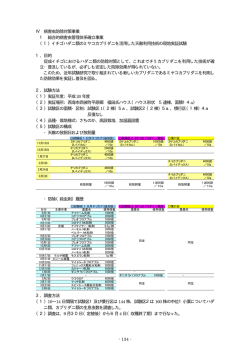
バイオコントロール 第15巻1号 目 次 IPMにおける寄生菌と寄生蜂 -最近の研究から- ·······························1 静岡大学 農学部 応用昆虫学教室 西東 力 神奈川県におけるIPM(総合的病害虫管理)の現状について ·······················6 神奈川県農業技術センター 藤代 岳雄 スモモヒメシンクイの生態と防除·················································11 山梨県果樹試験場 環境部 村上 芳照 ホウレンソウケナガコナダニの 簡易モニタリング装置(コナダニ見張番)の開発と被害予測による防除体系 ·········· 17 山口県農林総合技術センター 農業技術部 本田 善之 スワルスキーカブリダニに対する農薬の影響······································ 23 宮城県農業・園芸研究所 園芸環境部 宮田 將秀 ナシ黒星病の減農薬防除の取り組み·············································· 29 千葉県農林総合研究センター 病理昆虫研究室 金子 洋平 茶栽培におけるニーム散布剤の利用 ~ベトナムでの実践~ ······················· 34 東京農工大学 濱周吾 仲井まどか 熊本県におけるトマト黄化葉巻病の防除対策······································ 40 熊本県農業研究センター 生産環境研究所 樋口 聡志 ブラジルの生物的害虫防除について·············································· 47 サンパウロ州立大学 植物保護学部 昆虫学科教授 オダイル・フェルナンデス 在来種天敵の農薬としての登録は不必要·········································· 52 ジャパンIPMシステム代表 和田 哲夫 潮流は生物農薬の利用の方向に向かっているのか? ································ 54 ヴィレム・ラーフェンスベルグ Ph.D. Koppert Biological Systems 高知県の施設果菜類における天敵を利用したアザミウマ類防除の現状 ················ 59 高知県農業振興部 環境農業推進課 古味 一洋 天敵誘引物質と天敵の関係について·············································· 63 京都大学生態学研究センター 上船 雅義 バラのカブリダニを用いた省力防除法の普及······································ 69 神奈川県立フラワーセンター大船植物園 植栽課 関塚 知己 第 13 回国際生物防除機構(IOBC:International Organisation for Biological and Integrated Control of Noxious Animals and Plants)の 「昆虫病原体および昆虫寄生性線虫」ヨーロッパ会議に参加して ···················· 71 帯広畜産大学 相内 大悟/小池 正徳 ― 随 想 ― 「鳴かない虫と音楽」 (後編) ················································· 76 柏田 雄三 協 議 会 規約 ··································································· 84 お 知 ら せ ··································································· 85 資料 天敵に関する農薬の影響表(第20版) ········································ 綴じ込み 最新版 生物農薬登録状況 ···················································· 綴じ込み 表 紙:黄釣舟と虎丸花蜂 (キツリフネとトラマルハナバチ 和田 原図) 2000年頃増殖に成功したトラマルハナバチ。本来はツリフネソウやアザミ、ブルベリー の花などを好むが、トマトのハウスで働かせられるのは不本意であろう。 裏 表 紙:表4写真(古味 原図) IPMにおける寄生菌と寄生蜂 ー最近の研究からー 静岡大学 農学部 西東 力 はじめに 今日、重要害虫のほとんどが侵入害虫と言っても過言ではない。この 20 年間に限 ってみても、タバココナジラミ、マメハモグリバエ、ミカンキイロアザミウマ、トマ トハモグリバエなどがつぎつぎと侵入し、我が国の農業に甚大な被害を与えてきた。 いずれもコスモポリタンの害虫であり、彼らの世界も急速にグローバル化しつつある。 コスモポリタン害虫の共通点は、①殺虫剤抵抗性、②非休眠、③広食性という性質を 併せ持つことである。殺虫剤抵抗性については、大きな被害を及ぼす直接、そして最 大の要因となっている。非休眠については、年間を通して問題化する背景となってい る。広食性については、急速な分布拡大の要因となっており、コスモポリタンの所以 でもある。 生物的防除法を基幹技術とする IPM では、こうしたコスモポリタン害虫が主体と なっている。しかし、いざ IPM を実践しようとすると、農薬使用の制限という大き な壁に直面することになる。天敵に影響のある農薬は使用できないばかりか、使用法 を間違えればリサージェンスを招きかねない。薬剤抵抗性の天敵があれば、IPM を 足踏みさせているこの壁をブレイクスルーできるかもしれない。 ここでは、侵入害虫のその後の動向に触れたうえで、殺虫剤抵抗性の寄生バチと殺 菌剤耐性の昆虫病原菌について紹介したい。 1.侵入害虫の運命 1989 年と 1990 年は侵入害虫の当たり年であった。前述のタバココナジラミ、マメ ハモグリバエおよびミカンキイロアザミウマはこの 2 年間に相前後して確認された 侵入種である。侵入当初、高度の殺虫剤抵抗性と爆発的な増殖によって壊滅的な被害 が生じたが、今日、いずれも一般害虫とみなさるまでに密度が低下しており、かつて の異常発生の記憶も薄れつつある。これら3種のみならず、侵入害虫は、皆、同じ運 命をたどっている。同様の現象は海外でも知られている。こうした密度低下はどうし て起こるのか。関係者の努力と防除の徹底だけでは、到底、説明できない。侵入害虫 の増減には、人為的な要因を越えた何か大きな力が働いているように思えるが、この 謎を解く糸口は想像の域を出ない。 侵入害虫が一般害虫化すると、かつてのような異常発生は起こらなくなる。しかし、 殺虫剤抵抗性をはじめとする性質はそのまま保持していることから、気を抜けば大き な被害につながる。IPM の中心的な対象害虫であることに変わりない。 2.寄生バチ (1) 殺虫剤抵抗性の寄生バチ ハモグリバエとその寄生バチに及ぼす殺虫剤の影響を調べるなかで、殺虫剤を散布 すると密度が増加する寄生バチ( Halticoptera circulus)がみつかった(図1)。殺 虫剤に対する感受性検定においても、この寄生バチは他の寄生バチと比べて殺虫剤抵 抗性の高いことが確認された(表1)。本種の発育日数は、生物農薬として市販され ているハモグリコマユバチやイサエアヒメコバチと比べるとかなり長い(表2)。 H.circulus は、ナモグリバエ、トマトハモグリバエおよびマメハモグリバエに寄生 するが、通常ではマイナーの寄生バチである。マメハモグリバエに対する選好性はと くに低い。一方、ネギハモグリバエでは優占種のひとつに挙げられている(大井田ら、 2009; 井村、私信)。この理由は、ネギハモグリバエに対して選好性が高いためか、 ネギ畑で使用される殺虫剤に対して抵抗性を示すためであろう。H.circulus は海外で も知られているが、その生態はほとんど調べられていない。 D. isaea 80 60 40 20 0 Nov. 1-Dec Dec. 1-Jan Jan. 1-Oct Oct.1-Nov 2004 2005 Feb.2004 Mar. 2005Apr. 1-Feb 1-Mar 1-Apr 40 20 Feb. 2004 Mar. Apr. 2005 1-Feb 1-Mar 1-Apr 100 H. circulus 80 30 % Comosition % Composition 60 0 Nov. 1-Dec Dec. 1-Jan Jan. 1-Oct Oct. 1-Nov 2004 2005 C.pentheus 40 D.minoeus 80 % Composition % Composition 100 20 10 40 20 Saito et al., 2009 0 Nov. 1-Dec Dec. 1-Jan Jan. 1-Oct Oct. 1-Nov 2004 2005 60 Feb. 1-Feb Mar. Apr. 1-Mar 1-Apr 00 Nov. 1-Dec Dec. 2005 Jan. 1-Oct Oct. 2004 1-Nov 1-Jan 2004 2005 Feb. 1-Feb Mar. 1-Mar Apr. 1-Apr 図 1. 殺虫剤散布下におけるナモグリバエの4種寄生バチの消長(エンドウ畑)(Saito et al., 2009) マラチオン乳剤 トラロメトリン乳剤 無処理 表 1. ハモグリバエと寄生バチの殺虫剤感受性(LD50:ng/mg) (松田ら、2010) マラチオン ペルメトリン LD50 R比 LD50 R比 657 9,660 0.936 8.9 229 3,370 1.89 34.0 500 0.962 イサエアヒメコバチ 0.224 3.3 ハモグリヒメコバチ 0.104 カンムリヒメコバチ 0.076 ハモグリミドリヒメコバチ H. circulus イミダクロプリド LD50 エマメクチン 安息香酸塩 R比 LD50 R比 171 8.,550 33.8 338 18 69.6 3,480 0.266 2.7 9.2 18.4 920 5.86 58.6 1.35 12.9 0.172 8.6 1.12 11.2 1.5 0.197 1.9 0.020 1 0.124 1.2 1.1 0.105 1 0.059 3.0 0.304 3.0 0.068 1 0.184 1.8 0.056 2.8 0.100 1 8.58 126 ハモグリバエ マメハモグリバエ トマトハモグリバエ ナモグリバエ 寄生バチ 1) LD50値の単位はmg/g 2) R比はLD50値が最も低い虫を1とした場合の指数 表 2. H. circulus と生物農薬(ハモグリコマユバチ、イサエアヒメコバチ)の比較(剣持ら、2011) 寄生バチ 発育日数(卵-羽化) 発育零点(℃) 有効積算温度(日度) 寄生様式 H. circulus 27.9 (20℃) 8.4 330 飼い殺し ハモグリコマユバチ 15.7 (22℃) 7.2 256 飼い殺し 16.6 (20℃) 8.6 175 殺傷 (Malais,1992) イサエアヒメコバチ (Malais,1992) (2)ハモグリバエの人工飼料 市販の寄生バチの生産は、植物に寄生させたハモグリバエを用いて行われている。 ハモグリバエの人工飼料を開発できれば、寄生バチの大量生産に役立つかもしれない。 本研究は、①採卵装置の開発、②幼虫用飼料の開発、の2段階に分けて行っている。 ①については、アザミウマ用の採卵装置(Murai and Loomans, 2001)を改良した(図 2)。②については、市販の人工飼料を改良し、ふ化幼虫を蛹化まで発育させること ができた。しかし、蛹化個体は稀であり、羽化個体も得られていない。餌の改良を進 めているところである。 図 2. ハモグリバエの採卵装置(左)と装置内に産卵された卵(右)(溝口ら、2011) 3.昆虫病原菌 (1)殺菌剤耐性菌の育種 昆虫病原菌を利用に当たって殺菌剤の影響が問題となる。殺菌剤耐性の昆虫病原菌 を育種できれば、使用できる殺菌剤が増える。 代表的な昆虫病原菌( Beauveria bassiana、Isaria fumosorosea)に量子ビーム(イ オンビーム、ガンマ線)を照射した結果、ベノミル剤とトリフルミゾール剤に対する 耐性レベルが 4~5 倍高い変異体が得られた(表3、表4)。これらの変異体は他の殺 菌剤に対しても耐性レベルが上昇しており、交差耐性を示すことがわかった。殺菌剤 耐性変異体の病原力は低下傾向を示したが、元菌と同等の病原力を保持している変異 体も得られている。 表 3. I. fumosorosea 変異体におけるベノミルとトリ フルミゾールの MIC (ppm)(篠原ら、2011) 変異体 ベノミル 4-Ben34P1 * > 5,000 2-PB1 * 5,000 R1-1B8 ** > 5,000 トリフルミゾール > 1,200 表 4. B. bassiana 変異体におけるベノ ミルの MIC (ppm)(篠原ら、2011) 変異体 ベノミル 2-BB22 * > 5,000 2-BB24 * > 5,000 5-Ben47B * > 5,000 R1-2T20 ** > 1,200 R1-2B33-1 ** > 5,000 R1-2T22 ** > 1,200 元菌(BB1026) 1,000 300 * 単独照射 ** 2 段階照射 元 菌 1,000 (PF3110) * 単独照射 ** 2 段階照射 (2)輸入菌と国内菌の病原力の比較 天敵昆虫については国内の生物農薬が増えつつある。一方、天敵微生物については 依然として外国産が主流である。はたして外国の天敵微生物は国内のそれより優れて いるのであろうか。表5は I. fomosorosea の輸入製剤と保存菌株の病原力を比較した もので、病原力はほぼ同等であることがわかる。このことは、外国の天敵微生物に引 けを取らない天敵微生物が国内にも存在することを示している。 微生物農薬の開発には、素材そのものの力価はもとより、大量生産の難易などコス ト面の要因が大きくかかわってくる。また、マーケットが小さければ、オリジナル商 品の開発意欲は削がれ、すでに実績のある輸入品に頼ることになる。こうしたことを 考えると、天敵微生物の病原力が同等だからといって、直ちに国内の天敵微生物を生 物農薬として開発しようとするのは早計である。しかし、国内にも有望な天敵微生物 がいることを忘れてはならないと思う。 表5. 保存菌および微生物農薬のタバココナジラミに対する病原力 (Saito and Sugiyama, 2005) おわりに IPM は、その理念はともかく、現実的には生物農薬あるいは土着天敵を中心にすえ、 必要に応じて化学合成農薬を組み込んでいくのが一般的であろう。このため、IPM を 実践するうえでは天敵と化学合成農薬の関係の理解が欠かせない。こうした面から、 本稿では殺虫剤抵抗性の寄生バチと殺菌剤耐性の昆虫病原菌をとりあげたが、この分 野の研究はあまり進んでいない。使い勝手のよい天敵を探し出し、その力を十分に発 揮させることができるようになるまでには、まだまだ多くの課題が残されている。 神奈川県におけるIPM(総合的病害虫管理)の現状について 神奈川県農業技術センター 藤代岳雄 1 はじめに 神奈川県におけるIPM技術の導入は、大正5年にミカンのイセリアカイガラムシ対策と して、ペダリアテントウムシを導入・放飼したものが最初の大きな取組である。その後、 果樹では、黄色防蛾灯の利用、ウメ、ナシ等での性フェロモンによる交信攪乱剤の利用 (コンフューザーN、スカシバコンなど)、最近ではハウスミカンでスワルスキーカブリダ ニの導入などが試みられている。 野菜では、古くはマリーゴールドによるキタネグサレセンチュウ防除に始まり、オンシ ツツヤコバチによるオンシツコナジラミの防除、ハスモンヨトウの性フェロモンによる大 量誘殺、ナス畑周囲のバンカープランツ(ソルゴー)の栽植、イチゴの害虫に対する各種 天敵を利用したIPM防除(ハダニ類に対するチリカブリダニとミヤコカブリダニ、アブ ラムシ類に対するコレマンアブラムシ等)が導入され、現在、キュウリのスワルスキーカ ブリダニの導入等が試みられている。 このように、数々のIPM技術が県内の生産現場で導入されたが、広く普及したもの、 利用が限定的だったもの、発売直後はほとんど普及しなかったが、他の防除技術の発達や 技術の再評価がされて、再び日の目を見ることになった技術、ほとんど利用されなくなっ た技術など状況は様々である。今回、筆者が生産現場での普及指導に当たったイチゴ・キ ュウリの天敵利用防除の導入・普及の経過を中心に、普及指導という観点から神奈川県に おけるIPM技術の現状と導入に当たっての注意点等について紹介したい。 2 野菜 (1)施設イチゴ 施設イチゴの減収要因として、ハダニの被害は大きく、有効な薬剤が少ないこと、収 穫期に入ってからの防除が労力面で大変なこと、ミツバチによる交配が早くから普及し 薬剤散布が制限されることなどから、化学農薬に依らないIPM防除技術の導入が望ま れていた。1991年にチリカブリダニ剤が市販されると、各地のイチゴ産地で展示ほ等で 普及が図られたが、チリカブリダニ剤単独では効果の持続性に問題があり、また化学農 薬との組み合わせが不十分であったこともあり、海老名市の生産部会で利用されるに留 まった。その後、ミヤコカブリダニ剤が2003年に上市されると、農業技術センター農業 環境研究部での研究結果や天敵に影響の少ない化学農薬やチリカブリダニ剤との組み合 わせなどの知見を総合して、天敵導入マニュアルが作成され、スケジュール散布で防除 してもほぼ失敗なくできるようになった。現在では県内のほとんどの産地で、ハダニ天 敵の導入が進んでいる。また、先進地の海老名市では、カブリダニ類のほか、アザミウ マ防除のククメリスカブリダニ(ククメリス)、アブラムシ防除のコレマンアブラバチ 剤(アフィパール)と天敵の餌となるムギクビレアブラムシ(アフィバンク)、センチ ュウ対策としてギニアグラスの緑肥栽培などが導入されている。県内の他産地では、ハ ダニの天敵導入がほとんどで、アブラムシの天敵防除が一部で導入されているが、クク メリスは春先からのアザミウマの飛び込みに対応できず化学農薬を用いることが多い。 イチゴにおけるスワルスキーカブリダニの利用は、イチゴの栽培管理温度が比較的低い ことから、本圃での導入は困難と見られるが、ホコリダニ類の防除には極めて効果があ ることから、施設内育苗のナイアガラ方式で、導入を試みる生産者も一部ある。 2009年から、藤沢市や海老名市でオオタバコガ、ハスモンヨトウ防除のためフェロモ ン剤(コンフューザーV)を導入し始めている。施設内での増殖を抑えるための技術であ るが、導入者の評価は高く、今後普及の拡大が予想される。 (2)施設キュウリ 施設キュウリは、慢性的なネコブセンチュウの被害があり、D-D剤など化学農薬を用い た土壌消毒が行われてきた。2001年にパスツーリアペネトランス水和剤(パストリア水 和剤)の展示ほが海老名市内で組まれたが、単年度の試験では防除効果は不十分で、当 時は活用方法が十分に分かっていなかったこともあり、その後も導入戸数はごく少数で あった。一方、2004年頃から土壌還元消毒がネコブセンチュウを含む多くの土壌病害虫 に効果を示したことから、環境保全型農業に取り組む生産者を中心に普及が進んだ。そ の後、2008年頃に大磯町の生産者が、混住化の中で、臭気を気にせずにネコブセンチュ ウを防除できる方法として、パスツーリアペネトランス水和剤を導入し、工夫を加えて ネコブセンチュウ防除に成功した。現在、パスツーリアペネトランス水和剤の利用方法 についてJA湘南管内の農業青年グループでプロジェクト研究を行っている。 また、県内最大産地のJA湘南管内のキュウリ産地では、黄化えそ病を媒介するミナ ミキイロアザミウマ防除も重要問題である。化学農薬防除では薬剤抵抗性の獲得が早く、 またウイルスを獲得した蛹が土中に落ちるために、薬剤散布だけでは完全な防除が困難 である。普及では2005年から4年間、ミナミキイロアザミウマの総合防除対策に取り組み、 筆者も2007年の1年間担当した。 ミナミキイロアザミウマは県内の露地では越冬しないことから、①抑制栽培終了後に 加温して、早く蛹を孵化させ餓死させること、②施設内の雑草防除を徹底すること、③ 県央地域の施設メロン栽培で取り入れられていた、側窓や出入り口のほか、天窓下に防 虫ネットを展張する方法などを紹介した。防虫ネット被覆を試みた生産者のほ場では黄 化えそ病の他、CMVや他の害虫も激減し、化学殺虫剤散布回数は約1/3に低下し、優良事 例と考えられたが、その後に同技術を導入した施設は立地条件によって、通気が不十分 となり、普及は限定的であった。現在、アザミウマ類の薬剤抵抗性を考慮して、バーテ ィシリウム レカニ剤(マイコタール)など微生物殺虫剤を展示ほで実施し、通常防除や スワルスキーカブリダニと組み合わせた方法を今後検討していく予定である。 アザミウマ類やコナジラミ類などの防除を目的としたスワルスキーカブリダニの導入 は2008年から行われており、徐々に導入農家が増えてきている。キュウリはうどんこ病、 べと病、褐斑病対策のための薬剤散布回数が多く、スワルスキーカブリダニの増殖に問 題のある農薬も使われることが多いことや、導入時期が難しいことから、急速な普及に は至っていない。今後は、化学農薬とのマッチングを含め、防除暦のようにスケジュー ル散布モデルを組み立てることなどが必要である。 (3)施設トマト 神奈川県では、1991年にオンシツツヤコバチを利用したオンシツコナジラミの防除試 験に取り組み、1995年にオンシツツヤコバチ剤(エンストリップ)の農薬登録が取れ、 普及センターによる展示ほの設置などを行った結果、1999年農薬年度には出荷量の増加 が見られた。その背景には、篤農家の観察により、導入時期の判定等ができるようにな ったことが大きい。その後、オンシツコナジラミ防除にピリプロキシフェノン剤(ラノ ーテープ)が発売され、発売当初は導入が緩やかであったが、オンシツツヤコバチ剤と 比べて、導入時期の観察等が不要であることや、省力効果が大きいことから、生産現場 の利用は急速に拡大 200 した。一方で、オン 180 シツツヤコバチ剤の 160 140 から激減した。 120 その後、2005年11 月にトマト黄化葉巻 kg 利 用は 2000 農 薬年 度 100 80 60 病が、神奈川県で初 40 発生が確認され、そ 20 の後急速に被害が拡 0 1996 1997 1998 1999 2000 2001 2002 2003 2004 2005 2006 2007 2008 2009 大したことから、ト マト黄化葉巻病ウイ ルス(TYLCV)を媒介 農薬年度 第1図 オンシツツヤコバチの神奈川県への出荷 するタバココナジラミの要防除水準が大幅に低くなり、育苗期からの粒剤施用等の薬剤 防除が必須になるとともに、温室開口部への防虫ネットの被覆、TYLCV耐病性品種の利用 等の技術が普及した。現在は天敵農薬を利用したIPM防除は海老名市内の5戸で確認され るのみで、当初導入されていたオンシツツヤコバチ剤より、タバココナジラミとオンシ ツコナジラミの両方に効果があるサバクツヤコバチ剤の導入が増えているが、サバクツ ヤコバチ剤の方がやや高温性であるため、オンシツツヤコバチ剤と使い分けされている。 なお、ピリプロキシフェノン剤(ラノーテープ)はタバココナジラミへの効果が劣るこ とから、近年では、漸減傾向である。 土壌害虫防除技術としては2004~2005年にかけて土壌還元消毒、神奈川県が開発した 熱水土壌消毒が導入され、両者とも主にネコブセンチュウや褐色根腐病等の対策として 普及した。土壌還元消毒は大量の用水を要し、臭気の問題があり、熱水土壌消毒は燃油 コストが比較的高く、熱水が土壌深部に到達する必要があることから、現在の普及状況 は横ばいないし、やや漸減傾向である。 第1表 イチゴ天敵等の普及状況(導入農家戸数) 天敵種類 平成 16 年 平成 17 年 平成 18 年 平成 19 年 ハダニ類天敵 24(34) 49(72) 55(81) 57(88) アブラムシ類天敵 11(16) 7(10) 27(40) 33(51) バンカープランツ 0( 0) 3( 4) 26(38) 30(46) ピリプロキシフェノンテープ剤 5(22) 18(70) 19(83) 20(87) 注1)天敵、バンカープランツは秦野市、伊勢原市、海老名市の合計。 注2)ピリプロキシフェノンテープ剤は秦野市のみの集計。 注3)数字は導入農家戸数(戸)。( )内は生産者の導入割合(%) (4)露地野菜 露地ナスでは、横浜市、川崎市、平塚市、伊勢原市、藤沢市などで、土着天敵(ヒメ ハナカメムシ)によるアザミウマ類、アブラムシ類の防除や農薬飛散防止、風除けを兼 ねて、ナスほ場の周囲にソルゴー栽植が行われている。品種は、背が高く、太くて硬い 「風立」や「つちたろう」が用いられている。最も普及した平塚市では平成19年度から 現在まで23名の部会員全員で、ソルゴー栽植が行われている。 三浦半島では、1970年代からダイコンのネグサレセンチュウ防除と目的としたマリー ゴールドの栽植が行われてきたが、定植や雑草の防除の手間がかかること、マリーゴー ルドにオオタバコガなどが多く食入して、その後の栽培に問題が生じることから、環境 保全型農業に取り組む一部の農家が続ける程度となった。現在では、風による土壌の流 亡防止、有機物の補給、センチュウ密度の低減等を目的にエンバク、ギニアグラスなど が導入されている。また、近年は匍匐性マリーゴールドの品種が育成され、研究段階で はあるが、土壌の被覆面積が大きく、オオタバコガ寄生数も少ないことから、今後の導 入が期待されている。 その他、綾瀬市内の有機生産者の要望で、コレマンアブラバチを活用したハクサイI PMの展示ほを平成22年から行っている。 3 果樹類 果樹におけるフェロモン剤を利用したIPM防除技術としては、約20年前に小田原市に 栽植されているウメのコスカシバ防除にチェルトリア剤(スカシバコン)が導入され、そ の後カキのコスカシバ、厚木市のナシのナシヒメシンクイ防除にオリフルア・トートリル ア、ピーチフルア剤等(コンフューザーN)が導入された。現在でも、フェロモン剤は一 部利用されているが、最盛期に比べると利用は限定的である。その理由として、コスカシ バ防除については、コスカシバ以外の樹幹害虫(マダラメイガ)の出現があり、ナシヒメ シンクイ防除については、比較的産地にまとまりがあり、広い地域でないと効かないこと、 近年はカメムシの吸汁被害が拡大しており、農薬散布回数の削減につながりにくいといっ たことが挙げられる。 その他の技術として、ブルーベリーのコガネムシ防除にスタイナーネマ・グラセライ剤 (バイオトピア)が各地で検討されているが、効果はやや不安定である。また、神奈川県 北部(相模原市)で合成ピレスロイド剤を削減し、土着天敵を温存してIPMを実践する 試みも行われているが、果樹の場合は、花きと同様にわずかな害虫被害があっても、経営 的な影響が大きいため、土着天敵利用は今後の課題である。ハウスミカンのIPMについ ては、ミカンハダニ防除にスワルスキーカブリダニ、アザミウマ防除に光を利用した物理 的防除が、農業技術センター足柄地区事務所で精力的に研究が行われ、スワルスキーカブ リダニの導入が数戸で行われているが、生産現場での普及はまだ端序に付いたばかりであ る。 4 最後に 神奈川県におけるIPMの普及について述べてきたが、広く普及し、しかも現在まで 持続する要因として、生産者の誰でも導入が容易で、失敗が少なく、しかも省力効果が 高いことなどが挙げられる。一度導入に失敗すると、再度産地に導入することは困難に なることから、観察眼の優れる篤農的農家との連携により、導入技術の評価や修正が必 要である。海老名市のトマトやイチゴにおいて、天敵農薬が進んだ要因として、天敵農 薬のパイオニア的生産者の存在が大きい。また、IPMといえども、非化学農薬だけで なく、積極的に化学農薬等との組み合わせや最新の研究成果や知見の技術化を検討し、 粘り強く技術の修正を図っていくことも重要と考えられる。 引用文献 小林正伸 (2009) バイオロジカルコントロール 13(1):25-30 小林正伸 (2010)第25回報農会シンポジウム 57-62 スモモヒメシンクイの生態と防除 山梨県果樹試験場・環境部・病害虫科 村上芳照 はじめに スモモヒメシンクイ Grapholita dimorpha Komai の山梨県における発生は、1994 年 にスモモで初確認した。年次により発生の多少はあったものの、近年の被害は大きく 拡大している。本種の被害は、果実被害であるためスモモに発生する病害虫の中では 最も重要な問題となっている。また、収穫時には被害果と健全果の区別が付きにくい 場合も多く、市場や消費者からのクレームも問題となる。 スモモは、スモモヒメシンクイの被害を除けばモモやブドウに比べて病害虫が少な い。しかし、本種の被害が拡大するにつれ、防除回数は増え、生産者の労力も増加し 安定生産の大きな障害となっている。本種は岩手県の山間高冷地において古くから発 生していたようで、その他青森県一帯、および福島・長野県の高冷地で被害が認めら れていた。現在は、山梨県、長野県、福島県、青森県、北海道など東日本で発生が確 認されている。一般に早生種での被害は少なく、中晩生種、特に晩生種での被害が大 きく、‘太陽’などの晩生種では成熟期の被害が出やすい。毎年発生している園では、 密度が高まり、時として多発し甚大な被害が発生することも多い。近年、リンゴでの 被害も増加し大きな問題となっている産地も見られる。ここでは、山梨県のスモモに おける生態と防除について紹介したい。 生 態 成虫は、落葉果樹の重要害虫であるナシヒメシンクイ Grapholita molesta (Busck) に類似している。4月下旬から 10 月上旬まで年3回~4回発生する。各世代の山梨 県での発生時期は図1のとおりである。標高 300~450m前後の地域では、1回目が 4月下旬~5月下旬、2回目が6月上旬~7月上旬、3回目が7月中旬~8月下旬、 4回目が8月下旬~9月下旬となる。3~4回目の発生は連続して明確なピークが見 られない場合が多い。 成虫は羽化数日後から産卵を始める。産卵は果実表面に1~2卵ずつ行われ、大き さは、直径 0.6~0.7mm、円形または楕円形で、はじめ乳白色であるが後に黄色~赤 黄色を帯びる(図2)。ふ化した幼虫は産卵された場所の近くから短時間で果実内に 食入する。果実内に食入した後は、種子の近くまで食害する場合と、果皮の直下を食 害する場合があり、皮下を食害した場合果面に黒いスジ状の特有の症状が見られる (図3)。ふ化・食入率も高いため、密度が高まると重大な被害となる。 若齢幼虫は、頭部が黒色で、胴部は乳白色で、老熟してくると赤みを帯びる。十分 に発育すると、体長 12mm 程度となり、果皮に紡錘形の切れ込みを入れ、その内面に マユを作り蛹化する(図4)。成熟果では果実の外に脱出し、土粒等でマユを作る。 卵から成虫までの発育期間は、25℃で 27 日である。卵期間は4日と比較的短い。 1雌の産卵数は平均で 55 卵である。 標高300 m 第2世代 (南 ア ル プス 市 ) 第3 世 代 第 1 世代 越冬 世 代 標 高400 ~450 m (山 梨 市江 曽 原・ 笛 吹 市境 川 ) 標 高700 m 6 5 4 3 2 6 10.1 5 4 3 2 6 9.1 5 4 3 2 6 8.1 5 4 3 2 6 7.1 5 4 3 2 6 6.1 5 4 3 2 6 5.1 5 4.4 (甲 州 市神 金 ・ 山梨 市 牧 丘) 月 ・ 半旬 図1 スモモヒメシンクイ成虫の標高別発生消長(模式図) *フェロモントラップへの誘殺数(H14~17 年)をもとに作成 図2 スモモヒメシンクイの卵 図3 被害果の症状 (果皮直下の食害) 図4 老齢幼虫と果面上の蛹室 化学的、耕種的防除 本種の防除は殺虫剤による防除が主体となるが、それだけでは十分な効果が得られ ない事例も多い。このように難防除化している中で、より効果を上げるためには総合 的な対策が必要となっている。 ○殺虫剤による防除 幼虫が果実に食入した後では、殺虫剤の効果は期待できないため、幼虫の食入防止 が中心となる。成虫は、羽化後数日で交尾・産卵をすることや、卵期間短いことなど を考慮すると成虫発生盛期に薬剤を散布することが効果的である。 薬剤の効果については、スカウトフロアブル、ダイアジノン水和剤、ダーズバン水 和剤、モスピラン水溶剤について、室内試験した結果、いずれの薬剤もふ化直後の幼 虫および成虫に対して高い殺虫効果が認められた。しかしながら、圃場では十分な防 除効果を得られない事例が多い。理由として、スモモは果面に果粉が多く、薬剤の付 着を妨げる現象が挙げられる。特に収穫期近くの果実では、果実全体が果粉で厚く覆 われ、薬液の多くがはじかれてしまう。果粉を洗い流すように散布すれば、果実の汚 染・果粉の溶脱などの品質の問題が生じる。高品質な果実を生産を目的とする現在の 栽培体系では今後も避けられない重要な問題となる。 1回目の防除は、越冬世代成虫の発生期となる5月上中旬となる。6月中下旬は、 2回目の成虫発生期となる。防除薬剤は、合成ピレスロイド剤が主体となるが、その ほかにスピノサド剤、クロラントラニリプロール剤などが防除薬剤となっている。 ○耕種的防除 幼果、肥大期の果実ともに幼虫の加害を受けた果実は落下しやすくなる。落下した 被害果を放置すると発生源となるため、土中に埋める、肥料袋等に集める等適切に処 理する。樹上の被害果についても見つけたら直ちに取り除き処分することが重要であ る。 交信かく乱剤による防除 モモ圃場においてナシヒメシンクイに対する交信かく乱剤(コンフューザーMMな ど)は高い防除効果が認められた。そこで、本種に対しても効果が期待できると考え 試験を行った。交信かく乱剤の利用においては、できるだけ処理面積が広いことが望 ましい。また、処理地域の生産者全員の参加が前提となる。しかしながら、すべての 生産者の合意を得ることは容易ではない。本種に対する交信かく乱剤の利用において は、小面積での利用が可能か検討した。 ○スモモヒメシンクイに対する交信かく乱剤の効果 コンフューザーMMは、ナシヒメシンクイ、モモハモグリガ他4種を対象とした製 剤である。スモモではモモハモグリガは発生しないため、本試験ではコンフューザー Nを使用した。はじめにコンフューザーNのかく乱効果を調査するため、モニタリン グトラップへの誘引阻害を調査した。現地圃場の広域処理(1ha)および果試場内 の小面積処理(10a)における誘引阻害はともに高いことが確認された(表1)。 ○コンフューザーNの小面積処理による防除効果 果試場内のスモモ圃場において、小面積処理によるコンフューザーNの効果につい て検討した。処理圃場の面積は 10a、無処理圃場は6aで、両圃場とも毎年本種が 多発している。