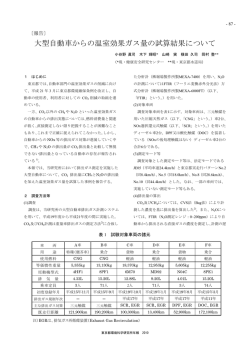
農業環境研究叢書 第18号 農業環境研究 2001― 2010 独立行政法人 農業環境技術研究所編 独立行政法人 農業環境技術研究所 はじめに 農業環境技術研究所は国立研究機関として 1983 年 12 月 1 日に新たに設立された.この発足に は,国民に対する食料の安定的な供給ということだけではなく,農林水産業が国土における人間の 生存全体を保証し得るような役割を果たすための技術を開発するという大きな期待が込められてい たことが,当時の農林水産技術会議事務局長の講演から窺い知ることができる.17 年が経過した 2001 年 4 月 1 日に,国立研究機関等の独立行政法人化が進められる中で,農業環境技術研究所も 独立行政法人として装いも新たに再出発した.そして,間もなく満 10 年を迎える. 再出発した農業環境技術研究所は,設立当初に期待された目的がいささか凝縮されてはいるが, 「農業生産の対象となる生物の生育環境に関する技術上の基礎的な調査および研究等を行うことに より,その生育環境の保全および改善に関する技術の向上に寄与する」ことを目的としている.こ の目的を遂行するに当たっては,具体には,国が策定する農林水産研究基本計画にそって5年の中 期目標期間ごとに国により当研究所に付与される中期目標に従い中期計画を立案し,実行すること となっている. 第 I 期および第 II 期にわたる 10 年間には数々の輝かしい成果を上げることができた.種々の学 術賞につながる数々の学術的成果はもとより,土壌モノリスを活用した土の理解増進に対する文 部科学大臣表彰受賞,ミニ農村の創造・展示による農村の生物多様性の理解増進に対する文部科 学大臣表彰受賞,気候変動に関する政府間パネル(IPCC)への貢献,さらには農林水産研究成果 10 大トピックスに 2005 年より連続して毎年選出されていること,あるいは残留性有機汚染物質 (POPs)を始め外来生物,遺伝子組換え作物などに関する各種の基本指針,実験指針,手引き,マ ニュアルなどに採用されていることなどは,その一例である.これらも含めこれまでに上げられた 研究成果は,研究成果発表会を始め折々に開催する農業環境シンポジウム,ワークショップ,公開 セミナー,研究会,国際シンポジウムやワークショップ,科学フェスティバル,アグリビジネス創 出フェアなどでの発表や展示,研究所年報(和文,英文),研究所報告,研究所資料,研究成果情 報,環境報告書,農環研ニュース,その他各種刊行物,さらにはホームページによって紹介をして いる. 独立行政法人として再出発してから 10 年が経過し,第Ⅱ期中期目標期間の終了を迎えるに当た り,この間に上げられた研究成果を一巻の書にまとめ公表し,読者諸賢の批評の目に晒すことは, 私たちに「離見」を与えるとともに,農業環境研究の一層の発展を図る上で大いなる意義あるもの と考え,ここに上梓し世に送る次第である. 本書が農業環境研究のさらなる発展と農業環境問題の解決に少しでも寄与することができるなら ば望外の喜びである. 2011 年2月 記 佐藤 洋平 目 次 序 章 農業をめぐる環境問題と環境研究(佐藤 洋平)… ……………………………………… 1 1.農業をめぐる環境問題:制御と循環 … ……………………………………………………………… 1 2.農業環境研究の枠組み … ……………………………………………………………………………… 3 3.本書の構成 … …………………………………………………………………………………………… 5 Ⅰ.農業生態系における有害化学物質の動態とリスク低減 - 食の安全・安心をめざして 第 1 章 重金属(カドミウム,ヒ素)汚染リスク評価と 汚染土壌修復技術(西尾 隆)……………………… 7 1.農産物中のカドミウム,ヒ素の基準値等を巡る国内外の動向 … ………………………………… 7 2.農作物におけるカドミウム,ヒ素の吸収・移行特性の解明 … …………………………………… 9 3.農作物の品種・系統間差や育種を利用したカドミウム低減技術の開発 … ……………………… 9 4.農耕地へのカドミウムの負荷 … …………………………………………………………………… 12 5.栽培管理による農作物のカドミウム,ヒ素低減技術 … ………………………………………… 12 6.農作物可食部のカドミウム濃度予測手法の開発 … ……………………………………………… 13 7.カドミウムの分析技術の開発 … …………………………………………………………………… 14 8.カドミウム低減のための土壌修復技術 … ………………………………………………………… 15 9.農地土壌中における有機ヒ素の動態 … …………………………………………………………… 18 第 2 章 POPs,農薬のリスク評価とリスク低減技術(與語 靖洋)… ……………………… 21 1.はじめに … …………………………………………………………………………………………… 21 2.POPs の動態および作物残留低減技術 … ………………………………………………………… 22 3.農薬の環境影響評価 … ……………………………………………………………………………… 26 4.農薬のリスク低減技術の開発 … …………………………………………………………………… 30 5.農薬や POPs の大気中挙動 … ……………………………………………………………………… 31 6.薬剤抵抗性と病害抵抗性誘導 … …………………………………………………………………… 33 7.今後の展望 … ………………………………………………………………………………………… 34 第 3 章 栄養塩類の動態と負荷低減(新藤 純子)… …………………………………………… 37 1.はじめに … …………………………………………………………………………………………… 37 2.栄養塩の土壌・地下水層での挙動と流出メカニズムの解明 … ………………………………… 37 3.流域スケールの栄養塩動態シミュレーションモデル … ………………………………………… 44 4.農地からのアンモニア揮散 … ……………………………………………………………………… 47 5.県・国・東アジアスケールの物質循環の評価と水質への影響 … ……………………………… 48 6.農業環境モニタリング … …………………………………………………………………………… 52 7.今後の展望 … ………………………………………………………………………………………… 53 ii Ⅱ.地球温暖化と農業 -気候変動にそなえる 第 4 章 気候変動と作物生産変動予測(宮田 明)… …………………………………………… 56 1.はじめに … …………………………………………………………………………………………… 56 2.農耕地観測地点における近年の気温変化の傾向 … ……………………………………………… 56 3.気候変化シナリオの統計的ダウンスケーリング … ……………………………………………… 57 4.日本およびアジアの主要生産地のコメ収量変動予測 … ………………………………………… 59 5.農業水利用を考慮した大陸スケールの水循環モデルの開発 … ………………………………… 64 6.今後の展望 … ………………………………………………………………………………………… 65 第 5 章 イネ・水田生態系の応答と適応戦略(谷山 一郎)… ………………………………… 67 1.はじめに … …………………………………………………………………………………………… 67 2.温暖化影響の把握 … ………………………………………………………………………………… 67 3.温度,CO2 濃度上昇の水稲への影響解析 … ……………………………………………………… 68 4.影響把握・予測のためのモデル開発・データベースの構築 … ………………………………… 71 5.温暖化に対するぜい弱性の予測 … ………………………………………………………………… 74 第 6 章 温室効果ガス発生抑制と土壌炭素蓄積(八木 一行)………………………………… 76 1.はじめに … …………………………………………………………………………………………… 76 2.温室効果ガス発生・土壌炭素蓄積メカニズムの解明 … ………………………………………… 77 3.モニタリング手法の開発 … ………………………………………………………………………… 78 4.発生量評価とモデル開発 … ………………………………………………………………………… 81 5.発生抑制技術の開発とその評価 … ………………………………………………………………… 87 6.今後の展望 … ………………………………………………………………………………………… 91 Ⅲ.農業と生物多様性 -豊かな農業・農村環境をめざして 第 7 章 農業生態系における生物多様性の評価(安田 耕司)………………………………… 96 1.はじめに … …………………………………………………………………………………………… 96 2.休耕田・耕作放棄地における生物多様性 … ……………………………………………………… 96 3.農業活動による生物多様性の維持 … ……………………………………………………………… 98 4.農業生態系の各種環境要因が生物多様性に及ぼす影響 … ……………………………………… 100 5.長期・広域スケールにおける生物多様性の評価 … ……………………………………………… 105 6.今後の展望 … ………………………………………………………………………………………… 106 第 8 章 外来生物のリスク評価(安田 耕司)… ………………………………………………… 109 1.はじめに … …………………………………………………………………………………………… 109 2.外来植物の定着と拡散 … …………………………………………………………………………… 109 3.外来生物の雑草リスクの評価 … …………………………………………………………………… 112 4.非意図的に導入された昆虫等に関する研究 … …………………………………………………… 114 5.意図的に導入された昆虫等に関する研究 … ……………………………………………………… 117 6.外来性病害微生物に関する研究 … ………………………………………………………………… 119 7.外来生物データベースの構築 … …………………………………………………………………… 120 iii 8.今後の展望 … ………………………………………………………………………………………… 121 第 9 章 遺伝子組換え作物の生態系影響評価と 共存のための管理手法の開発(田中 宥司)………………… 124 1.はじめに … …………………………………………………………………………………………… 124 2.ほ場条件下における遺伝子組換えダイズとツルマメとの自然交雑 … ………………………… 124 3.遺伝子組換えナタネのモニタリング … …………………………………………………………… 127 4.遺伝子組換え作物と一般栽培作物等との共存のための管理手法開発 … ……………………… 130 5.今後の展望 … ………………………………………………………………………………………… 134 第 10 章 環境生物の情報化学物質と機能利用(藤井 毅)… ………………………………… 137 1.はじめに … …………………………………………………………………………………………… 137 2.昆虫性フェロモンの利用に関わる研究 … ………………………………………………………… 138 3.植物のアレロケミカルの利用に関わる研究 … …………………………………………………… 140 4.微生物の有用機能の利用に関わる研究 … ………………………………………………………… 143 5.今後の展望 … ………………………………………………………………………………………… 149 Ⅳ.農業環境の把握と情報の活用 -農業環境を支える 第 11 章 リモートセンシング・地理情報システムの利用(三輪 哲久)…………………… 155 1.はじめに … …………………………………………………………………………………………… 155 2.リモートセンシング技術 … ………………………………………………………………………… 155 3.地理情報システムを利用した農業生態系の解析 … ……………………………………………… 160 4.時空間情報のデータベース化とインターネットでの利用 … …………………………………… 162 5.今後の展望 … ………………………………………………………………………………………… 164 第 12 章 農業環境のモニタリングと手法の開発 ………………………………………………… 167 1.物理環境・ガスフラックス変動のモニタリング(宮田 明)…………………………………… 167 2.農業環境中の放射性物質のモニタリング(西尾 隆)…………………………………………… 174 第 13 章 農業環境情報の活用(對馬 誠也)……………………………………………………… 182 1.はじめに … …………………………………………………………………………………………… 182 2.分 類 … ……………………………………………………………………………………………… 182 3.収集・保存 … ………………………………………………………………………………………… 186 4.データベース … ……………………………………………………………………………………… 188 5.今後の展望 … ………………………………………………………………………………………… 193 農業環境技術研究所 研究成果 … …………………………………………………………………… 201 農業環境問題と研究の流れ … ………………………………………………………………………… 211 序章 農業をめぐる環境問題と環境研究 1.農業をめぐる環境問題:制御と循環 20 世紀における科学・技術の目覚しい発展による技術開発,技術革新は,利便性と豊かさを実 現した.工業部門における生産性の向上,労働と資源の大量投入による生産の拡大,都市と地方の 開発,貿易の拡大などによる経済の成長によって実現した豊かな工業化社会は,他方で,大量生 産,大量消費,大量廃棄を伴い,肥大化し高度化した人間活動による環境負荷の累積は地球規模で の環境問題を生み出すようになった. 急速な工業化のもとで工場がもたらす大気,水質,土壌の汚染など産業公害としての環境問題 も,温室効果ガスの排出量の増加による地球の温暖化,化石燃料の燃焼に伴い排出される硫黄酸化 物などによる酸性雨,多様な生物の生息地域である森林の開発などによる生物多様性の減少,過放 牧による砂漠化の進行など地球規模の環境問題も,K. ボールディング(K. E. Boulding)が「宇宙 船地球号」の経済学を論じる中で「循環的生態システム」として把握する地球の複雑なシステムに 重大な影響を与えていることから生じている問題である.このシステムは,太陽エネルギーによっ てもたらされ,空間と時間の種々のスケールの生態システムの中で作用する物質循環によって支え られているので,環境問題はこの循環を損なうことによって生じている問題であるということがで きる. 農業をめぐる環境問題も同様であ 表 資源に対する農業の負荷 る. 20 世紀に発達した近代農業は農業 の工業化によるものであった.近代農 業は,肥料や農薬などを含め各種の 資源の種類 硝酸 水 土壌流出 土壌 行い,農産物などを産出してきた.そ アンモニア 大気 化(High Input, High Return)を特徴 としている.こうした農業は生産性の 向上と生産の拡大を実現したが,他方 で, 「循環的生態システム」への負荷 を強めるに至った. 表は,農業が循環利用する生態系を 資源の視点からとらえて整理し,水, 土壌,生物資源など生産条件としての 農業資源に対する農業による負荷の可 能性を表したものであるが,農業は, 生態系への負荷の可能性を常に内包し ている. 土壌圧縮 機能障害 れは,化学資材の大量投入と資源の 集約的利用による食料等生産の最大 リン 農薬 資材を投入し,土壌,水,大気など各 種資源を集約的に利用して生産活動を 農業による負荷の可能性 ①非生物的資源 メタン 一酸化二窒素 農薬 ②生物的資源 植物相 動物相 生息圏 生命力の減退 個体 個体群 生物種の減少 群集 生息基盤の奪取 生態系(エコシステム) ③美的資源 多様な景観要素 多様な景観像 景観像の単調化 分割耕地(小さい圃場) 圃場の拡大 輪作 輪作の単純化 農地の畔 道路の端 生け垣 農地の樹木 圃場の整備 小川 湖沼(池、沼) 小川・湖沼の除去 出典:四方康行ほか訳(1996).一部加筆修正. 農業は,周知のように,太陽エネルギーと生態系を繰り返し利用して食料,飼料,代替エネル ギーなどを生産する産業であるが,生態系の利用のあり方に再検討が迫られるようになった.農業 は,生態系を利用するための農耕において,人間の利益のために生態系の生産力を高めることを目 的としてこれを制御することを必然とする.この生態系への能動的な働きかけは,種の選別,品種 改良,遺伝子組換え作物の作出,病害虫の防御,除草などのようにたくさんの生物種を制御(生物 こううん かんがい 制御)するとともに,耕耘,加温,被覆,灌漑などのように生態系の物的要素を改変(物理制御) し,また,農作物の収穫によって奪われる土壌中の養分を補給する施肥(窒素,リン,カリ)のよ うに生態系の化学的要素を制御(化学制御)することを通じて,生態系における物質循環の過程に 介入する.農業にかかわる環境問題の特徴は,この生態系(生物の生育環境)の制御による物質循 環の過程への介入により引き起こされる物質循環の不全問題であるということができる.この不全 問題は,循環が破壊されることによって,あるいは阻害されることによって,さらには停滞するこ じゃっき とによって,または偏在(空間的,時間的)することによって惹起される.したがって,制御が異 なれば,生起される農業の環境問題は異なるし,他方,同一の制御が行われても,その対象となる 生態系が異なれば,農業の環境問題は異なるものとなる. 前掲の表において可能性として示されている農業による負荷の多くがすでに顕現していることか らも分かるように,これらの負荷の顕在化は,近代農業を性格づけている生態系の過度な利用と過 度な生態系の制御に起因している.農業の本質であるこの生態系の制御に伴う環境負荷が,生態系 の自己修復力(Resilience)を超える時,農業が依拠する生態系に重大な影響を与える.農業によ る負荷が顕在化する機構の解明,負荷を低減し,さらには削減し除去する技術開発などは,近代農 業に関する環境研究の原初的かつ今日的課題である. 上述したような生態系の制御を通じた農業の環境問題のほかに,農業の外的要因によって引き起 こされる問題がある.その一つは,食(農産物)の安全確保の視点から,土壌など農業資源(農業 の生態系)が汚染されていることが懸念される場合である.産業活動によりもたらされた農耕地土 壌の重金属(カドミウム,ヒ素)など有害化学物質による汚染はその一例である.その二つは,生 まんえん かくらん 態系の多様性を保全する視点から,侵入外来生物の蔓延が生態系を攪乱し,農業の生態系などに悪 影響を及ぼすことが懸念される場合である.交易等によってもたらされた特定外来生物カワヒバリ ガイの分布拡大によるパイプライン等灌漑施設の機能障碍はその一例である.その三は,地球規模 の環境問題が農業の生態系に影響を及ぼすことが懸念される場合である.オゾン層の破壊によって もたらされる紫外線量の増大が生態系を損傷する問題,大気中の二酸化炭素の増加や温暖化がイネ の生育や収量に影響を及ぼす問題などはその好例である. これらはいずれも生態系の変化にかかわる問題であり,そうした変化が農業に直接的影響を及ぼ すことによって,農業の持続性という問題を惹起する.ただし,先にも述べたように,生態系の変 化が自己修復力を超えるときに問題となる.また,農業の持続性の問題は「循環的生態システム」 において行われている物質循環の時間スケールが考慮されることによって認識される問題である. さらには,農業をめぐる社会経済条件も大いに関係する問題であることも付け加えておこう. 地球規模の,あるいは地域規模での農業内外の要因によって生態系に生じる問題は,農業におけ る環境問題であると同時に農業問題でもある. 土が密接な関係を結んでいる外部環境と相互の関係を示した中野政詩による地球表層の物質循環 の経路図(中野,1999)を参考に,農業(による生態系の制御)と地球表層における物質循環との 関係を整理すると図のように表される. 序 章 ᄢ᳇ 䇭䇭䇭 䇭䇭䇭䇭䇭䇭 ↢‛ ㄘᬺ ფ ㄘᬺ䈮䉋䉎ᓮ ‛⾰䈱ᓴⅣ ᳓ 図 地球表層における物質循環と農業との関係 農業が制御の対象とする生態系(農業生態系)は,大気圏,水圏(地表,地下),土壌圏(地 表,地下) ,生物圏(地上,地下)によって構成され,このそれぞれの間を太陽エネルギーがもた らす作用によって物質が循環している.ここでは,物質が循環するスケールあるいは生態系のス ケールは,取り上げる問題の視点によって変わってくる.農業が営まれる空間で起きている問題を 取り上げるならば,その生態系のスケールは,多くの場合,一定地域の広がりであろう.しかし, BSE(牛海綿状脳症)が引き金となって大豆ブームが始まり,アマゾンの熱帯林破壊を進めたよう ゆが に,生産と貿易の国際化が農産物輸出国の農業における生態系の制御のあり方に著しい歪みをもた らす.このように,土地が豊富な国でそれを集約的かつ収奪的に利用するなどによって生産費を低 く抑えて得られる農産物の大量・遠距離・一方向の移動は,輸出国の生態系を破壊するばかりでな く,地球規模での循環を攪乱することにもなる. 2.農業環境研究の枠組み ここに農業環境研究のあり方についてとりまとめた一つの資料がある.2001 年 4 月の独立行政 法人化に向けて農業環境技術研究所内に設けられた「将来検討委員会」のもとに農業環境研究のあ り方などについて検討したとりまとめである(西尾道徳,1999).そこでは,農業生産と環境保全 との調和を図りつつ,わが国の食料安全保障を確保し,国民の暮らしと国家社会の基盤である農 業・農村の有する多面的機能を維持・向上させることを目的とし,この目的を達成するために,以 下の6項目を農業環境研究として掲げている. (1)農業環境資源を維持・保全・活用しつつ,環境への負荷を最小限にするとともに,農業の持 つ公益的機能を最大限に発揮させながら,安全・高品質な食料を安定的に生産するための環 境管理技術の開発に向けた研究を行う. (2)農業環境資源の状態に関する情報を一元的に収集し,わが国の農業や自然条件等にふさわし い農業環境指標を用いて農業環境資源の変化を評価する. (3)農業にかかわる地域から地球規模までの空間について,食料の流通や消費の側面も含め,農 業を軸にして新しい時代のニーズに応える総合的な環境技術の開発戦略を策定する. (4)新たな施策提言に資する研究などを行う. (5)これらのために必要な問題については,農地のみならず,林地,水系,都市部等についても 研究対象を拡大するとともに, (6)研究実施に際しては経営・経済的側面も重視する. 2001 年 4 月に独立行政法人として再発足した農業環境技術研究所は,その独立行政法人個別法 において,前述の「将来検討委員会」で検討された目的および研究の範囲は大幅に縮減され,「農 業生産の対象となる生物の生育環境に関する技術上の基礎的な調査及び研究等を行うことにより, その生育環境の保全及び改善に関する技術上の向上に寄与する」ことを目的に与えられた.すなわ ち,研究の対象となる「農業環境」は,前者のように,「農業生産の対象となる地域」という産業 の視点でとらえた広い空間を基底に置く概念から,「農業生産の対象となる生物の生育環境」とい う狭い空間を基底に置く概念へと転換された. 環境研究は,現在の状態がどうなっているかを解明し,問題を解決するための知識を創造する営 為である.農業環境研究もこれと異なるものではない.ただし,対象とする環境が「農業生産の対 象となる生物の生育環境」に限定されているに過ぎない.環境研究には,その定義づけから導き出 されるように, 「問題がある」ことを発見し解明し解決するための研究(第 1 の研究)と「問題が ない」ことを解明し提示する研究(第 2 の研究)の二つがあることがわかる.第 1 の研究では,現 象を解明し,影響を評価し,必要な対策を導き出すことが研究の出口となる.農業環境研究では, 1.で述べたような農業にかかわる環境問題の解決を目指す研究はすべて第 1 の研究に属する.な お,すでに述べたように,農業の特殊性は,農業が自ら依拠する生態系を制御しつつ繰り返し利用 することにある.このことは,農業は一方では 1.で見たような環境問題を引き起こす原因となっ ているが,他方では,国土保全機能,環境保全機能,生物多様性保全機能,居住快適性機能,保健 休養機能など生態系が有する多面的な機能を維持し,増進するものともなっている.