food Standards Agency)は、ガーデンアンゼリカ根を伝統的な
通常の消化効果用に使用できるが、呼吸器系においては 1 日あたり 4.5 g の乾燥根相当を上
限としています。AD 試験で使用した ANM176Ⓡ に使用されているガーデンアンゼリカ根は
乾燥根に換算すると 1 日当たり 2gです。また、ANM176Ⓡ ではフロクマリン類の含量を規
格化しており、ANM176Ⓡ の標準使用量に含まれるフロクマリン類は 1 日当たり 0.8mg 以
内です。
商業的なガーデンアンゼリカの栽培は主に根の収穫を目的としており、栽培国はポーラン
ド、オランダ、フランス、ベルギー、スイス、以前のチェコスロバキアがあります。これら
の地域で生産されているガーデンアンゼリカに含まれるフロクマリン類の含量は大きくばら
ついており、前に述べた様にこれらのフロクマリン類は、収穫後に新たに根の中で合成され
る成分であるため、8 割方はほとんど含まれていません。一方、高濃度に含まれている場合
でも、フロクマリン類が 0.2%以上含まれているケースは稀です。
(ANM176Ⓡはアルツハイマー病の予防と改善に役立つ)
βアミロイドは記憶と関係がある神経伝達物質のアセチルコリン(ACh)量を増加させる
コリンアセチルトランスフェラーゼ(図 52)の活性を低下させ 610)、ACh 受容体の 1 つで
あるニコチン性アセチルコリン受容体(nAChR)を障害し
619,620)、これらなどがコリン作
動性ニューロンの働きに影響して 614,615)、マウスの記憶や学習力を低下させます 234,288)。ま
107
ANM176®が認知症の予防と改善に役立つ可能性について
たβアミロイドは、ニューロン樹状突起のスパイン膜にある足場タンパク質 PSD-95 の量
を減少させ、PSD-95 の PDZ ドメインに結合する NMDA 受容体など神経作用に関係する
様々なタンパク質の結合量を減少させ
279)(65
ページ)、長期記憶と関係するニューロンネッ
トワークの形成に悪影響を及ぼすと考えられます。ニューロンの細胞外で濃度が上昇した
βアミロイドは、インテグリンが関与するβアミロイド複合受容体(52 ページ)を刺激し、チ
ロシンシグナリングを引き起こして PI3K を活性化し、PIP3 を増産させます(53 ページ)。増
産した PIP3 は細胞内で酸化ストレスを引き起こしてニューロン膜の電子伝達系に影響し、
記憶や学習に影響を及ぼすと考えられます。このようにβアミロイドは、神経伝達やスパイ
ン形成などに影響して、アルツハイマー病(AD)の記憶や認知機能の低下を引き起こすと
考えられます。さらにβアミロイドは、脳内で酸化ストレスを引き起こし、慢性の神経炎症
を引き起こしてニューロンのアポトーシス(自己死)を誘導し、AD に見られる脳の萎縮と
認知機能の低下を進行させるのではないでしょうか(63 ページ「βアミロイドがアルツハイマー病
を発症させる仕組み」)。
現在の AD 用医薬品は AChE の活性を阻害して ACh 量を増加させ(75 ページ「現在のアル
ツハイマー病治療方法」)
、AD で低下している ACh 量
610,611)を補う作用があります。一方、β
アミロイドによるマウスの記憶や学習力の低下を抑制するフェルラ酸には、医薬品のよう
な AChE 活性を阻害する作用がない 821)にもかかわらず、βアミロイドによる ACh 量の低
下を抑制します(図 59)。また、フェルラ酸と特定クマリン類を含む ANM176Ⓡは、フェル
ラ酸単品や医薬品のドネペジル(75 ページ「現在のアルツハイマー病治療方法」)よりβアミロイ
ド神経毒性を抑制する効果が高い(データ非公開)ことが確認されています。このことは、
βアミロイドが ACh 量を低下させる経路には AChE 活性の抑制作用以外のルートがあり、
フェルラ酸は、このβアミロイドが ACh 量を低下させるルートを抑制していることを示唆
しています。AChE 阻害作用もしくは NMDA 受容体拮抗作用を利用した AD 用医薬品が
AD の進行を一時的に抑制する効果しかない理由は(75 ページ「現在のアルツハイマー病治療方
法」)
、ACh 量は神経活動によって制御されており、医薬品で制御した ACh 量は、その神経
活動によって短期間に修復されることによる、と考えられます。
神経系細胞の中ではアストロサイトにだけ観察される転写因子 Nrf2120)は、酸化ストレス
応答など様々な環境ストレスによって、その転写能力が活性し 69,122)、ストレス応答を統一
的に制御し、虚血耐性にも関係します 66,123)(32 ページ「ストレス耐性」)。Nrf2 の転写活性化
に関与するフェルラ酸 732)は、アストロサイトで高齢になって低下したストレス耐性(89 ペ
ージ)を強化し、βアミロイドによって低下した ACh 量を回復させ、βアミロイド神経毒性
を抑制しました 234)。
一方、このフェルラ酸によるβアミロイド神経毒性抑制効果を観察したマウスやラット
(以下では「げっ歯類」と言います)のニューロンの死は観察されず、また、βアミロイド
注入 8~10 日後にはβアミロイドによる記憶・学習低下作用も回復しました
234,288)。また、
βアミロイドの濃度が上昇する変異がある AD モデルマウスを使った試験で、フェルラ酸
108
ANM176®が認知症の予防と改善に役立つ可能性について
は脳内のβアミロイドの集積を減少させ、記憶や認知機能の低下を抑制しましたが 803)、ニ
ューロン死は見られません。このことは、げっ歯類の第三脳室に注入したβアミロイドの作
用は、ヒトの AD におけるβアミロイドの作用の全てを現していないことを示唆していま
す。AD では、ニューロンの神経作用が障害されるだけでなく、ニューロン死も進行するか
らです。βアミロイドは神経作用を障害するだけでなく、脳内の神経炎症やニューロン死の
進行も引き起こします(51 ページ「βアミロイド神経毒性はどのようにして起こるのか?」)。このこ
とから、げっ歯類の第三脳室に注入したβアミロイド神経毒性を抑制した試験は、フェルラ
酸が AD における記憶力や認知機能の低下の改善に役立つ可能性を示唆していますが、フ
ェルラ酸だけで AD で起こるニューロン死の進行抑制に十分かどうかは分かりません(95 ペ
ージ「フェルラ酸はアルツハイマー病に役立つか?」)。事実、トウキに含まれβアミロイド神経毒性
を抑制できる成分はフェルラ酸の他 12 種のクマリン類があったことは
726)、βアミロイド
神経毒性は様々なルートで起こっていることを示唆しています。
トウキ根やカンゾウ根が配合されている漢方薬は、βアミロイドがマウスで引き起す記
憶や学習力の障害や ACh 量の減少を抑制し 726,822,823)、AD の改善にも有効であることが臨
床試験で示されています 812)。また、韓国トウキを原料にβアミロイド神経毒性を抑制する
フェルラ酸と 12 種のクマリン化合物の含量が規格化された INM®176 は二重盲検法で AD
症状の進行抑制に有効であることが示されています 805)(図 64)。フェルラ酸と抗炎症作用
がある特定のクマリン類を配合した ANM176Ⓡは AD 患者に 9 ヵ月間使用した試験で症状
の抑制に有効なことが示唆されました(図 70,71)。一方、フェルラ酸だけによる AD 抑制
効果は、臨床試験では報告されていません。
そもそも ANM176Ⓡ は、Yan 博士らが行ったトウキによる AD 改善作用に関する研究が
ベースとなって開発された INM®176 をベースに、AD の予防や改善用のサプリメント用製
剤として開発されました。トウキやガーデンアンゼリカに含まれβアミロイド神経毒性を
抑制する 12 種のクマリン類の一部には炎症抑制作用があります
815)。しかし、抗炎症作用
だけでは AD に対する効果が十分に発揮できないだろうという事も、炎症を抑制する非ス
テロイド抗炎症剤(NSAID)が AD 用医薬品としての効果が実証されていないことから窺
えます(75 ページ「現在のアルツハイマー病治療方法」)。さらに、NSAID には長期使用に耐えら
れない副作用もあります(77 ペーシ)。 AD は多因子性疾患と考えられており、βアミロイド
は、直接的にニューロンを障害する作用だけでなく、脳内炎症にも関与して AD を発症させ
ると考えられています。AD に限らず、他の慢性炎症疾患にも言えますが、これらの病気を
引き起こしている炎症を抑制するには、炎症の原因と既に起こっている炎症の両方を抑制
することが重要です。原因を処置しないで炎症を抑制するだけの治療は、一時的な効果があ
っても、根本的な治療はできません。また、炎症は原因を増強することもあり、この原因と
炎症の悪循環を断ち切るためには、炎症を引き起こしている原因と炎症抑制の両方をバラ
ンスさせて抑制することが重要と考えられます。
AD 患者のほとんどは遺伝とは関係しない老年性アルツハイマー型認知症(SDAT)です
109
ANM176®が認知症の予防と改善に役立つ可能性について
が、高齢になるとβアミロイドの分泌とは直接関係ない他の遺伝的な要因が、SDAT の発症
に関連している可能性があります。例えば、βアミロイドの分解に関連している NEP(53
ページ)やβアミロイドの排出に関連している ApoE などの遺伝子多型が SDAT 発症のリス
クとなっていると考えられています(61 ページ「アルツハイマー病の発症リスク」)。さらに、高
齢になると、脳のインスリンシグナリングが低下し 824)、GSK3βをリン酸化して Nrf2 の転
写活性を上昇させる能力が低下します。ヒトの nrf2 遺伝子も多型です
458,825)。こうしてス
トレス耐性の能力が低下したところにβアミロイドが増加して、神経伝達系を直接的に、か
つ、炎症を通して障害します。一方、高齢になって低下するインスリンシグナリングを、フ
ェルラ酸などごくわずかな天然物が、インスリンシグナリングによる GSK3βのリン酸化
とは無関係な Keap1 のルートで、インスリンシグナリング経路が十分に活性化できなかっ
た Nrf2 の転写活性を補強すると考えられます。
βアミロイドによる神経毒性作用に様々な要因が加わって AD が発症する可能性があり、
ANM176Ⓡの成分管理は重要です。INMⓇ176 と ANM176Ⓡの製剤開発の過程で、市販され
ているトウキやガーデンアンゼリカの根に含まれ炎症抑制作用がある特定クマリン類の含
量が極端に異なることが判明しました(104 ページ)。AD に対しては継続的に炎症を抑制する
ことが重要であるにもかかわらず、トウキに含まれる抗炎症作用がある成分の含量がバラ
ついていたのです。また、漢方では、トウキは「もの忘れ」に有効という伝承にしか過ぎな
かったことや、漢方薬のトウキシャクヤクサンが AD に有効との臨床試験がある 812)にもか
かわらず、実際には AD に使用されてこなかった理由も分かってきました。結局、漢方薬と
しては AD の周辺症状を改善する「抑肝散」があるだけです。INMⓇ176 に配合された韓国
トウキや ANM176Ⓡに配合されたガーデンアンゼリカには抗炎症作用がある特定クマリン
類 815)の含量が規格化されています。
ANM176Ⓡに配合されているβアミロイド神経毒性を抑制できる特定クマリン類を確保
する方法はノウハウで、一般には公開されていないことから、単にフェルラ酸とガーデンア
ンゼリカを配合した商品に ANM176Ⓡ を使用した商品と同等の効果を期待することはでき
ないと考えられます。ANM176Ⓡにはβアミロイドの神経毒性を十分に抑制できる量のフェ
ルラ酸が配合され、同時に配合されているβアミロイド神経毒性を抑制できる 12 種のクマ
リン類は、既に起こってしまっている炎症を抑制できる作用があります。フェルラ酸は摂取
した後 7-9 時間にピークとなります
。炎症が関係していると考えられている AD の進
778)
行を抑制するには、朝・晩 ANM176Ⓡを 2 回に分けて食前か食間に使用し、長期にわたって
炎症を抑制する必要があると考えられます。また、Yan 博士らが示したように、フェルラ酸
には即効力がなく、14 日前から継続して使い続けないと効果が発揮されません 234)
(図 58)。
このフェルラ酸の効果は、フェルラ酸が Keap1 阻害して Nrf2 を活性化する(89 ページ)こ
とからも、時間がかかることが窺えます。ANM176Ⓡ が AD の予防や治療に役立つかにつ
いて以下のようにまとめることができます。
1) ANM176Ⓡ のβアミロイド神経毒性抑制力がフェルラ酸より高いことは(図 69)
、
110
ANM176®が認知症の予防と改善に役立つ可能性について
AD の予防や治療に対する効果もフェルラ酸より ANM176Ⓡ の方が高いことを示し
ています。
2) 脳内のβアミロイドは正常な分泌物として様々な酵素によって分解され、一定濃度を
維持しています。高齢になって分解の能力が弱まって脳内で増えたβアミロイド
は、分子シャペロンなどタンパク質品質管理システムの加齢による影響などが加わ
り、βアミロイド・オリゴマーを形成します。βアミロイド・オリゴマーの中にあ
るβシート構造はシナプス機能に直接作用して神経作用を障害するだけでなく、酸
化ストレスを引き起こしてシナプスの機能などに影響を及ぼすと考えられます。
ANM176Ⓡ に含まれるフェルラ酸はストレス耐性を増強しβアミロイド・オリゴマ
ーのβシート構造による酸化ストレスを抑制して、神経作用を保護すると考えられ
ます。
3) 脳内で増加したβアミロイドが集積し、その集積したβアミロイドを処理する時、
高齢によるストレス応答の低下や他の要因が加わってストレス応答の閾値を超え、
炎症が起こると考えられますが、ANM176Ⓡ はβアミロイドの集積を減少させ、高
齢になって低下するストレス応答を強化して炎症が起こらないようにできる可能性
があります。また、AD に進行してしまった後でも、新たなβアミロイドの集積を抑
制し、既に起こっている炎症を抑制できる可能性があります。
4) ANM176Ⓡに配合されたフェルラ酸は、本来、食事で補給できるレベルで、また、配
合されたガーデンアンゼリカも常識的な量で、ガーデンアンゼリカ根抽出物に含ま
れる特定成分は極めて安全なレベルになるように管理されています。
このように ANM176Ⓡ に配合されているフェルラ酸は、Keap1 経路を通して高齢による
インスリンシグナリングとそれに伴うストレス耐性の低下を抑制し(図 72)、健忘や AD の
予防に役立つ可能性があります。また、ANM176Ⓡ に微量ながら一定量含まれる特定クマリ
ン類は、脳内炎症を抑制して、AD の改善に役立つ可能性があります。
インスリンシグナリング経路に異常がある場合、さまざまな疾患を引き起こします。例え
ば、インスリン抵抗性はインスリンシグナリングを低下させ高血糖症、糖尿病 144)あるいは
脂質代謝異常
826)を引き起こします。高齢になると
Nrf2 の働きが関与するストレス耐性が
低下し、AD や糖尿病だけでなく、様々な生活習慣病の発病に関係すると考えられます。
ANM176Ⓡは高齢になると低下するストレス耐性を強化し、かつ、安定的な抗炎症作用
があります。ANM176Ⓡに配合されたフェルラ酸は事前に Nrf2 による転写を活性化して
732)ストレス耐性を増強し、特定なクマリン類は炎症を鎮静化する抗炎症作用があります。
この 2 通りの効果によって、ANM176Ⓡ は AD の予防や改善に役立つ可能性があるだけで
なく、高血圧や糖尿病など様々な成人病の改善にも同時に役立つ可能性が考えられます。
事実、フェルラ酸が配合された ANM176Ⓡ はグルタチオンを増加させると予測されていま
したが、143 名のアルツハイマー病患者で ANM176Ⓡ を 9 ヵ月間使用した今回の試験で、
肝機能を示すγ-GTP が低下し 9 ヵ月間にわたり安定していました(図 73)。また、
111
ANM176®が認知症の予防と改善に役立つ可能性について
ANM176Ⓡは、AD を発症し
てからの改善にも有用な可
健忘とアルツハイマー病の発症模式図
能性があります。早期に
ストレス耐性のレベル
ANM176Ⓡ を使用すれば
βアミロイドの濃度
アルツハイマー病
AD の進行を抑制できる可能
健忘
性が高くなることが示唆さ
れましたが、しかし、かな
正常
り病状が進行してしまって
いる場合でも、あるいはす
でにドネペジルの効果が失
われていると思われる場合
80才
60才
40才
でも、一部は改善できる可
能性が示されています(図
71)。
ANM176®を使った場合の模式図
ANM176®を使わなかったストレス耐性のレベル
ANM176Ⓡ は、本来であ
ANM176®を使ったストレス耐性のレベル
れば食事で摂り得る成分を
アルツハイマー病
ANM176®を使わなかったβアミロイドの濃度
ANM176®を使ったβアミロイドの濃度
補給するもので、副作用も
健忘
全くなく、毎日使っても極
めて安全で長期の使用に耐
正常
えられます。一方、食事で
取り得る常識的な量でβア
ミロイドによる神経毒性を
抑制できるフェルラ酸や特
定なクマリン類が、今日の
食事では、ほとんど摂れて
いないと言えます。AD に限
40才
60才
80才
図 72.ANM176®はストレス耐性の低下を抑制する
ANM176®は、インスリンシグナル経路と独立した Keap1経路で、高齢
による Nrf2 の転写活性とストレス耐性の低下を抑制し、健忘とアルツ
ハイマー病の発症年齢を高齢側にシフトできる可能性があります。
らず様々な要因が関与する
生活習慣病全般について
も、一旦、これらの病気を
発症すると根本的な治療
は、現状ではかなり困難で
す。その理由は、体に本来
ある恒常性維持のシステム
がなかなか元の状態に戻ら
ないためだと考えられるか
らです。以上に述べたよう
図 73.ANM176®によるγ-GTP に対する影響
9 ヵ月間飲用し、最後まで試験に参加した AD 患者 98 名のγ-GTP の群
全体の最大値、最小値と群の平均値の推移
112
ANM176®が認知症の予防と改善に役立つ可能性について
に、高齢になると低下するストレス耐性を強化して、AD に限らず糖尿病などの様々な生
活習慣病の予防にも役立つ可能性がある ANM176Ⓡ を、AD 患者の補助食品としてだけで
なく、AD も含めた様々な生活習慣病の予防としても広めることは、社会的にも意義があ
ると思っています。
最後に
タンパク質だけでも 3 万種が織りなす生命の仕組みは、当然のことながら全貌は解明さ
れていません。また、解明されていることについても、その全部を網羅することは不可能で
す。本紙で述べたポイントは、高齢化に伴う生活習慣病は共通してホメオスタシス(恒常性)
の破綻が関係し、そして、この破綻の原因として高齢によるストレス耐性の低下や生活環境
の変化による影響について考察し、多くの人が理解できるように努めて解説したものです。
2015/6/25
113
ANM176®が認知症の予防と改善に役立つ可能性について
(本文中の略号)( )内の数字は初めて登場するページ
Ach(acetyl choline:アセルコリン、35)、AChE(Acetylcholinesterase:アセルコリンエステ
ラーゼ、76)、AD(Alzheimer’s disease;アルツハイマー病、1)、Apo(Apo lipoprotein:アポ
リポタンパク質、61)、ApoJ(apolipoprotein J :アポリポタンパク J、51)、APP(amyloid
precursor protein:アミロイド前駆タンパク質、46)、ARE(antioxidant response element:
抗酸化応答エレメント、28)、ASK(Apotosis signaling kinase:アポトーシスシグナリングキ
ナーゼ、35)、BBB(blood brain barrier :血液脳関門、9)、COX(cyclooxygenese:プロスタ
グランジン合成酵素、30)、CRMP2(Collapsin response mediating protein 2:コラプシン反
応媒介タンパク質 2 、41)、CSF(cerebrospinal fluid 脳髄液:9)、DLB(dementia with lewy
bodies:レビー小体型認知症、6)、DM(Diabetes Mellitus:糖尿病、2)、eIF(eukaryotic initiation
factor:真核生物開始因子、20)、EPSP(excitatory postsynaptic potential:興奮性シナプス後
電位、37)、ER(endoplasmic reticulum:小胞体、20)、FAD(familial Alzheimer's disease:
家族性アルツハイマー病、44)、FTD(frontotemporal dementia:前頭側頭型認知症、6)、GFAP
(glial fibllial acids protein :グリア線維性酸性タンパク質、29)、GPCR(protein-coupled
receptor G:タンパク質共役受容体、15)、GS(glycogen synthetase:グリコーゲン合成酵素、
70)、GSH(glutathione:グルタチオン、27)、GSK3β(glycogen synthase kinase 3β:グ
リコーゲン合成酵素キナーゼ 3β、70)、GST(glutathione S-transferase :グルタチオン S 転移酵素性物、27)、HDL(high-density lipoprotein:高比重リポタンパク質、10)、HO-1(Heme
oxygenase-1:ヘムオキシゲナーゼ1、33)、Hsp(heat shock protein:熱ショックタンパク質、
50)、IDE(insulin degrading enzyme:インスリン分解酵素、53)、LDL(low-density lipoprotein:
低比重リポタンパク質、10)、IGF(insulin-like growth factor:インスリン様成長因子、10)、
IL(Interleukin:インターロイキン、13)、IR(insulin receptor:インスリン受容体、10)、IRS
(insulin receptor substrate:インスリン受容体基質、70)、LPS(lipopolysaccharide:リポ多
糖類、30)
、LTP(long term potentiation:長期増強、36)、MAP(microtubule-associated protein:
微小管結合タンパク質、41)、MAPK(mitogen-activated protein kinase:分裂促進因子活性化
タンパク質キナーゼ、16)、MCI(mild cognitive impairmenrt:軽度認知障害、5)、MCT(Mono
Carbonate Transporter:モノカルボン酸トランスポーター、10)、MT(microtubule:微小管、
40)、NADPH(nicotinamide adenine dinucleotide phosphate H:ニコチンアミドアデニンジ
ヌクレオチドリン酸、26)、NEP(neprilysin:ネプリライシン、53)
、NFT(neurofibrillary
tangle:神経原線維変化、44)、NSAID(non-steroidal anti-inflammatory drugs:非ステロイ
ド抗炎症剤、30)、PD(Parkinson’s disease:パーキンソン病、6)、PI3K(phosphatidyl-inositol
3-kinase:ホスファチジルイノシトール 3-キナーゼ、17)、PIP3(phosphatidyl-inositol 3,4,5trisphosphate:ホスファチジルイノシトール(3,4,5)三リン酸、13)、PSD(postsynaptic density:
後シナプス肥厚部、38)、RTK(receptor tyrosine kinase:チロシンキナーゼ型受容体、14)、
SDAT(senile dementia of Alzheimer type:老年性アルツハイマー型認知症、2)、SNP(singlenucleotide polymorphism:一塩基多型、19)
、TLR(Toll like receptor:Toll 様受容体、15)
TZD(thiazolidinedione:チアゾリジン、77)、UPR(unfold protein response:小胞体ストレ
ス応答、25)、VD(vascular dementia: 脳血管性認知症 、5)、4EBP(eIF4E binding protein:
eIF4E 結合タンパク質、21)
文献
1) 博野信次 [痴呆症診療における諸問題点] インターネット医科大学 ( (n.d.)).
2) Petersen RC, Smith GE, Waring SC, Ivnik RJ, Tangalos EG, Kokmen E [Mild cognitive impairment: clinical
characterization and outcome.] Arch. Neurol. (1999) 56:303–8.
3) 水上勝義 [軽度認知障害(MCI)症例にはどう対応すべきか?] 精神神經學雜誌 = Psychiatr. Neurol. Jpn. 日本精神神経学会
(2009) 111(1):26–30.
4) 下方浩史 [我が国の疫学統計] 日本臨床増刊号痴呆症候学 3 (2004) 62(増刊号):121–5.
5) 朝田隆 [都市部における認知症の有病率と認知症の生活機能障害への対応] 厚生労働省 平成 23 年度~24 年度総合研究報告
6) 涌谷陽介 [アルツハイマー病患者は増加している] 第 19 回「大学と科学」アルツハイマー病:治療の可能性を探る B セッ
ション●疫学調査から (2004).
7) Kosaka K [Diffuse Lewy body disease.] Neuropathology (2000) 20 Suppl:S73–8.
8) 若林孝一 [パーキンソン病(PD)] 日本神経病理学会 (2005).
9) Williams RW, Herrup K [The control of neuron number.] Annu. Rev. Neurosci. (1988) 11:423–53.
10) 深水昭吉 [時空間的な細胞内ネットワークの機能修飾] 細胞工学 (2002) 21(3):266–689.
11) Coba MP, Pocklington AJ, Collins MO, Kopanitsa M V, Uren RT, Swamy S, Croning MDR, Choudhary JS, Grant SGN
[Neurotransmitters drive combinatorial multistate postsynaptic density networks.] Sci. Signal. (2009) 2:ra19.
12) Kanda T [Blood-brain barrier and neurological disorders] J. Japan Epilepsy Soc. (2014) 32(1):47–50.
13) Goldstein GW, Betz AL [The blood-brain barrier.] Sci. Am. (1986) 255(3):74–83.
14) Zlokovic B V., Apuzzo MLJ [Cellular and Molecular Neurosurgery: Pathways from Concept to Reality-Part I: Target
Disorders and Concept Approaches to Gene Therapy of the Central Nervous System] Neurosurgery (1997) 40(4):789–
804.
15) Pardridge WM [Drug transport across the blood-brain barrier.] J. Cereb. Blood Flow Metab. (2012) 32:1959–72.
16) Zloković B V, Segal MB, Begley DJ, Davson H, Rakić L [Permeability of the blood-cerebrospinal fluid and blood-brain
barriers to thyrotropin-releasing hormone.] Brain Res. (1985) 358:191–9.
17) Pardridge WM, Eisenberg J, Yang J [Human blood-brain barrier insulin receptor.] J. Neurochem. (1985) 44:1771–8.
18) Duffy KR, Pardridge WM, Rosenfeld RG [Human blood-brain barrier insulin-like growth factor receptor.] Metabolism.
(1988) 37:136–40.
19) Golden PL, Maccagnan TJ, Pardridge WM [Human blood-brain barrier leptin receptor. Binding and endocytosis in
isolated human brain microvessels] J. Clin. Invest. (1997) 99:14–8.
20) Pardridge WM, Eisenberg J, Yang J [Human blood-brain barrier transferrin receptor.] Metabolism. (1987) 36:892–5.
21) 佐藤隆一郎 [脂質代謝制御の中心的役割を演じる転写因子 SREBP の分子基盤] Nutr. Rev. (2008) June.
22) Jaeger S, Pietrzik CU [Functional role of lipoprotein receptors in Alzheimer’s disease] Curr Alzheimer Res (2008)
5:15–25.
23) Kanekiyo T, Bu G [The low-density lipoprotein receptor-related protein 1 and amyloid-β clearance in Alzheimer’s
disease.] Front. Aging Neurosci. (2014) 6:93.
24) Dieckmann M, Dietrich MF, Herz J [Lipoprotein receptors-an evolutionarily ancient multifunctional receptor family]
Biol. Chem. (2010), 1341–63.
25) Boucher P, Herz J [Signaling through LRP1: Protection from atherosclerosis and beyond] Biochem. Pharmacol. (2011),
1–5.
26) Wang X, Lee S-R, Arai K, Lee S-R, Tsuji K, Rebeck GW, Lo EH [Lipoprotein receptor-mediated induction of matrix
metalloproteinase by tissue plasminogen activator.] Nat. Med. (2003) 9:1313–7.
27) Van Uden E, Mallory M, Veinbergs I, Alford M, Rockenstein E, Masliah E [Increased extracellular amyloid deposition
and neurodegeneration in human amyloid precursor protein transgenic mice deficient in receptor-associated protein.]
J. Neurosci. (2002) 22:9298–304.
28) Deane R, Bell RD, Sagare A, Zlokovic B V [Clearance of amyloid-beta peptide across the blood-brain barrier:
implication for therapies in Alzheimer’s disease.] CNS Neurol. Disord. Drug Targets (2009) 8:16–30.
29) Sagare AP, Deane R, Zlokovic B V [Low-density lipoprotein receptor-related protein 1: a physiological Aβ homeostatic
mechanism with multiple therapeutic opportunities.] Pharmacol. Ther. (2012) 136(1):94–105.
30) Burchett SA [Regulators of G protein signaling: a bestiary of modular protein binding domains.] J. Neurochem. (2000)
75:1335–51.
31) Rojas AM, Fuentes G, Rausell A, Valencia A [The Ras protein superfamily: Evolutionary tree and role of conserved
amino acids] J. Cell Biol. (2012), 189–201.
32) Clerk A, Sugden PH [Small guanine nucleotide-binding proteins and myocardial hypertrophy.] Circ. Res. (2000)
86:1019–23.
33) Etienne-Manneville S, Hall A [Rho GTPases in cell biology.] Nature (2002) 420:629–35.
34) Zarich N, Oliva JL, Martínez N, Jorge R, Ballester A, Gutiérrez-Eisman S, García-Vargas S, Rojas JM [Grb2 is a
negative modulator of the intrinsic Ras-GEF activity of hSos1.] Mol. Biol. Cell (2006) 17:3591–7.
35) Ojaniemi M, Vuori K [Epidermal growth factor modulates tyrosine phosphorylation of p130Cas. Involvement of
phosphatidylinositol 3’-kinase and actin cytoskeleton] J. Biol. Chem. (1997) 272:25993–8.
36) Chen Z, Gibson TB, Robinson F, Silvestro L, Pearson G, Xu BE, Wright A, Vanderbilt C, Cobb MH [MAP kinases]
Chem. Rev. (2001) 101:2449–76.
37) Schlessinger J [Cell signaling by receptor tyrosine kinases.] Cell (2000) 103:211–25.
38) Wang X, Martindale JL, Liu Y, Holbrook NJ [The cellular response to oxidative stress: influences of mitogen-activated
protein kinase signalling pathways on cell survival.] Biochem. J. (1998) 333 ( Pt 2:291–300.
39) Chang L, Karin M [Mammalian MAP kinase signalling cascades.] Nature (2001) 410:37–40.
40) Kyriakis JM, Avruch J [Mammalian mitogen-activated protein kinase signal transduction pathways activated by
stress and inflammation.] Physiol. Rev. (2001) 81:807–69.