コンフューザーNは4月下旬に設置した。6月 29 日、7月8日、7 月 19 日に薬剤散布を行い、被害果の調査は7月 26、27 日に行った。処理圃場は毎年 大きな被害が発生していたが、交信かく乱剤と薬剤散布を併用することにより高い防 除効果が得られた。 ○コンフューザーNの小面積処理現地実証試験 県内のスモモ産地の5~20aの 16 圃場においてコンフューザーNを処理した。防 除は園主慣行とし、第2世代幼虫の被害が見られる7月上旬、第3世代幼虫の被害が 見られる8月上旬に被害果の調査を行った。また、試験を依頼した 16 圃場の園主に コンフューザーNの被害抑制効果、今後の利用についてアンケート調査を行った。 現地の慣行防除に併用し、コンフューザーNを処理することにより8月上旬収穫の 中晩生種では被害抑制効果が認められた(データ略)。8月下旬収穫の晩生種では被 害がやや多かったが慣行防除に比較し被害抑制効果があるという回答が得られた。現 地実証を行った園主へのアンケート調査のまとめでは、効果があり、今後も利用した いとう評価が得られた(図5)。 表1 コンフューザーN処理下におけるフェロモントラップへの スモモヒメシンクイガの誘引阻害 試験場所 処理 総誘殺数/トラップ 面積 コン N 処理区 誘引阻 害率(%) 1) 無処理区 南アルプス市 1ha 4 491 99.2 果樹試 10a 5 292 98.3 1)誘引阻害率は次式より算出 誘引阻害率={1-(処理区誘殺/無処理誘殺)}*100 表2 スモモヒメシンクイに対するコンフューザーNの防除効果 調査圃場 調査樹 調査果数 コンフューザーN処理区 a 682 5.1 (10a) b 855 2.4 品種:太陽 c 645 2.9 a 363 29.8 (6a) b 189 55.6 品種:ソルダム c 381 29.1 無散布樹 415 75.4 無 処 理 区 被害果率 (%) *薬剤散布日:6月 29 日、7月8日、7月 19 日 *薬剤散布:aモスピラン水溶剤、bバリアード顆粒水和剤、cスカウトフロアブル *調査日:7月 26、27 日。 不明 ない 利用しない かなりある ある 利用したい 今後の利用について 効果について 図5 コンフューザーN小面積処理現地実証試験アンケート結果(16 戸) 試験圃場:圃場数 16 甲州市8 南アルプス市5 笛吹市3 品種:ソルダム、貴陽、太陽 面積:5~20a 栽培:棚栽培7、立木栽培9 環境:平地9、山付き7 おわりに 本稿のタイトルは、スモモヒメシンクイに対する交信かく乱剤を活用した減農薬防 除としたいところであるが、まだ減農薬という段階には至っていない。現実は、とに かく利用できる防除法を活用し、本種の被害を出来るだけ少なくしたいというのが生 産者の要望である。慣行防除と併用という形ではあるが、交信かく乱剤を使用するこ とにより被害の軽減が実感され、処理面積は拡大している。交信かく乱剤の効果は、 害虫の密度が低い場合により高くなる。毎年の防除を確実に行い、発生密度を低下さ せることにより交信かく乱剤の効果および総合的な防除対策もよい方向に向かって いくことを願う。交信かく乱剤の試験は、現地の圃場を使用し試験規模も大きい。試 験に当たってご協力を賜った生産者、JA職員およびメーカーの方々に厚くお礼申し 上げる。 ホウレンソウケナガコナダニの簡易モニタリング装置 (コナダニ見張番)の開発と被害予測による防除体系 山口県農林総合技術センター 本田善之 1.はじめに 山口県の中山間地域の主要野菜であるホウレンソウでは、近年ホウレンソウケナ ガコナダニ(以下コナダニと略す)の被害が拡大し、生産振興上の重要課題となっ ている。しかし、コナダニは土壌中に生息し、発生確認が困難で防除判断ができな かった。よって簡易なモニタリング方法の開発により的確な防除判断技術と効率的 防除を可能にする必要がある。 2.コナダニの被害の特徴 コナダニの被害は、ホウレンソウの新芽に成虫が移動・加害し、葉に小さな穴や コブ状の少突起が発生し、正常に展開しなくなる。発生密度が高い場合は新芽が褐 変し、生育が止まってしまうこともある。被害発生時期は周年栽培のホウレンソウ において、2月~6月、9月~11月で、夏期には発生しない。播種時~2葉期ま ではほとんど被害がないが、4葉期頃に成虫が侵入し、侵入後の展開葉は加害を受 けるが、葉の被害を確認してからの防除は手遅れとなることが多い。コナダニやそ の被害の特徴としては、以下のようなものが上げられる ・コナダニの被害は雨除け栽培で多く、露地では少ない。 ・有機質資材として、未熟堆肥やなたね油粕を投入すると被害が増大する。 ・コナダニはホウレンソウ以外のコナツナやレタスで飼育可能だが、被害が顕著 なのはホウレンソウのみである。 ・油粕などを放置しておくと、通常はケナガコナダニが発生するが、雨除けホウ レンソウ圃場ではホウレンソウケナガコナダニが優占的に発生する。 ・コナダニは高湿度を好み、低温に強く高温に弱い。 ・効果的な薬剤が少ない。 3.既存の調査方法とコナダニ見張番の開発・構造 こうしたコナダニの発生をモニタリングするため、千葉大学がコナダニを簡易に モニタリングするコナダニトラップ(乾燥酵母を利用)を開発している。山口県は それを改良し、灌水に影響されず安定した捕獲が可能なトラップ(コナダニ見張番) を開発し特許を取得した。「コナダニ見張番」は、上部シート(断熱性、防水性を 備えた素材)、下部シート(防水性を備えた素材)、誘引シート(黒画用紙等に両面 テープで乾燥酵母等のコナダニの餌を付着)で構成される。この構造により、上部 シートや下部シートの土壌接地面に は水滴が付くが、誘引シートは紙が水 分を吸収し餌周辺は高湿度となる。土 壌表面と「コナダニ見張番」の温湿度 を比較すると、コナダニの好む75% 以上の湿度割合が高く、コナダニの嫌 う高温にもなりにくいことがわかっ ている。コナダニ類の好む高湿度の空 間と、餌の両方で誘引する新しい発想 図1 のトラップである(図1)。 「コナダニ見張番」の構造 4.既存トラップとの精度確認 従来の「コナダニトラップ」(千葉大・春日氏開発)と「コナダニ見張番」によ るコナダニの発生確認率は、共に 25.6%で両方のトラップでコナダニ発生確認の 結果が一致する割合は約 95%であった。また、 「コナダニ見張番」は「コナダニト ラップ」に比べ土壌が多湿条件の時にも捕獲数が多く、「コナダニトラップ」と同 等以上にコナダニの発生状況を確認できた(表)。 表 コナダニ見張番とコナダニトラップ(春日式)のコナダニ捕獲状況 項目 灌水や灌水や雨の降り込みで土壌 が多湿条件の場合 多湿条件の場合 コナダニトラップ コナダニ見張番 (春日式) コナダニ見張番 5 調査点数 合計 土壌表面が乾燥条件の場合 コナダニトラップ (春日式) コナダニ見張番 38 コナダニトラップ 両トラップの一致 (春日式) 率(%) 43 平均コナダニ捕獲数 (頭/トラップ) 21.5a* 4.4b* 0.6a 0.3a 2.8a 0.7a - 発生確認率(%) 18.4 18.4 80.0 80.0 25.6(11/43) 25.6(11/43) 95.3 注1: 2005年6月29日~11月9日に周南市の4農家の6ほ場(雨除けハウス)において43回(月の上旬と下旬)調査を実施した。1回の調査で各トラッ プは3個設置し、その平均を求めて比較した。 注2:発生確認率=(コナダニの発生を確認した調査地点)/(全調査地点)×100 注3:両トラップの一致率=(両トラップでコナダ二の発生の有無が一致した調査点数)/(全調査点数)×100 *異なるアルファベット小文字はt検定及びフリードマン検定(ノンパラメトリック検定)で2者間に有意差あり(*p<0.05)。 5.コナダニ見張番の予測精度(播種後) 開発された「コナダニ見張番」のホウレンソウ被害との関係を調べた。試験場所 は山口県周南市のホウレンソウ農家のハウス、品種はトラッド、播種は、平成18 年2月28日、栽植距離:畝幅 15 ㎝×株間 10 ㎝ 、栽培管理は一般慣行によった。 試験区は1区 7.5 ㎡ (5×1.5m)500株2連制で行った。各区は無処理と粒剤2 種類を播種時に散布、散布剤3種類を2葉期、4葉期に散布してコナダニの密度が ばらつくような区を12区設置し、各区のハウス左右2地点で、 「 コナダニ見張番」、 コナダニトラップ、ツルグレン抽出によりコナダニ密度を計測した。 その結果、トラップ調査 1 週間後の被害度との相関は、「コナダニ見張番」>コ ナダニトラップ>ツルグレン抽出の順であった。調査時期別に 1 週間後の被害度と の相関を調べた結果、回帰式の傾きは本葉2葉時<本葉4葉時<本葉6葉時と大き くなり、生育後半になり密度が増加すると被害が加速する傾向が見られた(図2)。 図2 各種トラップのコナダニ捕獲数と 1 週間後の被害度との相関 6.コナダニ見張番の予測精度(播種前) 播種後の被害と「コナダニ見張番」の相関が高いことは確認できたが、コナダニ の発生が多いと播種後の薬剤散布だけではコナダニ被害を抑えきれない。従来から コナダニに対して効果が高いとされている、播種前の土壌消毒+2 回の散布防除を 実施するには播種前の被害予測が不可欠となる。 そこで、「コナダニ見張番」を前作収穫時に設置して、土壌消毒の必要性が判断 できるか、現地試験を実施した。試験は山口県周南市及び阿武町のホウレンソウ農 家の、合計 25 ホウレンソウハウスで行った。平成 21 年 9 月~10 月に実施した。 収穫時に「コナダニ見張番」を 1 ハウス 10 器設置し、1日設置後に捕獲された合 計コナダニ数を算出した。次作の被害程度(多発生:ハウス全体的に被害発生、中 発生:ハウス半分程度に被害発生、小発生:ハウスの一部で被害発生、なし:被害 発生が見られなかった。)、防除歴を聞き取り調査した。 その結果、「コナダニ見張番」で捕獲された頭数を5頭以上とそれ以下に分けた 場合、5頭に満たない場合は無防除でも被害の発生割合が少なかった。また、5頭 以上でも3回の防除を実施していれば、小発生以下となる場合が多かった(図3)。 図3 前作収穫時のコナダニ見張番の捕獲頭数と次作の被害程度(防除回数別) 7.コナダニ見張番の使い方と被害予測による防除体系 「コナダニ見張番」は播種前、播種後も防除判断可能であることを明らかにした が、ホウレンソウ農家は複数のホウレンソウハウスを周年で栽培しており、それら の作型すべてに「コナダニ見張番」を用いて防除判断するのは大変な作業となる。 そこで、コナダニの被害発生の特徴などから利用方法を整理した。 <年間・作型での使い方> コナダニが発生し始める2~3月又は9~10 月の収穫期に、 「コナダニ見張 番」を 1 ハウス当たり 10 個、1日間以上設置して、1ハウスで5頭/10個 以上のコナダニが確認された場合は、次作播種前の土壌消毒を実施する。防除 実施後も2葉期と4葉期を目安に「コナダニ見張番」を設置し、コナダニの発 生が確認されれば、直ちに追加防除を行う。その後の作型で、前作で被害が発 生したハウスでは土壌消毒+2 回防除を徹底し、発生がなかったハウスでは播 種前に「コナダニ見張番」を設置して、同様の防除判断を行う(図4)。 <発生状況に応じた使い方> ○被害はほとんど無いが、突発的に被害が発生することがある。あるいは、発 生は頻繁にあるが、どのハウスに発生するか不明な場合 2~3月又は9~10 月の収穫期に「コナダニ見張番」で被害予測を実施 する。上記の年間・作型での使い方に準ずる。 ○毎年、すべてのハウスにもある程度発生する場合 2~3月又は9~10 月の作型では必ず播種前の土壌消毒を実施する。播種 後は2葉期と4葉期を目安に「コナダニ見張番」を設置し、コナダニの発生が 確認されれば、直ちに追加防除を行う。 ○毎年、時期になると多発生するハウス 2~3月又は9~10 月の作型では必ず播種前の土壌消毒++2 回防除を徹 底する。 図4 「コナダニ見張番」による防除判断の方法(まとめ) 8.残された問題点 「コナダニ見張番」は、コナダニの防除判断に有効なツールとして、商品化を目 指しているが、コストの問題、同様品の精度の問題、設置場所、設置時期などのば らつきの問題、設置場所の土壌湿度による変動の問題等、汎用的に高い精度を求め るが故に超えなければならない問題点も残っている。 モニタリングの問題以外にも、困難と言われるコナダニ問題の解決のため、京都 大学を総括として平成22年度から「新たな農林水産政策を推進する実用技術開発 事業」で「環境保全型農業と両立する生物的相互関係を活用した難防除コナダニ類 新管理体系の確立」が取り組まれ、今までの土づくり技術及び有機質資材の管理を、 コナダニ等害虫面から見直す研究が進んでいる。複雑な土壌環境に発生するコナダ ニの増殖と有機質資材の関連性を軸に、物質面(京大)、微生物面(岐阜、広島)、 水分や物理面(奈良、山口)、天敵(京大、北海道)、モニタリング(サンケイ化学 ㈱、山口)、化学防除の影響(北海道、岐阜、奈良)などの各機関で多面的な試験 が行われている。今後の研究成果に期待したい。 スワルスキーカブリダニに対する農薬の影響 宮城県農業・園芸総合研究所 園芸環境部 宮田將秀 はじめに コナジラミ類やアザミウマ類などに対する天敵であるスワルスキーカブリダニ ( Amblyseius swirskii )は、2008 年 11 月に農薬登録されて以来、主にキュウリや ピーマン、ナスなどで高い防除効果が確認されている(宮田ら、2009;森田ら、2009; 岡崎ら、2010;柴尾ら、2009)。スワルスキーカブリダニを使った防除体系では、病 害のほか、アブラムシ類やハダニ類、チョウ目害虫など、本天敵の対象外となる病害 虫が発生した場合、薬剤散布などの防除を行う必要がある。その場合、スワルスキー カブリダニに悪影響のない薬剤を選ぶことが望ましいが、本天敵に対する薬剤の影響 についての知見はまだ少ない(柏尾、2009;桃下・山中、2008)。そこで、数種の薬 剤について本天敵への影響をナス圃場などで確認した。 薬剤(散布剤)の影響について 2008 年から 2010 年にかけて、スワルスキーカブリダニをあらかじめ放飼した施設 栽培のナス圃場で各薬剤を散布し、散布前と散布後の密度を調査した。供試薬剤のう ちスワルスキーカブリダニの密度に顕著な悪影響が認められた剤は、ピレスロイド系 殺虫剤のアクリナトリン、ビフェントリン、有機りん系殺虫剤のアセフェート、殺ダ ニ剤のミルベメクチン、ビフェナゼート、その他の系統であるスピノサドで、殺菌剤 ではマンゼブ、キノキサリン、プロピネブ、キャプタン+ホセチル(図 1)であった。 やや悪影響が認められた剤は、ネオニコチノイド系殺虫剤のアセタミプリド、殺菌剤 のポリオキシン複合体、TPN で、昆虫成長制御剤のノバルロンは散布 10 日後から密 度の低下が認められた(図 1)。散布直後に密度が一時的に低下した剤はイミダクロ プリド、チアメトキサム、エマメクチン安息香酸塩であった(図 1)。その他、ネオ ニコチノイド系殺虫剤のジノテフラン、殺ダニ剤のシフルメトフェン、シエノピラフ ェン、アセキノシル、昆虫成長制御剤のルフェヌロン、その他の系統のフロニカミド、 殺菌剤のクレソキシムメチル、ボスカリド、キャプタン、ジエトフェンカルブ+チオ ファネートメチル、トリフルミゾールについては、ほとんど影響はないと考えられた。 例えばイミダクロプリドやチアメトキサムなどでは、スワルスキーカブリダニ密度の 急減は認められなかったものの、増加することもなかった。さらに、ノバルロンでは、 その影響が散布 10 日後に観察されたが、これらの剤は卵の生存や産卵数に影響を及 ぼした可能性もある。今後、スワルスキーカブリダニの各ステージ毎に影響を精査す る必要があるだろう。 図1 スワルスキーカブリダニに対する薬剤の影響 (上図:2008 年、中図:2009 年、下図:2010 年) 散布剤の影響期間について 天敵利用の失敗の原因の一つとして、天敵に影響のある薬剤を散布し、その影響が 残っているうちに天敵を放飼してしまう事例がある。天敵放飼前の薬剤散布は盲点と なっている。そこで、散布剤(殺虫剤 1 剤と殺菌剤 3 剤)についてスワルスキーカブ リダニに対する影響期間を把握するための試験を実施した。 試験は施設栽培ナス圃場で実施した。9 月 16 日に各薬剤を散布し、その後、約 1 週間間隔でスワルスキーカブリダニを放飼した。調査は各放飼の 6 日後に、全葉に存 在するスワルスキーカブリダニを計数した。その結果、ポリカーバメートおよびポリ オキシンが悪影響を与える期間は 3 週間程度、マンゼブでは 4 週間程度、トルフェン ピラドでは 5 週間程度であった(図 2)。 図2 スワルスキーカブリダニに対する薬剤(散布剤)の影響期間 (*は調査なし。グラフは各放飼からそれぞれ 6 日後の値を示す。) 粒剤の影響期間について さらに、粒剤についての試験をポット植えのパプリカで実施した。1/5000a ワグネ ルポットに各粒剤を処理し、直ちにパプリカを定植、その後、約 1 週間間隔でスワル スキーカブリダニを放飼した。調査は散布剤の試験と同様である。その結果、イミダ クロプリド、アセタミプリド、ニテンピラム、チアメトキサム、クロチアニジンの各 粒剤の影響期間はおおむね 1 週間程度、ホスチアゼート、イミシアホス、カルボスル ファン、ジノテフランは 2 週間程度、ベンフラカルブは 4 週間程度、アセフェートで 6 週間程度であった(図 3)。 図3 スワルスキーカブリダニに対する薬剤(粒剤)の影響期間 (凡例は粒剤処理からの放飼の時期を示し、グラフは各放飼からそれぞれ 6 日後の値を示す。) 天敵と各薬剤の併用の考え方について 天敵を組み入れた防除体系では、その天敵の捕食(または寄生)対象外の病害虫に 対して薬剤で防除する場合、天敵に悪影響がない剤を選ぶということは言うまでもな い。しかし、天敵への影響の有無は一概に割り切れるものではなく、今回の試験結果 のように、悪影響が顕著な剤とほとんどない剤のほか、影響はあるものの、その程度 が低い剤もある。さらに悪影響が顕著であっても、その影響期間が長い剤と短い剤が ある。一方、薬剤はその対象病害虫に防除効果が高いものと、抵抗性害虫や耐性菌の 出現により効果が低下しているもの、さらにその中間的なものとに分けられる。その 中で天敵に影響がなく、対象害虫に効果が高い薬剤がもっとも使いやすいが、現状で はそのような薬剤は限られている。そのため、その他に分類される薬剤も状況に応じ て組み合わせざるを得ない。図 4 に薬剤を大まかに分類し、それぞれに分類された薬 剤がどのような場面で使用できるかを示した。A に分類される薬剤は栽培期間をとお していつでも使える剤である。しかし、このような剤はなるべく温存するということ を前提に考えると、害虫密度が急増した場合に限るべきであろう。天敵への影響が強 いものの、害虫に対する防除効果が高い C、C´に分類される薬剤は、場合によって は使わざるを得ない場面がある。つまり、C の薬剤は天敵への影響期間が短いため、 天敵放飼前に害虫密度を下げる場面、C´は栽培終期に、最後の散布と見込まれるタ イミングでの使用場面が想定される。次に D の薬剤であるが、天敵の放飼後、害虫 密度が増加し始める場合での使用場面が想定される。さらに B と E の薬剤であるが、 天敵が十分に圃場に定着し、かつその天敵が対象とする害虫密度がよく抑えられてい る場合で、その天敵の対象外の病害虫が急増した場面では B の使用を、その病害虫 の増加初期にはEの使用が想定される。 登録農薬をA~Fに当てはめてみる 天敵に 対して 対象病害虫に対して 効果が高い 効果が低い 影響がない A D やや影響がある B E 顕著な影響 がある C 天敵放飼 ↓ ↓ 残効が短い C 残効が長い C´ D ↓ A ↓ F E ↓ B ↓ C´ ↓ 天敵の対象外 害虫の発生 天敵放飼前に 害虫密度を下 げる。 害虫密度の増加 初期なら、Aを温 存して。 害虫密度が急増 してしまったらAを。 栽培終期ならば 天敵に影響が長 い剤で逃げ切り もあり。 天敵が十分に定着し、かつ天敵の対象害虫 の密度が低いことが前提で・・・ 天敵の対象外害虫の増加初期ならEを、 急増してしまったらBを。 図4 天敵と薬剤の併用についての考え方 おわりに 今回、スワルスキーカブリダニに対して数種の薬剤の影響について検討した。その 影響は「影響有り」と「影響無し」の二分割に割り切れるものではなく、剤によって 影響の程度や現れ方に違いが認められた。これは生育ステージによって影響の強弱が 違う可能性もあり、引き続き精査する必要がある。また、影響期間についてはさらに 他の薬剤についても把握する必要がある。天敵と薬剤の併用にあたって多くの知見が 蓄積され、より効果的、効率的なスワルスキーカブリダニと薬剤の併用体系が構築さ れることを望む。 引用文献 1) 柏尾具俊(2009):九州病虫研報 55:194(講要) 2) 宮田將秀ら(2009):第 53 回日本応用動物昆虫学会大会講要:65 3) 桃下光敏・山中聡(2008):第 52 回日本応用動物昆虫学会大会講要:12 4) 森田茂樹ら(2009):第 53 回日本応用動物昆虫学会講要:129 5) 岡崎真一郎ら(2010):第 54 回日本応用動物昆虫学会大会講要:81 6) 柴尾学ら(2009):関西病虫研報 51:1~3 ナシ黒星病の減農薬防除の取り組み 千葉県農林総合研究センター 病理昆虫研究室 金子洋平 1.はじめに 千葉県におけるニホンナシの収穫量は約 40,200t(平成 21 年)で全国第一位となって いる。主要品種は「幸水」 、 「豊水」であり、最も重要な地上部病害はナシ黒星病である。 一般にナシ栽培では減農薬防除は困難とされている。この理由としては、減農薬防除は 後述するような知識・技術が必要であり、労力もかかる。また、宿主となるナシが長期間 同じ場所に存在するため、病原菌も園内に定着しており、当年の防除の失敗が翌年にも影 響を与える等のリスクも大きい。このことから、通常は、ナシ防除指針(防除暦)に基づ いてスケジュール的な薬剤防除が行われている。 一方、近年は環境にやさしい農業の推進、安全・安心な農産物の提供の観点から、減農 薬が求められている。また、本県では、環境にやさしい農業を進めながら、慣行と比べて 農薬や化学肥料の使用を半分以下に減らした「ちばエコ農業」と呼ばれる千葉県独自の認 証制度による減農薬防除の取り組みが行われている。 ここでは、ナシ黒星病の発生生態、その減農薬防除を述べ、次にパソコン上で黒星病の 防除要否を判断できる「梨病害防除ナビゲーション」を解説し、これを用いて地域ぐるみ で「ちばエコ農業」に取り組んでいる事例を主に病害防除の面から紹介する。 2.ナシ黒星病の発生生態および防除 (1)伝染源(胞子の飛散開始時期) ナシ黒星病の第一次伝染源は、前年の被害落葉上に形成される子のう胞子と、罹病芽基 部上に形成される分生子である。子のう胞子の飛散は、3月下旬から4月上旬に始まり、 5月上旬から下旬に終息する。子のう胞子の飛散はナシの開花し始める時期と関係があり、 開花が早いほど、飛散も早い時期から始まる。一方、罹病芽基部上に形成される分生子も 開花始め頃から飛散が始まる。これ以降は、両胞子によって罹病した葉、幼果上に形成さ れる分生子が再び周囲に飛散し、二次伝染するため、生育期間中は常に黒星病の胞子が飛 散している可能性がある。 (2)ナシの黒星病に対する感受性 感受性とは、植物の持つ、病気に罹りやすい性質のことである。ナシは生育期間中は常 に感受性があり、防除が必要である。その中でも、 「幸水」 、 「豊水」の幼果は開花直後から 5月上旬頃までは感受性が高く、さらに、 「幸水」では6月中旬から7月中旬にも果実の感 受性が高まる期間がある。両期間は黒星病防除において重要な防除期間である。 (3)感染に好適な環境条件 ナシ黒星病菌の胞子が葉に付着した後、葉の組織に感染するためには、一定の温度条件 下において、 濡れた状態が継続する必要がある。 黒星病の感染好適温度は 15~25℃であり、 この条件下では9時間以上濡れた状態が継続すると感染に至る(図1)。また、5~30℃で も 12 時間以上濡れた状態が継続すると、感染する。以上のように、黒星病の感染・発病に 関する環境要因は温度と濡れ時間であり、これらの関係を基に、気象観測装置等を用いて 温度と濡れ時間を計測し、感染危険の度合いを計算により求めることもできる。また、こ の計算により算出された値を感染危険度とする。 80 5℃ 10℃ 15℃ 20℃ 25℃ 30℃ 発 病 60 度 40 20 0 0 10 20 30 40 濡れ継続時間(時間) 図1 温度、濡れ継続時間と黒星病発病度の関係 (4)ナシ黒星病の感染・発病の条件 一般に病気の三要因(主因、素因、誘因)が揃った時に病気は感染・発病する。ナシと 黒星病との関係の場合で述べると、感受性があるナシ(素因)において、胞子(主因)の 飛散が開始した状態で、感染危険度(誘因)がある時に、黒星病は感染・発病に至る。逆 に、これらの要因が一つでも欠ける時、あるいは各要因が揃った場合でも、その時に薬剤 による保護効果がある場合には、感染・発病は抑制される。 (5)ナシ黒星病防除の減農薬方法 慣行防除体系では防除暦にしたがって、農薬による効果がほぼ途切れることのないよう にスケジュール的に殺菌剤が散布される。減農薬栽培をする場合、まず、①被害落葉の処 理や罹病芽基部、罹病葉の切除等、耕種的防除を徹底する。次に、②ナシの感受性が高い 重要な防除期間には防除暦に従って確実に防除を行う。③その他の時期において、前述の 各要因が揃う時には薬剤の保護効果がある状態にすることが必要である。すなわち、図2 のように散布を行った後、その残効期間中は降雨があっても次の散布はしない。また、雨 が降らなければ残効期間が切れていても、次の散布はせず、その次の降雨直前に散布を行 う(図2)。これらにより、スケジュール散布に比べて減農薬できる。 ①散布を行った後、 その残効期間中は 降雨があっても散布 はしない ②雨が降らなけれ ば、残効期間が切 れていても、次の 散布はしない 散布A ③その次に、 雨の降る直 前に散布を 行う 雨 散布B 残効期間 残効期間 図2 減農薬防除における散布間隔の延長 3.「梨病害防除ナビゲーション」 これまで述べてきたとおり、黒星病の減農薬防除を行うためには、黒星病の発生生態を理解し、 気象条件やナシの感受性、薬剤の効果・残効期間等を総合的に評価し防除要否を判断する必要 がある。しかし、これは専門的な知識だ けでなく、長年の経験・勘といった曖昧 各ナシ園 パソコン上 農薬防除を継続的に行うことのできる生 微気象データ 迅速に情報を収集 産者はごく一部に限られていた。このた め、千葉県では、パソコン上で防除の要 胞子飛散状況 否や薬剤散布のタイミングを把握できる ナシ病害防除支援情報システム「梨病 害防除ナビゲーション」を開発した(図 3)。 農薬散布歴 総合的に評価 防除要否の判断 なものに頼るところも多く、黒星病の減 図3 「梨病害防除ナビゲーション」 「梨病害防除ナビゲーション」は、「マイクロソフトエクセル」上で稼働し、ナシ開花日、気象観測 データに基づく黒星病の感染危険度、薬剤散布日を入力すると、胞子の飛散状況、黒星病感染 危険度及び予測発病度、黒星病に対する果実の高感受性期、散布農薬の効果期間の目安等が チャート化(「病害防除支援チャート」)される(図4)。 このシステムは減農薬防除を実際に現場で普及、実践する普及指導員や生産者が農薬散布 の要否や散布時期を判断するために用いられている。この他にも、ナシ園における病害発生の危 険性や、防除状況が視覚的に把握でき、また、それらをシミュレーションできることから、病害発生 や防除の成否の要因解析等にも活用できる。 黒星病感染危険度 農薬散布日と 残効期間 胞子飛散有り 子のう胞子 累積飛散曲線 潜伏期間後の 予測発病度 「豊水」幼果 高感受性期間 「幸水」幼果 高感受性期間 図4 「病害防除支援チャート」表示画面 本システムを稼働させるに当たっては、小型温湿度記録計(サーモレコーダー)で気象 データ(温度と相対湿度)を随時収集し、エクセル上で黒星病の感染危険度を算出し、 「梨 病害防除ナビゲーション」にコピーペーストして入力する。千葉県市原市のナシ生産者の グループでは、 「梨病害防除ナビゲーション」を集団で利用して地域で「ちばエコ農業」に 取り組み成果を上げている。 4.市原市の「予察隊」による減農薬防除 市原市のナシ生産者のグループでは、 「ちばエコ農業」認証基準を目指した地域の生産者 が「予察隊」を編成し、指導機関と連携して、生育期間中 10 日に一度集まって病害虫の発 生調査を行っている。この時に「梨病害防除ナビゲーション」を稼働させる。発生調査及 び「梨病害防除ナビゲーション」の結果を踏まえて、今後の防除の方針を話し合い、一連 の情報を掲示板に提示する(図5)。この取り組みには、個人レベルでは困難な病害虫の発 生状況を地域で把握し、防除の目的意識を共有できる等のメリットがある。2008 年~2010 年にかけて、毎年複数の生産者が「ちばエコ農業」認証基準を達成している。 図5 「予察隊」の病害虫発生調査及びその結果の掲示の様子 5.終わりに これまで述べたように、ナシ栽培においても減農薬防除が求められているが、知識・技 術や多大な労力を必要とする。そこで、 「梨病害防除ナビゲーション」のように、パソコン を利用した病害防除支援情報システムや、あるいは「予察隊」のように、集団で防除の認 識を共有する取り組み事例を紹介した。 このような取り組みを通じて、減農薬防除を行うことにより、環境にやさしい農業の推 進、安全・安心な農産物の提供の一助となれば幸いである。 茶栽培におけるニーム散布剤の利用 ~ベトナムでの実践~ 濱周吾 東京農工大学 仲井まどか 【はじめに】 ベトナムにおけるニームの農業利用の実践を紹介する。ニーム(英名:Neem,学名: Azadirachta indica ,和名:インドセンダン)は、インドなど南アジアを原産地とする 樹木である。筆者(濱)は、2004 年から現在までニームを活用した茶栽培をベトナム で行ってきた。本稿では、ベトナムにおける茶栽培と害虫防除としてのニーム利用の 現状と展望を報告する。 【ベトナムにおける茶栽培の歴史】 ベトナムの茶文化は、古くから中国の影響を受け主に北部で発展してきた。ベトナ ム北西部は中国雲南省と同じく茶の原産地と考えられている。昔から茶は薬として扱 われており、今日でも高さ十数メートルに育った巨木茶樹から春に収穫される茶は薬 として珍重される。 1950 年頃からベトナムでは、先住民である H.Muong 族や Thai 族等の民族の住む山 岳部へ、政治的に主権を握る民族である Kinh 族(京族、別名:越族)の入植が行わ れてきた。この入植を円滑に進めるべく、収入源となる茶園の開発が全国的に行われ た。現在では大規模なプランテーションが建設され、緑茶や紅茶を輸出しており現在 では世界7位の茶生産国になった(FAOSTAT,2008)。 【ベトナムにおける茶栽培の現状と問題点】 ベトナムでは、緑茶として飲まれる 在来の雪山茶(Shan Tuyet)(図 1)や TP14 などの紅茶品種(アッサム品種) の栽培が政府から推奨されていたが、 最近では渋みが多いため敬遠され、そ のかわりに商品価値が高い台湾品種 とベトナム在来品種との交配品種や クーロン品種への移行が進んでいる。 これらの品種は、上述の在来品種に比 べ肥料要求性が高く、また病害虫に弱 い。そのため化学合成農薬の散布が必 要になり、農薬使用が広まった。不思 議なことに、これまで化学合成農薬散 図1 Shan Tuyet Tea の収穫(Lai Chau 省). 布の必要がなかった在来品種の茶園 でも殺虫剤の使用なしには茶が収穫 できない状態になっている。考えられる原因は、侵入害虫の影響であり、また天敵昆 虫の減少など生態系のバランスが崩れている可能性も考えられる。 現在の一般的なベトナム緑茶の中東諸国向け輸出価格は 1.5 から 2.5 USD/kg であ り、この価格では物価上昇を受けて生産原価が高騰している現在では、十分利益が得 られない。ベトナム茶が国際市場から評価されない理由の一つは、農薬管理の不徹底 であると考えられる。農薬の管理は Viet GAP の奨励などを通して政府も支援を続け ているが、管理徹底は未だ成果を上げているとは言われていない。 【世界におけるニーム利用の現状】 そのような状況の中、著者(濱)は、 2004 年から日系企業 Vina Suzuki Tea Co., Ltd において減農薬による高級 茶生産プロジェクトに取り組んだ(図 2)。その結果、化学合成農薬散布を 段階的に減らすことで 2 年後に 124 ヘ クタールの栽培面積のほぼ全域につ いて化学合成農薬に頼らない栽培を 実現した。この減農薬茶栽培を実現で きたのは、ベトナムに自生するニーム を利用した事による(図3)。ニームは、 熱帯地域に自生し、乾燥に強いため緑 化樹や薪材としても利用さ、アジア、 図2 Vina Suzuki Tea Co., Ltd.の茶園. アフリカ、中南米等にも広く分布して いる。ニームはインドやスリランカなどの南アジア地域で古くから医療に利用されて い る が 、 主 成 分 Azadirachtin が 昆 虫 に 対 し て 摂 食 阻 害 や 脱 皮 阻 害 能 を 示 す (Schmutterer,1990)。ニーム散布剤とは、ニームの種子からの抽出物やニームオイ ルをもとに製剤化した害虫防除資材である。インドやタイ、アフリカ諸国などでニー ムによる植物保護を推進する国は少なくない。日本では、まだ農薬登録を取得したニ ーム散布剤はないが、インド、ドイツ、米国などで公的機関の認可を受けて販売され ており、ドイツでは有機栽培における使用も認可されている。表1に、米国 EPA の認 可を受け販売されているニーム散布剤の一例を示す。 表1 米国で農薬登録されているニーム散布剤の一例 【ベトナムにおけるニーム生産】 ベトナムでは、中南部東海岸に 広がる乾燥地域のユーカリに代 替される緑化樹として 1990 年頃 からニームの植林が始められた。 ベトナムの研究者によって西ア フリカのセネガルから輸入され たニーム種子から苗が生産され、 これまでに Ninh Thoan 省および Binh Thoan 省を中心に 10,000ha 図3 ニーム果実(左) 及び種子核(右). に定植され、現在も定植は続けら れている。 しかしながらニーム種子は、海外からの需要がある程度あるものの国内需要は少な く、多くが収穫されず放置されている。ニーム種子がベトナム国内で認知され市場で 取引されるようになれば、これらの乾燥地域に住む人々にとって重要な収入源になる に違いない。 【日系茶企業における減農薬の取り組み】 Vina Suzuki Tea Co.,Ltd.の茶 園 は 南 部 高 原 Lam Dong 省 の 標 高 1000m という高地にあり、気候はモ ンスーンで雨季(5 月~10 月)と乾 期(11 月から4月)がある。