生態系が有す る多面的機能を解明し,評価し,維持し増進するための方策を導き出す研究もここに含めることが できる. 第 2 の研究では現象の解明と影響の評価が重要となる.農業が依拠する生態系をモニターし,そ の状態や変化を評価する研究である.農業がかかわる環境における放射性同位体の長期モニタリン グにより得られるデータのデータベースを構築し,その状態や変化を評価することはこの一例であ る. なお,生態系を構成する要素である土壌,植物,昆虫,微生物などを収集し分類し評価するこ と,気象や大気ガスを観測し評価すること,それらを情報化しデータベースを構築するとともに総 合化のための研究を実施すること,情報を収集し解析/分析し処理すること,さらには,リモート センシング(R/S) ,地理情報システム(GIS) ,統計解析など手法を開発することは,第 1 の研究 および第 2 の研究においてともに必要不可欠な基盤的研究である. 生産と貿易の国際化が農産物輸出国の農業による生態系の著しい改変に伴い種々の環境問題を引 序 章 き起こしていることが指摘されているが,他方で,農産物の輸入国においては窒素等が蓄積される ことによって生態系の富栄養化が進行するという現象も見られる.こうした国際経済社会とかかわ る環境問題に対処するために,さらには,農業による環境問題を回避しうる生態系制御のあり方を それぞれの地域,国において探求するためには,農業環境研究における国際共同研究の推進が重要 である. 2001 年 4 月から始まる第 1 期中期目標期間における中期計画においては,(1)農業生態系の持つ 自然循環機能に基づいた食料と環境の安全性の確保,(2)地球的規模での環境変化と農業生態系と の相互作用の解明, (3)生態学・環境科学を支える基盤技術に関する研究を柱に,22 の中課題に研 究を重点化した.これに続く 2006 年 4 月からの第 2 期中期目標期間における中期計画では,リス クの評価と管理に研究の焦点を当てて, (1) 農業環境のリスクの評価及び管理技術の開発,(2)自然 循環機能の発揮に向けた農業生態系の構造・機能の解明と管理技術の開発,(3)農業生態系の機能 の解明を支える基盤的研究を柱に,8 つの中課題に研究の重点を置いて実施した.独立行政法人化 以後の農業環境技術研究所における研究課題を,生態系制御が直接に介入する循環過程の規模をX 軸とし,自己修復力の範囲内に収まる程度の制御であるか否かといった生態系の自己修復力を基準 とする制御の相対的な強度をY軸とする平面上に位置づけて表すことを試みるならば,循環過程の 規模においては,植物の個体といったミクロなレベルから地球的規模のマクロレベルまでの広い範 囲にわたり,また生態系制御の相対強度においては残留性有機汚染物質(POPs)のリスクといっ た高いレベルから昆虫フェロモンのような低いレベルまでの広いレンジにわたる農業環境問題を研 究の対象として扱ってきていることがわかる. 3.本書の構成 本書は,序章およびそれに続く 4 部から構成されている.序章では,農業をめぐる環境問題を概 観し,農業環境研究の枠組みを提示している.農業生産環境の安全性を確保することは安全な食料 を生産するための必須要件である.第I部では,農業環境におけるカドミウム,ヒ素,残留性有機 汚染物質(POPs)などの有害化学物質や,肥料や有機質資材の施用に由来する窒素・リン等の栄 養塩類の農業環境における動態の解明,さらには,リスクあるいは負荷の評価とそれらの低減技術 について概説する.地球温暖化がわが国やアジア地域の主要作物であるコメの生産に甚大な影響を 及ぼすことが懸念されている.第Ⅱ部では,そうした懸念の解消に向けて,地球温暖化がもたらす コメの生産変動,イネおよび水田生態系の地球温暖化への応答など地球温暖化の影響評価や対応策 などについて述べる.また,農業活動による温室効果ガスの排出,農耕地土壌への炭素蓄積など, 農業における排出抑制や緩和策についても論じ,地球温暖化と農業との関係について全体像を明ら かにする.農業生態系の適正な管理および生態系の有する機能を活用することは農業の持続性に とって重要な要件である.第Ⅲ部では, 「豊かな生き物を育む場」としての農業生態系を維持し保 全することを目指して,そこにおける生物種の構成と動態および機能の解明,生物多様性の客観的 評価,侵入外来生物や組換え体作物が生態系に与える影響の評価などを述べる.また,農業生態系 では,多様な生物が,様々な生理活性物質を介して,互いに影響を及ぼしたり,環境中に存在する 化学物質を分解したりしている.それら情報化学物質とその機能についても述べる.農業生態系の 適切な評価と管理にとっては,生態系の長期にわたる継続的なデータの収集と有用なデータベース の構築,さらにはそれらのデータを分析し,解析する手法の開発が重要である.第Ⅳ部では,農業 生態系におけるガスフラックス,放射性物質など各種モニタリング,リモートセンシング(R/S) や地理情報システム(GIS)の利用,農業生態系に関する統合データベースについて述べる. 引用文献 1)中野政詩(1999)学術の動向 12 月号,日本学術会議,pp. 68-71 2)西尾道徳(1999)農業環境研究の将来方向(案),mimeo,pp. 7-8 3)四方康行,飯国芳明,谷口憲治 訳(1996)ドイツにおける農業と環境(Heissenhuber A., Meusel F., Katzek J., Ring H.Landwirtschaft und Umwelt, Economic Verlag, 1994),農政研究 センター国際部レポート,食料農業政策研究センター,p.58 Ⅰ.農業生態系における有害化学物質の動態とリスク低減 −食の安全 ・ 安心をめざして 第 1 章 重金属(カドミウム,ヒ素)汚染リスク評価と汚染土壌修復技術 1.農産物中のカドミウム,ヒ素の基準値等を巡る国内外の動向 カドミウムはヒトにとっては非必須元素であるにもかかわらず,必ず農産物には含まれており, 人体に毒性,蓄積性があるため,低濃度でも長期間にわたり一定量以上を摂取し続けると腎機能障 害などの健康被害を引き起こす可能性のあることが疫学調査等で明らかになっている.カドミウム の暫定耐容摂取量(PTWI:一生にわたってヒトがとり続けても健康への悪影響が現れないと見な される摂取量を体重 1kg,1 週間あたりで表した値)については,1998 年の JECFA(FAO/WHO 合同食品添加物専門家委員会)で 7μg/kg/週と設定された . また,2005 年 7 月にローマで開催さ れた第 28 回コーデックス委員会(FAO/WHO 合同食品規格委員会)総会では,食品中のカドミ ウム濃度の基準値が検討され,小麦の 0.2ppm,ばれいしょ(皮をむいたもの)0.1ppm,茎菜・根 菜(セロリアック・ばれいしょを除く)0.1ppm,葉菜 0.2ppm,その他の野菜(鱗茎類,アブラナ 科野菜,ウリ科果菜,その他果菜) (キノコ・トマトを除く)0.05ppm がそれぞれ国際基準値とし て採択された.しかし,わが国の主食である米については,0.4ppm を国際基準値案として予備採 択されることとなった. 一方,わが国では 1970 年に「食品衛生法」により玄米のカドミウム濃度を 1mg/kg 未満とする ことが定められ,その翌年から「土壌汚染防止法」によりカドミウム汚染地域が指定され客土事 業が開始された.今では,対策地域に指定された 6,428ha のほとんどで事業が完了している.しか し,今世紀になって,わが国の薬事・食品衛生審議会食品衛生分科会食品規格部会で再び審議が なされ,カドミウム摂取量において最も寄与率の高いコメについて,国内の含有実態に「ALARA の原則」 (合理的に達成可能な範囲でできる限り低く設定する(As low as reasonably achievable) との考え方)を適用し,国際規格に準じて基準値を 1.0ppm から 0.4ppm 以下に引き下げることが 決定され,2011 年 2 月 28 日から施行されている.これに伴いカドミウムに係る土壌環境基準(農 用地)の見直しも行われ,カドミウム汚染対策地域の指定要件等が改定された. また,コメ以外の農産物については,食品中カドミウム濃度の国際基準値制定の動きに対応して 国内の実態調査が行われ,様々な作物について国際基準値を超過する割合がもとめられている.今 回の国内基準値の検討では,コメ以外の品目について基準値を設定しないことで落着したが,政府 としては 3 〜 5 年後に国内実態,低減対策の実施状況について報告を求め,必要に応じて基準値の 設定を検討することになっているので,今後の動きには注意して行く必要がある. さらに,前述のカドミウムの暫定耐容摂取量については,国内基準値も食品安全委員会の審議を 経て 2009 年に 7μg/kg/週に設定されたが,2010 年 6 月に開催された第 73 回 JECFA では,カド ミウムの長期蓄積性を考慮すると耐容摂取量は月単位で表示するのが妥当であるとの考え方が示さ れ,また,基準値も従来のカドミウムの PTWI 7μg/kg/週よりは若干厳しい PTMI 25μg/kg/月 という値が提示されている. 厚生労働省国立医薬品食品衛生研究所の調査結果からの試算によると,日本人一人あたりの摂取 量は平均 3.0μg/kg/週程度と推定されている.したがって,日本の国民は平均的に耐容摂取量の 4 1.農産物中のカドミウム,ヒ素の基準値等を巡る国内外の動向 カドミウム 魚介類, その他, 0 11.5 野菜・海 藻, 14.7 米,43.4 有色野 菜, 10.9 図 1 日本人の食品群別カドミウ ム,ヒ素摂取割合(平成 12 〜 16 年度平均) ヒ素は総ヒ素摂取量の割合 (無機態ヒ素量の摂取割合で はコメは 17.2%) 豆・豆加工 品, 4.2 ヒ素 雑穀・ 芋, 9.7 その他, 3.3 魚介類, 55.9 米, 8.9 雑穀・芋, 1.2 豆・豆加工品, 0.6 有色野菜, 0.2 野菜・ 海藻, 29.9 割程度を摂取しているという計算になるが,確率論的な分布による推定をしてみると日本人のカド ミウム摂取量の 95 パーセンタイル値が 7μg/kg/週付近になるので,少なくとも国民の数%以上が 暫定耐容摂取量を超過しているという推測も成り立つ.いずれにせよカドミウムは国内外において 最も優先的にリスク管理を行うべき有害化学物質の評価を受けており,今後,さらなる低減化に向 けた研究を重点的に進めて行く必要があると考えられる. ヒ素については,わが国ではイネの生育障害防止の観点から農用地土壌汚染防止対策法を実施す る基準値が 15mg/kg と設定されているものの,食品中の基準値としては残留農薬基準値が決めら れているだけで,総ヒ素濃度や形態別の基準値の設定には至っていない.それは,食品中のヒ素の 化学形態や形態別毒性の解明が不十分であること,形態別の分析方法に問題があること,基準値設 定をどの形態に適用すべきか十分な根拠が得られてないこと等を主な理由としているが,わが国の 食品安全委員会でも 2009 年 3 月に「自ら評価を行う案件」に選定し,化学物質・汚染物質専門調 査会において食品健康影響評価を行っており,今後,厚生労働省において,コメを含めた基準値設 定に向けた検討が開始される可能性もある. JECFA によるヒ素の暫定耐容週間摂取量(PTWI)の設定は,1983 年に飲料水からの無機態ヒ 素摂取に係る複数地域の疫学的調査結果を基に 15μg/kg 体重/週とされた.しかし,2009 年 2 月 に出された JECFA の評価では,肺がんの発生に係る BMDL0.5(がんの発生率が 0.5%増加する無 機ヒ素の摂取量(これをベンチマーク用量:BMD という)の安全側の 95%信頼下限値)を飲料水 中の無機ヒ素濃度と肺がんに関する疫学調査から推定して,3.0μg/kg 体重/日(2 〜 7μg/kg 体 重/日)という値が設定された.食品に含まれるヒ素には様々な化学形態があり,その形態により 毒性が異なるが,ラットを用いた経口投与による LD50 値(50%致死量)では,無機ヒ素(亜ヒ 酸) ,無機ヒ素(ヒ酸) ,有機ヒ素(ジメチルアルシン酸)の順に急性毒性が高い.しかし,遺伝毒 性試験や発がん性試験では有機ヒ素も有害な作用の可能性が報告されている. 以上のように,ヒ素に関しては食品中の化学形態別の評価が定まっておらず,今後,食物に関す るより詳細な情報が必要とされる.わが国のトータルダイエット調査によれば,日本人の無機ヒ 第 1 章 重金属(カドミウム,ヒ素)汚染リスク評価と汚染土壌修復技術 素摂取量は 62.8μg/人/日と推定され,そのうちコメからの摂取量は 10.8μg/人/日(総ヒ素からの 換算係数として 0.86 を使用)と農産物の個別品目では最大で,主要な部分を占めている(「食品中 に含まれるヒ素の食品影響評価に関する調査」平成 20 年度調査報告,財団法人国際医学情報セン ター) .したがって,有機ヒ素の毒性に明確でない部分があるものの,無機ヒ素摂取源として寄与 率の高いコメに含まれるヒ素濃度を低減させることがわれわれの食生活におけるヒ素のリスク管理 にとって非常に重要なことは以上の結果から充分に結論できる. 2.農作物におけるカドミウム,ヒ素の吸収 ・ 移行特性の解明 農産物中のカドミウム,ヒ素等の重金属濃度を低減するためには,作物における重金属の吸収 ・ 移行特性,および環境要因との関係等を明らかにする必要がある.すなわち,対象としている重金 属が作物の生育のどの段階でどのくらい吸収され,それが可食部にどのようなメカニズムで蓄積さ れて行くのか,吸収は主にどのような環境要因に支配されているのか,作物品目 ・ 品種等において 吸収能の差異は何に由来するか等を解明することが技術開発を進めるための知見として不可欠であ る. 農業環境技術研究所では,最近 10 年間にわたり主にダイズとコメを中心にこれらのメカニズ ム解明を進めてきた.まず,研究手法における技術開発として,カドミウムの安定同位体である 113 Cd を直接土壌中に投与する簡便で精度の高いほ場試験法を開発した(平成 13 年度研究成果情 報,川崎ら) (川崎・織田,2005) .この技術を用いて,ダイズの生育前期に吸収されたカドミウム が生殖生長期以降に子実に移行することや,カドミウムの添加時期を変えてポットで生育させたイ ネの実験から,生殖生長期に添加したカドミウムは葉部に,それ以降に添加したカドミウムは茎部 に集積しやすいが,出穂直前に添加したカドミウムは穂に移行しやすいことなどが明らかにされて いる.また,113Cd で標識した化成肥料を試験的に製造し,土壌中に施用した化成肥料中に不純物 として含まれるカドミウムの作物への移行が調査された(平成 14 年度研究成果情報,織田ら).そ の結果,ダイズが吸収したカドミウムのうち,全体の 11%が肥料由来であったが,それは肥料に 含まれていたカドミウムの 0.13%にすぎず,大部分は土壌に蓄積するので,カドミウム含量の高い リン酸肥料や汚泥肥料を連用する場合には蓄積レベルの増加が要注意であることが指摘された. このように,カドミウムの安定同位体は,植物が吸収した時期や植物体内中で蓄積する部位を 特定する解析手段として使えるので,水耕栽培でもより詳細な解析が行われている(平成 16 年度 研究成果情報,川崎ら) (箭田ら,2004a,b) .水耕栽培したダイズ「エンレイ」 について調べた試 験によると,子実へのカドミウムの移行は着莢盛期(R4)から粒肥大盛期(R6)に多く,その時 期に吸収したカドミウムが全子実含量の約 50%を占め,子実のカドミウム濃度に最も影響するこ とがわかった.また,茎から転流したカドミウムの成熟期(R8)の子実含量に及ぼす影響は小さ く,子実の全カドミウム量の約 20%であった.これらの結果は,先のほ場試験で得られた移行パ ターンを支持するものであり,子実のカドミウム蓄積を抑制するには着莢盛期から粒肥大盛期まで の間にカドミウムの経根吸収を抑制することが肝要という結論が導かれた.カドミウムの安定同位 体は,後述する導管にカドミウムを輸送する能力の品種間差をナスで解析する際にも有効に利用さ れた. 3.農作物の品種 ・ 系統間差や育種を利用したカドミウム低減技術の開発 ⑴ 農作物の品種 ・ 系統間差の解明とその利用 農作物のカドミウム吸収能の品種・系統間差は,これまでいろいろな作目で調査されているが, 10 3.農作物の品種 ・ 系統間差や育種を利用したカドミウム低減技術の開発 どの作目も品種 ・ 系統によって大なり小なり違いがある.その違いを生じる原因を解明するために 子実のカドミウム濃度の低いダイズ品種の吸収・蓄積機構が調べられた(平成 14 年度研究成果情 報 , 阿江ら) (Arao et al., 2003) .その結果,子実カドミウム濃度の低い品種ほど根のカドミウム濃 度が高く,一般に低吸収系統といわれているものは,カドミウムが根の細胞壁に強固に結合して根 に蓄積し地上部への移行が妨げられていることが原因と考えられた.このことは,ダイズの低吸収 品種を台木として接ぎ木栽培をすると地上部(穂木)へのカドミウムの移行が抑制されることから も確認することができた(Sugiyama et al., 2007). このダイズの接ぎ木により得られた結果をさらに発展させた成果として,ナスの接ぎ木栽培を挙 げることができる.国内産ナスは,現在,半分以上が土壌病害防除等を目的として接ぎ木栽培が行 われているが,この接ぎ木栽培において,台木の品種選択と果実のカドミウム濃度の関係を調べて みると,ダイズと同様に台木に用いる品種をカドミウム吸収の少ない品種にしてやれば果実のカ ドミウム含量を確実に低く抑えられるということがわかった(平成 19 年度研究成果情報,荒尾ら) (竹田ら,2007;Arao et al., 2008) .すなわち,スズメナスビ系のトルバムビガーやトナシムを台 木として用いると,穂木の種類,土壌タイプ,作型等によらず果実へのカドミウムの移行を台木の ところでかなりブロックすることができて子実のカドミウム濃度を 1/3 以下に減少できるというこ とが示された.ナスの果実へのカドミウム集積の違いを決定する要因を取り込み速度から解析した ところ,根細胞から導管へカドミウムを輸送する能力の差異がカドミウム果実濃度の品種間差を支 配している主要因と考えられた(Mori et al., 2009a, b).このナス台木のカドミウム輸送能力の違 いを解明するために根の切片試料の表皮から中心柱を含めた領域についてシンクロトロン放射光源 マイクロビーム蛍光 X 線による元素マッピングが行われた.その結果,輸送能力の低いトルバム ビガーでは,根の先端からの距離にかかわらず,表皮および内皮にカドミウムが局在しており,中 心柱付近におけるカドミウム濃度が相対的に低いのに対し,それより輸送能力の高い台木品種(千 両二号)では中心柱付近においても表皮,内皮と同程度のカドミウムが存在していた.したがっ て,地上部へのカドミウム移行の抑制は,表皮と内皮に存在するカスパリー線における導管への移 行段階で起こることが示唆された(Yamaguchi et al., 2011). 一方,イネについては,世界各地の多様なイネ品種・系統をカドミウム汚染土壌で栽培し,玄米 カドミウム濃度を比較すると,2 倍〜 10 倍以上の品種間差異のあることが明らかとなった(平成 20 年度研究成果情報,石川ら) (Arao and Ae, 2003).このイネについても,ナスと同様に子実へ のカドミウム集積を支配する要因は,導管へのカドミウム輸送能力であることが「ササニシキ」 と 「ハバタキ」 のカドミウム取り込み速度の比較解析から証明されている(平成 21 年度研究成果情 報,石川ら) (Uraguchi et al., 2009) . 一般的に,イネのジャポニカ品種はインディカ品種に比べ子実のカドミウム含量が低いが,広く 調べてみると玄米カドミウム濃度が「コシヒカリ」等の約半分と日本の主要品種よりさらに低い 「LAC23」という熱帯ジャポニカ品種の存在することがわかった.この「LAC23」は出穂が遅く, 長稈,長粒,低収量であるため,そのままでは日本における実用的な食用イネ品種には適さない. そこで,草姿が良好な安定多収品種「ふくひびき」と交配して実用的な性質を付与しようと試みた ところ, 「LAC23」に比べて出穂が早く草丈が低い 5 系統(「羽系 1118」〜「羽系 1122」)を選抜 することができた(平成 20 年度研究成果情報,石川ら)(山口,2006).これらの系統は,東北地 方でも栽培可能で,また,わが国の在来食用品種に比べて玄米カドミウム濃度が 40 〜 50%程度低 くなったにもかかわらず,カドミウム以外の重金属ミネラル成分(銅,鉄,マンガン,亜鉛)につ いては影響がなかった.今後,実用的な低カドミウム食用品種を育成するためには,カドミウムの 第 1 章 重金属(カドミウム,ヒ素)汚染リスク評価と汚染土壌修復技術 11 イネの品種間差 低Cd集積品種(食用に利用) 低Cdだが、良食味ではない、 栽培し難い 高Cd集積品種(ファイレメ用) 実用性に 乏しい 不良形質の改良(育種) 高Cdだが、インディカ品種の ため、脱粒や倒伏等の栽培特 性に難あり ①従来の交雑育種法 ② DNA マーカー育種法 ③突然変異育種法 ④遺伝子組換え技術による育種 実用的なファイレメ品種、低Cd品種(食用)の開発 図 2 イネの Cd 吸収特性を利用した Cd 低減技術(石川覚・農環研) 低吸収や栽培の地域適応性にとどまらず,安定多収や良食味等の特性を付与して行くことが必要と なる.イネで実施されているこのようなカドミウム低吸収品種との交配による新品種・系統の育成 は,現在,ムギやダイズでもそれぞれ試みられている. ⑵ イネのカドミウム吸収の遺伝子解析 植物の遺伝形質は質的形質と量的形質に分けられるが,このうち量的遺伝形質に関与する遺伝子 座を QTL(Quantitative Trait Loci)といい,その解析手法を QTL 解析と呼んでいる.カドミウ ムの吸収・集積は,量的形質の一つと見なされるが,一般的に,量的形質は複数遺伝子が関与する 複雑な仕組みになっており解析が難しい. 遺伝的背景が「コシヒカリ」で,インド型品種「カサラス」 の染色体断片が各々 1 カ所のみ大き く置換された 39 系統群(農業生物資源研究所で作出)を利用して玄米カドミウム濃度に関わる遺 伝子座(QTL)を検索したところ,第 3,6,8 染色体上に関係する QTL の存在することが明らかと なった(平成 16 年度研究成果情報,石川ら) (Ishikawa et al., 2005).また,ジャポニカ品種「サ サニシキ」とインディカ品種「ハバタキ」の戻し交雑自殖系統群について,玄米のカドミウム集積 に関わる QTL を第 7 染色体の短腕側に同定した(平成 21 年研究成果情報,石川ら).さらに,こ の QTL は銅,鉄,マンガン,亜鉛の必須重金属の玄米集積には関与せず,カドミウムのみを特異 的に集積することを確認した. この第 7 染色体の遺伝子座に関しては,秋田県立大学,岡山大学等のグループで並行して解析が 進められ,最近,イネのカドミウム集積に関する品種間差のメカニズムとそれを支配する遺伝子が 明らかにされた(Ueno et al., 2010; Miyadate et al., 2010). それらによると,子実へのカドミウ ムの集積は,イネの根の液胞膜に存在する OsHMA3 タンパク質による液胞内への選択的な輸送と 隔離に支配されており,カドミウム高集積品種は,そのタンパク質遺伝子の機能欠損型であった. この膜タンパク質の遺伝子は塩基配列も明らかにされている.このようにカドミウムの集積機構に 関する遺伝子の解明が進んで来たことで,今後,カドミウム低吸収の食用品種の育成とともに,カ ドミウムを多量に集積するファイトレメディエーション用実用イネ品種の作出にも進展が大いに期 待される. 12 5.栽培管理による農作物のカドミウム,ヒ素低減技術 4.農耕地へのカドミウムの負荷 わが国は世界各国の中でもとりわけカドミウム(Cd)の生産量,消費量が多く(供給量(生産 量+輸入量)約 5 千トン,消費量 4650 トン:2001 年),1960 年代から 1970 年代にかけては顔料, メッキ,塩ビ安定剤など様々な用途に使われてきた.また,かつては主として亜鉛鉱石の採掘や亜 鉛製錬に付随して鉱排水やばいじんに含まれる形でカドミウムが周辺環境中を汚染してきた.しか し,近年,鉱山や精錬所の排出規制が厳しくなり,また,カドミウムの用途も 90%以上がニッケ ル-カドミウム電池に限定されるようになったので,新たな環境汚染は減少してきている. 農地におけるカドミウムの汚染も年々減少してきているが,つくば市真瀬地区のカドミウム非汚 染水田における年間収支を推定した結果,水田への供給は肥料由来が最も多く,降水,かんがい水 と合わせて 3.2g/ha/年となり,また,コメの収穫,排水,下方浸透等による排出は 0.8g/ha/年と なった(平成 19 年度研究成果情報,荒尾ら) .