41) Zhang D, Hu X, Qian L, Chen S-H, Zhou H, Wilson B, Miller DS, Hong J-S [Microglial MAC1 receptor and PI3K are
essential in mediating β-amyloid peptide-induced microglial activation and subsequent neurotoxicity.] J.
Neuroinflammation (2011) 8:3.
42) Sata M [Phosphatidylinositol 3-Kinase: A Key Regulator of Vascular Tone?] Circ. Res. (2002) 91(4):273–5.
I
43) Lee JS, Kim JH, Jang IH, Kim HS, Han JM, Kazlauskas A, Yagisawa H, Suh P-G, Ryu SH [Phosphatidylinositol
(3,4,5)-trisphosphate specifically interacts with the phox homology domain of phospholipase D1 and stimulates its
activity.] J. Cell Sci. (2005) 118:4405–13.
44) Stephens L, Anderson K, Stokoe D, Erdjument-Bromage H, Painter GF, Holmes AB, Gaffney PR, Reese CB,
McCormick F, Tempst P, Coadwell J, Hawkins PT [Protein kinase B kinases that mediate phosphatidylinositol 3,4,5trisphosphate-dependent activation of protein kinase B.] Science (1998) 279:710–4.
45) Vanhaesebroeck B, Alessi DR [The PI3K-PDK1 connection: more than just a road to PKB.] Biochem. J. (2000) 346 Pt
3:561–76.
46) Brazil DP, Hemmings BA [Ten years of protein kinase B signalling: A hard Akt to follow] Trends Biochem. Sci. (2001),
657–64.
47) Pickart CM, Fushman D [Polyubiquitin chains: Polymeric protein signals] Curr. Opin. Chem. Biol. (2004), 610–6.
48) Chiu Y-H, Zhao M, Chen ZJ [Ubiquitin in NF-kappaB signaling.] Chem. Rev. (2009) 109(4):1549–60.
49) Lander ES, Linton LM, Birren B, Nusbaum C, Zody MC, Baldwin J, Devon K, Dewar K, Doyle M, FitzHugh W, Funke
R, Gage D, Harris K, Heaford A, Howland J, Kann L, Lehoczky J, LeVine R, McEwan P, McKernan K, Meldrim J,
Mesirov JP, Miranda C, Morris W, Naylor J, Raymond C, Rosetti M, Santos R, Sheridan A, et al. [Initial sequencing
and analysis of the human genome.] Nature (2001) 409:860–921.
50) Thannickal VJ, Fanburg BL [Reactive oxygen species in cell signaling.] Am. J. Physiol. Lung Cell. Mol. Physiol. (2000)
279:L1005–28.
51) 山崎創 [核タンパク質 IκB-ζ を介した炎症性遺伝子の発現調節] 生化学 (2008) 80(8):758–62.
52) Le-Niculescu H, Bonfoco E, Kasuya Y, Claret FX, Green DR, Karin M [Withdrawal of survival factors results in
activation of the JNK pathway in neuronal cells leading to Fas ligand induction and cell death.] Mol. Cell. Biol. (1999)
19:751–63.
53) Serhan CN [Resolution phase of inflammation: novel endogenous anti-inflammatory and proresolving lipid mediators
and pathways.] Annu. Rev. Immunol. (2007) 25:101–37.
54) 日野真一郎 [小胞体ストレス誘導性オートファジーの制御による神経変性疾患の治療戦略開発] 上原記念生命科学財団研究
報告集 (2009) 23:133.
55) Ron D, Walter P [Signal integration in the endoplasmic reticulum unfolded protein response.] Nat. Rev. Mol. Cell Biol.
(2007) 8:519–29.
56) Kim I, Xu W, Reed JC [Cell death and endoplasmic reticulum stress: disease relevance and therapeutic opportunities.]
Nat. Rev. Drug Discov. (2008) 7:1013–30.
57) Lee AS [GRP78 induction in cancer: Therapeutic and prognostic implications] Cancer Res. (2007), 3496–9.
58) Yoshida H, Matsui T, Yamamoto A, Okada T, Mori K [XBP1 mRNA is induced by ATF6 and spliced by IRE1 in
response to ER stress to produce a highly active transcription factor] Cell (2001) 107:881–91.
59) Aderem A, Underhill DM [Mechanisms of phagocytosis in macrophages.] Annu. Rev. Immunol. (1999) 17:593–623.
60) Aderem A [How to eat something bigger than your head.] Cell (2002) 110:5–8.
61) Condliffe AM, Kitchen E, Chilvers ER [Neutrophil priming: pathophysiological consequences and underlying
mechanisms.] Clin. Sci. (Lond). (1998), 461–71.
62) 古澤新平 [臨床検査技術学] (2003).
63) Whitlock JP [Induction of cytochrome P4501A1.] Annu. Rev. Pharmacol. Toxicol. (1999) 39:103–25.
64) Moi P, Chan K, Asunis I, Cao A, Kan YW [Isolation of NF-E2-related factor 2 (Nrf2), a NF-E2-like basic leucine zipper
transcriptional activator that binds to the tandem NF-E2/AP1 repeat of the beta-globin locus control region.] Proc.
Natl. Acad. Sci. U. S. A. (1994) 91:9926–30.
65) Chen X-L, Varner SE, Rao AS, Grey JY, Thomas S, Cook CK, Wasserman MA, Medford RM, Jaiswal AK, Kunsch C
[Laminar flow induction of antioxidant response element-mediated genes in endothelial cells. A novel antiinflammatory mechanism.] J. Biol. Chem. (2003) 278:703–11.
66) 伊東健 [Nrf2 酸化ストレス応答系による病態制御] 生化學. 日本生化学会 (2009) 81(6):447–55.
67) Jaiswal AK [Antioxidant response element] Biochem. Pharmacol. (1994) 48(3):439–44.
68) Wasserman WW, Fahl WE [Functional antioxidant responsive elements.] Proc. Natl. Acad. Sci. U. S. A. (1997)
94:5361–6.
69) Sykiotis GP, Bohmann D [Stress-activated cap’n'collar transcription factors in aging and human disease.] Sci. Signal.
(2010) 3:re3.
70) 山本雅之 [環境適応の遺伝子発現制御機構 Regulatory Mechanisms of Gene Expression for Adaptation] 開始未来開拓学
術研究推進事業研究プロジェクト (1998).
71) 榮長裕晴, 吉原栄治, 松尾禎之, 淀井淳司 [酸化ストレスとレドックス制御∼ タンパク質の酸化的修飾と活性調整∼]
plaza.umin.ac.jp (2009) 32(4):265–72.
72) 北川一夫, 八木田佳樹, 佐々木勉, 佐古田三郎 [神経細胞死研究の成果と臨床応用] 脳卒中 (2008) 30(6):862–8.
73) Kitagawa K [脳梗塞:炎症の制御を標的とした治療戦略の可能性 Anti-inflammation strategy: potential brain protection
in cerebral ischemia] Folia Pharmacol. Jpn. 公益社団法人 日本薬理学会 (2009) 134(4):202–6.
74) Sasaki T, Kitagawa K, Yamagata K, Takemiya T, Tanaka S, Omura-Matsuoka E, Sugiura S, Matsumoto M, Hori M
[Amelioration of hippocampal neuronal damage after transient forebrain ischemia in cyclooxygenase-2-deficient mice.]
J. Cereb. Blood Flow Metab. (2004) 24:107–13.
75) Ma XL, Kumar S, Gao F, Louden CS, Lopez BL, Christopher TA, Wang C, Lee JC, Feuerstein GZ, Yue TL [Inhibition
of p38 mitogen-activated protein kinase decreases cardiomyocyte apoptosis and improves cardiac function after
myocardial ischemia and reperfusion.] Circulation (1999) 99:1685–91.
76) Sucher R, Gehwolf P, Kaier T, Hermann M, Maglione M, Oberhuber R, Ratschiller T, Kuznetsov A V, Bösch F, Kozlov
A V, Ashraf MI, Schneeberger S, Brandacher G, Ollinger R, Margreiter R, Troppmair J [Intracellular signaling
pathways control mitochondrial events associated with the development of ischemia/ reperfusion-associated damage.]
Transpl. Int. (2009) 22:922–30.
77) Shioda N, Han F, Moriguchi S, Fukunaga K [Constitutively active calcineurin mediates delayed neuronal death
through Fas-ligand expression via activation of NFAT and FKHR transcriptional activities in mouse brain ischemia]
J. Neurochem. (2007) 102:1506–17.
II
78) Shioda N, Ishigami T, Han F, Moriguchi S, Shibuya M, Iwabuchi Y, Fukunaga K [Activation of phosphatidylinositol 3kinase/protein kinase B pathway by a vanadyl compound mediates its neuroprotective effect in mouse brain ischemia]
Neuroscience (2007) 148:221–9.
79) Tanaka H, Katoh A, Oguro K, Shimazaki K, Gomi H, Itohara S, Masuzawa T, Kawai N [Disturbance of hippocampal
long-term potentiation after transient ischemia in GFAP deficient mice] J. Neurosci. Res. (2002) 67:11–20.
80) Eng LF [Glial fibrillary acidic protein (GFAP): the major protein of glial intermediate filaments in differentiated
astrocytes.] J. Neuroimmunol. (1985) 8(4-6):203–14.
81) Nielsen AL, Holm IE, Johansen M, Bonven B, Jørgensen P, Jørgensen AL [A new splice variant of glial fibrillary acidic
protein, GFAP epsilon, interacts with the presenilin proteins.] J. Biol. Chem. (2002) 277:29983–91.
82) Doyle SE, Vaidya SA, O’Connell R, Dadgostar H, Dempsey PW, Wu TT, Rao G, Sun R, Haberland ME, Modlin RL,
Cheng G [IRF3 Mediates a TLR3/TLR4-Specific Antiviral Gene Program] Immunity (2002) 17:251–63.
83) Ramirez-Carrozzi VR, Nazarian AA, Li CC, Gore SL, Sridharan R, Imbalzano AN, Smale ST [Selective and
antagonistic functions of SWI/SNF and Mi-2beta nucleosome remodeling complexes during an inflammatory response]
Genes Dev. (2006) 20:282–96.
84) Kono H, Rock KL [How dying cells alert the immune system to danger.] Nat. Rev. Immunol. (2008) 8:279–89.
85) Crutcher KA, Gendelman HE, Kipnis J, Perez-Polo JR, Perry VH, Popovich PG, Weaver LC [Debate: “Is increasing
neuroinflammation beneficial for neural repair?”] J. Neuroimmune Pharmacol. (2006), 195–211.
86) Popovich PG, Longbrake EE [Can the immune system be harnessed to repair the CNS?] Nat. Rev. Neurosci. (2008)
9:481–93.
87) Frank-Cannon TC, Alto LT, McAlpine FE, Tansey MG [Does neuroinflammation fan the flame in neurodegenerative
diseases?] Mol. Neurodegener. (2009) 4:47.
88) Rivest S [Regulation of innate immune responses in the brain.] Nat. Rev. Immunol. (2009) 9:429–39.
89) McGeer PL, McGeer EG [Inflammation of the brain in Alzheimer’s disease: implications for therapy.] J. Leukoc. Biol.
(1999) 65:409–15.
90) 尾内康臣 [PET をもちいた脳ミクログリアの画像化] 臨床神経学 (2009) 49:925―928.
91) Mabuchi T, Kitagawa K, Ohtsuki T, Kuwabara K, Yagita Y, Yanagihara T, Hori M, Matsumoto M [Contribution of
microglia/macrophages to expansion of infarction and response of oligodendrocytes after focal cerebral ischemia in
rats.] Stroke. (2000) 31:1735–43.
92) Kalimo H, Paljärvi L, Vapalahti M [The early ultrastructural alterations in the rabbit cerebral and cerebellar cortex
after compression ischaemia.] Neuropathol. Appl. Neurobiol. (1979) 5(3):211–23.
93) Thun MJ, Henley SJ, Patrono C [Nonsteroidal anti-inflammatory drugs as anticancer agents: mechanistic,
pharmacologic, and clinical issues.] J. Natl. Cancer Inst. (2002) 94:252–66.
94) Morimoto K, Shirata N, Taketomi Y, Tsuchiya S, Segi-Nishida E, Inazumi T, Kabashima K, Tanaka S, Murakami M,
Narumiya S, Sugimoto Y [Prostaglandin E2-EP3 signaling induces inflammatory swelling by mast cell activation.] J.
Immunol. (2014) 192(3):1130–7.
95) Kurumbail RG, Kiefer JR, Marnett LJ [Cyclooxygenase enzymes: catalysis and inhibition.] Curr. Opin. Struct. Biol.
(2001) 11:752–60.
96) Cory AH, Owen TC, Barltrop JA, Cory JG [Use of an aqueous soluble tetrazolium/formazan assay for cell growth
assays in culture.] Cancer Commun. (1991) 3:207–12.
97) Marshall NJ, Goodwin CJ, Holt SJ [A critical assessment of the use of microculture tetrazolium assays to measure cell
growth and function.] Growth Regul. (1995) 5:69–84.
98) Kaneko I, Yamada N, Sakuraba Y, Kamenosono M, Tutumi S [Suppression of mitochondrial succinate dehydrogenase,
a primary target of beta-amyloid, and its derivative racemized at Ser residue.] J. Neurochem. (1995) 65:2585–93.
99) Berridge M V, Tan AS [Characterization of the cellular reduction of 3-(4,5-dimethylthiazol-2-yl)-2,5diphenyltetrazolium bromide (MTT): subcellular localization, substrate dependence, and involvement of mitochondrial
electron transport in MTT reduction.] Arch. Biochem. Biophys. (1993) 303:474–82.
100) Goodwin CJ, Holt SJ, Riley PA, Downes S, Marshall NJ [Growth hormone-responsive DT-diaphorase-mediated
bioreduction of tetrazolium salts.] Biochem. Biophys. Res. Commun. (1996) 226:935–41.
101) Green TR, Pratt KL [Detection and isolation of the NADPH-binding protein of the NADPH:O2 oxidoreductase
complex of human neutrophils] J. Biol. Chem. (1990) 265:19324–9.
102) Bernas T, Dobrucki J [Mitochondrial and nonmitochondrial reduction of MTT: Interaction of MTT with TMRE, JC-1,
and NAO mitochondrial fluorescent probes] Cytometry (2002) 47:236–42.
103) Bamberger ME, Harris ME, McDonald DR, Husemann J, Landreth GE [A cell surface receptor complex for fibrillar
beta-amyloid mediates microglial activation.] J. Neurosci. (2003) 23:2665–74.
104) Babior BM [NADPH oxidase: an update.] Blood. American Society of Hematology (1999) 93(5):1464–76.
105) Hall A [G proteins and small GTPases: distant relatives keep in touch.] Science (1998) 280(5372):2074–5.
106) Takenawa T, Miki H [WASP and WAVE family proteins: key molecules for rapid rearrangement of cortical actin
filaments and cell movement.] J. Cell Sci. (2001) 114:1801–9.
107) Kitamura Y, Shibagaki K, Takata K, Tsuchiya D, Taniguchi T, Gebicke-Haerter PJ, Miki H, Takenawa T,
Shimohama S [Involvement of Wiskott-Aldrich syndrome protein family verprolin-homologous protein (WAVE) and
Rac1 in the phagocytosis of amyloid-beta(1-42) in rat microglia.] J. Pharmacol. Sci. (2003) 92:115–23.
108) Coxon A, Rieu P, Barkalow FJ, Askari S, Sharpe AH, von Andrian UH, Arnaout MA, Mayadas TN [A novel role for
the beta 2 integrin CD11b/CD18 in neutrophil apoptosis: a homeostatic mechanism in inflammation.] Immunity
(1996) 5(6):653–66.
109) Persaud-Sawin D-A, Banach L, Harry GJ [Raft aggregation with specific receptor recruitment is required for
microglial phagocytosis of Abeta42.] Glia (2009) 57(3):320–35.
110) Moncayo J, de Freitas GR, Bogousslavsky J, Altieri M, van Melle G [Do transient ischemic attacks have a
neuroprotective effect?] Neurology (2000) 54:2089–94.
111) Kitagawa K, Matsumoto M, Tagaya M, Kuwabara K, Hata R, Handa N, Fukunaga R, Kimura K, Kamada T
[Hyperthermia-induced neuronal protection against ischemic injury in gerbils.] J. Cereb. Blood Flow Metab. (1991)
11:449–52.
112) Van der Worp HB, Sena ES, Donnan GA, Howells DW, Macleod MR [Hypothermia in animal models of acute
ischaemic stroke: a systematic review and meta-analysis.] Brain (2007) 130:3063–74.
III
113) Li GC, Werb Z [Correlation between synthesis of heat shock proteins and development of thermotolerance in Chinese
hamster fibroblasts.] Proc. Natl. Acad. Sci. U. S. A. (1982) 79:3218–22.
114) 川口昌彦, 古家仁 [脳虚血に対するプレコンディショニング] Anet. Mail (2009) 13(1):14–8.
115) Tohyama M, Ogawa T [Stress response of cultured astrocytes to hypoxia/reoxygenation] Nippon yakurigaku zasshi.
Folia Pharmacol. Jpn. (1997) 109(3):145–51.
116) Hori O, Ichinoda F, Tamatani T, Yamaguchi A, Sato N, Ozawa K, Kitao Y, Miyazaki M, Harding HP, Ron D,
Tohyama M, M Stern D, Ogawa S [Transmission of cell stress from endoplasmic reticulum to mitochondria: enhanced
expression of Lon protease.] J. Cell Biol. (2002) 157:1151–60.
117) Bailey D, O’Hare P [Transmembrane bZIP transcription factors in ER stress signaling and the unfolded protein
response.] Antioxid. Redox Signal. (2007) 9:2305–21.
118) Imaizumi K, Tohyama M [The regulation of unfolded protein response by OASIS, a transmembrane bZIP
transcription factor, in astrocytes] Nippon yakurigaku zasshi. Folia Pharmacol. Jpn. (2004) 124(6):383–90.
119) Luo S, Baumeister P, Yang S, Abcouwer SF, Lee AS [Induction of Grp78/BiP by translational block: Activation of the
Grp78 promoter by ATF4 through an upstream ATF/CRE site independent of the endoplasmic reticulum stress
elements] J. Biol. Chem. (2003) 278:37375–85.
120) Shih AY, Johnson DA, Wong G, Kraft AD, Jiang L, Erb H, Johnson JA, Murphy TH [Coordinate regulation of
glutathione biosynthesis and release by Nrf2-expressing glia potently protects neurons from oxidative stress.] J.
Neurosci. (2003) 23:3394–406.
121) Lee J-M, Shih AY, Murphy TH, Johnson JA [NF-E2-related factor-2 mediates neuroprotection against mitochondrial
complex I inhibitors and increased concentrations of intracellular calcium in primary cortical neurons.] J. Biol. Chem.
(2003) 278:37948–56.
122) Motohashi H, O’Connor T, Katsuoka F, Engel JD, Yamamoto M [Integration and diversity of the regulatory network
composed of Maf and CNC families of transcription factors] Gene (2002), 1–12.
123) 伊東健, 山本雅之 [異物代謝系第 2 相酵素群の発現制御機構 (環境応答・適応の分子機構) – (動物における環境適応の分
子機構)] 蛋白質核酸酵素. 共立出版 (1999) 44(15):2370–6.
124) Motohashi H, Yamamoto M [Nrf2-Keap1 defines a physiologically important stress response mechanism] Trends Mol.
Med. (2004), 549–57.
125) Thimmulappa RK, Lee H, Rangasamy T, Reddy SP, Yamamoto M, Kensler TW, Biswal S [Nrf2 is a critical regulator
of the innate immune response and survival during experimental sepsis.] J. Clin. Invest. (2006) 116:984–95.
126) Gong JS, Kobayashi M, Hayashi H, Zou K, Sawamura N, Fujita SC, Yanagisawa K, Michikawa M [Apolipoprotein E
(ApoE) isoform-dependent lipid release from astrocytes prepared from human ApoE3 and ApoE4 knock-in mice.] J.
Biol. Chem. (2002) 277:29919–26.
127) Maines MD [The heme oxygenase system and its functions in the brain.] Cell. Mol. Biol. (2000), 573–85.
128) Chen K, Gunter K, Maines MD [Neurons overexpressing heme oxygenase-1 resist oxidative stress-mediated cell
death.] J. Neurochem. (2000) 75:304–13.
129) Maines MD [The heme oxygenase system: a regulator of second messenger gases.] Annu. Rev. Pharmacol. Toxicol.
(1997) 37:517–54.
130) Chen J, Tu Y, Moon C, Nagata E, Ronnett G V [Heme oxygenase-1 and heme oxygenase-2 have distinct roles in the
proliferation and survival of olfactory receptor neurons mediated by cGMP and bilirubin, respectively.] J. Neurochem.
(2003) 85:1247–61.
131) Kapinya KJ, Harms U, Harms C, Blei K, Katchanov J, Dirnagl U, Hörtnagl H [Role of NAD(P)H:quinone
oxidoreductase in the progression of neuronal cell death in vitro and following cerebral ischaemia in vivo] J.
Neurochem. (2003) 84(5):1028–39.
132) Heydari AR, Wu B, Takahashi R, Strong R, Richardson A [Expression of heat shock protein 70 is altered by age and
diet at the level of transcription.] Mol. Cell. Biol. (1993) 13:2909–18.
133) Masoro EJ [Hormesis and the antiaging action of dietary restriction] Exp. Gerontol. (1998), 61–6.
134) Malinin NL, West XZ, Byzova T V. [Oxidation as “The Stress of Life”] Aging (Albany. NY). (2011) 3(9):906–10.
135) Sena LA, Chandel NS [Physiological roles of mitochondrial reactive oxygen species] Mol. Cell (2012), 158–66.
136) Groeger G, Quiney C, Cotter TG [Hydrogen peroxide as a cell-survival signaling molecule.] Antioxid. Redox Signal.
(2009) 11(11):2655–71.
137) Knoefler D, Thamsen M, Koniczek M, Niemuth NJ, Diederich AK, Jakob U [Quantitative In Vivo Redox Sensors
Uncover Oxidative Stress as an Early Event in Life] Mol. Cell (2012) 47(5):767–76.
138) Del Bello B, Paolicchi A, Comporti M, Pompella A, Maellaro E [Hydrogen peroxide produced during gamma-glutamyl
transpeptidase activity is involved in prevention of apoptosis and maintainance of proliferation in U937 cells.] FASEB
J. (1999) 13:69–79.
139) Clément M V, Stamenkovic I [Superoxide anion is a natural inhibitor of FAS-mediated cell death.] EMBO J. (1996)
15:216–25.
140) Burdon RH [Superoxide and hydrogen peroxide in relation to mammalian cell proliferation] Free Radic. Biol. Med.
(1995) 18:775–94.
141) Suzuki YJ, Forman HJ, Sevanian A [Oxidants as stimulators of signal transduction] Free Radic. Biol. Med. (1996),
269–85.
142) Manning G, Whyte DB, Martinez R, Hunter T, Sudarsanam S [The protein kinase complement of the human
genome.] Science (2002) 298:1912–34.
143) Hotamisligil G [Role of endoplasmic reticulum stress and c-Jun NH2-terminal kinase pathways in inflammation and
origin of obesity and diabetes] Diabetes (2005) 54 Suppl 2:S73–8.
144) 鎌田勝雄 [糖尿病の基礎知識] (2010).
145) Salminen A, Kauppinen A, Suuronen T, Kaarniranta K, Ojala J [ER stress in Alzheimer’s disease: a novel neuronal
trigger for inflammation and Alzheimer's pathology.] J. Neuroinflammation (2009) 6:41.
146) Sekine Y, Takeda K, Ichijo H [The ASK1-MAP kinase signaling in ER stress and neurodegenerative diseases.] Curr.
Mol. Med. (2006) 6:87–97.
147) O’Donnell E, Vereker E, Lynch MA [Age-related impairment in LTP is accompanied by enhanced activity of stressactivated protein kinases: analysis of underlying mechanisms.] Eur. J. Neurosci. (2000) 12:345–52.
148) Fujino G, Noguchi T, Takeda K, Ichijo H [Thioredoxin and protein kinases in redox signaling] Semin. Cancer Biol.
(2006), 427–35.
IV
149) Hattori K, Naguro I, Runchel C, Ichijo H [The roles of ASK family proteins in stress responses and diseases.] Cell
Commun. Signal. (2009) 7:9.
150) Hayakawa T, Matsuzawa A, Noguchi T, Takeda K, Ichijo H [The ASK1-MAP kinase pathways in immune and stress
responses.] Microbes Infect. (2006) 8:1098–107.
151) Ni M, Lee AS [ER chaperones in mammalian development and human diseases] FEBS Lett. (2007), 3641–51.
152) Yoshida H [ER stress and diseases] FEBS J. (2007), 630–58.
153) Cajal SRY [The Croonian Lecture: La Fine Structure des Centres Nerveux] Proc. R. Soc. London (1894), 444–68.
154) Hebb DO [The organization of behavior: a neuropsychological theory] Sci. Educ. (1949) 44:335.
155) Pizzorusso T, Medini P, Berardi N, Chierzi S, Fawcett JW, Maffei L [Reactivation of ocular dominance plasticity in
the adult visual cortex.] Science (2002) 298(5596):1248–51.
156) Luo L, O’Leary DDM [Axon retraction and degeneration in development and disease.] Annu. Rev. Neurosci. (2005)
28:127–56.
157) Lømo T [The discovery of long-term potentiation.] Philos. Trans. R. Soc. Lond. B. Biol. Sci. (2003) 358:617–20.
158) West AE, Chen WG, Dalva MB, Dolmetsch RE, Kornhauser JM, Shaywitz AJ, Takasu MA, Tao X, Greenberg ME
[Calcium regulation of neuronal gene expression.] Proc. Natl. Acad. Sci. U. S. A. (2001) 98:11024–31.
159) 奥野浩行 [シナプスから核へのシグナリングとシナプス活動依存的遺伝子発現:前初期遺伝子Arcの発現制御メカニズ
ムを中心に] 生化学 (2010) 82(9):841–6.
160) Hosokawa T, Rusakov DA, Bliss T V, Fine A [Repeated confocal imaging of individual dendritic spines in the living
hippocampal slice: evidence for changes in length and orientation associated with chemically induced LTP.] J.
Neurosci. (1995) 15:5560–73.
161) Toni N, Buchs PA, Nikonenko I, Bron CR, Muller D [LTP promotes formation of multiple spine synapses between a
single axon terminal and a dendrite.] Nature (1999) 402:421–5.
162) Huveneers S, Danen EHJ [Adhesion signaling - crosstalk between integrins, Src and Rho.] J. Cell Sci. (2009)
122:1059–69.
163) Tanaka H [Cadherin-induced remodeling of the synaptic junction] Folia Pharmacol. Jpn. 公益社団法人 日本薬理学会
(2012) 139(3):93–8.
164) O’Keefe J, Dostrovsky J [The hippocampus as a spatial map. Preliminary evidence from unit activity in the freelymoving rat.] Brain Res. (1971) 34:171–5.
165) Ekstrom AD, Kahana MJ, Caplan JB, Fields TA, Isham EA, Newman EL, Fried I [Cellular networks underlying
human spatial navigation.] Nature (2003) 425:184–8.
166) Rolls ET, O’Mara SM [View-responsive neurons in the primate hippocampal complex] Hippocampus (1995) 5:409–24.
167) Rolls ET, Robertson RG, Georges-François P [Spatial view cells in the primate hippocampus] Eur. J. Neurosci. (1997)
9:1789–94.
168) Solstad T, Boccara CN, Kropff E, Moser M-B, Moser EI [Representation of geometric borders in the entorhinal
cortex.] Science (2008) 322(5909):1865–8.
169) MacDonald CJ, Lepage KQ, Eden UT, Eichenbaum H [Hippocampal “time cells” bridge the gap in memory for
discontiguous events] Neuron (2011) 71:737–49.
170) 松本信圭, 坂口哲也, 池谷裕二 [海馬回路演算の機能と意義 : 基礎から最新の知見まで (特集 海馬の情報処理)] 心理学評
論. 心理学評論刊行会 (2013) 56(2):157–85.
171) Mizunuma M, Norimoto H, Tao K, Egawa T, Hanaoka K, Sakaguchi T, Hioki H, Kaneko T, Yamaguchi S, Nagano T,
Matsuki N, Ikegaya Y [Unbalanced excitability underlies offline reactivation of behaviorally activated neurons.] Nat.
Neurosci. (2014) 17:503–5.
172) Silva AJ, Paylor R, Wehner JM, Tonegawa S [Impaired spatial learning in alpha-calcium-calmodulin kinase II
mutant mice.] Science (1992) 257:206–11.
173) Kullmann DM, Lamsa KP [Long-term synaptic plasticity in hippocampal interneurons.] Nat. Rev. Neurosci. (2007)
8:687–99.
174) Gray EG [Axo-somatic and axo-dendritic synapses of the cerebral cortex] J. Anat. (1959) 93:420–33.
175) Kreienkamp H-J [Organisation of G-protein-coupled receptor signalling complexes by scaffolding proteins.] Curr.
Opin. Pharmacol. (2002) 2:581–6.
176) Vessey JP, Karra D [More than just synaptic building blocks: Scaffolding proteins of the post-synaptic density
regulate dendritic patterning] J. Neurochem. (2007), 324–32.
177) Richardson JS [The anatomy and taxonomy of protein structure.] Adv. Protein Chem. (1981) 34:167–339.
178) Burack WR, Shaw AS [Signal transduction: Hanging on a scaffold] Curr. Opin. Cell Biol. (2000), 211–6.
179) Snyder EM, Nong Y, Almeida CG, Paul S, Moran T, Choi EY, Nairn AC, Salter MW, Lombroso PJ, Gouras GK,
Greengard P [Regulation of NMDA receptor trafficking by amyloid-beta.] Nat. Neurosci. (2005) 8:1051–8.
180) Hsieh H, Boehm J, Sato C, Iwatsubo T, Tomita T, Sisodia S, Malinow R [AMPAR removal underlies Abeta-induced
synaptic depression and dendritic spine loss.] Neuron (2006) 52(5):831–43.
181) Nonaka M, Doi T, Fujiyoshi Y, Takemoto-Kimura S, Bito H [Essential contribution of the ligand-binding beta B/beta
C loop of PDZ1 and PDZ2 in the regulation of postsynaptic clustering, scaffolding, and localization of postsynaptic
density-95.] J. Neurosci. (2006) 26:763–74.
182) Spaller MR [Act globally, think locally: systems biology addresses the PDZ domain.] ACS Chem. Biol. (2006), 207–10.