栽培 品種は金萱や四季春といった台湾 ウーロン茶品種であった。当時この 茶園では、ダニ、アブラムシ、アザ ミウマ、ハマキガ、ヨコバイ、Tea Mosquito Bug ( 学 名 : Helopeltis theivora ,半翅目)といった害虫が 問題になっていた。 ニーム種子を利用した減農薬の 試 み は 2005 年 か ら 始 め ら れ た 。 Lam Dong 省 の 東 に 位 置 す る Ninh Thoan 省で生産されるニーム種子を 原料に自社で散布剤を調整した。当 時ベトナムではニーム由来の殺虫 剤が登録、販売されていなかったた め、散布剤を独白に開発する以外に 方法はなかった。ニーム由来散布剤 の開発方針として、安価原料で容易 な方法を目指した。その結果、米や 米糠など安価で手に入る原料を利 用した方法を確立した。 2005 牛 3 月から開始したニーム 散布剤によって、ダニ、アブラムシ、 アザミウマなどには効果が高くこ 図4 化学合成農薬散布延べ面積(ha/月)の推移. 図5 Tea Mosquito Bug( Helopeltis theivora ). れらの防除に使用していた化学合成農薬を使用する必要がなくなった。2005 年(1 年目)は、ニーム種子に米から製造した含水エタノール溶液を添加し抽出した散布剤 を使用したが、ハマキガやヨコバイには効果がなく、化学合成農薬の散布を継続した。 2006 年(2年目)には、このニーム散布剤に酵素分解米糠の水抽出物を加え、乳化 性と展着性を改善した。その結果、ハマキガやヨコバイにも効果が認められた。 2005 年から 2007 年までの農薬散布面積の推移を図4に示す。グラフ縦軸は月毎の農薬散 布延べ面積を示し、農薬成分は色分けした。 2007 年にはほぼ農薬を使用しない防除 に移行したことが示されている。一方で、全くニーム散布剤による防除効果が見られ ない害虫もあった。その一つが Tea Mosquito Bug である(図5)。これは台湾以南に 生息する茶園重要害虫の一つである。この害虫に関しては化学合成農薬(ジノテフラ ン剤)によって防除したが、Tea Mosquito Bug が残す若葉吸汁痕をたよりに化学合 成農薬をごく少量スポット散布する工夫により、その農薬使用量を軽減した。このこ とによって、2006 年末からは化学合成殺虫剤を全面に散布する必要がなくなり、茶 園生態系のかく乱を抑えるシステムが確立した。 【ベトナムにおけるニーム散布剤登録】 ベトナムでは、この数年の間にニームに含まれる活性成分である Azadirachtin を 主成分とする殺虫剤が多数登録されている。 2010 年の農薬登録に関する資料によれ ば22製品がニーム散布剤として登録されており、化学合成農薬との混合剤も6製品 が登録されている(表2)。しかしながら、どの程度市場に受け入れられているかは 不明である。 表2 ベトナムにおける農薬登録された Azadirachtin を主成分とする製品の一例 【ベトナム北西部における実践例】 著者(濱)は、Vina Suzuki Tea Co., Ltd. の経験を生かして、北西部の茶園でもニー ム散布剤を用いた茶栽培を行った。ベトナ ム最北西端の Lai Chau 省は茶の原産地と 呼ばれる中国雲南省に隣接しており、樹齢 数百年の茶樹が点在していることでも有 名である。 省都 Lai Chau 市から南方 60km に位置す る Than Uyen 郡に製茶会社 Than Uyen Tea JSC.がある。標高 1000m に位置するこの元 図6 Lai Chau 省 Than Uyen 郡の茶園. 国営製茶会社は、契約面積を含めると 1100ha の茶園を管理し、収穫される茶葉から緑 茶を生産し、主に中東諸国に輸出している(図6)。 この茶園において、2009 年から現在までニーム散布剤のみを防除資材として使用 する試験を行った。その結果、2010 年には毎年春に発生する主要害虫ヒメヨコバイ による被害が、ニーム散布剤により周辺の農薬散布区と同等まで減少した。そのため、 この茶園では 2011 年からニーム散布剤のみで防除する栽培区の面積を拡大した。 この茶園で観察した特筆すべき点は、捕食性天敵であるクモ類の数が周辺の茶園に 比べて多いことである。一般に、ニーム散布剤はクモの生育に悪影響を与えないと言 われているが、2010 年の筆者らの調査によっても、クモの種類と個体数が多くなる 傾向が確認された。このことから、ニーム散布剤による防除効果には直接的な害虫抑 制に留まらず、天敵による密度制御効果が化学合成農薬に比べて高くなる可能性があ る。 【ニーム散布剤と天敵利用との組み合わせ】 ニーム散布剤による防除によって 土着天敵による害虫抑制効果が促進 される例は、野菜栽培でも見られた。 著者(濱)は、南部の先進集約農業地 帯である Lam Dong 省 Dong Duong 郡の パプリカ圃場でも、ニーム散布剤を使 用した栽培を指導した。この地域は、 標高が高いためベトナム有数の集約 的栽培地域で農薬の使用量が極めて 高いところである。この地域の慣行の 化学合成農薬防除圃場では害虫の抵 抗性の発達やリサージェンス等が問 図7 確認されたコヒメカメムシの成虫. 題になっており、天敵の密度も極めて 低い。しかし、ニーム散布剤を使用し た実験圃場には、慣行の化学合成農薬散布圃場では見られないコヒメハナカメムシの 存在が確認された(図7)。この圃場では、アブラムシやダニの密度も低く、アザミ ウマによる被害果実率が低く抑えられることが確認された。アブラムシやダニなどは、 ニーム散布剤に感受性が高く散布による直接的効果があったと考えられるが、アザミ ウマの場合、花の内部に生息するためニーム散布剤が届かない。しかし、殺虫スペク トラムの広い化学合成農薬をやめたことにより天敵(コヒメハナカメムシなど)が回 復し、これらの害虫の密度が低下した可能性がある。このニーム散布区では、若齢の 若虫から成虫までさまざまなステージのコヒメハマカメムシが観察された。今後、ど のようなしくみで天敵の回復がみられたのか調査する必要がある。土着天敵保護によ る生物的防除は、殺虫スペクトラムの広い化学合成農薬の使用を避け、選択性の高い 殺虫剤を使用して天敵を保護する(大野,2009)。ニーム散布剤を土着天敵保護によ る防除体系に組み込むことが可能なのか、さらなる調査研究が必要である。 【ベトナムにおけるニーム利用の課題と展望】 近年、益々安全な農産物生産の必要性が高まっている。これまで筆者らは、ニーム 散布剤について研究開発を進めてきたが、防除資材としての最も大きな問題点は、効 果が遅効性であり不安定であることと考えている。製剤の調整法を改変して安定的な 製剤を作出することが必要である。またニーム散布剤は、一般的に天敵に影響が少な いと言われているが(Schmutterer,1990)、その調査結果も少ない。さらに、散布方 法についての開発が十分に行われていないと考えられる。本稿では、土着天敵保護に よる防除体系にニーム散布剤を組み入れる可能性について述べたが、今後さらに詳細 な研究を行いニーム散布剤の天敵への直接的な影響や適切な使用方法を明らかにす る必要がある。 ニームは緑化樹として、また薬効のある有用樹としてアフリカ、中米、アジアなど 世界中の熱帯地域に広まっている。ベトナムにおいて、特に乾燥地域において栽培が 拡大している背景には、緑化の必要性と乾燥地帯住民の所得向上に寄与することが期 待されているからである。今後は、農村支援の枠組みの中で自給できる防除資材とし てニームが普及し、その利用技術体系が開発されることを望む。また、ベトナムのみ ならず熱帯地域の多くの国々で化学合成農薬に依存した農業からの脱却、乾燥地帯の 住民の所得向上、緑化促進など、ニームを利用することによる複合的な効果を期待す る。 参考文献 Schmutterer,H.(1990)Properties and potential of natural pesticide from the neem tree, Azadirachta indica,Annual Review of Entomology ,35,271-97. FAOSTAT (2008)http://faostat.fao.org/ 大野和朗 (2009)土着天敵保護による生物的防除,バイ才ロジカルコントロール ―害虫管理と天敵の生物学―(仲井まどか他 編),朝倉書店,p51- 64. 熊本県におけるトマト黄化葉巻病の防除対策 熊本県農業研究センター 生産環境研究所 樋口聡志 1 はじめに 熊本県は、トマト、スイカ、メロンおよびイチゴ等、施設果菜類の産地である。特 にトマトの作付面積は 1,150ha と全国一位であり、夏秋および冬春作ともに産地が形 成されている(農林水産省 平成 21 年産野菜生産出荷統計)。トマト栽培で最も警戒 する病害虫は,タバココナジラミ Bemisia tabaci が媒介する Tomato yellow leaf curl virus (TYLCV)を病原とするトマト黄化葉巻病である。トマト黄化葉巻病の発病 株は,新葉が葉巻症状とともに小葉化し,葉縁から黄化する。また、株全体が萎縮し 開花や結実が阻害されるため、収穫量が激減して農家の経営を圧迫する。 本県では、1999 年に有明海や八代海沿岸のトマト産地で初確認された。その後, 発生地域が拡大し,2003 年には県内全域のトマト産地で確認された。当時は栽培初 期から多発生し、トマト栽培を中止する生産者も見られた。一方、トマト黄化葉巻病 に関する調査、研究が進むとともに、取るべき対策が明らかになった。その後、対策 が指導されて普及した結果、トマト黄化葉巻病の発病株率が 20%を超えるほ場面積 は、2005 年では 2004 年に比べて半減し、2006 年では 0 になるなど、対策の効果が認 められている。本稿では,本県でこれまで取り組み、成果を収めたトマト黄化葉巻病 の防除対策を中心に紹介する。 2 TYLCV と 媒 介 虫 の 越 冬 場所はハウス内である ハウス内や地域における TYLCV の拡がり方 トマト黄化葉巻病の防除対策 を構築するためには、TYLCV の感 TYLCV 染の拡がり方や伝染環を理解す 冬 る必要がある。トマトのハウスで 秋 起こる TYLCV の感染には、一次感 染と二次感染がある。一次感染は、 春 TYLCV TYLCV 保毒したタバココナジラミが野 TYLCV 外からハウス内へ侵入し、トマト TYLCV 夏 を吸汁するときに起こる。これら の一次感染したトマト株に寄生 TYLCV することでタバココナジラミが 保毒虫となり、周囲に分散し、吸 野良生え トマト 図1 TYLCV の主要な伝染環 汁することで健全株が感染する。これが二次感染である。これらの繰り返しにより, 感染株と保毒虫は増えていく。 次に TYLCV の主要な伝染環のモデルを図1に示した。タバココナジラミやトマトは 低温に弱く,野外で越冬できない。TYLCV とタバココナジラミの越冬場所はトマトの 栽培ハウスである。気温が上がる春季の保毒虫は,ハウス内で増加して野外に飛び出 し,周辺ハウスのトマトや廃棄された果実などが生育したハウス周辺の野良生えトマ トを感染させる。その後,秋季まで TYLCV はハウスや露地のトマト,野外の野良生え トマトを行き来する保毒虫により媒介されている。このようにハウスに侵入する保毒 虫の飛来源は,周辺ハウスのトマトまたは野良生えトマトなどである。TYLCV の感染 は,トルコギキョウや雑草でも確認されているが,感染源にはなりにくい,あるいは ならない。 3 熊本県で取り組んだ防除対策 トマト黄化葉巻病の防除対策のポイントは,TYLCV の伝染環を断ち切ることである。 そのためには,ハウス内や野外の感染トマトや保毒虫を減らすことが重要である。こ れらの対策には,個別農家が取り組む対策と地域で取り組む対策がある。 (1)個別農家が取り組む対策 先に述べたとおり,ハウス内 表1 個別農家が取り組む対策 の被害は,一次感染と二次感染 ハウス開口部の防虫ネット展張 野良生えトマトの除去 により拡大していく。これを阻 止するためには,ハウス内へ保 入れない対策 近紫外線除去フィルムの展張 毒虫を入れない対策,ハウス内 光反射シートの設置 で保毒虫を増やさない対策,ハ 薬剤防除 感染株の抜き取り ウス内から野外へ保毒虫を出 さない対策が必要である。具体 増やさない対策 ハウス内の除草 粘着板の設置 的に個別農家が取り組む対策 を表1に示した。 ハウス周辺の除草 抵抗性品種 出さない対策 栽培終了時のハウス密閉処理 ハウス開口部の防虫ネット展張 1)入れない対策 ハウス内への保毒虫の侵入を完全に防げば,一次感染は起こらず,TYLCV の被害も 発生しない。入れない対策は基本であり,特にハウス開口部への防虫ネットの展張が 重要である。防虫ネットは目合い 0.8mm で侵入防止効果が認められ,侵入を完全に阻 止するには,0.4mm 未満の目合いが必要である(松浦ら,2004)。目合いが細かいネ ットほど侵入防止効果が高くなるが,ハウス内の気温は高くなるため,地域でのトマ ト黄化葉巻病の発生状況や作型に応じて防虫ネットの目合いを選択する。本県では, 目合い 0.8mm 以下の防虫ネットの使用を推奨している。導入当初の防虫ネットは目合 い 0.8mm が主体であったが,十分な効果が認められないハウスもあった。そのために, 現在ではより侵入防止効果が高い目合い 0.4mm の普及割合が高くなっている。また, 野外の保毒虫を減少させてハウス内への侵入量を減らすために,野良生えトマトの除 去も重要な対策として取り組む必要がある。雑草でもタバココナジラミが増殖するた め,ハウス周辺の除草により媒介虫の発生量を減少させる。 近紫外線除去フィルムの展張や光反射シートの設置は,ハウスへのコナジラミ類の 侵入抑制が確認されている(嶋田,1994;長塚,2000)。ただし,現在のところ本県 では両資材ともにトマト黄化葉巻病対策としての普及率は低い。 2)増やさない対策 増やさない対策は,ハウス内での保毒虫の個体数を減少させ二次感染を阻止するこ とが目的であり,薬剤防除が基本である。TYLCV に直接効果のある薬剤はないため, 病原ウイルスを媒介するタバココナジラミを防除する必要がある。薬剤防除を考える 上で,タバココナジラミの発育速度と野外からの侵入の有無が重要である。長期栽培 の促成トマトを例に,発生消長モデルと防除の考え方を図2に示した。タバココナジ ラミが野外で活動するためには、平均気温 10℃以上が必要である(行徳ら,2008)。 野外から ハウス内へ侵入 マルハナバチ導入 タバココナジ ラミ発生量 速い増殖 重要な 防除時期 速い増殖 ゆっくり増殖 定植 9月 10 11 【時期Ⅰ】 感染防止のた め,殺成虫効果 のある薬剤を選 択する。 図2 12 1 2 3 【時期Ⅱ】 媒介虫を増やさないことが重 要。幼虫を対象とした薬剤で も効果がある。マルハナバチ への影響を考慮する。 4 5 6 【時期Ⅲ】 感染防止および野外へ 保毒虫を出さないため に、殺成虫効果のある 薬剤を選択する 促成栽培トマトでのタバココナジラミ発生消長モデルと各時期での防除の考え方 熊本県では、11 月中旬に平均気温が 10℃以下となり、野外からハウス内への侵入量 が少なくなる。また,タバココナジラミは気温が高いと発育期間が短くなり、「定植 から秋期」および「春先から収穫終了」 の期間では増殖が速い。逆に、気温が も遅い。促成栽培では、①侵入の有無 と量、②増殖の速さで 3 つの時期に分 けることができる。すなわち、常に侵 入があり増殖も速い【時期Ⅰ】、侵入 はなく増殖が遅い【時期Ⅱ】、侵入は 成虫+4齢幼虫数/3複葉 低くなる冬期は発育期間が長く、増殖 15 10 5 0 少ないが増殖が速い【時期Ⅲ】である。 9/1 10/1 11/1 12/1 1/1 2/1 3/1 2008年 2009年 このように侵入量や増殖の速さが異 なる時期Ⅰ~Ⅲでは、使用薬剤や防除 11/14 散布 図3 間隔が変わる。また、マルハナバチの タバココナジラミバイオタイプ Q に対する 11 月中旬散布の防除効果 導入時期も防除対策を考えるときに ●:散布区 考慮する必要がある。 定植時粒剤:両区にジノテフラン粒剤 1g/株 【時期Ⅰ】では,TYLCV に対する感 4/1 ○:無散布区 定植:2008 年 9 月 16 日 11 月 14 日の散布薬剤:ピリダベン水和剤 1000 倍 染抑制効果が認められている粒剤を 定植時期に処理することが基本であ る(大矢・植草,2009)。その後も保 は成虫に効果のある薬剤を中心に散 布する。 【時期Ⅱ】では,野外からハウス内 への侵入が少なくなる時期に高い効 果の薬剤で防除することがポイント 成虫+4齢幼虫数/45葉 毒虫による感染を防ぐため,10 月まで 150 100 50 0 である。熊本県では 11 月中旬の散布 0 7 13 20 27 1回目散布後日数 により,ハウス内の密度が低くなり, 時期Ⅱの防除管理が行いやすくなる (図3)。侵入がないため,タバココ ナジラミを増やさないことが重要で あり,幼虫を対象とした薬剤でも十分 な効果が期待できる。 【時期Ⅲ】では気温が高くなり,タ バココナジラミの増殖が早くなるた め,感染防止および野外へ成虫を出さ 図4 タバココナジラミバイオタイプ Q に対する ボーベリア バシアーナ乳剤(ボタニガード ES) の防除効果 ○:ボーベリア バシアーナ乳剤 500 倍 △:ピリダベンフロアブル 1000 倍 ↓:薬剤散布 ●:無処理 散布は,ボーベリア バシアーナ乳剤で 3 回,ピリダベンでは初回の 1 回のみであった。 ないためにも殺成虫効果のある薬剤を主体に選択する。 本県の施設果菜類に発生しているタバココナジラミは,ほとんどがバイオタイプ Q である(樋口ら,2007)。本種は薬剤が効きにくく,有効な薬剤が少ない。このこと により同一系統薬剤の連用による抵抗性発達や使用回数制限による長期栽培での薬 剤不足などの問題を生じやすい。対策として薬剤抵抗性が発達しにくく,使用回数制 限のない微生物殺虫剤や気門封鎖剤の活用があり,バイオタイプ Q に対する密度抑制 効果が認められた(図4)。対照とした化学薬剤に劣る効果や残効を考慮して, 【時期 Ⅱ】に使用する。 薬剤防除以外の増やさない対策もある。ハウス内に感染株が存在すると伝染源とな るため,発病株は抜き取る。また,雑草でタバココナジラミが増殖するので,ハウス 内も除草する。黄色粘着資材での成虫捕殺によるコナジラミ類の密度抑制も可能であ る(徳丸ら,2009)。トマト黄化葉巻病に対する抵抗性品種の利用もあり,県内では 数十ヘクタールで栽培されている。国内で市販されている抵抗性品種は,抵抗性遺伝 子 Ty-1,Ty-2,Ty-3 および Ty-3a を単独または複数保有している(斎藤ら,2008)。 熊本県で発生している TYLCV のイスラエル系統長崎株に対して,Ty-2 を保有してい る抵抗性品種では感染および発病抑制効果が認められている(堤,2010)。しかし, 現在の抵抗性品種は完全な免疫性ではなく TYLCV が感染するため,周辺に感受性品種 が栽培されているとその伝染源になる可能性がある。本県では,抵抗性品種を栽培す る際もタバココナジラミを防除するように指導している。 3)出さない対策 できる。出さない対策では,ハウ ス密閉処理が基本である。密閉処 理は栽培終了後にトマトの株元 誘殺数/1トラップ TYLCV の伝染環を断ち切ることが 400 40 200 ウス内を除草しておくこと,媒介 虫が野外へ飛び出さないように 20 100 0 0 3 ることでタバココナジラミを死 ントは,生存虫をなくすためにハ 60 300 を切断し,ハウス開口部を閉め切 滅させる(図5)。この際のポイ 80 500 虫を野外へ出さないことにより, 9 24 48 ハウス内の最高気温(℃) 栽培終了時にハウス内の保毒 72 密閉処理後の経過時間 図5 3 連棟ビニルハウス栽培トマトでのタバココナ ラジミバイオタイプ Q 成虫の誘殺数とハウス内最 高気温の推移 :成虫の誘殺数(10×10cm の IT シート) ビニルの破損などの開口部をな ●:ハウス内の最高気温(高さ 150cm) くすことである。また,タバココ 2006 年 6 月 5 日に,トマト株元を切断して午前 8 時か ら密閉処理を開始した。図中のバーは,標準誤差を示す。 ナジラミを確実に死滅させるた めに,トマト株が枯死する 1 週間以上の密閉期間を確保する。 (2)地域で取り組む対策 これまで述べたように TYLCV は 100 熊本市 つまり,地域でトマトが栽培されて いない期間を設ければ,TYLCV の伝 染環を断ち切ることができる。そこ で,2003 年 12 月に立ち上げられた 保毒虫率(%) トマトからトマトへ伝染している。 玉名市 八代市 75 50 25 県,農業団体および関係機関などで 構成される熊本県緊急病害虫防除 0 7月 対策会議で,トマト作型の統一によ 8月 9月 トマトを栽培 しない期間 る防除対策が検討された。作型統一 とは,各地域で定植の開始日と栽培 終了の最終日を取り決め,トマトを 図6 野外におけるタバココナジラミの 栽培しない期間を 1 か月以上設け TYLCV 保毒虫率の推移(2007 年) ることである。当初,困難と考えら 野外にトマト苗を設置し,寄生したタバコ れていた作型統一であったが,地域 コナジラミを調査した。平成 20 年度に病 での話し合いにより取り組まれる 害虫防除所が発表した技術情報第 2 号より ようになった。その結果,各地域の 抜粋した。 野外の保毒虫率が低下し(図6), 発生面積は確実に減少していった。 地域に存在する家庭菜園のトマトは,感染源になる可能性がある。そのために,市 町村や JA の広報誌やパンフレット等を用いて,家庭菜園での発病株除去の協力を呼 びかけ,非農家を含む地域全体での取り組みにより感染源を減らしている。 4 おわりに トマト黄化葉巻病の防除対策は,個別農家が取り組む対策だけでなく,地域全体で の取り組みが重要である。TYLCV の感染株や保毒虫を地域全体で低く抑えていくこと が,トマト黄化葉巻病などの虫媒伝染ウイルス病に対する最も効果的な対策となる。 本県では,2004 年に新規ウイルス Cucurbit clorotic yellows virus(CCYV)による ウリ類退緑黄化病が確認された(行徳ら,2009)。本ウイルス病も虫媒伝染ウイルス 病であり,媒介虫はタバココナジラミである。現在,ウリ類退緑黄化病に対する防除 対策にも取り組んでいるが,トマト黄化葉巻病で得られた対策を活用している。今後, 新たな侵入病害虫が問題となることも予想されるが,地域一丸となって安定生産に取 り組んでいきたい。 引用文献 行徳 裕ら(2008)熊本農研セ研究報告 15:50-61. 行徳 裕ら(2009)日植病報 75:109-111. 樋口聡志ら(2007)九病虫研会報 53:59-65. 松浦 明ら(2005)九病虫研会報 51:64-68. 長塚 久(2000)植物防疫 54(9):7-10. 大矢武志・植草秀敏(2009)今月の農業 53(2):17~21. 斎藤 新ら(2008)園学研 7 号(別冊 1):107. 嶋田知英(1994)関東病虫研報 41:213-216. 徳丸 堤 晋ら(2009)関西病虫研報 51:87-88. 泰之(2010)植物防疫 64:390-396. ブラジルの生物的害虫防除について サンパウロ州立大学 植物保護学部 昆虫学科教授 オダイル・フェルナンデス 1. ブラジルの農薬市場規模について 68億米ドル (5500億円 @80円) この内、殺虫剤が34%、殺菌剤27%、除草剤 40% 生物的防除としては、昆虫、菌、ウイルスの大量放飼法が主流。 病害対策としては、交配による耐性付与が主流。 GMO(遺伝子操作植物)としては、ラウンドアップ耐性と Bt 植物は導入されている。 2. ブラジルでの1960年以降の病害虫防除の進化について 1960年代より 高投入高収量時代 (グリーン革命) 1970年代より サトウキビ生物防除開始 1970年代後半 麦での伝統的生物防除プログラム開始 1980年代より 大豆およびワタでの IPM プログラム開始 1980年代半ばより 柑橘での IPM プログラム開始 1990年代より トマトでの生物防除開始 2000年代初めより GMO の導入 3. 生物的防除・・・成功した生物防除プログラム 3-1 麦 アブラムシの生物防除 1960年代後半よりアブラムシの問題が発生。 アブラムシの種類 (和名は漢字混交とした) Metopolophium dirhodum 麦ウスイロアブラムシ Sitobion avenae 麦ヒゲナガアブラムシ Schizaphis graminum 麦緑アブラムシ Rhopalosiphum padi ムギクビレアブラムシ Rhopalosiphum rufiabdominale 陸稲の赤アブラムシ 上記のアブラムシに対して下記の天敵昆虫、寄生天敵、捕食天敵を導入した。 寄生天敵 Aphidius uzbekistanikus ウズベクアブラバチ Praon gallicum プラオン属アブラバチ Praon volucre プラオン属アブラバチ Ephedrus plagiator クロアブラバチ Lysiphlebus testaceipes リシフレブス属アブラバチ Aphelinus asychis アブラコバチ 捕食天敵 Hippodamia convergens サカハチテントウ Coccinella septempunctata 七星天道虫 麦におけるアブラムシの生物的防除の結果: 目標:15%の面積での生物防除の達成 2010 年現在、 生物防除を行っている面積は全面積 200 万ヘクタールの 94%に達している。 3-2 柑橘 柑橘のハモグリ (絵描き虫)の生物防除 害虫名: Phyllocnistis citrella ミカンコハモグリ 1996-1998 にわたり、ハモグリ防除用の殺虫剤の使用が増大した。 カンキツ潰よう病の発生 (これが農薬使用増大と関係あるかは不明:訳者注) 1998 年からフロリダの寄生天敵を導入。 寄生蜂 : Ageniaspis citricola トビコバチ科の寄生蜂 和名なし 天敵導入の結果: 2010/2011 カンキツグリーニング病感染原因であるキジラミ防除の殺虫剤使用にも拘わ らず、40%の生存率を記録した。 農薬無散布地区では、80%の生存率を記録。 南部ブラジルに分布するようになった。 3-3 トマトでの生物防除 キバガの一種である Tuta absoluta トマトハモグリ、英名 Tomato pin worm の被害 が 1980 年代より増大。 生物防除を含むいくつかの対策がとられた。 卵寄生蜂トリコグラマ・プレティオスム trichogramma pretiosum の大量増殖 2010 年現在 民間企業によるトリコグラマの増殖が継続している。 殺虫剤・殺菌剤は継続的に大量に使用されている。 3-4 大豆での生物的防除 大豆の重要害虫 Nezara viridula ミナミアオカメムシ Anticarsia gemmatalis ビロードマメケムシ(大豆 青虫)英名 Velvet bean caterpillar Piezodorus guildinii カメムシ 1970-1980 年代は主に有機リン剤による防除。 1シーズンに 4 種の有機リン剤を散布。 1980 年半ば以降 Anticarsia ビロードマメケムシについては、昆虫ウイルスを散布するようになる。 カメムシについては Trissolucus basalis というタマゴクロバチ科のカメムシの卵寄生蜂 を利用。1ヘクタールあたり 5000 寄生蜂(寄生卵塊を16個ほど付着させたカードを1ヘ クタールあたり3枚の割合=1 カード当たり 100 卵程度となる)を孵化 2 日前ごろに 圃場に吊るす。宅急便にて配達。 ブラジルの大豆面積は 2 千万ヘクタール。ウイルス製剤は最大その10%まで 使われたが、現在は 5%の面積で使用されている。30 トンの冷凍昆虫を利用。 EMBRAPA (国立ブラジル農業研究所)および EMATER(ブラジルの普及機関)の 関与が重要であった。 寄生蜂は 20,000 ヘクタールで現在利用されている。ミナミアオカメムシの密度は低下。 3-4 サトウキビでの生物防除 害虫:サトウキビズイムシ(メイガ) Diatraea saccharalis 天敵昆虫:Cotesia flavipes 幼虫寄生蜂ズイムシサムライコマユバチ 放飼方法:フィールドスカウティングによりメイチュウの密度を調査。生存数と寄生率を 調査の上、放飼タイミングを決定。放飼寄生蜂数 6000 頭/ha. 50mごとに放飼。 35 年間放飼を継続。現在 200 万ヘクタールで放飼している。 現在はアワフキムシの一種が問題である。Mahanarva fimbriolata (Hem.: Cercopidae) メタリジウム菌を使った防除 400 万ヘクタールで使用。 ブラジルの各作物における生物防除の比率(最大値) 2010 100 90 80 70 60 50 化学防除 40 生物防除 30 20 10 0 小麦 カンキツ トマト 大豆 サトウキビ 4. 生物防除の成功の秘訣 政府機関の初期投資が不可欠。 研究機関(大学、農水省関係)の協力が不可欠。 生物防除は化学農薬防除にくらべコストは高い。天敵コマユバチ Cotesia の場合 700 円/ha 化学農薬コストは不明。 製糖会社は自社の天敵増殖設備を保有している。 ブラジルではバイオエタノール生産で追い風が吹いている。生物防除のコストを負担 出来る状況。 5. 将来性のある生物防除種 センチュウ捕捉・寄生性菌 Arthrobotrys oligospora A. musiformis Dactylella leptospora Monacrosporium robustum Paecilmyces lilacinus 6. 生物防除の今後 社会は環境に優しい病害虫防除を要求している。 Society claims for ecologically sound management tools. 世界の天敵生物はまだ十分に研究、開発されていない。 Diversity of Natural enemies was not fully explointed yet. 保全的生物防除:まだ幼児期にある。研究領域は広い。 農薬は今後も使われるが、削減の傾向は続く。 散布回数、投与量。代替方法の模索も続くであろう。 化学農薬も含めた全体的な植物保護システムの統合を図る必要がある。 生物防除に関わる研究者、教育者、学生、生産者の増加が必要。 生物防除に関する法律、規制などは、これまでの化学農薬の規制と同じではなく むしろ進めていく方向に政府は転換するべき。 生物防除資材の品質管理は必要。 協力:筑波大学、東京農工大学 その他関係研究機関など。 2010 年春来日。 (翻訳および文責 和田哲夫 ジャパンアイピーエムシステム) 在来種天敵の農薬としての登録は不必要 天敵利用研究家 和田哲夫 日本における天敵昆虫の登録要件については、1990年代初頭より、明確なガイドラ インがなく、民間より化学農薬のガイドラインを参考にガイドライン案などを自主的に提 出してきた。 農水省の研究者と日本植物防疫協会の専門家などと、相談しながら、それぞれの天敵種に ついての使用方法などについて、協議を重ねたのは記憶に新しい。 当時はオランダ産の天敵昆虫で日本にとっては、外来種(exotic insect)であるオンシ ツコナジラミとチリカブリダニ、コレマンアブラバチ、ククメリスカブリダニが登録対象 であった。オランダ産と言っても、チリカブリダニは地中海沿岸が原産地であり、他の天 敵にしてもかならずしもオランダ原産というわけでもなかった。 オンシツコナジラミは九州大学の梶田先生、チリカブリダニは森鷗外の孫としても知ら れていた森ハンス教授らの研究と導入の蓄積があり、日本においては、登録なしで各地の 試験場や農家で使用されていたのである。静岡県の試験場では、チリカブリダニが、三重 県の旧野菜茶業試験場では、オンシツツヤコバチが増殖されていたのを見たことがある。 これらは、農水省の補助金などで行われきたため、特例として、登録なしで利用できたの である。鹿児島県でのチャハマキ防除用のハマキ天敵が登録なくして地元農協で増殖され、 広い面積で使用されてきたのと同様である。 天敵昆虫の登録要件のなかでもっとも障害となったのは、生物効果試験の実施であった。 化学農薬と同様に作物毎、対象害虫ごとに6例づつ試験をする必要があった。現在のコ ストは1試験33万円程度で、化学農薬の試験のコストより高い。各国政府が生物農薬の 普及を促進するため、登録コストを下げるようにしているのとは対照的である。 コストもさることながら、当時は天敵昆虫の試験をしてくれる試験場の研究者の数が圧 倒的に少なく、また経験もない方も多く、試験の実施が困難であり、とても2年間で有効 例6データを積み上げることはできなかった。このため有効な生物効果データ6例を得る までに3-4年もかかってしまった。実際にこれらの試験場で実施した試験が、現場の農 家のハウスでも再現が可能であったかといえば、ハウスの大きさ、温度条件などにより、 再現性は極めてひくかったのである。 現場での指導はまた別物であり、技術情報はもっぱらオランダからのノウハウであっ た。またこれらの天敵を触ったことのある県の技術者たちの助言もきわめて有効であった。 現場では、成功例より失敗例が多く、これは初期の天敵類の使い勝手の悪さに起因する ものが多かった。環境条件をととのえ、放飼タイミングがベストであったとしても、また 別の理由により効果のでないこともしばしばであったのである。このため、この時期に天 敵利用を見限った農家も多かったのである。しかし中には、研究熱心な農家、研究者、普 及員、流通の方々の努力により、成功例もだんだんと増加してきた。 効果試験に年数がかかりすぎて、防除のタイミングを失うという悲喜劇もあった。 ハモグリバエの天敵ハモグリコマユバチ、イサエアヒメコバチでの例がそうである。 開発試験をしている間は、トマトなどでのハモグリに被害は甚大であった、4年程度経過 して、いざ登録がおりた時点では、すでにハモグリバエの被害はなくなっていたのである。 たぶん国産天敵の捕食、寄生により、ハモグリの密度が激減したのだった。 このように、天敵利用においては、問題害虫に対するスピーディな対応がもとめられる。 日本産の在来種天敵とは、日本原産で生息する在来種の天敵類のことであるが、外来種 が定着した種はとりあえず除外する。 在来種天敵で以前から利用が期待されていた種はたとえば、フツウカブリダニ、ケナガ カブリダニ(茶)、ニセラーゴカブリダニ(果樹)、多くのツヤコバチ、サシガメ(鱗翅目 捕食者)ヒメオオメカメムシ(捕食者)など多くの種が天敵昆虫として期待されていた。 近年では、クロヒョウタンおよびタバコカスミカメ(温室)やキイカブリダニ、ミドリ カスミカメ、カタグロミドリカスミカメ(飛来天敵:水稲)など重要で、もし使えるよう になれば、日本作物保護が大きくかわる可能性のある天敵昆虫が目白押しである。 