ここで稲わら,もみがら等をほ場に還元したとして も,土壌カドミウム濃度の上昇は 1.6μg/kg/ 年と推定され,土壌総カドミウム濃度(0.35mg/kg) と比較して 2 桁ほど低いことがわかった. このように非汚染水田においては,カドミウムの負荷は非常に小さいものの,農地に有機性廃棄 物を連用する場合,カドミウムの負荷量が作物中のカドミウム濃度増加につながる可能性もある. そこで,北海道と熊本で畑地に施用される有機性廃棄物により持ち込まれるカドミウムの作物への 影響が調査された(平成 19 年度 研究成果情報,川崎ら) .有機性 単位;mg ha -1 廃棄物の中でもホタテ中腸腺やイ カ肝臓など水産廃棄物にはカドミ ウム濃度が高いものもあったが, 玄米 128 その他の有機性廃棄物のカドミウ 灌漑水 降水 172 1020 稲わら 1008 ム濃度の中央値は 2mg/kg 以下で あった.有機性廃棄物由来の堆肥 を 5 年間連用した農地ではカドミ 排水 573 ウム負荷量が持ち出し量を上回っ たが,作物中のカドミウム濃度は 増加しなかった.その理由として は土壌 pH の上昇と施用有機物に よるカドミウムの不溶化が考えら 浸透水 91 土壌Cd増加量 2400 (0.0016mg kg -1) 肥料 2000 図 3 真瀬水田における年間カドミウム収支の試算 れた. 5.栽培管理による農作物のカドミウム,ヒ素低減技術 日本人のカドミウム摂取量全体に占める食品別の内訳をみると,コメ由来のカドミウムが全摂取 量の 43%と半分近くを占めているので,コメのカドミウム濃度を合理的な範囲でできるだけ低くす ること(ALARA の原則)が重要である.これまで湛水処理によって玄米のカドミウム濃度の上昇 を大幅に抑制できることが多くの研究で明らかとなっているが,湛水条件下では水田土壌の還元化 が進み,土壌溶液中のカドミウムイオンは硫化カドミウム等の化合物を形成して不溶化するため, 水稲のカドミウム吸収が抑制される.特に,出穂期前後のカドミウム吸収が最も盛んな時期に湛水 管理を継続すると玄米中のカドミウム濃度は非常に低いレベルに抑えられる(稲原ら,2007).農 第 1 章 重金属(カドミウム,ヒ素)汚染リスク評価と汚染土壌修復技術 13 業環境技術研究所では,この湛水処理を技術の核とする「水稲のカドミウム吸収抑制のための技術 確立マニュアル(初版 2002 年,改訂版 2005 年) 」を農林水産省と連名で作成した.このマニュア ルは農林水産省の事業やカドミウムの基準値超過が懸念される地域等で広く活用されている. しかし,湛水管理に関しては,二つの大きな問題がある.第一に,水稲のカドミウム 吸収抑制 のために長期間にわたって水田を湛水状態に保っておくと,水田の地耐力が低下して大型機械を 使った収穫作業等に著しい支障を来す.第二に,水稲に含まれる有害性重金属としてはカドミウム 以外にヒ素も問題となるが,水稲のヒ素吸収はカドミウム吸収とトレードオフの関係にあり,水田 の湛水処理により土壌の還元化が進むほど毒性の強い亜ヒ酸の土壌溶液中への溶出と作物による 吸収が促進される.最近のポット試験等による結果では,土壌中のヒ素濃度や水田土壌の湛水時期 等の違いによって,玄米の総ヒ素濃度だけでなく,全体に占める無機ヒ素と有機ヒ素(ジメチル アルシン酸)の割合も大きく変化し,また出穂前よりも出穂後の湛水の有無の影響が大きかった (Arao et al., 2009) . OH OH HO As III OH 亜ヒ酸 OH HO As V O H 3C A s OH O H3 C A s OH OH ヒ酸 V モノメチルアルソン酸 V O C H3 ジメチルアルシン酸 図 4 ヒ素の化学形態と構造式 食品に含まれるヒ素については,化学形態別の割合やそれぞれの形態の健康影響(無機ヒ素に比 べ毒性の低い有機ヒ素も発がん性の懸念がある)の基礎的なデータ等が不十分なことから,まだ基 準値の設定に至っていないが,日本人にとって摂取寄与の大きなコメについてカドミウムとヒ素を 同時に低減することは非常に重要と考えられるので,今後,技術開発が急がれる. 土壌の還元条件以外でカドミウム吸収抑制に著しい効果を及ぼす要因には,土壌 pH,EC など がある.特に,土壌が酸性側に傾くと土壌 pH の低下とともにカドミウムが土壌液相中に溶けやす くなるので,吸収抑制のためには石灰等のアルカリ性の肥料・資材を施用し,土壌の pH を上げる ことが有効な手段となる.様々な事例においてアルカリ性資材の効果が報告されているが,それを 実際の生産現場で使うとなった場合,一方では土壌の pH を持続的に安定してコントロールするこ とや作物根圏全体の pH 調整が難しいことがあり,また,他方では pH の調整による影響が他にも 多岐にわたっているため,良好な作物生育・収量をどう確保するかについて栽培技術上の工夫が必 要である.また,アルカリ性資材とともに,堆肥等の有機物施用のカドミウム吸収抑制効果も数多 く報告されている.このような効果の認められる原因として,部分的には,土壌 pH の酸性化を抑 制する有機物の緩衝作用や,有機物により生育が促進されることによる希釈効果がまず考えられる が,そのほかに有機物のカドミウム吸着作用など,いくつかのメカニズムも考慮する余地があるの で,メカニズムを明らかにした上で安定的な技術開発を進めて行く必要があろう.以上の研究は, 農業環境技術研究所においても農林水産省の委託プロジェクト「生産・流通・加工工程における体 系的な危害要因の特性解明とリスク低減技術の開発」等で重点的に進められている. 6.農作物可食部のカドミウム濃度予測手法の開発 農作物収穫物中のカドミウム,ヒ素等の重金属濃度を低減するために,作物の収穫前に収穫物の 14 7.カドミウムの分析技術の開発 カドミウム濃度を予測する技術を開発することも重要である.農用地土壌汚染防止法の公定法で は,0.1M 塩酸抽出法が土壌中のカドミウム(Cd)測定法となっているが,必ずしも収穫物可食部 中のカドミウム濃度との相関性は高くない.そこで,0.1M 塩酸抽出法以外の様々な抽出法につい て,畑作物可食部の Cd 濃度を予測することを目的として最適の土壌抽出法の選抜や,ダイズやコ ムギの子実 Cd 汚染リスク予測式の開発が現在検討されているところである. 別の視点からの方法としては,土壌に添加した安定同位体(111Cd)が土壌溶液と固相で平衡状 態に達するまでの経時変化から植物に吸収されうる潜在的な量を推定する手法が提案されている (川崎・箭田,2007;Kawasaki and Yada, 2008) (平成 20 年度研究成果情報,川崎ら).すなわち, 土壌中のカドミウムは,溶存態,交換態,結合態の 3 つに大別できるが,植物は土壌溶液に溶解し ているカドミウムを吸収するので,溶存態と交換態の合計量が植物に吸収されうる潜在的な量と考 えられる.この手法では,土壌に 111Cd を添加して 111Cd 比率の経時的変化から植物の吸収可能量 (以下 E 値という)を推定しているが,推定結果をみると E 値は土壌中の全カドミウムの 4 〜 6 割 に相当することや,微量のカドミウムを含む有機質の資材を 5 年間連用しても E 値がほとんど変 化しないことなどが明らかになった. 生育がある程度進んでからの予測であれば,イネについては,出穂期以降の土壌溶液中のカドミ ウム濃度と玄米中カドミウム濃度の間に相関が認められるので,ポーラスカップにより採取した土 壌溶液の分析値により,玄米中カドミウム濃度の推定が可能であることが示された(平成 14 年度 研究成果情報,櫻井ら) . 一方,カドミウム汚染土壌を充填したポットで播種後 3 週間まで栽培したダイズ幼植物体中のカ ドミウム/亜鉛の濃度比とダイズ子実中のカドミウム濃度は相関性が高く,ダイズ幼植物の段階で 子実カドミウム蓄積性の高い品種を簡易に検定できることが示された(Sugiyama et al., 2007)(平 成 15 年度研究成果情報,杉山ら) .この知見は,カドミウム蓄積性の品種間差を効率的に予測でき る手法としてダイズ有望品種の選抜等に活用できる. 7.カドミウムの分析技術の開発 作物体可食部中のカドミウムは,近年,高精度の ICP 発光分光法や ICP 質量分析法が用いられ ているが,前処理に強酸での分解や煩雑な操作が必要なため短時間に多数の検体を分析することが 困難なことや,分析機器が高額なため農業現場近傍などで手軽な分析ができないという問題があっ た. そこで,分析の迅速化,効率化を図るために,コメの粉砕,強酸分解の工程を省略し,玄米粒の ままの状態でカドミウム濃度を分析するレーザーアブレーション ICP 質量分析器を用いたカドミ ウム分析法を開発した(Baba et al., 2003) .この方法では,極めて短時間に玄米中のカドミウム濃 度を評価することができたが,濃度が 0.3ppm になると直接分析が不可能であった. また,農業の生産現場や農産物の流通段階で利用できる簡便な方法としては,イムノクロマト アッセイ法とボルタンメトリー法が開発された.イムノクロマトアッセイは,Cd-EDTA 錯体とこ れに特異的に反応する抗 Cd-EDTA 抗体との抗原抗体反応を利用した検出キットを用いるもので, 玄米の場合は希塩酸でカドミウムの抽出,カラム精製をした後に,検出キット上の反応後の発色値 からカドミウム濃度を簡易・迅速に測定することができる(阿部ら,2006).現在では,コメ以外 の作物への適用も検討されており,基本的に,野菜は生でミキサー磨さい後,希塩酸抽出ベースの 前処理法で測定可能となっている.前処理工程での操作法の工夫(Cd 分離精製カラム操作や緩衝 液による希釈倍率変更など)によってトータルの測定感度を向上させることにより,国際基準値の もっとも低い野菜(0.05mg/kg)な どにも適用できるように改良が進 められた(平成 22 年度研究成果情 報,阿部ら) .また,品目によって は測定が困難なものもあるが,たと えば脂質が多く分離困難なダイズで は,焼成処理や濾過促進剤を希塩酸 抽出に併用することにより問題を解 決した. ボルタンメトリー法は,電解液中 に分極性の電極と非分極性の対極を -1 イムノクロマト測定によるCd濃度(mg kgFW ) 第 1 章 重金属(カドミウム,ヒ素)汚染リスク評価と汚染土壌修復技術 15 入れ電極間の電圧を連続的に変化さ ホウレンソウ生 2.0 1.5 y=1.010x R 2=0.933 1.0 0.5 0.1mol 塩酸 1:5 抽出 0.0 0.0 0.5 1.0 1.5 2.0 酸分解ICP-MS 測定によるCd濃度(mgkgFW -1 ) 図 5 生ホウレンソウのイムノクロマト測定 せて得られる電流変化曲線から重金 属イオンの定性,定量を行う手法であるが,これについても,コムギ,ダイズの前処理法として Cd 分離精製カラムや簡易硝酸分解法を用いて,最適条件等を明らかにした.コムギやダイズの子 実中のカドミウムは 0.2mg/kg まで,変動係数 10%以下で測定可能となっている. 8.カドミウム低減のための土壌修復技術 農作物のカドミウム吸収を低減する栽培技術はこれまで見てきたように様々な方法があり,それ ぞれ有効に活用できる場面がある.しかし,現実にはそれらの技術を取り入れることが労力やコス ト面での負担になる場合も多く,また,カドミウム低減化のために導入した技術が他の重要な要因 とトレードオフの関係にあったり,副次的な悪影響を生み出したりする可能性もある.そのような 点を考慮すると,やはりカドミウム基準値超過のリスクを抱えている農地には,土壌修復(浄化) による抜本的な対策を検討すべきである. これまで,一般的に重金属汚染土壌の修復法としては,客土が行われてきた.カドミウム汚染土 壌の場合も,主に 20 〜 40cm の非汚染土の盛り土をする上乗せ客土により,土壌汚染防止法にお けるカドミウム汚染の指定対策地域への対策が講じられている.しかし,この客土法は非常にコス トがかかる上に,最近では,客土に適当な非汚染土の確保が難しくなってきている.さらに,肥沃 度の低い山土等を用いると生産力を回復するまで何年もかかることになる.このような問題点があ るために,その代替法として農業環境技術研究所で開発したのが土壌洗浄法と植物修復法(ファイ トレメディエーション)である. ⑴ 土壌洗浄法 土壌洗浄法は,化学薬剤を溶かした用水を流し込んで湛水した水田で土壌を撹拌することにより カドミウムを溶出させた後,溶出水の浄化とカドミウムの回収をオンサイトで行うことにより土壌 浄化する技術である.カドミウムを溶出する薬剤としては,比較検討の結果,溶出効率が高く土壌 への作用が比較的穏やかという点から塩化第二鉄が最も適切であるという結論が得られた(平成 16 年度研究成果情報,牧野ら) (Makino et al., 2008).この薬剤を用いたことと,水田を撹拌する ときの水深を 43 〜 45cm と深くしたことにより,水田土壌のカドミウム濃度を 60 〜 80%程度, そこで生産される玄米中のカドミウム濃度を 70 〜 90%程度まで低減できるということが全国のい くつかのほ場レベルの実証試験で明らかにされている(平成 22 年度研究成果情報).この手法で は,客土法に匹敵するカドミウムの低減化が達成されており注目に値するが,一方ではカドミウム 16 8.カドミウム低減のための土壌修復技術 溶出の副作用としてマグネシウムやカルシウム等の塩基類までも同時に溶出してしまうという問題 も派生する.また,土壌の入念な撹拌処理により土壌物理性が多少悪化する懸念もある.しかし, これらの問題は資材施用による塩基類の補填や堆肥施用による物理性改良等で問題にならないレベ ルまで回復可能であることが,処理後の作物栽培試験等による検討で確かめられている.また,処 理水放流による周辺水生生物への影響も 2 倍以上に希釈すれば問題のないことも確認された.以上 の点から判断すると,この手法は土壌修復の手段として,今後,有力な選択肢の一つになると考え られるが,コストの試算結果では 10a あたり約 300 万円と高めなので,低コスト化に向けて手法の 改良が検討されているところである. ⑵ ファイトレメディエーション もう一つの修復法で あるファイトレメディ エーションは,カドミ ウム吸収能力の高い植 物を栽培して土壌のカ ドミウムを除去しよう ①薬剤散布(塩化第二鉄) ②攪拌(Cdの抽出) ③静置・排水 ④排水処理 ⑤Cd含有汚泥の回収 ⑥処理水の放流 という方法である.こ の目的に適する作物種 を選定するために,複 数の異なる土壌タイプ について,イネ,ダイ ス, ト ウ モ ロ コ シ な 図 6 土壌洗浄技術の概要 どの地上部へのカド ミウム集積量が畑条件で比較検討された(平成 15 年度研究成果情報,村上ら)(Murakami et al., 2007) .その結果,調査した作物の中では,イネのカドミウム吸収量が最も多く,カドミウムの可 溶性画分だけでなく難溶性画分の吸収も多いことがわかった.また,温暖地(九州地方)の現地圃 場におけるカドミウム吸収試験においても,密陽 23 号,IR8 などのカドミウム吸収能の高いイネ 品種が適していることが明らかになった. 従来,ファイトレメディエーションには,重金属で汚染されている鉱山跡地などに生育している 重金属に耐性のある重金属超集積植物を使用することが検討されていた.しかし,わが国の農地の 汚染土壌はカドミウム濃度が数 ppm を超えるような高濃度の汚染土壌はほとんどなく,適用する 修復植物に必ずしも高濃度の耐性は必要ではない.むしろ,土壌からのカドミウム吸収量は植物生 育量(バイオマス)と植物体カドミウム濃度の積で決まる訳であるから,高濃度重金属の選択圧の ない農地で通常生育がかなり劣る重金属集積植物を使うよりは,突出した重金属蓄積能はなくても おう盛な植物生育で盛んにカドミウムを吸収する植物を利用した方が有利であるといえる.さら に,従来の植物修復がうまくいかなかった理由として収穫残渣の処理システムまで考えに入れてい なかったという問題があるが,収穫した植物体を効率的に処理・利用しうる機械化システムをいか に構築するかということが,実用技術を開発する上でもう一つの肝要な点である.イネであれば, わが国の農家は栽培に慣れており,既存の機械が利用可能,地域適応性の高い品種の候補も豊富 で,連作可能等の多くのメリットがある.以上の観点から,農業環境技術研究所では,現在,長香 穀や IR8 などのカドミウム高吸収イネを中心に技術開発を進めている. 技術開発のポイントとしては,カドミウム吸収量を最大化する栽培法と収穫した植物体の効率 第 1 章 重金属(カドミウム,ヒ素)汚染リスク評価と汚染土壌修復技術 17 的な処理法が重要で 効率的な Cd吸収・除去 あ る. 前 者 に 関 し て 収穫、乾燥および在庫技術 Cd回収技術と有効利用 は,苗移植後 1 か月か ら 1.5 か月は湛水栽培 してイネの生育を確保 し,その後落水を継続 ◆収穫・天日乾燥 してカドミウム吸収を ◆ 高Cd吸収品種の選抜 促 進 す る「 早 期 落 水 ◆ Cd吸収を高めるための 栽培技術 法」が提案された.ま 落水・収穫時期、 最適窒素施肥量 た,後者に関しては, ◆焼却(Cd回収) 残渣 熱エネルギー バイオマス有効利用 ◆回収・梱包機械化作業 ◆ バイオエタノール ◆ 生分解性プラスチック ◆ 土壌修復効果の検証 最初にもみだけを収穫 ◆ バイオマスガス化等 し,その後天日で乾燥 した稲わらをロール状 ◆収穫植物の在庫乾燥 にまるめて収穫する 図 7 ファイトレメディエーション一貫システム 「もみ・わら分別収穫 法」と,ロールの上部を透湿防水シートでおおって水田に数か月置く「現場乾燥法」により,効率 的に低コストで焼却処理できる植物体一貫処理システムを開発した.このようなシステムでカドミ ウム高吸収イネを 2 〜 3 作栽培することにより,土壌中のカドミウム濃度を 20 〜 40%,後作食用 イネ玄米のカドミウム濃度を 40 〜 50%低減化することに成功した(Murakami at al., 2009)(平成 21 年度研究成果情報,村上ら) . 以上に示した客土法,土壌洗浄法,ファイトレメディエーションの 3 つの土壌修復法の長所,短 所を表 1 にまとめた.それぞれの方法に短所,長所があるので,修復の条件によりケース・バイ・ ケースで手法を選択して行くことが現時点では最も適切と考えられる. 表 1 土壌のカドミウム汚染修復技術の比較 客土 土壌洗浄 ファイトレメディエーション メリット ・抜本対策として確実 な効果が得られる ・客土工事自体は 1 年 間で終了 ・短期間で工事が終了 ・事前の室内試験で浄 化効果を精度良く予 測できる ・コストが安い (25 万円 /10a/ 年程度) ・修復による土壌の劣化 がほとんどない ・広い面積に適用できる デメリット ・コストがかかる ・ 水持ちの悪い水田では (300-600 万円 /10a) 施工が困難 ・客土に利用できる土 ・対策時(大抵は冬場) の確保が困難 に大量の水が必要 ・周辺インフラの整備 ・ カ リ ウ ム、 マ グ ネ シ (水路嵩上げ、農道整 ウム等の土壌成分の 備等)が必要 流出がある ・浄化に時間がかかる (5 年前後を想定) 留意事項 ・客土に用いる土は土 壌改良が必要な場合 が多い ・収穫物の処理が必要 ・気象や栽培条件により 効果が変動 ・コスト的には客土 と同等以下の水準 (300 万円 /10a) 18 引用文献 9.農地土壌中における有機ヒ素の動態 2004 年に茨城県神栖町(現,神栖市)の農業用井戸水と収穫された米からジフェニルアルシン 酸(DPAA)等の有機ヒ素化合物が検出された.これらの化合物は自然界に存在せず過去に遺棄 された有機ヒ素系化学兵器に由来すると推測されている.これらにより農耕地土壌や地下水が汚染 された地域での有機ヒ素化合物のヒトへのリスク評価が喫緊の課題となったが,農耕地土壌・農作 物における DPAA 関連有機ヒ素化合物の化学形態変化については,これまで研究の実例がほとん どなかった. 農業環境技術研究所では,この問題に緊急に取り組み,まず分析手法面では,逆相クロマトグラ フィーと誘導結合プラズマ質量分析装置を用いた分析により,水溶性の無機ヒ素である亜ヒ酸,お よびヒ酸から疎水性の高いフェニル置換ヒ素化合物まで一度に定量できる手法を開発した(平成 18 年度研究成果情報,馬場ら) (Baba et al., 2008).その分析手法等を用いて解析を進めた結果, 土壌中の DPAA は,メチル化や脱フェニルによって経時的に化学形態変化を生じ,その変化は畑 条件下に比べて水田条件下の方が顕著であることが明らかとなった(Arao et al., 2009b; Maejima et al., 2011) (平成 22 年度研究成果情報,前島ら).次いで,化学変化にともなう土壌中における 鉛直方向への移動性の差異や土壌タイプとそれら化合物の移動性との関係が明らかになった.一 方,イネにおける DPAA 等有機ヒ素化合物の吸収,玄米への蓄積は畑条件下に比べて水田条件下 の方が顕著であり,また,有機ヒ素化合物のメチル化は主に土壌中で進み,脱フェニルは稲体中で も進むと考えられた.有機ヒ素化合物の応急処置的な対策としては,活性炭の施用や節水栽培に よりメチルフェニルアルシン酸(MPAA)の玄米への蓄積を低減することができた(Arao et al., 2009b) . 図 8 土壌中におけるジフェニルアルシン酸の代謝経路 引用文献 1)阿部 薫,石川 覚,櫻井泰弘,奥山 亮,佐々木和裕,俵田 啓(2006)カドミウム検出用 イムノクロマトキットによる玄米中カドミウム濃度簡易測定の試み.日本土壌肥料学雑誌,77 (6) ,679-682 第 1 章 重金属(カドミウム,ヒ素)汚染リスク評価と汚染土壌修復技術 19 2)Arao, T. and Ae, N.(2003)Genotypic variations in cadmium levels of rice grain. Soil Science and Plant Nutrition, 49,473-479 3)Arao, T., Ae, N., Sugiyama, M. and Takahashi, M.(2003)Genotypic differences in cadmium uptake and distribution in soybeans. Plant and Soil, 251, 247-253 4)Arao, T., Takeda, H. and Nishihara, E.(2008)Reduction of cadmium translocation from roots to shoots in Eggplant(Solanum melongena)by grafting onto Solanum torvum rootstock. Soil Science and Plant Nutrition, 54, 555-559 5)Arao, T., Kawasaki, A., Baba, K., Mori, S. and Matsumoto, S.(2009)Effects of water management on cadmium and arsenic accumulation and dimethylarsinic acid concentrations in Japanese rice. Environmental Science & Technology, 43(24),9361-9367 6)Arao, T., Maejima, Y., and Baba, K.(2009)Uptake of aromatic arsenicals from soil contaminated with diphenylarsinic acid by rice. Environmental Science & Technology, 43, 1097-1101 7)Baba, K., Watanabe, E., Eun H. and Ishizaka, M.(2003)Direct determination of cadmium in rice flour by laser ablation-ICP-MS. Journal of Analytical Atomic Spectrometry, 18, 1485 8)Baba, K., Arao, T., Maejima, Y., Watanabe, E., Eun, H. and Ishizaka, M.(2008)Arsenic speciation in rice and soil containing related compounds of chemical warfare agents. Analytical Chemistry, 80, 5768-5775 9)稲原 誠,雄川洋子,東 英男(2007)生育後期の湛水管理による水稲のカドミウム吸収抑 制.日本土壌肥料科学雑誌,78,149-155 10)Ishikawa, S., Ae, N., Sugiyama, M., Murakami, M. and Arao, T.