183) Saras J, Heldin C-H [PDZ domains bind carboxy-terminal sequences of target proteins] Trends Biochem. Sci. (1996)
21(12):455–8.
184) Cowburn D [Peptide recognition by PTB and PDZ domains.] Curr. Opin. Struct. Biol. (1997) 7:835–8.
185) Hashimoto K, Fukaya M, Qiao X, Sakimura K, Watanabe M, Kano M [Impairment of AMPA receptor function in
cerebellar granule cells of ataxic mutant mouse stargazer.] J. Neurosci. (1999) 19:6027–36.
186) Lee HK, Barbarosie M, Kameyama K, Bear MF, Huganir RL [Regulation of distinct AMPA receptor phosphorylation
sites during bidirectional synaptic plasticity.] Nature (2000) 405:955–9.
187) 昆隆英, 栗栖源嗣 [補助サブユニットによるグルタミン酸受容体の機能制御] 生化学. 日本生化学会 (2013) 85(4):275–8.
188) Hayashi Y, Shi SH, Esteban JA, Piccini A, Poncer JC, Malinow R [Driving AMPA receptors into synapses by LTP and
CaMKII: requirement for GluR1 and PDZ domain interaction.] Science (2000) 287(5461):2262–7.
189) Nakagawa T, Futai K, Lashuel HA, Lo I, Okamoto K, Walz T, Hayashi Y, Sheng M [Quaternary structure, protein
dynamics, and synaptic function of SAP97 controlled by L27 domain interactions] Neuron (2004) 44:453–67.
V
190) Ziv NE, Smith SJ [Evidence for a role of dendritic filopodia in synaptogenesis and spine formation] Neuron (1996)
17(1):91–102.
191) 根東覚 [興奮性シナプスの形成とその分子構築] 福岡医学雑誌 = Fukuoka acta medica. 福岡医学会 (2013) 104(9):263–
74.
192) Chowdhury S, Shepherd JD, Okuno H, Lyford G, Petralia RS, Plath N, Kuhl D, Huganir RL, Worley PF [Arc/Arg3.1
Interacts with the Endocytic Machinery to Regulate AMPA Receptor Trafficking] Neuron (2006) 52:445–59.
193) Star EN, Kwiatkowski DJ, Murthy VN [Rapid turnover of actin in dendritic spines and its regulation by activity.]
Nat. Neurosci. (2002) 5:239–46.
194) Okamoto K-I, Nagai T, Miyawaki A, Hayashi Y [Rapid and persistent modulation of actin dynamics regulates
postsynaptic reorganization underlying bidirectional plasticity.] Nat. Neurosci. (2004) 7(10):1104–12.
195) Okamoto K, Hayashi Y [Visualization of F-actin and G-actin equilibrium using fluorescence resonance energy
transfer (FRET) in cultured cells and neurons in slices.] Nat. Protoc. (2006) 1(2):911–9.
196) Narumiya S, Ishizaki T, Watanabe N [Rho effectors and reorganization of actin cytoskeleton.] FEBS Lett. (1997)
410:68–72.
197) 尾藤晴彦 [神経形態とその可塑性を制御するシグナル伝達機構の探索] 生化学. 日本生化学会 (2007) 79(1):5–15.
198) Mori Y, Matsui T, Furutani Y, Yoshihara Y, Fukuda M [Small GTPase Rab17 regulates dendritic morphogenesis and
postsynaptic development of hippocampal neurons] J. Biol. Chem. (2012) 287:8963–73.
199) Bradke F, Dotti CG [Establishment of neuronal polarity: Lessons from cultured hippocampal neurons] Curr. Opin.
Neurobiol. (2000), 574–81.
200) Arimura N, Kaibuchi K [Neuronal polarity: from extracellular signals to intracellular mechanisms.] Nat. Rev.
Neurosci. (2007) 8:194–205.
201) Hensley K, Venkova K, Christov A, Gunning W, Park J [Collapsin response mediator protein-2: An emerging
pathologic feature and therapeutic target for neurodisease indications] Mol. Neurobiol. (2011) 43:180–91.
202) Gomi F, Uchida Y [MAP1B 1-126 interacts with tubulin isoforms and induces neurite outgrowth and neuronal death
of cultured cortical neurons] Brain Res. (2012) 1433:1–8.
203) Fukata Y, Itoh TJ, Kimura T, Ménager C, Nishimura T, Shiromizu T, Watanabe H, Inagaki N, Iwamatsu A, Hotani
H, Kaibuchi K [CRMP-2 binds to tubulin heterodimers to promote microtubule assembly.] Nat. Cell Biol. (2002)
4:583–91.
204) Inagaki N, Chihara K, Arimura N, Ménager C, Kawano Y, Matsuo N, Nishimura T, Amano M, Kaibuchi K [CRMP-2
induces axons in cultured hippocampal neurons.] Nat. Neurosci. (2001) 4:781–2.
205) Yoshimura T, Kawano Y, Arimura N, Kawabata S, Kikuchi A, Kaibuchi K [GSK-3β regulates phosphorylation of
CRMP-2 and neuronal polarity] Cell (2005) 120:137–49.
206) Shore P, Sharrocks AD [The MADS-box family of transcription factors.] Eur. J. Biochem. (1995) 229:1–13.
207) Ramanan N, Shen Y, Sarsfield S, Lemberger T, Schütz G, Linden DJ, Ginty DD [SRF mediates activity-induced gene
expression and synaptic plasticity but not neuronal viability.] Nat. Neurosci. (2005) 8:759–67.
208) Knöll B, Kretz O, Fiedler C, Alberti S, Schütz G, Frotscher M, Nordheim A [Serum response factor controls neuronal
circuit assembly in the hippocampus.] Nat. Neurosci. (2006) 9(2):195–204.
209) Lu PPY, Ramanan N [Serum Response Factor Is Required for Cortical Axon Growth But Is Dispensable for
Neurogenesis and Neocortical Lamination] J. Neurosci. (2011), 16651–64.
210) Posern G, Treisman R [Actin’ together: serum response factor, its cofactors and the link to signal transduction]
Trends Cell Biol. (2006), 588–96.
211) Glenner GG, Wong CW [Alzheimer’s disease: initial report of the purification and characterization of a novel
cerebrovascular amyloid protein.] Biochem. Biophys. Res. Commun. (1984) 120:885–90.
212) Glenner GG, Wong CW [Alzheimer’s disease and Down's syndrome: sharing of a unique cerebrovascular amyloid
fibril protein.] Biochem. Biophys. Res. Commun. (1984) 122:1131–5.
213) 秋山治彦 [アルツハイマー病(Alzheimer disease)] 日本神経病理学会 (2006).
214) Terry RD, Hansen LA, DeTeresa R, Davies P, Tobias H, Katzman R [Senile dementia of the Alzheimer type without
neocortical neurofibrillary tangles] J Neuropathol Exp Neurol (1987) 46:262–8.
215) Brion JP, Anderton BH, Authelet M, Dayanandan R, Leroy K, Lovestone S, Octave JN, Pradier L, Touchet N, Tremp
G [Neurofibrillary tangles and tau phosphorylation.] Biochem Soc Symp (2001) 81–8.
216) 箕浦克彦 [タウタンパク質に存在する微小管結合ドメインの溶液構造とフィラメント形成機構に関する研究] 大阪薬科大
学紀要. 大阪薬科大学 (2007) (1):159–70.
217) Combs B, Voss K, Gamblin TC [Pseudohyperphosphorylation has differential effects on polymerization and function
of tau isoforms] Biochemistry (2011) 50:9446–56.
218) Rossor MN [Molecular pathology of Alzheimer’s disease.] J. Neurol. Neurosurg. Psychiatry. BMJ Group (1993)
56(6):583–6.
219) 須貝佑一 [アルツハイマー型および脳血管性痴呆患者の生命予後と死因] 日本医事新報 (1991) 3513:13–6.
220) Näslund J, Schierhorn A, Hellman U, Lannfelt L, Roses AD, Tjernberg LO, Silberring J, Gandy SE, Winblad B,
Greengard P [Relative abundance of Alzheimer A beta amyloid peptide variants in Alzheimer disease and normal
aging.] Proc. Natl. Acad. Sci. U. S. A. (1994) 91:8378–82.
221) Näslund J, Haroutunian V, Mohs R, Davis KL, Davies P, Greengard P, Buxbaum JD [Correlation between elevated
levels of amyloid beta-peptide in the brain and cognitive decline.] JAMA (2000) 283:1571–7.
222) Lue LF, Kuo YM, Roher AE, Brachova L, Shen Y, Sue L, Beach T, Kurth JH, Rydel RE, Rogers J [Soluble amyloid
beta peptide concentration as a predictor of synaptic change in Alzheimer’s disease.] Am. J. Pathol. (1999) 155:853–
62.
223) Mayeux R, Tang MX, Jacobs DM, Manly J, Bell K, Merchant C, Small SA, Stern Y, Wisniewski HM, Mehta PD
[Plasma amyloid beta-peptide 1-42 and incipient Alzheimer’s disease] Ann. Neurol. (1999) 46:412–6.
224) Mehta PD, Pirttilä T, Mehta SP, Sersen EA, Aisen PS, Wisniewski HM [Plasma and cerebrospinal fluid levels of
amyloid beta proteins 1-40 and 1-42 in Alzheimer disease.] Arch. Neurol. (2000) 57:100–5.
225) Kawarabayashi T, Younkin LH, Saido TC, Shoji M, Ashe KH, Younkin SG [Age-dependent changes in brain, CSF,
and plasma amyloid (beta) protein in the Tg2576 transgenic mouse model of Alzheimer’s disease] J Neurosci (2001)
21:372–81.
226) Hardy J [Amyloid, the presenilins and Alzheimer’s disease] Trends Neurosci. (1997), 154–9.
VI
227) Hardy J, Selkoe DJ [The amyloid hypothesis of Alzheimer’s disease: progress and problems on the road to
therapeutics.] Science (2002) 297:353–6.
228) Schupf N, Patel B, Silverman W, Zigman WB, Zhong N, Tycko B, Mehta PD, Mayeux R [Elevated plasma amyloid βpeptide 1-42 and onset of dementia in adults with Down syndrome] Neurosci. Lett. (2001) 301:199–203.
229) Mann DM [Alzheimer’s disease and Down's syndrome.] Histopathology (1988) 13:125–37.
230) Gyure KA, Durham R, Stewart WF, Smialek JE, Troncoso JC [Intraneuronal Aβ-amyloid precedes development of
amyloid plaques in Down syndrome] Arch. Pathol. Lab. Med. (2001) 125:489–92.
231) Shamas-Ud-Din S [Genetics of Down’s syndrome and Alzheimer's disease: Comment.] Br. J. Psychiatry (2002)
181:167–8.
232) Li S, Jin M, Koeglsperger T, Shepardson NE, Shankar GM, Selkoe DJ [Soluble Aβ oligomers inhibit long-term
potentiation through a mechanism involving excessive activation of extrasynaptic NR2B-containing NMDA
receptors.] J. Neurosci. (2011) 31:6627–38.
233) Spires TL, Meyer-Luehmann M, Stern EA, McLean PJ, Skoch J, Nguyen PT, Bacskai BJ, Hyman BT [Dendritic spine
abnormalities in amyloid precursor protein transgenic mice demonstrated by gene transfer and intravital multiphoton
microscopy.] J. Neurosci. (2005) 25:7278–87.
234) Yan JJ, Cho JY, Kim HS, Kim KL, Jung JS, Huh SO, Suh HW, Kim YH, Song DK [Protection against beta-amyloid
peptide toxicity in vivo with long-term administration of ferulic acid.] Br. J. Pharmacol. (2001) 133:89–96.
235) Stern EA, Bacskai BJ, Hickey GA, Attenello FJ, Lombardo JA, Hyman BT [Cortical synaptic integration in vivo is
disrupted by amyloid-beta plaques.] J. Neurosci. (2004) 24:4535–40.
236) Kanai M, Matsubara E, Isoe K, Urakami K, Nakashima K, Arai H, Sasaki H, Abe K, Iwatsubo T, Kosaka T,
Watanabe M, Tomidokoro Y, Shizuka M, Mizushima K, Nakamura T, Igeta Y, Ikeda Y, Amari M, Kawarabayashi T,
Ishiguro K, Harigaya Y, Wakabayashi K, Okamoto K, Hirai S, Shoji M [Longitudinal study of cerebrospinal fluid
levels of tau, A beta1-40, and A beta1-42(43) in Alzheimer’s disease: a study in Japan.] Ann. Neurol. (1998) 44:17–26.
237) Arai H, Terajima M, Miura M, Higuchi S, Muramatsu T, Machida N, Seiki H, Takase S, Clark CM, Lee VM [Tau in
cerebrospinal fluid: a potential diagnostic marker in Alzheimer’s disease.] Ann. Neurol. (1995) 38:649–52.
238) Braak H, Braak E [Neuropathological stageing of Alzheimer-related changes.] Acta Neuropathol. (1991) 82:239–59.
239) Busciglio J, Lorenzo A, Yeh J, Yankner BA [beta-amyloid fibrils induce tau phosphorylation and loss of microtubule
binding.] Neuron (1995) 14:879–88.
240) Geula C, Wu CK, Saroff D, Lorenzo A, Yuan M, Yankner BA [Aging renders the brain vulnerable to amyloid betaprotein neurotoxicity.] Nat. Med. (1998) 4:827–31.
241) Zempel H, Thies E, Mandelkow E, Mandelkow E-M [Abeta oligomers cause localized Ca(2+) elevation, missorting of
endogenous Tau into dendrites, Tau phosphorylation, and destruction of microtubules and spines.] J. Neurosci. (2010)
30:11938–50.
242) Kang DE, Pietrzik CU, Baum L, Chevallier N, Merriam DE, Kounnas MZ, Wagner SL, Troncoso JC, Kawas CH,
Katzman R, Koo EH [Modulation of amyloid β-protein clearance and Alzheimer’s disease susceptibility by the LDL
receptor-related protein pathway] J. Clin. Invest. (2000) 106:1159–66.
243) Shibata M, Yamada S, Ram Kumar S, Calero M, Bading J, Frangione B, Holtzman DM, Miller CA, Strickland DK,
Ghiso J, Zlokovic B V. [Clearance of Alzheimer’s amyloid-β1-40 peptide from brain by LDL receptor-related protein-1
at the blood-brain barrier] J. Clin. Invest. (2000) 106:1489–99.
244) Zipser BD, Johanson CE, Gonzalez L, Berzin TM, Tavares R, Hulette CM, Vitek MP, Hovanesian V, Stopa EG
[Microvascular injury and blood-brain barrier leakage in Alzheimer’s disease] Neurobiol. Aging (2007) 28:977–86.
245) Farrall AJ, Wardlaw JM [Blood-brain barrier: ageing and microvascular disease--systematic review and metaanalysis.] Neurobiol. Aging (2009) 30:337–52.
246) Sjögren M, Vanderstichele H, Agren H, Zachrisson O, Edsbagge M, Wikkelsø C, Skoog I, Wallin A, Wahlund LO,
Marcusson J, Nägga K, Andreasen N, Davidsson P, Vanmechelen E, Blennow K [Tau and Abeta42 in cerebrospinal
fluid from healthy adults 21-93 years of age: establishment of reference values.] Clin. Chem. (2001) 47:1776–81.
247) Fukumoto H, Tennis M, Locascio JJ, Hyman BT, Growdon JH, Irizarry MC [Age but not diagnosis is the main
predictor of plasma amyloid beta-protein levels.] Arch. Neurol. (2003) 60:958–64.
248) Dickson DW [The pathogenesis of senile plaques.] J. Neuropathol. Exp. Neurol. (1997) 56:321–39.
249) Yamada M, Itoh Y, Otomo E, Suematsu N, Matsushita M [Dementia of the Alzheimer type and related dementias in
the aged: DAT subgroups and senile dementia of the neurofibrillary tangle type] Neuropathology (1996) 16(2):89–98.
250) Jellinger KA, Bancher C [Senile dementia with tangles (tangle predominant form of senile dementia).] Brain Pathol.
(1998) 8:367–76.
251) Nilsberth C, Westlind-Danielsson A, Eckman CB, Condron MM, Axelman K, Forsell C, Stenh C, Luthman J, Teplow
DB, Younkin SG, Näslund J, Lannfelt L [The “Arctic” APP mutation (E693G) causes Alzheimer’s disease by enhanced
Abeta protofibril formation.] Nat. Neurosci. (2001) 4:887–93.
252) Van Nostrand WE, Melchor JP, Cho HS, Greenberg SM, Rebeck GW [Pathogenic Effects of D23N Iowa Mutant
Amyloid β-Protein] J. Biol. Chem. (2001) 276:32860–6.
253) Kang J, Lemaire HG, Unterbeck A, Salbaum JM, Masters CL, Grzeschik KH, Multhaup G, Beyreuther K, MüllerHill B [The precursor of Alzheimer’s disease amyloid A4 protein resembles a cell-surface receptor.] Nature (1987)
325(6106):733–6.
254) Tanzi RE, Gusella JF, Watkins PC, Bruns GA, St George-Hyslop P, Van Keuren ML, Patterson D, Pagan S, Kurnit
DM, Neve RL [Amyloid beta protein gene: cDNA, mRNA distribution, and genetic linkage near the Alzheimer locus.]
Science (1987) 235:880–4.
255) Selkoe DJ [Clearing the brain’s amyloid cobwebs] Neuron (2001), 177–80.
256) O’Brien RJ, Wong PC [Amyloid precursor protein processing and Alzheimer’s disease.] Annu. Rev. Neurosci. (2011)
34:185–204.
257) Gralle M, Botelho MM, de Oliveira CLP, Torriani I, Ferreira ST [Solution studies and structural model of the
extracellular domain of the human amyloid precursor protein.] Biophys. J. (2002) 83(6):3513–24.
258) Goldgaber D, Lerman MI, McBride OW, Saffiotti U, Gajdusek DC [Characterization and chromosomal localization of
a cDNA encoding brain amyloid of Alzheimer’s disease.] Science (1987) 235:877–80.
259) Kang J, Müller-Hill B [Differential splicing of Alzheimer’s disease amyloid A4 precursor RNA in rat tissues:
PreA4(695) mRNA is predominantly produced in rat and human brain.] Biochem. Biophys. Res. Commun. (1990)
166:1192–200.
VII
260) De Strooper B, Annaert W [Proteolytic processing and cell biological functions of the amyloid precursor protein.] J.
Cell Sci. (2000) 113 ( Pt 1:1857–70.
261) Selkoe DJ, Schenk D [Alzheimer’s disease: molecular understanding predicts amyloid-based therapeutics.] Annu.
Rev. Pharmacol. Toxicol. (2003) 43:545–84.
262) Masters CL, Simms G, Weinman NA, Multhaup G, McDonald BL, Beyreuther K [Amyloid plaque core protein in
Alzheimer disease and Down syndrome.] Proc. Natl. Acad. Sci. U. S. A. (1985) 82:4245–9.
263) Lammich S, Kojro E, Postina R, Gilbert S, Pfeiffer R, Jasionowski M, Haass C, Fahrenholz F [Constitutive and
regulated α-secretase cleavage of Alzheimer’s amyloid precursor protein by a disintegrin metalloprotease] Proc. Natl.
Acad. Sci. U. S. A. (1999) 96:3922–7.
264) Allinson TMJ, Parkin ET, Turner AJ, Hooper NM [ADAMs family members as amyloid precursor protein alphasecretases.] J. Neurosci. Res. (2003) 74:342–52.
265) 榎本和生 [神経ネットワーク再編の鍵を握る細胞外マトリックス分解機構] 生化學. 日本生化学会 (2010) 82(10):972–8.
266) Zheng H, Koo EH [The amyloid precursor protein: beyond amyloid.] Mol. Neurodegener. (2006) 1(1):5.
267) Hung AY, Koo EH, Haass C, Selkoe DJ [Increased expression of beta-amyloid precursor protein during neuronal
differentiation is not accompanied by secretory cleavage.] Proc. Natl. Acad. Sci. U. S. A. (1992) 89:9439–43.
268) Bibel M, Richter J, Schrenk K, Tucker KL, Staiger V, Korte M, Goetz M, Barde Y-A [Differentiation of mouse
embryonic stem cells into a defined neuronal lineage.] Nat. Neurosci. (2004) 7:1003–9.
269) Van Den Heuvel C, Blumbergs P, Finnie J, Manavis J, Lewis S, Jones N, Reilly P, Pereira R [Upregulation of
amyloid precursor protein and its mRNA in an experimental model of paediatric head injury] J Clin Neurosci (2000)
7:140–5.
270) Van den Heuvel C, Blumbergs PC, Finnie JW, Manavis J, Jones NR, Reilly PL, Pereira RA [Upregulation of amyloid
precursor protein messenger RNA in response to traumatic brain injury: an ovine head impact model.] Exp. Neurol.
(1999) 159:441–50.
271) Leyssen M, Ayaz D, Hébert SS, Reeve S, De Strooper B, Hassan BA [Amyloid precursor protein promotes postdevelopmental neurite arborization in the Drosophila brain.] EMBO J. (2005) 24:2944–55.
272) Zheng H, Koo EH [Biology and pathophysiology of the amyloid precursor protein.] Mol. Neurodegener. (2011) 6:27.
273) Teller JK, Russo C, DeBusk LM, Angelini G, Zaccheo D, Dagna-Bricarelli F, Scartezzini P, Bertolini S, Mann DM,
Tabaton M, Gambetti P [Presence of soluble amyloid beta-peptide precedes amyloid plaque formation in Down’s
syndrome.] Nat. Med. (1996) 2:93–5.
274) Figueroa DJ, Morris JA, Ma L, Kandpal G, Chen E, Li Y-M, Austin CP [Presenilin-dependent gamma-secretase
activity modulates neurite outgrowth.] Neurobiol. Dis. (2002) 9:49–60.
275) Rushworth J V, Hooper NM [Lipid Rafts: Linking Alzheimer’s Amyloid-β Production, Aggregation, and Toxicity at
Neuronal Membranes.] Int. J. Alzheimers. Dis. (2010) 2011:603052.
276) Von Arnim CAF, Spoelgen R, Peltan ID, Deng M, Courchesne S, Koker M, Matsui T, Kowa H, Lichtenthaler SF,
Irizarry MC, Hyman BT [GGA1 acts as a spatial switch altering amyloid precursor protein trafficking and
processing.] J. Neurosci. (2006) 26:9913–22.
277) Plant LD, Boyle JP, Smith IF, Peers C, Pearson HA [The production of amyloid beta peptide is a critical requirement
for the viability of central neurons.] J. Neurosci. (2003) 23:5531–5.
278) Mucke L, Selkoe DJ [Neurotoxicity of amyloid β-protein: Synaptic and network dysfunction] Cold Spring Harb.
Perspect. Med. (2012) 2.
279) Revett TJ, Baker GB, Jhamandas J, Kar S [Glutamate system, amyloid beta peptides and tau protein: Functional
interrelationships and relevance to Alzheimer disease pathology] J. Psychiatry Neurosci. (2013), 6–23.
280) Puzzo D, Privitera L, Fa’ M, Staniszewski A, Hashimoto G, Aziz F, Sakurai M, Ribe EM, Troy CM, Mercken M, Jung
SS, Palmeri A, Arancio O [Endogenous amyloid-β is necessary for hippocampal synaptic plasticity and memory] Ann.
Neurol. (2011) 69:819–30.
281) Cirrito JR, Yamada KA, Finn MB, Sloviter RS, Bales KR, May PC, Schoepp DD, Paul SM, Mennerick S, Holtzman
DM [Synaptic activity regulates interstitial fluid amyloid-beta levels in vivo.] Neuron (2005) 48:913–22.
282) Selkoe DJ [Soluble oligomers of the amyloid beta-protein impair synaptic plasticity and behavior] Behav. Brain Res.
(2008), 106–13.
283) Tang BL [Neuronal protein trafficking associated with Alzheimer disease: From APP and BACE1 to glutamate
receptors] Cell Adhes. Migr. (2009), 118–28.
284) Abramov E, Dolev I, Fogel H, Ciccotosto GD, Ruff E, Slutsky I [Amyloid-beta as a positive endogenous regulator of
release probability at hippocampal synapses.] Nat. Neurosci. (2009) 12(12):1567–76.
285) Shankar GM, Li S, Mehta TH, Garcia-Munoz A, Shepardson NE, Smith I, Brett FM, Farrell MA, Rowan MJ, Lemere
CA, Regan CM, Walsh DM, Sabatini BL, Selkoe DJ [Amyloid-beta protein dimers isolated directly from Alzheimer’s
brains impair synaptic plasticity and memory.] Nat. Med. (2008) 14:837–42.
286) Murakami K, Masuda Y, Shirasawa T, Shimizu T, Irie K [The turn formation at positions 22 and 23 in the 42-mer
amyloid β peptide: The emerging role in the pathogenesis of Alzheimer’s disease] Geriatr. Gerontol. Int. (2010), S169–
79.
287) Saito T, Suemoto T, Brouwers N, Sleegers K, Funamoto S, Mihira N, Matsuba Y, Yamada K, Nilsson P, Takano J,
Nishimura M, Iwata N, Van Broeckhoven C, Ihara Y, Saido TC [Potent amyloidogenicity and pathogenicity of Aβ43.]
Nat. Neurosci. (2011) 14:1023–32.
288) Balducci C, Beeg M, Stravalaci M, Bastone A, Sclip A, Biasini E, Tapella L, Colombo L, Manzoni C, Borsello T,
Chiesa R, Gobbi M, Salmona M, Forloni G [Synthetic amyloid-beta oligomers impair long-term memory independently
of cellular prion protein.] Proc. Natl. Acad. Sci. U. S. A. (2010) 107:2295–300.
289) Zhang Y, McLaughlin R, Goodyer C, LeBlanc A [Selective cytotoxicity of intracellular amyloid β peptide1-42 through
p53 and Bax in cultured primary human neurons] J. Cell Biol. (2002) 156:519–29.
290) Walsh DM, Klyubin I, Fadeeva J V, Cullen WK, Anwyl R, Wolfe MS, Rowan MJ, Selkoe DJ [Naturally secreted
oligomers of amyloid beta protein potently inhibit hippocampal long-term potentiation in vivo.] Nature (2002)
416:535–9.
291) Ellgaard L, Helenius A [Quality control in the endoplasmic reticulum.] Nat Rev Mol Cell Biol (2003) 4:181–91.
292) 藤本充章, 中井彰 [熱ストレス応答による恒常性維持の機構] 生化學. 日本生化学会 (2009) 81(6):465–73.
293) Benndorf R, Welsh MJ [Shocking degeneration.] Nat. Genet. (2004) 36(6):547–8.
VIII
294) Brenner M, Johnson AB, Boespflug-Tanguy O, Rodriguez D, Goldman JE, Messing A [Mutations in GFAP, encoding
glial fibrillary acidic protein, are associated with Alexander disease.] Nat. Genet. (2001) 27:117–20.
295) Cohen E, Bieschke J, Perciavalle RM, Kelly JW, Dillin A [Opposing activities protect against age-onset
proteotoxicity.] Science (2006) 313:1604–10.
296) Fonte V, Kipp DR, Yerg J, Merin D, Forrestal M, Wagner E, Roberts CM, Link CD [Suppression of in vivo betaamyloid peptide toxicity by overexpression of the HSP-16.2 small chaperone protein.] J. Biol. Chem. (2008)
283(2):784–91.
297) Okochi M, Nomura T, Zako T, Arakawa T, Iizuka R, Ueda H, Funatsu T, Leroux M, Yohda M [Kinetics and binding
sites for interaction of the prefoldin with a group II chaperonin: contiguous non-native substrate and chaperonin
binding sites in the archaeal prefoldin.] J. Biol. Chem. (2004) 279:31788–95.
298) Zako T, Iizuka R, Okochi M, Nomura T, Ueno T, Tadakuma H, Yohda M, Funatsu T [Facilitated release of substrate
protein from prefoldin by chaperonin] FEBS Lett. (2005) 579:3718–24.
299) Zako T, Murase Y, Iizuka R, Yoshida T, Kanzaki T, Ide N, Maeda M, Funatsu T, Yohda M [Localization of Prefoldin
Interaction Sites in the Hyperthermophilic Group II Chaperonin and Correlations between Binding Rate and Protein
Transfer Rate] J. Mol. Biol. (2006) 364:110–20.
300) Zhang J, Liu L, Zhang X, Jin F, Chen J, Ji C, Gu S, Xie Y, Mao Y [Cloning and characterization of a novel human
prefoldin and SPEC domain protein gene (PFD6L) from the fetal brain.] Biochem. Genet. (2006) 44(1-2):69–74.
301) Sakono M, Zako T, Ueda H, Yohda M, Maeda M [Formation of highly toxic soluble amyloid beta oligomers by the
molecular chaperone prefoldin] FEBS J. (2008) 275:5982–93.
302) Faccio L, Fusco C, Chen A, Martinotti S, Bonventre J V., Zervos AS [Characterization of a novel human serine
protease that has extensive homology to bacterial heat shock endoprotease HtrA and is regulated by kidney ischemia]
J. Biol. Chem. (2000) 275:2581–8.
303) Park HJ, Seong YM, Choi JY, Kang S, Rhim H [Alzheimer’s disease-associated amyloid beta interacts with the
human serine protease HtrA2/Omi] Neurosci. Lett. (2004) 357:63–7.
304) Liu LZ, Huq S, Xu WM [Targeting Cyclooxygenase and Nitric Oxide Pathway Cross-Talk: A New Signal
Transduction Pathway for Developing More Effective Anti-Inflammatory Drugs] Curr. Signal Transduct. Ther. (2009)
4:66–75.
305) Oda T, Wals P, Osterburg HH, Johnson SA, Pasinetti GM, Morgan TE, Rozovsky I, Stine WB, Snyder SW, Holzman
TF [Clusterin (apoJ) alters the aggregation of amyloid beta-peptide (A beta 1-42) and forms slowly sedimenting A beta
complexes that cause oxidative stress.] Exp. Neurol. (1995) 136:22–31.
306) Lambert MP, Barlow AK, Chromy BA, Edwards C, Freed R, Liosatos M, Morgan TE, Rozovsky I, Trommer B, Viola
KL, Wals P, Zhang C, Finch CE, Krafft GA, Klein WL [Diffusible, nonfibrillar ligands derived from Abeta1-42 are
potent central nervous system neurotoxins.] Proc. Natl. Acad. Sci. U. S. A. (1998) 95:6448–53.