海外では原則、EUでも米国でも天敵昆虫に登録は必要がない。日本の昆虫学者はせっ かくの研究成果を実現できない状況である。また日本が生物防除をリードする国としても、 日本の在来種天敵については、すくなくとも生物効果試験の除外を早急に実施すべきと考 えます。 天敵増殖会社が困難な増殖に注力することができるようになるように、また小さな会社で も天敵防除に参加できるようにするためにはどうしても必要な改善でありましょう。 潮流は生物農薬の利用の方向に向かっているか? ヴィレム・ラーフェンスベルグ Ph.D. Koppert Biological Systems はじめに 病害虫防除に影響をあたえるファクターはさまざまであるが、生物防除、IPM に おいては法令、環境および食品安全性についての懸念などがあげられる。各国の政策 が直接、間接に植物保護に影響を与えている。 生物防除を阻害するファクター 政策 既存の化学農薬が微生物農薬より価格が安いかぎり、植物保護において化学農薬が マジョリティを占め続けるであろう。しかしながらよく知られているように化学合成 農薬には、隠れた環境コストが存在しているのである。 微生物農薬には化学農薬のような環境コストは存在しない。政府による微生物農薬 利用を促進するような政策がうちだされなければ、いつまでもこの状況は変わらない であろう。 生物多様性条約における発展途上国と先進国間での資源利益配分ルールは生物防 除への阻害要因となりうる。2010 年 10 月の名古屋議定書が批准されれば、関係国の 間での契約が必要となってくる。政府間の現実的な取り組みがなければ持続可能な病 害虫防除を継続することが困難になるであろう。 社会および市場からのネガティブファクター 天敵昆虫や微生物農薬が広汎に使用されると、昆虫およびカビという一般大衆から は否定的に受け取られていることによるリスクが生ずる可能性がある。情報を十分に 社会に流すことでこのような嫌悪感を払しょくすることが可能であろう。 化学農薬というひとつの方向からだけの技術に頼るのではなく、生物農薬を IPM のなかでの中心素材として組み入れるべきである。それらは化学農薬の代用としてで はなく、また化学農薬と競合するものとして期待されるものではないのである。 生産者も防除パターンを変えるべきであり、総合的に防除を考えるべきなのである。 生産者が生物防除に切り替えるためには、コストのカバーと技術力の蓄積が必要であ る。このためには、消費者は生物防除を支援するためのコストを相応に負担しなけれ ばならない。持続的農業を希望するならば。 侵入病害虫 気候変動と貿易の増加により病害虫が世界的に伝播しつつある。このために化学農 薬を多用することが生物防除、IPM を阻害している。 生物防除を促進するファクター 社会、市場からのポジティブファクター 環境保護グループや消費者団体は生鮮食品における化学農薬の残留量を減らすよ うスーパーマーケットや生産グループに強く要望しつづけている。結果として、多く のスーパーマーケットは各国政府が規定する水準以下に残留量を減らすようガイド ラインを設定している。また使用される化学農薬の原体数(種類数)自体を減少する べく指導している。 G-GAP グローバルギャップというスーパーマーケットと出荷グループによって 構成された世界的な機関は、IPM の利用を含めた多くの「義務」を設定している。 このスーパーマーケットによる運動は化学農薬を減らす重要なドライビングフォー ス(牽引力)となっている。生物防除はこのギャップを埋めることができる。 科学技術的ファクター 近年既知の微生物殺虫剤について新たな利用方法が提唱されてきている。例えば、 植物病原菌に対するアンタゴニスト、センチュウ防除、害虫に対するエンドファイト、 植物成長促進などへの用途としてである。 いくつかの病害防除用の微生物剤は植食性昆虫に対する植物の防御システムのト リッガーとなることも知られてきている。 数種類の微生物殺虫剤のハイブリダイゼイションにより、対象害虫の拡大が可能と なる。 メタリジウム・アニソプリエ菌では、土壌害虫防除の効果をあげる microsclerotia を生産するストレインの増殖方法が発見されている。また BT 剤のシネルギスト(効 果を上げる物質)としてある種のタンパクが発見されておりスペクトラムの拡大に貢 献する可能性がある。 遺伝子工学も重要な分野であるが、本稿では省略する。 各国政府の環境の変化 微生物農薬の登録のハードルはいまだに高いといわざるを得ない。これは化学農薬 の登録ガイドラインを不適切に使用しているからにほかならない。 ヨーロッパにおいては、EU での評価と、各国登録と二つの評価を通過しなければ ならないという弊害がある。 EU の新ルールである EC/1107/2009 では、ローリスクな物質、たとえば微生物農 薬などについては、カテゴリーが作られたものの、細則はまだ決まっていない。 イギリスの微生物農薬イニシアチブ、オランダのゲヌーフ・プロジェクトなどにより、 新規微生物農薬が多く承認されている。 イギリス政府は微生物専門の担当(Champion)制度があらたに制度化され、申請 会社と政府窓口とのコミュニケイションの向上に役立っている。 他の EU 各国でも同様の動きがみられ、デンマークでは、化学農薬以外の申請者へ の補助制度が 2011 年より発効している。 OECD 微生物農薬ステアリングコミッティーにおいては、微生物農薬、植物抽出 物、フェロモンについてのガイドラインを作成しているが、世界的にきわめて重要な 作業である。 内容的には下記のようである。 同定法、種としての毒性の評価、ヒトへの暴露評価、代謝物の作物残留、効果試験、 提出データの免除の条件など。 もう一つの新しい 2009/128/EC の法律では、各国は IPM を促進し、化学農薬への 依存を下げる技術を開発すべきとある。 これらの各国での施行は 2014 年からとなっている。 生物多様性と環境への考慮 生物多様性条約の順守においては、化学農薬にだけ頼る方法は、生物防除を阻害し かねず、まず生物防除を採用したのちに、化学農薬による防除を考慮するという思考 パターンが必要になってくる。 生物な農薬産業における潮流 生物農薬産業においては、近年、生産効率が飛躍的に向上している。 製剤技術、品質管理、生物効果、処理方法、マーケティングなどにおいての過去 10 年での進歩は目覚ましいものがある。 生物農薬の信頼性は向上し、価格も安定し、マーケットは拡大している。 欧州と米国以外での生物農薬産業の全体像は把握が困難であるが、ヨーロッパにおい ては 1995 年に設立された IBMA (国際生物農薬生産協会)は約 160 社が加盟して いる。この内生産会社は 30 社である。 米国では BPIA(生物農薬工業会)は 60 社の会員を擁しており、その内約 10 社が 微生物農薬の製造会社であり、その他は天敵昆虫、フェロモン、植物抽出物や生化学 的植物保護剤に関わっている。近年は大手化学農薬メーカーのこの分野への参入も見 られる。 2009 年にドイツのバイエル社はセンチュウ防除用のアグログリーン社のバイオネ ム剤( B.firmus )及びポストハーヴェスト用殺菌剤である Shemer( Metschnikowia fructicola )を買収している。 2009 年には、やはりドイツの BASF 社は米国 Agraquest 社の微生物殺菌剤セレナ ーデの全世界での販売権を取得し、バイエル社は同剤の米国での家庭園芸用分野での 販売権を得ている。日本では、アリスタライフサイエンス、三井物産、住友化学など が生物農薬の関連ビジネスに強く関与している。すなわちフランス NPP 社、Certis 社、Valent 社である。 最近の侵入害虫 化学農薬の効果が低い侵入害虫が近年増加している。 ヨーロッパでは、コーンルートウォーム( Diabrotica virgifera virgifera )と南米 から侵入したトマトピンウォーム Tuta absoluta である。後者には BT 剤が大量に使 われているが、天敵昆虫であるタバコカスミカメ Nesidiocoris tenuis の効果が高 いことが知られている。またヤシの木を食害する難防除害虫であるゾウムシ Rhynchophorus ferrugineus はスタイナーネマによる防除が増えている。 市場規模と成長 世界の生物農薬(天敵昆虫を含めない)マーケットサイズは 2008 年で 4 億ドル程 度である。成長速度は毎年 15-20%と推定される。 基本的にニッチでかつ、高価格の作物に使用が伸びている。しかしながら一般の農 業分野でも、特に病害分野での利用が期待される。 森林、都市での緑の空間、緑地、果樹園、蚊、ハエなどの防除にも利用されている。 BT 剤以外に、バチルス・スブチルス剤、シュードモナス菌もより使われるようにな るであろう。 害虫防除には、バキュロウイルスも果樹および温室での利用が増えている。スタイ ナーネマのような昆虫病原性センチュウは着実に果樹、温室分野で新しい害虫を含め 各国において伸長している。欧州では、病害には Trichoderma spp .製剤が急速に増 加している。 結語 持続可能で農薬残留のない世界を実現するためには、生物防除が植物保護において 大きな部分をカバーするようになることが十分予想される。 (翻訳・文責 和田哲夫) (訳者注 ラーフェンスベルグ博士はコパート社における微生物農薬開発の責任者で ある。化学農薬についてはかなり辛口な語り口ではある。) ヨーロッパとオランダにおける微生物農薬の利用状況 (2009) ヨーロッパ全体 5% 17% 17% BT剤 寄生性センチュウ 天敵ウイルス 5% 21% 微生物殺虫剤 微生物殺菌剤 その他 35% オランダ 6% 11% BT剤 4% 寄生性センチュウ 天敵ウイルス 52% 13% 微生物殺虫剤 微生物殺菌剤 その他 14% W.J. Ravensberg, 2011. A Roadmap to the Successful Development and Commercialization of Microbial Pest Control Products for Control of Arthropods. Springer, Netherlands. 高知県の施設果菜類における天敵を利用したアザミウマ類防除の現状 高知県農業振興部 環境農業推進課 古味一洋 はじめに 高知県ではミナミキイロアザミウマやヒラズハナアザミウマの薬剤感受性低下が 顕在化し、安定的な防除効果の期待できる薬剤は極めて少ない。そのため栽培期間が 長期にわたる促成栽培の施設ナス科果菜類などの栽培では環境保全型農業の推進と ともに薬剤感受性の低下したアザミウマ類対策として天敵類が利用されている。ここ では、高知県で栽培されている主要な果菜類について天敵を用いたアザミウマ類やそ の他の害虫防除について紹介したい。 施設ナス 施設ナスでは平成 9 年頃から天敵利用によるミナミキイロアザミウマ防除が始ま った。これはホルモン剤の単花処理にかかる労力軽減のため、受粉用のマルハナバチ を導入したことで殺虫剤の使用が制限されたことが契機となった。当初はアザミウマ 類に対してククメリスカブリダニのみが利用可能な天敵であり、アザミウマ類に対す る効果の不安定性とともに、チャノホコリダニ対策にも苦慮する状況であった。その 後、タイリクヒメハナカメムシが利用できるようになると、定植時の粒剤処理、選択 性殺虫剤ラノー乳剤の効果的な使用と併せてミナミキイロアザミウマに対する防除 効果が安定した。他の害虫に対する天敵利用、黄色蛍光灯や防虫ネットなど物理的防 除技術の導入なども進み総合的な害虫管理ができるようになってきた。このため、平 成 16 年度には施設ナスでの天敵導入面積率が 43%に達した。しかし、この時期から タバココナジラミバイオタイプ Q の発生が問題となってきた。この当時の IPM 体系 ではタバココナジラミに対しては無防備であったため、やむなく天敵の導入をあきら めた生産者も多く、平成 17、18 年には天敵導入面積率が 27%、29%と大きく低下し た(図 1)。 このタバココナジラミの危機を救ったのが土着天敵タバコカスミカメ(写真 1)で あった。本種は若干の食害をナスに与えるものの、タバココナジラミに対する十分な 防除効果を示したため施設ナスでの利用が急速に進んだ。また、本種はタバココナジ ラミだけでなく、個体数が増えればミナミキイロアザミウマの防除も可能であること、 ゴマ(写真 2)で比較的簡単に維持、増殖できるため当初利用が進んだ安芸地区から 県内全域へと利用が拡大した。このため、天敵導入面積が急速に高まり平成 22 年に は 59%となった(図 1)。 この他、スワルスキーカブリダニの利用も進んでいるが、後述する施設ピーマン・ シシトウ比較して冬期の管理温度が低い施設ナスでは活動できる期間が短いことか らミナミキイロアザミウマ、タバココナジラミに対しては補助的な天敵となっている。 ただし、チャノホコリダニに対する防除効果が認められており存在価値は高い。 90% 施設ナス 天敵導入面積率(%) 80% 施設ピーマン類 70% 施設キュウリ 60% 50% 40% 30% 20% 10% 0% H13 H14 H15 H16 H17 H18 H19 H20 H21 図1 高知県の主要施設果菜類における天敵導入面積率の推移 注)高知県農業振興部環境農業推進課取りまとめ。 写真 1 タバココナジラミ 写真 2 ゴマ 施設ピーマン類 施設ピーマン類についても定植時の粒剤処理とタイリクヒメハナカメムシ放飼、選 択性殺虫剤および物理的防除などを組み合わせることでアザミウマ類や各種害虫に 対する防除体系が確立されていた。しかしながら、施設ナスと同様にタバココナジラ ミバイオタイプ Q の出現により、大きく防除体系が崩れることになった。 施設ナスにおいてこの危機を救ったのはタバコカスミカメであったが、施設ピーマ ン、シシトウではスワルスキーカブリダニとクロヒョウタンカスミカメ(写真 3)で あった。スワルスキーカブリダニはチリカブリダニのように対象害虫よりも高い増殖 能力をもって押さえ込むタイプではなく、花粉を代替餌として対象害虫がいない状態 でも個体数を維持しながら長期間害虫の密度を押さえ込む天敵である。本種は花粉量 が多く、冬期の管理温度が高いピーマン(18℃)、シシトウ(20℃)での定着性は非 常に高く、タバココナジラミに対する十分な防除効果を示したため急速に導入が進ん だ。また、スワルスキーカブリダニのチャノホコリダニに対する防除効果は高く、施 設ピーマン・シシトウでの被害はほとんどみられなくなった。現在のところ、一部産 地を除き、タイリクヒメハナカメムシとスワルスキーカブリダニを用いた IPM 体系 で主要害虫に対する防除は安定している。ただし、スワルスキーカブリダニはテトラ ニカス族のハダニ類に対する密度抑制効果が期待できないため、注意が必要となる。 また、クロヒョウタンカスミカメはア ザミウマ類、コナジラミ類、ハダニ類な どを捕食できる広食性の天敵である。本 種は、タバコカスミカメのように簡単に 維持、増殖することが難しく、野外から の 採 集 や 温存 ハ ウ ス で維 持 し た もの を 利用している。このため、クロヒョウタ ン カ ス ミ カメ の 放 飼 量を 確 保 す るの は 難しいものの、ピーマン、シシトウを加 害しないことが評価されている。この他、 薬 剤 に 弱 いこ と も あ り使 い こ な すの が 写真 3 クロヒョウタンカスミカメ 難しい天敵といえる。 施設ウリ科果菜類 施設キュウリにおいてはスワルスキーカブリダニの販売開始と同時に天敵利用が 始まった。前述した施設ナス、ピーマン類で天敵類の導入が進んだ状況とウリ科果菜 類の状況は大きく異なり、平成 21 年の天敵導入面積率 1.8%(施設キュウリ)とな っている。これは、アザミウマ媒介性のウイルス病の発生程度の影響が大きい。施設 ナス、ピーマン類ではアザミウマ媒介性ウイルスであるトマト黄化えそウイルスによ る病害(TSWV)の発生がほとんどなく、アザミウマ類の防除はその直接的な被害を 抑えればよい。一方、施設ウリ科果菜類では、依然としてミナミキイロアザミウマが 媒介するメロン黄化えそウイルスによる病害(MYSV)が猛威をふるっている。元々、 ミナミキイロアザミウマの直接的な果実被害を受けにくいウリ類の方が現状ではア ザミウマの被害許容水準が低い状況になっている。 施設キュウリにおいてスワルスキーカブリダニを利用したミナミキイロアザミウ マ防除は取り組み始めたばかりで IPM 体系の確立まではかなりの時間がかかると思 われる。また、スワルスキーカブリダニだけでなくアザミウマ類に対する密度抑制効 果の高いタイリクヒメハナカメムシなどの導入が必要となるかもしれない。しかし、 タイリクヒメハナカメムシに影響がなくミナミキイロアザミウマに対する防除効果 の高い薬剤は皆無といってよい状況にあり、MYSV の感染が拡がっている圃場での対 応に苦慮することになる。また、施設メロン、スイカではキュウリよりも作期が短い (3 ヶ月)ため毎作期に天敵を再導入しなければならずコスト増が避けられない状況 にあり、天敵導入のハードルは高い。しかし、施設キュウリ同様に抵抗性を獲得した ミナミキイロアザミウマ対策に打つ手はないため、天敵導入へと検討を進めなければ ならない。 おわりに 施設ウリ科果菜類では天敵を用いた害虫防除が始まったばかりで早急に IPM 体系 を確立する必要がある。施設ナス科果菜類では主要害虫に対する IPM 体系は一定確 立しているものの、問題がないわけではない。近年、施設ピーマン・シシトウに寄生 するチャノキイロアザミウマによる被害(写真 4)が発生し問題となっている。この 他、クリバネアザミウマ(写真 5)、モトジロアザミウマ(写真 6)といった新たなア ザミウマ害虫の発生もみられている。この他にもチリカブリダニ、ミヤコカブリダニ では防除できないミツユビナミハダニの発生も確認されており、既存の IPM 体系の 見直しをしていく必要がある。新たな害 虫が問題となる一方、新規の天敵開発に 期待したい。本県で有望と見いだされた ヘヤカブリダニ、キイカブリダニや当初 タ イ リ ク ヒメ ハ ナ カ メム シ の ブ ース タ ー 天 敵 と して 研 究 さ れた ア カ メ ガシ ワ クダアザミウマなどが期待できる。今後 これらが登録され、さらに IPM 体系が 強 固 な も のと な る よ う関 係 機 関 と連 携 を密にして行きたい。 写真 5 クリバネアザミウマ 写真 4 チャノキイロアザミウマ によるシシトウ被害 写真 6 モトジロアザミウマ 天敵誘引物質と天敵の関係について 京都大学 生態学研究センター 上船雅義 1 はじめに 天敵が餌や寄主を探す際に、何を利用しているだろうか?天敵などの昆虫が住む世 界を人間社会に例えるならば、植物が生い茂った環境はまるで東京のようにビルが建 ち並ぶ都会のような状況だろう。つまり、人間であれば建物で周りが見渡せない状況 下で飲食店を探すようなものである。そのような状況下で、まず我々はカレーや珈琲 などの匂いを発見し、この匂いを手がかりに飲食店の方へ近づいていき、看板などを 発見して最終的にお目当ての飲食店へたどり着くだろう。まさに天敵も同じように、 自分の餌が存在する植物を発見するための手がかりとして匂いを利用している。そし て、この匂いこそが天敵誘引物質である。では、天敵誘引物質はどこから放出されて いるのだろうか?そこでは、餌からでる天敵誘引物質と植物からでる天敵誘引物質と いう2つの場合が存在する。 2 天敵誘引物質の放出源 (1)天敵の餌や寄主由来の天敵誘引物質 天敵が手かがりの匂いとして考えられるのは、餌や寄主が出すフェロモンである。 性フェロモンは、交尾相手を誘引する物質であり、アブラムシの天敵である寄生蜂が 誘引されることが研究により分かっている(Hardie et al., 1994)。集合フェロモンは、 同種の仲間を集めるフェロモンであるが、カメムシの寄生蜂が誘引されることが報告 されている(水谷, 2006)。 天敵が手がかりとして利用しているもう一つの匂いとして植食者の糞や脱皮殻が 考えられるが、これらの匂いにはハダニの天敵が誘引されない場合が報告されている (Maeda et al., 1999)。しかし、天敵は、植食者の糞や脱皮殻を全く餌の手がかりと して利用しない訳ではない。天敵は、直接糞などに触角で触れることで認識する物質 (接触刺激物質、人で言えば味覚で感じる物質)を餌の手がかりとして利用している。 (2)植物由来の天敵誘引物質 植物は、植食者から食害を受けると特異的な匂い(植食者誘導性植物揮発性物質 (Herbivore Induced Plant Volatiles)、略称 HIPVs)を生産し、放出する。この匂い を手がかりに食害した植食者を食べる天敵がやってくる。HIPVs は、食害した植食 者の種によってその匂いが変化するため、天敵は HIPVs から自分の餌が植物を食べ ているかどうかを判断することができる。植食者種による HIPVs の匂いの違いは、 人間でも密閉した容器に食害植物を入れておくと分かるだろう。私の経験では、ミナ ミキイロアザミウマに食べられたナスは渋い匂いがするが、カンザワハダニが食べる と甘い匂いがする。また、食害している植食者密度の変化によっても HIPVs は変化 し、天敵は植食者密度が高い植物の匂いを選好することが知られている。つまり、 HIPVs は、植食者の種類だけでなく数までわかる情報を持つ天敵誘引物質と言える だろう。 3 天敵誘引物質に対する天敵の反応 天敵誘引物質の受け手である天敵はどのように天敵誘引物質に反応しているのだ ろうか?このメカニズムを理解できれば、天敵誘引物質を用いて畑に来てほしい天敵 を誘引することが可能となる。しかし、この話をする前に、天敵誘引物質がどのよう なものか知る必要がある。フェロモンにしても HIPVs にしても他の匂いと異なるこ とで情報源となり、天敵を誘引する。フェロモンや HIPVs は、複数の物質からなり、 それらに含まれる物質の種類の違いとブレンド比の違いからの特異性が生まれる。で は、天敵はどのように反応するだろうか? (1)天敵が反応している物質は単品なのか?ブレンドなのか? モンシロチョウの天敵アオムシサムライコマユバチは、モンシロチョウ幼虫が食害 したキャベツの HIPVs に含まれるヘキサナールやヘキセニルアセテートの物質を単 品で与えた場合に誘引されることが明らかになっている。しかし、コナガの天敵コナ ガサムライコマユバチは、コナガ幼虫が食害したキャベツの HIPVs に含まれる 4 成 分をそれぞれ単品で与えても誘引されないが、4 成分のブレンドを与えると誘引され る。つまり、天敵の種類によって単品で誘引できたり、ブレンドでないと誘引できな い場合がある。このため、天敵誘引物質で天敵を誘引したい場合は、対象としている 天敵の誘引条件が単品なのかブレンドなのか調べる必要がある。 (2)天敵が反応する物質の濃度が影響するか? 物質の濃度の違いも昆虫にとっては重要な情報源となっている。例えば、カメムシ が出す匂いは、濃度が高いときは警戒フェロモンになり逃避行動を引き起こすが、濃 度が低いと集合フェロモンとなる。天敵誘引物質でこのような情報の変化が起こって いるかは不明であるが、コナガサムライコマユバチは誘引される濃度が決まっており、 それより低すぎても高すぎても反応しない。このため、天敵誘引物質を用いて天敵を 誘引したい場合には濃度も検討する必要がある。 4 害虫防除への天敵誘引物質の利用 作物が害虫に食害を受ければ、天敵誘引物質が放出される。そして、天敵が誘引さ れ、害虫を食べてくれる。しかし、これだけでは害虫による経済的な被害が出ないよ うにするには難しい。最大の理由の1つは、天敵を誘引する匂いを放するときには、 後手の防衛システムであるため、害虫から食害を受けており、天敵が来たときには害 虫個体群の増殖を抑えきれないことである。そこで、天敵をできるだけ早く誘引しよ うとする試みが、世界中で広く行われている。 (1)天敵誘引物質を放出する植物の利用 バンカー植物、障壁作物、コンパニオンプランツは、これらの正確な目的は違うが、 アブラムシなどが食害することで天敵誘引物質が放出され天敵を誘引する能力を持 っている。天敵を放飼する場合もあるが、この植物由来の天敵誘引物質により畑周辺 の天敵が誘引され、維持されている。さらに、コンパニオンプランツなどの花は、蜜 や花粉が寄生蜂やアブ類などの天敵の餌資源になっており、花の匂いも天敵誘引物質 になっていると考えられる。 (2)植物ホルモンの利用 食害応答に関係する植物ホルモンを用いて、食害を受ける前から植物に天敵誘引物 質を放出させたり、食害を受けた場合には通常より多くの量の天敵誘引物質を放出さ せる手法が研究されている。例えば、ジャスモン酸は、植物が昆虫に食害されると増 え、防御反応を誘導する物質である。ジャスモン酸を健全なトウモロコシに処理する と、アワヨトウが食害した時と同じ物質が放出され、さらに処理されたトウモロコシ がアワヨトウに食害されるとより多くの量が放出される(Ozawa et al., 2004)。また、 ジャスモン酸をトマトに処理することでシロイチモジヨトウへの寄生蜂の寄生率が 向上する(Thaler, 1999)。サリチル酸は、主に病原菌が感染すると合成され、病原 菌に対する抵抗性を誘導する物質だが、ナミハダニが食害しても合成される。健全な リママメにジャスモン酸とメチル化したサリチル酸を処理すると、ナミハダニの食害 で放出される匂いと同じ成分が放出される(Ozawa et al., 2000)。 (3)植物の選抜 シロイヌナズナやタバコでは、遺伝子組換えによって天敵誘引成分の 1 つであるオ シメンを恒常的に放出する個体が作成されており、遺伝子組換えや育種によって、食 害の有無にかかわらず天敵誘引物質を常に放出するものを作りだすことで害虫防除 を行える可能性は十分にある。しかし、BT 作物のような遺伝子組換え作物は、健康 への安全性や生態系へ与える影響が問題視され我が国ではほとんど栽培されていな い。このような経緯もあって、天敵誘引物質の放出を強化した遺伝子組換え作物を栽 培し販売することは難しいと考えられる。 (4)合成した天敵誘引物質の利用 合成した HIPVs を用いて来てほしい天敵を誘引できれば、誘引した天敵を用いて 防除したい害虫を防除できると考えられる。また、害虫の発生有無にかかわらず、合 成した HIPVs を用いて常に天敵を少数呼び寄せられれば、天敵放飼のための害虫モ ニタリングの労力が不必要になる。しかし、対象害虫と天敵を絞った研究はほとんど ない。James and Grasswitz(2005)は、多くの植物の HIPVs 内で共通して認めら れる物質や植物ホルモンをブレンドして作成した HIPVs を畑に設置して、寄生蜂の 個体数を増加させた。 特定の天敵を誘引して対象害虫を防除した成果は、筆者らの研究グループが出して おり、アブラナ科の重要害虫コナガの防除するために、コナガの優占的な天敵である コナガサムライコマユバチを誘引することができる合成 HIPVs(天敵誘引剤)の開 発に成功している。そして、ミズナ栽培ハウスにおけるコナガ防除試験では、天敵に 餌を与える装置と一緒に天敵誘引剤を設置したミズナ栽培ハウスでは、設置しなかっ たハウスに比べてコナガの発生数が半分以下になった。このことにより、天敵誘引剤 を用いて防除対象の害虫を抑えることができることを実証している(上船ら, 未発 表)。 5 天敵誘引物質の他の効果 植物由来の天敵誘引物質(HIPVs)は、天敵を植物上に誘引した後でも、天敵はこ の物質にさらされている。このため、天敵は植物上へ降り立った後でも HIPVs の影 響を受ける可能性がある。ワタアブラムシの寄生蜂は、HIPVs を生息環境の評価の 手 が か り と し て 利 用 し て お り 、 植 物 上 で の 滞 在 時 間 に HIPVs が 影 響 し て い た (Tentelier and Fauvergue, 2007)。コナガサムライコマユバチでは、誘引される合 成 HIPVs が存在することで植物上での滞在時間が長くなるため、天敵を誘引する機 能を持つ物質は、植物上での天敵の滞在時間を長くする機能もあると考えられた (Uefune et al., 2011)。 6 合成した天敵誘引物質(天敵誘引剤)を用いた害虫防除の今後の課題 (1)技術の適応拡大 天敵誘引剤は、防除対象害虫の天敵を誘引する技術であるため、すべての天敵を誘 引できるわけではない。このため、対象となる害虫ごとに天敵誘引剤を開発していく ことが一般的だと考えられる。現在、天敵誘引剤を用いた害虫防除技術はコナガのみ でしか開発できておらず、防除できる害虫が少ない。今後、ハダニ類、アザミウマ類、 アブラムシ類など問題となっている害虫を抑制できる天敵誘引剤が開発できれば、天 敵誘引剤を用いた技術は有望な害虫防除技術の1つとなるだろう。 コナガに対する天敵誘引剤を用いた害虫防除技術は、ハウス栽培のミズナで開発さ れてきたが、現在では露地キャベツでもコナガに対する防除効果が認められてきた。 このため、露地栽培条件でも天敵誘引剤を使用して対象害虫を防除できるかどうか評 価するためにさらなる研究が必要である。 (2)農薬登録 現在、天敵誘引剤は、食害植物の匂いにも含まれる天然化合物であり、さらに作 物体に散布しないにも関わらず、害虫防除効果を謳うため農薬登録を取る必要が出 てきた。農薬登録には、薬効試験など得る必要があり、取得までに時間と費用を要 する。この新しい害虫防除技術である天敵誘引剤の農薬登録において登録のハード ルをどのくらい下げられたかが今後の技術普及や技術拡大に影響するため、農薬登 録に向けた活動は慎重に進めていく必要があると考えている。 現在、コナガ防除のための天敵誘引剤を農薬登録するための薬効試験では、コナ ガの個体数が減るということを示すことを優先して行っているが、技術普及を達成 するためには収量がどのくらい向上し、生産者の利益がどのくらい向上するかを調 べていく必要がある。 7 おわりに 天敵誘引剤を用いた害虫防除技術は、畑周辺に土着天敵が存在して初めて成り立つ。 このため、畑周辺に天敵が生息するように環境を保全管理する必要がある。環境の保 全管理によってより多くの天敵が維持できれば、天敵誘引剤を用いた害虫防除の効果 はより安定的なものになると予想される。最終的な目標として、害虫は畑から周辺環 境へ押し出し、天敵は周辺環境から畑で移動させることを目指したい。このための天 敵の行動操作に天敵誘引物質が活躍できると考えている。 8 引用文献 James, DG. and TR. Grasswitz (2005) Synthetic herbivore-induced plant volatiles increase field captures of parasitic wasps. BioControl. 50: 871-880. Hardie, J., AJ. Hick, C. Holler, J. Mann, L. Merritt, SF. Nottingham, W. Powell, LJ. Wadhams, J. Witthinrich and AF. Wright (1994) The responses of Praon spp. parasitoids to aphid sex pheromone components in the field. Entomologia Experimentalis et Applicata. 71: 95-99. Maeda, T., J. Takabayashi, S. Yano, and A. Takafuji (1999) Response of the predatory mite, Amblyseius womersleyi (Acari : Phytoseiidae), toward herbivore-induced plant volatiles: Variation in response between two local populations. Applied Entomology and Zoology. 34: 449-454. 水谷信夫 (2006) カメム シ類雄成虫 のフェロモ ンとそれに よる天敵捕 食寄生者の 誘 引. 日本応用動物昆虫学会誌. 50: 87-99. Ozawa, R., G. Arimura, J. Takabayashi, T. Shimoda and T. Nishioka (2000) Involvement of Jasmonate- and Salicylate-Related Signaling Pathways for the Production of Specific Herbivore-Induced Volatiles in Plants. Plant and Cell Physiology. 41: 391-398. Ozawa, R., K. Shiojiri, MW. Sabelis,G. Arimura,T. Nishioka and J. Takabayashi (2004) Corn plants treated with jasmonic acid attract more specialist parasitoids, thereby increasing parasitization of the common armyworm. Journal of Chemical Ecology. 30: 1797-1808. Tentelier, C. and X. Fauvergue (2007) Herbivore-induced plant volatiles as cues for habitat assessment by a foraging parasitoid. Journal of Animal Ecology. 76, 1–8. Thaler, J. (1999) Jasmonate-inducible plant defences cause increased parasitism of herbivores. Nature 399: 686–688. Uefune, M., S. Kugimiya, K. Sano and J. Takabayashi (2011) Herbivore-induced plant volatiles enhance the ability of parasitic wasps to find hosts on a plant. Journal of Applied Entomology. Article first published online. バラのカブリダニを用いた省力防除法の普及 神奈川県立フラワーセンター大船植物園 植栽課 関塚知己 1.