(2005)Geno-typic variation in shoot cadmium concentration in rice and soybean in soils with different leavels of cadmium concentration. Soil Science & Plant Nutrition, 51, 101-108 11)Ishikawa, S., Ae, N. and Yano, Y.(2005)Chromosomal regions with quantitative trait loci controlling cadmium concentration in brown rice(Oryza sativa). New Phytologist., 168, 345-350 12)Ishikawa, S., Abe, T., Kuramata, M., Yamaguchi, M., Ando, T., Yamamoto, T. and Yano, M.(2010)A major quantitative trait locus for increasing cadmium-specific concentration in rice grain is located on the short arm of chromosome 7. Journal of Experimental Botany, 61, 923-934 13)川崎 晃,織田久男(2005)カドミウム安定同位体(113Cd)トレーサーのダイズ圃場試験へ の適用.日本土壌肥料学雑誌,76,261-267 14)川崎 晃,箭田佐衣子(2007)同位体希釈法による農地土壌中の可給性カドミウム量の評価. 分析化学,52,1081-1087 15)Kawasaki, A. and Yada, S.(2008)Determination of labile cadmium in agricultural soils by isotope dilution plasma mass spectrometry and a coprecipitation separation technique, Journal of Nuclear Science and Technology, Sup., 6, 138-142 16)Maejima, Y., Arao, T. and Baba, K.(2011)Transformation of diphenylarsinic acid in agricultural soils. Journal of Environment Quality, 40, 76-82 17)Makino, T., Takano, H., Kamiya, T., Itou, T., Sekiya, N., Inahara, M. and Sakurai, Y.(2008) Restoration of cadmium-contaminated paddy soils by washing with ferric chloride: Cd 20 引用文献 extraction mechanism and bench-scale verifeication. Chemosphere, 70, 1035-1043 18)Miyadate, H., Adachi, S., Hiraizumi, A., Tezuka, K., Nakazawa, N., Kawamoto, T., Katou, K., Kodama, I., Sakurai, K., Takahashi, H., Satoh, N., Watanabe, A., Fujimura, T. and Akagi, H. (2011) OsHMA3, a P1B-type of ATPase affects root-to-shoot cadmium translocation in rice by mediating efflux into vacuoles. New Phytologist, 189(1),190–199 19)Mori, S., Uraguchi, S., Ishikawa, S. and Arao, T.(2009a)Xylem loading process is a critical factor in determining Cd accumulation in the shoots of Solanum melongena and Solanum torvum. Environmental and Experimental Botany, 67, 127-132 20)Mori, S., Kawasaki, A., Ishikawa, S. and Arao, T.(2009b)A new method for evaluating symplastic cadmium absorption in the roots of Solanum melongena using enriched isotopes 113 Cd and 114 Cd. Soil Science and Plant Nutrition, 55, 294-299 21)Murakami, M., Ae, N., Ishikawa, S., Ibaraki, T. and Ito, M.(2008)Phytoextraction by a highCd-accumulating rice: Reduction of Cd content of soybean seeds. Environmental Science and Technology, 42, 6167-6172 22)Murakami, M., Nakagawa, F., Ae, N., Ito, M. and Arao, T.(2009)Phytoextraction by rice capable of accumulating Cd at high levels: Reduction of Cd content of rice grain. Environmental Science and Technology, 43, 5878-5883 23)Murakami, M., Ae, N. and Ishikawa, S.(2007)Phytoextraction of cadmium by rice(Oryza sativa L.) , soybean(Glycine max[L.]Merr.), and maize(Zea mays L.). Environmental Pollution, 145, 96-103 24)Sugiyama, M., Ae, N. and Arao, T.(2007)Role of roots in differences in seed cadmium concentration among soybean cultivars - proof by grafting experiment. Plant and Soil, 295, 1-11 25)竹田宏行,佐藤 淳,西原英治,荒尾知人(2007)スズメノナスビ(Solanum torvum)を台 木とした接ぎ木栽培によるナス果実中カドミウムの低減技術 . 日本土壌肥料学雑誌,78,581586 26)Ueno, D., Yamaji, N., Kono, I., Huang, C. F., Ando, T., Yanoc, M. and Maa, J. F.(2010)Gene limiting cadmium accumulation in rice. Proceedings of the National Academy of Sciences, 107, 16500-16505 27)Uraguchi, S., Mori, S., Kuramata, M., Kawasaki, A., Arao, T. and Ishikawa, S.(2009)Rootto-shoot Cd translocation via the xylem is the major process determining shoot and grain cadmium accumulation in rice. Journal of Experimental Botany, 60, 2677-2688 28)箭田佐衣子,織田久男,川崎 晃(2004a)水耕栽培ダイズの生育時期別に曝露したカドミウ ムの子実への移行特性.Biomedical Research on Trace Elements, 15, 289-291 29)箭田佐衣子,織田久男,川崎 晃(2004b)耕栽培ダイズの生育時初期に曝露したカドミウム の子実への移行経路.Biomedical Research on Trace Elements, 15, 292-294 30)山口誠之(2006)カドミウム低吸収性,高吸収イネ品種の育成.農林水産技術研究ジャーナ ル,29, 11-14 31)Yamaguchi, N., Mori, S., Baba, K. Yada, S., Arao, T., Kitajima, N. Hokura, A. and Terada, Y.(2011)Cadmium distribution in the root tissues of solanaceous plants with contrasting root-to-shoot Cd translocation efficiencies. Environmental and Experimental Botany, in press 21 第 2 章 POPs,農薬のリスク評価とリスク低減技術 1.はじめに 農薬には,有機・無機,天然・合成,さらに天敵も含めると様々な種類がある.いずれも農作物 を健康に育成する目的で,病害虫・雑草を防除,または農作物の成長を制御する手段である.中で も,1938 年の DDT の発見から始まった有機化学合成農薬(以後“農薬”とする)は,わが国だけ でなく,世界の食料安定供給に多大な貢献をしており,その重要性は今後も変わらないであろう. しかし,初期の農薬の開発目標は,薬害が生じない範囲で充分な薬効を出すことに重点が置かれ ていた.その後,1962 年にレイチェル・カーソン女史の「沈黙の春」が出版されたように,環境 影響に対する意識が高まり始め,これまでに膨大な研究成果や調査データが蓄積されている. 残留性有機汚染物質(Persistent Organic Pollutants, POPs)は,毒性,難分解性,生物蓄積性, 長距離移動性の 4 つの性質を有しており,2004 年 5 月に POPs 条約(ストックホルム条約)が発 効され,12 物質が指定された.2009 年 5 月の第 4 回締約国会議において,さらに 9 種類の物質が 追加され,現在 21 物質が指定されているが,そのうちの約半分が農薬を主たる用途としている. わが国では,POPs の一つであるダイオキシン対策特別措置法等,各種法令の制定や,廃棄物処 理等の各種対策が推進され,ヒトの健康や環境の安全性を確保している.しかし,近年,POPs に 指定されているディルドリンやヘプタクロル類がウリ科野菜やイモ類から検出されている.それ は,これらが,土壌中の消失速度がきわめて遅く,わが国で使用禁止になってから 30 年以上経過 した今でも農耕地に残留していることによる. 農薬は,1948 年に農薬取締法が制定され,その後改訂を繰り返している.現在,農薬を登録・ 販売するには,薬効・薬害だけでなく,毒性や環境中挙動に関して数多くの試験項目をクリアす る必要がある.また,2003 年食品安全委員会が発足し,食品衛生法改正(2006 年施行)におけ るポジティブリスト制度の導入(2006 年)や,作物残留の適正実験規範(GLP, Good Laboratory Practice)の導入(2011 年)など,食の安全に係わる規制も厳しさを増している.さらに,水道法 における規制強化によって,飲料水の安全性も確保されている. しかし,このように規制を厳しくした結果,無登録農薬問題は終息したものの,輸入農産物やヤ マトシジミ等の魚介類に基準値を超える農薬が検出され,輸入禁止や出荷の自粛,農薬登録の見直 しなどの対応を余儀なくされた.さらに農薬散布時のドリフトによる周辺作物への残留や,前作に 処理した農薬の後作物への残留も懸念されている. その他には, 「農地・水・環境保全向上対策事業」など,近年環境保全型農業の推進が図られた 結果,農薬の過剰使用は抑制されたものの,環境保全への効果を科学的な根拠に基づいて評価して いるとは言い難い.また,中央環境審議会・水環境部会・環境基準健康項目専門委員会における要 監視項目の設定において,農薬は工場や家庭からの排水と異なり,季節性のある面源汚染であるた め,監視方法を適切に示すことが難しく,懸案事項のままとなっている. 農薬はその使用目的や用途から必然的に環境中に拡散する.その環境は,農耕地から周辺の自然 や住宅,土壌・水(河川・湖沼等) ・大気,気象・地形・地域等,様々にある複合系(混合系)で ある.また,農薬や POPs は,それぞれに様々な物理・化学・生物学的性質を持ち,その代謝物も 異なる性質を有する.そこに時間と空間の要因を絡めると,農薬の挙動とそこにおけるヒト健康や 環境に対する影響を科学的に裏付けるのはきわめて難しい.ここでは,農業環境技術研究所で実施 22 2.POPs の動態および作物残留低減技術 された農薬と POPs のリスク評価およびリスク管理に関する最近 10 年間の研究成果を紹介する. 2.POPs の動態および作物残留低減技術 ⑴ 分析法の開発 ダイオキシン類は,ポリ塩化ジベンゾパラジオキシン(PCDD)とポリ塩化ジベンゾフラン (PCDF)の総称であり,両者を合わせて 135 種類ある.その分子種間で人畜毒性の違いが 10 万倍 あるため,個々のダイオキシンの毒性を,毒性が最も強い 2,3,7,8- テトラクロロジベンゾ -1,4- ジオ キシン(2,3,7,8-TCDD)に置き換えた毒性等量(Toxicity Equivalency Quantity, TEQ)で示す. わが国では,ダイオキシン類の耐容一日摂取量(TDI)を 4 p(10−12)g-TEQ と設定している. 一方,環境省の調査(2001 年)によれば,ダイオキシン類は,残留量が最も高い底質や土壌で も平均濃度で 6 〜 8pg-TEQ/g と,環境中でごくわずかにしか存在しないため,超微量多成分一斉 分析が求められる.そのため環境省では,大気・底質・土壌における分析法マニュアルを公開して いる.ダイオキシンはその由来によって分子種の組成が異なることから,主成分分析,クラスター 解析,レセプターモデル等を駆使して解析することで,分析結果から由来を推定することができる (清家ら,2007) . 一方,ディルドリンやヘプタクロル類などのわが国で農薬として過去に使用されていた POPs の 使用履歴に関する農耕地ごとの情報は皆無であり,POPs の基準値を超えた作物残留を未然に防ぐ には,農耕地土壌の汚染の分布を詳細に知ることが求められる.それに対応する分析法として,メ タノールと水の容積比を 50%にして抽出した場合の土壌中ディルドリン濃度によって,土壌が異 なっていてもキュウリ植物体中ディルドリン残留濃度を予測できることを示した(平成 20 年度研 究成果情報,清家ら) (図 1) . 図 1 異なる土壌における 50%メタノール・水で抽出したディルドリン濃度とキュウリ茎葉中濃度との相関 第 2 章 POPs,農薬のリスク評価とリスク低減技術 23 また,河川などの水系における POPs,農薬(および代謝物),医薬品,ホルモン類などの人工 合成有機化学物質に対して多成分一斉分析方法を開発した(平成 21 年度研究成果情報 b,殷ら) (WARC et al., 2008; WARC et al., 2009) . ⑵ ダイオキシン類 1 作物および環境中動態の解析 水田における,代かき,水稲移植期の落水,降雨時等によって,ダイオキシン類は懸濁物質 (SS)に吸着して河川に流出し湖沼・海洋まで達するとともに,食物連鎖を介して水生生物等に影 響を与える危険性がある(平成 13 年度研究成果情報,芝野ら).そこで,連続遠心分離法を改良し て土壌を粒径別に迅速・大量に分取して調べると,SS における粒径別のダイオキシン類濃度は, 灰色低地土や黄色土では粒径が小さくなると低下し,黒ボク土では逆に高くなった(平成 14 年度 研究成果情報,牧野ら) .また,牛久沼流域では,1,3,6,8-TCDD と 2,3,7,8-TCDD の濃度比に基づい て推定した底質におけるダイオキシン類の TEQ への土地利用別寄与率は,林地,水田および畑・ 市街地がそれぞれ,2,32 および 66%であった(平成 14 年度研究成果情報,谷山ら). 一方,ダイオキシン類は,廃棄物の燃焼・焼却による生成や,農薬に含まれる不純物としての農 耕地への投入等,様々な要因で農業生態系に負荷され,農耕地土壌中に蓄積する.水稲で検出され るダイオキシン類の異性体の多くは大気中のコプラナー PCB であり,維管束液中からは検出され ないことから,土壌からの吸収ではなく,大気が水稲におけるダイオキシンの主な汚染源であるこ とを明らかにした(平成 14 年度研究成果情報,桑原ら)(図 2). ⨍䋺0.045pg-TEQ/g ⪲䋺4.1pg-TEQ/g ☭Ზ䋺0.38pg-TEQ/g ⛽▤᧤ᶧ <0.0001pg-TEQ/g ₵☨䋺0.0011pg-TEQ/g ფ䋺120pg-TEQ/g 図 2 水稲の各部位におけるダイオキシン類の濃度 水田土壌中のダイオキシン類濃度は,1960 年代から急激に上昇し,1970 年前後をピークに現在 まで緩やかに減少した.ダイオキシン類の起源については,1960 年以前は燃焼・焼却過程からの 寄与が主なものであり,1960 〜 1970 年代に入って PCP 製剤と CNP 製剤の不純物が主な要因とな り,近年は再び燃焼・焼却過程からの寄与が増加していることを示した(平成 15 年度研究成果情 報,清家ら) .また,水田土壌におけるダイオキシン類の年間負荷量は 5.3n(10−9)g-TEQ/m2,消 失量は 686ng-TEQ/m2 と試算され,その増減は現在の残留量 24,600ng-TEQ/m2 と比較するとごく わずかであるため,今後もきわめて緩慢に減少していくものと予想された(平成 16 年度研究成果 情報,上垣ら) .このことは,その他の POPs についても同様であり,各地農耕地土壌の分析結果 24 2.POPs の動態および作物残留低減技術 から,当初の減衰は早いものの,その後はきわめて緩慢に減少するホッケースティック型減衰曲線 を示した. 2 リスク低減技術の開発 野菜類におけるダイオキシン類の残留は,土壌粒子と大気粉じんの付着が主原因であり,地上 部(葉菜や果菜)はマルチ被覆やトンネル栽培,根部(根菜)は皮層の除去によって汚染が軽減さ れた(平成 13 年度研究成果情報,殷ら) .また,紫外線領域を太陽光に近似させたソーラーシミュ レーターを用いて,土壌表層中のダイオキシン類の光分解反応実験装置を試作し,粘土鉱物に添 加した八塩素化ダイオキシン(OCDD)に光照射すると,OCDD の脱塩素化によって TEQ が増加 するものの,二酸化チタンを添加することで分解が促進された(平成 14 年度研究成果情報,小原 ら) .土壌中のダイオキシン類は,酸化カルシウムを基材とする化学資材を混和すると,ラジカル 反応によって分解された(平成 14 年度研究成果情報,清家ら). ダイオキシン類の水田から河川への流出抑制対策としては,節水または無代かきが有効であった (平成 13 年度研究成果情報,芝野) .代かきをする場合は,凝集剤として塩化カルシウムまたは塩 化カリウムを施用すると,田面水の SS を速やかに沈降させ,SS に吸着しているダイオキシン類の 水田系外への流出を大幅に軽減できた(平成 15 年度研究成果情報,牧野ら)(図 3). ήᣉ↪ Ⴎൻ䍔䍷䍚䍑䍯䋨15䌫䌧/10a) Ⴎൻ䍔䍷䍚䍑䍯䋨30䌫䌧/10a) Ⴎൻ䍔䊥䍑䍯䋨15䌫䌧/10a) Ⴎൻ䍔䊥䍑䍯䋨30䌫䌧/10a) SS㊂(g/L) 16 12 8 4 0 1ᤨ㑆 3ᤨ㑆 ઍហ䈐ᓟ䈱⚻ㆊᤨ㑆 24ᤨ㑆 図 3 塩化カリウムおよび塩化カルシウムの懸濁物質(SS)流出抑制効果 ⑶ ドリン類 1 作物および環境中動態の解析 ドリン類およびヘプタクロル類は,ウリ科作物のみが特異的に吸収して,地上部へ移行させる (Otani et al., 2007) .これらが導管流に乗って地上部に移行する際,何らかのタンパク質がトラン スポーターとして関与すると考えられている(Murano et al., 2010). 果菜類であるキュウリ果実中ディルドリンや,カボチャ果実中ヘプタクロル濃度分布は,乾重あ たりに換算すると濃度差はみられなかった.根菜類であるニンジンおよびジャガイモにおけるディ ルドリンの残留は,ダイオキシン類同様土壌の付着と考えられた.また,ディルドリンが残留した キュウリを塩化ナトリウム水溶液に浸漬(漬物を想定)した場合と,カボチャ果実を煮沸(煮物を 想定)した場合,それぞれ約 5%と 1%しか溶出せず,加工調理工程では,ウリ科作物におけるド リン類の汚染低減対策を立てることは困難であった. 第 2 章 POPs,農薬のリスク評価とリスク低減技術 25 また,各種作物の可食部へのディルドリンの移行率を各作物の残留基準値(MRL)で除して, 基準超過指標値を求めることにより,ディルドリン汚染圃場における栽培適性を評価した(平成 22 年度研究成果情報,清家ら) . 一方,ドリン類およびヘプタクロル類のわが国の農耕地土壌における残留程度を俯瞰的に把握 することは重要である.そこで,これらの土壌中消長,作物体への吸収・移行特性,過去の出荷 状況,土壌特性(全炭素含有率および仮比重) ,気象条件等の要因に基づいて,モンテカルロ法に よって,土壌残留や作物残留の経年変化を推定する手法を開発した. 2 リスク低減技術の開発 POPs で汚染された農耕地土壌におけるリスク低減対策は,汚染程度に応じて講じる.具体的に は,農耕地として利用しない,作土層を客土する,POPs を吸収しにくい作物に切り替える,土壌 を浄化する,POPs の吸収を抑制する等の対応が考えられる. まず,ドリン類の吸収を抑制する技術として,接ぎ木と活性炭の土壌混和がある.接ぎ木につい ては,キュウリのディルドリン吸収性は,低吸収性の台木カボチャ品種を選ぶことで,穂木キュウ リの果実中濃度を 30 〜 50%程度低減することができる(平成 18 年度研究成果情報,大谷ら)(図 4) .活性炭は,木炭や木材チップに比べてディルドリンの吸着資材としてきわめてすぐれているも のの,農耕地土壌における汚染程度や土壌の種類によって投入する活性炭の最適量は大幅に異な る.しかし,この量は汚染土壌から 50%メタノールで抽出したディルドリン濃度から推察できる. 図 4 キュウリ果実中のディルドリン濃度に及ぼす台木と穂木の影響 次に,汚染土壌の浄化技術である.これには,物理的,化学的,生物的技術に分けることができ るが,ここでは物理的浄化技術については割愛する. 化学的浄化技術には分解と溶出がある.分解については,鉄資材(FeB, MT-V3)を,ディルド リンを含む水溶液や汚染土壌に添加することで分解できることが阻害要因とともに明らかにされて いる.溶出については,汚染土壌からの溶出力は,アセトン,エタノール,ジエチルエーテル,プ ロパノール,メタノールの順で溶出力が高かった.界面活性剤にも溶出力があるが,環境に対す る影響が少ない生分解性の剤では,ソホロリピッド,ラムノリピッド,ペグノール ST の順であっ た. 生物的浄化技術は,植物を利用したファイトレメディエーションと,微生物を利用したバイオ 26 3.農薬の環境影響評価 レメディエーションがある.ドリン類で汚染された土壌のファイトレメディエーションに最適な 植物は,ディルドリンを高吸収するズッキーニである.この植物の土壌浄化効果は,室内試験で キュウリの果実中濃度を 40 〜 60%低減できた(平成 20 年度研究成果情報 a,大谷ら).カボチャ へのヘプタクロル類残留対策にも,低吸収性品種や活性炭の土壌混和による吸収抑制など,キュウ リのディルドリン対策で用いたリスク低減技術が適用できる(平成 20 年度研究成果情報 b,大谷 ら) .次に,微生物を利用したバイオレメディエーションについては,エンドスルファン連用畑土 壌から,ディルドリンを炭素源とする土壌・木炭還流法により,好気的に分解する糸状菌(Mucor racemosus)を単離した(平成 21 年度研究成果情報 a,高木ら)(図 5).同様に,ペンタクロロニ トロベンゼン(PCNB)を炭素・窒素源として単離した細菌 PD653 株(Nocardioides 属)は POPs である HCB を好気的に PCP(ペンタクロロフェノール)に変換し,さらに脱塩素分解を経て無機 䊂䉞䊦 䊄䊥䊮 Ớᐲ (mg/kgੇ) 化した(平成 20 年度研究成果情報,高木ら) . 6 5 䈸䈜䉁 (ფ᳓ಽ40䋦) 4 䈸䈜䉁 (ფ᳓ಽ50䋦) 3 DDF+䈸䈜䉁 (ფ᳓ಽ40䋦) 2 1 DDF+䈸䈜䉁 (ფ᳓ಽ50䋦) 0 0 5 10 15 20 25 ၭ㙃ᣣᢙ 図 5 糸状菌によるディルドリンの分解 3.