307) Wilson MR, Easterbrook-Smith SB [Clusterin is a secreted mammalian chaperone.] Trends Biochem. Sci. (2000)
25(3):95–8.
308) Dahlgren KN, Manelli AM, Blaine Stine W, Baker LK, Krafft GA, Ladu MJ [Oligomeric and fibrillar species of
amyloid-beta peptides differentially affect neuronal viability] J. Biol. Chem. (2002) 277(35):32046–53.
309) Murakami K, Irie K, Morimoto A, Ohigashi H, Shindo M, Nagao M, Shimizu T, Shirasawa T [Synthesis, aggregation,
neurotoxicity, and secondary structure of various Aβ1-42 mutants of familial Alzheimer’s disease at positions 21-23]
Biochem. Biophys. Res. Commun. (2002) 294:5–10.
310) Murakami K, Irie K, Morimoto A, Ohigashi H, Shindo M, Nagao M, Shimizu T, Shirasawa T [Neurotoxicity and
physicochemical properties of Aβ mutant peptides from cerebral amyloid angiopathy: Implication for the pathogenesis
of cerebral amyloid angiopathy and Alzheimer’s disease] J. Biol. Chem. (2003) 278:46179–87.
311) Kodali R, Wetzel R [Polymorphism in the intermediates and products of amyloid assembly] Curr. Opin. Struct. Biol.
(2007), 48–57.
312) Jarrett JT, Berger EP, Lansbury PT [The carboxy terminus of the beta amyloid protein is critical for the seeding of
amyloid formation: implications for the pathogenesis of Alzheimer’s disease.] Biochemistry (1993) 32(18):4693–7.
313) Muramatsu H, Yokoi K, Chen L, Ichihara-Tanaka K, Kimura T, Muramatsu T [Midkine as a factor to counteract the
deposition of amyloid β-peptide plaques: in vitro analysis and examination in knockout mice.] Int. Arch. Med. (2011)
4(1):1.
314) Cantara S, Thorpe PE, Ziche M, Donnini S [TAT-BH4 counteracts Aβ toxicity on capillary endothelium] FEBS Lett.
Elsevier (2007) 581(4):702–6.
315) Lorenzo A, Yankner BA [Beta-amyloid neurotoxicity requires fibril formation and is inhibited by congo red.] Proc.
Natl. Acad. Sci. U. S. A. (1994) 91:12243–7.
316) Takeuchi O, Akira S [Recognition of viruses by innate immunity] Immunol. Rev. (2007), 214–24.
317) Ehlenberger AG, Nussenzweig V [The role of membrane receptors for C3b and C3d in phagocytosis.] J. Exp. Med.
(1977) 145:357–71.
318) Le Cabec V, Carréno S, Moisand A, Bordier C, Maridonneau-Parini I [Complement receptor 3 (CD11b/CD18)
mediates type I and type II phagocytosis during nonopsonic and opsonic phagocytosis, respectively.] J. Immunol.
(2002) 169:2003–9.
319) Luo B-H, Carman C V, Springer TA [Structural basis of integrin regulation and signaling.] Annu. Rev. Immunol.
(2007) 25:619–47.
320) Ross GD, Vĕtvicka V [CR3 (CD11b, CD18): a phagocyte and NK cell membrane receptor with multiple ligand
specificities and functions.] Clin. Exp. Immunol. (1993) 92:181–4.
321) Akiyama H, McGeer PL [Brain microglia constitutively express beta-2 integrins.] J. Neuroimmunol. (1990) 30:81–93.
322) Flannagan RS, Cosío G, Grinstein S [Antimicrobial mechanisms of phagocytes and bacterial evasion strategies.] Nat.
Rev. Microbiol. (2009) 7:355–66.
323) Vieira O V, Botelho RJ, Rameh L, Brachmann SM, Matsuo T, Davidson HW, Schreiber A, Backer JM, Cantley LC,
Grinstein S [Distinct roles of class I and class III phosphatidylinositol 3-kinases in phagosome formation and
maturation.] J. Cell Biol. (2001) 155:19–25.
324) Minakami R, Maehara Y, Kamakura S, Kumano O, Miyano K, Sumimoto H [Membrane phospholipid metabolism
during phagocytosis in human neutrophils.] Genes Cells (2010) 15:409–24.
IX
325) Wilkinson B, Koenigsknecht-Talboo J, Grommes C, Lee CYD, Landreth G [Fibrillar β-amyloid-stimulated
intracellular signaling cascades require Vav for induction of respiratory burst and phagocytosis in monocytes and
microglia] J. Biol. Chem. (2006) 281:20842–50.
326) Kaneko I, Tutumi S [Matters arising: conformations of β-amyloid in solution] J. Neurochem. (1997) 68(1):437–9.
327) Brown EJ, Frazier WA [Integrin-associated protein (CD47) and its ligands.] Trends Cell Biol. (2001) 11:130–5.
328) Koenigsknecht J, Landreth G [Microglial phagocytosis of fibrillar beta-amyloid through a beta1 integrin-dependent
mechanism.] J. Neurosci. (2004) 24:9838–46.
329) Pike CJ, Cummings BJ, Monzavi R, Cotman CW [Beta-amyloid-induced changes in cultured astrocytes parallel
reactive astrocytosis associated with senile plaques in Alzheimer’s disease.] Neuroscience (1994) 63:517–31.
330) Gitter BD, Cox LM, Rydel RE, May PC [Amyloid beta peptide potentiates cytokine secretion by interleukin-1 betaactivated human astrocytoma cells.] Proc. Natl. Acad. Sci. U. S. A. (1995) 92:10738–41.
331) Beaty CD, Franklin TL, Uehara Y, Wilson CB [Lipopolysaccharide-induced cytokine production in human monocytes:
role of tyrosine phosphorylation in transmembrane signal transduction.] Eur. J. Immunol. (1994) 24:1278–84.
332) Ilyin SE, Gayle D, Flynn MC, Plata-Salamán CR [Interleukin-1beta system (ligand, receptor type I, receptor
accessory protein and receptor antagonist), TNF-alpha, TGF-beta1 and neuropeptide Y mRNAs in specific brain
regions during bacterial LPS-induced anorexia.] Brain Res. Bull. (1998) 45:507–15.
333) Nolan Y, Martin D, Campbell VA, Lynch MA [Evidence of a protective effect of phosphatidylserine-containing
liposomes on lipopolysaccharide-induced impairment of long-term potentiation in the rat hippocampus] J.
Neuroimmunol. (2004) 151:12–23.
334) Palop JJ, Mucke L [Amyloid-beta-induced neuronal dysfunction in Alzheimer’s disease: from synapses toward neural
networks.] Nat. Neurosci. (2010) 13:812–8.
335) Vereker E, Campbell V, Roche E, McEntee E, Lynch MA [Lipopolysaccharide inhibits long term potentiation in the
rat dentate gyrus by activating caspase-1.] J. Biol. Chem. (2000) 275:26252–8.
336) Shaw KN, Commins S, O’Mara SM [Lipopolysaccharide causes deficits in spatial learning in the watermaze but not
in BDNF expression in the rat dentate gyrus] Behav. Brain Res. (2001) 124:47–54.
337) Lynch AM, Walsh C, Delaney A, Nolan Y, Campbell VA, Lynch MA [Lipopolysaccharide-induced increase in
signalling in hippocampus is abrogated by IL-10--a role for IL-1 beta?] J. Neurochem. (2004) 88:635–46.
338) Nick JA, Avdi NJ, Young SK, McDonald PP, Billstrom MA, Henson PM, Johnson GL, Worthen GS [An intracellular
signaling pathway linking lipopolysaccharide stimulation to cellular responses of the human neutrophil: the p38 MAP
kinase cascade and its functional significance.] Chest (1999) 116(1 Suppl):54S – 55S.
339) Inoue T, Boyle DL, Corr M, Hammaker D, Davis RJ, Flavell RA, Firestein GS [Mitogen-activated protein kinase
kinase 3 is a pivotal pathway regulating p38 activation in inflammatory arthritis.] Proc. Natl. Acad. Sci. U. S. A.
(2006) 103:5484–9.
340) Olson CM, Hedrick MN, Izadi H, Bates TC, Olivera ER, Anguita J [p38 mitogen-activated protein kinase controls
NF-κB transcriptional activation and tumor necrosis factor alpha production through RelA phosphorylation mediated
by mitogen- and stress-activated protein kinase 1 in response to Borrelia burgdorferi antigens] Infect. Immun. (2007)
75(1):270–7.
341) Oltmanns U, Issa R, Sukkar MB, John M, Chung KF [Role of c-jun N-terminal kinase in the induced release of GMCSF, RANTES and IL-8 from human airway smooth muscle cells.] Br. J. Pharmacol. (2003) 139:1228–34.
342) McDonald DR, Brunden KR, Landreth GE [Amyloid fibrils activate tyrosine kinase-dependent signaling and
superoxide production in microglia.] J. Neurosci. (1997) 17:2284–94.
343) Pyo H, Jou I, Jung S, Hong S, Joe EH [Mitogen-activated protein kinases activated by lipopolysaccharide and betaamyloid in cultured rat microglia.] Neuroreport (1998) 9:871–4.
344) Iwasaki K, Sunderland T, Kusiak JW, Wolozin B [Changes in gene transcription during a beta-mediated cell death]
Mol Psychiatry (1996) 1:65–71.
345) Estus S, Tucker HM, van Rooyen C, Wright S, Brigham EF, Wogulis M, Rydel RE [Aggregated amyloid-beta protein
induces cortical neuronal apoptosis and concomitant “apoptotic” pattern of gene induction.] J. Neurosci. (1997)
17:7736–45.
346) Combs CK, Johnson DE, Karlo JC, Cannady SB, Landreth GE [Inflammatory mechanisms in Alzheimer’s disease:
inhibition of beta-amyloid-stimulated proinflammatory responses and neurotoxicity by PPARgamma agonists.] J.
Neurosci. (2000) 20(2):558–67.
347) Ma TC, Zhu XZ [Suppression of lipopolysaccharide-induced impairment of active avoidance and interleukin-6induced increase of prostaglandin E2 release in rats by indometacin.] Arzneimittelforschung. (1997) 47:595–7.
348) Iwata N, Mizukami H, Shirotani K, Takaki Y, Muramatsu S, Lu B, Gerard NP, Gerard C, Ozawa K, Saido TC
[Presynaptic localization of neprilysin contributes to efficient clearance of amyloid-beta peptide in mouse brain.] J.
Neurosci. (2004) 24:991–8.
349) Leissring MA, Farris W, Chang AY, Walsh DM, Wu X, Sun X, Frosch MP, Selkoe DJ [Enhanced proteolysis of betaamyloid in APP transgenic mice prevents plaque formation, secondary pathology, and premature death] Neuron
(2003) 40:1087–93.
350) Van Nostrand WE, Porter M [Plasmin cleavage of the amyloid beta-protein: alteration of secondary structure and
stimulation of tissue plasminogen activator activity.] Biochemistry (1999) 38:11570–6.
351) Qiu WQ, Walsh DM, Ye Z, Vekrellis K, Zhang J, Podlisny MB, Rosner MR, Safavi A, Hersh LB, Selkoe DJ [Insulindegrading enzyme regulates extracellular levels of amyloid beta- protein by degradation] J. Biol. Chem. (1998)
273:32730–8.
352) Miller BC, Eckman EA, Sambamurti K, Dobbs N, Chow KM, Eckman CB, Hersh LB, Thiele DL [Amyloid-beta
peptide levels in brain are inversely correlated with insulysin activity levels in vivo.] Proc. Natl. Acad. Sci. U. S. A.
(2003) 100:6221–6.
353) Hamazaki H [Cathepsin D is involved in the clearance of Alzheimer’s beta-amyloid protein.] FEBS Lett. (1996) 396(23):139–42.
354) Glabe C [Intracellular mechanisms of amyloid accumulation and pathogenesis in Alzheimer’s disease.] J. Mol.
Neurosci. (2001) 17:137–45.
355) Vartak DG, Gemeinhart RA [Matrix metalloproteases: underutilized targets for drug delivery.] J. Drug Target.
(2007) 15:1–20.
X
356) Kirschner RJ, Goldberg AL [A high molecular weight metalloendoprotease from the cytosol of mammalian cells.] J.
Biol. Chem. (1983) 258:967–76.
357) Duckworth WC, Bennett RG, Hamel FG [Insulin degradation: Progress and potential] Endocr. Rev. (1998), 608–24.
358) Hersh LB [The insulysin (insulin degrading enzyme) enigma] Cell. Mol. Life Sci. (2006), 2432–4.
359) Kurochkin I V. [Insulin-degrading enzyme: Embarking on amyloid destruction] Trends Biochem. Sci. (2001), 421–5.
360) Ciaccio C, Tundo GR, Grasso G, Spoto G, Marasco D, Ruvo M, Gioia M, Rizzarelli E, Coletta M [Somatostatin: A
Novel Substrate and a Modulator of Insulin-Degrading Enzyme Activity] J. Mol. Biol. (2009) 385:1556–67.
361) Ralat LA, Kalas V, Zheng Z, Goldman RD, Sosnick TR, Tang WJ [Ubiquitin is a novel substrate for human insulindegrading enzyme] J. Mol. Biol. (2011) 406:454–66.
362) Kakimura J-I, Kitamura Y, Takata K, Umeki M, Suzuki S, Shibagaki K, Taniguchi T, Nomura Y, Gebicke-Haerter
PJ, Smith MA, Perry G, Shimohama S [Microglial activation and amyloid-beta clearance induced by exogenous heatshock proteins.] FASEB J. (2002) 16:601–3.
363) Iwata N, Higuchi M, Saido TC [Metabolism of amyloid-beta peptide and Alzheimer’s disease.] Pharmacol. Ther.
(2005) 108:129–48.
364) Matsas R, Kenny AJ, Turner AJ [The metabolism of neuropeptides. The hydrolysis of peptides, including
enkephalins, tachykinins and their analogues, by endopeptidase-24.11.] Biochem. J. (1984) 223:433–40.
365) Fukami S, Watanabe K, Iwata N, Haraoka J, Lu B, Gerard NP, Gerard C, Fraser P, Westaway D, St. George-Hyslop
P, Saido TC [Aβ-degrading endopeptidase, neprilysin, in mouse brain: Synaptic and axonal localization inversely
correlating with Aβ pathology] Neurosci. Res. (2002) 43:39–56.
366) Hama E, Shirotani K, Masumoto H, Sekine-Aizawa Y, Aizawa H, Saido TC [Clearance of extracellular and cellassociated amyloid beta peptide through viral expression of neprilysin in primary neurons.] J. Biochem. (2001)
130:721–6.
367) Guan H, Chow KM, Shah R, Rhodes CJ, Hersh LB [Degradation of islet amyloid polypeptide by neprilysin]
Diabetologia (2012) 55:2989–98.
368) Hersh LB, Rodgers DW [Neprilysin and amyloid beta peptide degradation.] Curr. Alzheimer Res. (2008) 5:225–31.
369) Saito T, Iwata N, Tsubuki S, Takaki Y, Takano J, Huang S-M, Suemoto T, Higuchi M, Saido TC [Somatostatin
regulates brain amyloid beta peptide Abeta42 through modulation of proteolytic degradation.] Nat. Med. (2005)
11:434–9.
370) Mizushima N [Autophagy: process and function.] Genes Dev. (2007) 21:2861–73.
371) 江崎淳二, 上野隆 [老年医学の展望 オートファジー減弱と老化の関わり] 日本老年医学会雑誌. 日本老年医学会 (2011)
48(6):606–12.
372) He C, Klionsky DJ [Regulation mechanisms and signaling pathways of autophagy.] Annu. Rev. Genet. (2009) 43:67–
93.
373) Bjørkøy G, Lamark T, Brech A, Outzen H, Perander M, Øvervatn A, Stenmark H, Johansen T, Overvatn A
[p62/SQSTM1 forms protein aggregates degraded by autophagy and has a protective effect on huntingtin-induced cell
death.] J. Cell Biol. (2005) 171:603–14.
374) Komatsu M, Waguri S, Koike M, Sou Y-S, Ueno T, Hara T, Mizushima N, Iwata J-I, Ezaki J, Murata S, Hamazaki J,
Nishito Y, Iemura S-I, Natsume T, Yanagawa T, Uwayama J, Warabi E, Yoshida H, Ishii T, Kobayashi A, Yamamoto
M, Yue Z, Uchiyama Y, Kominami E, Tanaka K [Homeostatic levels of p62 control cytoplasmic inclusion body
formation in autophagy-deficient mice.] Cell (2007) 131:1149–63.
375) Johansen T, Lamark T [Selective autophagy mediated by autophagic adapter proteins.] Autophagy (2011) 7:279–96.
376) Weidberg H, Shvets E, Elazar Z [Biogenesis and cargo selectivity of autophagosomes.] Annu. Rev. Biochem. (2011)
80:125–56.
377) Jung CH, Ro S-HH, Cao J, Otto NM, Kim D-H, Kim Do-Hyung DH [MTOR regulation of autophagy] FEBS Lett.
(2010) 584:1287–95.
378) Wang Z, Zhong J, Inuzuka H, Gao D, Shaik S, Sarkar FH, Wei W [An evolving role for DEPTOR in tumor
development and progression.] Neoplasia (2012) 14:368–75.
379) Hara T, Takamura A, Kishi C, Iemura S-I, Natsume T, Guan J-L, Mizushima N [FIP200, a ULK-interacting protein,
is required for autophagosome formation in mammalian cells.] J. Cell Biol. (2008) 181:497–510.
380) Hosokawa N, Hara T, Kaizuka T, Kishi C, Takamura A, Miura Y, Iemura S, Natsume T, Takehana K, Yamada N,
Guan J-L, Oshiro N, Mizushima N [Nutrient-dependent mTORC1 association with the ULK1-Atg13-FIP200 complex
required for autophagy.] Mol. Biol. Cell (2009) 20:1981–91.
381) Yu L, McPhee CK, Zheng L, Mardones GA, Rong Y, Peng J, Mi N, Zhao Y, Liu Z, Wan F, Hailey DW, Oorschot V,
Klumperman J, Baehrecke EH, Lenardo MJ [Termination of autophagy and reformation of lysosomes regulated by
mTOR.] Nature (2010) 465:942–6.
382) Heitman J, Movva NR, Hall MN [Targets for cell cycle arrest by the immunosuppressant rapamycin in yeast.]
Science (1991) 253:905–9.
383) Benton BM [A novel FK506- and rapamycin-binding protein (FPR3 gene product) in the yeast Saccharomyces
cerevisiae is a proline rotamase localized to the nucleolus] J. Cell Biol. (1994) 127(3):623–39.
384) Bové J, Martínez-Vicente M, Vila M [Fighting neurodegeneration with rapamycin: mechanistic insights.] Nat. Rev.
Neurosci. (2011) 12:437–52.
385) Tian Y, Bustos V, Flajolet M, Greengard P [A small-molecule enhancer of autophagy decreases levels of Abeta and
APP-CTF via Atg5-dependent autophagy pathway.] FASEB J. (2011) 25:1934–42.
386) Kovacina KS, Park GY, Bae SS, Guzzetta AW, Schaefer E, Birnbaum MJ, Roth RA [Identification of a proline-rich
Akt substrate as a 14-3-3 binding partner.] J. Biol. Chem. (2003) 278:10189–94.
387) Vander Haar E, Lee S-I, Bandhakavi S, Griffin TJ, Kim D-H [Insulin signalling to mTOR mediated by the Akt/PKB
substrate PRAS40.] Nat. Cell Biol. (2007) 9:316–23.
388) Sarbassov DD, Guertin DA, Ali SM, Sabatini DM [Phosphorylation and regulation of Akt/PKB by the rictor-mTOR
complex.] Science (2005) 307:1098–101.
389) Cafferkey R, Young PR, McLaughlin MM, Bergsma DJ, Koltin Y, Sathe GM, Faucette L, Eng WK, Johnson RK, Livi
GP [Dominant missense mutations in a novel yeast protein related to mammalian phosphatidylinositol 3-kinase and
VPS34 abrogate rapamycin cytotoxicity.] Mol. Cell. Biol. (1993) 13:6012–23.
390) Pause A, Belsham GJ, Gingras AC, Donzé O, Lin TA, Lawrence JC, Sonenberg N [Insulin-dependent stimulation of
protein synthesis by phosphorylation of a regulator of 5’-cap function.] Nature (1994) 371:762–7.
XI
391) Zoncu R, Efeyan A, Sabatini DM [mTOR: from growth signal integration to cancer, diabetes and ageing.] Nat. Rev.
Mol. Cell Biol. (2011) 12:21–35.
392) Nakatogawa H, Suzuki K, Kamada Y, Ohsumi Y [Dynamics and diversity in autophagy mechanisms: lessons from
yeast.] Nat. Rev. Mol. Cell Biol. (2009) 10:458–67.
393) Vabulas RM, Hartl FU [Protein synthesis upon acute nutrient restriction relies on proteasome function.] Science
(2005) 310:1960–3.
394) Onodera J, Ohsumi Y [Autophagy is required for maintenance of amino acid levels and protein synthesis under
nitrogen starvation.] J. Biol. Chem. (2005) 280:31582–6.
395) Suzuki SW, Onodera J, Ohsumi Y [Starvation induced cell death in autophagy-defective yeast mutants is caused by
mitochondria dysfunction] PLoS One (2011) 6.
396) Rujano MA, Bosveld F, Salomons FA, Dijk F, Van Waarde MAWH, Van Der Want JJL, De Vos RAI, Brunt ER, Sibon
OCM, Kampinga HH [Polarised asymmetric inheritance of accumulated protein damage in higher eukaryotes] PLoS
Biol. (2006) 4:2325–35.
397) 水島 昇 [細胞が自分を食べる オートファジーの謎] PHP Science World OS1 (2011).
398) Lau A, Wang X-J, Zhao F, Villeneuve NF, Wu T, Jiang T, Sun Z, White E, Zhang DD [A noncanonical mechanism of
Nrf2 activation by autophagy deficiency: direct interaction between Keap1 and p62.] Mol. Cell. Biol. (2010) 30:3275–
85.
399) Hampe J, Franke A, Rosenstiel P, Till A, Teuber M, Huse K, Albrecht M, Mayr G, De La Vega FM, Briggs J, Günther
S, Prescott NJ, Onnie CM, Häsler R, Sipos B, Fölsch UR, Lengauer T, Platzer M, Mathew CG, Krawczak M, Schreiber
S [A genome-wide association scan of nonsynonymous SNPs identifies a susceptibility variant for Crohn disease in
ATG16L1.] Nat. Genet. (2007) 39:207–11.
400) Zatloukal K, Stumptner C, Fuchsbichler A, Heid H, Schnoelzer M, Kenner L, Kleinert R, Prinz M, Aguzzi A, Denk H
[p62 Is a common component of cytoplasmic inclusions in protein aggregation diseases.] Am. J. Pathol. (2002)
160:255–63.
401) Jaeger PA, Pickford F, Sun C-H, Lucin KM, Masliah E, Wyss-Coray T [Regulation of amyloid precursor protein
processing by the Beclin 1 complex.] PLoS One (2010) 5:e11102.
402) Lee J-H, Yu WH, Kumar A, Lee S, Mohan PS, Peterhoff CM, Wolfe DM, Martinez-Vicente M, Massey AC, Sovak G,
Uchiyama Y, Westaway D, Cuervo AM, Nixon RA [Lysosomal proteolysis and autophagy require presenilin 1 and are
disrupted by Alzheimer-related PS1 mutations.] Cell (2010) 141:1146–58.
403) Margolis RL, Ross CA [Diagnosis of Huntington disease] Clin. Chem. (2003), 1726–32.
404) Lue LF, Brachova L, Civin WH, Rogers J [Inflammation, A beta deposition, and neurofibrillary tangle formation as
correlates of Alzheimer’s disease neurodegeneration.] J. Neuropathol. Exp. Neurol. (1996) 55:1083–8.
405) Akiyama H, Barger S, Barnum S, Bradt B, Bauer J, Cole GM, Cooper NR, Eikelenboom P, Emmerling M, Fiebich BL,
Finch CE, Frautschy S, Griffin WS, Hampel H, Hull M, Landreth G, Lue L, Mrak R, Mackenzie IR, McGeer PL,
O’Banion MK, Pachter J, Pasinetti G, Plata-Salaman C, Rogers J, Rydel R, Shen Y, Streit W, Strohmeyer R, et al.
[Inflammation and Alzheimer’s disease.] Neurobiol. Aging (2000) 21(3):383–421.
406) Griffin WST [Inflammation and neurodegenerative diseases] Am J Clin Nutr (2006) 83(2):470S – 474.
407) Donato R, Sorci G, Riuzzi F, Arcuri C, Bianchi R, Brozzi F, Tubaro C, Giambanco I [S100B’s double life: Intracellular
regulator and extracellular signal] Biochim. Biophys. Acta - Mol. Cell Res. (2009), 1008–22.
408) Sen J, Belli A [S100B in neuropathologic states: The CRP of the brain?] J. Neurosci. Res. (2007) 85:1373–80.
409) Bianchi R, Garbuglia M, Verzini M, Giambanco I, Ivanenkov V V., Dimlich R V, Jamieson GA, Donato R [S-100
(alpha and beta) binding peptide (TRTK-12) blocks S-100/GFAP interaction: identification of a putative S-100 target
epitope within the head domain of GFAP.] Biochim. Biophys. Acta (1996) 1313(3):258–67.
410) Gilquin B, Cannon BR, Hubstenberger A, Moulouel B, Falk E, Merle N, Assard N, Kieffer S, Rousseau D, Wilder PT,
Weber DJ, Baudier J [The calcium-dependent interaction between S100B and the mitochondrial AAA ATPase
ATAD3A and the role of this complex in the cytoplasmic processing of ATAD3A.] Mol. Cell. Biol. (2010) 30:2724–36.
411) Versijpt J, Van Laere K, Dierckx RA, Dumont F, De Deyn PP, Slegers G, Korf J [Scintigraphic visualization of
inflammation in neurodegenerative disorders.] Nucl. Med. Commun. (2003) 24:209–21.
412) 小野寺理 [脊髄小脳変性症の分子病態機序の解明] 臨床神経学. 日本神経学会 (2009) 49(11):750–2.
413) Muchowski PJ [Protein misfolding, amyloid formation, and neurodegeneration: A critical role for molecular
chaperones?] Neuron (2002), 9–12.
414) Barral JM, Broadley SA, Schaffar G, Hartl FU [Roles of molecular chaperones in protein misfolding diseases.] Semin.
Cell Dev. Biol. (2004) 15:17–29.
415) Perutz MF, Johnson T, Suzuki M, Finch JT [Glutamine repeats as polar zippers: their possible role in inherited
neurodegenerative diseases.] Proc. Natl. Acad. Sci. U. S. A. (1994) 91:5355–8.
416) Strong MJ, Kesavapany S, Pant HC [The pathobiology of amyotrophic lateral sclerosis: a proteinopathy?] J.
Neuropathol. Exp. Neurol. (2005) 64:649–64.
417) Miyata T, Oda O, Inagi R, Iida Y, Araki N, Yamada N, Horiuchi S, Taniguchi N, Maeda K, Kinoshita T [β2Microglobulin modified with advanced glycation end products is a major component of hemodialysis-associated
amyloidosis] J. Clin. Invest. (1993) 92:1243–52.
418) Tanji N, Markowitz GS, Fu C, Kislinger T, Taguchi A, Pischetsrieder M, Stern D, Schmidt AM, D’Agati VD
[Expression of advanced glycation end products and their cellular receptor RAGE in diabetic nephropathy and
nondiabetic renal disease.] J. Am. Soc. Nephrol. (2000) 11:1656–66.
419) Araki N, Ueno N, Chakrabarti B, Morino Y, Horiuchi S [Immunochemical evidence for the presence of advanced
glycation end products in human lens proteins and its positive correlation with aging.] J. Biol. Chem. (1992)
267:10211–4.
420) Saishoji T, Higashi T, Ikeda K, Sano H, Jinnouchi Y, Ogawa M, Horiuchi S [Advanced glycation end products
stimulate plasminogen activator activity via GM-CSF in RAW 264.7 cells.] Biochem. Biophys. Res. Commun. (1995)
217:278–85.
421) Westermark GT, Westermark P [Endocrine amyloid--a subject of increasing interest for the next century.] Amyloid
(2000) 7(1):19–22.
422) Rambaran RN, Serpell LC [Amyloid fibrils: abnormal protein assembly.] Prion (2008) 2:112–7.
XII
423) Nishitoh H, Matsuzawa A, Tobiume K, Saegusa K, Takeda K, Inoue K, Hori S, Kakizuka A, Ichijo H [ASK1 is
essential for endoplasmic reticulum stress-induced neuronal cell death triggered by expanded polyglutamine repeats.]
Genes Dev. (2002) 16:1345–55.
424) Hsu A-L, Murphy CT, Kenyon C [Regulation of aging and age-related disease by DAF-16 and heat-shock factor.]
Science (2003) 300:1142–5.
425) Wyttenbach A, Sauvageot O, Carmichael J, Diaz-Latoud C, Arrigo A-P, Rubinsztein DC [Heat shock protein 27
prevents cellular polyglutamine toxicity and suppresses the increase of reactive oxygen species caused by huntingtin.]
Hum. Mol. Genet. (2002) 11:1137–51.
426) 今泉和則 [疾患と ER ストレス] 血管医学 (2008) 9(4):31–7.
427) Bossy-Wetzel E, Schwarzenbacher R, Lipton SA [Molecular pathways to neurodegeneration.] Nat. Med. (2004) 10
Suppl:S2–9.
428) Beck JA, Poulter M, Campbell TA, Uphill JB, Adamson G, Geddes JF, Revesz T, Davis MB, Wood NW, Collinge J,
Tabrizi SJ [Somatic and germline mosaicism in sporadic early-onset Alzheimer’s disease] Hum. Mol. Genet. (2004)
13:1219–24.
429) Guerreiro RJ, Gustafson DR, Hardy J [The genetic architecture of Alzheimer’s disease: Beyond APP, PSENS and
APOE] Neurobiol. Aging (2012), 437–56.
430) Pitas RE, Boyles JK, Lee SH, Hui D, Weisgraber KH [Lipoproteins and their receptors in the central nervous system.
Characterization of the lipoproteins in cerebrospinal fluid and identification of apolipoprotein B,E(LDL) receptors in
the brain.] J. Biol. Chem. (1987) 262:14352–60.