背景 平成17年頃、国庫補助金所管部署に居た際に、生産者と普及機関がイチゴの天敵利用に 取り組んだのを手伝い、その効果を実感した。 その直後、普及機関(農業技術センター普及指導部)へ異動になり、若手のリーダー達と 今後の技術について語らったところ、天敵利用に強い興味を示した方が居た。その方と打 ち合わせを重ねて行った取り組みである。 2.経過 今までの農薬使用歴を聞きとりながら戦略を練った。花卉なので食料と違い、減農薬な どが有利販売につながるとは思えず、目的を「春先の収穫増時の防除労力削減」を第1目 標とした。他にこちらから候補としてあげたのは「水を使わないので灰色かび病対策にな る」「薬剤抵抗性発達を遅らせることができる」などを候補としたが、生産者の反応を見 て前述の通り決めた。 また、バラの場合、イチゴと違い、一端植えたら3~5年間はその株を使用するため、 施設内に常にバラがある状態になる。このため、年間ローテーション(春~夏に天敵を使用 し、夏には雑害虫対策の意味もあって影響の長い剤中心、秋から準備に入る)を練った。 天敵を実際に使用する直前に、生産者主催の研究会で天敵を取り上げてもらえることに なり、浜村先生を講師にお呼びした。先生の体系だった話のお陰で、実際に取り組む生産 者は2人増えて3人になった。 なお、この年は3人とも、3~7月頃まで、殺ダニ剤の散布回数がほぼゼロとなった。 3.問題点 初年度こそ、大成功となったが翌年から雨が多い気候となり、うどんこ病が多発した。 このため、硫黄燻煙がネックとなり、最初の3人以外は途中で挫折した。最初の3人も薬 剤散布のみでうどんこ病に対処したので、あまり労力削減にはならなかった。 現在でも、これが天敵利用上の最大の問題である。 4.発展 2年目からは最初に関心を示した生産者(横田氏)が食用バラの生産を始めた。初年度に 余程の手応えを感じたようである。この年からスワルスキーが上市され、横田氏の食用バ ラでの利用にも着手した。苗からのアザミウマ持ち込みがあり、春~夏まではアザミウマ が多発したものの、暑さとともにアザミウマが減り、その後、秋になってもアザミウマの 被害はほとんど発生しなくなった。翌年の春以降も、アザミウマの被害は激減し、時折、 飛び込みが原因と思われる発生が見られたが一時的なもので済んだ。もちろん、コナジラ ミはほぼ全く発生しなかった。 その後も同じ傾向が続いている。なお、「ブルーリバー」という1品種のみアザミウマ が多発したため、農業環境研究部に同定を依頼したところ、ネギアザミウマの寄生だった。 (バラで発生するアザミウマはほとんどがミカンキイロ) 5.その後 最大の問題点は、うどんこ病対策との兼ね合いである。一時期、ケイ素添加を検討した りもしたが、採算が合う効果にはならないようである。ヒートポンプを利用した微気象制 御が他県を中心に普及しつつあるので、これがもっとも効果的だと思われるが、残念なが ら神奈川県内ではヒートポンプの導入が遅れている現状である。 また、横田氏の食用バラ(当然、ほぼ無農薬)生産では最終的な課題としてカイガラムシ の発生に行き着いているようだ。これは有機リン系殺虫剤が禁止されてしまったオランダ でも同様になっており、有機リン系殺虫剤の登録維持は天敵利用の面でも重要だろう。 BIOCONTROL 2011 VOL.15-NO.1 有機栽培で使用できる数種殺菌剤のサヤエンドウの うどんこ病に対する防除効果と残効期間 北海道立総合研究機構 道南農業試験場 生産環境グループ 三澤知央 はじめに 北海道における 2008 年のサヤエンドウの栽培面積は 135ha、生産量は 896t である。道 内では、道南の上ノ国町、道央の石狩市などを中心に全道各地に産地が点在している。 道産サヤエンドウは、高品質であるため市場の評価が高く、東京市場では 8~9 月、大 阪市場では 7~8 月にシェアが 50%を超えている。 サヤエンドウ栽培における最重 要病害はうどんこ病(写真 1)であ り、生産地では本病を対象に多数回 の薬剤散布が実施されている。一 方、近年安心・安全な農作物に対す る消費者の関心が高まり、これに対 応して減化学農薬栽培の技術開発 が全国各地で行われている。北海道 においても、有機栽培および減化学 農薬栽培の技術開発に取り組んで 写真1 サヤエンドウのうどんこ病 きた。しかし、サヤエンドウうどん こ病に対する減化学農薬栽培技術 葉及び茎の発病(左) 莢(がく)の発病(右) については、道内および国内において未確立である。 本稿では、サヤエンドウを含め野菜類のうどんこ病に対して登録を有する薬剤のなか から、有機栽培において使用可能である 5 薬剤について、サヤエンドウうどんこ病に対 する防除効果と残効期間を検討した結果について紹介する。 各薬剤の防除効果と残効期間 試験は 2008~2010 年の 3 年間、北海道北斗市の道南農業試験場露地圃場で実施した。 いずれの年次も、品種「白花砂糖」を供試した。3 年間の播種日および収穫期間は、表 1 に示した。 表1 耕種概要および薬剤散布月日 試験年次 播種日 収穫期間 薬剤散布月日 2008 7/17 8/29~10/15 8/13, 21, 27, 9/5, 12, 19 2009 7/15 8/27~10/22 8/14, 21, 26, 9/3, 10, 18, 25, 10/1 2010 7/21 8/30~10/22 8/6, 13, 20, 27, 9/ 2, 10, 16, 24, 10/1 1 BIOCONTROL 2011 VOL.15-NO.1 試験には、有機栽培で使用できる 5 薬剤および対照薬剤として化学農薬であるトリフル ミゾール水和剤を供試し(表 2),表 1 に示した日程で薬剤散布を行った。 表2 有機栽培で使用できる 5 薬剤の葉・茎・莢に対する防除効果(2008~2010 年) 発病調査月日 2008年 希釈 葉 茎 莢 9/29 9/29 8/29- 9/29 供試薬剤 倍数 発病度 防除価 調査莢数 病莢率(%) 防除価 水和硫黄剤 ×500 0.0 100 0.0 100 500 0.2 99 - 炭酸水素ナトリウム・銅水和剤 ×750 0.9 99 2.9 96 425 0.4 99 - 脂肪酸グリセリド乳剤 ×300 1.1 98 18.6 72 492 0.6 98 ± 炭酸水素ナトリウム水溶剤 ×800 4.4 94 33.9 49 489 1.9 94 - バチルス・ズブチリス水和剤 ×500 7.9 88 38.6 42 414 4.6 86 - 対照)トリフルミゾール水和剤 ×3,000 0.0 100 0.0 100 443 0.4 99 - 382 33.4 無散布 発病割合 防除価 68.2 66.8 薬害 発病調査月日 2009年 希釈 葉 茎 莢 10/7 10/7 8/27- 10/8 供試薬剤 倍数 発病度 防除価 調査莢数 病莢率(%) 防除価 薬害 水和硫黄剤 ×500 0.0 100 0.0 100 763 0.4 100 - 炭酸水素ナトリウム・銅水和剤 ×750 0.3 99.6 11.8 82 905 3.0 97 - 脂肪酸グリセリド乳剤 ×300 0.0 100 4.2 94 732 1.4 98 ± 炭酸水素ナトリウム水溶剤 ×800 6.2 92 22.2 67 813 7.7 91 - バチルス・ズブチリス水和剤 ×500 9.2 88 18.1 73 722 6.8 92 - 対照)トリフルミゾール水和剤 ×3,000 0.0 100 0.0 100 626 0.5 99 - 751 88.6 無散布 発病割合 防除価 74.6 66.6 発病調査月日 2010年 希釈 葉 茎 莢 10/8 10/8 8/30-10/7 供試薬剤 倍数 発病度 防除価 調査莢数 病莢率(%) 防除価 薬害 水和硫黄剤 ×500 0.0 100 0.0 100 672 0.3 99 - 炭酸水素ナトリウム・銅水和剤 ×750 0.0 100 3.5 95 497 1.3 95 - 脂肪酸グリセリド乳剤 ×300 0.0 100 5.5 92 608 1.4 94 ± 脂肪酸グリセリド乳剤 ×600 1.1 98 25.3 64 577 2.2 91 - 炭酸水素ナトリウム水溶剤 ×800 0.3 99 22.7 68 548 2.7 89 - バチルス・ズブチリス水和剤 ×500 7.5 85 36.2 48 631 4.5 82 - 対照)トリフルミゾール水和剤 ×3,000 0.0 100 0.0 100 600 0.6 98 - 446 25.4 無散布 発病割合 防除価 50.2 70.1 本病は葉、茎、莢に発生するため、それぞれの部位の発病状況を調査した。葉の発病 は、野菜類殺菌剤圃場試験法(日本植物防疫協会)の基準に従って調査し、発病度を算出 した。茎の発病は、草丈を測定するとともに、各株から最も上位まで発病した茎を選ん で地際部から連続して白色となった高さ(茎発病到達位置)を測定し、次式により発病 割合を算出した。発病割合=茎発病到達位置÷草丈×100。莢の発病は、全収穫莢につ いて発病の有無を調査し、病莢率を算出した。 2 BIOCONTROL 2011 VOL.15-NO.1 薬剤の最終散布約 1 週間後に発病調査を実施し、調査結果から各薬剤の防除効果を判 定した。さらに最終散布 3 週間後まで調査を継続し、残効期間を判定したが、紙面の都 合上、最終散布 1 週間後のデータのみを掲載した。詳細は三澤(2011)を参照されたい。 うどんこ病の初発は 2008 年は 8 月 26 日(初回散布 13 日後)、2009 年は 9 月 1 日(同 18 日後)、2010 年は 8 月 16 日(同 10 日後)に茎地際部または下位葉で認められ、いずれの 年次も予防的な散布条件下での試験となった。 水和硫黄剤の葉および莢に対する防除価は 99~100 であり、供試薬剤のなかで最も防 除効果が高かった。残効期間は 2 週間であった。 炭酸水素ナトリウム・銅水和剤および脂肪酸グリセリド乳剤(300 倍)の葉の発病に対 する防除価は、99~100 および 98~100、莢の発病に対する防除価は 95~99、94~98 で あり、水和硫黄剤に次いで高い防除効果を示した。残効期間は 2 週間であった。なお、 いずれの年次においても、脂肪酸グリセリド乳剤(300 倍)散布区において、葉および莢 が光沢を帯びる薬害が発生したが、実用上問題ないと判断した。なお、データは示して いないが供試品種「白花砂糖」以外にも「華夏絹莢」および「あずみ野 30 日絹莢 PMR」 の 2 品種においても同様の薬害が発生することを確認している。 炭酸水素ナトリウム水溶剤の葉の発病に対する防除価は 94、92、99、莢の発病に対す る防除価は 94、91、89 であった。同剤は茎の発病において、3 ヶ年とも前述の 3 剤と比 較して顕著に発病が多く、前述の 3 剤より効果が低かった。残効期間は 1~2 週間であ った。 バチルスズブチリス水和剤(インプレッション水和剤)の葉の発病に対する防除価は 88、88、85、莢の発病に対する防除価は 86、92、82 であり、供試薬剤のなかでは最も 防除価が低かった。残効期間は 1 週間であった。 脂肪酸グリセリド乳剤(600 倍)は単年度試験ではあるが、同剤の 300 倍液散布と比較 して防除効果が劣り、その防除効果は炭酸水素ナトリウム水溶剤と同程度であり、残効 期間は 1 週間であった。また、同剤 300 倍液散布でみられた薬害は発生しなかった。 発病調査方法について サヤエンドウのうどんこ病の薬剤試験においては、通常葉の発病のみが調査対象とな っているが、本試験で独自の調査基準を設定して茎、莢の発病も調査した。最終散布 1 週間後における葉、茎、莢の発病に対する防除効果を水和硫黄剤と他剤との間で比較す ると、茎での防除価に顕著な違いが認められた。その上、葉および莢の発病に対する防 除効果の序列と茎の発病に対する防除効果の序列は同じであった。このことから、茎の 発病調査は薬剤間の防除効果の差を検出しやすい調査方法であることが明らかとなっ た。茎の調査は簡便(調査時間は、葉の調査の 10 分の 1 程度)で防除効果の差を検出し やすいため、今後活用が期待される。 莢の発病は主にがくに発生し、直接商品の外観品質を低下させるため、最も被害が大 きい部位である。しかし、 2~3 日おきに収穫した全莢の発病を調査することは、多く 3 BIOCONTROL 2011 VOL.15-NO.1 労力を要する作業である。莢の発病に対する防除価は葉の発病に対する防除価と近似し ていることから、葉の発病調査で代用できると考えられた。 「野菜類うどんこ病」に対して登録のある薬剤の効果 本試験に供試した薬剤については、ニガウリ、イチゴ、ワサビのうどんこ病に対して、 本試験と同様に高い防除効果を示した事例が報告されている(芳賀ら 2010,宮田・増田 2006, 尾松・新屋敷 2010)。一方、トマトのうどんこ病に対する炭酸水素ナトリウム・ 銅水和剤および脂肪酸グリセリド乳剤、ワサビのうどんこ病に対するバチルスズブチリ ス水和剤では、防除効果が低い事例も報告されている(芳賀ら 2010,末長ら 2007)。した がって、本試験に供試した薬剤はいずれも野菜類のうどんこ病に対して農薬登録を有し ているものの、薬剤および作物毎に防除効果が大きく異なることから、個別の試験事例 を積み重ねる必要性が示唆された。 散布体系の確立と現場での活用 本試験に供試した 5 薬剤は、薬剤間で防除効果に差はあるものの、いずれも葉および 莢に対して防除価 82 以上の高い防除効果を示した。さらに,初発後(5 日以内)に薬剤散 布を開始した試験においても、初発前散布とほぼ同程度の防除効果を示した(データ未 記載)。 これらは農薬登録において散布回数に制限がない上、耐性菌の発生報告がないことか ら、耐性菌発生の可能性は極めて低いと考えられる。このため、同一薬剤の連用が可能 であり、各薬剤の防除効果を評価した本試験の散布体系を即現場で活用できる。 これらの薬剤は化学農薬の散布回数としてカウントされないため,北海道が推進する 有機栽培および減化学農薬栽培において活用できる。サヤエンドウのうどんこ病の防除 におけるこれら薬剤の使用は、①高い防除効果を維持しながら、②減化学農薬栽培が実 現でき、③薬剤耐性菌発生リスクを低減できる。そのため、これらの薬剤の積極的な導 入が期待される。 引用文献 芳賀 一ら(2010)関西病虫研報 52:69-71. 宮田將秀・増田俊雄(2006)北日本病虫研報 57:182-184. 三澤知央(2011)北日本病虫研報 62:(印刷中). 尾松直志・新屋敷生男(2010)鹿児島農総セ報(耕種) 4:11-16. 末永寛子ら(2007)岡山県農試県報 25:29-32. 4 「鳴かない虫と音楽」(後編) 柏田 雄三 バッタ目(直翅目) 生誕 350 年以上が経つアラン・マレ(1656~1728)の「異国趣味の音楽(1717 年)」 から「バッタ(La Sauterelle)」。ヴィオールとギターの二重奏、後半にはヴィオール もピチカート奏法を駆使してバッタが飛び交っているような掛け合いがあり何とも 楽しい曲だ。同じくフランスのフランシス・プーランク(1899~1963)の「イナゴ (La Sauterelle) 」。動物小話集(または「オルフェウスのお供」 )中の一曲。詩人が 自らをイナゴと比較し、はたして聖人に食べてもらえるだろうかと謙虚に歌うわずか 20 秒ほどの曲。昆虫食である。歌詞はかのアポリネール。前にも紹介したルーズ・ ランゴー(1893~1952)のピアノ曲集「インセクタリウム」の一曲「Locust」はう ろうろと飛ぶ飛蝗の描写である。 クロード・ラプハム(1891~1957)の「虫の歌」 は赤蜻蛉、蝗、蛍、鈴虫、蝉、蝶々の 6 曲からな るピアノ作品で滞日中の 1934 年に作られた。当 時のアメリカ人が虫の描写を通じ日本人の情感を どのように捉えたかを窺えて興味深い。エリカ・ ヘルツォークさんの弾く「日本の思ひ出」という アルバムで聴いた。「蝗(イナゴ)」は飄々とした 風情の曲である。 我が坂本龍一(1952~)のピアノ曲「バッタ (Grasshoppers)」。ピアノデュオ版が原曲だが、超絶技巧ピアニストとして知られ る岡城千歳さんのソロ編曲で聴く。はじめはバッタの動きを示すような 3 拍子系で、 キース・ジャレットのケルンコンサートを思わせるやや叙情的な中間部を経たのち冒 頭テーマをミニマル音楽風に展開させ 5 分強の曲を終える。虫好きだという山田栄二 (1948~ )の「ファーブル昆虫記」は 8 本のホルンのための組曲。名手アンサン ブル「つの笛集団」の CD を聴く。その中の「バッタ」では大発生したバッタが植物 を食い荒らしながら進軍する。 フィンランドのカレヴィ・アホ(1949~)の「Insect Symphony」と名づけられた 交響曲第 7 番には全楽章虫の名前が付されていて、第 4 楽章は「The Grasshoppers」。 曲はカレル・チャペックの戯曲「虫の生活から」から発想されている。戯曲中での虫 はコオロギだから、鳴かない虫として取り上げるのは妥当でないかもしれない。 カマキリ目(蟷螂目) オーストラリアの作曲家フィリップ・ホートン(1954~)の「月とカマキリ」はギタ ーのための二重奏曲である。なぜカマキリなのかライナー・ノーツを見ても判らない。 山田栄二「ファーブル昆虫記」の「ウスバカマキリ」はピアソラ風のユーモラスなタ ンゴである。 ハエ目(双翅目) フィニ・ヘンリケス(ヘンリク) (1867~1940)はデンマークの作曲家である。そ の手になる「蚊の踊り」は 1 分半ほどの小曲で、管弦楽をバックにクラリネット(ま たはリコーダー)が活躍し、リムスキー=コルサコフの「熊蜂の飛行(マルハナバチ の飛行)」と同様に無窮動風の音の連なりにより蚊の飛ぶ様を描く。 ロシアの作曲家アレクサンダー・スクリャービン(1872~1915)には「蚊」の通 称を持つ 1 分程のピアノの練習曲作品 42-3。蚊を連想させるトリルが難しそうだ。 ハンガリーのベラ・バルトーク(1881~1945)の「2 つのヴァイオリンのための 44 の二重奏曲」中の「蚊の歌」は更に短い 30 秒ほどの曲で、のんびりと蚊が飛ぶ様子 が示される。モデスト・ムソルグスキー(1839~1881)の「ボリス・ゴドゥノフ」は 彼の天才ぶりを感じさせる傑作歌劇である。この中のアリア「蚊の踊り」はロシア皇 帝の皇子と皇女の乳母がひょうきんに歌う。蚊、南京虫、トンボが登場し、南京虫の 振り回した薪に当たった蚊が肋骨を 3 本折ってしまったというたわいのない内容で 伴奏のヴァイオリンが蚊の羽音を擬している。椎名誠氏著の「蚊學ノ書」によると、 シベリアの蚊は大きさや発生数が日本の常識では考えられないものらしいので、肋骨 を折る蚊がいても不思議ではない気がしてくる。この本では蚊について薀蓄がさまざ ま語られるが残念なことに音楽との関連は薄く、栗原毅氏著の「蚊の博物誌」でも同 様である。 メル・ボニス(1858~1937)をご存じだろうか。時折名前を目にするようになっ たフランスの女性作曲家でピアノ曲の「蚊」。めまぐるしいがダンスのようでこれな ら刺されても悔しくなさそうだ。 ブラジルが生んだ南米最高のクラシック音楽家エイトール・ヴィラ=ロボス(1887 ~1959)の「蚊の踊り」。大規模な管弦楽曲で、蚊が飛び回る様子を表す。ホルンが 音を大きく跳躍させて活躍し演奏がすごく難しそうだ。蚊が飛んでイライラさせる様 子はヴァイオリンの高音でよく表現されている。 チェコのレオシュ・ヤナーチェク(1854~1928)のオペラ「利口な女狐の物語」に も蚊が登場する。このオペラには人間と動物、昆虫が一緒に出てくるので自然賛歌だ と言われるが、こういうことにかけては日本だって負けていない。音楽ではないが、 狂言の「蚊相撲」は大名が「蚊の精」と相撲を取ると言う奇想天外な内容だ。野村萬 斎が大名役を演ずる舞台を遠くまで観に出かけたが実に面白かった。蚊の精が「ぷー ん」と言いながら大名を刺しに行く場面では客席から笑い声が上がる。大名は相手の 男が江州守山の出身だと聞いただけで正体を蚊の精だと見破る。琵琶湖を囲む地域は 昔マラリアの多発地帯だったことが知られている。蚊の名所という訳なのだ。 喧しい若者がたむろしないよう、彼らにしか聞こえない高周波数の音(17,600Hz) を発する装置がイギリスの街角に設置され賛否を呼んでいる。このような高音はモス キート音と呼ばれ、この装置は日本でも「モスキート」という名前で販売されてコン ビニや公園などに試験的に設置されているようである。 人間の耳は最高で 20,000Hz 位までしか聞こえないとされており、CD の規格が最高 20,000Hz となっている理由の一つである。加齢により高い周波数の音がだんだんと 聞こえ辛くなるというので、どのくらいの高さまで聞こえるかをテスト CD で試し、 その結果に愕然としたことがある。精神衛生上あまりお勧めはできないが、耳が若い と自信をお持ちの向きは試してみられたい。 音楽の再生に 20,000Hz より高い周波数が必要でないという考えには異論もあり、 再生周波数の範囲が 100,000Hz までと広くダイナミックレンジも大きい SACD が製 品化されている。最近インターネット上で従来のマルチビットのリニア PCM 方式で はなく DSD 規格による配信が始まり、環境を整えれば SACD に匹敵する音が簡単に 入手できると言う大変な時代になった。耳で直接聞こえない周波数の存在によってな ぜ音が良くなるのか、倍音成分が影響しているといわれるがそれを論ずる場ではない ので先を急ごう。 蚊の音楽はどのぐらいの高さで演奏されるのだろうか。因みに 88 鍵のピアノの最 高音 C8 は 4,186Hz なので、うるさい若者撃退装置の周波数より 2 オクターブ以上低 いことになる。 安富和男氏の著書によると蚊の羽音周波数は 350~600Hz と意外に低いのである。 種や雌雄によって周波数が決まっていて、メスでは、コガタアカイエカ 380Hz、シナ ハマダラカ 340Hz、ヒトスジシマカ 512Hz だそうだ。 「モスキート音」の命名は過大 (過高)評価されていることになる。いわゆる鳴く虫の音は大体が 4,000~6,000Hz なのでこちらのほうがずっと高い。 バルトークによるピアノ曲集「ミクロコスモス」に「蝿の日記より」という小曲が ある。 「ミクロコスモス」は 153 の小曲からなる子供のための練習曲集で、6 つの巻 に分かれており後になるにつれ演奏が難しくなる。これは最も難しい第 6 巻に収録さ れた 142 番目の曲である。わずか 1 分半ほどの曲で、ぶんぶん飛ぶハエの音を密集し た位置で白鍵と黒鍵を二つの声部に分けて運動させるという方法で表している。弦同 士が共鳴するような不思議な響きであるが、ソステヌートペダルを多用しているのか もしれない。磯崎敦博(?~ )の「虫の謝肉祭」はクラリネットアンサンブルの曲。 虫の名前の童謡やクラシックの曲が多数メドレーでごく短時間ずつ現れ、「蠅の日記 より」と後述の「蚤の歌」も出て来る。なんとまあという印象だ。ポーランドのアレ クサンドル・タンスマン(1897~1986)の「ピアノを弾く子供たち」の第 1 巻「ミ レイユと動物たち」 。ミレイユとは愛嬢の名前である。その第 6 曲「小さな犬とハエ」。 彼は瞠目すべき描写力を持つ作曲家で、この曲では犬が吠えながらハエをくるくると 追いかける様子が目に見えるようだ。 曲の名前には出ないが、アントニオ・ヴィヴァルディ(1678~1741)の有名な「四 季」の夏の第 2 楽章ではハエが描かれる。四季の各楽章にはソネット(十四行詩)が 添えられている。この楽章は農夫がハエの群に脅かされるという内容で、曲ではわん わんと五月蝿い様子がヴァイオリンで奏される。木下牧子(1956~)の歌曲「一群の ぶよ」は虫の飛ぶ様を示す伴奏部分がプジョールのギター曲「くまんばち」に似てい るが、ヴィヴァルディのハエとは異なるややおっとりした雰囲気を漂わせている。 ルイ 14 世から 15 世の時代ヴェルサイユ宮殿で活躍した音楽家フランスのフランソ ワ・クープラン(1668~1733)はチェンバロのための曲をたくさん作っており、そ のうちの一曲に「羽虫」がある。ここで言う「羽虫」の原題は「Le moucheron」、 「あ ぶ」のことらしい。翅の動きを擬した小曲である。 ポール・モーリス(1910~1967)と言うフランスの女性作曲家による「プロヴァン スの風景」中の一曲「あぶ」。サキソフォンのためのオリジナル曲で、無窮動風に 3 分以上も飛び回る。プロヴァンスの抜ける青空が見えてくるようだ。 一転してソ連のドミートリイ・ショスタコーヴィッチ(1906~1975)に「馬あぶ」 という映画音楽がある。15 曲にも及ぶヘヴィーな彼の交響曲は苦手なのだが、映画 音楽は別の世界だ。この「馬あぶ」の中の「ロマンス」や「序奏」の美しいメロディ を初めて聴いて、誰が作曲者の名を当てられようか。 高柳未来さんが演奏する「マンドリン・レボリュ ーション」という題名のアルバムに齋藤秀雄作曲 の「蚊トンボ」という曲を見つける。齋藤は音楽 教育家や演奏家として近代日本音楽史に巨大な足 跡を残した人である。 彼はチェロと並んで若いころ「オルケストル・エ トワール」と言うクラブを作りマンドリンの演奏 も熱心に行っていたのだ。 「蚊トンボ」とはもちろんガガンボのことである。 この CD にはベートーヴェンの秘曲ともいえるマ ンドリンのオリジナル曲、ギターのラスゲアード様の奏法を取り入れたフラメンコ調 の曲、そして何と萩原朔太郎のオリジナル曲なども収録されていて興味が尽きない。 この「蚊トンボ」には「Kleine Libelle」と言う曲名が併記されているのだが、これ ではガガンボではなく小さなトンボになってしまう。発売元に問い合わせたところ、 齋藤の草稿は日本語表示のみで、ドイツ語はあとから誰かによって付け加えられたが、 それが作曲者本人によるかどうかはわからないとのこと。マンドリンの二重奏により ガガンボの飛ぶ様を表したおおらかな曲、手で叩かれるような音を経て終わる。ラン ゴーの「インセクタリウム」中の一曲「ガガンボ」ではすこし頼りなげに飛ぶ。こち らもガガンボの風情そのものである。 ノミ目(隠翅目) まずはムソルグスキーの手になる「蚤の歌」。20 世紀最高のバス歌手といわれたシ ャリアピンの歌唱でこの曲を知った。 「ウフフフフ・・・」という印象的な笑い声の部分を聞き、江戸時代の川柳「蚤一つあ ったら娘を裸にし」 ( 「蟻一つ娘盛りを裸にし」ともいう)と同様に蚤に刺された男が 痒さを堪える様子を歌っているものとばかり思っていた。この曲の歌詞はゲーテの 「ファウスト」を基にしたもので、王様が蚤を大臣に取り立て、勲章まで与えて可愛 がったという風刺の歌だった。正式には「アウエルバッハの酒蔵でのメフィストフェ レスの歌」と言う曲名で、学生が歌う「鼠の歌」を受けて悪魔のメフィストフェレス が歌う。 同じ詩の歌をルードヴィヒ・ファン・ベートーヴェン(1770~1827)は歌曲とし て、ルイ・エクトル・ベルリオーズ(1803~1869)は劇的物語「ファウストの劫罰」 の中でのアリアとして作曲している。 ベートーヴェンの曲「メフィストの蚤の歌」 (1809 年)はピアノの前奏部分に蚤を 思わせる細かい音の動きがあり、最後もユーモラスに終わらせようとするが、糞真面 目な男が慣れない冗談で笑わせようと無理しているような印象を受ける。 ベルリオーズの曲も歌詞にこめられた毒の部分が薄められており、やはりムソルグ スキーに軍配が上がる。天才のなせる業だ。或いは彼の命を縮めた酒毒のなせる業だ ろうか。ムソルグスキーの「蚤の歌」の歌詞は、ストルゴフシチコフという人がロシ ア語訳したもので、有名な「ウフフフフ・・・」の部分はもとには無く、ムソルグスキ ー自身が作曲のときに付け加えたと言われる。 初めて名前を聞くロシアのユリ・シャポーリン(1887~1966)の「ノミ」という曲 が入った CD を見つける。出だしはサーカスのジンタ風で蚤が跳ねる様子を表したよ うに聞こえる。6 曲の小曲からなり、マンドリンを単弦にしたようなドムラやアコー ディオンの兄弟分バヤーンという民俗楽器が活躍して気張らずに聴ける。この曲は 10 枚組の CD 中の一曲として収録されていて、この曲だけのために苦手なショスタ コーヴィッチの交響曲などが大半を占める CD セットを求めるのは酔狂としか言いよ うがないが、昆虫の珍しい曲を聴くためなので致し方ない。 ぐっと時代をさかのぼると、蚤の生態をよく描写した音楽にジョゼフ・ボダン・ド・ ボワモルティエ(1689~1755)のチェンバロ曲「のみ」 。リュート・ストップと言う 演奏法を使って蚤の飛躍を表す。ユーモラスな小曲である。 河盛好蔵のエッセイ「パリの芝居」には 19 世紀末パリのカジノ座で「蚤」とい うパントマイムが上演されたことが書かれている。アンジェール・エラールという女 優が着物を脱いでノミを捜しまわるストリップで、大当たりしてヨーロッパ中を興行 したそうだ。先の川柳を地で行くような内容で、上演されれば観てみたいものである。 シラミ目(咀顎目、裸尾目) 團伊玖磨(1924~2001)は本職である作曲家のほかに随筆家としても名高い。代 表作「パイプのけむり」シリーズは題名をたくみに工夫して全 27 巻にも及んだが、 その第 1 巻「パイプのけむり」 (朝日新聞社)に「ソラ・ソラ・シラミ」という一文 がある。陸軍軍楽隊の上等兵であった團と芥川也寸志の二人が机の上をシラミが這っ ているのを見て曲を作ったことが書かれている。シラミの三つの文字が音階の中に含 まれているのに気付いたという訳だ。その歌詞は、 「そらそら虱! どらどら虱! 見れど見れど虱! 虱!」 自分で適当に歌ってみると、シラミの部分がロシア風で趣がある。 さだまさしには「シラミ騒動」という同趣向の曲がありこちらのほうが長い。このよ うな曲は階名ソングと呼ばれている。曲と歌詞が同時にできるわけだ。 甲虫目(鞘翅目) 最大の種類数を誇る甲虫目であるが、曲の数はチョウ目に及ばないようである。 ムソルグスキーの歌曲集「子供部屋」に「カブトムシ」という一曲。カブトムシが飛 んできて自分に当たって動かなくなったがどうしたのだろうという子供らしい可愛 い歌詞の曲である。デパートで買ったカブトムシが死んでしまうと電池を入れ替えて くれと親にせがんだりする最近の子供とは違う。ブルガリアの名バス、ボリス・クリ ストフの歌唱で聴く。子供の気持を思っての歌い方だが、不自然でもある。 ヴィラ=ロボスのピアノ曲中の「赤ちゃんの一族」という組曲。ここで言う「一族」 とは赤ちゃんの親や兄弟ではなく、赤ちゃんが遊ぶおもちゃなどを指す。第 1 集は「お 人形たち」、第 2 集は「小さい動物たち」、楽譜が失われた第 3 集は「遊び」がテーマ である。第二集「小さい動物たち」は大胆な不協和音が使われた難曲だ。その冒頭が 「紙のカブトムシ」。第二集は第一集と異なり描写よりも暗示に重きが置かれている とされるが、この曲に限っては右手の 16 分音符の細かい音型が虫のぎこちない動き を示すようで、ある程度描写的である。ヴィラ=ロボスは代表作「ブラジル風バッハ」 や「ショーロス」、ギター音楽の数々で我々に親しいが、このようなピアノ曲を聴い て終生個性的であろうとした彼の音楽の一端に触れるのもよいだろう。 ヨハン・シュトラウス二世(1825~1899)のワルツ「テントウムシ」 。彼は膨大な 数のワルツやポルカを残した。有名でないこの曲は、CD52 枚組セットの中でしか聴 くことができず、廉価盤であるとはいえかなりの勇気をもって求めた。このセットに はほかに「蛍」「蝶」「蛾」がある。「テントウムシ」は彼の多くの曲と同じくこれと いった特徴がない。 「カルメン」や「アルルの女」で有名なジョルジュ・ビゼー(1838~1875)には「テ ントウムシ」という歌曲。この曲をテノール歌手武田正雄氏の「わが動物詩集」と銘 打ったリサイタルで聴く。詩はヴィクトル・ユーゴー。「彼女にうなじに止まったテ ントウムシを取った代わりに彼女とのキスを逃がしてしまった」というほほえましい 少年の歌だ。「テントウムシ」のほかにフランスの作曲家たちによる昆虫や動物の歌 が何曲も歌われたこのリサイタルは大変楽しかった。ランゴーの「インセクタリウム」 には甲虫目として「Cockchafer」と「Death Watch Beetle」の二種類が取り上げら れている。特に「Death Watch Beetle」でピアノの蓋を叩くという特殊奏法によりシ バンムシが木の中で立てる音をあらわしていることは以前も書いた。 ヨハン・シュトラウス二世のワルツ「蛍」を先ほどの CD セットで聴く。「蛍」と は言ってもヨーロッパの蛍はゲンジボタルなどと異なり翅のない成虫だけが光る種 類のようで、この曲名も Johannis-K äferln(Glow-Worms)である。調べると優美 な姿とは言い難い。曲は光る様子を表したのかもしれないが個性的でない。 アメリカの作曲家ルドルフ・フリムル(1879 ~ 1972)には、オペレッタの「蛍(The firefly) 」。序曲のみやっと手に出来たがこれといった特徴が無くがっかりだ。 フランスのウジェーヌ・ボザ(1905~1991)の 6 本のクラリネットのための「蛍」。 「熊蜂の飛行(マルハナバチの飛行)」にヒントを得た曲らしく、ドビュッシーが混 合されたような洒落た響きだ。でも蛍にしては少し忙しい。三連符の高速の音型の受 け渡しが難しいそうだが、それを感じさせないアンサンブルである。 バッタ目で紹介したラプハムの「蛍」。空中で明滅を繰り返しながら飛ぶ蛍の様子 が表された美しい曲である。 山田栄二「ファーブル昆虫記」には「ツチハンミョウ」と「ムナゲモンシデムシ」。 前者は不格好な姿でのそのそと歩き回る様子、後者はスカベンジャーの葬送の様子で ある。上手いものだ。アホの交響曲第 7 番第 3 楽章は糞虫(The Dung Beetles)だ。 橋本國彦(1904~1949)には傑作歌曲 「斑猫」がある。彼の歌は親しみやすい「朝 はどこから」や「お菓子と娘」ばかりでは ないのである。 曲は「斑猫です・・」に始まるハンミョウ の形態と生態とを巧みに織り込んだ深尾 須磨子の歌詞を前衛的な旋律が包み込み 離れ、また無調に傾いたりもしながら、思 いがけぬ弧を描いて進む。彼の「黴」とと も に 日 本 歌 曲 の 大 き な 転 機 に な っ た と 言 わ れ る 名 曲 で あ る 。 (写真 有吉立) そのハンミョウの口器の拡大写真を見ると、まるで鋼鉄で出来ているようだ。肉食だ けあってのんびりした歌詞とは異なる凶暴な姿に思わずたじろぐ。 これで 2 回にわたった「鳴かない虫」に関する音楽の紹介を終わる。その多くは有 名曲の花籠からは零れた曲であった。誌面の都合で紹介できなかった曲もあるし、原 稿締め切り後に出会った曲もある。カゲロウ目、カメムシ目(半翅目)、ハサミムシ 目(革翅目)の曲は割愛した。それらの紹介は他の機会に譲りたい。 「虫と音楽」 (バイオコントロール 2008 Vol.12 No1)の文中で Mosquito の学名 は Culex pipiens が正しいことを指摘したが、その後技術士の安東和彦氏から Cockchafer の属名は Melonetha ではなく、Melolontha であるとの指摘を受けた。 ㈱キングインターナショナルの音楽プロデューサー宮山幸久氏からランゴーの「イン セクタリウム」の貴重な楽譜(The Society for Publication of Danish Music & The Royal Library 発行)を見せていただく機会を得た。蚊については pipiens と正しい 種小名が、コフキコガネについては Melonetha と誤った属名が記載されていた。し かし作曲者がどのように綴ったのかは判らない。 安東氏と宮山氏、写真を提供してくださった有吉氏に謝意を表して筆を擱きたい。 (元武田薬品工業株式会社) <<殺虫・殺ダニ剤>> 種類名 アカリタッチ アクタラ(粒) アクタラ(顆粒水溶) アクテリック ア-デント アグロスリン アタブロン アニキ アディオン アドバンテ-ジ(粒) アドマイヤアドマイヤ-(粒) アファ-ム アプロ-ド ウララDF エビセクト エンセダン オサダン オマイト オリオン オルトラン(水) オルトラン(粒) オレ-ト ガ-ドサイド ガードホープ(液剤) カ-ラ カスケ-ド ガスタ-ド(粒) カネマイト カルホス キルバ-ル クロ-ルピクリン ケルセン コテツ コロマイト コロマイト(EC) コロマイト(WP) サイハロン サニ-フィ-ルド サブリナフロアブル サンクリスタル乳剤 サンマイト ジェットロン ジメトエ-ト 除虫菊乳剤 シラトップ スカウト スタ-クル スピノエ-ス スプラサイド スミサイジン混剤 スミチオン ゼンタ-リ ダ-ズバン ダイアジノン(乳・水) ダイアジノン(粒) ダイシストン(粒) ダニカット ダニサラバ ダニトロン ダントツ チェス D-D D-D92 DDVP(乳) DDVP(煙) ショクガ タマバエ 幼 - - - - - × - - × - × ◎ - △ - - - ○ - - - - - - - - - - - - - ◎ - - - 成 - - - × - × - ◎ × - × ◎ - △ - - ◎ ◎ - × - - × - ◎ ◎ - - - - - × - - ◎ 残 - - - - - 84 - 0 84 - - 0 - 7 - - - - - 28 - - - - - - - - - - - - - - 0 - - - - - - △ - - - - - - × - - - × - - - - - - ◎ - - - - - - - - - ○ × - - - - △ × - - × × - - - - - - ◎ - - × - - - - - - - 14 - - - - - 84 - - - 56 - - - - - - 0 - - 7 - コレマン アブラバチ マ ◎ - - × - × ◎ 成 ◎ - - × - × ◎ 残 0 - - - - 84 0 × - × ◎ ◎ ◎ ◎ ○ - ◎ ◎ - - - - - ◎ ◎ - - ◎ - - - - - - × - × ◎ × ◎ ◎ × - ◎ △ - × - - × ◎ ◎ ◎ - - - - - △ - - 84 - - 0 7 0 0 - - 0 - - - - - - 0 0 - - - - - - - - - - - - - - ◎ × - - - - - × × - - × × - - - - - - ◎ - - - - - - ◎ ○ × ◎ × × - - - - × × - - × × - - - ◎ - - ◎ - - × - - - - 0 - 0 - - - - - - - 84 - - - - - - - - - - 0 - - - - 天敵等への殺虫・殺ダニ剤の影響の目安 ミヤコ カブリダニ 卵 ◎ - ◎ - - - ◎ - - - ◎ ◎ × ◎ ◎ - - ◎ - - - - ◎ - ◎ ◎ △ - ◎ - - - - - - - - - - - - - - - - - - ○ △ - - - - - - - - - - ◎ ◎ - - ◎ - 成 ○ - ◎ × - - ○ × △ - ◎ ◎ × ◎ ◎ - - ◎ △ - × - ◎ - ◎ ◎ ◎ - ◎ - △ - × - ○ △ △ - - - ○ △ - × - - - - ◎ × - - - △ ○ - - × ◎ ◎ ◎ ◎ - - ◎ - 残 - - 0 - - - 9 3 - - 0 0 - 0 - - 0 - - 21 - 0 - 0 0 - - 0 - - - - 7 - 1 1 - - - 0 - - - - - - - - - - - - 14 14 - - 21 - - 0 0 - - 0 - チリ カブリダニ 卵 ◎ - ◎ × - × ◎ 成 ◎ - △ × - × ○ 残 0 - - 28 - 84 1 × ○ ◎ ◎ × ◎ ◎ - - ◎ × - - - ◎ ◎ ◎ ◎ ◎ - ◎ - - - △ - - × ○ ◎ ◎ × ○ ◎ ○ - ◎ △ - × - ◎ ○ ◎ ◎ ◎ - ◎ - × - △ - △ 84 7 0 0 - 0 7 - 0 0 - 28 - 0 0 0 0 0 - 0 - - - 14 - - - - - ○ - - × ◎ - - - ○ × × - - ◎ ◎ - - × - - ◎ ◎ - - ◎ - - - - ○ - × × × - - - ◎ × × × - △ ◎ - - × - - ○ ◎ - - × - - - - - - * 56 7 - - - - 21 84 - - 7 7 - - 21 - - - 0 - - 7 - ククメリス カブリダニ 卵 ◎ - - × × × ◎ - × - ◎ ◎ ◎ ◎ ◎ - - ◎ - × × - - × ◎ ◎ ◎ - - - - - - ◎ - - - - - - - - × ◎ - - - - × × × - × ◎ - - - - - - ◎ - - ◎ - スワルスキ-カ タイリクヒメハナカ ブリダニ メムシ 成 残 卵 - - - - - - - - ◎ × 56 - × 21↑ - × 84 - × 9 - × 3 - × 84 - - - - ◎ 0 △ ◎ 0 - ○ 6 - ◎ 0 - ◎ - ○ - - - - - ◎ 0 ◎ × - - - - - × 28 - - - - - - - × 56 - ◎ 0 - ◎ 0 - ◎ 0 ◎ - - - - - - - - - - - - - - - × 28 - × 6 - - - ○ × 7 - × - - - - - - - - - - - ○ 0 - × - × - - - × 84 - × 7 - - - - - - - - - - - - × × 56 - × 84 - × 56 - - - - × 56 - × 21 - - - - - - - × 28 - ◎ - - - - - - - - ◎ 0 ◎ - - - - - - × 7 - - - - 成 - - ◎ - - - - × - - ○ - - ○ - - - ◎ - - - - - - - - ◎ - - - - - - - × × × - - - - △ - - - - - - × - - - - - - - - - - - - ◎ - - - - 残 - - - - - - - 3 - - - - - - - - - - - - - - - - - - 0 - - - - - - - - 1 1 - - - - - - - - - - - - - - - - - - - - - - - - - - - - - 幼 ◎ - - - - × × - × - × - - ◎ ◎ - × ○ - × × - - × ◎ - △ - - - × - ◎ ◎ - - 成 ◎ - - - - × × ◎ × - × - × ◎ ◎ - × ○ △ - × - - × ◎ ◎ ◎ - - - × - ◎ ◎ - ◎ 残 0 - - - - 84 14↑ 0 84 - 14↑ - 7 0 0 - 56 0 - - - - - - 0 0 28 - - - - - 0 0 - 0 - - ◎ ◎ × × ◎ - - - - × × - - ○ × - - ○ ◎ ◎ - ○ - - △ - - - ◎ ◎ × - × ◎ - - - - × × × - × × - - △ - ◎ - ○ - - × - - - - - 14 - - 0 - - - - 14 84 - - - - - - 21 - 0 - - - - 7 - アリガタシマ アザミウマ オンシツ ツヤコバチ 幼 ◎ - - - - - - 成 ◎ - - - - - - 残 0 - - - - - - - - △ - × ◎ - - ◎ - - ◎ - - - - - × - ◎ - - - - △ ◎ - - △ - × ◎ - - - ◎ - - △ - - - - - ◎ - ◎ - - - - △ × - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - × - - - ◎ - - - - - - ◎ - ◎ - - × - - - - - - - - - - - - △ - - - ◎ - - - - - - ◎ - ◎ - - × - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - サバク ツヤコバチ 蛹 ◎ - - × - × ◎ ○ × - ◎ ◎ - ○ ◎ ○ ◎ ◎ △ - × ○ ◎ × ◎ ◎ ◎ - - × - - ◎ - - - 成 ◎ - - × - × ◎ × × - △ × × ◎ ◎ ○ - ○ △ - × × ◎ × ◎ ◎ ◎ - - - × - × - - - 残 0 - - 56 - 84 0 28 84 - 35 30 21 7 0 7 - 0 7 - 28 30 0 84 0 0 0 - - - - - 14 - - 1 蛹 - - - - - × - ◎ × - - ◎ - ◎ - - ◎ - - × - - - - ◎ ◎ - - - - - - - - ◎ 成 - - - - - × - - × - - ◎ - ◎ - △ ◎ - - × - - - - ◎ - - - - × - - - - - 残 - - - - - 84 - 0 84 - - 0 - 0 - - 0 - - 28 - - - - 0 - - - - - - - - - 0 - - ◎ - △ ○ × ◎ - - - - × × △ - △ ○ - - × ◎ - - ◎ - - × × - - ◎ - × ◎ × × - - × × × × × - × × - - × - - - ◎ - - × × - - - - 21 0 84 3 - - - 42 56 84 56 - 84 42 - - 21 - - - 0 - - 10 10 - - - - - - - - - - - - - × - - - - - - ○ - - - ◎ - - - - - - - - - - - - - - × - - × - - - - - - ○ - - - ◎ - - × - - - - - - - - - - - - - - 84 - - - - - - 14 - - - 0 - - 7 - チチュウカイ ツヤコバチ 蛹 - - - - - - - 成 - - - - - - - 残 - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - × - × ◎ - - - - - - - - - - - ◎ - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - △ × - - - - - × × - - - - - - - - - - - × ◎ - - - - - - - 0 - - - - - - - - - - - - - - - - - - - - - - - - - イサエアヒメコバチ ハモグリコマユバチ デジェネランス カブリダニ クサカゲロウ 類 幼 ◎ - - - - - ◎ - - - ◎ - - ◎ ◎ ○ - ◎ - - - - ◎ - - ◎ ◎ - - - - - - - - - 成 ◎ - - × - × ◎ ○ × × × ◎ × ◎ ◎ × - ○ △ - × × ◎ × ○ ◎ ◎ - - × - - △ × - - 残 0 - - - - 84 0 3 84 - 14 21 - 0 0 - - 0 - - 28 49 - - 22 0 0 - - 49 - - - - - 3 卵 - - - × - × - 成 - - - × - × - 残 - - - 56 - 84 - 幼 - - - × - × - 成 - - - × - × - × - ◎ ◎ - ◎ - - ◎ - - × - - × - ◎ - - - - - - - - - × - ◎ ◎ - ◎ ○ - △ × - × - - × - ◎ ◎ - - - - - × - - 84 - 0 0 - 0 - - 0 - - 28 - - 56 - 0 - - - - - - 28 - - × - × ◎ - △ ○ × ◎ ◎ - × - - - - ◎ △ - ◎ - △ - ○ - - - - ◎ - ○ - - - - - - - - × - - × - - - - - - - ◎ - - - - - - ◎ - △ - × - - - - × × - - × × - - - - × - ◎ - - × - - - - - 21 - - 7 - - - - - 84 - - - - - - - - - - 0 - - 7 - - - - - - - × ◎ - - - - × × × - × ◎ - - - - - - ◎ - - ○ - - - - - × × × - - - - × × × - × × - - × - - - ◎ - - × - - - - - - - 84 7 - - - - 56 84 56 - 56 21 - - 28 - - - 0 - - 7 - - - ◎ ◎ - × ○ - - - - × × △ - × × - - - - - - ◎ - - × - 日本バイオロジカルコントロール協議会・2011年9月作成・第20版 ヨトウタマゴバチ類 ネマト-ダ 類 ボ-ベリア バシア-ナ バ-ティシリウム レカニ バチルス ズブチリス エルビニア カロトボ-ラ シュ-ドモナス フルオレッセンス 残 - - - 56 - 84 - 蛹 - - - × - × - 成 - - - × - × - 残 - - - 28 - 84 - 幼 - - - - - ◎ - 残 - - - - - 0 - 分生子 ◎ - 胞子 - - - ◎ - ◎ - 芽胞 ◎ - ◎ ◎ ◎ ◎ ◎ 菌 - - - - ◎ ◎ × 菌 ◎ × - 巣 - × × × × × × 残 - 21 42 14 3 20↑ 4 × - × ◎ - △ - - - ◎ - × - - - - ◎ × - - - △ - ○ - - 84 - - 0 - 7 - - - - - 28 - - - - 0 - - - - - - 7 - - × - - ◎ - ◎ △ - ◎ ◎ - ○ - - - - ◎ - - - - ◎ - △ - - × - - ◎ - ◎ △ - ◎ ◎ - × - - - - ◎ ◎ - - - × - △ - - 84 - - 0 - 0 14 - - - - - - - - - 0 - - - - 28 - 21 - - ◎ ○ - - - ◎ - ○ - ◎ - ◎ - - - - - - - - 0 - - - 0 - 0 - - - ◎ ◎ ◎ ◎ - ◎ ◎ - - - ◎ ◎ - ◎ × - △ - - - ◎ - ◎ - ◎ ◎ - - - ◎ - - ◎ - ◎ - 水◎乳× ◎ ◎ ◎ - - ◎ - - - ◎ ◎ ◎ - - ◎ × × ◎ - × × × × △ ○ × × ○ ◎ - × × ○ × 20↑ 21 30↑ 35↑ 2 1 3 - 1 - 10~20 14~30 1 14 - - - - ○ - - ◎ - - - - - - 30 - - 0 - - ◎ ◎ ◎ ◎ - - - ◎ - ◎ - ◎ - ◎ - ◎ - - - ◎ ◎ ◎ - - - - ◎ × - ◎ ◎ × ◎ - ○ △ × - × × × ○ × - 1 2 21 - 14 - 28 1 9 - - - ◎ - ◎ × ○ - - - - × × × - × × - - ◎ - - - ◎ - - × - - - - - - - 84 7 - - - - 56 84 - - 84 28 - - - - - - 0 - - 7 - - - - ○ - - × - - - - - × × × - × × - - ○ - - - ◎ - - - - - - - - - - × × - - - - × × × - × × - - × - - - ◎ - - × - - - - - - - 42 - - - - - 28 84 70 - 28 14 - - 28 - - - 0 - - 7 - - ◎ - - - ○ - ◎ - - - ◎ ◎ ○ - ○ ○ - - ◎ - - - ◎ - - ◎ - - 0 - - - - - 0 - - - - - 1 - 14 14 - - 0 - - - 0 - - 0 - - ◎ ◎ ◎ ◎ ◎ × ◎ ◎ ◎ ◎ - - - - - - ◎ - - - - - × ◎ × - △ × - - △ - - - - - - ◎ - ◎ - - - ◎ - ◎ ◎ - ◎ ◎ ◎ ◎ - ◎ ◎ - ◎ - - - - ◎ ◎ ◎ - - ◎ - 水◎乳× - - ◎ - - × - ◎ ◎ ◎ × - - - - ◎ - - - - - ◎ ◎ - - × - - ◎ ◎ ◎ ◎ ◎ - × - - × ◎ × △ - △ - × × × × - × × × ○ ◎ ◎ ○ - ◎ × × × × 4 - - 0 1~4 0 20↑ 2 - 2 - 3~7 30 30↑ 20↑ - 30↑ 15~30 30 1 マルハナバチ - 1 - 28 28 7↑ 7 <<殺虫・殺ダニ剤>> 種類名 ディトラペックス ディプテレックス テデオン デミリン テルスタ-(水) テルスタ-(煙) デルフィン水和剤 トクチオン トリガ-ド トレボン ニッソラン ネマトリン ネマトリンエース(粒) 粘着くん ノ-モルト ハチハチ ハッパ乳剤 バイスロイド バイデ-ト(粒) パダン バリア-ド バロック BT剤 ピラニカ ファルコン プレオ プレバソン フェニックス ペイオフ ベストガ-ド(水) ベストガ-ド(粒) ペンタック マイコタ-ル マイトコ-ネ マシン油 マッチ マトリック マブリック(水) マブリック(煙) マラソン マリックス ミクロデナポン Mr.ジョ-カミルベノック モスピラン(水) モスピラン(煙) モスピラン(粒) ラ-ビン ラノランネ-ト リラ-ク ルビトックス レルダン ロディ-(乳) ロディ-(煙) ロムダン ショクガ タマバエ コレマン アブラバチ 天敵等への殺虫・殺ダニ剤の影響の目安 ミヤコ カブリダニ チリ カブリダニ 幼 - - - ◎ × - - - - - - - - - - 成 - - ◎ ◎ × - - - ◎ - - - - - - 残 - - - 0 84 - - - 0 - - - - 0 - - マ - - - ◎ × - - - ◎ - ◎ - ◎ × - - 成 - × ○ ◎ × - - - ◎ - ◎ - ◎ ◎ - 残 - - - 0 84 - - - - - 0 - 0 * - - 卵 - - - ◎ - - - ◎ - - - ◎ ◎ ◎ - 成 - - - ◎ - - ◎ - ◎ - ◎ - ○ ◎ - 残 - - - 0 - - - 0 - 0 - 21 * 0 14 卵 - × ◎ ◎ × - - ◎ - ◎ ◎ ◎ ◎ ◎ - × - - - - ◎ - - - - ◎ - - - - - - - - - - - △ △ △ - - - - - × - × - ○ - × - - × - - - - ◎ - - - - ◎ ◎ - - - - - ◎ △ ◎ - - △ × × - - - - - × - × - - × × - - 84 - - - - 0 - - - - 0 - - - - - - - - - - - 14 - - - - - - - - - 84 - - - 84 - - × - - - - ◎ - - - - ◎ - - - - - - - - - - - × △ - - - - - - × - × - - × × - - × ◎ - - - ◎ - - - ◎ ◎ × - - - - - - - - ○ - × - × - ○ ◎ - - × - × - - × × - - 84 - - - - 0 - - - 0 0 - - - - - - - - - - - 84 - - - - - - - - - 84 - - - 84 - - - - - ◎ × ◎ × - ◎ - - - △ - × - ◎ - ◎ ◎ × - - - - - ○ ○ - - × - - - - ◎ - - ◎ - - - ○ ◎ ◎ × - ◎ - - - ○ - ◎ ◎ ◎ ○ ◎ ◎ × - - - - - ○ ◎ - - × - - - × ◎ - - ◎ - - - - - 0 14 - 0 - - - - - - - 0 28 0 0 - - - - - - - - - - - - - - - 0 - - 0 × ◎ - △ × ◎ × - ◎ ◎ - × × - × ◎ ◎ - ◎ ◎ × - × - - - × ○ - - × ◎ △ - - ◎ × - ◎ 成 - × ◎ ◎ × ○ ◎ - ◎ ○ ◎ ◎ ◎ ◎ - ○ × ◎ - ○ ◎ ◎ × - ◎ ◎ - × × - ◎ ◎ ◎ △ ◎ ◎ × - × × × - △ ◎ - - × ◎ × - △ △ × - ◎ 残 - 14 0 0 84 7 - 0 - 0 0 0 * 0 - 0 84 0 - - - 0 - - 0 0 - 42 5 - - 0 0 - 0 0 42 - 14 14 14 - - - - - - 0 28 - - - 84 - 0 ククメリス カブリダニ スワルスキ-カ タイリクヒメハナカ ブリダニ メムシ 卵 - × - ◎ × - - × ◎ - ◎ - ◎ ◎ ◎ - 成 - × ◎ ◎ × - - - ◎ - ◎ - ◎ ◎ - 残 - 14 - 0 84 - - - 0 - 0 - 0 * 0 14 卵 - - - - - - - - - - - - - - ◎ - 成 - - - - - - - - - - - - - - ◎ - 残 - - - - - - - - - - - - - - - - 幼 - × - ○ × - - × - × ◎ - ◎ ◎ × - × ◎ - - - ◎ - - - - - × - - - ◎ - - ◎ ◎ × - × × - - - - - - × - × - - - × - - × ◎ - - - ◎ × - - ◎ - × - - - ◎ - - ◎ ◎ × - × × × - △ ◎ - - × ◎ × - × - × - - 84 0 - - - 0 - - - 0 - - - - - 0 - - 0 0 - - 84 56 56 - - 0 - - - - 56 - - - 84 - - - - - ◎ - - - ◎ - ◎ - - - - - ◎ ◎ - ◎ - - - - - - - - - - - - - - - - - - - - - - - ◎ - ◎ - ◎ - ◎ ◎ - - - - ◎ ◎ - ◎ - - - - - - - - - - - - ○ - - - - - - - - - - - - - - - - 0 0 - - - - - - - - - - - - - - - - - - - - - - - - - - - - × - - - - ◎ × - - ◎ - - - - - ◎ - - △ - × - × △ × × ◎ × - - × ◎ × - × × × - ◎ 成 - × - ◎ × - - - ◎ × ◎ - ◎ △ ◎ - ◎ × × - - - ◎ × - - ◎ ◎ - - - - ○ - ◎ △ ◎ × - × × × - ◎ - - - × ◎ × - × × × - - 残 - - - 0 84 - - - - 14↑ 0 - 0 0 14 - 0 84 - - - - 0 7 - - 0 - - - - - - - - 14 - - - - - 14↑ - 0 - - - - 0 84 - 14 - 84 - 0 アリガタシマ アザミウマ オンシツ ツヤコバチ サバク ツヤコバチ 幼 - - - - - - - - - × ◎ - - △ × - 成 - - - - - - - - - △ ◎ - - × ◎ - 残 - - - - - - - - - - - - - - - - 蛹 - - ◎ ◎ ◎ - - - ○ - ◎ - ◎ ◎ ◎ - 成 - - ○ ◎ × - - - ○ × ◎ - - △ ◎ × 残 - - 7 0 36 - - - 0 35 0 - - 0 0 - 蛹 - - - - × - - - ◎ - ◎ - - ◎ ◎ - - - - - △ - △ - - - - - × - - - - - × - - - - - - - - × - - - - - - - - - - - - - - - ◎ - ◎ - - - - - × - - - - - ◎ - - - - - - - - △ - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - × ○ - - - ◎ - - - - ◎ × △ ○ - ◎ - ◎ ◎ ◎ ○ - × ◎ △ - - △ △ - × - × - × △ × - - × ◎ - × - ◎ - - - ◎ ◎ × × × - ◎ - ◎ ◎ ◎ × - × × × - - × × - × ◎ × - × × × - - 84 0 - 3 - 0 - - - 0 0 - 30 28 - 0 - 0 - 0 7 - 84 84 28 - - 24 24 - - 1 70 - 84 84 84 - - × - - - - ◎ - - - - - - - - - - - - ◎ - × - × - - - - - - - × - × - - × × - - 成 - - - - × - - - ◎ - ◎ - - △ ◎ × ○ × - - - - ◎ - - - - - - - - - - - - ◎ - × - × - - - - - - - × - × - - × × - - 残 - - - - 84 - - - 0 - 0 - - 0 0 - 0 84 - - - - 0 - - - - - - - - - - - - 0 - - - 84 - - - - - - - - - 84 - - - 84 - - チチュウカイ ツヤコバチ 蛹 - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - 成 - - - - - - - - ◎ - - - - - - × △ - - - ○ - - - - - - - - × - - - - - ◎ - - - - - - - - - - - - - - - - - - - - 残 - - - - - - - - - - - - - - - - 0 - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - イサエアヒメコバチ ハモグリコマユバチ デジェネランス カブリダニ クサカゲロウ 類 幼 - - - - × - - - ◎ △ ◎ - - ◎ ◎ - 卵 - × - ◎ × - - - ◎ - ◎ - - ◎ ◎ - - × ◎ - - - ◎ - - - - - × - - - ◎ - - ◎ ◎ × - × × - - - - - - × - × - - - × - - 幼 - - ◎ × × - - - × - ◎ - - × - - × - - - - ◎ - - - ◎ ◎ - - - - - - ◎ × ◎ × - × ◎ △ - - - - - × ◎ × - ○ - × - - × - - - - ◎ - - - - - - - - - - - - - - - - - - - - ◎ - - - × ○ × - - - × - - 成 - × ◎ ◎ × - - × ◎ × ◎ × ○ ○ ◎ - ○ × ◎ × - - ◎ ○ - - ◎ ◎ - × - - ◎ - △ - ◎ × - × × × - × △ - - × ◎ × - - × × - - 残 - - 0 0 84 - - - 0 21 0 42 19 0 0 - 0 84 0 21 - - 0 - - - 0 - - - - - 0 - - - - - - 84 - - - - - - - - - 84 - - - 84 - - 成 - × ◎ ◎ × - - - ◎ - ◎ - - ◎ - - × ◎ - - - ◎ × - - - - × - - - ◎ - △ ◎ ◎ × - × × × - - - - - × ○ × - × - × - - 残 - 14 - 0 84 - - - 0 - 0 - - * 0 - - 84 0 - - - 0 - - - - - - - - - 0 - - 0 0 - - 84 56 56 - - - - - - - 56 - - - 84 - - 成 - - ◎ △ × - - - × - ◎ - - △ - - × ○ - - - ◎ ○ - - - - ○ - - - - - ◎ ◎ - × - × × × - - - - - × ◎ × - ○ - × - - 残 - - - - 84 - - - - - 0 - - 0 - - - 84 0 - - - 0 - - - 0 - - - - - - - 0 - - - - - - 28 - - - - - - 0 84 - - - 84 - - ヨトウタマゴバチ類 ネマト-ダ 類 ボ-ベリア バシア-ナ バ-ティシリウム レカニ バチルス ズブチリス エルビニア カロトボ-ラ シュ-ドモナス フルオレッセンス 蛹 - ○ ◎ - × - - - - - ◎ - - ◎ ◎ - - × - - - - ◎ - - - - - - - - - - - - - ◎ × - × × × - - - - - × - × - △ - × - - 幼 - ◎ - ○ ◎ - - - ◎ - ○ - - ◎ - - - × - - - ◎ - - - - - - - - - - - ◎ - - ◎ - ◎ ◎ ◎ - - - - - × - × ◎ - - △ - - 分生子 ◎ ◎ ◎ ◎ ◎ ◎ ◎ ◎ × × ◎ ◎ - 胞子 - ◎ △ ◎ - - - - ◎ - ○ - - - - - - - - - ◎ - - - - - ◎ - - - ◎ ◎ ◎ - ◎ - - △ ◎ × - - - - - - - ◎ - ◎ - ◎ - - 芽胞 - ◎ ◎ - ◎ - - ◎ ◎ ◎ ◎ - - ◎ ◎ ◎ - - - ◎ ◎ ◎ ◎ - ◎ - - - ◎ - - - ◎ - ◎ ◎ ◎ - ◎ - - - - ◎ - - - - ◎ - - - ◎ - ◎ 菌 - - - - ◎ - - - ◎ - - - - - - - - - ◎ ◎ - - - - - - ◎ ◎ - - - ◎ - - ◎ - - - × - - ◎ - ◎ - ◎ - - - × - - 菌 - ◎ ◎ ◎ ◎ × × × ◎ - 成 - × △ ◎ × - - - ◎ - ◎ - - ◎ - - × - - - - ◎ - - - - - - - - - - - - ○ - × - × × × - - - - - × ○ × - × - × - - 残 - - 14 - 84 - - - - - 0 - - 0 - - - 84 - - - - 0 - - - - - - - - - - - - - - 42 - 84 56 - - - - - - - - 84 - 42 - 84 - - 残 - 0 - 0 - - - - 0 - - - - 0 0 - - - 7 - - - 0 - - - - - - - - - - - 0 - - - - - 0 7 - - - - - - - 7 0 - - - - - ・本評価表は正会員、賛助会員、個人会員の負担により維持、訂正が行われています。転載にあたっては所定の転載料を事務局までお支払くださるようお願いもうしあげます。 注)卵:卵に、幼:幼虫に、成:成虫に、マ:マミ-に、蛹:蛹に、胞子:胞子に、巣:巣箱の蜂のコロニ-に対する影響 残:その農薬が天敵に対して影響のなくなるまでの期間で単位は日数です。数字の横に↑があるものはその日数以上の影響がある農薬です。 *は薬液乾燥後に天敵を導入する場合には影響がないが、天敵が存在する場合には影響がでる恐れがあります。 記号:天敵等に対する影響は◎:死亡率0~25%、○:25~50%、△:50~75%、×:75~100%(野外・半野外試験)、◎:死亡率0~30%、○:30~80%、△:80~99%、×:99~100%(室内試験) マルハナバチに対する影響は◎:影響なし、○:影響1日、△:影響2日、×:影響3日以上 マルハナバチに対して影響がある農薬については、その期間以上巣箱を施設の外に出す必要があります。影響がない農薬でも、散布にあたっては蜂を巣箱に回収し、薬液が乾いてから活動させて下さい。 ・本評価表は会員の負担により維持、訂正が行われています。転載にあたっては所定の転載料を事務局までお支払くださるようお願いもうしあげます。 ・表中のエルビニア カロトボ-ラは乳剤との混用はできませんが、3日以上の散布期間であれば近接散布が可能です。またバチルス ズブチリスは混用できない剤とでも、翌日以降の近接散布は可能です。 ・表中の影響の程度及び残効期間はあくまでも目安であり、気象条件(温度、降雨、紫外線の程度及び換気条件等)により変化します。 上記の理由により、この表が原因で事故が発生しても、当協議会としては一切責任を負いかねますのでご了承の上、ご使用下さい。 この表はエクセルver7で作成していますので、パソコンで参照する場合にはver7以上で使用して下さい。セルにカ-ソルを近づけると出典が表示されます。表示のない場合はIOBCの資料です。 