農薬の環境影響評価 ⑴ 分析法の開発 農薬の分析は,対象となる化学物質,試料,定量限界値,担当者,分析に要する時間などによっ て選択する.国民の食の安全性に対する関心の高まりとともに,農作物や加工食品中の残留農薬 の把握がますます重要な課題となっており,収穫から出荷までの流通を円滑に行うため,短時間 に農薬の残留量を把握することが要求され,機器分析についても高度化が図られている(石井, 2004) . 抗原抗体反応を利用したイムノアッセイは,ガスクロマトグラフ(GC)や高速液体クロマトグ ラフ(HPLC)のような機器分析に比べて,短時間かつ簡便な測定手順で分析ができ,導入や運用 コストが安価であるため,生産や流通などより現場に近い場面で,スクリーニングを目的として多 量の試料を扱うのに適しており,農薬分野では現在数 10 種類のキットが市販されている. しかし,イムノアッセイにおける抽出・精製操作は,一般に簡便である反面,夾雑物を完全に除 去しないため,それらの抗原-抗体反応への影響評価,機器分析とのバリデーション,擬陽性・擬 第 2 章 POPs,農薬のリスク評価とリスク低減技術 27 陰性の確認とともに,実際に運用するには,農薬の抽出・精製という前処理と一体化したマニュア ルが必要である.一例として,殺虫剤イミダクロプリドのイムノアッセイキットを用いて,イムノ アッセイに分析段階で共存できるメタノールを利用した簡易抽出法が開発された(平成 16 年度研 究成果情報,渡邉ら) . ⑵ 環境中動態解析 農耕地に処理された農薬は,土壌の固相への吸着とそこからの脱着を繰り返しつつ,水ととも に移動する.その吸着様式には,疎水結合,水素結合,共有結合等,10 種類程度想定され,土壌 と農薬の性質によって異なると考えられる.たとえば除草剤 2,4- ジクロロフェノキシ酢酸(2,4-D) と火山灰土壌の組み合わせでは,その構成成分であるアロフェン等の鉱物や鉄・アルミニウム ― 腐植複合体に,配位子(リガンド)交換反応によって強く吸着される(平成 13 年度研究成果情 報,平舘ら) . 農薬の水田環境中挙動モデル(Watanabe et al., 2007)については,いくつか開発されている が,わが国では,水田 ― 河川環境における農薬の挙動予測モデル(PADDY(図 6)や PADDYLarge(稲生,2004) ) ,PCPF-1(Watanabe et al., 2000)がある.農薬の代謝物については,水稲 用除草剤ピリミノバックメチルを対象に水田における光異性化および代謝分解を考慮した挙動予測 が行えるように PADDY モデルが改良されている(Inao et al., 2009).水管理については,田面水 余剰貯留水深(EWSD)の概念(Phong et al., 2008)や,農薬処理後の止水期間の延長(石井ら, 2004)が農薬の水田からの流出抑制に有効である. さらに,地理情報システム(GIS)によって河川流域特性を解析するとともに,流域内での農薬 の使用量や使用時期を反映させることで,農薬の河川中濃度の季節変化を予測できる GIS 結合型 PADDY - Large モデルを開発した(平成 22 年度研究成果情報,稲生ら). 図 6 水田における水の動きと農薬の挙動 28 3.農薬の環境影響評価 ⑶ 環境毒性評価法の開発 農薬の環境毒性は,生態系に生息する生物に対する影響であり,OECD テストガイドラインにお いて様々な試験法が提示されている.わが国の農薬取締法においても,平成 17 年度に改正された 「水産動植物の被害防止に係る農薬登録保留基準(以下水産登録保留基準)」に基づいて,農薬の水 生生物に対するリスク評価を行い,魚類(メダカまたはコイ),甲殻類(オオミジンコ),藻類(緑 藻:Pseudokirchneriella subcapitata) ,いわゆる「3 点セット」への影響について調べることになっ ている.これらに対して,毒性評価として,急性影響濃度(Acute Effect Concentration,AEC) と,段階的評価による河川における環境中予測濃度(Predicted Environmental Concentration, PEC)との比較から登録の可否を判定する. わが国では,農耕地の約半分が水田であり,水田は河川と直結しているため,水田農薬の河川へ の流出が最も懸念される.わが国の河川においては,生産者や第一次消費者として,それぞれ付着 性珪藻・藍藻やトビケラ類が優占しており,OECD テストガイドラインで推奨されている緑藻類 やミジンコ類は,ほとんど分布していないため,上記 3 点セットだけでわが国の河川生態系に生息 する生物に及ぼす影響を正確に評価できない. わが国に生息する藻類について,OECD テストガイドラインに則って 14 種類の水田除草剤に対 する薬剤感受性を調べてみると,わが国の付着性珪藻や藍藻の薬剤感受性は,緑藻類とは大きく異 なっていた(平成 15 年度研究成果情報,石原ら)(図 7). 1000 : amide compound : triazine compound : sulfonylurea compound : thiocarbamate compound : miscellaneous compound 100 & 10 : EC50 values are higher than their concentration bentazone daimuron dimepiperate molinate imazosulfuron Herbicides quinoclamine esprocarb thiobencarb mefenacet Bensulfuronmethyl simetryn dimethametryn 0.1 cafenstrole 1 pretilachlor : P. subcapitata : A. minutissima 72-h EC50 values (ug/L) : C. Vulgaris : M. tenuissima 10000 図 7 除草剤に対する感受性の藻類種間差 実際の河川では,単一の藻類ではなく,様々な藻類が群集を形成している.しかし,群集内の藻 類をすべて評価するには,それぞれ単独で培養して毒性試験を実施する必要があり,多大な労力と 長い期間を要する.そこで,河川や水田で生息している珪藻群集をそのまま利用することで,地域 性を考慮したより現場に近い藻類への影響評価を開発した(平成 17 年度研究成果情報,石原ら). 他の植物では,水稲用スルホニルウレア系除草剤に対して高感受性のジュンサイやスイタグワ イの展開葉が奇形する症状は,除草剤の系外流出の生物指標として利用できる(平成 13 年度研究 第 2 章 POPs,農薬のリスク評価とリスク低減技術 29 成果情報,伊藤ら) .絶滅危惧種である浮遊シダ植物のサンショウモも,これらと同様に,スルホ ニルウレア系除草剤に対して緑藻類に比べて高感受性を示した(平成 17 年度研究成果情報,池田 ら) . 次に昆虫類についてみると,広くわが国の河川に高密度で分布するコガタシマトビケラは,農薬 の毒性試験のための生物種として最適であるが,試験生物種として利用するためには,標準系統を 恒常的に提供する必要がある.しかし,この昆虫は湖沼等の止水域に生息するミジンコ類と異な り,流水域に生息し,ライフサイクルも長いため,室内で飼育することはきわめて難しい.そこ で,全国から採集したコガタシマトビケラの殺虫剤感受性を比較して,最も感受性が高いグループ から標準系統を選定するとともに,室内人工飼育法を確立した(平成 16 年度研究成果情報,大津 ら) (図 8) (横山ら,2008) . Breeding aquaria (Large glass petri dish) in the 3rd- to 5th-instar stage Larger pebbles are added in the breeding aquarium. Sufficient aeration and water current supply oxygen to the larva efficiently. The larva grow well by feeding algae and a commercial fish food. Emergence Cage Emerged adults are transferred to a custommade aquarium for oviposition. Adult Larvae in the1st- to 2nd-instar stage Pupae After hatching, 1st-instar larva build nests with algae and small glass beads quickly. A copulation The stirrer creates a water current. Hatching Rocks or bricks Egg masses Egg masses on substrates such as rock or brick Eggs are incubated in a large glass petri dish with sufficient aeration. Custom-made aquarium for oviposition Matured females swim to bottom rocks, where they lay eggs in flattened clusters. 図 8 水生昆虫の人工飼育法(コガタシマトビケラ) 魚類については,農薬等化学物質の内分泌かく乱作用,いわゆる環境ホルモンとして疑われてい ることに対応して,女性ホルモン様作用に着目して,ふ化したばかりのメダカ(淡水性魚類)を化 学物質に曝露し,オスからメスへの性転換を短期間(2 か月以内)に検出できる試験系を開発した (平成 14 年度研究成果情報,堀尾) . ⑷ 環境リスク評価法 環境中に放出された農薬の生態系影響の評価は,毒性(有害性)評価と曝露評価の組み合わせに よって行うが,リスク評価には決定論的方法と確率論的方法がある. 決定論的リスク評価では AEC と PEC を単純に比較するが,水稲用除草剤の場合,河川水中濃 度は,わが国の河川に生息する付着性藻類も含めて,半数影響濃度(EC50)より低いため,水田 30 4.農薬のリスク低減技術の開発 から流出した除草剤の河川における藻類への影響は小さい(石原,2008).水稲用除草剤の分解物 についても,河川水中濃度が親化合物よりも高まることがあるものの,その濃度は藻類の EC50 よ りも低く,親化合物と同様に藻類への影響は小さいと推測される(平成 20 年度研究成果情報,岩 船ら) .また,殺虫剤やその分解物については,トビケラの 1 齢幼虫を用いた農薬の急性毒性試験 法を開発し(平成 19 年度研究成果情報,横山ら)(横山ら,2008),水生節足動物(ミジンコやト ビケラ)への影響を評価した.いくつかの殺虫剤で分解物の水中最高濃度が殺虫剤そのものよりも 高いものの,親化合物や分解物の河川水中最高濃度は水生節足動物の EC50 よりも明らかに低く, 水生節足動物への影響は小さいことを示した(平成 21 年度研究成果情報,横山ら). 一方,確率論的評価では,水生生物に対する農薬の毒性の生物種ごとのバラツキや,農薬の河川 水中濃度の地域的なバラツキを解析することで,農薬による環境リスクの大きさを計算する(平成 21 年度研究成果情報,永井ら) .その方法を使ってシメトリンで解析したところ,5%の種が影響 を受ける確率は 1.5%と計算され,曝露を受ける確率と組み合わせたリスクカーブから,水生生物 に対する期待多様性影響は全国平均で 0.3%と試算された(永井ら,2008). 4.農薬のリスク低減技術の開発 ⑴ 系外流出抑制および水質浄化技術 農薬は,土壌,水系,大気および生物の間を,様々に代謝を受けながら複雑に挙動する. 水田から河川への農薬の流出を抑制するために,先に述べた PCPF-1 モデルを活用して,水稲用 除草剤の吸着材としてのモミガラ成形炭について,粒子径,炭化温度,施用量を最適化した農薬の 水田系外流出削減技術を開発した(平成 14 年度研究成果情報 a,高木ら).一方,地球温暖化や化 石燃料の高騰をきっかけに,バイオマスエネルギーとして利用されるもみがら燃料のガス化発電の 残渣として大量に排出される炭化物の処理が大きな課題となっている.そこで,水田農業における ゼロエミッションをめざして,もみがらガス化残渣を水稲用農薬の系外流出防止技術に組み込むこ とで,CO2 削減などに寄与する米生産システムを開発した(平成 19 年度研究成果情報,高木ら). 一方,水の汚染問題が近年世界各地で深刻化し,良質な水の確保が重要な課題となっている.農 薬,POPs,工業廃物等の有機合成化合物は,河川や湖沼だけでなく,地下水を含む幅広い水系を 汚染する危険性がある.そこで,それらを強力に吸着する微粉末活性炭をタブレット(錠剤)化 し,簡単・迅速・安全に水を浄化する技術を開発した(平成 21 年度研究成果情報 a,殷ら).ま た,活性炭シートは,POPs 等によって汚染された農耕地土壌への適用が期待されている. ⑵ 微生物による分解 農薬の土壌中減衰は,先に述べた POPs 同様,ホッケースティック型減衰曲線を描き,後半の緩 やかな減衰は,土壌微生物による分解が主な要因と考えられている. 近年,農耕地土壌等で広範囲に低濃度で分布している難分解性有機汚染物質の分解除去技術とし てバイオレメディエーションが注目されている.しかし,微生物による農薬の分解については,多 くの研究事例があるものの,現場では複数の有機化合物で汚染されていることが多く,これらの 汚染物質を同時に原位置で完全分解する技術は開発されていない.土壌環流法によって土壌から 分離したシマジン分解細菌群(CD7)は,3 種類の細菌 Arthrobacter sp.(CD7w),Bradyrhizobium japonicum(CSB1) ,新種のβ-Proteobacteria(CDB21)で構成され,CDB21 のみがシマジン分解活 性を有している(平成 16 年度研究成果情報,高木ら).シメトリン分解細菌とシマジン分解細菌な どを組み合わせた複合微生物系を木質炭化素材に集積させることで,この系統の農薬を同時に分 解・無機化した(平成 17 年度研究成果情報,高木ら)(図 9).CD7 菌群を集積させた木質炭化素 第 2 章 POPs,農薬のリスク評価とリスク低減技術 31 材を,ゴルフ場を想定した芝生の下層に敷き詰めて,シマジンの炭化素材への吸着と分解を実証し た(平成 14 年度研究成果情報 b,高木ら) .樹脂,塗料の原料として利用されるメラミンの廃棄物 を再生利用するため,人体にとって有害なメラミンや分解物であるシアヌル酸まで分解する細菌 (Nocardioides 属の新種)を土壌中から単離した(平成 21 年度研究成果情報 b,高木ら). a. CDB21ᩣ b. Fjᩣ O Cl R1 N N H O N N N H R2 R1 䉪䊨䊨䊃䊥䉝䉳䊮 N N N H N N H O SCH3 R2 䉴䊦䊖䊮 R1 N R1 N H N H R2 䉴䊦䊖䉨䉲䊄 R1 N N H N N N H R2 䊜䉼䊦䉼䉥䊃䊥䉝䉳䊮 OH N N SCH3 N N N H OH N SCH3 N H 2-䊍䊄䊨䉨䉲 R2 N HO N N OH ੑ㉄ൻ⚛ ᳓ 䉝䊮䊝䊆䉝 䉲䉝䊇䊷䊦㉄ 図 9 複合微生物系によるトリアジン系化合物の分解様式 5.農薬や POPs の大気中挙動 散布された農薬は,直接,またはいったん作物や土壌に落下したものが,光分解を受けながら大 気中を拡散するが,土壌中減衰や河川中挙動に比べて大気の流動性はきわめて高いため,その中に 混在する農薬の挙動を把握することはきわめて難しい.大気中挙動については,現在も使用されて いる農薬は分解性も高いため,主にヒトが活動する範囲の挙動,POPs は環境中の残留性と長距離 移動性が高いため,主に地球規模の挙動を把握する必要がある. ⑴ モニタリングと動態モデル 散布された農薬は,ほとんどがその場所にとどまり,標的となる病害虫・雑草,作物または土壌 に落ちるが,一部は空中を漂流・揮散する.これをドリフトという.ドリフトにはスプレードリフ トとベーパードリフトの 2 種類ある(與語,2006).前者は散布時のドリフトのことであり,後者 は一度作物や土壌などに吸着したものが,徐々に大気中に移行・拡散するものである. スプレードリフトについては,欧米において,農薬の大気中挙動に関する各種シミュレーション モデルがあるが,純水を想定したものであり,農薬の製剤や希釈倍率等は考慮されていない.一 方,わが国では,農薬処理後の大気中拡散のモニタリングに関する長年の研究実績を蓄積するにと どまっている.スプレーノズルによって生成する噴霧粒子(一次液滴)のうち,ドリフトへの寄与 が大きいと考えられる微小粒子(< 30μm)生成への影響を調べるための試験装置を作製して調べ たところ,農薬製剤に含まれるまたは散布時に添加する界面活性剤の種類によって,微小粒子の分 布が異なることを示した. ベーパードリフトについては,農薬を降雨によって洗い流すことで回収して分析する手法と,大 32 5.農薬や POPs の大気中挙動 気を直接回収する方法がある.後者には拡散原理を利用したパッシブサンプラーと吸引ポンプを利 用したアクティブサンプラーがあり,いずれも吸着材で農薬を捕集する.前者は電源が不要なた め,農薬や POPs の広域分布における追跡に,後者は限られた場所における,より精緻な解析に適 している.パッシブサンプラーによって,東アジアの各地域における大気中の農薬・POPs をモニ タリングし,バックトラジェクトリー(後方流跡線)解析したところ,一部の化学物質はアジア大 陸からわが国まで移動していることが示された(小原,2010).一方,アクティブサンプラーによっ て,除草剤の大気移行量は,ガス状および粒子状ともに急激に減少すること,両者の比は薬剤に よって大きく異なるものの,土壌や季節の影響を受けないことを示した(與語,2006). 農薬や POPs を地球規模で把握するために,欧米ではフガシティー(逃散能,逸散度)という圧 力で表された化学ポテンシャルの概念によるシミュレーションモデルが開発されている.しかし, 欧米のモデルは,畑作農業を念頭に置いたものであり,アジアにおける主要な水田農業やそこで 使われる農薬には対応していない.そこで,農薬や POPs の水田農業を組み込んだ地球規模の挙 動を予測するシミュレーションモデル(MIAES-MMM)を新たに開発した(平成 19 年度研究成果 情報,西森ら) (図 10) .このモデルは,地球を複数の方格(グリッド)に分け,その中を大気, 海洋水,水田等を含む 9 つのコンパートメントに分けて,その間の移動をフガシティーの概念で 結んだ非定常モデルである.このモデルによって,1960 〜 1970 年代にわが国の水田で使用された POPs の一部が北極にまで到達していることを明らかにした(西森ら,2010). 図 10 農薬の地球規模の挙動予測のためのフガシティーモデル(NIAES-MMM) 第 2 章 POPs,農薬のリスク評価とリスク低減技術 33 ⑵ 臭化メチル 土壌くん蒸剤として農耕地で使用されている臭化メチルは,オゾン層を破壊する物質として,モ ントリオール議定書締約国会議において,2005 年に不可欠用途を除く全廃,2013 年にはそれも含 めた全廃が決定されている.わが国では,これまで園芸農業を中心に臭化メチルに大きく依存して 病害虫・雑草を防除してきたため,臭化メチルの代替技術の早期開発が強く望まれているものの, 現時点で,臭化メチルに完全に置き換わることができる技術は開発されていない. 実際,慣行の農業用ポリエチレンフィルムを用いた場合,臭化メチルの大気放出量は施用量の 63.8%であったが,遮光資材の併用で 56.2%,ガスバリアー性被覆資材で 24.1%,光触媒含有積層 被覆資材で 1%以下まで削減できた(平成 13 年度研究成果情報,小原ら). 臭化メチルの代替技術として,低濃度エタノールを利用した土壌還元消毒がある.本技術は, エタノールを水で 2%以下に薄めて畑土壌が湛水状態になるまで灌水した後,農業用ポリエチレン フィルムで土壌表面を覆い,1 週間以上放置するという簡便なものであり,環境への負荷も少ない と考えられる(小原ら 2007) (図 11) .本技術には農薬のような殺菌・殺虫・除草の直接的効果は ないものの,幅広い範囲の土壌病害虫や雑草に対する防除効果が確認されている.その作用機構 は,フスマなどと同様に,還元(嫌気的)土壌環境への変化や有機酸濃度の増加等が想定されてい る. ಣℂ㧔ࠛ࠲ࡁ࡞㧝㧑ṁᶧ㧕 ήಣℂ 図 11 低濃度エタノールの効果の一例(キュウリ根のネコブセンチュウ) 6.薬剤抵抗性と病害抵抗性誘導 ⑴ 薬剤抵抗性 農薬は病害虫・雑草に対して大きなストレスを与える.病害虫や雑草には様々なバイオタイプが あり,それらが混在(交雑・交配などを含む)して個体群を形成している.そのため,農薬の連用 によってその影響を受けにくいバイオタイプが生存しやすくなり,農薬に対する感受性を基軸とし たシフトが個体群に生じる.それが薬剤抵抗性の発現である. 薬剤抵抗性の検出方法は In vivo から In vitro まで様々にある.遺伝子診断による方法の一例とし て,イネいもち病菌のシタロン脱水酵素阻害剤(MBI-D 剤)耐性の場合,当該酵素遺伝子の 1 塩 34 7.今後の展望 基変異を PCR-Luminex 法で解析することで迅速かつ多検体処理が可能となる(平成 13 年度研究 成果情報 2003b,石井ら) .また,ワタアブラムシの有機リン剤抵抗性の原因となる解毒酵素(カ ルボキシルエステラーゼ)遺伝子の塩基配列の系統間の違いから抵抗性系統と感受性系統を明確に 識別できる(平成 14 年度研究成果情報,鈴木) . 雑草の除草剤抵抗性も一般に 1 塩基変異によって発現するとともに,感受性遺伝子に対して優性 を示す.たとえば,イヌホタルイのスルホニルウレア系除草剤抵抗性遺伝子は,他殖率および感受 性の埋土種子集団の存在にかかわらず,この除草剤が持つ高い淘汰圧によって集団中に急速に広ま る可能性がある(平成 15 年度研究成果情報,池田ら). ⑵ 病害抵抗性誘導 植物は生育する環境から動くことができないため,各種環境ストレスに対して応答して耐性を獲 得する機構を有している.最も知られているのが気孔の開閉であるが,農薬を含む薬剤や病害虫に 対しても応答する.その際,特定の農薬や病害虫だけに耐性を有するよりも,同様のストレスに対 して耐性を獲得することが多く,それを利用した技術がある.雑草では除草剤耐性遺伝子組換え作 物や解毒剤(セーフナー) ,害虫では Bt タンパク質産生遺伝子組換え作物,病原菌では病害抵抗 性誘導剤がある.病害抵抗性誘導剤は植物が本来持つ生体防御機能を活性化して,各種病害に対す る複合抵抗性を全身に発現させるが,その分子機構も解明されつつある. アシベンゾラル S メチルが黒星病抵抗性を誘導する仕組みは,抵抗性が誘導される過程で,ポリ ガラクツロナーゼ阻害タンパク質遺伝子の転写量が増大するとともに(平成 15 年度研究成果情報 a,石井) ,他の殺菌剤と異なり,作物の葉面糸状菌相に影響しない(平成 16 年度研究成果情報, 石井) . 7.今後の展望 農業環境を考える場合,農業が環境に与える影響と環境が農業に与える影響の二つの視点があ る.農業を農薬および POPs に置き換えると,環境への影響については生物が曝露することによる 影響である.一方,環境が農薬および POPs に与える影響については,その残留を含む環境中動態 への影響であり,温度による分解や降雨や風による吸着体である土壌の移動に伴う移動が考えられ る. 当面は,農薬や POPs が環境に与える影響が研究課題であろうが,中でも重点化すべきことはリ スク分析である.この要素としてリスク評価とリスク管理があるが,一つの危害要因に対してリス 分析を講じる場合,今後様々な視点を念頭に置いて研究を推進することが求められる. 第一にリスクトレードオフである.ここには,農薬を使用しない,または POPs で汚染された農 耕地に何も講じないことによるリスクと,農薬以外の代替技術,または POPs で汚染された農耕地 に何らかのリスク低減技術を講じることによるリスクがある.これまで開発した技術を実用化する ためには,それらが持つ技術的マイナス面(デメリット)も考慮すべきである. 第二にリスク ― ベネフィットやコスト ― ベネフィットの概念である.農薬を使うベネフィット は,その用途である病害虫・雑草防除が効率的かつ確実に実施することであり,結果として安定し た収量と品質を確保することができる.遺伝子組換え作物では,除草剤耐性作物導入による土壌侵 食防止や,Bt タンパク質産生作物による農薬処理作業の軽減・低減も挙げられる.一方,農薬や POPs のリスク低減対策を講じるためのコストがベネフィットに見合うものかも見きわめなければ ならない.そのためにも,これらを主眼に置いた作物栽培(農耕地)における農薬や POPs のライ フサイクルアセスメントやレギュラトリーサイエンスの概念も必要であろう. 