431) Weisgraber KH, Roses AD, Strittmatter WJ [The role of apolipoprotein E in the nervous system.] Curr. Opin. Lipidol.
(1994) 5:110–6.
432) Das HK, McPherson J, Bruns GA, Karathanasis SK, Breslow JL [Isolation, characterization, and mapping to
chromosome 19 of the human apolipoprotein E gene.] J. Biol. Chem. (1985) 260:6240–7.
433) Schmechel DE, Saunders AM, Strittmatter WJ, Crain BJ, Hulette CM, Joo SH, Pericak-Vance MA, Goldgaber D,
Roses AD [Increased amyloid beta-peptide deposition in cerebral cortex as a consequence of apolipoprotein E genotype
in late-onset Alzheimer disease.] Proc. Natl. Acad. Sci. U. S. A. (1993) 90:9649–53.
434) Greenberg SM, Rebeck GW, Vonsattel JP, Gomez-Isla T, Hyman BT [Apolipoprotein E epsilon 4 and cerebral
hemorrhage associated with amyloid angiopathy.] Ann. Neurol. (1995) 38:254–9.
435) Dai XY, Nanko S, Hattori M, Fukuda R, Nagata K, Isse K, Ueki A, Kazamatsuri H [Association of apolipoprotein E4
with sporadic Alzheimer’s disease is more pronounced in early onset type.] Neurosci. Lett. (1994).
436) Yoshizawa T, Yamakawa-Kobayashi K, Komatsuzaki Y, Arinami T, Oguni E, Mizusawa H, Shoji S, Hamaguchi H
[Dose-dependent association of apolipoprotein E allele epsilon 4 with late-onset, sporadic Alzheimer’s disease.] Ann.
Neurol. (1994) 36:656–9.
437) Strittmatter WJ, Saunders AM, Schmechel D, Pericak-Vance M, Enghild J, Salvesen GS, Roses AD [Apolipoprotein
E: high-avidity binding to beta-amyloid and increased frequency of type 4 allele in late-onset familial Alzheimer
disease.] Proc. Natl. Acad. Sci. U. S. A. (1993) 90:1977–81.
438) Corder EH, Saunders AM, Strittmatter WJ, Schmechel DE, Gaskell PC, Small GW, Roses AD, Haines JL,
Pericakvance MA [Gene Dose of Apolipoprotein-E Type-4 Allele and the Risk of Alzheimers-Disease in Late-Onset
Families] Science (80-. ). (1993) 261:921–3.
439) Slooter AJ, Cruts M, Kalmijn S, Hofman A, Breteler MM, Van Broeckhoven C, van Duijn CM [Risk estimates of
dementia by apolipoprotein E genotypes from a population-based incidence study: the Rotterdam Study.] Arch.
Neurol. (1998).
440) Minagawa H, Gong JS, Jung CG, Watanabe A, Lund-Katz S, Phillips MC, Saito H, Michikawa M [Mechanism
underlying apolipoprotein E (ApoE) isoform-dependent lipid efflux from neural cells in culture] J. Neurosci. Res.
(2009) 87:2498–508.
441) Goldstein JL, Brown MS [The LDL receptor locus and the genetics of familial hypercholesterolemia.] Annu. Rev.
Genet. (1979) 13:259–89.
442) Parkyn CJ, Vermeulen EGM, Mootoosamy RC, Sunyach C, Jacobsen C, Oxvig C, Moestrup S, Liu Q, Bu G, Jen A,
Morris RJ [LRP1 controls biosynthetic and endocytic trafficking of neuronal prion protein.] J. Cell Sci. (2008)
121:773–83.
443) Jen A, Parkyn CJ, Mootoosamy RC, Ford MJ, Warley A, Liu Q, Bu G, Baskakov I V, Moestrup S, McGuinness L,
Emptage N, Morris RJ [Neuronal low-density lipoprotein receptor-related protein 1 binds and endocytoses prion
fibrils via receptor cluster 4.] J. Cell Sci. (2010) 123:246–55.
444) Deane R, Sagare A, Hamm K, Parisi M, Lane S, Finn MB, Holtzman DM, Zlokovic B V. [apoE isoform-specific
disruption of amyloid beta peptide clearance from mouse brain.] J. Clin. Invest. (2008) 118(12):4002–13.
445) Gonias SL, Campana WM [LDL receptor-related protein-1: A regulator of inflammation in atherosclerosis, cancer,
and injury to the nervous system] Am. J. Pathol. (2014), 18–27.
446) Prasansuklab A, Tencomnao T [Amyloidosis in Alzheimer’s Disease: The Toxicity of Amyloid Beta (A β ), Mechanisms
of Its Accumulation and Implications of Medicinal Plants for Therapy.] Evid. Based. Complement. Alternat. Med.
(2013) 2013:413808.
447) Willcox BJ, Donlon TA, He Q, Chen R, Grove JS, Yano K, Masaki KH, Willcox DC, Rodriguez B, Curb JD [FOXO3A
genotype is strongly associated with human longevity.] Proc. Natl. Acad. Sci. U. S. A. (2008) 105:13987–92.
448) Flachsbart F, Caliebe A, Kleindorp R, Blanché H, von Eller-Eberstein H, Nikolaus S, Schreiber S, Nebel A
[Association of FOXO3A variation with human longevity confirmed in German centenarians.] Proc. Natl. Acad. Sci. U.
S. A. (2009) 106:2700–5.
449) Hara K, Boutin P, Mori Y, Tobe K, Dina C, Yasuda K, Yamauchi T, Otabe S, Okada T, Eto K, Kadowaki H, Hagura
R, Akanuma Y, Yazaki Y, Nagai R, Taniyama M, Matsubara K, Yoda M, Nakano Y, Tomita M, Kimura S, Ito C,
Froguel P, Kadowaki T [Genetic variation in the gene encoding adiponectin is associated with an increased risk of
type 2 diabetes in the Japanese population.] Diabetes (2002) 51:536–40.
450) 門脇孝 [脂肪細胞によるインスリン抵抗性の分子機構] 第 124 回日本医学会シンポジウム記録集 肥満の科学 (2003)
110–8.
451) Sodeyama N, Mizusawa H, Yamada M, Itoh Y, Otomo E, Matsushita M [Lack of association of neprilysin
polymorphism with Alzheimer’s disease and Alzheimer's disease-type neuropathological changes.] J. Neurol.
Neurosurg. Psychiatry (2001), 817–8.
XIII
452) Elkins JS, Douglas VC, Johnston SC [Alzheimer disease risk and genetic variation in ACE: a meta-analysis.]
Neurology (2004) 62:363–8.
453) Lehmann DJ, Cortina-Borja M, Warden DR, Smith AD, Sleegers K, Prince JA, Van Duijn CM, Kehoe PG [Large
meta-analysis establishes the ACE insertion-deletion polymorphism as a marker of Alzheimer’s disease] Am. J.
Epidemiol. (2005) 162:305–17.
454) Tiret L, Poirier O, Hallet V, McDonagh TA, Morrison C, McMurray JJ, Dargie HJ, Arveiler D, Ruidavets JB, Luc G,
Evans A, Cambien F [The Lys198Asn polymorphism in the endothelin-1 gene is associated with blood pressure in
overweight people.] Hypertension (1999) 33:1169–74.
455) Ezzidi I, Turki A, Messaoudi S, Chaieb M, Kacem M, Al-Khateeb GM, Mahjoub T, Almawi WY, Mtiraoui N [Common
polymorphisms of calpain-10 and the risk of Type 2 Diabetes in a Tunisian Arab population: a case-control study.]
BMC Med. Genet. (2010) 11:75.
456) Karamohamed S, Demissie S, Volcjak J, Liu CY, Heard-Costa N, Liu J, Shoemaker CM, Panhuysen CI, Meigs JB,
Wilson P, Atwood LD, Cupples LA, Herbert A [Polymorphisms in the insulin-degrading enzyme gene are associated
with type 2 diabetes in men from the NHLBI Framingham Heart Study] Diabetes (2003) 52:1562–7.
457) Jeong B-H, Lee K-H, Lee Y-J, Yun J, Park Y-J, Bae Y, Kim Y-H, Cho Y-S, Choi E-K, Carp RI, Kim Y-S [Genetic
association of a cathepsin D polymorphism and sporadic Creutzfeldt-Jakob disease.] Dement. Geriatr. Cogn. Disord.
(2009) 28:302–6.
458) Marzec JM, Christie JD, Reddy SP, Jedlicka AE, Vuong H, Lanken PN, Aplenc R, Yamamoto T, Yamamoto M, Cho
H-Y, Kleeberger SR [Functional polymorphisms in the transcription factor NRF2 in humans increase the risk of acute
lung injury.] FASEB J. (2007) 21:2237–46.
459) Vitek MP, Bhattacharya K, Glendening JM, Stopa E, Vlassara H, Bucala R, Manogue K, Cerami A [Advanced
glycation end products contribute to amyloidosis in Alzheimer disease.] Proc. Natl. Acad. Sci. U. S. A. (1994) 91:4766–
70.
460) 山口晴保 [アルツハイマー病における β アミロイド沈着と PHF 形成] 静岡総合研究機構 (1996).
461) Shirotani K, Tsubuki S, Iwata N, Takaki Y, Harigaya W, Maruyama K, Kiryu-Seo S, Kiyama H, Iwata H, Tomita T,
Iwatsubo T, Saido TC [Neprilysin degrades both amyloid beta peptides 1-40 and 1-42 most rapidly and efficiently
among thiorphan- and phosphoramidon-sensitive endopeptidases.] J. Biol. Chem. (2001) 276(24):21895–901.
462) Kanemitsu H, Tomiyama T, Mori H [Human neprilysin is capable of degrading amyloid β peptide not only in the
monomeric form but also the pathological oligomeric form] Neurosci. Lett. (2003) 350:113–6.
463) Grimm MOW, Mett J, Stahlmann CP, Haupenthal VJ, Zimmer VC, Hartmann T [Neprilysin and Aβ Clearance:
Impact of the APP Intracellular Domain in NEP Regulation and Implications in Alzheimer’s Disease.] Front. Aging
Neurosci. (2013) 5:98.
464) Hayashi M, Yamashita A, Shimizu K [Somatostatin and brain-derived neurotrophic factor mRNA expression in the
primate brain: decreased levels of mRNAs during aging.] Brain Res. (1997) 749:283–9.
465) Lu T, Pan Y, Kao SY, Li C, Kohane I, Chan J, Yankner BA [Gene regulation and DNA damage in the ageing human
brain] Nature (2004) 429:883–91.
466) Deane R, Du Yan S, Submamaryan RK, LaRue B, Jovanovic S, Hogg E, Welch D, Manness L, Lin C, Yu J, Zhu H,
Ghiso J, Frangione B, Stern A, Schmidt AM, Armstrong DL, Arnold B, Liliensiek B, Nawroth P, Hofman F, Kindy M,
Stern D, Zlokovic B [RAGE mediates amyloid-beta peptide transport across the blood-brain barrier and accumulation
in brain.] Nat. Med. (2003) 9:907–13.
467) Dries DR, Yu G, Herz J [Extracting β-amyloid from Alzheimer’s disease.] Proc. Natl. Acad. Sci. U. S. A. (2012)
109(9):3199–200.
468) Yan SD, Chen X, Fu J, Chen M, Zhu H, Roher A, Slattery T, Zhao L, Nagashima M, Morser J, Migheli A, Nawroth P,
Stern D, Schmidt AM [RAGE and amyloid-beta peptide neurotoxicity in Alzheimer’s disease.] Nature (1996) 382:685–
91.
469) Seubert P, Vigo-Pelfrey C, Esch F, Lee M, Dovey H, Davis D, Sinha S, Schlossmacher M, Whaley J, Swindlehurst C
[Isolation and quantification of soluble Alzheimer’s beta-peptide from biological fluids.] Nature (1992) 359:325–7.
470) Shoji M, Golde TE, Ghiso J, Cheung TT, Estus S, Shaffer LM, Cai XD, McKay DM, Tintner R, Frangione B
[Production of the Alzheimer amyloid beta protein by normal proteolytic processing.] Science (1992) 258:126–9.
471) Shoji M, Kanai M, Matsubara E, Ikeda M, Harigaya Y, Okamoto K, Hirai S [Taps to Alzheimer’s patients: a
continuous Japanese study of cerebrospinal fluid biomarkers.] Ann. Neurol. (2000) 48(3):402.
472) Deane R, Wu Z, Sagare A, Davis J, Du Yan S, Hamm K, Xu F, Parisi M, LaRue B, Hu HW, Spijkers P, Guo H, Song
X, Lenting PJ, Van Nostrand WE, Zlokovic B V. [LRP/amyloid beta-peptide interaction mediates differential brain
efflux of Abeta isoforms.] Neuron (2004) 43(3):333–44.
473) Hammad SM, Ranganathan S, Loukinova E, Twal WO, Argraves WS [Interaction of apolipoprotein J-amyloid betapeptide complex with low density lipoprotein receptor-related protein-2/megalin. A mechanism to prevent pathological
accumulation of amyloid beta-peptide.] J. Biol. Chem. (1997) 272(30):18644–9.
474) Sousa MM, Norden AGW, Jacobsen C, Willnow TE, Christensen EI, Thakker R V., Verroust PJ, Moestrup SK,
Saraiva MJ [Evidence for the role of megalin in renal uptake of transthyretin] J. Biol. Chem. (2000) 275:38176–81.
475) Carro E, Spuch C, Trejo JL, Antequera D, Torres-Aleman I [Choroid plexus megalin is involved in neuroprotection by
serum insulin-like growth factor I.] J. Neurosci. (2005) 25:10884–93.
476) Dietrich MO, Spuch C, Antequera D, Rodal I, de Yébenes JG, Molina JA, Bermejo F, Carro E [Megalin mediates the
transport of leptin across the blood-CSF barrier] Neurobiol. Aging (2008) 29:902–12.
477) Calero M, Rostagno A, Matsubara E, Zlokovic B, Frangione B, Ghiso J [Apolipoprotein J (clusterin) and Alzheimer’s
disease] Microsc. Res. Tech. (2000), 305–15.
478) Bell RD, Sagare AP, Friedman AE, Bedi GS, Holtzman DM, Deane R, Zlokovic B V [Transport pathways for
clearance of human Alzheimer’s amyloid beta-peptide and apolipoproteins E and J in the mouse central nervous
system.] J. Cereb. Blood Flow Metab. (2007) 27:909–18.
479) Christoforidis M, Schober R, Krohn K [Genetic-morphologic association study: association between the low density
lipoprotein-receptor related protein (LRP) and cerebral amyloid angiopathy] Neuropathol Appl Neurobiol (2005)
31:11–9.
480) Tsubuki S, Takaki Y, Saido TC [Dutch, Flemish, Italian, and Arctic mutations of APP and resistance of Abeta to
physiologically relevant proteolytic degradation] Lancet (2003) 361:1957–8.
XIV
481) Betts V, Leissring MA, Dolios G, Wang R, Selkoe DJ, Walsh DM [Aggregation and catabolism of disease-associated
intra-Abeta mutations: reduced proteolysis of AbetaA21G by neprilysin.] Neurobiol. Dis. (2008) 31(3):442–50.
482) Blackstone C, Sheng M [Protein targeting and calcium signaling microdomains in neuronal cells.] Cell Calcium
(1999) 26:181–92.
483) Garner CC, Nash J, Huganir RL [PDZ domains in synapse assembly and signalling] Trends Cell Biol. (2000), 274–80.
484) Almeida CG, Tampellini D, Takahashi RH, Greengard P, Lin MT, Snyder EM, Gouras GK [Beta-amyloid
accumulation in APP mutant neurons reduces PSD-95 and GluR1 in synapses.] Neurobiol. Dis. (2005) 20(2):187–98.
485) Roselli F, Tirard M, Lu J, Hutzler P, Lamberti P, Livrea P, Morabito M, Almeida OFX [Soluble beta-amyloid1-40
induces NMDA-dependent degradation of postsynaptic density-95 at glutamatergic synapses.] J. Neurosci. (2005)
25:11061–70.
486) Wei W, Nguyen LN, Kessels HW, Hagiwara H, Sisodia S, Malinow R [Amyloid beta from axons and dendrites reduces
local spine number and plasticity.] Nat. Neurosci. (2010) 13:190–6.
487) Ittner LM, Ke YD, Delerue F, Bi M, Gladbach A, van Eersel J, Wölfing H, Chieng BC, Christie MJ, Napier IA, Eckert
A, Staufenbiel M, Hardeman E, Götz J [Dendritic function of tau mediates amyloid-β toxicity in alzheimer’s disease
mouse models] Cell (2010) 142:387–97.
488) Liu M-L, Liu M-J, Shen Y-F, Ryu H, Kim H-J, Klupsch K, Downward J, Hong S-T [Omi is a mammalian heat-shock
protein that selectively binds and detoxifies oligomeric amyloid-beta.] J. Cell Sci. (2009) 122:1917–26.
489) Csermely P [Chaperone overload is a possible contributor to “civilization diseases”] Trends Genet. (2001), 701–4.
490) Nardai G ábor, Csermely P, Söti C [Chaperone function and chaperone overload in the aged. A preliminary analysis]
Exp. Gerontol. (2002), 1257–62.
491) Gagliano N, Grizzi F, Annoni G [Mechanisms of aging and liver functions.] Dig. Dis. (2007) 25:118–23.
492) 藤本充章, 井上幸江, 中井彰 [熱ショック転写因子群の生理機能] 生化学. 日本生化学会 (2004) 76(5):419–28.
493) Morimoto RI [Proteotoxic stress and inducible chaperone networks in neurodegenerative disease and aging.] Genes
Dev. (2008) 22:1427–38.
494) Tu S, Okamoto S-I, Lipton SA, Xu H [Oligomeric Aβ-induced synaptic dysfunction in Alzheimer’s disease.] Mol.
Neurodegener. (2014) 9(1):48.
495) Deshpande A, Mina E, Glabe C, Busciglio J [Different conformations of amyloid beta induce neurotoxicity by distinct
mechanisms in human cortical neurons.] J. Neurosci. (2006) 26:6011–8.
496) Gong Y, Chang L, Viola KL, Lacor PN, Lambert MP, Finch CE, Krafft GA, Klein WL [Alzheimer’s disease-affected
brain: presence of oligomeric A beta ligands (ADDLs) suggests a molecular basis for reversible memory loss.] Proc.
Natl. Acad. Sci. U. S. A. (2003) 100:10417–22.
497) Fukumoto H, Tokuda T, Kasai T, Ishigami N, Hidaka H, Kondo M, Allsop D, Nakagawa M [High-molecular-weight
beta-amyloid oligomers are elevated in cerebrospinal fluid of Alzheimer patients.] FASEB J. (2010) 24:2716–26.
498) Bjorklund NL, Reese LC, Sadagoparamanujam V-M, Ghirardi V, Woltjer RL, Taglialatela G [Absence of amyloid β
oligomers at the postsynapse and regulated synaptic Zn2+ in cognitively intact aged individuals with Alzheimer’s
disease neuropathology.] Mol. Neurodegener. (2012) 7:23.
499) Keller JN, Hanni KB, Markesbery WR [Impaired proteasome function in Alzheimer’s disease.] J. Neurochem. (2000)
75:436–9.
500) Keller JN [Age-related neuropathology, cognitive decline, and Alzheimer’s disease.] Ageing Res. Rev. (2006) 5:1–13.
501) Breusing N, Grune T [Regulation of proteasome-mediated protein degradation during oxidative stress and aging]
Biol. Chem. (2008) 389:203–9.
502) Flanary BE, Sammons NW, Nguyen C, Walker D, Streit WJ [Evidence that aging and amyloid promote microglial
cell senescence.] Rejuvenation Res. (2007) 10:61–74.
503) Mawuenyega KG, Sigurdson W, Ovod V, Munsell L, Kasten T, Morris JC, Yarasheski KE, Bateman RJ [Decreased
clearance of CNS beta-amyloid in Alzheimer’s disease.] Science (2010) 330(6012):1774.
504) Eva R, Dassie E, Caswell PT, Dick G, ffrench-Constant C, Norman JC, Fawcett JW [Rab11 and its effector Rab
coupling protein contribute to the trafficking of beta 1 integrins during axon growth in adult dorsal root ganglion
neurons and PC12 cells.] J. Neurosci. (2010) 30:11654–69.
505) Cleary JP, Walsh DM, Hofmeister JJ, Shankar GM, Kuskowski MA, Selkoe DJ, Ashe KH [Natural oligomers of the
amyloid-beta protein specifically disrupt cognitive function.] Nat. Neurosci. (2005) 8:79–84.
506) Du Y, Dodel RC, Eastwood BJ, Bales KR, Gao F, Lohmüller F, Müller U, Kurz A, Zimmer R, Evans RM, Hake A,
Gasser T, Oertel WH, Griffin WS, Paul SM, Farlow MR [Association of an interleukin 1 alpha polymorphism with
Alzheimer’s disease.] Neurology (2000) 55:480–3.
507) Grimaldi LM, Casadei VM, Ferri C, Veglia F, Licastro F, Annoni G, Biunno I, De Bellis G, Sorbi S, Mariani C, Canal
N, Griffin WS, Franceschi M [Association of early-onset Alzheimer’s disease with an interleukin- 1alpha gene
polymorphism] Ann.Neurol. (2000) 47:361–5.
508) Nicoll JAR, Mrak RE, Graham DI, Stewart J, Wilcock G, MacGowan S, Esiri MM, Murray LS, Dewar D, Love S,
Moss T, Griffin WST [Association of interleukin-1 gene polymorphisms with Alzheimer’s disease] Ann. Neurol. (2000)
47:365–8.
509) Grupe A, Li Y, Rowland C, Nowotny P, Hinrichs AL, Smemo S, Kauwe JSK, Maxwell TJ, Cherny S, Doil L, Tacey K,
van Luchene R, Myers A, Wavrant-De Vrièze F, Kaleem M, Hollingworth P, Jehu L, Foy C, Archer N, Hamilton G,
Holmans P, Morris CM, Catanese J, Sninsky J, White TJ, Powell J, Hardy J, O’Donovan M, Lovestone S, et al. [A
scan of chromosome 10 identifies a novel locus showing strong association with late-onset Alzheimer disease.] Am. J.
Hum. Genet. (2006) 78:78–88.
510) Hoozemans JJM, van Haastert ES, Nijholt DAT, Rozemuller AJM, Eikelenboom P, Scheper W [The unfolded protein
response is activated in pretangle neurons in Alzheimer’s disease hippocampus.] Am. J. Pathol. (2009) 174:1241–51.
511) Hoshino T, Nakaya T, Araki W, Suzuki K, Suzuki T, Mizushima T [Endoplasmic reticulum chaperones inhibit the
production of amyloid-beta peptides.] Biochem. J. (2007) 402:581–9.
512) Lee DY, Lee K-S, Lee HJ, Kim DH, Noh YH, Yu K, Jung H-Y, Lee SH, Lee JY, Youn YC, Jeong Y, Kim DK, Lee WB,
Kim SS [Activation of PERK signaling attenuates Abeta-mediated ER stress.] PLoS One (2010) 5:e10489.
513) Kim H-J, Cho H-K, Kwon YH [Synergistic induction of ER stress by homocysteine and beta-amyloid in SH-SY5Y
cells.] J. Nutr. Biochem. (2008) 19:754–61.
514) Meng Y, Xu H, Wang R, Ji Z, Yu S, Zhou J, Sheng S [Impairment of signal transduction pathway on neuronal
survival in brains of Alzheimer’s disease] Zhonghua Bing Li Xue Za Zhi (2002) 31:502–5.
XV
515) 大田秀隆 [長寿遺伝子による血管老化制御機構] 日本老年医学会雑誌. 日本老年医学会 (2007) 44(2):194–7.
516) Fargnoli J, Kunisada T, Fornace AJ, Schneider EL, Holbrook NJ [Decreased expression of heat shock protein 70
mRNA and protein after heat treatment in cells of aged rats.] Proc. Natl. Acad. Sci. U. S. A. (1990) 87:846–50.
517) Saraiva LM, Seixas da Silva GS, Galina A, Da-Silva WS, Klein WL, Ferreira ST, De Felice FG [Amyloid-β triggers
the release of neuronal hexokinase 1 from mitochondria.] PLoS One (2010) 5(12):e15230.
518) Alberdi E, Sánchez-Gómez MV, Cavaliere F, Pérez-Samartín A, Zugaza JL, Trullas R, Domercq M, Matute C
[Amyloid beta oligomers induce Ca2+ dysregulation and neuronal death through activation of ionotropic glutamate
receptors.] Cell Calcium (2010) 47(3):264–72.
519) De Felice FG [Alzheimer’s disease and insulin resistance: translating basic science into clinical applications.] J. Clin.
Invest. (2013) 123:531–9.
520) McGeer PL, Schulzer M, McGeer EG [Arthritis and anti-inflammatory agents as possible protective factors for
Alzheimer’s disease: a review of 17 epidemiologic studies.] Neurology (1996) 47:425–32.
521) Patrono C, Patrignani P, García Rodríguez LA [Cyclooxygenase-selective inhibition of prostanoid formation:
transducing biochemical selectivity into clinical read-outs.] J. Clin. Invest. American Society for Clinical Investigation
(2001) 108(1):7–13.
522) Zhou Y, Su Y, Li B, Liu F, Ryder JW, Wu X, Gonzalez-DeWhitt PA, Gelfanova V, Hale JE, May PC, Paul SM, Ni B
[Nonsteroidal anti-inflammatory drugs can lower amyloidogenic Abeta42 by inhibiting Rho.] Science (2003) 302:1215–
7.
523) Tang BL [Alzheimer’s disease: Channeling APP to non-amyloidogenic processing] Biochem. Biophys. Res. Commun.
(2005), 375–8.
524) Weggen S, Eriksen JL, Das P, Sagi SA, Wang R, Pietrzik CU, Findlay KA, Smith TE, Murphy MP, Bulter T, Kang
DE, Marquez-Sterling N, Golde TE, Koo EH [A subset of NSAIDs lower amyloidogenic Abeta42 independently of
cyclooxygenase activity.] Nature (2001) 414:212–6.
525) Pei Z, Pang H, Qian L, Yang S, Wang T, Zhang W, Wu X, Dallas S, Wilson B, Reece JM, Miller DS, Hong JS, Block
ML [MAC1 mediates LPS-induced production of superoxide by microglia: The role of pattern recognition receptors in
dopaminergic neurotoxicity] Glia (2007) 55:1362–73.
526) Hu X, Zhang D, Pang H, Caudle WM, Li Y, Gao H, Liu Y, Qian L, Wilson B, Di Monte DA, Ali SF, Zhang J, Block
ML, Hong J-S [Macrophage antigen complex-1 mediates reactive microgliosis and progressive dopaminergic
neurodegeneration in the MPTP model of Parkinson’s disease.] J. Immunol. (2008) 181:7194–204.
527) Cameron B, Landreth GE [Inflammation, microglia, and alzheimer’s disease] Neurobiol. Dis. (2010), 503–9.
528) 斎藤博久 [アレルギーはなぜおこるか?] ブルーバックス (2003).
529) Biessels GJ, Kamal A, Ramakers GM, Urban IJ, Spruijt BM, Erkelens DW, Gispen WH [Place learning and
hippocampal synaptic plasticity in streptozotocin-induced diabetic rats.] Diabetes (1996) 45:1259–66.
530) Van den Berg E, Reijmer YD, de Bresser J, Kessels RP, Kappelle LJ, Biessels GJ, Utrecht Diabetic Encephalopathy
Study G [A 4 year follow-up study of cognitive functioning in patients with type 2 diabetes mellitus] Diabetologia
(2010) 53:58–65.
531) Matsuzaki T, Sasaki K, Tanizaki Y, Hata J, Fujimi K, Matsui Y, Sekita A, Suzuki SO, Kanba S, Kiyohara Y, Iwaki T
[Insulin resistance is associated with the pathology of Alzheimer disease: The Hisayama study] Neurology (2010)
75:764–70.
532) Kumari M, Marmot M [Diabetes and cognitive function in a middle-aged cohort: Findings from the Whitehall II
study] Neurology (2005) 65:1597–603.
533) Ott A, Stolk RP, Hofman A, Van Harskamp F, Grobbee DE, Breteler MMB [Association of diabetes mellitus and
dementia: The Rotterdam Study] Diabetologia (1996) 39:1392–7.
534) De la Monte SM, Tong M, Lester-Coll N, Plater M, Wands JR [Therapeutic rescue of neurodegeneration in
experimental type 3 diabetes: relevance to Alzheimer’s disease.] J. Alzheimers. Dis. (2006) 10:89–109.
535) Van Obberghen E, Baron V, Delahaye L, Emanuelli B, Filippa N, Giorgetti-Peraldi S, Lebrun P, Mothe-Satney I,
Peraldi P, Rocchi S, Sawka-Verhelle D, Tartare-Deckert S, Giudicelli J [Surfing the insulin signaling web] Eur. J.
Clin. Invest. (2001), 966–77.
536) Johnston AM, Pirola L, Van Obberghen E [Molecular mechanisms of insulin receptor substrate protein-mediated
modulation of insulin signalling] FEBS Lett. (2003), 32–6.
537) Bevan P [Insulin signalling] J. Cell Sci. (2001) 114(8):1429–30.
538) Lizcano JM, Alessi DR [The insulin signalling pathway.] Curr. Biol. (2002) 12(7):R236–8.
539) Cohen P [The hormonal control of glycogen metabolism in mammalian muscle by multivalent phosphorylation.]
Biochem. Soc. Trans. (1979) 7(3):459–80.
540) Embi N, Rylatt DB, Cohen P [Glycogen synthase kinase-3 from rabbit skeletal muscle. Separation from cyclic-AMPdependent protein kinase and phosphorylase kinase.] Eur. J. Biochem. (1980) 107(2):519–27.
541) 荒木栄一, 近藤龍也 [インスリンシグナリングの調節とその障害] 日本薬理学雑誌 (2005) 125:120–4.
542) Woodgett JR [cDNA cloning and properties of glycogen synthase kinase-3.] Methods Enzymol. (1991) 200:564–77.
543) Woodgett JR [Molecular cloning and expression of glycogen synthase kinase-3/factor A.] EMBO J. (1990) 9(8):2431–8.
544) Plyte SE, Hughes K, Nikolakaki E, Pulverer BJ, Woodgett JR [Glycogen synthase kinase-3: Functions in oncogenesis
and development] Biochim. Biophys. Acta - Rev. Cancer (1992), 147–62.