日本バイオロジカルコントロール協議会・2011年9月作成・第20版 マルハナバチ 巣 × × ○ × × - - - ○ × ○ ◎ ◎ ◎ ○ - × × × - - ◎ ○ - - ○ - × × × - ◎ - ○ ◎ - × × × × × - - ○ ○ ー × ◎ × - △ × × × ◎ 残 28 - 1 - 30 - - - 1 20↑ 1 - - 1 5 - 14 3 - - 1 - - 1 1 28 10↑ 30↑ - - 1 - 2~3 2~3 30 7 3 - - 1 1 1 4 0 14 - 2 28 14 14 <<殺菌剤>> 種類名 天敵等への殺菌剤・除草剤の影響の目安 ショクガ タマバエ コレマン アブラバチ ミヤコ カブリダニ チリ カブリダニ ククメリス カブリダニ スワルスキ-カ ハナカメムシ類 ブリダニ アリガタシマ アザミウマ オンシツ ツヤコバチ サバク ツヤコバチ チチュウカイ ツヤコバチ イサエアヒメコバチ ハモグリコマユバチ デジェネランス クサカゲロウ類 カブリダニ 幼 成 残 マ 成 残 卵 - - - - - - - - - - - - - - ◎ - - - - ◎ - - ◎ ◎ 0 - ◎ - - ○ ○ - ○ ○ - - - - - - - - - ◎ ◎ 0 - - - ◎ - - - - - - - - - - - - - - - - - - - - - - - - ◎ ◎ 0 ◎ ー ◎ ー ◎ 0 - - - - - - - - - - - - - ー - - - - - - - - - - - - - - - - - - - - - ◎ ○ - ◎ ◎ 0 ◎ - - - - - - - - - ◎ ◎ 0 - ヨトウタマゴバチ類 ネマト-ダ類 ボ-ベリア バ-ティシリウム バシア-ナ レカニ 成 残 卵 成 残 卵 成 残 卵 成 残 幼 成 残 幼 成 残 蛹 成 残 蛹 成 残 蛹 成 残 幼 成 残 卵 成 残 幼 成 残 蛹 成 残 幼 残 分生子 アミスタ- - ◎ ◎ 0 - - - ◎ ◎ - - - - ◎ ◎ - ◎ ◎ 0 - - - - - - ◎ ◎ 0 - - - - - - - - - - × アフェット ◎ - - ◎ 0 - ◎ - アリエッティ◎ 0 ◎ ◎ 0 - - - - - - ◎ - - ◎ ◎ - - ◎ - - - - - - - - ◎ 0 - - - - - - - - - ◎ 0 ◎ アントラコ-ル - - × × 7 - △ - - - - - ◎ - - - - ◎ × 28 ◎ ◎ 0 - - - - × - - △ - ◎ ◎ - - ○ - - - × アンビル ◎ - ◎ ◎ 0 - ◎ - - - - - ◎ - - - - - - - - - - - - - - - - - ◎ - ◎ ◎ 0 - ◎ - ◎ 0 イオウフロアブル - - ◎ ◎ 0 - ○ - - - - ◎ ◎ 0 - - - ◎ ○ 3 ◎ △ 7 - - - - △ 7 - ○ - ◎ ◎ - ◎ × - △ 0 ◎ オ-シャイン - - - ◎ - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - オ-ソサイド ◎ 0 ◎ ◎ 0 - ◎ 0 - - - ◎ ◎ 0 ◎ ◎ - ◎ ◎ 0 ◎ ◎ 0 - - - ◎ △ - - ◎ 0 ◎ ◎ 0 - - - - - △ カスミン - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - カスミンボルド- - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - ◎ カ-ゼ-トPZ - - - △ - - - - - - - - - - - - - - ○ - - - - - - - - - - - - - - - - - - - - - カリグリ-ン ◎ 0 ◎ ◎ ◎ ◎ ◎ ◎ - - - ◎ ◎ 0 - - - ◎ ○ - - ◎ - - - - ◎ ◎ 0 - ◎ - - - - - - - - - カンタス ◎ 0 ◎ 0 ◎ 0 ◎ 0 キノンド- - - - - - - - - - - - - - ◎ ◎ - - - - - - - - - - - - - - - - - - - - - - - - グラステン - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - ◎ 0 グランサ- - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - ◎ 0 ゲッタ- - - - - - - - - - - - - - - - - ◎ ◎ 0 - - - - - - - - - - - - - - - - - - - - △ クリ-ンヒッタ- - - - - - - - - - - - - - ◎ ◎ - - - - - - - - - - - - - - - - - - - - - - - - サプロ-ル ◎ 0 ◎ ○ 0 ◎ ○ 7 - - - ◎ ◎ 0 - - - ◎ ◎ 0 ◎ ◎ 0 - - - - ◎ 0 ◎ ○ 7 ◎ ◎ 0 - ◎ - ◎ 0 × サルバト-レME - - ◎ - ◎ - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - × サンヨ-ル - - ◎ - 0 - - - - - - ○ ◎ - - - - ◎ ◎ 0 - - - - - - - - - - - - - - - - - - - - シグナム ◎ 0 ◎ 0 ジマンダイセン - - - ◎ ◎ 0 ◎ ◎ 0 ○ ○ 0 - ◎ 0 - × - ○ ◎ 0 ◎ ◎ - ◎ ◎ 0 ◎ ◎ 0 - - - ◎ ◎ 0 - ◎ - ○ ○ - - × - ◎ 0 × ジャストミ-ト - - - ◎ ◎ 0 - - - ◎ ◎ 0 - - - - - - - - - - - - ◎ ◎ 0 - - - - - - - ◎ 0 - - - - - - - - - - - × スコア - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - ◎ - - - - - - - - - - - - ストロビ- ◎ - - ◎ 0 - - - - ◎ 0 - - - - ◎ - ◎ - 0 - - - - ◎ - - - - - - - - ◎ 0 - - - - - - - - - - - △ スミレックス ○ ○ - ◎ 0 0 ◎ ◎ 0 ◎ ◎ 0 - ◎ 0 - - - ◎ △ 0 ◎ ◎ - ◎ ◎ 0 ◎ ◎ 0 - - - - ◎ 0 - ◎ - ◎ ◎ 0 - ◎ - ◎ 0 ○ スミブレンド - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - ◎ - - - - - - - - - - - - セイビア- - - - - - - - - ◎ ◎ 0 - - - - - - - - - ◎ ◎ - - ◎ 0 - - - - - - - - - - - - - - - - - - - - × ダイセン - ◎ - - ◎ - - - - ◎ ◎ 0 ◎ ◎ 0 - - - ◎ - 0 - - - ◎ ◎ 0 ◎ ◎ 0 - - - ◎ ○ - ◎ ◎ 0 ◎ ◎ 0 - ◎ - - - ダコグリ-ン - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - ◎ 0 ダコニ-ル ◎ ◎ 0 ◎ ◎ 0 ◎ ◎ 0 ◎ ◎ 0 - ○ 0 - - - ◎ ◎ 0 ◎ ◎ - ◎ ◎ 0 ◎ ◎ 0 - - - - ◎ 0 - ○ - - ◎ - - - - ◎ 0 × チウラム ○ ◎ - ◎ ○ - - ◎ - ○ ○ 0 △ ◎ - - - - ◎ ○ - - - - ◎ △ 7 ◎ ○ 7 - - - - ◎ 0 △ ◎ - ○ ◎ - - - - ◎ 0 チルト ◎ × - ◎ ◎ - - - - - ◎ - - - - - - - ○ ◎ - - - - ◎ ◎ - - - - - - - - ◎ 0 - - - - ◎ - - - - ◎ 0 デラン - - - - ◎ - - ◎ - - ◎ - - - - - - - - ◎ - - - - ◎ ◎ 0 - - - - - - - - - - - - ◎ ◎ - - ◎ - ◎ 0 ◎ 銅剤 - ◎ - - ◎ - ◎ ◎ 0 ◎ ◎ 0 - ◎ 0 - - - ◎ ◎ 0 - - - ◎ ◎ 0 ◎ ◎ 0 - - - - - - - ◎ - ◎ ○ - ◎ ◎ 0 ◎ 0 ◎ トップジンM - - - ◎ ◎ - - - - ○ △ 21 ○ △ 21 - - - ◎ ◎ 0 ◎ ◎ - ◎ ◎ 0 ◎ ◎ 0 - - - - ◎ 0 ○ △ 21 ◎ ◎ - ○ ○ 14 ○ - △ トリアジン - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - ◎ - - - - - - - - - - - - トリフミン ◎ ◎ 0 ◎ ◎ 0 ◎ ◎ 0 ◎ ◎ 0 ◎ ◎ 0 - ◎ - ◎ ◎ 0 ○ ◎ - ◎ ◎ 0 ◎ ◎ 0 - - - - ◎ - ◎ ◎ 0 - - - - - - ○ 0 △ ナリア ◎ ◎ 0 ◎ 0 ◎ 0 ◎ 0 バイコラ-ル ◎ - - ◎ ◎ 0 ◎ ◎ 0 ◎ ◎ 0 ◎ ◎ 0 - ◎ - ○ ◎ 0 - - - ◎ ◎ 0 ◎ ◎ 0 - - - - ◎ 0 ◎ ◎ 0 ◎ ◎ 0 ◎ ◎ 0 ◎ 0 ○ バイレトン - - - ◎ ◎ 0 ◎ ◎ 0 ◎ ◎ 0 - ◎ 0 - - - ◎ ◎ 0 ◎ ◎ - ◎ ◎ 0 ◎ ◎ 0 - - - - ◎ 0 - ◎ - ◎ ◎ - - - - ○ 0 △ バシタック - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - パスポ-ト ◎ ◎ 0 ◎ ◎ 0 ◎ ◎ 0 ◎ ◎ 0 - ○ - - - - ◎ ◎ 0 - - - ◎ ◎ 0 ◎ ◎ 0 - - - - ◎ 0 - ○ - - ◎ - - - - ◎ 0 パンソイル灌注 - - - - - - ◎ ◎ 0 ◎ ◎ 0 - - - - - - - - - - - - ◎ ◎ 0 ◎ ◎ 0 - - - - - - - - - - - - - - - - - ビスダイセン - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - フェスティバル - - - - ◎ 0 - - - - ◎ 0 - - - - - - ◎ - 0 - - - - ◎ 0 - - - - - - - - - - - - - - - - - - - フルピカ - - - - ◎ - - - - - - - - - - - - - - - - ◎ ◎ - ◎ ◎ 0 - - - - - - - - - - - - - - - - - - - ◎ ベフラン - - - - - - - - - - - - - - - - - - ◎ ◎ 0 - - - - - - - - - - - - - - - - - - - - - - - - - - ベルク-ト - - - - ◎ - - - - - - - ◎ ◎ 0 - - - - - - ◎ ◎ - ◎ - - - - - - - - - - - - - - - - - - - - - - × ベンレ-ト ◎ ◎ 0 ◎ ◎ 0 - - - ◎ △ 21 ◎ △ 21 - - - ◎ ◎ 0 ◎ ◎ - ◎ ◎ 0 ◎ ◎ 0 - - - ◎ ◎ 0 ◎ △ 21 ◎ ○ - - - - ◎ 0 × ポリオキシンAL - - - - - - - - - - - - - - - - - - ◎ ◎ 0 ◎ ◎ - - - - - - - - - - - ◎ - - - - - - - - - - - - ◎ マンネブダイセンM ◎ △ - ◎ × - ◎ ◎ 0 ◎ ◎ 0 ◎ ◎ 0 - - - - × - - - - ◎ △ 0 ◎ ◎ 0 - - - - ◎ 0 ◎ ◎ 0 ◎ ◎ 0 ◎ × - ◎ 0 ミルカ-ブ灌注 - - - - - - ◎ ◎ 0 ◎ ◎ 0 - - - - - - - - - - - - ◎ ◎ 0 ◎ ◎ 0 - - - - - - - - - - - - - - - - - モレスタン △ △ - ◎ ◎ - - △ - × × 28 - ◎ 0 - - - ◎ - 0 △ ◎ - ◎ △ 5 - - - - △ - ◎ ○ 0 - ◎ - △ △ - - ◎ - - - ユ-パレン ◎ ◎ - ○ ○ - ◎ ◎ 0 ◎ ◎ 0 - ◎ 0 - - - ◎ ◎ 0 ◎ ◎ - ○ × 7 ◎ ◎ 0 - - - - ○ - - ○ - ○ ○ - ◎ × 28 - - ヨネポン - - - - - - - - - - - - - ◎ 0 - - - ◎ ◎ 0 - - - - - - - - - - - - - - - - - - - - - - - - - - ラリ- - - - - - ◎ ◎ 0 ◎ ◎ 0 - ◎ 0 - - - ◎ ◎ 0 - - - - - - - - - - - - - - - - - - - - - - - - ○ - × ランマンフロアブル - - - ◎ ◎ 0 ◎ ◎ 0 ◎ ◎ 0 ◎ ◎ 0 - - - ◎ ◎ 0 - - - ◎ ◎ 0 - - - - - - ◎ ◎ 0 - - - - - - - - - - - リゾレックス - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - リドミルMZ - - - - - - ◎ ◎ 0 ○ ○ 0 - ◎ - - × - - - - - - - ◎ ◎ - - - - - - - - - - - ◎ - - - - - - - - - ルビゲン - - - ◎ ◎ 0 ◎ ◎ 0 ◎ ◎ 0 ◎ ◎ 0 - ◎ - ◎ ◎ 0 - - - ◎ ◎ 0 ◎ ◎ 0 - - - - ◎ 0 ◎ ◎ 0 - ○ - ◎ ◎ 0 ◎ 0 ○ ロブラ-ル ◎ ◎ 0 ◎ ◎ 0 ◎ ◎ 0 ◎ ◎ 0 ◎ ◎ 0 - ◎ - ◎ ◎ 0 ◎ ◎ - ◎ ◎ 0 ◎ ◎ 0 - - - - ◎ 0 ◎ ◎ 0 ◎ ◎ 0 ◎ ◎ 0 ◎ 0 × 注)卵:卵に、幼:幼虫に、成:成虫に、マ:マミ-に、蛹:蛹に、胞子:胞子に、巣:巣箱の蜂のコロニ-に対する影響 残:その農薬が天敵に対して影響のなくなるまでの期間で単位は日数です。数字の横に↑があるものはその日数以上の影響がある農薬です。 *は薬液乾燥後に天敵を導入する場合には影響がないが、天敵が存在する場合には影響がでる恐れがあります。 記号:天敵等に対する影響は◎:死亡率0~25%、○:25~50%、△:50~75%、×:75~100%(野外・半野外試験)、◎:死亡率0~30%、○:30~80%、△:80~99%、×:99~100%(室内試験) マルハナバチに対する影響は◎:影響なし、○:影響1日、△:影響2日、×:影響3日以上 マルハナバチに対して影響がある農薬については、その期間以上巣箱を施設の外に出す必要がありまあす。影響がない農薬でも、散布にあたっては蜂を巣箱に回収し、薬液が乾いてから活動させて下さい。 ・表中の網掛けの部分はその農薬の開発メ-カ-の提供による知見です。 ・表中のエルビニア カロトボ-ラは乳剤との混用はできませんが、3日以上の散布期間であれば近接散布が可能です。またバチルス ズブチリスは混用できない剤とでも、翌日以降の近接散布は可能です。 ・表中の影響の程度及び残効期間はあくまでも目安であり、気象条件(温度、降雨、紫外線の程度及び換気条件等)により変化します。 上記の理由により、この表が原因で事故が発生しても、当協議会としては一切責任を負いかねますのでご了承の上、ご使用下さい。 この表はエクセルver7で作成していますので、パソコンで参照する場合にはver7以上で使用して下さい。セルにカ-ソルを近づけると出典が表示されます。表示のない場合はIOBCの資料です。 <<除草剤>> (バイオセ-フと混用可能な除草剤は下記のとおり) クサブロック、スタッカ-、バナフィン、カ-ブ、クサレス、タ-ザイン、ウエイアップ、ディクトラン 日本バイオロジカルコントロール協議会・2011年9月作成・第20版 バチルス ズブチリス エルビニア シュ-ドモナス カロトボ-ラ フルオレッセンス マルハナバチ 胞子 - 芽胞 ◎ 菌 ◎ 菌 ◎ 巣 ◎ 残 1 × ○ × × - × - - - - × × ◎ × ◎ ◎ ◎ × ◎ - × × × × - - - - - - - - - × △ ○ ◎ ◎ - ◎ ◎ ◎ - ◎ 2 1 0 - 0 0 - 0 - - - - - △ - ◎ ◎ ◎ ◎ ◎ × ◎ ◎ ◎ - × - - - - - - - - - - ◎ - ◎ ◎ ◎ - - - 0 - 0 0 × - - - ◎ - - × - × × × △ × ◎ - - × ◎ ◎ ◎ ◎ ◎ ◎ × ◎ ◎ ◎ ◎ × ◎ ◎ ◎ ◎ ◎ × ◎ × × × ◎ × ◎ × - - - - - ◎ - - ◎ - - - × ◎ - - ◎ ◎ - - ◎ - ◎ ◎ - ◎ ◎ ◎ ◎ ◎ ◎ - ○ 0 0 - - 0 - - 0 0 0 - 1 × △ - × ◎ - - ◎ - - △ - × ◎ × × - × - - ◎ ○ ◎ ◎ ◎ ◎ ◎ ◎ ◎ × ◎ ◎ ◎ ◎ ◎ ◎ ◎ × ◎ ◎ × ◎ ◎ × × ◎ × × ◎ 水◎ - - - - - - - - - ◎ ◎ - - - - - - - - ◎ × - ◎ ◎ ◎ ○ ◎ ◎ ◎ ◎ ◎ - ◎ ◎ ◎ ◎ - × ◎ - ◎ ◎ ◎ ◎ ◎ 1 0 0 0 - 0 0 0 0 3~5 - 0 0 0 天敵昆虫・ダニ製剤 登録状況 農薬の種類 イ サエ ア ヒ メ コ バチ 農薬の名称 マ イ ネッ ク ス 0 97.12.24 アリスタライフサイエンス㈱ 03/ 3/ 5変更 野菜類( 施設栽培) ハモ グリ バエ 類 01. 4.16 アリスタライフサイエンス㈱ 03/ 3/ 5変更 ヒ メ コ バチ D I 野菜類( 施設栽培) ハモ グリ バエ 類 02. 9.17 シンジェンタ ジャパン㈱ 03/ 3/ 5変更 ヒメト ップ 野菜類( 施設栽培) ハモ グリ バエ 類 02. 9. 3 ㈱アグリ総研 03/ 3/ 5変更 石原イ サパラ リ ト マ ト ( 施設栽培) ハモ グリ バエ 類 06. 1.25 石原産業㈱ 06/ 1/25新規登録 コ マ ユバチ D S ト マ ト ( 施設栽培) 06. 1.25 マ メ ハモ グリ バエ ミ ニ ト マ ト ( 施設栽培) ハモ グリ ミ ド リ ヒ メ コ バチ 剤 取り扱い会社 ハモ グリ バエ 類 ミ ニ ト マ ト ( 施設栽培) ハモ グリ コ マ ユバチ 剤 初年度登録 野菜類( 施設栽培) ・ ハモ グリ コ マ ユバチ 剤 マ イ ネッ ク ス 9 1 イ サエ ア ヒ メ コ バチ 剤 2011年8月22日現在 JPPAまとめ 対象作物 ミ ドリヒメ 野菜類( 施設栽培) 02. 9.17 シンジェンタ ジャパン㈱ 03. 3. 5 ハモ グリ バエ 類 05. 6.22 住友化学㈱ 琉球産経㈱ オン シ ツ ツ ヤコ バチ 剤 エン スト リ ッ プ 野菜類( 施設栽培) コ ナジ ラ ミ 類 ポイ ン セ チ ア ( 施設栽培) ツ ヤコ バチ E F ト マ ト ( 施設栽培) 05/ 6/22新規登録 03/ 3/ 7変更 04. 7. 8 オン シ ツ コ ナジ ラ ミ ミ ニ ト マ ト ( 施設栽培) サバク ツ ヤコ バチ 剤 95. 3.10 アリスタライフサイエンス㈱ 05/ 6/22新規登録 02. 9.17 シンジェンタ ジャパン㈱ 08/9/17失効 03. 3. 5 08/9/17失効 ツ ヤコ バチ E F 3 0 野菜類( 施設栽培) コ ナジ ラ ミ 類 02. 8.13 シンジェンタ ジャパン㈱ 03/ 3/ 5変更 ツ ヤト ッ プ 野菜類( 施設栽培) オン シ ツ コ ナジ ラ ミ 01. 1.30 ㈱アグリ総研 03/ 3/ 5変更 ツ ヤト ッ プ 2 5 野菜類( 施設栽培) オン シ ツ コ ナジ ラ ミ 10. 7.21 ㈱アグリ総研 10/ 7/21新規登録 石原ツ ヤパラ リ 野菜類( 施設栽培) コ ナジ ラ ミ 類 05.11.30 石原産業㈱ 07/ 7/18変更 エ ルカ ード 野菜類( 施設栽培) コ ナジ ラ ミ 類 03. 5. 7 アリスタライフサイエンス㈱ 03/10/ 8変更 サバク ト ッ プ 野菜類( 施設栽培) コ ナジ ラ ミ 類 05. 6. 1 ㈱アグリ総研 06/ 8/16変更 チ チ ュ ウカ イ ツ ヤコ バチ 剤 ベミ パール 野菜類( 施設栽培) タ バコ コ ナジ ラ ミ 類 ( シ ルバーリ ーフ コ ナ ジ ラ ミ を 含む) 07.07.04 アリスタライフサイエンス㈱ 07/ 7/ 4新規登録 コ レ マ ン ア ブ ラ バチ 剤 ア フ ィ パール 野菜類( 施設栽培) ア ブ ラ ムシ 類 98. 4. 6 アリスタライフサイエンス㈱ 11/4/6使用量変更 ア ブ ラ バチ A C 野菜類( 施設栽培) ア ブ ラ ムシ 類 02. 9. 3 シンジェンタ ジャパン㈱ 03/ 3/ 5変更 コ レト ッ プ 野菜類( 施設栽培) ア ブ ラ ムシ 類 02. 9. 3 ㈱アグリ総研 石原コ レ パラ リ 野菜類( 施設栽培) ア ブ ラ ムシ 類 07.6. 6 石原産業㈱ 07/ 6/ 6新規登録 チ ャ バラ ア ブ ラ コ バチ 剤 チ ャ バラ 野菜類( 施設栽培) ア ブ ラ ムシ 類 09.12.16 住化テクノサービス㈱ 09/12/16新規登録 シ ョ ク ガタ マ バエ 剤 ア フ ィ デン ト 野菜類( 施設栽培) ア ブ ラ ムシ 類 98. 4. 6 アリスタライフサイエンス㈱ 03/ 3/ 5変更 ナミ テ ン ト ウ剤 ナミ ト ッ プ 野菜類( 施設栽培) ア ブ ラ ムシ 類 02.11.26 ㈱アグリ総研 03/ 3/ 5変更 ナミ ト ッ プ 2 0 野菜類( 施設栽培) ア ブ ラ ムシ 類 05. 2. 9 ㈱アグリ総研 05/ 2/ 9追加 ス パイ デッ ク ス 野菜類( 施設栽培) 06. 4. 5 アリスタライフサイエンス㈱ 06/ 4/20新規登録 豆類( 種実) ( 施設栽培) 08.6.11 08/ 6/11変更 チ リ カ ブ リ ダニ 剤 いんげんま め( 施設栽培) ハダニ 類 08.6.11 果樹類( 施設栽培) 06. 4. 5 野菜類( 施設栽培) 07.11.28 ハダニ 類 ばら ( 施設栽培) 石原チ リ ガブ リ 07/11/28変更 02. 9. 3 シンジェンタ ジャパン㈱ 03/ 3/ 5変更 03. 8.20 ナミ ハダニ 02. 9. 3 野菜類( 施設栽培) ハダニ 類 06.10.18 石原産業㈱ 07/ 5/ 9変更 07. 9.19 07/ 9/19追加 チリ ト ッ プ 野菜類( 施設栽培) ハダニ 類 02. 6.18 ㈱アグリ総研 03/ 3/ 5変更 チ リ カ ・ ワーカ ー 野菜類( 施設栽培) ハダニ 類 09.12.02 小泉製麻(株) 09/12/ 2新規登録 ククメ リス 野菜類( 施設栽培) ア ザミ ウマ 類 98. 4. 6 アリスタライフサイエンス㈱ 03/ 3/ 7変更 シ ク ラ メ ン ( 施設栽培) ミ ヤコ カ ブ リ ダニ 剤 08/ 6/11追加 おう と う ( 施設栽培) ばら ( 施設栽培) ク ク メ リ ス カ ブ リ ダニ 剤 06. 4. 5 いも 類( 施設栽培) 花き 類・ 観葉植物( 施設栽培) カ ブ リ ダニ P P 03/ 3/ 5変更 02.10.16 ほう れんそう ( 施設栽培) ケナガコ ナダニ 03. 3. 7 メリト ップ 野菜類( 施設栽培) ア ザミ ウマ 類 02. 6.18 ㈱アグリ総研 03/ 3/ 7変更 ス パイ カ ル 野菜類( 施設栽培) ハダニ 類 03. 6. 3 アリスタライフサイエンス㈱ 03/10/ 8変更 豆類( 種実) ( 施設栽培) 08.6.11 08/ 6/11変更 いも 類( 施設栽培) 08.6.11 08/ 6/11変更 農薬の種類 ミ ヤコ カ ブ リ ダニ 剤 農薬の名称 ス パイ カ ル 対象作物 果樹類 0 花き 類・ 観葉植物( 施設栽培) ス パイ カ ルE X ス ワルス キーカ ブ リ ダニ 剤 ナミ ヒ メ ハナカ メ ムシ 剤 タ イ リ ク ヒ メ ハナカ メ ムシ 剤 初年度登録 ハダニ 類 取り扱い会社 アリスタライフサイエンス㈱ 08.6.11 08/ 6/11変更 08. 8.27 アリスタライフサイエンス㈱ 08/ 8/27新規登録 豆類( 種実) ( 施設栽培) 08. 8.27 08/ 8/27新規登録 いも 類( 施設栽培) 08. 8.27 08/ 8/27新規登録 果樹類 08. 8.27 08/ 8/27新規登録 花き 類・ 観葉植物( 施設栽培) 08. 8.27 08/ 8/27新規登録 野菜類( 施設栽培) ハダニ 類 茶 カ ン ザワハダニ 09. 5.13 2009/5/13変更 ミ ヤコ ト ッ プ いち ご ( 施設栽培) ハダ ニ 類 11. 3. 2 アグリ総研 11/3/2新規登録 ス ワルス キー 野菜類( 施設栽培) ア ザミ ウマ 類 08.11.19 アリスタライフサイエンス㈱ 豆類( 種実) ( 施設栽培) コナジラミ類 いも 類( 施設栽培) チャノホコリダニ 10. 5.26 10/ 5/26追加 果樹類(施設栽培) ミカンハダニ 10. 2.17 11/ 6/22変更 マンゴー(施設栽培) チャノキイロアザミウマ 10. 5.26 10/ 5/26追加 花き類・観葉植物(施設栽培) アザミウマ類 11. 6.22 11/ 6/22追加 ピ ーマ ン ( 施設栽培) ミ カ ン キイ ロ ア ザミ ウマ 98. 7.29 住友化学㈱ ミ ナミ キイ ロ ア ザミ ウマ 98. 7.29 オリ ス タ ー 08/11/19新規登録 10/ 5/26変更 オリ ス タ ーA 野菜類( 施設栽培) ア ザミ ウマ 類 01. 1.30 住友化学㈱ 03/ 3/ 5変更 ト ス パッ ク 野菜類( 施設栽培) ア ザミ ウマ 類 04.10. 6 協友アグリ㈱ タイリ ク 野菜類( 施設栽培) ア ザミ ウマ 類 01. 6.22 アリスタライフサイエンス㈱ 03/ 3/ 7変更 リクト ッ プ 野菜類( 施設栽培) ア ザミ ウマ 類 05. 8.31 ㈱アグリ総研 05/ 8/31新規登録 ヤマ ト ク サカ ゲロ ウ剤 カ ゲタ ロ ウ 野菜類( 施設栽培) ア ブ ラ ムシ 類 01. 3.14 アグロスター㈲ 03/ 3/ 5変更 ア リ ガタ シ マ ア ザミ ウマ 剤 ア リ ガタ 野菜類( 施設栽培) ア ザミ ウマ 類 03. 4.22 アリスタライフサイエンス㈱ 03/10/ 8変更 04. 1.28 琉球産経㈱ 04/ 1/28変更 サンケイ化学㈱ 微生物製剤 登録状況 殺虫剤 (BT剤を除く) 農薬の種類 バーテ ィ シ リ ウム・ レ カ ニ水和剤 農薬の名称 2011年8月22日現在 JPPAまとめ 対象作物 対象病害虫 登録年月日 取り扱い会社 バータ レ ッ ク 野菜類(施設栽培) ア ブ ラ ムシ 類 00. 8.15 アリスタライフサイエンス㈱ 03/ 3/ 5変更 マイ コ タ ール 野菜類(施設栽培) コ ナジ ラ ミ 類 06. 4. 5 アリスタライフサイエンス㈱ 06/ 4/ 5変更 き く ( 施設栽培) ミ カ ン キイ ロ ア ザミ ウマ 02. 7.30 トルコギ キョウ( 施設栽培) ミ カ ン キイ ロ ア ザミ ウマ 11. 2. 2 11/2/2追加 ペキロ マイ セス・ フ モ ソ ロ セウス水和剤 プ リ フ ァ ード 水和剤 野菜類(施設栽培) コ ナジ ラ ミ 類 01. 6.11 東海物産㈱ 03/ 3/ 7変更 ワタ ア ブ ラ ムシ 04. 7.21 06/ 6/ 7変更 ペキロ マイ セス・ テ ヌ イ ペス乳剤 ゴッ ツA 野菜類(施設栽培) コ ナジ ラ ミ 類 08.6.11 住友化学㈱ 08/ 6/11新規登録 野菜類(施設栽培) ア ブ ラ ムシ 類 10.5.26 出光興産㈱ 11/ 7/20変更 98.12. 4 サンケイ化学㈱ 04/ 8/13変更 98.12. 4 08/11/19変更 パスツ ーリ ア ペネト ラ ン ス水和剤 パスト リ ア 水和剤 野菜類 いも 類 ネコ ブ セン チュ ウ いち じ く ボーベリ ア ・ ブ ロ ン ニア テ ィ 剤 バイ オリ サ・ カ ミ キリ 00. 4.12 桑 キボシ カ ミ キリ 95.11.17 出光興産㈱ 果樹類 カ ミ キリ ムシ 類 95.11.17 かえで ゴ マダラ カ ミ キリ 98.12.10 うど セン ノ カ ミ キリ 03. 3. 7 たら のき ボーベリ ア ・ バシ ア ーナ乳剤 ボタ ニガード E S 02. 7. 2 しいたけ ハラアカコブカミキリ 07.12.12 野菜類 コ ナジ ラ ミ 類 06. 3. 8 アリスタライフサイエンス㈱ 06/ 3/ 8変更 ア ザミ ウマ類 04.12.22 04/12/22変更 コ ナガ 02.11.12 03/ 3/ 5変更 コ ナジ ラ ミ 類 06. 3. 8 10/ 5/26濃度変更 06. 3. 8 10/ 5/26濃度変更 ト マト ミ ニト マト ボーベリ ア ・ バシ ア ーナ剤 チャ ハマキ顆粒病ウイ ルス・ リ ン ゴ コ カ ク モ ン ハマキ顆粒病ウイ ルス水和剤 07/12/12追加 キャ ベツ ア オムシ 09. 8. 5 2009/8/ 5追加 レタ ス オオタ バコ ガ 09. 8. 5 2009/8/ 5追加 茶 ク ワシ ロ カ イ ガラ ムシ 09. 8. 5 2009/8/ 5追加 マン ゴ ー チャ ノ キイ ロ ア ザミ ウマ 09.12.16 2009/12/16追加 バイ オリ サ・ マダラ ま つ( 枯損木) マツ ノ マダラ カ ミ キリ 07.2.21 出光興産㈱ 07/ 2/21新規登録 ボーベリ ア ン ま つ( 枯損木) マツ ノ マダラ カ ミ キリ 07.2.21 井筒屋化学産業㈱ 07/ 2/21新規登録 ハマキ天敵 りんご リ ン ゴ コ カ ク モ ン ハマキ 03. 3.20 アリスタライフサイエンス㈱ 茶 チャ ハマキ 03. 3.20 チャ ノ コ カ ク モ ン ハマキ 03. 3.20 りんご リ ン ゴ コ カ ク モ ン ハマキ 04. 4. 7 日本化薬㈱ 10/4/7失効 茶 チャ ハマキ 04. 4. 7 10/4/7失効 チャ ノ コ カ ク モ ン ハマキ 04. 4. 7 10/4/7失効 サツ マイ モ ネコ ブ セン チュ ウ 90. 7.24 エムシー・ファーティコム㈱ カ ヤク ハマキ天敵 モ ナク ロ スポリ ウム・ フ ィ マト パガム剤 03/ 3/ 7変更 ネマヒ ト ン たばこ ト マト 01.12. 3 ミ ニト マト ハスモ ン ヨ ト ウ核多角体病ウイ ルス水和剤 ハスモ ン 天敵 ア スパラ ガス 03. 3. 5 ハスモ ン ヨ ト ウ ( 散布) 09. 8. 5 日本化薬㈱ 09/10/ 7使用方法変更 食用ぎく 10. 4.21 10/ 4/21追加 つる むら さ き 10. 4.21 10/ 4/21追加 いち ご 07.3.7 11/5/25濃度追加 キャ ベツ 08. 7. 9 08/ 7/ 9追加 しそ 08. 7. 9 08/ 7/ 9追加 し そ ( 花穂) 11. 5.25 11/5/25追加 レタ ス 07.3.7 えだま め 07.3.7 だいず 07.3.7 き く ( 葉) 11. 5.25 11/5/25追加 08. 7. 9 09/10/ 7使用方法変更 えだま め ハスモ ン ヨ ト ウ ( 無人ヘリコプ ターによ る 散布) 線虫製剤 農薬の種類 スタ イ ナーネマ・ カ ーポカ プ サエ 剤 農薬の名称 バイ オセーフ 対象作物 芝 対象病害虫 登録年月日 取り扱い会社 シ バオサゾ ウムシ 幼虫 05. 4.27 ㈱エス・ディー・エス バイオテック タ マナヤガ 05. 4.27 かんし ょ ア リ モ ド キゾ ウムシ 05. 4.27 かんし ょ ( 茎葉) イモゾウムシ 05. 4.27 2009/ 3/30記載内容整理 06. 7. 5 2009/ 3/30記載内容整理 08. 9.10 2008/ 9/10追加 08. 9.10 2008/ 9/10追加 野菜類 豆類( 種実) ハスモ ン ヨ ト ウ いも 類 いち じ く キボシ カ ミ キリ ( 幼虫) 05. 4.27 花き 類・ 観葉植物 キン ケク チブ ト ゾ ウムシ 05. 4.27 ハスモ ン ヨ ト ウ 09. 2.18 2009/ 2/18追加 モモシン ク イ ガ 05. 4.27 2007/ 4/ 4変更 07.10.31 2008/ 9/10追加 08. 9.10 2008/ 9/10追加 ネク タ リ ン 09. 2.18 2009/ 2/18追加 さく ら 09. 2.18 2009/ 2/18追加 08. 9.10 2008/ 9/10追加 09. 2.18 2009/ 2/18追加 果樹類 もも おう と う なし コ スカ シ バ ヒ メ ボク ト ウ り んご たら のき セン ノ カ ミ キリ 幼虫 08. 9.10 2008/ 9/10追加 オリ ーブ オリ ーブ ア ナア キゾ ウムシ 幼虫 08. 9.10 2008/ 9/10追加 オリ ーブ ( 葉) スタ イ ナーネマ・ グラ セラ イ 剤 バイ オト ピ ア やし ヤシ オオオサゾ ウムシ 幼虫 06. 1.25 2008/ 9/10変更 野菜類 ネキリ ムシ 類 11. 5.25 ㈱エス・ディー・エス バイオテック 11/5/25追加 コ ガネムシ 類幼虫 00.10.11 豆類( 種実) いも 類 かんし ょ 芝 00.10.11 シ バオサゾ ウムシ 幼虫 00. 8. 1 シ バツ ト ガ 03. 9. 3 スジ キリ ヨ ト ウ 03. 9. 3 タ マナヤガ 04. 8. 4 ハスカ ッ プ ナガチャ コ ガネ幼虫 08. 5.28 2011/2/16使用方法変更 ブ ルーベリ ー いち ご ヒ メ コ ガネ幼虫 ハス モ ン ヨ ト ウ 03. 3. 5 11. 2.16 2011/2/16使用方法変更 11/2/16追加 2011/2/16希釈水量変更 2008/ 5/28追加 殺菌剤 農薬の種類 ア グロ バク テ リ ウム・ ラ ジ オバク タ ー剤 農薬の名称 バクテローズ 対象作物 果樹類(りんごを除く) 対象病害虫 根頭がんしゅ病 登録年月日 10/3/3使用回数変更 02. 7. 2 日本農薬㈱ ばら 89.12. 