第 2 章 POPs,農薬のリスク評価とリスク低減技術 35 第三に行政や現場との関連である.研究成果は,行政では規制,現場では意思決定に活用され る.規制においては,一日許容摂取量(ADI)や最大残留基準(MRL)のように,ある固有の値 に対する決定論的判断が求められる.一方,指導や啓発活動は,最終的にはいずれかに意思決定す るものの,その判断基準として確率が入ってもよい.リスク指標は,本来確率論的概念がベースに なっていることから,研究成果の移転に際して配慮する必要があろう. 次に重点化すべきことに複合影響や組み合わせ効果がある.複合(混合)系であり,かつ非定常 の環境に対する農薬や POPs の影響を調べる際には,化学物質どうしの対象生物に対する相互作用 を念頭に置く必要がある.ここでは,異なる農薬間の相互作用と肥料など農薬以外の化学物質との 関係が考えられるが,農薬の場合,特に光学異性体や代謝物間で環境中動態や生理活性(毒性)が 異なることがあり,これらの割合が経時的に変化しつつ,同一環境中に混在するため,さらに複雑 な解析が求められる.それらに対応して,これまでに開発した技術を実用化するには,単なる組み 合わせでなく,病原微生物のリスク管理におけるハードルテクノロジーのように,めざすべきレベ ルを明確にした適切な総合的管理を開発する必要がある. 同様に,農薬の環境中挙動に関するシミュレーションモデルは,水田河川生態系において,時 空間を把握できるレベルになった.一方,非対象水生生物に対する毒性評価は数少ない指標生物 の急性毒性をようやく把握できる段階であり,今後リスク評価対象として,個体群,生態系,さ らには生物多様性への影響まで発展させるためには,種の感受性分布(SSD: Species Sensitivity Distribution)の考え方の導入や,メソコズム研究を含む膨大な農薬の環境影響および各種モニタ リングのデータとの関連性を解析する必要がある.さらに,底質における農薬の動態と底生生物へ の影響,農薬の土壌中挙動と陸域生態系への影響,大気中挙動とヒト健康および環境に対する影響 等,課題は山積みである. 今後,農薬の新規有効成分の開発や環境汚染の低減を目指した製剤開発とともに,植物保護に限 らず,農作物栽培における幅広い管理技術もあわせて,総合的作物管理技術(ICM)の中で研究や 技術開発を推進することが肝要である. 引用文献 1)石井康雄(2004)水稲収穫期における有機リン系およびカーバメート系殺虫剤の残留特性.農 業環境技術研究所報告,第 23 号,1-14 2)石井康雄,稲生圭哉,小原裕三(2004)田面水および土壌中における水田除草剤の経時的変化 に基づく止水期間の検討.農業環境技術研究所報告,第 23 号,15-25 3)稲生圭哉(2004)水田環境における農薬の挙動予測モデルの開発と有効性の検証.農業環境技 術研究所報告,第 23 号,27-76 4)Inao, K., Mizutani, H., Yogo, Y. and Ikeda, M.(2009)Improved PADDY model including photoisomerization and metabolic pathways for predicting pesticide behavior in paddy fields: Application to the herbicide pyriminobac-methyl, Journal of Pesticide. Science, 34(4),273–282 5)石原 悟(2008)河川生態系の一次生産性に及ぼす除草剤の影響評価手法開発.農業環境技術 研究所報告,第 25 号,1-92 6)小原裕三,清家伸康,西森基貴,益永茂樹,細見正明(2010)日本全域におけるパッシブエ アサンプリング法による有機塩素系農薬類の広域同時モニタリング.環境科学会誌,23(1), 3-17 36 引用文献 7)Murano, H., Otani, T. and Seike, N.(2010)Dieldrin-dissolving abilities of the xylem saps of several plant families, particularly Cucurbita pepo L. Environmental Toxicology and Chemistry, 29 (10) , 2269-2277 8)永井孝志,稲生圭哉,堀尾 剛(2008)不確実性を考慮した農薬の確率論的生態リスク評価: 水稲用除草剤シメトリンのケーススタディー.農薬学会誌,33(4),393-402 9)西森基貴,小原裕三,魏永芬(2010)日本で使用された有機塩素系農薬類の北極域への移動に 関するマルチメディアモデル.環境工学研究論文集,47,135-140 10)Otani, T., Seike, N. and Sakata, Y.(2007)Differential uptake of dieldrin and endrin from soil by several plant families and Cucurbita genera. Soil Science & Plant Nutrition, 53, 86-94 11)Phong, T.K., Watanabe, H., Hien, T.Q., Vu, S.H., Tanaka, T., Nhung, D.T.T. and Motobayashi, T. (2008)Excess water storage depth − a water management practice to control simetryn and thiobencarb runoff from paddy fields. Journal of Pesticide. Science, 33(2),159–165 12)清家伸康,稲生圭哉(2007)土壌中の POPs の残留分析と要因解析.第 24 回農薬環境動態研 究会資料,17-24 13)WARC, TZW, NIAES, OCWD(2008)Report on international collaborative research(Volume I) : Development of Multi-residual Analytical Methods for Pharmaceuticals, Perfluorinated Compounds, Nitrosamines, Hormones and POPs in Water, pp.237 14)WARC, TZW, NIAES, OCWD(2009)Report on international collaborative research(Volume II) : Development of Multi-residual Analytical Methods for Emerging Contaminants in Water, pp.148 15)Watanabe, H. and Takagi, K. (2000) A simulation model for predicting pesticide concentrations in paddy water and surface soil I. Model development. Environmental Technology 21, 1379-1391 16)Watanabe, H., Inao, K., Vu, S.H., Ishihara, S., Takagi, K. and Tournebize, J.(2007)Chapter 8 - Pesticide Exposure Assessment in Rice Paddy Areas: A Japanese Perspective,“Pesticide Risk Assessment in Rice Paddies: Theory and Practice”, eds. by E. Capri and D. Karpouzas Elsevier Ltd., 167-214 17)與語靖洋(2006)大気中に拡がる農薬 ― その拡散と制御を考える ― .農業環境技術研究所 研究成果発表会 2006 講演要旨,15-24 18)横山淳史,大津和久,堀尾 剛,遠藤正造,石原 悟(2008)コガタシマトビケラの飼育法マ ニュアル,pp.28 19)横山淳史,大津和久,堀尾 剛,遠藤正造,石原 悟(2008)コガタシマトビケラ 1 齢幼虫を 用いた農薬の急性毒性試験法マニュアル,pp.26 37 第 3 章 栄養塩類の動態と負荷低減 1.はじめに 20 世紀初頭に約 16 億人だった世界の人口は,1961 年には 30.8 億人,2007 年には 66.7 億人まで 増加した.人の生存を支える最も重要な食料である穀物の 1900 年の生産量は約 3 億 t であったが, 1961 年に 8.8 億 t,2007 年に 23.5 億tと人口増加を上回る速度で増加した.特に中国,インドを含 むアジアでの生産量の増加が顕著で, 2007 年の生産量は 1961 年の 3.4 倍となった(人口は 2.3 倍). これによって世界平均の一人当たり穀物消費量は,285 kg(1961 年)から 349 kg(2007 年)まで 上昇した.この急激な生産拡大を実現させるために,品種改良などに加えて,大量の化学肥料を投 入して単収を増大させることが必要であった.農業由来の栄養塩類が地下水の硝酸汚染を引き起こ し,河川湖沼の水質を悪化させ,富栄養化を招いたのは,人口の増加を支えるための必然であった とも言えるであろう. 欧米では 1980 年代,地下水や河川水の汚染が深刻となり,農業・畜産がその主たる原因である ことが認識されるようになった.EU では,1991 年に硝酸指令を公布して,地下水や表流水中の硝 酸イオン濃度が WHO の飲料水基準である 50ppm(硝酸性窒素の濃度では,11.3mgN/L)以下と することを定め,メンバー国に,施肥の時期・量,作付け体系,家畜ふん尿の貯蔵,農地への投入 などの適正化を図ること,また国内の水質モニタリングを実施して硝酸イオン濃度や地形等に基づ いて脆弱地域を設定し,より厳しい対策をとることなどを求めている.このような対策にもかかわ らず,水質の改善は見られるものの,地下水は全体の 15%の地点で,表流水は 3%の地点で硝酸性 窒素濃度が 11.3mgN/L を超えており,依然として高い窒素濃度を示す地域が広く分布している. これまでの長期間継続的な負荷の影響からの回復は容易ではないことがわかる. わが国においても 1999 年に公共用水域および地下水における硝酸性窒素および亜硝酸性窒素の 水質環境基準(10mgN/L)が設定され,現在約 5%の井戸でこれを満たしていない.また閉鎖性水 域の水環境改善を目指した総量規制においても農業など面源負荷の削減が課題となっている.日本 は,農地面積の減少により肥料の総使用量は減少傾向にあるが,耕地面積当たりでは 1961 年から 一貫して 100kg/ha から 150kg/ha 程度の施肥量(N,P2O5 のいずれも)であり,特にリン酸肥料 の面積当たり投入量は世界の中で最高水準である.このような状況の下,この 10 年間,栄養塩に よる水質汚染の解明を目的として,長期間投入され続けた窒素やリンを含めて,栄養塩が土壌から 地下水,さらに河川へ流出するプロセスの解明の研究が進められるとともに,そのモデル化が取り 組まれた.また,負荷削減のための施策を考えるための基礎情報の提供を目的として,農業由来の 窒素,リンの環境への負荷量,あるいは食生活からも含めた負荷量の国や地域スケールでの評価が 行われた.これらの研究の基礎となるデータベースの作成やモニタリング手法の開発なども実施さ れた.以下にこれらの研究成果を紹介する. 2.栄養塩の土壌・地下水層での挙動と流出メカニズムの解明 施肥などによる農地への窒素インプットと作物吸収などによるアウトプットとの差から計算され る地表面収支から推定した窒素負荷ポテンシャルと実際の河川からの流出負荷量は,良く一致する 場合と大きく異なる場合がある.また,河川流出負荷量のピークは窒素負荷のピーク年に数年〜数 十年遅れて発生する場合も多い.これらは,土壌中および地下水中での水および窒素の輸送過程の 38 2.栄養塩の土壌・地下水層での挙動と流出メカニズムの解明 違いによると考えられ,土壌や地形による輸送過程の詳細な解明の研究が行われた. ⑴ 陰イオン吸着定量法の改良とその利用 黒ボク土は陰イオン吸着能を有し,硝酸イオン(NO3− )を吸着するために,土層内の NO3− の 移動速度は水移動速度に比べて小さくなるので吸着能の正確な測定が重要である.従来用いられて きたバッチ法では土壌に加えられる溶液量が多いために,吸着態硫酸イオン(SO42−)とのイオン 交換による NO3−の吸着も同時に進み,NO3−吸着容量を過大評価していた.そこで,硝酸塩溶液を 混和した土壌を充填したカラムに水を浸潤させる方法(カラム浸潤法)が開発され,吸着態 SO42− の脱離をほとんど引き起こすことなく,ほ場条件に近い吸着態イオン組成を持つ黒ボク土による NO3−吸着容量の測定を可能とした(平成 13 年度研究成果情報 , 加藤)(Katou et al., 2001).この 方法を用いて,黒ボク土への硝酸イオンと塩化物イオンの競合吸着の特性が明らかにされ(Katou, 2004) ,親水性有機物の流出過程にも応用された(Ahmad et al., 2005).また,北海道の主要な黒 ボク土壌について,硝酸イオンの水移動に対する相対移動速度と土壌化学的特性の関係が検討さ れ,水に対する相対移動速度はほぼ 0.5 〜 1 の範囲にあること,陰イオン吸着能の大きい層位ほど 相対移動速度が小さいこと,相対移動速度は酸性シュウ酸塩可溶ケイ素含量と高い負の相関を示す ことが示された(三木ら , 2009) .これらから土壌への NO3−の吸着による相対移動速度の低下が, 施用された NO3−の下方移動の抑制に実効的に働くことが明らかになった. ⑵ 黒ボク土層から地下水層への水と溶質の移動過程 土壌水分状態(30 分間隔の体積含水率,圧力ポテンシャル)の観測に基づいて,圃場条件下の 土壌中における不均一な水移動を,マトリックス流(ダルシー式に従い巨視的に見て水平方向に均 一とみなせる流れ)と選択流(粗大孔隙や亀裂を通して流下する部分的に速い流れ)に区別して 定量化する手法が開発された(平成 19 年度研究成果情報 , 江口ら).この手法と土壌水中硝酸イオ ン濃度の観測値を用いて,農業環境技術研究所内の黒ボク土ほ場における水と硝酸性窒素の根域 土壌から浅層地下水面への移動,さらに難透水層を通した溶脱の挙動が明らかにされた.黒ボク土 畑ほ場からの流出は,鉛直一次元のダルシー式で近似できるマトリックス流が主体であり,選択流 は年に数回しか発生しない.しかし選択流発生時の日流出量は 50mm/日以上と大きく,選択流は 年間流出量の 16 〜 27%を占める(平成 19 年度研究成果情報 , 江口ら)(江口 , 2006;Eguchi and Hasegawa, 2008) (図 1) .前述のように硝酸イオンは黒ボク土壌に吸着され流出が遅延する(本対 象ほ場でも,硝酸イオンの半分は吸着態であった)が,図 2 に示したように,硝酸イオンは水の流 出速度とほぼ同じ速度で 1 mまで達しており,硝酸性窒素の土層内移動速度は水の移動速度と見か け上ほぼ一致した.これは選択流の寄与により見かけの流出速度が速まったためと考えられた(平 成 15 年度研究成果情報 , 江口ら) (江口 , 2006) .根群域下端から溶脱した硝酸性窒素が浸透水とと もに下方移動して浅層地下水面に到達するのに要する時間は年次変動が大きいが,平均 2.8 年(年 ごとの降水量により 1.6 〜 6.0 年)と推定された(平成 15 年度研究成果情報 , 江口ら). 浅層地下水に到達した後の挙動,すなわち,難透水層である常総粘土層を通した鉛直方向への流 出と水平方向への流出の割合が,浅層地下水位と硝酸イオン濃度の面的分布の通年観測に基づい て,難透水層深度分布(Eguchi et al., 2009b)と土層別の間隙率および飽和透水係数を考慮した数 値解析により推定された.対象ほ場では常総粘土層を通じた漏水量は従来の報告より 1 桁大きな値 (2001 年 11 月 25 日〜 2002 年 10 月 11 日の間に 188 mm,浸透水量の 85%)と推定された(平成 14 年度研究成果情報 , 江口ら) . 30ಽ㑆㒠᳓㊂ (mm) 第 3 章 栄養塩類の動態と負荷低減 39 0 2 4 2001ᐕ1010ᣣ 3:30~ 㒠᳓㊂ ว⸘119.8 mm 㒠㔎ᒝᐲ ᐔဋ5.83 mm h–1 6 8 120 SE[Dp] 100 ㆬᛯᵹ㊂ Dp ᳓㊂ (mm) 80 60 㒠᳓㊂ P Dm Dm + S0–100 40 20 ᷓ䈘1 m䉁䈪 䈱ጀౝ ᳓㊂ᄌൻ S0–100 䊙䊃䊥䉾䉪䉴 ᵹ㊂ Dm 0 t1 –20 0 t2 t 3 t4 12 24 36 㒠㔎㐿ᆎ䈎䉌䈱ᤨ㑆 (h) 䊙䊃䊥䉾䉪䉴ᵹ䋨Dm䋩䈲䇮ᷓ䈘1m䈱Ⓧ ᳓₸䈱㑐ᢙ䈪䈅䉎ਇ㘻ㅘ᳓ଥᢙ䈫䇮 ᷓ䈘90䈍䉋䈶110cm䈱ജ᳓㗡䈎䉌᳞ 䉁䉎േ᳓൨㈩䈱Ⓧ䋨䉻䊦䉲䊷ᑼ䋩䉋䉍▚ 䈚䇮ㆬᛯᵹ䋨Dp䋩䈲㒠㔎㐿ᆎ䈎䉌⚳ੌ ⋥ᓟ䉁䈪䈱ᦼ㑆䈱ጀౝ᳓ᡰ䈮䉋䉍 ផቯ䈚䈢䇯䈖䈱ᤨ䇮ጀౝ䈱᳓㊂ᄌ ൻȴS0-100䈲䇮ᷓ䈘1m䉁䈪䈱Ⓧ᳓₸ 䈱ᄌൻ䈎䉌᳞䉄䉎䇯ฦ㒠᳓䉟䊔䊮䊃䈮䈧 䈇䈩䇮ㆬᛯᵹ㊂䈏䈠䈱ᮡḰ⺋ᏅSE[Dp] 䈱୯䉕࿁䈦䈢ᤨ䈱䉂ㆬᛯᵹ䈏⊒↢ 䈚 䈢䈫್ᢿ䈚䈢䇯 48 図 1 土壌中の不均一な水の流れの定量化(降水量 P から Dm と△ S0-100 を差し引いて選択流 Dp を求める) ᷓ䈘㩷㩷㫑 㩷㩷㩿㫄㪀 㪇 㫑㫇 㪈 㪉 㪊 㪈㪇 㪄㪈 㪥㪦㪊 㩿㫄㫄㫆㫃 㪺㩷㪣 㪀 D E ᷓ䈘㪈㪅㪌㩷㫄 ᷓ䈘㪈㩷㫄 㪄㩷 㪌 㪿 㪇 㪈㪐㪐㪌㪈㪐㪐㪌 㪈㪐㪐㪍 㪈㪐㪐㪍 㪈㪐㪐㪎 㪈㪐㪐㪏 㪈㪐㪐㪏 㪈㪐㪐㪐 㪈㪐㪐㪐 図 2 黒ボク土畑の (a) 作土からの浸透水の到達深さ zp と浅層地下水位 h の変化と,(b) 深さ 1 および 1.5 m の 液相 NO3−濃度の変化 (1995 年秋〜 1996 年秋に作土から移動し始めた水は,1997 年夏に深さ 1 m,1998 年夏には深さ 1.5 m に 到達し,各々の深さ zp への到達時期は,各々の深さの液相 NO3−濃度ピークとほぼ一致した。) 40 2.栄養塩の土壌・地下水層での挙動と流出メカニズムの解明 ⑶ 水畔域における脱窒による窒素除去能の評価 窒素が土壌中,地下水へ流出する過程で,脱窒による窒素の除去が重要な役割を果たしている場 合がある(江口 , 2008) .三河湾への流入河川である矢作川下流域では,洪積台地上で茶の栽培が 盛んで施肥由来の窒素による地下水汚染が顕著である.一方,その茶園に囲まれた沖積地の水田の 地下水は硝酸イオンが高濃度の場所と低濃度の場所が存在している.この矢作川下流域を対象とし て,水・窒素の流動と脱窒の寄与が検討された.水田の縁部および内部に複数の深度の井戸を設置 し,毎月の硝酸イオン濃度,0.5 m 深ごとに採取した土壌試料の脱窒活性(アセチレンブロック法 による) ,および地下水と水田周辺の茶園内の地表面下 0.8 m で採取した土壌浸透水の NO3-N の窒 素安定同位体比(δ15N)などが測定された(恒川ら , 2006).また地下水位と地下水の水素及び酸 素の安定同位体比(δD-H2O およびδ18O-H2O)の鉛直分布の測定結果から,各深さの地下水はそ の上下の地下水がどのような比率で混合したものであるかを推定した.これらの測定結果から, 脱窒の起こっている地点と深度が特定された(平成 19 年度研究成果情報 , 江口ら)(Eguchi et al., 2009a) .茶園に接する位置にある井戸について各パラメータの鉛直プロファイルを図 3 に示す. この地点の深度 2 m 付近には高い脱窒活性と高い全炭素含量が認められた.また地下水の混合比 から推定した値に比べて,硝酸イオン濃度の低下,δ15N-NO3−の上昇および HCO3−濃度の上昇が 見られ,この位置で従属栄養型脱窒反応が起こっていることを,見かけ上ほぼ定量的に説明でき た.硝酸イオンの除去速度は,NO3−濃度分布と地下水流速を考慮した物質収支に基づき 32.6 mg N/m2/日(119 kgN/ha/年)と推定された.以上の結果より,茶栽培の盛んな洪積台地に隣接した 沖積水田下の浅層地下水中における脱窒が農業集水域からの硝酸性窒素除去に大きく寄与している ことが定量的に示された. ᷓ䈘 P 12± PPRO /± G112± Å D +&2± PPRO /± E F G ో⓸⚛ 䋨71䋩 ో⚛ 䋨7&䋩 ⣕⓸䈮䉋䉎 Ớᐲ ⣕⓸䈮䉋䉎 Ớᐲૐਅ H 䉝䉶䉼䊧䊮㒖ኂᴺ 䈮䉋䉎ᣂ㞲ფ䈱 ⣕⓸ᵴᕈ᷹ቯ ਅ᳓ᷙว 䈱䉂䈱⚿ᨐ 7&71㊂ JNJ± ⣕⓸ᵴᕈ P/ 12J± ੇ '>12±@ ±PPRO /± ᓥዻᩕ㙃ဳ䈱 ⣕⓸ᔕᑼ P0 .12ṁᶧ 䉕ᷝട䈚䋬᷷ᐲ㷄 ⣕⓸ㆊ⒟䈪䈱േ 䈪ᤨ㑆ህ᳇ၭ㙃䈚 ⊛หಽല 䈢䈫䈐䈱12↢ᚑ㊂ ᨐ䈮䉋䉎୯ '>+&2±@ PPRO /± &+ 2 12 + &2 1 + 2 図 3 洪積層と沖積層の境界付近における脱窒の定量的評価 有機物含量が高く,土壌培養による脱窒活性が高い洪積層と沖積層の境界付近における硝酸イオン除去 速度は,物質収支より,32.6mgN/m2/ 日と推定された。また,その反応は見かけ上従属栄養型の脱窒反 応式によりほぼ説明できた。 第 3 章 栄養塩類の動態と負荷低減 41 このように,安定同位体自然存在比は流域内での物質の挙動を解明するための有力な手がかり となり,硝酸イオンの動態解明には硝酸イオンの窒素安定同位体比(δ15N-NO3−)を用いた手法が 使われてきた.しかしδ15N-NO3−値は窒素負荷物質の起源によって異なる(化学肥料由来の窒素 は 0‰に近い値,有機質由来はそれよりも高い値)ため,負荷物質の種類と脱窒の進行程度を同時 に明らかにするのは困難である.そこで,酸素安定同位体比(δ18O-NO3−)も同時に測定すること により,これらの同時推定が行われた(平成 22 年度研究成果情報 , 中島ら).脱窒に伴う窒素およ び酸素の安定同位体比の変化は,各々硝酸イオン濃度の変化と表 1 に示したレイリー式に従う.こ こでは,土壌カラムを用いた培養実験を行い,土壌溶液中のδ15N-NO3−値,δ18O-NO3−値の変化を 測定し,これらと硝酸イオン濃度の対数との間に直線関係があることから脱窒が進行していること を確認するとともに,その傾きから同位体濃縮係数(ε)が推定された.亜酸化窒素還元酵素を 欠いた脱窒菌を用いた方法(Silva et al., 2000)を利用することにより,微量試料の同位体比の測 定が可能となり,世界で初めて濃縮係数を精度よく求めることに成功した.その結果,窒素と酸素 の同位体濃縮係数は同程度の大きさであった(15ε=−18.1〜−21.9,18ε=−17.6〜−22.3).これは 脱窒過程におけるδ15N-NO3−の変化とδ18O-NO3−の変化の比,すなわち 15ε: 18 εがほぼ 1:1 であ ることを意味しており,定説の 2:1 とは大きく異なる値であった.さらに,現場の地下水などの δ15N-NO3−とδ18O-NO3−および硝酸イオン濃度を測定し,脱窒前のδ18O-NO3−が一般に 0 〜 5‰程 度の値をとることを利用して,レイリー式と先に求めた同位体濃縮係数に基づいて窒素負荷物質の 脱窒前δ15N-NO3−を求めることができ,負荷物質の起源を推定できる.台地畑 ― 谷津田連鎖系の 地下水調査では,集水域内上流の台地畑には起源の異なる硝酸イオンが存在すること,下流の谷津 田では脱窒がさかんに起こっていることが確認された(図 4). 表 1 脱窒過程のレイリー式 δ15N-NO3−=δ15N0-NO3−+ 15εln(C/Co) δ18O-NO3−=δ18O0-NO3−+ 18εln(C/Co) δ15N-NO3−:試料の窒素安定同位体比 δ18O-NO3−:試料の酸素安定同位体比 δ15N0-NO3−:脱窒前の試料の窒素安定同位体比 δ18O0-NO3−:脱窒前の試料の酸素安定同位体比 15 18 ε:窒素安定同位体濃縮係数 ε:酸素安定同位体濃縮係数 C:試料の濃度,C0:脱窒前の試料の濃度 2.栄養塩の土壌・地下水層での挙動と流出メカニズムの解明 ⣕⓸ࠍฃߌߚ01 ᵹၞਅㇱߩ⼱ᵤ↰ 20 100 G118O-NO3- (‰) 15 80 10 60 5 40 0 20 -5 0 5 ൻቇ⢈ᢱ↱᧪01 ᵹၞᵹߩ⇌ 10 15 N NO3- (‰) G15 N-NO 20 25 NO3- Ớᐲ (mgN L-11) 42 0 ᯏ⾰↱᧪01 ᵹၞᵹߩ⇌ 図 4 台地畑-谷津田連鎖系 ( 茨城県石岡市 ) における浅層地下水の硝酸性窒素濃度および窒素・酸素安定同位 体自然存在比の空間分布 流域上流の台地畑では起源の異なる窒素が存在していること,また下流の谷津田では NO3−濃度の低下お よびδ15N-NO3−とδ18O-NO3−の上昇から脱窒がさかんに起こっていることが分かった. (トピックス)台地畑における浅層地下水中の亜酸化窒素の由来 ― 硝酸イオンの安定同位体比を 用いた窒素循環の推定 肥料や家畜ふん尿由来の窒素は地下水へ流出し,脱窒あるいは硝化のプロセスにともなって一酸 化二窒素(N2O)の間接発生の原因となる.