545) Frame S, Cohen P [GSK3 takes centre stage more than 20 years after its discovery.] Biochem. J. (2001) 359:1–16.
546) Hemmings BA, Restuccia DF [PI3K-PKB/Akt pathway.] Cold Spring Harb. Perspect. Biol. (2012) 4(9):a011189.
547) Rada P, Rojo AI, Chowdhry S, McMahon M, Hayes JD, Cuadrado A [SCF/{beta}-TrCP promotes glycogen synthase
kinase 3-dependent degradation of the Nrf2 transcription factor in a Keap1-independent manner.] Mol. Cell. Biol.
(2011) 31:1121–33.
548) Salazar M, Rojo AI, Velasco D, de Sagarra RM, Cuadrado A [Glycogen synthase kinase-3beta inhibits the xenobiotic
and antioxidant cell response by direct phosphorylation and nuclear exclusion of the transcription factor Nrf2.] J.
Biol. Chem. (2006) 281:14841–51.
549) Kosaka K, Hagura R, Kuzuya T [Insulin Responses in Equivocal and Definite Diabetes, with Special Reference to
Subjects Who Had Mild Glucose Intolerance but Later Developed Definite Diabetes] Diabetes (1977) 26(10):944–52.
550) Kahn SE, D’Alessio DA, Schwartz MW, Fujimoto WY, Ensinck JW, Taborsky GJ, Porte D [Evidence of cosecretion of
islet amyloid polypeptide and insulin bv beta-cells] Diabetes (1990) 39(5):634–8.
XVI
551) Moore CX, Cooper GJ [Co-secretion of amylin and insulin from cultured islet beta-cells: modulation by nutrient
secretagogues, islet hormones and hypoglycemic agents.] Biochem. Biophys. Res. Commun. (1991) 179:1–9.
552) Sanke T, Bell GI, Sample C, Rubenstein AH, Steiner DF [An islet amyloid peptide is derived from an 89-amino acid
precursor by proteolytic processing] J. Biol. Chem. (1988) 263:17243–6.
553) Haataja L, Gurlo T, Huang CJ, Butler PC [Islet amyloid in type 2 diabetes, and the toxic oligomer hypothesis]
Endocr. Rev. (2008), 303–16.
554) 浦野文彦 [小胞体ストレス(ER ストレス)シグナリングの膵臓 β 細胞における役割] 生化学. 日本生化学会 (2007)
79(11):1055–9.
555) Robertson RP, Harmon JS [Pancreatic islet beta-cell and oxidative stress: the importance of glutathione peroxidase.]
FEBS Lett. (2007) 581:3743–8.
556) 石原寿光 [膵 β 細胞のストレス応答・生存と mRNA 翻訳制御] 生化學. 日本生化学会 (2009) 81(6):474–85.
557) Lipson KL, Fonseca SG, Urano F [Endoplasmic reticulum stress-induced apoptosis and auto-immunity in diabetes.]
Curr. Mol. Med. (2006) 6:71–7.
558) Tuncman G, Hirosumi J, Solinas G, Chang L, Karin M, Hotamisligil GS [Functional in vivo interactions between
JNK1 and JNK2 isoforms in obesity and insulin resistance.] Proc. Natl. Acad. Sci. U. S. A. (2006) 103:10741–6.
559) Vallerie SN, Furuhashi M, Fucho R, Hotamisligil GS [A predominant role for parenchymal c-Jun amino terminal
kinase (JNK) in the regulation of systemic insulin sensitivity] PLoS One (2008) 3.
560) Huang CJ, Lin CY, Haataja L, Gurlo T, Butler AE, Rizza RA, Butler PC [High expression rates of human islet
amyloid polypeptide induce endoplasmic reticulum stress mediated beta-cell apoptosis, a characteristic of humans
with type 2 but not type 1 diabetes] Diabetes (2007) 56:2016–27.
561) Peters A [The selfish brain: Competition for energy resources] Am. J. Hum. Biol. (2011), 29–34.
562) Pardridge WM, Boado RJ, Farrell CR [Brain-type glucose transporter (GLUT-1) is selectively localized to the bloodbrain barrier: Studies with quantitative Western blotting and in situ hybridization] J. Biol. Chem. (1990) 265:18035–
40.
563) Nagamatsu S, Sawa H, Kamada K, Nakamichi Y, Yoshimoto K, Hoshino T [Neuron-specific glucose transporter
(NSGT): CNS distribution of GLUT3 rat glucose transporter (RGT3) in rat central neurons] FEBS Lett. (1993)
334:289–95.
564) Neumann KF, Rojo L, Navarrete LP, Farías G, Reyes P, Maccioni RB [Insulin resistance and Alzheimer’s disease:
molecular links & clinical implications.] Curr. Alzheimer Res. (2008) 5:438–47.
565) Banks WA, Jaspan JB, Huang W, Kastin AJ [Transport of insulin across the blood-brain barrier: saturability at
euglycemic doses of insulin.] Peptides (1997) 18(9):1423–9.
566) Rulifson EJ, Kim SK, Nusse R [Ablation of insulin-producing neurons in flies: growth and diabetic phenotypes.]
Science (2002) 296:1118–20.
567) Heidenreich KA [Insulin and IGF-I receptor signaling in cultured neurons.] Ann. N. Y. Acad. Sci. (1993) 692:72–88.
568) Robinson LJ, Leitner W, Draznin B, Heidenreich KA [Evidence that p21ras mediates the neurotrophic effects of
insulin and insulin-like growth factor I in chick forebrain neurons] Endocrinology (1994) 135:2568–73.
569) Talbot K, Wang HY, Kazi H, Han LY, Bakshi KP, Stucky A, Fuino RL, Kawaguchi KR, Samoyedny AJ, Wilson RS,
Arvanitakis Z, Schneider JA, Wolf BA, Bennett DA, Trojanowski JQ, Arnold SE [Demonstrated brain insulin
resistance in Alzheimer’s disease patients is associated with IGF-1 resistance, IRS-1 dysregulation, and cognitive
decline] J. Clin. Invest. (2012) 122:1316–38.
570) Fernandez AM, Torres-Alemán I [The many faces of insulin-like peptide signalling in the brain] Nat. Rev. Neurosci.
(2012), 225–39.
571) Ishihara H, Takeda S, Tamura A, Takahashi R, Yamaguchi S, Takei D, Yamada T, Inoue H, Soga H, Katagiri H,
Tanizawa Y, Oka Y [Disruption of the WFS1 gene in mice causes progressive β-cell loss and impaired stimulus Secretion coupling in insulin secretion] Hum. Mol. Genet. (2004) 13:1159–70.
572) Yamada T, Ishihara H, Tamura A, Takahashi R, Yamaguchi S, Takei D, Tokita A, Satake C, Tashiro F, Katagiri H,
Aburatani H, Miyazaki JI, Oka Y [WFS1-deficiency increases endoplasmic reticulum stress, impairs cell cycle
progression and triggers the apoptotic pathway specifically in pancreatic β-cells] Hum. Mol. Genet. (2006) 15:1600–9.
573) DuRose JB, Scheuner D, Kaufman RJ, Rothblum LI, Niwa M [Phosphorylation of eukaryotic translation initiation
factor 2alpha coordinates rRNA transcription and translation inhibition during endoplasmic reticulum stress.] Mol.
Cell. Biol. (2009) 29:4295–307.
574) Takashima A [GSK-3 is essential in the pathogenesis of Alzheimer’s disease.] J. Alzheimers. Dis. (2006) 9:309–17.
575) Baltzis D, Pluquet O, Papadakis AI, Kazemi S, Qu L-K, Koromilas AE [The eIF2alpha kinases PERK and PKR
activate glycogen synthase kinase 3 to promote the proteasomal degradation of p53.] J. Biol. Chem. (2007)
282(43):31675–87.
576) Cross DAE, Culbert AA, Chalmers KA, Facci L, Skaper SD, Reith AD [Selective small-molecule inhibitors of glycogen
synthase kinase-3 activity protect primary neurones from death] J. Neurochem. (2001) 77:94–102.
577) Pap M, Cooper GM [Role of glycogen synthase kinase-3 in the phosphatidylinositol 3-Kinase/Akt cell survival
pathway.] J. Biol. Chem. (1998) 273:19929–32.
578) Zhao W-Q, Chen H, Quon MJ, Alkon DL [Insulin and the insulin receptor in experimental models of learning and
memory.] Eur. J. Pharmacol. (2004) 490:71–81.
579) Zhao W-Q, Alkon DL [Role of insulin and insulin receptor in learning and memory] Mol. Cell. Endocrinol. (2001),
125–34.
580) Zhao W-Q, De Felice FG, Fernandez S, Chen H, Lambert MP, Quon MJ, Krafft GA, Klein WL [Amyloid beta
oligomers induce impairment of neuronal insulin receptors.] FASEB J. (2008) 22:246–60.
581) De Felice FG, Vieira MNN, Bomfim TR, Decker H, Velasco PT, Lambert MP, Viola KL, Zhao W-Q, Ferreira ST, Klein
WL [Protection of synapses against Alzheimer’s-linked toxins: insulin signaling prevents the pathogenic binding of
Abeta oligomers.] Proc. Natl. Acad. Sci. U. S. A. (2009) 106:1971–6.
582) Takeda S, Sato N, Uchio-Yamada K, Sawada K, Kunieda T, Takeuchi D, Kurinami H, Shinohara M, Rakugi H,
Morishita R [Diabetes-accelerated memory dysfunction via cerebrovascular inflammation and Abeta deposition in an
Alzheimer mouse model with diabetes.] Proc. Natl. Acad. Sci. U. S. A. (2010) 107:7036–41.
583) Bomfim TR, Forny-Germano L, Sathler LB, Brito-Moreira J, Houzel JC, Decker H, Silverman MA, Kazi H, Melo HM,
McClean PL, Holscher C, Arnold SE, Talbot K, Klein WL, Munoz DP, Ferreira ST, De Felice FG [An anti-diabetes
XVII
agent protects the mouse brain from defective insulin signaling caused by Alzheimer’s disease-associated Aβ
oligomers] J. Clin. Invest. (2012) 122:1339–53.
584) Salkovic-Petrisic M, Hoyer S [Central insulin resistance as a trigger for sporadic Alzheimer-like pathology: an
experimental approach.] J. Neural Transm. Suppl. (2007) (72):217–33.
585) Carro E, Trejo JL, Gerber A, Loetscher H, Torrado J, Metzger F, Torres-Aleman I [Therapeutic actions of insulin-like
growth factor I on APP/PS2 mice with severe brain amyloidosis.] Neurobiol. Aging (2006) 27(9):1250–7.
586) Kovacs P, Hajnal A [In vivo electrophysiological effects of insulin in the rat brain] Neuropeptides (2009) 43:283–93.
587) Craft S [Alzheimer disease: Insulin resistance and AD—extending the translational path] Nat. Rev. Neurol. (2012),
360–2.
588) Van Der Heide LP, Kamal A, Artola A, Gispen WH, Ramakers GMJ [Insulin modulates hippocampal activitydependent synaptic plasticity in a N-methyl-D-aspartate receptor and phosphatidyl-inositol-3-kinase-dependent
manner] J. Neurochem. (2005) 94:1158–66.
589) Chiu SL, Chen CM, Cline HT [Insulin Receptor Signaling Regulates Synapse Number, Dendritic Plasticity, and
Circuit Function In Vivo] Neuron (2008) 58:708–19.
590) Niswender KD, Morrison CD, Clegg DJ, Olson R, Baskin DG, Myers MG, Seeley RJ, Schwartz MW [Insulin
activation of phosphatidylinositol 3-kinase in the hypothalamic arcuate nucleus: A key mediator of insulin-induced
anorexia] Diabetes (2003) 52:227–31.
591) Ménager C, Arimura N, Fukata Y, Kaibuchi K [PIP3 is involved in neuronal polarization and axon formation.] J.
Neurochem. (2004) 89:109–18.
592) Ma Q-L, Yang F, Rosario ER, Ubeda OJ, Beech W, Gant DJ, Chen PP, Hudspeth B, Chen C, Zhao Y, Vinters H V,
Frautschy SA, Cole GM [Beta-amyloid oligomers induce phosphorylation of tau and inactivation of insulin receptor
substrate via c-Jun N-terminal kinase signaling: suppression by omega-3 fatty acids and curcumin.] J. Neurosci.
(2009) 29:9078–89.
593) Gasparini L, Xu H [Potential roles of insulin and IGF-1 in Alzheimer’s disease] Trends Neurosci. (2003), 404–6.
594) Carro E, Torres-Aleman I [The role of insulin and insulin-like growth factor I in the molecular and cellular
mechanisms underlying the pathology of Alzheimer’s disease] Eur J Pharmacol (2004) 490:127–33.
595) Hong M, Lee VM [Insulin and insulin-like growth factor-1 regulate tau phosphorylation in cultured human neurons.]
J. Biol. Chem. (1997) 272:19547–53.
596) Cheng CM, Tseng V, Wang J, Wang D, Matyakhina L, Bondy CA [Tau is hyperphosphorylated in the insulin-like
growth factor-I null brain.] Endocrinology (2005) 146(12):5086–91.
597) 若月修二, 荒木敏之 [ユビキチン・プロテアソーム系によって制御される軸索変性の分子メカニズム] 生化學. 日本生化
学会 (2012) 84(6):463–71.
598) Zhou F-Q, Snider WD [Cell biology. GSK-3beta and microtubule assembly in axons.] Science (2005) 308:211–4.
599) Yoshimura T, Arimura N, Kaibuchi K [Signaling networks in neuronal polarization.] J. Neurosci. (2006) 26:10626–
30.
600) Martín D, Salinas M, López-Valdaliso R, Serrano E, Recuero M, Cuadrado A [Effect of the Alzheimer amyloid
fragment Aβ(25-35) on Akt/PKB kinase and survival of PC12 cells] J. Neurochem. (2001) 78:1000–8.
601) Wei W, Wang X, Kusiak JW [Signaling events in amyloid beta-peptide-induced neuronal death and insulin-like
growth factor I protection.] J. Biol. Chem. (2002) 277:17649–56.
602) Beurel E, Michalek SM, Jope RS [Innate and adaptive immune responses regulated by glycogen synthase kinase-3
(GSK3)] Trends Immunol. (2010) 31:24–31.
603) Iwata N, Takaki Y, Fukami S, Tsubuki S, Saido TC [Region-specific reduction of Aβ-degrading endopeptidase,
neprilysin, in mouse hippocampus upon aging] J. Neurosci. Res. (2002) 70(3):493–500.
604) Wang S, Wang R, Chen L, Bennett DA, Dickson DW, Wang DS [Expression and functional profiling of neprilysin,
insulin-degrading enzyme, and endothelin-converting enzyme in prospectively studied elderly and Alzheimer’s brain]
J. Neurochem. (2010) 115:47–57.
605) Miners JS, Baig S, Tayler H, Kehoe PG, Love S [Neprilysin and insulin-degrading enzyme levels are increased in
Alzheimer disease in relation to disease severity.] J. Neuropathol. Exp. Neurol. (2009) 68(8):902–14.
606) Miners JS, Barua N, Kehoe PG, Gill S, Love S [Aβ-Degrading Enzymes: Potential for Treatment of Alzheimer
Disease] J. Neuropathol. Exp. Neurol. (2011), 944–59.
607) Nguyen HT, Kirk JK, Arcury TA, Ip EH, Grzywacz JG, Saldana SJ, Bell RA, Quandt SA [Cognitive function is a risk
for health literacy in older adults with diabetes] Diabetes Res. Clin. Pract. (2013) 101:141–7.
608) Rivera EJ, Goldin A, Fulmer N, Tavares R, Wands JR, de la Monte SM [Insulin and insulin-like growth factor
expression and function deteriorate with progression of Alzheimer’s disease: link to brain reductions in acetylcholine.]
J. Alzheimers. Dis. (2005) 8:247–68.
609) Schiöth HB, Craft S, Brooks SJ, Frey WH, Benedict C [Brain insulin signaling and Alzheimer’s disease: Current
evidence and future directions] Mol. Neurobiol. (2012) 46:4–10.
610) Perry EK, Gibson PH, Blessed G, Perry RH, Tomlinson BE [Neurotransmitter enzyme abnormalities in senile
dementia. Choline acetyltransferase and glutamic acid decarboxylase activities in necropsy brain tissue] J. Neurol.
Sci. (1977) 34:247–65.
611) Auld DS, Kornecook TJ, Bastianetto S, Quirion R [Alzheimer’s disease and the basal forebrain cholinergic system:
relations to beta-amyloid peptides, cognition, and treatment strategies.] Prog. Neurobiol. (2002) 68:209–45.
612) Perry EK, Haroutunian V, Davis KL, Levy R, Lantos P, Eagger S, Honavar M, Dean A, Griffiths M, McKeith IG
[Neocortical cholinergic activities differentiate Lewy body dementia from classical Alzheimer’s disease.] Neuroreport
(1994).
613) Tiraboschi P, Hansen LA, Alford M, Merdes A, Masliah E, Thal LJ, Corey-Bloom J [Early and widespread cholinergic
losses differentiate dementia with Lewy bodies from Alzheimer disease.] Arch. Gen. Psychiatry (2002) 59:946–51.
614) Kar S, Slowikowski SPM, Westaway D, Mount HTJ [Interactions between β-amyloid and central cholinergic neurons :
implications for Alzheimer ’ s disease] J. Psychiatry Neurosci. (2004) 29:427–42.
615) Bobich JA, Zheng Q, Campbell A [Incubation of nerve endings with a physiological concentration of Abeta1-42
activates CaV2.2(N-Type)-voltage operated calcium channels and acutely increases glutamate and noradrenaline
release.] J. Alzheimers. Dis. (2004) 6:243–55.
616) Appleyard ME, Smith AD, Wilcock GK, Esiri MM [Decreased CSF acetylcholinesterase activity in Alzheimer’s
disease.] Lancet (1983), 452.
XVIII
617) Pedersen WA, Kloczewiak MA, Blusztajn JK [Amyloid beta-protein reduces acetylcholine synthesis in a cell line
derived from cholinergic neurons of the basal forebrain.] Proc. Natl. Acad. Sci. U. S. A. (1996) 93:8068–71.
618) Price DL, Whitehouse PJ, Struble RG [Alzheimer’s disease.] Annu. Rev. Med. Annual Reviews 4139 El Camino Way,
P.O. Box 10139, Palo Alto, CA 94303-0139, USA (1985) 36:349–56.
619) Wang HY, Lee DH, D’Andrea MR, Peterson PA, Shank RP, Reitz AB [beta-Amyloid(1-42) binds to alpha7 nicotinic
acetylcholine receptor with high affinity. Implications for Alzheimer’s disease pathology.] J. Biol. Chem. (2000)
275:5626–32.
620) Pettit DL, Shao Z, Yakel JL [beta-Amyloid(1-42) peptide directly modulates nicotinic receptors in the rat
hippocampal slice.] J. Neurosci. (2001) 21:RC120.
621) Coyle JT, Price DL, DeLong MR [Alzheimer’s disease: a disorder of cortical cholinergic innervation.] Science (1983)
219:1184–90.
622) Fulga TA, Elson-Schwab I, Khurana V, Steinhilb ML, Spires TL, Hyman BT, Feany MB [Abnormal bundling and
accumulation of F-actin mediates tau-induced neuronal degeneration in vivo.] Nat. Cell Biol. (2007) 9:139–48.
623) Whiteman IT, Gervasio OL, Cullen KM, Guillemin GJ, Jeong E V, Witting PK, Antao ST, Minamide LS, Bamburg
JR, Goldsbury C [Activated actin-depolymerizing factor/cofilin sequesters phosphorylated microtubule-associated
protein during the assembly of alzheimer-like neuritic cytoskeletal striations.] J. Neurosci. (2009) 29(41):12994–3005.
624) Beglinger LJ, Gaydos BL, Kareken DA, Tangphao-Daniels O, Siemers ER, Mohs RC [Neuropsychological test
performance in healthy volunteers before and after donepezil administration.] J. Psychopharmacol. (2004).
625) Doody RS, Geldmacher DS, Gordon B, Perdomo CA, Pratt RD [Open-label, multicenter, phase 3 extension study of
the safety and efficacy of donepezil in patients with Alzheimer disease.] Arch. Neurol. (2001).
626) Ahmed MM, Hoshino H, Chikuma T, Yamada M, Kato T [Effect of memantine on the levels of glial cells,
neuropeptides, and peptide-degrading enzymes in rat brain regions of ibotenic acid-treated Alzheimer’s disease
model] Neuroscience (2004) 126(3):639–49.
627) Reisberg B, Doody R, Stöffler A, Schmitt F, Ferris S, Möbius HJ [Memantine in moderate-to-severe Alzheimer’s
disease.] N. Engl. J. Med. (2003).
628) Raina P, Santaguida P, Ismaila A, Patterson C, Cowan D, Levine M, Booker L, Oremus M [Effectiveness of
cholinesterase inhibitors and memantine for treating dementia: Evidence review for a clinical practice guideline] Ann.
Intern. Med. (2008), 379–97.
629) Reger MA, Watson GS, Green PS, Baker LD, Cholerton B, Fishel MA, Plymate SR, Cherrier MM, Schellenberg GD,
Frey WH, Craft S [Intranasal insulin administration dose-dependently modulates verbal memory and plasma
amyloid-beta in memory-impaired older adults.] J. Alzheimers. Dis. (2008) 13:323–31.
630) Ohtsuki S, Hirayama M, Ito S, Uchida Y, Tachikawa M, Terasaki T [Quantitative targeted proteomics for
understanding the blood-brain barrier: towards pharmacoproteomics.] Expert Rev. Proteomics (2014) 11(3):303–13.
631) Lehmann JM, Moore LB, Smith-Oliver TA, Wilkison WO, Willson TM, Kliewer SA [An antidiabetic thiazolidinedione
is a high affinity ligand for peroxisome proliferator-activated receptor gamma (PPAR gamma).] J. Biol. Chem. (1995)
270:12953–6.
632) Berger J, Moller DE [The mechanisms of action of PPARs.] Annu. Rev. Med. (2002) 53:409–35.
633) Wang K, Wan Y-JY [Nuclear receptors and inflammatory diseases.] Exp. Biol. Med. (Maywood). (2008) 233:496–506.
634) Longo FM, Massa SM [Neuroprotective strategies in Alzheimer’s disease.] NeuroRx (2004) 1:117–27.
635) Citron M, Diehl TS, Gordon G, Biere AL, Seubert P, Selkoe DJ [Evidence that the 42- and 40-amino acid forms of
amyloid beta protein are generated from the beta-amyloid precursor protein by different protease activities.] Proc.
Natl. Acad. Sci. U. S. A. (1996) 93:13170–5.
636) Giacobini E [Cholinesterase inhibitor therapy stabilizes symptoms of Alzheimer disease.] Alzheimer Dis. Assoc.
Disord. (2000) 14 Suppl 1:S3–10.
637) Brune K, Neubert A [Pharmacokinetic and pharmacodynamic aspects of the ideal COX-2 inhibitor: a
pharmacologist’s perspective.] Clin. Exp. Rheumatol. 19(6 Suppl 25):S51–7.
638) 延吉正清, 野坂秀行, 鈴木紳, 斎藤滋, 鈴木孝彦, 光藤和明 [冠動脈造影時の冠れん縮に対するNK843(ニトログリセ
リン注射剤)の臨床評価 冠動脈内投与による後期第II相試験] 月刊臨床と研究 (2000) 77(12):2387–96.
639) Bhat V, Weiner WJ [Parkinson’s disease: Diagnosis and the initiation of therapy] Minerva Med. (2005), 145–54.
640) Scharf S, Mander A, Ugoni A, Vajda F, Christophidis N [A double-blind, placebo-controlled trial of
diclofenac/misoprostol in Alzheimer’s disease.] Neurology (1999).
641) Aisen PS, Schafer KA, Grundman M, Pfeiffer E, Sano M, Davis KL, Farlow MR, Jin S, Thomas RG, Thal LJ [Effects
of rofecoxib or naproxen vs placebo on Alzheimer disease progression: a randomized controlled trial.] JAMA (2003).
642) Bem D, Yoshimura SI, Nunes-Bastos R, Bond FF, Kurian MA, Rahman F, Handley MTW, Hadzhiev Y, Masood I,
Straatman-Iwanowska AA, Cullinane AR, McNeill A, Pasha SS, Kirby GA, Foster K, Ahmed Z, Morton JE, Williams
D, Graham JM, Dobyns WB, Burglen L, Ainsworth JR, Gissen P, Müller F, Maher ER, Barr FA, Aligianis IA [Loss-offunction mutations in RAB18 cause Warburg micro syndrome] Am. J. Hum. Genet. (2011) 88:499–507.
643) Kobayashi H, Fukuda M [Rab35 regulates Arf6 activity through centaurin- 2 (ACAP2) during neurite outgrowth] J.
Cell Sci. (2012), 2235–43.
644) Fukuhara S, Chikumi H, Gutkind JS [RGS-containing RhoGEFs: the missing link between transforming G proteins
and Rho?] Oncogene. Nature Publishing Group (2001) 20(13):1661–8.
645) Labouesse M [Epithelium-mesenchyme: a balancing act of RhoGAP and RhoGEF.] Curr. Biol. (2004) 14(13):R508–10.
646) Takashima A, Noguchi K, Michel G, Mercken M, Hoshi M, Ishiguro K, Imahori K [Exposure of rat hippocampal
neurons to amyloid beta peptide (25-35) induces the inactivation of phosphatidyl inositol-3 kinase and the activation
of tau protein kinase I/glycogen synthase kinase-3 beta] Neurosci. Lett. (1996) 203(1):33–6.
647) Pawson T, Nash P [Assembly of cell regulatory systems through protein interaction domains.] Science (2003)
300:445–52.
648) Kobayashi A, Kang M-I, Okawa H, Ohtsuji M, Zenke Y, Chiba T, Igarashi K, Yamamoto M [Oxidative stress sensor
Keap1 functions as an adaptor for Cul3-based E3 ligase to regulate proteasomal degradation of Nrf2.] Mol. Cell. Biol.
(2004) 24:7130–9.
649) Dinkova-Kostova A [Protection Against Cancer by Plant Phenylpropenoids: Induction of Mammalian
Anticarcinogenic Enzymes] Mini-Reviews Med. Chem. Bentham Science Publishers (2002) 2(6):595–610.
XIX
650) Dinkova-Kostova AT, Liby KT, Stephenson KK, Holtzclaw WD, Gao X, Suh N, Williams C, Risingsong R, Honda T,
Gribble GW, Sporn MB, Talalay P [Extremely potent triterpenoid inducers of the phase 2 response: correlations of
protection against oxidant and inflammatory stress.] Proc. Natl. Acad. Sci. U. S. A. (2005) 102:4584–9.
651) Itoh K, Mochizuki M, Ishii Y, Ishii T, Shibata T, Kawamoto Y, Kelly V, Sekizawa K, Uchida K, Yamamoto M
[Transcription factor Nrf2 regulates inflammation by mediating the effect of 15-deoxy-Delta(12,14)-prostaglandin
j(2).] Mol. Cell. Biol. (2004) 24:36–45.
652) Sykiotis GP, Bohmann D [Keap1/Nrf2 Signaling Regulates Oxidative Stress Tolerance and Lifespan in Drosophila]
Dev. Cell (2008) 14(1):76–85.
653) Onken B, Driscoll M [Metformin induces a dietary restriction-like state and the oxidative stress response to extend C.
elegans healthspan via AMPK, LKB1, and SKN-1] PLoS One (2010) 5(1):e8758.
654) Bishop NA, Guarente L [Two neurons mediate diet-restriction-induced longevity in C. elegans.] Nature (2007)
447(7144):545–9.
655) Przybysz AJ, Choe KP, Roberts LJ, Strange K [Increased age reduces DAF-16 and SKN-1 signaling and the hormetic
response of Caenorhabditis elegans to the xenobiotic juglone] Mech. Ageing Dev. (2009) 130(6):357–69.
656) Ungvari Z, Bailey-Downs L, Gautam T, Sosnowska D, Wang M, Monticone RE, Telljohann R, Pinto JT, De Cabo R,
Sonntag WE, Lakatta EG, Csiszar A [Age-associated vascular oxidative stress, Nrf2 dysfunction, and NF-kB
activation in the nonhuman primate macaca mulatta] Journals Gerontol. - Ser. A Biol. Sci. Med. Sci. (2011) 66
A(8):866–75.
657) Frenkel-Denkberg G, Gershon D, Levy AP [The function of hypoxia-inducible factor 1 (HIF-1) is impaired in
senescent mice] FEBS Lett. (1999) 462(3):341–4.
658) Hsieh CC, Rosenblatt JI, Papaconstantinou J [Age-associated changes in SAPK/JNK and p38 MAPK signaling in
response to the generation of ROS by 3-nitropropionic acid] Mech. Ageing Dev. (2003) 124(6):733–46.
659) Liu P, Kemper LJ, Wang J, Zahs KR, Ashe KH, Pasinetti GM [Grape seed polyphenolic extract specifically decreases
Aβ*56 in the brains of Tg2576 mice] J. Alzheimer’s Dis. (2011) 26:657–66.
660) Wang J, Ho L, Zhao Z, Seror I, Humala N, Dickstein DL, Thiyagarajan M, Percival SS, Talcott ST, Pasinetti GM
[Moderate consumption of Cabernet Sauvignon attenuates Abeta neuropathology in a mouse model of Alzheimer’s
disease.] FASEB J. (2006) 20:2313–20.
661) Ho L, Chen LH, Wang J, Zhao W, Talcott ST, Ono K, Teplow D, Humala N, Cheng A, Percival SS, Ferruzzi M, Janle
E, Dickstein DL, Pasinetti GM [Heterogeneity in red wine polyphenolic contents differentially influences Alzheimer’s
disease-type neuropathology and cognitive deterioration.] J. Alzheimers. Dis. (2009) 16(1):59–72.
662) Hartman RE, Shah A, Fagan AM, Schwetye KE, Parsadanian M, Schulman RN, Finn MB, Holtzman DM
[Pomegranate juice decreases amyloid load and improves behavior in a mouse model of Alzheimer’s disease]
Neurobiol. Dis. (2006) 24:506–15.
663) Shukitt-Hale B, Cheng V, Joseph JA [Effects of blackberries on motor and cognitive function in aged rats.] Nutr.
Neurosci. (2009) 12:135–40.