1 きく 95. 7.31 10/3/3使用方法変更 02. 8.13 出光興産㈱ 08/ 7/23追加 08. 7.23 日本農薬㈱ 03/ 3/ 5変更 00. 7.27 03/ 3/ 5変更 うどんこ病 (散布) 00. 7.27 07/ 3/20変更 なし 黒星病 (散布) 07. 3.20 09/ 8/13追加 稲 いもち病 (散布) 09. 8.13 08/ 7/23変更 野菜類(いちご,ピーマン除く) 灰色かび病 (ダクト内投入) 08. 7.23 07/ 9/19変更 花き類・観葉植物 07. 9.19 09/ 8/13追加 ぶどう 11. 2. 2 11/2/2追加 かんきつ、マンゴー 09. 8.13 07/ 9/19変更 07. 9.19 08/ 7/23追加 08. 7.23 03/3/5変更 りんご バチルス ズブ チリ ス水和剤 ボトキラー水和剤 ぶどう かんきつ、マンゴー 灰色かび病 (散布) 野菜類 いちご インプレッション水和剤 ピーマン うどんこ病 (ダクト内投入) 野菜類 灰色かび病 (常温煙霧) 野菜類 うどんこ病 03.10. 8 ㈱エス・ディー・エス バイオテック 09/ 3/30処理方法追加 10/ 4/21処理方法追加:「散布(上 澄液)」を追加。 灰色かび病 03. 5. 7 10/ 4/21追加 豆類(種実) うどんこ病 10. 4.21 10/ 4/21追加 いも類 灰色かび病 10. 4.21 07/ 9/19変更 かぼちゃ うどんこ病 05.11. 2 07/ 9/19変更 トマト 葉かび病 05. 7.20 07/ 9/19変更 ミニトマト 05. 7.20 09/ 8/11処理方法追加 にら 白斑葉枯病 08. 5.28 08/ 5/28追加 ししとう 黒枯病 08. 5.28 08/ 5/28追加 食用ゆり 葉枯病 08. 5.28 09/ 6/24追加 からしな(種子) うどんこ病 09. 6.24 09/ 6/24追加 灰色かび病 09. 6.24 灰色かび病 03. 5. 7 07/ 9/19追加 07. 9.19 07/ 9/19追加 灰色かび病 07. 9.19 10/ 4/21追加 うどんこ病 10. 4.21 07/ 9/19追加 灰星病 07. 9.19 07/ 9/19追加 07. 9.19 08/ 5/28追加 08. 5.28 08/ 5/28追加 08. 5.28 08/ 5/28追加 10/ 4/21追加 ぶどう ハスカップ ホップ もも ネクタリン おうとう 灰星病 すもも バイオワーク水和剤 ブルーベリー 斑点病 08. 5.28 いちご うどんこ病 10. 4.21 野菜類 うどんこ病 04. 9.22 丸和バイオケミカル㈱ 07/ 5/ 9変更 (トマト、ミニトマトを除く) 灰色かび病 06. 5.24 07/ 5/ 9追加 トマト うどんこ病 07.5. 9 07/ 5/ 9追加 灰色かび病 07.5. 9 07/ 5/ 9追加 葉かび病 07.5. 9 07/ 5/ 9追加 うどんこ病 07.5. 9 07/ 5/ 9追加 灰色かび病 07.5. 9 07/ 5/ 9追加 葉かび病 07.5. 9 06/ 3/ 8変更 野菜類 灰色かび病 06. 3. 8 出光興産㈱ いちご うどんこ病 06. 3. 8 にら 白斑葉枯病 10.10.27 10/10/27追加 野菜類 灰色かび病 05.10. 5 クミアイ化学工業㈱ 10/5/12希釈倍数変更 かんきつ 05.10. 5 09/11/18追加 ブルーベリー 09.11.18 09/11/18追加 ハスカップ 09.11.18 ぶどう 05.10. 5 ミニトマト ボトピカ水和剤 ピーマン エコショット 06. 3. 8 07/ 7/18変更 11/8/ 3希釈倍数変更 →1000~2000倍) うどんこ病 11.8. 3 11/8/ 3追加 トマト、ミニトマト 葉かび病 06. 6.21 06/ 6/21変更 なし 黒星病 07. 7.18 07/ 7/18追加 (1000倍 農薬の種類 バチルス ズブ チリ ス水和剤 農薬の名称 対象作物 対象病害虫 登録年月日 黒斑病 11.8. 3 クミアイ化学工業㈱ 11/8/ 3追加 にら 白斑葉枯病 09.11.18 09/11/18追加 セルリー 斑点病 10.5.12 10/5/12追加 おうとう 灰星病 10.5.12 10/5/12追加 パセリ 斑点病 11.8. 3 11/8/ 3追加 11/8/ 3追加 エコショット クリーンフルピカ (バチルス ズブチリス・メパニピリム混合 しそ 斑点病 11.8. 3 きゅうり うどんこ病 09. 5.27 クリーンカップ 野菜類 うどんこ病 10. 7. 7 10/ 7/ 7追加 (銅・バチルス ズブチリス混合剤) トマト、ミニトマト 葉かび病 09. 5.27 09/ 5/27追加 疫病 10. 1.20 10/ 1/20追加 灰色かび病 10. 7. 7 10/ 7/ 7追加 灰色かび病 10. 3. 3 10/ 3/ 3新規登録 (バチルス ズブチリス・ 葉かび病 10. 3. 3 10/ 3/ 3新規登録 ポリオキシン混合剤) うどんこ病 11. 6.22 11/ 6/22追加 すすかび病 10. 3. 3 10/ 3/ 3新規登録 灰色かび病 10. 3. 3 10/ 3/ 3新規登録 うどんこ病 10.11.24 10/11/24追加 ハダニ類 10. 3. 3 10/ 3/ 3新規登録 うどんこ病 10. 3. 3 10/ 3/ 3新規登録 灰色かび病 10. 3. 3 10/ 3/ 3新規登録 ハダニ類 10. 3. 3 10/ 3/ 3新規登録 うどんこ病 10. 3. 3 10/ 3/ 3新規登録 ハダニ類 10. 3. 3 10/ 3/ 3新規登録 ハダニ類 10. 3. 3 10/ 3/ 3新規登録 野菜類(トマト、ミニトマト、食用 灰色かび病 ゆりを除く) うどんこ病 09.11.18 日本曹達㈱ 10/ 5/26濃度変更 09.11.18 10/ 5/26濃度変更 トマト、ミニトマト 灰色かび病 09.11.18 10/ 5/26濃度変更 うどんこ病 09.11.18 10/ 5/26濃度変更 葉かび病 09.11.18 09/11/18新規登録 灰色かび病 09.11.18 10/ 5/26追加 うどんこ病 09.11.18 10/ 5/26追加 葉枯病 09.11.18 10/ 5/26追加 うどんこ病 10. 5.26 10/ 5/26追加 10. 5.26 10/ 5/26追加 10/ 5/26追加 クリーンサポート トマト なす きゅうり ばら カーネーション アグロケア水和剤 食用ゆり ガーベラ ペチュニア なし 黒星病 10. 5.26 かんきつ 灰色かび病 09.11.18 09/11/18新規登録 もみ枯細菌病 06.12.13 セントラル硝子㈱ 06/12/13新規登録 苗立枯細菌病 06.12.13 ばか苗病 03. 1.28 クミアイ化学工業㈱ もみ枯細菌病 03. 1.28 ばか苗病 03. 1.28 もみ枯細菌病 03. 1.28 苗立枯細菌病 03. 1.28 いもち病 04. 8.13 苗立枯病(リゾープス菌) 04. 8.13 ごま葉枯病 05.12.27 ばか苗病 04.11.24 クミアイ化学工業㈱ もみ枯細菌病 04.11.24 苗立枯細菌病 04.11.24 いもち病 05.12.27 05/12/27追加 苗立枯病(リゾープス菌) 05.12.27 05/12/27追加 バチルス シ ン プ レ ク ス水和剤 モミホープ水和剤 稲 ト リ コ デルマ・ ア ト ロ ビ リ デ水和剤 エコホープ 稲(箱育苗) 稲 エコホープドライ 稲 11/7/6追加 05/12/27追加 農薬の種類 ト リ コ デルマ・ ア ト ロ ビ リ デ水和剤 農薬の名称 エコホープDJ 非病原性エ ルビ ニア ・ カ ロ ト ボーラ 水和剤 バイオキーパー水和剤 エコメイト 対象作物 稲 対象病害虫 登録年月日 ばか苗病 07. 2. 7 クミアイ化学工業㈱ もみ枯細菌病 07. 2. 7 苗立枯細菌病 07. 2. 7 褐条病 07. 2. 7 いもち病 07. 2. 7 苗立枯病(リゾープス菌) 07. 2. 7 07/ 2/ 7新規登録 アスパラガス 紫紋羽病 10. 1.20 10/ 1/20追加 たばこ 白絹病 10.11.24 10/11/24追加 ばれいしょ 軟腐病 98.11. 9 セントラル硝子㈱ 野菜類 97. 7.25 日産化学工業㈱ シクラメン 07. 2.21 03/ 3/ 5変更 07/ 2/21追加 かんきつ かいよう病 07. 2.21 07/ 2/21追加 野菜類 軟腐病 06.6. 7 クミアイ化学工業㈱ 06/ 6/ 7新規登録 07. 2.21 07/ 2/21追加 ばれいしょ シクラメン かんきつ かいよう病 07. 2.21 07/ 2/21追加 非病原性フ ザリ ウム オキシ スポラ ム水和剤マルカライト かんしょ つる割病 02. 6.18 エーザイ生科研㈱ 08/7/1失効 シ ュ ード モ ナス・ フ ルオレ ッ セン ス剤 トマト セル苗元気 青枯病 01. 6.22 多木化学㈱ 根腐萎凋病 02. 2. 1 アリスタライフサイエンス㈱ 育苗期の伸長抑制 07. 7.18 青枯病 03. 3. 5 根腐萎凋病 03. 3. 3 育苗期の伸長抑制 07.07.18 07/ 7/18追加 青枯病 05. 9.21 05/ 9/21追加 ピーマン 05. 9.21 05/ 9/21追加 とうがらし類 06. 7.31 06/ 7/31変更 07/ 7/18追加 ミニトマト なす シ ュ ード モ ナス・ フ ルオレ ッ セン ス水和剤 ベジキーパー水和剤 にがうり つる割病 07. 7.18 レタス 腐敗病 05. 8.31 セントラル硝子㈱ 05/ 8/31新規登録 06. 5.24 06/ 5/24変更 非結球レタス キャベツ 黒腐病 ブロッコリー 花蕾腐敗病 シュードモナスCAB-02水和剤 モミゲンキ水和剤 稲 タ ラ ロ マイ セス・ フ ラ バス水和剤 バイオトラスト水和剤 いちご タフパール 野菜類 もみ枯細菌病 苗立枯細菌病 モミキーパー コ ニオチリ ウム ミ ニタ ン ス水和剤 ミニタンWG バリ オボラ ッ ク ス パラ ド ク ス水和剤 フィールドキーパー水和剤 06. 5.24 06/ 5/24変更 09. 6.10 09/6/10追加 09. 6.10 09/6/10追加 01.10.22 セントラル硝子㈱、日産化学工業㈱ 10/10/22失効 01.10.22 炭疽病 01. 7.12 出光興産㈱ うどんこ病 01. 7.12 うどんこ病 07.3.7 09/ 9/2追加 いちご 炭疽病 07. 9. 5 07/ 9/ 5追加 トマト 葉かび病 07. 9. 5 07/ 9/ 5追加 07. 9. 5 07/ 9/ 5追加 ミニトマト タフブロック 11/07/30失効(アリスタのみ) なす すすかび病 10. 8.25 10/ 8/25追加 稲 褐条病 07.3.7 07/ 3/ 8新規登録 ばか苗病 07. 9. 5 いもち病 07. 9. 5 苗立枯細菌病 07. 9. 5 もみ枯細菌病 07. 9. 5 苗立枯病(リゾープス菌) 08. 9.10 2008/ 9/10追加 苗立枯病(フザリウム菌) 08. 9.10 2008/ 9/10追加 苗立枯病(トリコデルマ菌) 10. 7. 7 10/ 7/ 7追加 稲(箱育苗) 苗立枯病(リゾープス菌) 08. 9.10 2008/ 9/10追加 稲 ばか苗病 10. 4.21 セントラル硝子㈱ 11/8/3使用方法変更 いもち病 10. 4.21 10/ 4/21新規登録 苗立枯病(リゾープス菌) 10. 4.21 11/8/3使用方法変更 苗立枯病(トリコデルマ菌) 10. 4.21 10/ 4/21新規登録 苗立枯病(フザリウム菌) 11. 8. 3 11/ 8/ 3追加 苗立枯細菌病 11. 8. 3 11/ 8/ 3追加 もみ枯細菌病 11. 8. 3 11/ 8/ 3追加 稲(箱育苗) ばか苗病 11. 8. 3 11/ 8/ 3追加 キャベツ 菌核病 07. 6. 6 石原産業㈱ 07/ 6/ 6新規登録 08. 3.19 セントラル硝子㈱ 09/ 8/11処理方法追加 11. 2.16 11/2/16追加 10/ 7/ 7使用方法変更 レタス はくさい キャベツ 根こぶ病 抗ウイルス剤 農薬の種類 農薬の名称 ズッ キーニ黄斑モ ザイ ク ウイ ルス 弱毒株水溶 "京都微研"キュービオZY 対象作物 きゅうり 対象病害虫 ズ ッキーニ黄斑モザ イクウイルス 登録年月日 03. 5. 7 ㈱微生物化学研究所 の感染によ る 萎凋症 ズッ キーニ黄斑モ ザイ ク ウイ ルス 弱毒株水溶 "京都微研"キュービオZY-02 きゅうり ズ ッキーニ黄斑モザ イクウイルスの感染 09/ 5/ 7失効 08. 4.23 ㈱微生物化学研究所 によるモザイク症および萎凋症 除草剤 農薬の種類 ザン ト モ ナス・ キャ ン ペスト リ ス液剤 農薬の名称 キャンペリコ液剤 対象作物 対象病害虫 西洋芝(ブルーグラス) 西洋芝(ベントグラス) 99. 9.27 多木化学㈱ スズメノカタビラ 日本芝(こうらいしば) ド レ ク スレ ラ ・ モ ノ セラ ス剤 タスマート 移植水稲 登録年月日 99. 9.27 97. 5.20 ノビエ 04. 1.28 三井化学㈱ 08/ 4/23新規登録 BT製剤登録状況(アイウエオ順) 農薬の名称 エコマスターBT エスマルクDF 濃度 だいず ハスモンヨトウ 1000倍 シロイチモジヨトウ 1000倍 キャベツ ハイマダラノメイガ 1000倍 にんにく ネギコガ 1000倍~2000倍 野菜類 アオムシ 1000~2000倍 オオタバコガ 1000倍 コナガ 1000~2000倍 ハスモンヨトウ 1000倍 ヨトウムシ 1000倍 JPPAまとめ 取り扱い会社 クミアイ化学㈱ 07/ 4/ 4新規登録 住友化学㈱ 08/ 2/ 6追加 きく オオタバコガ 1000倍 とうもろこし アワノメイガ 2000倍 かぶ ハイマダラノメイガ 1000倍 キャベツ ハイマダラノメイガ 1000倍 きく オオタバコガ 1000倍 しょうが アワノメイガ 2000倍 06/ 9/21変更 果樹類(りんごを除く) シャクトリムシ類 2000倍 06/ 9/21変更 11/ 8/ 3追加 ハマキムシ類 2000~3000倍 06/ 9/21変更 ケムシ類 2000倍 09/ 4/ 8追加 シャクトリムシ類 2000~4000倍 06/ 9/21変更 ハマキムシ類 2000~3000倍 06/ 9/21変更 ケムシ類 2000~3000倍 06/ 9/21変更 かき(葉) ハマキムシ類 2000~3000倍 10/7/21追加 茶 チャノコカクモンハマキ 1000倍 チャノホソガ 1000倍 チャハマキ 1000倍 アオムシ 1000~2000倍 オオタバコガ 1000倍 コナガ 1000~2000倍 ヨトウムシ 1000倍 野菜類 樹木類 サブリナフロアブル 対象病害虫 ねぎ りんご クオークフロアブル 2011年8月22日現在 対象作物 シャクトリムシ類 2000倍 11/ 5/25変更 ケムシ類 2000倍 11/ 5/25追加 花き類・観葉植物 コナガ 1000倍 きゅうり ウリノメイガ 400倍 10/ 7/21追加 住友化学㈱ そば ハスモンヨトウ 400倍 06/ 9/20変更 きく ハスモンヨトウ 400倍 06/ 9/20変更 果樹類 ハマキムシ類 400倍 野菜類 アオムシ 400~800倍 オオタバコガ 400倍 コナガ 400~800倍 ハスモンヨトウ 400倍 ヨトウムシ 400~800倍 野菜類(はくさいを除く) はくさい からしな(種子) 豆類(種実) コナガ 1000倍~1500倍 Meiji Seikaファルマ㈱ 10/ 4/ 7変更 アオムシ 1000倍 サンケイ化学㈱ 10/ 4/ 7変更 ハスモンヨトウ 500~750倍 10/ 4/ 7変更 ヨトウムシ 1000倍 10/ 4/ 7変更 10/ 4/ 7変更 オオタバコガ 500倍 コナガ 1000倍~1500倍 08/ 5/14変更 アオムシ 1000倍 07/ 1/24変更 ヨトウムシ 1000倍 07/ 6/20追加 コナガ 1000倍 08/ 7/ 9追加 アオムシ 1000倍 08/ 7/ 9追加 ヨトウムシ 1000倍 08/ 7/ 9追加 ハスモンヨトウ 500~750倍 08/ 7/ 9追加 ヨトウムシ 1000倍 09/ 4/ 8追加 農薬の名称 対象作物 サブリナフロアブル いも類 ジャックポット顆粒水和剤 ゼンターリ顆粒水和剤 濃度 取り扱い会社 500~750倍 Meiji Seikaファルマ㈱ 09/ 4/ 8追加 オオタバコガ 500倍 サンケイ化学㈱ 09/ 4/ 8追加 ヨトウムシ 1000倍 09/ 4/ 8追加 ハスモンヨトウ 500~750倍 09/ 4/ 8追加 オオタバコガ 500倍 09/ 4/ 8追加 雑穀類 アワノメイガ 1000倍 08/ 5/14追加 茶 チャノコカクモンハマキ 1000倍 チャハマキ 1000倍 野菜類 コナガ 1000倍 アオムシ 1000倍 アリスタライフサイエンス㈱ オオタバコガ 1000倍 キャベツ ヨトウムシ 1000倍 ハスモンヨトウ 1000倍 日本なし ハマキムシ類 1000倍 いも類 ハスモンヨトウ 1000倍 北興化学工業㈱ うり科野菜類 ウリノメイガ 1000倍 住友化学㈱ えんどうまめ ウリノメイガ 1000倍 住友化学園芸㈱ カーネーション ハスモンヨトウ 1000倍 きく ハスモンヨトウ 1000倍 さやえんどう ウリノメイガ 1000倍 しいたけ シイタケオオヒロズコガ 1000倍 シイタケオオヒロズコガ 200倍 ストック コナガ 1000倍 そば ハスモンヨトウ 2000倍 とうもろこし オオタバコガ 1000倍 はくさい アオムシ 2000倍 コナガ 2000倍 ヨトウムシ 2000倍 パセリ キアゲハ 1000倍 ふじまめ シロイチモジマダラメイガ 1000倍 果樹類 ハマキムシ類 1000倍 実えんどう ウリノメイガ 1000倍 芝 シバツトガ 1000~2000倍 スジキリヨトウ 1000~2000倍 タマナヤガ 1000倍 食用ほおずき タバコガ 1000倍 茶 チャノコカクモンハマキ 1000倍 チャハマキ 1000倍 ヨモギエダシャク 1000倍 豆類(種実) ハスモンヨトウ 1000倍 野菜類(はくさいを除く) アオムシ 1000~2000倍 オオタバコガ 1000倍 コナガ 1000~2000倍 シロイチモジヨトウ 1000倍 ハスモンヨトウ 1000倍 ヨトウムシ 1000~2000倍 からしな(種子) ダイポール水和剤 対象病害虫 ハスモンヨトウ 10/ 9/ 8追加 07/ 5/ 9追加 アオムシ 1000~2000倍 08/ 5/28追加 コナガ 1000~2000倍 08/ 5/28追加 08/ 5/28追加 ヨトウムシ 1000~2000倍 樹木類 ケムシ類 1000倍 あぶらな科野菜類 タマナギンウワバ 1000倍 かき イラガ類 1000倍 カキノヘタムシガ 1000倍 アメリカシロヒトリ 1000~2000倍 モンクロシャチホコ 1000倍 さくら 10/ 4/ 7新規登録 11/ 8/ 3追加 住友化学㈱ 農薬の名称 ダイポール水和剤 対象作物 さざんか ストック コナガ 1000倍 チャドクガ 1000倍 プラタナス アメリカシロヒトリ 1000~2000倍 りんご アメリカシロヒトリ 1000倍 ハマキムシ類 1000倍 ヒメシロモンドクガ 1000倍 シバツトガ 500~1000倍 スジキリヨトウ 500~1000倍 タマナヤガ 1000倍 チャノコカクモンハマキ 500~1000倍 チャノホソガ 500倍 チャハマキ 500倍 ヨモギエダシャク 500倍 アオムシ 1000~2000倍 コナガ 1000~2000倍 ヨトウムシ 500倍 つばき ハスオビエダシャク 500~1000倍 食用つばき(種子) チャドクガ 1000倍 ハスオビエダシャク 500~1000倍 茶 野菜類 あぶらな科野菜 アオムシ 1000~2000倍 コナガ 1000~2000倍 タマナギンウワバ 1000倍 ヨトウムシ 500倍 イラガ類 1000倍 カキノヘタムシガ 1000倍 アメリカシロヒトリ 1000~2000倍 モンクロシャチホコ 1000倍 さざんか チャドクガ 1000倍 さつき ベニモンアオリンガ 1000倍 ストック コナガ 1000倍 つつじ ベニモンアオリンガ 1000倍 つばき チャドクガ 1000倍 プラタナス アメリカシロヒトリ 1000~2000倍 りんご アメリカシロヒトリ 1000倍 ハマキムシ類 1000倍 ヒメシロモンドクガ 1000倍 シバツトガ 500~1000倍 スジキリヨトウ 500~1000倍 タマナヤガ 1000倍 チャノコカクモンハマキ 500倍~1000倍 チャノホソガ 500倍 チャハマキ 500倍 ヨモギエダシャク 500倍 かき さくら 芝 茶 チューレックス顆粒水和剤 チューンアップ顆粒水和剤 濃度 1000倍 つばき 芝 チューリサイド水和剤 対象病害虫 チャドクガ 野菜類 コナガ 1000倍 アオムシ 1000倍 取り扱い会社 住友化学㈱ 三井物産㈱ 三井物産㈱ 10/ 4/ 7新規登録 10/ 4/ 7新規登録 オオタバコガ 1000倍 10/ 9/ 8追加 キャベツ ヨトウムシ 1000倍 10/ 4/ 7新規登録 ハスモンヨトウ 1000倍 10/ 4/ 7新規登録 日本なし ハマキムシ類 1000倍 10/ 4/ 7新規登録 野菜類 ハイマダラノメイガ 2000~3000倍 いも類 アオムシ 2000~3000倍 豆類(種実) コナガ 2000~3000倍 オオタバコガ 2000~3000倍 ヨトウムシ 2000倍 ㈱エス・ディー・エスバイオテック 09/ 8/11変更 07/ 8/15追加 農薬の名称 対象作物 チューンアップ顆粒水和剤 デルフィン顆粒水和剤 取り扱い会社 果樹類 ハマキムシ類 4000倍 05/ 6/22変更 りんご ヨモギエダシャク 4000倍 09/ 8/11追加 ㈱エス・ディー・エスバイオテック 05/ 6/22追加 かんきつ アゲハ類 2000倍~4000倍 10/5/12希釈倍数変更 からしな(種子) アオムシ 2000倍~3000倍 10/5/12追加 コナガ 2000倍~3000倍 10/5/12追加 オオタバコガ 2000倍~3000倍 10/5/12追加 ハイマダラノメイガ 2000倍~3000倍 10/5/12追加 ヨトウムシ 2000倍 10/5/12追加 ウリノメイガ 3000倍 10/5/12追加 えんどうまめ シロイチモジヨトウ 1000倍 三井物産㈱ 10/12/8全適用作物に 使用液量設定 かんしょ ハスモンヨトウ 1000倍 きく オオタバコガ 1000倍 そらまめ シロイチモジヨトウ 1000倍 たばこ タバコアオムシ 1000~2000倍 ヨトウムシ 1000~2000倍 とうもろこし オオタバコガ 1000倍 なし ケムシ類 1000~2000倍 かんきつ アゲハ 2000倍 08/ 7/ 9追加 オリーブ(葉) ケムシ類 1000倍 06/ 3/ 8追加 ハマキムシ類 2000倍 06/ 3/ 8追加 ブルーベリー イラガ類 1000倍 やまのいも シロイチモジヨトウ 1000倍 りんご シャクトリムシ類 2000~4000倍 ハマキムシ類 2000~3000倍 ケムシ類 1000~3000倍 ケムシ類 1000倍 ハマキムシ類 2000倍 シバツトガ 1000~2000倍 スジキリヨトウ 1000~2000倍 タマナヤガ 1000~2000倍 チャノコカクモンハマキ 1000~2000倍 チャハマキ 1000倍 ヨモギエダシャク 1000倍 アオムシ 1000倍 ウリノメイガ 1000倍 オオタバコガ 1000倍 コナガ 1000倍 シロイチモジヨトウ 1000倍 ハスモンヨトウ 1000倍 さくら アメリカシロヒトリ 1000倍 つばき チャドクガ 1000倍 のざわな ヨトウムシ 500倍 芝 シバツトガ 500~1000倍 スジキリヨトウ 500~1000倍 アオムシ 1000~2000倍 オオタバコガ 500~1000倍 コナガ 1000~2000倍 ストック コナガ 1000倍 たばこ アオムシ 1000~2000倍 ヨトウムシ 500~1000倍 ハスモンヨトウ 500倍 芝 茶 野菜類 野菜類 トアロー水和剤CT 濃度 3000倍 果樹類 トアロー フロアブルCT 対象病害虫 ウリノメイガ パセリ 10/12/8追加 大塚アグリテクノ㈱ 大塚アグリテクノ㈱ アオムシ 1000~2000倍 07/ 1/24追加 コナガ 1000~2000倍 07/ 1/24追加 ヨトウムシ 500~1000倍 07/ 1/24追加 農薬の名称 トアロー水和剤CT 対象作物 ひえ 1000倍 11/ 6/22追加 雑穀類(ひえを除く) アワノメイガ 1000倍 07/ 9/19変更 りんご ヒメシロモンドクガ 500~1000倍 ハマキムシ類 500~1000倍 07/ 1/24追加 果樹類(りんごを除く) ハマキムシ類 500~1000倍 07/ 1/24変更 樹木類(つばき類を除く) アメリカシロヒトリ 1000~2000倍 07/ 9/19変更 イラガ類 1000倍 08/ 6/11追加 アメリカシロヒトリ 1000~2000倍 07/ 9/19変更 チャドクガ 1000倍 野菜類(パセリを除く) 果樹類(りんごを除く) りんご 樹木類 かき イラガ類 1000倍 チャノコカクモンハマキ 500~1000倍 ヨモギエダシャク 500~1000倍 1000~2000倍 07/ 1/24変更 07/ 1/24変更 ヨトウムシ 500~1000倍 シャクトリムシ類 2000倍 07/ 1/24変更 住化グリーン㈱ 06/ 9/21変更 ハマキムシ類 2000~3000倍 06/ 9/21変更 ケムシ類 2000倍 09/ 6/ 1変更 シャクトリムシ類 2000~4000倍 06/ 9/21変更 ハマキムシ類 2000~3000倍 06/ 9/21変更 ケムシ類 2000~3000倍 06/ 9/21変更 シャクトリムシ類 2000倍 11/ 5/25変更 ケムシ類 2000倍 11/ 5/25追加 1000倍 アメリカシロヒトリ 1000~2000倍 モンクロシャチホコ 1000倍 さざんか チャドクガ 1000倍 さんしょう(実) アゲハ類 1000倍 ストック コナガ 1000倍 つばき チャドクガ 1000倍 フェニックス・ロベレニー トビモンオオエダシャク 500~1000倍 プラタナス アメリカシロヒトリ 1000~2000倍 りんご アメリカシロヒトリ 1000倍 ハマキムシ類 1000倍 ヒメシロモンドクガ 1000倍 シバツトガ 500~1000倍 スジキリヨトウ 500~1000倍 タマナヤガ 1000倍 樹木類 トビモンオオエダシャク 500~1000倍 茶 チャノコカクモンハマキ 500倍~1000倍 チャノホソガ 500倍 チャハマキ 500倍 ヨモギエダシャク 500倍 アオムシ 1000~2000倍 コナガ 1000~2000倍 タマナギンウワバ 1000倍 ハスモンヨトウ 500倍 ヨトウムシ 500倍 なし ケムシ類 1000~2000倍 りんご シャクトリムシ類 2000~4000倍 かんきつ 08/ 6/11追加 1000~2000倍 1000倍 野菜類 07/ 1/24変更 コナガ イラガ類 芝 大塚アグリテクノ㈱ アオムシ カキノヘタムシガ さくら ファイブスター顆粒水和剤 取り扱い会社 イネヨトウ 茶 バシレックス水和剤 濃度 500倍~1000倍 つばき類 バイオマックスDF 対象病害虫 アワノメイガ ㈱エス・ディー・エス・バイオテック アグロカネショウ㈱ 10/12/8全適用作物に 使用液量設定 ハマキムシ類 2000~3000倍 ケムシ類 1000~3000倍 10/12/8追加 アゲハ 2000倍 08/ 7/ 9追加 農薬の名称 対象作物 ファイブスター顆粒水和剤 果樹類 ブイハンターフロアブル いも類 茶 対象病害虫 濃度 ケムシ類 1000倍 ハマキムシ類 2000倍 コガネムシ類幼虫 25~50倍 ナガチャコガネ幼虫 125倍 取り扱い会社 住友化学㈱ ブイハンター粒剤 かんしょ コガネムシ類幼虫 20㎏/10a 住友化学㈱ フローバックDF だいず ハスモンヨトウ 1000倍 住友化学㈱ ねぎ シロイチモジヨトウ 1000倍 キャベツ ハイマダラノメイガ 1000倍 にんにく ネギコガ 1000倍~2000倍 野菜類 アオムシ 1000~2000倍 オオタバコガ 1000倍 コナガ 1000~2000倍 ハスモンヨトウ 1000倍 ヨトウムシ 1000倍 オオタバコガ 1000倍 きく 05/ 6/22追加 11/ 8/ 3追加 合成性フェロモン剤 登録状況 2011年8月22日現在 JPPAまとめ ※合成性フェロモン剤の登録はこれ以外にもございます。 詳しくは当協会刊「生物農薬ガイドブック2006」 p.12~p.16等をご覧下さい。 農薬の種類 オリフルア・トートリ ルア・ピーチフルア 剤 農薬の名称 コンフューザーN 対象作物 使用目的 果樹類 交尾阻害 対象病害虫 使用量 03. 1.28 信越化学工業㈱ モモシンクイガ 150~200本/10a 03. 1.28 チャハマキ 03. 1.28 チャノコカクモンハマキ 03. 1.28 リンゴコカクモンハマキ 03. 4. 8 すもも スモモヒメシンクイ 果樹類 モモシンクイガ 交尾阻害 オリフルア・トートリ コンフューザーMM ルア・ピーチフルア・ ピリマルア剤 オリフルア剤 果樹類 交尾阻害 ナシヒメシンクイ 100~120本/10a 02. 4.12 ミダレカクモンハマキ 03. 4. 8 リンゴモンハマキ 03. 4. 8 ナシヒメシンクイ 05. 3.23 リンゴコカクモンハマキ 100本~120本/10a 05. 3.23 信越化学工業㈱ モモハモグリガ 05. 3.23 モモシンクイガ 07. 4.11 チャノコカクモンハマキ 120本/10a 10. 3.17 10/ 3/17追加 09/ 5/13追加 ナシヒメコン 交尾阻害 ナシヒメシンクイ 50~100本/10a 10. 8.25 信越化学工業㈱ スモモヒメシンクイ 50~100本/10a スカシバコン 50~150本/10a 果樹類 交尾阻害 コナガコン アルミゲルア・ダイア コナガコン-プラス モルア剤 誘引 コナガが加害する農作物 等 交尾阻害 アメリカシロヒトリ コナガ コナガ、オオタバコガが加 害する農作物等 オオタバコガ、コナガ コナガ、オオタバコガ、ヨトウガが 交尾阻害 加害する農作物等 コナガ 果樹類 リンゴコカクモンハマキ オオタバコガ 直線使用(街路樹な ど)40m当り1個以上 面使用(公園など)10a 当り3個以上 ハウス:100~400m/10a (100mリール) 露地:100~110m/10a (100mリール) 露地:200本/10a (20cmチューブ) 100~120本/10a (22g/100本製剤) 100本/10a (22g/100本製剤) 茶 交尾阻害 交尾阻害 加害作物栽培地帯 アリマルア・オリフル コンフューザー-A ア・テトラデセニルア セテート・ピーチフル ア剤 りんご 01. 6.11 出光興産㈱ 03/ 3/20変更 01. 6.11 03/ 3/20変更 交信阻害 サンケイ化学㈱ 89. 2.27 02. 2. 1 信越化学工業㈱ 09/ 8/ 5変更 03. 4. 8 09. 8. 5 100~150本/10a 03. 4. 8 リンゴモンハマキ (54g/150本製剤) 03. 4. 8 チャハマキ 加害する農作物 シロイチモジヨトウの 05/ 2/ 9追加 ミダレカクモンハマキ 03/ 4/ 8変更 03. 4. 8 チャノコカクモンハマキ ヨトウコン-S 03/ 3/ 5変更 05. 2. 9 01. 1.30 信越化学工業㈱ チャハマキ ハスモンヨトウが 88.12.22 信越化学工業㈱ ヒメコスカシバ雄成虫 交尾阻害 ヨトウコン-H 11/ 8/ 3処理量変更 10/ 8/25新規登録 コスカシバ雄成虫 ヨトウガ ハマキコン-N 09/ 8/ 5変更 07/ 4/11変更 09. 5.13 サンケイ化学㈱ フォールウェブルア剤 ニトルアー<アメシロ> 樹木類 ビートアーミルア剤 02. 4.12 150~200粒/10a かき リトルア剤 03/ 4/ 8変更 リンゴコカクモンハマキ 食用さくら(葉) トートリルア剤 10/ 6/23変更 02. 4.12 信越化学工業㈱ ナシヒメシンクイ さくら ダイアモルア剤 06.10.18 交尾阻害 果樹類 10/ 6/23変更 03. 4. 8 200本/10a ラブストップヒメシン 果樹類 すもも チェリトルア剤 取り扱い会社 50~200本/10a リンゴモンハマキ コンフューザーR 登録年月日 ナシヒメシンクイ 03. 4. 8 150~250本/10a チャノコカクモンハマキ (90g/250本製剤) 20~200m/10a(20㎝ ハスモンヨトウ チューブの場合100~ 1000本) 露地の場合:100~500 シロイチモジヨトウ 本(20㎝チューブ)/10a ハウスの場合:100~ 140m(20㎝チューブの場 合は500~700本)/10a モモシンクイガ 01. 1.30 03/ 4/ 8変更 03. 4. 8 96. 7.22 信越化学工業㈱ 03/ 4/ 8変更 90. 7. 4 信越化学工業㈱ 96. 2.26 信越化学工業㈱ ナシヒメシンクイ キンモンホソガ ミダレカクモンハマキ 150~240枚(又は 本)/10a リンゴコカクモンハマキ リンゴモンハマキ アルミゲルア・ウワ バルア・ダイアモル ア・ビートアーミル ア・リトルア剤 コンフューザーV ピーチフルア剤 シンクイコン ブルウェルア・ロウカ コンフューザー-G ルア剤 野菜類 交信阻害 コナガ 04. 9. 1 信越化学工業㈱ 100~200本/10a (41g/100本製剤) 09/ 8/ 5変更 いも類 オオタバコガ 04. 9. 1 豆類(種実) ハスモンヨトウ 04. 9. 1 花き類・観葉植物 タマナギンウワバ 04. 9. 1 イラクサギンウワバ 09. 8. 5 11/6/22処理量変更 ヨトウガ 04. 9. 1 11/6/22処理量変更 シロイチモジヨトウ 100本/10a (41g/100本製剤) 04. 9. 1 100本~150本/10a 20~40m(20cmチューブに 換算して100~200 本)/10a 93. 9.30 信越化学工業㈱ 果樹類 交尾阻害 モモシンクイガ雄成虫 芝 交尾阻害 シバツトガ スジキリヨトウ 07. 4.11 信越化学工業㈱ 07/ 4/11変更
© Copyright 2026 Paperzz