硝酸イオンや N2O の窒素と酸素の同位体比の測定は, N2O の起源と生成プロセスの解明にも有効である. 茨城県内の豚舎跡地付近の台地畑は,厚層黒ボク土であり,畑地では主に化学肥料を施用し, また近傍の養豚農家では 1999 年頃まで素堀りによって家畜ふん尿を廃棄していた.3 年間にわた る浅層地下水の観測の結果,ある観測井戸で高い窒素濃度と N2O 濃度(20–5630μg N/mL)が観 測された.流下する浅層地下水中の N2O の窒素同位体比(δ15N-N2O)は特に高水域において全 窒素の同位体比(δ15N-TN)よりかなり低い(図 5a).脱窒により生成する N2O の同位体比は, N2O から N2 への脱窒の過程で重くなるため,基質(硝酸イオン)の同位体比と同程度となるの で,ここで観測された N2O は硝酸化成の過程で生成していると考えられた.また,δ15N-N2O, δ18O-N2O の水位による変動(図 5b)の要因としては,2 種類の N2O の混合,あるいは脱窒等によ る N2O の還元が考えられる.一般の脱窒におけるδ15N-N2O とδ18O-N2O の変化量の比はおよそ 1:2 であるのに対して,この観測結果による比は 1:1 に近く,肥料由来の軽い N2O と家畜ふん尿由来の 重い N2O の混合比の変化も起因していることがわかった(平成 16 年度研究成果情報 , 中島ら). 第 3 章 栄養塩類の動態と負荷低減 43 (a) 8 6 ɷ15N TN -20 -40 -2.8 4 2 ɷ15N N2O -2.6 -2.4 15 ɷ18O N2O 60 40 20 0 -2.0 ૐ᳓ᦼ ኅ⇓䈸䉖ዩ ↱᧪㪥2 O 10 N2 㪦ᷙว 0 -10 㜞᳓ᦼ ⣕⓸ -20 0 -2.2 (b) 20 10 㱐 N(‰) 100 80 㱐15N TN (‰) 㱐18O, 㱐15N N 2O (‰) 30 -30 -1.8 ᣉ⢈↱᧪㪥2 O 30 ਅ᳓䋨䌭䋩 40 50 60 70 18 㱐 O(‰) 80 90 図 5 茨城県台地畑における地下水観測結果. (a)地下水位と安定同位体比の関係,(b)N2O のδ15N-δ18O プ ロット ⑷ リンの流出過程 リンは溶解度が低く土壌粒子に強く収着されるため,土層内を下方移動しにくい.これまで水田 から流出するリンの 6 〜 8 割は表面流出による,また,畑・樹園地からの流出はきわめて少ないと 考えられてきた.しかし,下層土に亀裂を有する粘土質転換畑における観測の結果,土壌粒子に 吸着されたリンが懸濁態として下方移動し,暗渠へ流出することが明らかになった(平成 16 年度 研究成果情報,鈴木ら) (鈴木ら,2005) .新潟県内の細粒質斑鉄型グライ低地土(畑転換 9 年目, 大豆作付け,施肥量 12 kg-P/ha)における,降雨時の土壌水分状態,暗渠流出水および表面流出 水の量,流出水中の懸濁態(>0.1μm 画分) ・溶存態リン濃度の連続観測の結果,降雨により作土 と耕盤層の境界面(深さ 20 cm)の土壌水の圧力ポテンシャルが上昇して正になると暗渠流出が始 まることが見いだされた(図 6a) .暗渠流出水中のリン濃度は表面流出水中の濃度よりも高く(図 6b) ,最高 1.3 mg-P/L に達した.作付け期間を通した暗渠流出量は施肥量の約 5%で(その約 9 割 が懸濁態) ,表面流出量(施肥量の約 2%)を大きく上回った. 㒠㔎ᒝᐲ ᥧᷯᵹᵹ㊂ -200 -250 IJķıশ IJĸıশ IJĹıশ 2004㩷 ᐕ7 30 3 20 2 10 1 0 0 IJĺıশ 䊥䊮Ớ Ớᐲ䋨mg-P L 䋩 -150 150 ᷓߐ20 cmߩ ജࡐ࠹ࡦࠪࡖ࡞ -100 4 㕙ᵹᵹ㊂ 0.6 -1 40 -50 25 0.7 5 ᥧᷯᵹ ᵹᵹ㊂䋨mm h-1䋩 50 㔎ᒝᐲ䋨mm h-1䋩 㒠㔎 ᷓ䈘20 cm䈱ജ䊘䊁䊮䉲䊞䊦 䊦䋨cm䋩 㩷 0 0.5 20 㩷䊥䊮Ớᐲ䋨ᥧᷯᵹ䋩 㩷䊥䊮Ớᐲ䋨㕙ᵹ䋩 0.4 15 0.3 10 0.2 5 0.1 ᥧᷯᵹᵹ㊂ 0.0 IJķıশ IJĸıশ IJĹıশ 0 -1 ᥧᷯ䈍䉋䈶 㕙ᵹᵹ㊂䋨 mm h 䋩 IJĺıশ 2004ᐕ7 図 6 降雨時のリンの流出.(a) 作土と耕盤層の境界付近(深さ 20 cm)の圧力ポテンシャルと暗渠流出流量の 変化,および (b) 暗渠流出水・表面流出水中のリン濃度の変化 44 3.流域スケールの栄養塩動態シミュレーションモデル 3.流域スケールの栄養塩動態シミュレーションモデル ⑴ 農地由来の硝酸性窒素による地下水汚染リスクの推定モデル さまざまな負荷削減シナリオによる硝酸性窒素汚染リスクの低減効果を予測するために,農地か ら流出した硝酸性窒素による地下水汚染リスクを,対象地域の土壌・地形的特性に基づいて面的 に予測するためのリスク評価システム RealN(Risk evaluation for agricultural leached Nitrogen) が開発された(平成 22 年度研究成果情報 , 板橋ら).ここで,地下水汚染リスクとは,ある地点の 地下水の硝酸性窒素濃度が,気象条件の年々変動の下で基準(10 mgN/L)を超える確率を表す. RealN は,①各地点における硝酸性窒素の農地土壌から地下水への流出濃度,流出量を推定するモ デルと,②その結果を基に流域の地形的特性を反映して混合や脱窒を経た後の地下水中の硝酸性窒 素濃度の面的分布を推定するモデルとからなる. ①のモデルとして,LEACHM モデル(Hutson, 2003)が改良の後用いられた.LEACHM モデ ルは,土壌中の水の移動,窒素形態変化・移動,作物吸収などが記述されており,対象地点におけ る土壌・気象特性や栽培・施肥管理を入力データとして与えることにより,保水性・透水性や有機 物含量の異なる土壌からの硝酸性窒素の地下水流出濃度・流出量を予測する.モデルの利用に当 たっては,黒ボク土の有機物分解プロセスを,当研究所で得られた最新の知見を反映した RothC モデルと同様のものに変更するなどの改良が行われた. ②のモデルとして,浅層地下水中での環境負荷物質の混合・脱窒などによる減衰の過程から濃度 分布を面的に予測するための GIS モデル MacT(Mixing areal chemical Transport)が開発された (平成 22 年度研究成果情報,板橋ら) .対象地域を任意のグリッドサイズにメッシュ化し,各メッ シュの浅層地下水への土壌浸透水量および環境負荷物質(硝酸性窒素)の流入負荷量を入力データ として与え,デジタル標高データおよび土壌類型情報に基づいて,浅層地下水の流動,地下水混合 による濃度変化や生物化学的過程による濃度減衰を推定し,環境負荷物質の地下水中濃度分布を面 的に予測する. 関東地方の畑作地域を対象として,現況の土地利用・作付状況と施肥基準をもとに過去 31 年間 の気象データを用いて,RealN により地下水中硝酸性窒素濃度の面的分布の予測が行われた.得ら れた結果は地方公共団体による実測値とよく一致し,地下水中硝酸性窒素濃度が地下水環境基準値 の 10 ppm を超過する確率(地下水汚染リスク)が高いことが示された.対象地域では化学肥料と 堆肥が投入されているが,化学肥料のみ施用量を 50%削減しても,硝酸性窒素濃度を基準値以下 に低下させることは困難であること,一方,化学肥料を 50%削減し,堆肥施用をその代替分のみ に限れば(堆肥施用量は約 4 割削減) ,地下水汚染リスクを効果的に低減できると予測された(図 7) . なお,①と同様の目的のモデルとして,英国ローザムステッド農業試験場で開発された窒素ター ンオーバーモデルを基本としたモデルも作成された(平成 16 年度研究成果情報 , 竹内ら). 第 3 章 栄養塩類の動態と負荷低減 45 NP Ě͘↪࿑ Ě͘ઍ⊛䈭ᩱၭ‛ 図 7 RealN で予測した負荷削減シナリオの地下水汚染リスク低減効果 ⑵ 水,窒素の溶脱時間の推定 上記の研究(3(1) )では,RealN を用いて負荷が長期間継続した場合の地下水への流出濃度・ 量を推定した.一方,2 節で示したように,地表面で生じた窒素負荷が地下水面まで到達する時間 は水の移動・流出速度とともに,硝酸イオンの土壌への吸着などによる遅延に影響を受ける.水分 移動予測モデル(HYDRUS-1D)を用いて,土壌の種類を考慮した硝酸性窒素の地下水到達時間が 面的に推定された(平成 19 年度研究成果情報 , 加藤ら).土壌特性と土層構成が異なる多くの土壌 が分布する利根川流域を対象に,土壌を 14 種類の土層に分類して,各々の水分保持・透水特性と 陰イオン吸着に関するパラメータを求め,また,流域内の浅層地下水位と年間の正味浸入水量(降 水量と蒸発散量の差)を,国土交通省水基本調査データと利根川水系内の気象観測値から内挿によ り求めた.これらをもとに,土層構成が既知の農耕地(水田を除く)213 地点について,土壌表面 に存在した水が浅層地下水面に到達するのに要する時間を求めた.さらに,遅延係数(陰イオン吸 着による硝酸性窒素の水に対する移動の遅れ)の土層内分布から,硝酸性窒素の地下水到達時間を 推定した.降水量や土壌特性・土層構成,地下水面の深さなどの違いのために,予測した地下水到 達時間には約 0.4 〜 31 年と大きな差が見られること,また,難透水性の粘土層が分布する地域で は,降雨時に浸透水の水平方向への流出がみられ,地下水到達時間も短いことが示された(図 8) . 46 3.流域スケールの栄養塩動態シミュレーションモデル 図 8 利根川流域の農耕地(水田を除く)における硝酸性窒素の地下水到達時間の分布(HYDRUS-1D モデル による推定結果) ⑶ 懸濁物質の流出負荷モデル 硝酸性窒素は溶存態として土壌から地下水へ浸透し河川へ流出するが,リン酸,重金属,ダイオ キシンなどが土壌粒子に吸着した懸濁物質は異なる経路で輸送される.これら懸濁物質の,河川へ の流出負荷量を,土地利用連鎖を考慮して流域単位で算定するモデルが作成された(平成 14 年度 研究成果情報 , 板橋ら) .牛久沼集水域を対象に,地形図や土地利用図などに基づいて各地目にお ける懸濁物質の発生量を実測データや降水データなどに基づいて推定し,それらの河川までの流出 経路(水田排水路経由,あるいは河川への直接流出)を特定して,河川へ流出する懸濁物質負荷量 を流域単位で推定した. ⑷ 日本の農地土壌の物理的性質データベースの作成 前述のようなモデルを用いて,農地土壌中における水移動やそれに伴う環境負荷物質の輸送過程 を広域で予測するためには,土壌の基本的な物理的性質に関するデータが不可欠である.そこで, 農林水産省が実施した土壌環境基礎調査事業の定点調査データ(1979 〜 1998 年まで日本全国の農 地約 2 万地点を対象に,5 年に 1 回の土壌調査・分析を実施)を用いて,信頼性の低いエラー値な どを除くための品質管理を行った後,土壌統群別・層位別・地目別に集計した農地土壌の物理的性 質データベース SolphyJ(Agricultural soil-profile physical properties database, Japan)が作成さ れた.データベースには,土壌の基本的な物理的性質である表 2 に示す 17 の調査・分析項目が, 60 の土壌統群別(農耕地土壌分類第二次案による),最大で第 6 層までの層位別,および 5 つの地 目別(水田、普通畑、樹園地、牧草畑、施設)に,全国平均値,標準偏差,中央値,25 および 75 パーセンタイル値といった基本的な統計値とともに,度数分布図や正規分布曲線のグラフなどが収 納されている(図 9a) .収録データはさまざまな土壌・地目における層位別の保水性・透水性に関 わるパラメータの推定(図 9b)や,水移動にともなう環境負荷物質の輸送過程を解析・予測する ための調査研究などに利用できる. 第 3 章 栄養塩類の動態と負荷低減 47 表 2 農耕地土壌の物理的性質データベース SolphyJ の収録項目 < 17 の調査・分析項目> 各層位の厚さ,かさ密度(仮比重),土粒子密度(真比重) , 土壌硬度, 粗砂含量, 細砂含量, 砂 (粗砂+細砂) 含 量,シルト含量,粘土含量,土壌試料採取時の体積含水率,間隙率,土壌水分吸引圧32 cm(pF1.5),5.0 × 102 cm(pF2.7),1.6 × 104 cm(pF4.2),3.2 × 105 cm(pF5.5, 風乾土) における体積含水率, 飽和透水係数, 全 炭素含量 ;ďͿ Ⓧ᳓₸ P P± ;ĂͿ ⇌䇮╙ਃጀ ጀ⣣ᬀ⾰ 㤥䊗䉪 ⍾ਐᧂᾫ Ϭ ʹ ๆᒁ FP 図 9 データベース SolphyJ の収録データの例 (a) と活用事例 (b) (a) は,かさ密度(水田,厚層腐植質多湿黒ボク土[SSG04b],第一層)の度数分布とそれに当てはめた 正規分布曲線,(b) は,土壌の保水性曲線の推定(図中の数値はデータ数,エラーバーは標準偏差) 4.農地からのアンモニア揮散 農地に施用された窒素肥料と家畜ふん尿はアンモニアのもっとも大きな発生源であり,窒素循環 においても大きく寄与している.肥料からのアンモニア揮散は施肥効率の低下を招くとともに,揮 散したアンモニアの移流・沈着により陸域や水域の富栄養化や硝酸性窒素による汚染など,様々な 広域的環境問題に関与する.施肥窒素からのアンモニア揮散率に関して,主として欧米でのデータ に基づいた推定が行われているが(Bouwman et al., 1997),気象条件や土壌,営農形態の異なる日 本およびアジアに対して妥当な推定値を得ることが課題とされている.2009 年版の欧州環境庁に よるエミッションインベントリーガイドブックには,農耕地への施肥に伴う化学肥料からのアンモ ニア発生係数が春季の平均気温の一次式として示されており(Hutchings et al., 2009) ,気温を指標 とした気候の相違を考慮することができるため,わが国への適用も可能と期待される.しかし,日 本の重要な農耕地である水田や,アンモニア揮散を抑制するはたらきが解明されつつある火山灰土 壌(黒ボク土)が考慮されていない. わが国の水田からのアンモニア揮散量の推定とアンモニア揮散に影響を与える要因の解明のため に,水田(灰色低地土ライシメータ)への施肥に伴う揮散率の測定が行われた(平成 19 年度研究 成果情報 , 林ら) (Hayashi et al., 2008) .代かき時に基肥として尿素 50 kg N/ha を全層施肥,追肥 2 回(1 回目:中干し前に窒素換算で 30 kg N/ha を表面施肥,2 回目:出穂前に窒素換算で 10kg N/ha を表面施肥)を行い,田面とイネ(品種:日本晴)を覆う風洞を用いて大気-水田間のアン 48 5.県・国・東アジアスケールの物質循環の評価と水質への影響 モニア交換フラックスが測定された.基肥後のアンモニアの揮散率は 2.1%であったが,1 回目の追 肥後の揮散率は 21%に達し,一方,2 回目の追肥後はアンモニアが吸収された(図 10),全期間を 通しての揮散率は 8.2%だった.アンモニアの揮散を抑制するためには,全層施肥が有効であるこ と,一時的に窒素過剰の状態を招くような多量の追肥を避けることが必要であると考えられる.な お,田面水からのアンモニア揮散は物理的な現象であるため,揮散量は,田面水のアンモニア性 窒素濃度と pH,水温,風速などに基づく数値モデルにより推定することが可能で,イネを植えな い水田において観測した揮散量は推定値と高い相関を示した.図 10 に,田面水からの揮散量の推 定値も示したが,1 回目の追肥後の揮散量のかなりの部分は,イネから放出されていると示唆され た.また,黒ボク土に関しては,畑における野外実験と培養実験が実施され,施肥に伴うアンモニ ア揮散が非常に小さいことが確認された. 図 10 追肥に伴うアンモニア交換フラックスの推移 これらの結果に基づいて水田における揮散率の推定式を作成し,EEA による推定式とともに用 いることにより,わが国の化学肥料消費量の統計情報(FAOSTAT)を用いて,化学肥料の施肥 由来のアンモニア発生インベントリーが算定された(平成 22 年度研究成果情報,林).なお,黒ボ ク土畑における発生係数は EEA の推定式で求めた値の 1/10 と仮定している.この結果,昭和 46 年〜平成 14 年の全国の化学肥料の消費量 46.3 〜 82.1 万 tN に対して,アンモニアの発生量は 1.4 〜 2.3 万 tN と推定された. また,ここで示した観測やその他の観測結果およびアジアにおけるアンモニア揮散に関する論文 のレビューに基づいて,主として日本と中国における農業からのアンモニア発生量がまとめられた (Hayashi and Yan, 2010) .主たる発生源は,日本は集約的な畜産であり,中国は窒素肥料の過剰 投入であることが示されたが,今後これらのデータに基づいて,それぞれの地域の発生量及び発生 係数の推定をより精緻化する必要がある. 5.県・国・東アジアスケールの物質循環の評価と水質への影響 食料の需給の変化は栄養塩のフロー・収支を大きく変えてきた.食料生産・消費の変化と環境負 荷の関係を大局的に把握することは,将来の影響予測や負荷削減のための対策を考える上で重要で ある.都道府県や国を単位として統計データに基づいて栄養塩の収支を推定する研究が行われた. 第 3 章 栄養塩類の動態と負荷低減 49 ⑴ 農地における窒素とリンのバランス(地表面収支の推定) 農地における窒素やリンのバランスは,農業が環境に与える影響を広域的に評価するための有用 な指標である.農業・畜産に起因する水質汚濁が深刻であった欧米各国において,環境を考慮した 農業を促進するために 1990 年頃から推定が開始され,日本を含む OECD 加盟国について農地への 肥料などによる窒素インプットと作物吸収などによるアウトプットに基づくバランス(余剰量)が 数年に一度公表されている(OECD, 2008) .農地面積(草地を含む)当たりの 2002 年〜 2004 年の わが国の余剰量は欧米諸国と比較して多い(窒素は加盟国中 4 位,リンは 1 位).わが国に関して 類似の方法(図 11)で,余剰窒素および余剰リンの経年的な変化,また,地域的な特徴が明らか にされた.また,養分収支を都道府県・市区町村単位で推定するためのデータベースおよび表計算 ソフト(Microsoft EXCEL)ベースの算出システムが作成された(平成 15 年度研究成果情報 , 三 島ら;平成 21 年度研究成果情報,三島) . 2 種類のデータベースが作成され,各々次のデータが収録されている:① 1997 年の各都道府県・ 市区町村を対象に,耕地面積,70 種の作物の作付け面積と生産量,5 種の家畜の飼養頭羽数,各作 物の養分含有率,各作物への化学肥料施用量,家畜ふん尿排出量および畜種別ふん尿の農地への施 用量・廃棄率のデータ,および,② 1985 年〜 2005 年の 5 年ごとの各都道府県を対象に,6 作目(水 稲,畑作物,野菜,果樹,茶,牧草)5 畜種について作付け面積,飼養頭数,生産量等の基本デー タと窒素とリンの収支推定結果. これらのデータに基づいた窒素に関する推定結果(図 11 の各フローおよび余剰量)を図 12 に 示した.余剰量(=肥料+窒素固定+堆肥+未利用畜産ふん尿-作物吸収(含副産物))は,窒 素,リンとも 1985 年以降減少し続けており,特に化学肥料の施用量の減少によるところが大きい (Mishima et al. 2009, 2010) .都道府県別では,集約的畜産地帯で,未利用の家畜ふん尿が増加傾 向にあること,作目別では,水稲,畑作物,果樹,茶では減少傾向,野菜は増加傾向であることが 示された.ここで用いた推定法は OECD による手法と多少異なるため,特に窒素の余剰量がかな り小さく見積もられている.これは,主として家畜ふん尿の堆肥化過程におけるアンモニア揮散な どによる窒素消失率に関する仮定の違いによるもので,今後これを含むパラメータの精緻化が求め られる. 化学肥料や畜産ふん尿の余剰は,水環境への潜在的な負荷となる.たとえば,西尾(2001)によ り,茨城県と長野県の市区町村を対象に推定した施肥 N 負荷指数(農地面積当たりの化学肥料中 窒素量と,有機質肥料と堆肥から無機化される窒素量の合計から,作物の地上部全体に吸収される ⓸⚛ᶖᄬ ኅ⇓䈸䉖䊶ዩ ᱷ ᧂ↪ ၸ⢈ ൻቇ⢈ᢱ ⥄ὼ䈱 䉍 ㄘ ᡰ 図 11 わが国の農地の窒素余剰量推定に用いた窒素フロー ‛ᅓ 5.県・国・東アジアスケールの物質循環の評価と水質への影響 350 50 0 50 100 kgN ha-1 150 200 250 300 : ࿖ᐔဋ : ㇺᐭ⋵䈱 䊜䊂䉞䉝䊮 : ᭂᄖ୯ 2005 2000 1995 1990 1985 2005 2000 1995 1990 1985 2005 2000 1995 1990 1985 2005 2000 1995 1990 1985 2005 2000 1995 1990 1985 2005 2000 1995 1990 1985 2005 2000 1995 1990 1985 ൻቇ⢈ᢱ ၸ⢈ ‛ᅓ ⥄ὼ䈱⓸⚛ᡰ ᧂ↪ 䈸䉖ዩ ⓸⚛ 図 12 1985 年〜 2005 年までの耕地面積当たりの窒素の流れと余剰窒素量の変化 (kgN/ha) 窒素量を差し引いた値を市区町村の全面積で除した値)が,市区町村ごとの硝酸性窒素濃度に関す る基準超過井戸の割合,あるいは硝酸性と亜硝酸性窒素の合計濃度と高い相関を示すことが示され ている.前記のデータに基づいて都道府県ごとに推定した余剰量を,降水量から蒸発散量を差し引 いた余剰降水量(正味浸入水量)で除して求めた推定濃度は,県別の地下水硝酸性窒素の環境基準 超過率(環境省による全国調査結果)と正の相関が認められ(R = 0.59),農地における栄養塩の 余剰量は,流域の地下水質に影響を及ぼしていることが確認された(Mishima et al., 2009). ⑵ 食飼料の需給にともなう国レベルでの窒素フローの推定 国内での生産および輸入によりわが国に流入した食飼料が,食生活,畜産業,加工業を経て消 費・廃棄に至るまでの物流による窒素のフローを推定する「全国版養分収支算定システム」が作成 され,1982 年〜 1997 年までの 5 年ごとの窒素フローの変遷が示された(平成 15 年度研究成果情 報 , 織田) (織田 , 2006) .推定は,農林水産省による食料需給表,流通飼料便覧,畜産基本調査結 果(畜産統計) ,耕地及び作付面積統計などの統計データ,日本食品標準成分表(科学技術庁資源 調査会)や日本標準飼料成分表(農林水産技術会議事務局)などのデータに基づいている.国内生 産により供給される食飼料中窒素は 1987 年以降減少し,一方輸入される食飼料中窒素は増加を続 け,1997 年には生産量の 2 倍以上となった(国内生産 51.0,輸入 121.2 万 tN).最終的に廃棄(あ るいは農地に投入される)食飼料起源の窒素は,畜産業からが最も多く(1997 年は 80.2 万 tN), 食生活からがこれに続く(同 64.3 万 tN) .総廃棄量は 1992 年をピークとして 1997 年は若干減少 していることが示された. ⑶ 食料生産・消費の変化に基づく東アジアの窒素フローと水質変動予測 東アジアでは近年の経済発展に伴う食生活の質的・量的な変化により,食料の生産・消費に由来 する窒素フロー変化と環境への負荷の増大が予想される.FAO 統計やその他国際機関等による公 第 3 章 栄養塩類の動態と負荷低減 51 表データを用いることにより,各国共通の手法で窒素フローを推定するモデルが作成された(平 成 17 年度研究成果情報 , 新藤ら) .日本,中国,インド,インドネシア,タイに関しては国内行政 区別のデータも副次的に使われている.モデルは作物生産,畜産,食料の貿易,消費による窒素 フローの推定に加えて,アンモニアの揮散と沈着,作物残渣や家畜ふん尿など施用された有機物の 窒素無機化過程,農地および人から流出した窒素の脱窒と河川から海への流出の過程を含む(図 13) .1961 年〜 2005 年の毎年の窒素フローと地下水及び河川水の窒素濃度分布の変化が推定された. わが国に関する窒素フローの経年的な変化は, (1)で推定された余剰窒素量の変化や, (2)の食 料需給の変化の傾向と同様であり,近年は作物輸入による窒素の流入が最も大きい.河川水質も 1990 年頃から改善に向かっていることが推定された(Shindo et al., 2009).8 km グリッドで推定 された地下水,河川の窒素濃度の都道府県平均値は,全国公共用水域の水質調査結果などの実測 データの県平均濃度と高い相関を示した.東アジア,特に中国,インド,ベトナムなどの窒素フ ローはわが国と異なり,窒素流入の大部分を化学肥料が占め,その割合,量は年々増加を示し,中 国東部,東北部などで深刻な水質汚染が起こっている可能性が指摘された(図 14).