664) Papandreou MA, Dimakopoulou A, Linardaki ZI, Cordopatis P, Klimis-Zacas D, Margarity M, Lamari FN [Effect of a
polyphenol-rich wild blueberry extract on cognitive performance of mice, brain antioxidant markers and
acetylcholinesterase activity] Behav. Brain Res. (2009) 198:352–8.
665) Chan Y-C, Hosoda K, Tsai C-J, Yamamoto S, Wang M-F [Favorable effects of tea on reducing the cognitive deficits
and brain morphological changes in senescence-accelerated mice.] J. Nutr. Sci. Vitaminol. (Tokyo). (2006) 52:266–73.
666) Assunção M, Santos-Marques MJ, Carvalho F, Lukoyanov N V., Andrade JP [Chronic green tea consumption
prevents age-related changes in rat hippocampal formation] Neurobiol. Aging (2011) 32:707–17.
667) Farr SA, Price TO, Dominguez LJ, Motisi A, Saiano F, Niehoff ML, Morley JE, Banks WA, Ercal N, Barbagallo M
[Extra virgin olive oil improves learning and memory in SAMP8 mice] J. Alzheimer’s Dis. (2012) 28:81–92.
668) Krikorian R, Shidler MD, Nash TA, Kalt W, Vinqvist-Tymchuk MR, Shukitt-Hale B, Joseph JA [Blueberry
supplementation improves memory in older adults] J. Agric. Food Chem. (2010) 58:3996–4000.
669) Krikorian R, Nash TA, Shidler MD, Shukitt-Hale B, Joseph JA [Concord grape juice supplementation improves
memory function in older adults with mild cognitive impairment.] Br. J. Nutr. (2010) 103:730–4.
670) Morris MC, Evans DA, Bienias JL, Tangney CC, Bennett DA, Aggarwal N, Wilson RS, Scherr PA [Dietary intake of
antioxidant nutrients and the risk of incident Alzheimer disease in a biracial community study.] JAMA (2002)
287:3230–7.
671) Sung S, Yao Y, Uryu K, Yang H, Lee VMY, Trojanowski JQ, Praticò D [Early vitamin E supplementation in young
but not aged mice reduces Abeta levels and amyloid deposition in a transgenic model of Alzheimer’s disease.] FASEB
J. (2004) 18:323–5.
672) Sano M, Ernesto C, Thomas RG, Klauber MR, Schafer K, Grundman M, Woodbury P, Growdon J, Cotman CW,
Pfeiffer E, Schneider LS, Thal LJ [A controlled trial of selegiline, alpha-tocopherol, or both as treatment for
Alzheimer’s disease. The Alzheimer's Disease Cooperative Study.] N. Engl. J. Med. (1997).
673) Petersen RC, Thomas RG, Grundman M, Bennett D, Doody R, Ferris S, Galasko D, Jin S, Kaye J, Levey A, Pfeiffer E,
Sano M, van Dyck CH, Thal LJ [Vitamin E and donepezil for the treatment of mild cognitive impairment.] N. Engl. J.
Med. (2005).
674) Spurgeon D [Vitamin E doesn’t slow progression to Alzheimer's disease] BMJ (2005) 330(7497):923.
675) Galasko DR, Peskind E, Clark CM, Quinn JF, Ringman JM, Jicha GA, Cotman C, Cottrell B, Montine TJ, Thomas
RG, Aisen P [Antioxidants for Alzheimer disease: a randomized clinical trial with cerebrospinal fluid biomarker
measures.] Arch. Neurol. (2012) 69(7):836–41.
676) Cesarani A, Meloni F, Alpini D, Barozzi S, Verderio L, Boscani PF [Ginkgo biloba (EGb 761) in the treatment of
equilibrium disorders.] Adv. Ther. (1998).
677) Ernst E [The risk-benefit profile of commonly used herbal therapies: Ginkgo, St. John’s wort, ginseng, echinacea, saw
palmetto, and kava] Ann. Intern. Med. (2002) 136:42–53.
678) DeKosky ST, Williamson JD, Fitzpatrick AL, Kronmal RA, Ives DG, Saxton JA, Lopez OL, Burke G, Carlson MC,
Fried LP, Kuller LH, Robbins JA, Tracy RP, Woolard NF, Dunn L, Snitz BE, Nahin RL, Furberg CD [Ginkgo biloba
for prevention of dementia: a randomized controlled trial.] JAMA (2008) 300:2253–62.
679) Hamazaki K, Hamazaki T, Inadera H [Abnormalities in the fatty acid composition of the postmortem entorhinal
cortex of patients with schizophrenia, bipolar disorder, and major depressive disorder] Psychiatry Res. (2013)
210:346–50.
XX
680) Bazan NG, Molina MF, Gordon WC [Docosahexaenoic Acid signalolipidomics in nutrition: significance in aging,
neuroinflammation, macular degeneration, Alzheimer’s, and other neurodegenerative diseases.] Annu. Rev. Nutr.
(2011) 31:321–51.
681) 橋本道男 [ω3 系脂肪酸と認知機能 (特集 生活習慣病と認知機能 : 認知症の予防を見据えた生活習慣病の治療を目指して)
– (基礎知見)] 日本臨床. 日本臨床社 (2014) 72(4):648–56.
682) Hashimoto M, Inoue T, Katakura M, Tanabe Y, Hossain S, Tsuchikura S, Shido O [Prescription n-3 fatty acids, but
not eicosapentaenoic acid alone, improve reference memory-related learning ability by increasing brain-derived
neurotrophic factor levels in SHR.Cg-Lepr(cp)/NDmcr rats, a metabolic syndrome model.] Neurochem. Res. (2013)
38(10):2124–35.
683) 橋本道男 [多価不飽和脂肪酸と疾患 2 中枢神経系] 治療学 (2009) 43(8):838–44.
684) Hashimoto M, Hossain S [Neuroprotective and ameliorative actions of polyunsaturated fatty acids against neuronal
diseases: beneficial effect of docosahexaenoic acid on cognitive decline in Alzheimer’s disease.] J. Pharmacol. Sci.
(2011) 116(2):150–62.
685) 有田誠, 磯部洋輔 [ω3 系脂肪酸由来の抗炎症性代謝物の構造と機能] 生化學. 日本生化学会 (2008) 80(11):1042–6.
686) 橋本道男 [食生活による認知症と生活習慣病の予防 (特集 生活習慣病と認知症 : 負の連鎖を断ち切るために)] Med. = メ
ディシーナル 生活習慣病の薬物療法・治療薬を中心とする臨床医学誌. 医学出版 (2012) 2(9):101–11.
687) Hashimoto M, Yamashita K, Kato S [Beneficial effects of daily dietary omega-3 polyunsaturated fatty acid
supplementation on age-related cognitive decline in elderly Japanese with very mild] J Aging Res Clin … (2012)
1:193–201.
688) Han Y-S, Zheng W-H, Bastianetto S, Chabot J-G, Quirion R [Neuroprotective effects of resveratrol against betaamyloid-induced neurotoxicity in rat hippocampal neurons: involvement of protein kinase C.] Br. J. Pharmacol. (2004)
141:997–1005.
689) Kim D, Nguyen MD, Dobbin MM, Fischer A, Sananbenesi F, Rodgers JT, Delalle I, Baur JA, Sui G, Armour SM,
Puigserver P, Sinclair DA, Tsai L-H [SIRT1 deacetylase protects against neurodegeneration in models for Alzheimer’s
disease and amyotrophic lateral sclerosis.] EMBO J. (2007) 26:3169–79.
690) Witte A V, Fobker M, Gellner R, Knecht S, Flöel A [Caloric restriction improves memory in elderly humans.] Proc.
Natl. Acad. Sci. U. S. A. (2009).
691) Bäckesjö C-M, Li Y, Lindgren U, Haldosén L-A [Activation of Sirt1 decreases adipocyte formation during osteoblast
differentiation of mesenchymal stem cells.] Cells. Tissues. Organs (2009) 189:93–7.
692) Lin SJ, Defossez PA, Guarente L [Requirement of NAD and SIR2 for life-span extension by calorie restriction in
Saccharomyces cerevisiae.] Science (2000) 289:2126–8.
693) Rogina B, Helfand SL [Sir2 mediates longevity in the fly through a pathway related to calorie restriction.] Proc. Natl.
Acad. Sci. U. S. A. (2004) 101:15998–6003.
694) Haigis MC, Yankner BA [The aging stress response.] Mol. Cell (2010) 40:333–44.
695) Burnett C, Valentini S, Cabreiro F, Goss M, Somogyvári M, Piper MD, Hoddinott M, Sutphin GL, Leko V, McElwee
JJ, Vazquez-Manrique RP, Orfila A-M, Ackerman D, Au C, Vinti G, Riesen M, Howard K, Neri C, Bedalov A,
Kaeberlein M, Soti C, Partridge L, Gems D [Absence of effects of Sir2 overexpression on lifespan in C. elegans and
Drosophila.] Nature (2011) 477(7365):482–5.
696) Massudi H, Grant R, Braidy N, Guest J, Farnsworth B, Guillemin GJ [Age-associated changes in oxidative stress and
NAD+ metabolism in human tissue] PLoS One (2012) 7(7):e42357.
697) Boily G, Seifert EL, Bevilacqua L, He XH, Sabourin G, Estey C, Moffat C, Crawford S, Saliba S, Jardine K, Xuan J,
Evans M, Harper M-E, McBurney MW [SirT1 regulates energy metabolism and response to caloric restriction in
mice.] PLoS One (2008) 3(3):e1759.
698) Seifert EL, Caron AZ, Morin K, Coulombe J, Hong He X, Jardine K, Dewar-Darch D, Boekelheide K, Harper M-E,
McBurney MW [SirT1 catalytic activity is required for male fertility and metabolic homeostasis in mice] FASEB J.
(2012), 555–66.
699) Tissenbaum HA, Guarente L [Increased dosage of a sir-2 gene extends lifespan in Caenorhabditis elegans.] Nature
(2001) 410:227–30.
700) Garsin DA, Villanueva JM, Begun J, Kim DH, Sifri CD, Calderwood SB, Ruvkun G, Ausubel FM [Long-lived C.
elegans daf-2 mutants are resistant to bacterial pathogens.] Science (2003) 300:1921.
701) Brunet A, Sweeney LB, Sturgill JF, Chua KF, Greer PL, Lin Y, Tran H, Ross SE, Mostoslavsky R, Cohen HY, Hu LS,
Cheng H-L, Jedrychowski MP, Gygi SP, Sinclair DA, Alt FW, Greenberg ME [Stress-dependent regulation of FOXO
transcription factors by the SIRT1 deacetylase.] Science (2004) 303:2011–5.
702) Rodgers JT, Lerin C, Haas W, Gygi SP, Spiegelman BM, Puigserver P [Nutrient control of glucose homeostasis
through a complex of PGC-1alpha and SIRT1.] Nature (2005) 434(7029):113–8.
703) Moynihan KA, Grimm AA, Plueger MM, Bernal-Mizrachi E, Ford E, Cras-Méneur C, Permutt MA, Imai SI
[Increased dosage of mammalian Sir2 in pancreatic β cells enhances glucose-stimulated insulin secretion in mice] Cell
Metab. (2005) 2:105–17.
704) Rodgers JT, Puigserver P [Fasting-dependent glucose and lipid metabolic response through hepatic sirtuin 1.] Proc.
Natl. Acad. Sci. U. S. A. (2007) 104:12861–6.
705) Liu Y, Dentin R, Chen D, Hedrick S, Ravnskjaer K, Schenk S, Milne J, Meyers DJ, Cole P, Yates J, Olefsky J,
Guarente L, Montminy M [A fasting inducible switch modulates gluconeogenesis via activator/coactivator exchange.]
Nature (2008) 456:269–73.
706) Milne JC, Lambert PD, Schenk S, Carney DP, Smith JJ, Gagne DJ, Jin L, Boss O, Perni RB, Vu CB, Bemis JE, Xie
R, Disch JS, Ng PY, Nunes JJ, Lynch A V, Yang H, Galonek H, Israelian K, Choy W, Iffland A, Lavu S, Medvedik O,
Sinclair DA, Olefsky JM, Jirousek MR, Elliott PJ, Westphal CH [Small molecule activators of SIRT1 as therapeutics
for the treatment of type 2 diabetes.] Nature (2007) 450:712–6.
707) Pfluger PT, Herranz D, Velasco-Miguel S, Serrano M, Tschöp MH [Sirt1 protects against high-fat diet-induced
metabolic damage.] Proc. Natl. Acad. Sci. U. S. A. (2008) 105:9793–8.
708) Yang Y, Fu W, Chen J, Olashaw N, Zhang X, Nicosia S V, Bhalla K, Bai W [SIRT1 sumoylation regulates its
deacetylase activity and cellular response to genotoxic stress.] Nat. Cell Biol. (2007) 9:1253–62.
709) Julien C, Tremblay C, Emond V, Lebbadi M, Salem N, Bennett DA, Calon F [Sirtuin 1 reduction parallels the
accumulation of tau in Alzheimer disease.] J. Neuropathol. Exp. Neurol. (2009) 68:48–58.
XXI
710) Zhang L, Fiala M, Cashman J, Sayre J, Espinosa A, Mahanian M, Zaghi J, Badmaev V, Graves MC, Bernard G,
Rosenthal M [Curcuminoids enhance amyloid-beta uptake by macrophages of Alzheimer’s disease patients.] J.
Alzheimers. Dis. (2006) 10:1–7.
711) Cashman JR, Ghirmai S, Abel KJ, Fiala M [Immune defects in Alzheimer’s disease: new medications development.]
BMC Neurosci. (2008) 9 Suppl 2:S13.
712) Lim GP, Chu T, Yang F, Beech W, Frautschy SA, Cole GM [The curry spice curcumin reduces oxidative damage and
amyloid pathology in an Alzheimer transgenic mouse.] J. Neurosci. (2001) 21(21):8370–7.
713) Yang F, Lim GP, Begum AN, Ubeda OJ, Simmons MR, Ambegaokar SS, Chen P, Kayed R, Glabe CG, Frautschy SA,
Cole GM [Curcumin inhibits formation of amyloid β oligomers and fibrils, binds plaques, and reduces amyloid in vivo]
J. Biol. Chem. (2005) 280:5892–901.
714) Chandra V, Pandav R, Dodge HH, Johnston JM, Belle SH, DeKosky ST, Ganguli M [Incidence of Alzheimer’s disease
in a rural community in India: the Indo-US study.] Neurology (2001) 57(6):985–9.
715) Shoba G, Joy D, Joseph T, Majeed M, Rajendran R, Srinivas PSSR [Influence of piperine on the pharmacokinetics of
curcumin in animals and human volunteers] Planta Med. (1998) 64:353–6.
716) Jurenka JS [Anti-inflammatory properties of curcumin, a major constituent of Curcuma longa: a review of preclinical
and clinical research.] Altern. Med. Rev. (2009) 14(2):141–53.
717) Kawanishi S, Oikawa S, Murata M [Evaluation for safety of antioxidant chemopreventive agents.] Antioxid. Redox
Signal. 7(11-12):1728–39.
718) Kozikowski AP, Tückmantel W [Chemistry, pharmacology, and clinical efficacy of the Chinese nootropic agent
huperzine A] Acc. Chem. Res. (1999), 641–50.
719) Wang BS, Wang H, Wei ZH, Song YY, Zhang L, Chen HZ [Efficacy and safety of natural acetylcholinesterase
inhibitor huperzine A in the treatment of Alzheimer’s disease: an updated meta-analysis] J. Neural Transm. (2009)
116:457–65.
720) Tang XC, He XC, Bai DL [Huperzine A: A novel acetylcholinesterase inhibitor] Drugs Future (1999), 647–63.
721) Cheng DH, Ren H, Tang XC [Huperzine A, a novel promising acetylcholinesterase inhibitor.] Neuroreport (1996)
8:97–101.
722) Yoshinaga K, Ishizuka J, Evers BM, Townsend CM, Thompson JC [Age-related changes in polyamine biosynthesis
after fasting and refeeding.] Exp. Gerontol. 28(6):565–72.
723) Soda K, Kano Y, Sakuragi M, Takao K, Lefor A, Konishi F [Long-term oral polyamine intake increases blood
polyamine concentrations.] J. Nutr. Sci. Vitaminol. (Tokyo). (2009) 55(4):361–6.
724) 姉帯正樹, 柴田敏郎, 畠山好雄 [北海道産当帰の調製法と化学的品質評価(第 1 報) : 調製条件とショ糖および希エタノール
エキス含量の変動] 生薬學雜誌. 日本生薬学会 (1997) 51(4):331–4.
725) Laursen SE, Belknap JK [Intracerebroventricular injections in mice. Some methodological refinements.] J.
Pharmacol. Methods (1986) 16(4):355–7.
726) Yan JJ, Kim DH, Moon YS, Jung JS, Ahn EM, Baek NI, Song DK [Protection against beta-amyloid peptide-induced
memory impairment with long-term administration of extract of Angelica gigas or decursinol in mice] Prog. NeuroPsychopharmacology Biol. Psychiatry (2004) 28:25–30.
727) Kitagawa K, Matsumoto M, Tagaya M, Hata R, Ueda H, Niinobe M, Handa N, Fukunaga R, Kimura K, Mikoshiba K
[“Ischemic tolerance” phenomenon found in the brain.] Brain Res. (1990) 528:21–4.
728) Chopp M, Chen H, Ho KL, Dereski MO, Brown E, Hetzel FW, Welch KM [Transient hyperthermia protects against
subsequent forebrain ischemic cell damage in the rat.] Neurology (1989) 39:1396–8.
729) Tasaki K, Ruetzler CA, Ohtsuki T, Martin D, Nawashiro H, Hallenbeck JM [Lipopolysaccharide pre-treatment
induces resistance against subsequent focal cerebral ischemic damage in spontaneously hypertensive rats] Brain Res.
(1997) 748:267–70.
730) Kato K, Hasegawa K, Goto S, Inaguma Y [Dissociation as a result of phosphorylation of an aggregated form of the
small stress protein, hsp27.] J. Biol. Chem. (1994) 269:11274–8.
731) Matsushima K, Schmidt-Kastner R, Hogan MJ, Hakim AM [Cortical spreading depression activates trophic factor
expression in neurons and astrocytes and protects against subsequent focal brain ischemia] Brain Res. (1998) 807(12):47–60.
732) Ma Z-C, Hong Q, Wang Y-G, Tan H-L, Xiao C-R, Liang Q-D, Zhang B-L, Gao Y [Ferulic acid protects human
umbilical vein endothelial cells from radiation induced oxidative stress by phosphatidylinositol 3-kinase and
extracellular signal-regulated kinase pathways.] Biol. Pharm. Bull. (2010) 33:29–34.
733) Shahidi F, Wanasundara PK [Phenolic antioxidants.] Crit. Rev. Food Sci. Nutr. Taylor & Francis Group (1992)
32(1):67–103.
734) 加藤陽治 [植物細胞壁多糖の微細構造解析と機能に関する研究] J. Appl. Glycosci. 日本応用糖質科学会 (2008) 55(1):35–
43.
735) Ralph J, Garcia Conesa MT, Williamson G [Simple Preparation of 8−5-Coupled Diferulate] J. Agric. Food Chem.
(1998) 46:2531–2.
736) Grabber JH, Hatfield RD, Ralph J [Diferulate cross-links impede the enzymatic degradation of non-lignified maize
walls] J. Sci. Food Agric. (1998) 77:193–200.
737) MacAdam JW, Grabber JH [Relationship of growth cessation with the formation of diferulate cross-links and pcoumaroylated lignins in tall fescue leaf blades] Planta (2002) 215:785–93.
738) Manach C, Scalbert A, Morand C, Rémésy C, Jiménez L [Polyphenols: Food sources and bioavailability] Am. J. Clin.
Nutr. (2004), 727–47.
739) Terry W [The Wild Rose Scientific Herbal] (1991).
740) Sellappan S, Akoh CC, Krewer G [Phenolic compounds and antioxidant capacity of Georgia-grown blueberries and
blackberries] J. Agric. Food Chem. (2002) 50:2432–8.
741) Kato H, Ohta T, Tsugita T, Hosaka Y [Effect of parboiling on texture and flavor components of cooked rice] J. Agric.
Food Chem. (1983) 31(4):818–23.
742) Herrmann K [Occurrence and content of hydroxycinnamic and hydroxybenzoic acid compounds in foods.] Crit. Rev.
Food Sci. Nutr. (1989) 28:315–47.
743) 谷口久次 [米ぬかを原料とするフェルラ酸の製造技術] JST ニュース (2003) 83:9.
744) Hatcher DW, Kruger JE [Simple phenolic acids in flours prepared from Canadian wheat: Relationship to ash content,
color, and polyphenol oxidase activity] Cereal Chem. (1997) 74(3):337–43.
XXII
745) Adam A, Crespy V, Levrat-Verny M-A, Leenhardt F, Leuillet M, Demigné C, Rémésy C [The bioavailability of ferulic
acid is governed primarily by the food matrix rather than its metabolism in intestine and liver in rats.] J. Nutr. (2002)
132:1962–8.
746) NISHIZAWA C, OHTA T, EGASHIRA Y, SANADA H [Ferulic Acid Contents in Typical Cereals.] Nippon
SHOKUHIN KAGAKU KOGAKU KAISHI. 社団法人 日本食品科学工学会 (1998), 499–503.
747) Norton RA [Quantitation of steryl ferulate and p-coumarate esters from corn and rice] Lipids (1995), 269–74.
748) Akihisa T, Yasukawa K, Yamaura M, Ukiya M, Kimura Y, Shimizu N, Arai K [Triterpene alcohol and sterol ferulates
from rice bran and their anti- inflammatory effects] J. Agric. Food Chem. (2000) 48:2313–9.
749) Pussayanawin V, Wetzel DL, Fulcher RG [Fluorescence Detection and Measurement of Ferulic Acid in Wheat Milling
Fractions by Microscopy and HPLC] J. Agric. Food Chem. American Chemical Society (1988) 36(3):515–20.
750) Hakala P, Lampi AM, Ollilainen V, Werner U, Murkovic M, Wähälä K, Karkola S, Piironen V [Steryl phenolic acid
esters in cereals and their milling fractions] J. Agric. Food Chem. (2002) 50:5300–7.
751) Troszyńska A, Ciska E [Phenolic compounds of seed coats of white and coloured varieties of pea (Pisum sativum L.)
and their total antioxidant activity] Czech J. Food Sci. (2002) 20:15–22.
752) Colpas FT, Ono EO, Rodrigues JD, Passos JRDS [Effects of some phenolic compounds on soybean seed germination
and on seed-borne fungi] Brazilian Arch. Biol. Technol. (2003) 46:155–61.
753) Yang C-M, Chang I-F, Lin S-J, Chou C-H [Effects of Three Allelopathic Phenolics on Chlorophyll Accumulation of
Rice (Oryza Sativa) Seedlings: II. Stimulation of Consumption-Orientation] Bot. Bull. Acad. Sin. 中央研究院植物研究
所 (2004) 45(2):119–25.
754) Emmons CL, Peterson DM [Antioxidant activity and phenolic contents of oat groats and hulls] Cereal Chem. The
American Association of Cereal Chemists, Inc. (1999) 76(6):902–6.
755) Eun HJ, Sung RK, In KH, Tae YH [Hypoglycemic effects of a phenolic acid fraction of rice bran and ferulic acid in
C57BL/KsJ-db/db mice] J. Agric. Food Chem. (2007) 55:9800–4.
756) Fujita A, Sasaki H, Doi A, Okamoto K, Matsuno S, Furuta H, Nishi M, Nakao T, Tsuno T, Taniguchi H, Nanjo K
[Ferulic acid prevents pathological and functional abnormalities of the kidney in Otsuka Long-Evans Tokushima
Fatty diabetic rats] Diabetes Res. Clin. Pract. (2008) 79:11–7.
757) Hudson EA, Dinh PA, Kokubun T, Simmonds MS, Gescher A [Characterization of potentially chemopreventive
phenols in extracts of brown rice that inhibit the growth of human breast and colon cancer cells.] Cancer Epidemiol.
Biomarkers Prev. (2000) 9(11):1163–70.
758) Kampa M, Alexaki V-I, Notas G, Nifli A-P, Nistikaki A, Hatzoglou A, Bakogeorgou E, Kouimtzoglou E, Blekas G,
Boskou D, Gravanis A, Castanas E [Antiproliferative and apoptotic effects of selective phenolic acids on T47D human
breast cancer cells: potential mechanisms of action.] Breast Cancer Res. (2004) 6:R63–74.
759) Ou L, Kong L-Y, Zhang X-M, Niwa M [Oxidation of ferulic acid by Momordica charantia peroxidase and related antiinflammation activity changes.] Biol. Pharm. Bull. (2003) 26:1511–6.
760) Zhu H, Liang Q-H, Xiong X-G, Chen J, Wu D, Wang Y, Yang B, Zhang Y, Zhang Y, Huang X [Anti-Inflammatory
Effects of the Bioactive Compound Ferulic Acid Contained in Oldenlandia diffusa on Collagen-Induced Arthritis in
Rats.] Evid. Based. Complement. Alternat. Med. (2014) 2014:573801.
761) Castelluccio C, Bolwell GP, Gerrish C, Rice-Evans C [Differential distribution of ferulic acid to the major plasma
constituents in relation to its potential as an antioxidant.] Biochem. J. (1996) 316 ( Pt 2:691–4.
762) Uchida M, Nakajin S, Toyoshima S, Shinoda M [Antioxidative effect of sesamol and related compounds on lipid
peroxidation.] Biol. Pharm. Bull. (1996) 19:623–6.
763) Wang H, Peng RX [Sodium ferulate alleviated paracetamol-induced liver toxicity in mice] Zhongguo Yao Li Xue Bao
(1994) 15:81–3.
764) Gao SW, Chen ZJ [[Effects of sodium ferulate on platelet aggregation and platelet thromboxane A2 in patients with
coronary heart disease].] Zhong Xi Yi Jie He Za Zhi (1988) 8(5):263–5, 259.
765) Wang Q, Xiong LZ, Chen SY [[Effect of sodium ferulate on activation of extracellular signal regulated kinase after
cerebral ischemia/reperfusion injury in rats]] Zhongguo Zhong Xi Yi Jie He Za Zhi (2003) 23:918–21.
766) Li G, Wang J, Wang J, Liu Y, Jin Y [Effect of ferulic acid on the proliferation of nerve cells of retinas in vitro]
Zhonghua. Yan Ke Za Zhi. (2003) 39:650–4.
767) Okamoto R, Sakamoto S, Noguchi K [Effects of ferulic acid on FSH, LH and prolactin levels in serum and pituitary
tissue of male rats (author’s transl)] Nihon Naibunpi Gakkai Zasshi (1976) 52(9):953–8.
768) Gorewit RC [Pituitary and thyroid hormone responses of heifers after ferulic acid administration.] J. Dairy Sci.
(1983) 66:624–9.
769) Suzuki A, Yamamoto M, Jokura H, Fujii A, Tokimitsu I, Hase T, Saito I [Ferulic Acid Restores EndotheliumDependent Vasodilation in Aortas of Spontaneously Hypertensive Rats] Am. J. Hypertens. (2007) 20(5):508–13.
770) Ardiansyah, Ohsaki Y, Shirakawa H, Koseki T, Komai M [Novel effects of a single administration of ferulic acid on
the regulation of blood pressure and the hepatic lipid metabolic profile in stroke-prone spontaneously hypertensive
rats] J. Agric. Food Chem. (2008) 56:2825–30.
771) Zhao Z, Egashira Y, Sanada H [Digestion and absorption of ferulic acid sugar esters in rat gastrointestinal tract] J.
Agric. Food Chem. (2003) 51:5534–9.
772) Zhao Z, Egashira Y, Sanada H [Ferulic acid sugar esters are recovered in rat plasma and urine mainly as the
sulfoglucuronide of ferulic acid.] J. Nutr. (2003) 133:1355–61.
773) Zhao Z, Egashira Y, Sanada H [Ferulic acid is quickly absorbed from rat stomach as the free form and then
conjugated mainly in liver.] J. Nutr. (2004) 134:3083–8.
774) Poole RC, Halestrap AP [Transport of lactate and other monocarboxylates across mammalian plasma membranes.]
Am. J. Physiol. (1993) 264:C761–82.
775) Konishi Y, Hagiwara K, Shimizu M [Transepithelial transport of fluorescein in Caco-2 cell monolayers and use of
such transport in in vitro evaluation of phenolic acid availability.] Biosci. Biotechnol. Biochem. (2002) 66(11):2449–57.
776) BOOTH AN, MASRI MS, ROBBINS DJ, EMERSON OH, JONES FT, DEEDS F [Urinary metabolites of coumarin
and omicron-coumaric acid.] J. Biol. Chem. (1959) 234(4):946–8.
777) Rondini L, Peyrat-Maillard MN, Marsset-Baglieri A, Berset C [Sulfated ferulic acid is the main in vivo metabolite
found after short-term ingestion of free ferulic acid in rats] J. Agric. Food Chem. (2002) 50(10):3037–41.
778) Bourne LC, Rice-Evans C [Bioavailability of ferulic acid.] Biochem. Biophys. Res. Commun. (1998) 253(2):222–7.
XXIII
779) Henriksen A, Smith AT, Gajhede M [The structures of the horseradish peroxidase C-ferulic acid complex and the
ternary complex with cyanide suggest how peroxidases oxidize small phenolic substrates.] J. Biol. Chem. (1999)
274:35005–11.
780) Henriksen A, Mirza O, Indiani C, Teilum K, Smulevich G, Welinder KG, Gajhede M [Structure of soybean seed coat
peroxidase: a plant peroxidase with unusual stability and haem-apoprotein interactions.] Protein Sci. (2001) 10:108–
15.
781) Garcia R, Rakotozafy L, Telef N, Potus J, Nicolas J [Oxidation of ferulic acid or arabinose-esterified ferulic acid by
wheat germ peroxidase] J. Agric. Food Chem. (2002) 50:3290–8.
782) Kanski J, Aksenova M, Stoyanova A, Butterfield DA [Ferulic acid antioxidant protection against hydroxyl and
peroxyl radical oxidation in synaptosomal and neuronal cell culture systems in vitro: Structure-activity studies] J.
Nutr. Biochem. (2002) 13:273–81.
783) Ogiwara T, Satoh K, Kadoma Y, Murakami Y, Unten S, Atsumi T, Sakagami H, Fujisawa S [Radical scavenging
activity and cytotoxicity of ferulic acid.] Anticancer Res. (2002) 22(5):2711–7.
784) Prasad NR, Srinivasan M, Pugalendi K V, Menon VP [Protective effect of ferulic acid on gamma-radiation-induced
micronuclei, dicentric aberration and lipid peroxidation in human lymphocytes.] Mutat. Res. (2006) 603:129–34.
785) Usami E, Kusano G, Katayose T, Wachi H, Seyama Y [Assessment of antioxidant activity of natural compound by
water- and lipid-soluble antioxidant factor] Yakugaku Zasshi (2004) 124(11):847–50.
786) Trombino S, Cassano R, Bloise E, Muzzalupo R, Leta S, Puoci F, Picci N [Design and synthesis of cellulose
derivatives with antioxidant activity] Macromol. Biosci. (2008) 8:86–95.
787) Dringen R, Gutterer JM, Hirrlinger J [Glutathione metabolism in brain metabolic interaction between astrocytes and
neurons in the defense against reactive oxygen species.] Eur. J. Biochem. (2000) 267:4912–6.
788) Balasubashini MS, Rukkumani R, Viswanathan P, Menon VP [Ferulic Acid Alleviates Lipid Peroxidation in Diabetic
Rats] Phyther. Res. (2004) 18:310–4.
789) Murphy TH, De Long MJ, Coyle JT [Enhanced NAD(P)H:quinone reductase activity prevents glutamate toxicity
produced by oxidative stress.] J. Neurochem. (1991) 56:990–5.
790) Murphy TH, Yu J, Ng R, Johnson DA, Shen H, Honey CR, Johnson JA [Preferential expression of antioxidant
response element mediated gene expression in astrocytes] J. Neurochem. (2001) 76(6):1670–8.
791) Moehlenkamp JD, Johnson JA [Activation of antioxidant/electrophile-responsive elements in IMR-32 human
neuroblastoma cells.] Arch. Biochem. Biophys. (1999) 363:98–106.
792) Koh PO [Ferulic acid prevents the cerebral ischemic injury-induced decrease of Akt and Bad phosphorylation]
Neurosci. Lett. (2012) 507(2):156–60.
793) Sung J-H, Cho E-H, Cho J-H, Won C-K, Kim M-O, Koh P-O [Identification of proteins regulated by ferulic acid in a
middle cerebral artery occlusion animal model-a proteomics approach.] J. Vet. Med. Sci. (2012) 74(11):1401–7.
794) Bian Z, Furuya N, Zheng D-M, Oliva Trejo JA, Tada N, Ezaki J, Ueno T [Ferulic Acid induces Mammalian target of
rapamycin inactivation in cultured Mammalian cells.] Biol. Pharm. Bull. (2013) 36:120–4.
795) Talavéra S, Felgines C, Texier O, Besson C, Lamaison J-L, Rémésy C [Anthocyanins are efficiently absorbed from the
stomach in anesthetized rats.] J. Nutr. (2003) 133:4178–82.
796) Passamonti S, Vrhovsek U, Vanzo A, Mattivi F [The stomach as a site for anthocyanins absorption from food] FEBS
Lett. (2003) 544:210–3.
797) Jin Y, Yan E, Fan Y, Zong Z, Qi Z, Li Z [Sodium ferulate prevents amyloid-beta-induced neurotoxicity through
suppression of p38 MAPK and upregulation of ERK-1/2 and Akt/protein kinase B in rat hippocampus.] Acta
Pharmacol. Sin. (2005) 26:943–51.
798) Jin Y, Fan Y, Yan EZ, Liu Z, Zong ZH, Qi ZM [Effects of sodium ferulate on amyloid-beta-induced MKK3/MKK6-p38
MAPK-Hsp27 signal pathway and apoptosis in rat hippocampus] Acta Pharmacol. Sin. (2006) 27:1309–16.
799) Pappolla MA, Omar RA, Kim KS, Robakis NK [Immunohistochemical evidence of oxidative [corrected] stress in
Alzheimer’s disease.] Am. J. Pathol. (1992) 140:621–8.
800) Behl C, Davis JB, Lesley R, Schubert D [Hydrogen peroxide mediates amyloid β protein toxicity] Cell (1994)
77(6):817–27.
801) Goodman Y, Steiner MR, Steiner SM, Mattson MP [Nordihydroguaiaretic acid protects hippocampal neurons against
amyloid beta-peptide toxicity, and attenuates free radical and calcium accumulation.] Brain Res. (1994) 654:171–6.
802) Zhang Z, Rydel RE, Drzewiecki GJ, Fuson K, Wright S, Wogulis M, Audia JE, May PC, Hyslop PA [Amyloid betamediated oxidative and metabolic stress in rat cortical neurons: no direct evidence for a role for H2O2 generation.] J.
Neurochem. (1996) 67:1595–606.
803) Yan J-J, Jung J-S, Kim T-K, Hasan A, Hong C-W, Nam J-S, Song D-K [Protective effects of ferulic acid in amyloid
precursor protein plus presenilin-1 transgenic mouse model of Alzheimer disease.] Biol. Pharm. Bull. (2013)
36(1):140–3.
804) Kang SY, Kim YC [Neuroprotective coumarins from the root of Angelica gigas: structure-activity relationships.] Arch.
Pharm. Res. (2007) 30:1368–73.
805) Kim JH, Koh SK, Koh HJ, Kwon YA, Kim SH, Kim JG, Kim TE, Park JW, Seo MY, Song YR, Lee I, Kim DK [A Three
Month Placebo-Controlled Clinical Trial of INM 176 in the Old Aged Subjects with Memory Impairment] J. Korean
Neuropsychiatr. Assoc. (2003) 42(2):254–62.
806) 姉帯正樹, 増田隆広, 高杉光雄 [当帰の調製法と化学的品質評価(第 6 報)フロクマリンの指標物質としての可能性につい
て] 北海道立衛生研究所報. 北海道立衛生研究所 (2002) (52):19–23.
807) Wahlin B [Nordiska medicinalväxter : med uppgifter om deras ursprung, utbredning i Norden ...] Alnarp: Nordiska
genbanken (1994).
808) 中村重信, 佐々木健, 阿瀬川.孝治, 伊丹昭, 伊藤達彦, 清原龍夫, 河野和彦, 松田桜子, 水野裕, 宮原覚, 折笠秀樹, 遠藤英
俊 [Ferulic acid と garden angelica 根抽出物製剤 ANM176TM がアルツハイマー病患者の認知機能に及ぼす影響]
Geriat. Med (2008) 46:1511–9.
809) Smith Doody R [Update on Alzheimer drugs (donepezil).] Neurologist (2003) 9(5):225–9.
810) Caro JJ, Getsios D, Migliaccio-Walle K, Raggio G, Ward A [Assessment of health economics in Alzheimer’s disease
(AHEAD) based on need for full-time care.] Neurology (2001) 57:964–71.
811) Rogers SL, Friedhoff LT [Long-term efficacy and safety of donepezil in the treatment of Alzheimer’s disease: an
interim analysis of the results of a US multicentre open label extension study.] Eur. Neuropsychopharmacol. (1998).
812) 稲永和豊 [老年期認知症障害の当帰芍薬散による治療効果] Prog. Med. (1996) 16:293–300.
XXIV
813) 萩野信義 [脳に対する当帰芍薬散 (TJ-23) の作用様式-特にアルツハイマー病について-] 精神神経薬理 (1990) 12:229–
34.
814) 小山嵩夫, 荻野信義 [アルツハイマー型痴呆、脳内ニコチン・アセチルコリン・レセプターに対する当帰芍薬散の作用]
JAMA (1998) 4:22−3.
815) Venugopala KN, Rashmi V, Odhav B [Review on natural coumarin lead compounds for their pharmacological
activity] Biomed Res. Int. (2013).
816) Iranshahi M, Askari M, Sahebkar A, Adjipavlou-Litina D [Evaluation of antioxidant, anti-inflammatory and
lipoxygenase inhibitory activities of the prenylated coumarin umbelliprenin] DARU J. Pharm. Sci. (2009), 99–103.
817) 多田幸恵, 池田虎雄, 高橋博 [天然添加物フェルラ酸の F344 ラットによる亜慢性毒性試験] 東京都立衛生研究所研究年
報. 東京都立衛生研究所 (2001) (52):272–8.
818) James A. D [Handbook of medicinal herbs] Boca Raton: CRC Press Inc, USA (1987).
819) Barraja P, Spanò V, Patrizia D, Carbone A, Cirrincione G, Vedaldi D, Salvador A, Viola G, Dall’acqua F [Pyrano[2,3e]isoindol-2-ones, new angelicin heteroanalogues.] Bioorg. Med. Chem. Lett. (2009) 19(6):1711–4.
820) Tinel M, Belghiti J, Descatoire V, Amouyal G, Letteron P, Geneve J, Larrey D, Pessayre D [Inactivation of human
liver cytochrome P-450 by the drug methoxsalen and other psoralen derivatives.] Biochem. Pharmacol. (1987)
36(6):951–5.
821) Szwajgier D, Borowiec K [Phenolic acids from malt are efficient acetylcholinesterase and butyrylcholinesterase
inhibitors] J. Inst. Brew. (2012) 118(1):40–8.
822) 秋吉祐樹, 江頭伸昭, 岩崎克典, 新井隆史, 高垣祐紀, 三島健一, 藤原道弘 [β-アミロイド脳室内投与によるアルツハイマ
ー病モデルを用いた当帰芍薬散の薬効評価] 日本薬理学雑誌 (2004) 123(3):63.
823) Ahn J, Um M, Choi W, Kim S, Ha T [Protective effects of Glycyrrhiza uralensis Fisch. on the cognitive deficits caused
by β-amyloid peptide 25-35 in young mice] Biogerontology (2006) 7(4):239–47.
824) Cole GM, Frautschy SA [The role of insulin and neurotrophic factor signaling in brain aging and Alzheimer’s
Disease] Exp. Gerontol. (2007) 42:10–21.
825) Yamamoto T, Yoh K, Kobayashi A, Ishii Y, Kure S, Koyama A, Sakamoto T, Sekizawa K, Motohashi H, Yamamoto M
[Identification of polymorphisms in the promoter region of the human NRF2 gene] Biochem. Biophys. Res. Commun.
(2004) 321(1):72–9.
826) Twickler MTB, Cramer M-JM, Koppeschaar HPF [Unraveling Reaven’s syndrome X: serum insulin-like growth
factor-I and cardiovascular disease.] Circulation (2003) 107(20):e190–2; author reply e190–2.
827) Magee JC, Johnston D [A synaptically controlled, associative signal for Hebbian plasticity in hippocampal neurons.]
Science (1997) 275:209–13.
828) Lledo PM, Hjelmstad GO, Mukherji S, Soderling TR, Malenka RC, Nicoll RA [Calcium/calmodulin-dependent kinase
II and long-term potentiation enhance synaptic transmission by the same mechanism.] Proc. Natl. Acad. Sci. U. S. A.
(1995) 92:11175–9.
829) Barria A, Muller D, Derkach V, Griffith LC, Soderling TR [Regulatory phosphorylation of AMPA-type glutamate
receptors by CaM-KII during long-term potentiation.] Science (1997) 276:2042–5.
830) 工藤卓, 林勲, 田口隆久 [神経細胞とシナプス可塑性(<特集>脳と知覚)] 知能と情報 日本知能情報ファジィ学会誌 J.
Japan Soc. Fuzzy Theory Intell. Informatics. 日本知能情報ファジィ学会 (2006) 18(3):362–8.
831) Buchwalter G, Gross C, Wasylyk B [Ets ternary complex transcription factors] Gene (2004), 1–14.
832) Olson EN, Nordheim A [Linking actin dynamics and gene transcription to drive cellular motile functions.] Nat. Rev.
Mol. Cell Biol. (2010) 11:353–65.
833) Miralles F, Posern G, Zaromytidou AI, Treisman R [Actin dynamics control SRF activity by regulation of its
coactivator MAL] Cell (2003) 113(3):329–42.
XXV
(索引)
ページ
アクチン ----------------------------------------------------------------------------------- 32,39,40,51,58,65
アストロサイト -------------------------------------------------------- 7,9,29,33,47,58,66,88,89,94,95
アセチルコリン -------------------------------------------------------------------------------- 35,75,88,107
アポトーシス -------------------------------------- 13,16,25,26,34,35,57,58,60,66,71,74,94,95,108
アミリン ------------------------------------------------------------------------------- 53,54,69,72,73,75,80
アルツハイマー病 ----------------------------------------------------- 1,5,43,50,58,61,63,67,74,78,82
アロステリック制御 ------------------------------------------------------------------------ 15,16,25,34,80
インテグリン ------------------------------------------------------------------------------------- 14,15,52,68
インスリン ---------------------------------------------- 10,14,35,68,69,70,71,72,73,74,75,77,79,80
インスリン受容体 --------------------------------------------------------------------------- 10,69,70,73,74
インスリン抵抗性 ----------------------------------------------------------------- 35,62,71,72,73,77,111
インスリン分解酵素 -------------------------------------------------------------------------------- 53,62,77
インスリンシグナリング ------------------------------ 2,3,70,71,73,74,75,79,80,83,93,94,95,111
オートファジー -------------------------------------------------------------------------- 17,53,55,56,57,71
カテプシン D ------------------------------------------------------------------------------------------ 53,62,64
ガーデンアンゼリカ ------------------------------------- 1,3,97,100,101,105,106,107,109,110,111
モノカルボン酸トランスポーター ------------------------------------------------------------------ 10,92
キナーゼ ----------------------------------------------------------------------------- 14,15,16,34,41,56
クマリン -------------------------------------------- 3,79,87,96,98,99,101,104,105,109,110,111,112
グリコーゲン ----------------------------------------------------------------------------- 23,68,70,71,73,89
グリコーゲン合成酵素キナーゼ 3β ---------------------------------------------------------------- 70,73
グルコース ------------------------------------------------------- 10,35,50,57,68,69,70,71,72,73,80,89
グルタミン酸 --------------------------------------------------------------------------------------- 8,37,59,77
グルタチオン ---------------------------------------------------------------------------------------- 27,94,111
クロス・トレランス ------------------------------------------------------------------------------------- 34,89
クロスβシート ------------------------------------------------------------------------------------------ 50,67
コリン作動性ニューロン ---------------------------------------------------------------- 35,76,77,95,107
コレステロール ------------------------------------------------------------------------------ 10,11,13,61,92
シグナリング -- 2,11,12,13,14,15,16,17,24,25,28,29,32,34,38,40,41,42,47,53,54,65,66,
73,81,91,95
シナプス ---------------- 8,35,36,37,38,39,40,42,45,46,47,48,49,50,53,61,64,65,70,74,76,111
シナプス間隙 ------------------------------------------------------------------------------------------- 8,38,41
シナプス後膜 ----------------------------------------------------------------------------------- 8,37,38,64,65
シナプス前膜------------------------------------------------------------------------------------------------- 8,38
ストレス応答 ------------- 2,16,24,25,28,29,30,33,35,43,57,66,67,71,73,80,81,89,94,108,111
a
ストレス耐性 ------------------------------------------- 2,32,33,34,81,85,89,94,95,108,110,111,113
スパイン ------------------------------------------------------------------------------------- 37,39,64,65,108
セリン・スレオニンキナーゼ ------------------------------------------------------------- 16,17,35,56,60
セルロース ---------------------------------------------------------------------------------------- 89,90,91,93
ソマトスタチン ------------------------------------------------------------------------------ 53,54,63,69,75
タウ ------------------------------------------------------------------------ 43,44,45,57,58,62,65,74,75,81
チロシンキナーゼ ----------------------------------------------------------------------- 14,15,16,52,69,74
トウキ ----------------------------------------------- 1,3,79,86,87,91,96,97, 99,100,104,105,109,110
ニューロン ------- 3,7,8,9,10,28,29,32,33,35,36,37,38,40,41,43,45,46,47,48,49,50,53,54,58,
59,61,63,64,65,74,77,78,80,84,92,95,97,108,109
ニューロンネットワーク --------------------------------------------- 8,35,36,37,39,47,65,66,73,108
ヌカ ------------------------------------------------------------------------------------------------------- 1,91,97
パーキンソン病 --------------------------------------------------------------------------- 6,50,58,59,67,78
パターン認識受容体 -------------------------------------------------------------------------------- 14,15,52
ハンチントン病 ---------------------------------------------------------------------------------- 58,59,60,67
フェノール酸 --------------------------------------------------------------------------------- 87,89,93,96,97
フェルラ酸 ------- 1,2,3,10,79,81,87,88,89,90,91,92,93,94,95,96,97,98,99,100,101,104,105,
106,108,109,110,111,112
フェルラ酸アラビノース -------------------------------------------------------------------------- 89,92,93
フェルラ酸ダイマー ------------------------------------------------------------------------------------ 90,93
フロクマリン -------------------------------------------------------------------------------- 99,104,105,107
プロテアソーム ------------------------------------------------------------------------------ 17,50,65,80,94
ヘム ------------------------------------------------------------------------------------------------- 22,26,33,93
ヘム酵素 ----------------------------------------------------------------------------------------------- 26,30,93
ペルオキシダーゼ ------------------------------------------------------------------------------- 27,28,93,95
ホメオスタシス ----------------------------------------------- 21,24,28,29,35,75,79,80,81,82,83,113
マクロファージ -------------------------------------------------------------------------------- 8,26,32,40,52
マトリックス ----------------------------------------------------------------------------------------- 13,47,53
ミクログリア ------------------------------------------------------------------------------------------- 8,30,95
ミトコンドリア -------------------------------------------------------------------------- 22,31,32,53,65,81
ユビキチン ----------------------------------------------------------------------- 17,15,55,56,58,80,81,94
リソソーム ------------------------------------------------------------------------------------ 17,52,53,54,55
リポ多糖類 -------------------------------------------------------------------------------------------- 30,51,89
レビー小体型認知症 ------------------------------------------------------------------------- 6,58,59,76,78
一塩基多型 ------------------------------------------------------------------------------------------------ 19,62
遺伝子 ------------------- 10,11,16,17,18,19,22,23,24,28,33,45,46,59,61,62,63,64,79,81,90,94
遺伝子多型 -------------------------------------------------------------------------------------- 19,62,64,110
b
炎症 -------- 2,3,13,24,28,29,30,35,40,53,57,58,59,65,66,73,79,81,85,89,94,95,103,107,109,
110,111
炎症性サイトカイン ------------------------------------------------------- 13,29,30,52,53,66,73,88,91
獲得免疫 ------------------------------------------------------------------------------------------- 24,29,32,56
家族性アルツハイマー病 -------------------------------------------------------------------------- 44,61,64
間質液 ---------------------------------------------------------------------------------------------------- 9,49,63
虚血耐性 ----------------------------------------------------------------------------- 32,33,34,89,94,95,108
筋萎縮性側索硬化症 -------------------------------------------------------------------------------- 58,60,78
原核生物 --------------------------------------------------------------------------------------------- 10,22,32
恒常性維持 ---------------------------------------------------------------------------------- 21,22,35,57,112
抗炎症 ------------------------------------------------------------ 3,30,53,66,77,84,91,104,109,110,111
好中球 ---------------------------------------------------------------------------------------------- 26,27,32,52
呼吸バースト ----------------------------------------------------------------------------------------- 27,32,52
細胞核 --------------------------------------------------------------------------- 2,10,18,20,31,71,80,90,94
細胞質 ------------------------------------ 10,11,13,17,20,31,33,38,50,51,55,56,57,59,74,80,93,94
細胞膜 -------- 8,10,12,13,15,16,17,22,24,30,31,32,34,39,40,46,47,51,52,53,54,55,61,65,66,
69,73,74,84,89,90
酸化ストレス --------------------------------------------------------------- 17,28,33,34,71,73,80,81,108
酸化ストレス応答 ----------------------------------------------------------------- 26,28,29,53,66,80,108
酸化ストレス耐性 ------------------------------------------------------------------------------- 34,89,94,95
自然免疫 ----------------------------------------------------------------------------------------------- 24,29,94
受動的回避試験 --------------------------------------------------------------------------------- 48,97,98
受容体 ------------------------- 8,10,13,14,15,16,24,34,37,38,39,40,45,51,52,53,63,64,65,76,77
小胞体 --------------------------------------------------------------------- 20,25,31,32,33,35,40,50,51,72
小胞体ストレス応答 ----------------------------------------------------------- 24,25,29,30,33,35,66,73
生活習慣病 -------------------------------------- 2,3,6,24,67,76,78,79,80,81,82,83,92,111,112,113
線維状βアミロイド -------------------------------------------------------------------------------- 49,53,65
真核生物 ----------------------------------------------------------------------------------- 10,20,22,23,25,31
神経原線維変化 ------------------------------------------------------------------------------------------ 43,58
生長 -------------------------------------------------------------------------------- 2,3,23,34,57,70,71,79,91
選択的スプライシング --------------------------------------------------------------------------------- 23,46
染色体 ------------------------------------------------------------------------------------------ 18,45,46,59,62
第Ⅰ相系 -------------------------------------------------------------------------- 26,27,28,32,51,71,93,94
第Ⅱ相系 ---------------------------------------------------------------------- 26,27,28,32,51,58,71,93,94
多型 ----------------------------------------------------------------------------------------------- 19,62,66,110
多発性硬化症 ----------------------------------------------------------------------------------------- 50,59,60
低酸素応答 ------------------------------------------------------------------------------------------------ 25,28
c
電子伝達系 ------------------------------------------------------------------------------ 31,32,51,52,53,108
糖尿病 --------------------------------------------- 2,9,10,14,35,60,62,67,68,71,76,78,79,91,111,113
転写エレメント ---------------------------------------------------------------------------------- 18,19,28,33
熱ショックタンパク質 ----------------------------------------------------------------------------- 50,60,63
脳関門 ----------------------------------------------------------------------------------- 9,45,61,73,77,84,92
脳虚血 -------------------------------------------------------------------------------------------------- 28,89,94
脳髄液 -------------------------------------------------------------------------------------------- 9,45,49,62,63
微小管 -------------------------------------------------------------------------------------- 40,44,45,65,70,75
微小管結合タンパク質 ------------------------------------------------------------- 41,43,58,62,65,70,75
非ステロイド抗炎症剤 ----------------------------------------------------------------------- 30,66,77,109
慢性炎症 ------------------------------------------------------------------------------- 77,79,82,92,104,109
老年性アルツハイマー型認知症 --------------------------------------------------------- 2,44,61,81,109
ACh ------------------------------------------------------------------------------------ 35,75,76,107,108,109
AChE ---------------------------------------------------------------------------------------------- 76,77,85,108
AD -------- 1,2,3,5,6,34,43,44,45,47,57,58,59,60,61,62,63,64,65,66,67,68,74,75,76,77,78,79,
80,81,82,83,84,85,95,96,97,98,99,100,101,102,104,108,109,110,111,112,113
Akt ------------------------------------------------------------- 17,28,43,57,62,66,70,71,75,80,81,94,95
ALS ---------------------------------------------------------------------------------------------------------- 58,60
AMPA ----------------------------------------------------------------------------------------------- 37,38,39,64
ANM176® ---------------------------------- 1,3,97,100,101,102,104,105,106,108,110,111,112,113
Apo --------------------------------------------------------------------------------------------- 61,62,63,66,110
ApoJ ----------------------------------------------------------------------------------------------------- 51,63,64
APP ---------------------------------------------------------------------------------------------- 46,47,48,53,64
ARE ------------------------------------------------------------------------------------------------- 28,71,93,94
ASK ------------------------------------------------------------------------------------------------------ 35,60,67
Aβ40 ------------------------------------------------------------------------------------ 47,48,49,51,54,63,64
Aβ42 -------------------------------------------------------------------------------- 47,49,51,54,62,63,64,65
BBB ----------------------------------------------------- 9,10,29,45,58,61,62,63,64,65,73,75,77,84,92
COX ---------------------------------------------------------------------------------------------------------- 30,66
CSF ---------------------------------------------------------------------------------------------------- 9,45,62,63
CRMP2 ----------------------------------------------------------------------------------------- 41,42,65,70,75
DLB --------------------------------------------------------------------------------------------------- 6,59,60,76
DM ------------------------------------------------------------------------- 2,3,67,68,71,72,73,75,79,80,81
DNA ---------- 10,11,14,18,19,20,22,23,26,28,29,30,31,34,46,50,59,62,64,71,72,77,85,93,94
ER --------------------------------------------------------------------------- 20,25,26,33,35,40,50,60,67,72
FAD -------------------------------------------------------------------------------------------------- 44,45,61,64
d
G タンパク質 --------------------------- 14,15,16,17,32,40,41,42,43,51,52,59,65,66,70,74,75,79
G タンパク質共役受容体 --------------------------------------------------------------------------- 15,40,65
GEF ---------------------------------------------------------------------------------------------------------- 15,16
GFAP ---------------------------------------------------------------------------------------------------- 29,50,58
GRP78 ------------------------------------------------------------------------------------------------------ 26,66
GSH ----------------------------------------------------------------------------------------------------- 27,94,95
GSK3β --------------------------------------------------------------- 3,70,71,73,74,75,80,81,93,94,110
HDL --------------------------------------------------------------------------------------------------------- 10,61
Hsp ---------------------------------------------------------------------------------------------- 50,51,60,63,64
IDE -------------------------------------------------------------------------------------------------- 53,62,64,77
IGF ------------------------------------------------------------------------------------------ 10,14,63,70,73,74
IL-1β ------------------------------------------------------------------------- 28,30,35,52,53,66,88,91,95
INMⓇ176 ------------------------------------------------------------ 97,98,99,100,101,103,104,105,110
IR ----------------------------------------------------------------------------------------------------- 10,69,73,74
IREl ---------------------------------------------------------------------------------------------------------- 26,72
JNK --------------------------------------------------------------------------------------------- 16,17,60,67,73
Keap1 ------------------------------------------------------------------------------- 2,3,58,80,81,94,110,111
LC3 ---------------------------------------------------------------------------------------------------------- 55,58
LDL ----------------------------------------------------------------------------------------- 10,11,61,62,92,94
LDL 受容体 --------------------------------------------------------------------------------------- 11,15,61,62
LRP1 -------------------------------------------------------------------------------------------- 11,45,62,63,64
LRP2 -------------------------------------------------------------------------------------------------------- 63,64
LPS ---------------------------------------------------------------------------------------------- 30,51,52,53,89
LTP ------------------------------------------------------------------------- 36,37,38,39,45,47,48,50,53,74
MAC1L ------------------------------------------------------------------------------------------------- 52,65,67
MAP ------------------------------------------------------------------------------------ 41,44,58,62,65,70,75
MAP1 ----------------------------------------------------------------------------------------------- 41,58,71,75
MAPK ----------------------------------------------------------------------------------------------- 16,28,43,53
MCI ---------------------------------------------------------------------------------------------- 5,65,83,85,103
MCT --------------------------------------------------------------------------------------------------------- 10,92
mRNA ---------------------------------------------------------------------------------- 18,20,21,23,25,57,75
MT ---------------------------------------------------------------------------------- 40,41,44,57,58,65,70,75
mTOR ------------------------------------------------------------------------------------------- 56,57,71,81,95
mTOR1 ------------------------------------------------------------------------------------------------- 56,57,71
MTT 法 ------------------------------------------------------------------------------------- 31,32,51,53,64,95
NADPH ------------------------------------------------------------------------------------ 27,32,33,51,94,95
NADPH オキシダーゼ ----------------------------------------------------------------------------- 32,51,52
e
NEP ------------------------------------------------------------------------------- 53,54,62,63,64,65,75,110
NF-κB ----------------------------------------------------------------------------------------- 16,53,73,74,77
NFT ------------------------------------------------------------------------------------------------------ 43,45,58
NMDA ---------------------------------------------------------------------------- 37,38,39,65,74,77,85,108
Nrf2 ---------------------------------- 2,3,28,33,34,58,62,70,71,80,81,89,93,94,95,96,108,110,111
NSAID -------------------------------------------------------------------------------------------- 30,66,77,109
PDK1 ---------------------------------------------------------------------------------------------------- 17,57,70
PDZ ---------------------------------------------------------------------------------------- 38,39,51,64,65,108
PERK --------------------------------------------------------------------------------------------------- 26,72,73
PI3K ------------------------------------------------------------------17,32,51,53,69,70,74,79,80,95,108
PIP3 ----------------------------------------------------------------------- 13,17,32,51,52,53,69,70,74,108
PSD-95 ---------------------------------------------------------------------------------------- 38,39,64,65,108
p38 ------------------------------------------------------------------------------------------- 16,17,28,53,66,81
p62 ------------------------------------------------------------------------------------------------------- 55,57,58
p450 ------------------------------------------------------------------------------------------------ 23,27,28,107
Rab ----------------------------------------------------------------------------------------------------------- 40,65
Ras --------------------------------------------------------------------------------------------------- 16,40,42,65
Rho ---------------------------------------------------------------------------------------------- 40,41,43,66,75
ROS -------------------------------------------------------------------------------------------------- 34,35,92,94
RTK ------------------------------------------------------------------------------------------------------ 14,15,16
SDAT ------------------------------------------------------------------------- 2,44,45,61,62,78,81,109,110
SNP ---------------------------------------------------------------------------------------------------------- 19,62
TLR -------------------------------------------------------------------------------------------------- 15,16,51,53
UPR --------------------------------------------------------------------------------- 25,26,30,33,35,66,72,73
αシヌクレイン ------------------------------------------------------------------------------------------ 59,67
βアミロイド --- 1,2,3,11,43,44,45,46,47,48,49,51,53,54,57,58,50,60,61,62,63,64,65,66,
67,73,75,76,78,80,81,82,84,85,86,87,88,89,95,96,97,98,107,108,109,
111,112
βアミロイド・モノマー ------------------------------------------------------------------------ 49,51,63,64
βアミロイド・オリゴマー ----------------------------- 49,50,51,54,63,64,65,66,74,76,80,84,111
βアミロイド凝集体 ---------------------------------------------------------------------------- 51,52,53,65
βアミロイド神経毒性 ------- 1,2,3,49,51,64,74,75,80,86,87,88,89,95,96,97,98,99,100,101,
104,105,108,109,110
βアミロイド複合受容体 ---------------------------------------------------------------- 52,65,66,67,108
βシート構造 ---------------------------------------------- 38,49,50,52,53,59,60,64,65,67,80,81,111
4EBP -------------------------------------------------------------------------------------------------------- 21,57
f
© Copyright 2026 Paperzz