また,今後の 生産効率の向上を考慮しない場合,食生活の変化(肉消費量の増大)にともなう飼料作物の生産拡 䉝䊮䊝䊆䉝ᴉ⌕ ੱ ャ ‛ 䉝䊮䊝 䊆䉝 ャ ャ 㜞䉺䊮䊌䉪 㘺ᢱ 㝼㘃 ⇓↥ຠ ᵹ ᓴⅣ ⢈ᢱ ኅ⇓ ⓸⚛࿕ቯ ‛ᱷᷲ ㄘ ㄘ䈎䉌 䉝䊮䊝 䊆䉝 ੱ䈎䉌 ኅ⇓䈸䉖ዩ ᵹ ਅ᳓ಣℂ ფᯏ‛ ήᯏൻ Ⅳ Ⴚ䋨ਅ᳓䇮ᴡᎹ䈻䋩 図 13 食料生産・消費に伴う窒素フローモデルの模式図 ϭϵϴϴ ϮϬϬϱ 図 14 窒素フローモデルにより推定した河川水の窒素濃度分布 (mgN/L) 52 6.農業環境モニタリング 大のために,2020 年の窒素負荷は現在の 1.5 倍程度となると予測された(Shindo et al., 2006). 同モデルにより,食料生産・消費の様々なシナリオのもとで,将来の窒素負荷の予測を行うこと が可能であり,東南アジアにおいてエネルギー作物を生産した場合の窒素負荷の変化についても検 討された(平成 20 年度研究成果情報 , 新藤ら) . 6.農業環境モニタリング 前述のとおり,栄養塩は,圃場,流域,国など様々なスケールで循環し,土壌-地下水-河川- 海へと形態変化,相互作用を繰り返しながら移動している.この動態を把握するためには,適切な モニタリングが不可欠である.また広域の現象を対象とする場合には,他機関で測定されている データを有効に利用することが必要である. 水質データに関しては,環境省による公共用水域測定データなど,膨大な量のデータが蓄積され ているが,多くは年度ごとの数値データとしてまとめられているだけであり,利用しにくい.これ らのデータを簡易に表示・検索するとともに,他の気象データなどとの関連解析が可能となるよう なデータベース管理システムが作成された(平成 14 年度研究成果情報 , 竹内ら). また,サンプリングデータの精度に関する検討や,サンプリングや調査方法の提案も行われた. ①大雨や豪雨による侵食や洪水,水田代かき時などに生じる農耕地から水系への懸濁物質(SS) の流出は短期間に集中して生じる.このようなイベント時に集中的に発生する SS は流水中に不均 一に分布している可能性が指摘され,通常の定期的モニタリング時の採取手法(断面における複数 位置の採水とその混合)は,イベント時では適用できないことが懸念されていた.そこで,懸濁物 質濃度の採水位置による濃度変動を確認するために,農業地域を流れる中規模河川(桜川)を対象 に,降雨直後に 4 断面で観測が行われた(平成 17 年度研究成果情報 , 坂西ら).その結果,SS は河 川断面の鉛直方向にほぼ均一に分布していること,TN および TP 濃度はばらつきが大きいが,深 さによる傾向は見られないこと,SS 濃度の川幅方向の変動係数は約 10%であることが示され,通 常の採取方法が有効であると考えられた.②降雨時の土壌浸食に伴うリンの流出挙動を把握するた ᣣ , ㄘᎿ⎇ߩᢳᨒ㧦 ᢳᐲ, ᢳ㕙㐳O ᣣ , ረᕜㄘ႐ߩᢳᨒ ᢳᐲ, ᢳ㕙㐳O 図 15 傾斜枠のカメラ映像と撮影時前後の流出水量及び SS 濃度 ○が写真撮影時を表す。 第 3 章 栄養塩類の動態と負荷低減 53 め,露地野菜畑からの土壌流亡を観測する自動システムが開発された(平成 21 年度研究成果情報 , 坂西ら) .実際の圃場に傾斜枠を設置し,栄養塩流出量の測定と試料採取を行うと同時に,降雨感 知式ビデオカメラにより土壌浸食の状況を記録する.農村工学研究所内ほ場と嬬恋農場の 2 か所で 本システムを稼働させ,降雨による出水時に SS とリンの濃度変動がとらえられた(図 15). さらに,本章で紹介した研究で用いられた多くのモニタリング手法や養分収支調査法,その他, 農業活動が水環境へ及ぼす影響の解明,評価のために利用できる各種の手法や,水質基準と測定方 法に関する情報などをまとめて, 「水環境保全のための農業環境モニタリングマニュアル改訂版」 を発行した(平成 18 年度研究成果情報 , 齋藤ら) . 7.今後の展望 食料生産量の確保と環境負荷の低減を目指した栄養塩管理は,古くて新しい問題である.この 10 年間の研究により,土壌中や流域での窒素やリンの挙動がかなり明らかとなり,それに基づい たモデル化がすすめられた.また,国スケールの窒素循環の評価から農業が広域的に水質に影響を 与えていること,食料需要や輸入量の変化により環境負荷が変化してきたことが示された. 環境保全や安全な食品への指向により,農業も変化しつつある.化学肥料に代わる有機肥料の利 用や堆肥の投入,温暖化対策として進められようとしている農地への炭素蓄積,このために投入さ れる有機物,これらが栄養塩の挙動,流出をどのように変化させるか,環境負荷の視点から取り組 む必要がある.炭素動態に関する研究と栄養塩類研究の連携が重要であろう.硝化抑制剤や緩効性 肥料など,窒素負荷低減を目的とした対策技術に関してもその効果の定量的な評価が必要であろう. これまで,栄養塩研究における窒素は,土と水と作物の中の挙動を中心に研究されてきたが,大 気との間の交換も重要である.5(1)で述べたように,家畜ふん尿の堆肥化の過程におけるアンモ ニア揮散による窒素の損失率は不明な部分が多い.また肥料からの揮散率の測定と,それに基づく 日本やアジアの気象・土壌条件における発生係数および発生量の推定がすすめられているが,デー タをさらに蓄積することにより,さらなる精度の向上が期待される.グローバルな窒素循環の妥当 性はこのようなパラメータの信頼性に依存しているので,研究の強化が望まれる.また,温室効果 ガスである N2O の発生は,陸域における窒素の動態と密接に関係している.これらを含めた窒素 の循環を総合的に評価する研究が必要と考えられる. 引用文献 1)Ahmad, R., Katou, H. and Kookana, R. S.(2005)Measuring sorption of hydrophilic organic compounds in soils by an unsaturated transient flow method. Journal of Environmental Quality, 34, 1045-1054 2)Bouwman, A.F., Lee, D.S., Asman, W.A.H., Dentener, F.J., Van der Hoek, K.W., Olivier, J.G.J. (1997)A global high-resolution emission inventory for ammonia. Global Biogeochemical Cycles, 11, 561–587 3)江口定夫(2006)黒ボク土畑圃場における水移動と硝酸塩の溶脱.土壌の物理性 , 102, 19-30 4)江口定夫(2008)モデルによる土壌,農耕地,流域における窒素動態の理解 3. 地形連鎖系ス ケールの窒素動態 ― 調査技法とモデル化手法 ― .日本土壌肥料学雑誌 , 79, 213-227 5)Eguchi, S. and Hasegawa, S.(2008)Determination and characterization of preferential water flow in unsaturated subsoil of Andisol. Soil Science Society of America Journal, 72, 320-330 54 引用文献 6)Eguchi, S., Nakajima, Y., Yabusaki, S., Kasuya, M., Shibayama, H., Tsunekawa, A. and Imai, K. (2009a)Denitrification during vertical upwelling at an alluvium-diluvium interface below the upland perimeter of a riparian paddy. Journal of Environmental Quality, 38, 2198-2209 7)Eguchi, S., Sawamoto, M. and Shiba, M.(2009b)Field scale determination of the aquifer– aquitard boundary in Joso Clay by cone penetrometry. Soil Science and Plant Nutrition, 55, 337-348 8)Hayashi, K., Nishimura, S. and Yagi, K.(2008)Ammonia volatilization from a paddy field following applications of urea: Rice plants are both an absorber and an emitter for atmospheric ammonia. Science of the Total Environment, 390, 485-494 9)Hayashi, K., Yan, X.(2010)Airborne nitrogen load in Japanese and Chinese agroecosystems. Soil Science and Plant Nutrition, 56, 2-18 10)Hutchings,N., Webb, J. and Amon, B(2009)4.D Crop production and agricultural soils. In EMEP/EEA emission inventory guidebook 2009. 39pp 11)Hutson, J. L.(2003)LEACHM, Model description and user's guide. School of chemistry, physics, and earth sciences. The Flinders University of South Australia, Adelaide. 12)Katou, H.(2004)Determining competitive nitrate and chloride adsorption in an Andisol by the unsaturated transient flow method. Soil Science and Plant Nutrition, 50, 119-127 13)Katou, H., Uchimura, K. and Clothier, B. E.(2001)An unsaturated transient flow method for determining solute adsorption by variable-charge soils. Soil Science Society of America Journal, 65, 283-290 14)三木直倫,松本武彦,加藤英孝(2009)北海道に分布する各種黒ボク土による硝酸イオンの吸 着が移動速度に及ぼす影響.日本土壌肥料学雑誌 , 80, 365-378 15)Mishima, S., Endo, A. and Kohyama, K.(2009)Recent trend in residual nitrogen on national and regional scales in Japan and its relation with groundwater quality. Nutrient Cycling in Agroecosystems, 83, 1-11 16)Mishima, S., Endo, A. and Kohyama, K.(2010)Recent trends in phosphate balance nationally and by region in Japan. Nutrient Cycling in Agroecosystems, 86(1),69-77 17)西尾道徳(2001)作物種類別の施肥窒素負荷量に基づく地下水の硝酸性窒素汚染リスクの評価 手法.日本土壌肥料学雑誌 , 72, 522-528 18)織田健次郎(2006)我が国の食飼料システムにおける 1980 年代以降の窒素動態の変遷.土壌肥 料学雑誌 , 77, 517-524 19)OECD(2008)Environmental Performance of Agriculture in OECD countries since 1990, Paris, France 20)Shindo, J., Okamoto, K. and Kawashima, H.(2006)Prediction of the environmental effects of excess nitrogen caused by increasing food demand with rapid economic growth in eastern Asian countries, 1961–2020. Ecological Modelling, 193, 703-720 21)Shindo, J., Okamoto, K., Kawashima, H. and Konohira, E.(2009)Nitrogen flow associated with food production and consumption and its effect on water quality in Japan from 1961 to 2005. Soil Science and Plant Nutrition, 55, 532–545 22)Silva, S.R., Kendall, C., Wilkison, D. H., Ziegler, A. C., Chang, C. C. Y. and Avanzino, R. J.(2000) A new method for collection of nitrate from fresh water and the analysis of nitrogen and 第 3 章 栄養塩類の動態と負荷低減 55 oxygen isotope ratios, J. Hydrol., 228, 22-36 23)鈴木克拓,足立一日出,関口哲生,吉田修一郎,中野恵子,加藤英孝(2005)亀裂の発達した 転換畑における懸濁物質およびリンの暗渠流出.日本土壌肥料学雑誌 , 76, 43-47 24)恒川 歩 , 今井克彦 , 柴山浩子 , 糟谷真宏 , 江口定夫(2006)矢作川流域の水田下層土における 脱窒.日本土壌肥料学雑誌 , 77, 207-211 56 Ⅱ.地球温暖化と農業 ― 気候変動にそなえる 第 4 章 気候変動と作物生産変動予測 1.はじめに 2007 年に発表された気候変動に関する政府間パネル第 4 次評価報告書(IPCC AR4)では,気候 システムの温暖化には疑う余地がなく,20 世紀半ば以降の世界平均気温の上昇は,その大部分が 人間活動による温室効果ガスの増加によってもたらされた可能性が非常に高い,また現在の気候変 動緩和政策および関連する持続可能な開発の実践では,世界の温室効果ガス排出量は今後数十年間 増加し続けるという意見の一致度は高く,多くの証拠がある,とされている.つまり,21 世紀の 世界の農業は,大気中の二酸化炭素濃度の上昇と温暖化の進行,そしてそれに付随する水循環の変 化や極端な気象現象の頻発などの生産環境の変化に直面していると言える. 大気中の二酸化炭素濃度の上昇は,植物の光合成速度を増加させることから,適度な環境条件下 では作物の生長と収量を増加させ,同時に葉の気孔を閉じ気味にすることにより,水利用効率を高 める.一方,温暖化は低温が問題となっている地域では冷害などの低温ストレスを軽減し,栽培可 能期間を延長させるが,一般には生育期間の短縮,呼吸量の増加,高温ストレスの増加を招く.気 候変動にともなう作物生産変動は,これらの影響の相互作用を含めたバランスで決まり,そのバラ ンスは時期と地域によって異なる.気候変動にともなう作物生産変動予測では,これらの影響に関 する定性的な予測を精緻化し,不確実性を示しつつ定量化することが求められている.また,気候 変動に対する農業の適応技術の効果も含めた予測が必要となる. 気候変動にともなう作物生産変動予測は,作物統計や,実験室レベル・圃場スケールでの作物の 生理・生態反応に関する研究成果を利用して作物生産モデルを構築し,予想される気候変化シナリ オに基づいてモデル計算を実行して,将来の収量を予測することが基本となる.このモデル計算を 広域に展開するためには,モデル入力データとしての気象や土壌,品種,肥培管理に関するデータ の収集とデータセットの構築や,作物生産モデルに結合する微気象モデルや水収支モデル,窒素吸 収モデルなどの開発も必要となる.現在,農業環境技術研究所では,過去の研究の蓄積を生かし て,作物生産変動予測に関するさまざまな研究が行われている.本章では,「農業環境研究 20 年の 歩み(農業環境研究叢書第 16 号) 」との重複は避けつつ,地域スケールでの作物生産変動予測に関 するこの 10 年間の研究について紹介する.なお,気候変動に対する作物群落スケールでの応答や 適応戦略に関する研究については第 5 章で述べる. 2.農耕地観測地点における近年の気温変化の傾向 近年,異常高温や低日照など,年々の気象の変化の大きさに注目が集まっている(第 5 章参照) が,数十年という時間スケールで考えると,わが国の農耕地の気温はどの程度変化しているのだろ うか.これは,わが国で長年にわたって蓄積された作物統計データを解析するうえでも,また農耕 地の気温のモニタリングという観点からも重要な設問である.長期観測データから気温の永年変化 傾向を算出する際には,都市化の影響を除去する必要がある.気象庁は日本の平均気温の算出に, 統計の切断がなく都市化の影響も少ない気象官署 17 地点を用いている(JMA-17).しかし,気象 第 4 章 気候変動と作物生産変動予測 57 官署は小都市の場合でもその都心部にあ ることが多く,都市化の影響をすべて回 避できるわけではない.そこで,わが国 の農耕地における気温変化の実態を明 らかにするため,先行研究を参考にしつ つ,気象官署およびアメダス地点から, 農耕地における気温変化傾向の実態を 把握するのに有効な 19 地点(Rural-19) を 新 た に 選 定 し た( 西 森 ら,2009) . Rural-19 と大都市の気象官署(Metro-7) や JMA-17 を 比 較 す る と,1980 年 以 降 の都市化の影響は,日最低気温では春, 秋に,日最高気温では夏に顕著に表れて おり,Rural-19 の昇温トレンドは,日最 高気温では 2 月と 7 月に大きいことがわ かった(図 1) . 日最高・最低気温が観測所周辺の狭い 図 1 大都市(Metro-7),JMA-17 および Rural-19 観測地点 における,月別日最高(上図 )・日最低気温(下図)の 25 年間の変化率.1980 年から 2004 年までの線形トレ ンドを抽出した.●および▲は有意水準 99%,95%で トレンドが有意であることを表す(西森ら,2009). 領域の土地利用の影響を受けている可能 性や,水田などの存在による気候緩和効果にも留意して,今後も農耕地観測地点の気温の動向を注 視していく必要がある.また,農林水産省所管の独立行政法人試験研究機関や,農業関連の公設試 験研究機関の気象観測データの有効活用も検討すべきである.このうちの前者については,農林 水産研究情報総合センターが「農業研究気象観測ポータル」(http://www.affrc.go.jp/ja/info/scs/ ndb/agriresmet/index.html)を設置している. 3.気候変化シナリオの統計的ダウンスケーリング 気候変動の影響評価研究では,大気・海洋結合大循環モデル(CGCM)や領域気候モデル (RCM)などの気候モデルによる出力に基づいて作成された,気候変化シナリオを利用する.世界 各国の研究機関は,それぞれが開発した CGCM を用いて将来の気候のモデル計算を行っている. IPCC は,温室効果ガスの排出シナリオに関する特別報告(SRES)のなかで,A1B シナリオ(高 成長型社会シナリオ:各エネルギー源のバランス重視),A2 シナリオ(多元化社会シナリオ)な どの排出シナリオを用意しており,それぞれのシナリオに基づいて,将来の気候のモデル計算が行 われている.世界気候研究計画(WCRP)によるこれらの気候モデルの相互比較(第 3 次結合モデ ル相互比較プロジェクト,CMIP3)の結果が IPCC AR4 で採用され,CMIP3 マルチモデルデータ として公表されている. ところで,気候モデルの出力値を影響評価に利用するためには,気候モデルの出力値の時間・空 間解像度を上げ,バイアス(系統誤差)を補正する必要がある.統計的ダウンスケーリングは,気 候モデルの出力値と影響評価モデルに入力する気候データとの間の隔たりを埋め,影響評価のため の気候変化シナリオを作成するための手法である.その主な機能として,時間詳細化,空間詳細 化,要素推定,バイアス補正があげられる(飯泉ら,2010). 統計的ダウンスケーリングによる要素推定の例として,日射量の推定を紹介する.日射量は農業 影響評価に重要な要素だが,気候モデルからは出力されていなかった.そこで,多くの気候モデル 58 3.気候変化シナリオの統計的ダウンスケーリング で出力されており,観測値での検証が 比較的容易である気温日較差(日最高 気温と日最低気温との差)と相対湿度 という二つの要素から,全天日射量を 推定する統計モデルを構築した.この 統計モデルを気象研究所による RCM20 シナリオに適用し,温暖化後(2081 〜 2100 年 ) の 日 射 量 の 変 化 を 推 定 し た (Iizumi et al., 2008) . こ こ で,RCM20 シナリオとは,SRES-A2 シナリオに基 づく気象研究所の CGCM によるモデル 予測値を,同研究所が力学的ダウンス ケールの手法を用いて 20km メッシュに 展開したものである.温暖化後の日射 量の推定結果から,冬には北日本と東 日本で減少し,春の西日本と夏の東日 本で増加すること,特に夏の東日本で は増加率が現在の値に対し 10%以上と 大きいこと,秋には北海道を除いてや や減少することなど,温暖化によって 日射環境が変化する可能性が示唆され た(図 2) . 統計的ダウンスケーリングによる要 図 2 統計的ダウンスケーリング手法による将来のわが国の 全天日射量の推定値.1981 〜 2000 年の平均値に対す る,2081 〜 2100 年の平均値の増減を示す.赤色系は 日射量が増加,青色系は減少する地域を表す(西森, 2007). 素推定は,対象とする気候モデルの出 力要素値が他の要素に比べて精度の劣る場合にも適用される.特に,地点降水量は農業水資源予測 などの影響評価にとって重要な要素だが,気温や気圧に比べて気候モデルの出力値の精度が低い. そこで,比較的精度の高い気候モデル出力値である地上気圧を用いて,東アジア地域の 10km ス ケールの降水量を季節ごとに推定する手法を開発した(西森,2003).この研究は,月降水量や豪 雨変化の指標となる降水日数,降水強度の将来予測へと発展している. 統計的ダウンスケーリングの研究と並行して,農業環境技術研究所では影響評価研究での利用を 想定して,日本域の空間解像度 10km の「気候変化メッシュデータ」を作成,配布した.このデー タセットは,4 つの CGCM の出力値に対し,距離の逆数による重み付け平均による線形補間を行っ たものである(Yokozawa et al., 2003) .その改訂版である「気候変化メッシュデータ Ver. 2」は, CMIP3 マルチモデルデータセットを 10km メッシュ(高解像度モデルでは 1km メッシュ)に線形 補間したものであり, SRES-A1B と A2 シナリオを想定した 7 モデルによる計 9 シナリオに基づく. このメッシュデータには,日平均・最高・最低地上気温,日降水量,日積算日射量の 5 要素の日別 値が含まれ,データの期間は 1981 〜 2000 年, 2046 〜 2065 年,2081 〜 2100 年の 3 期間である(Okada et al., 2009) . 第 4 章 気候変動と作物生産変動予測 59 4.日本およびアジアの主要生産地のコメ収量変動予測 ⑴ わが国の将来のコメ収量変動予測 わが国の将来のコメ収量の変動予測は,行政ニーズも,また社会的関心も高い研究テーマであ る.利用可能な統計資料,土地利用分布ならびに気象メッシュデータを用いて,わが国のコメ収 量を都道府県別に推定するモデル(Process-based Regional rice Yield Simulator with Bayesian Inference,PRYSBI)を作成した(Iizumi et al., 2009).このモデルは収量の年々変動を再現し, モデルに内在する不確実性も考慮できる.PRYSBI に「気候変化メッシュデータ Ver.2」(第 3 節参 照)の気候変化シナリオを入力して,1981 〜 2000 年,2046 〜 2065 年,2081 〜 2100 年の 3 つの 期間について,各都道府県の年々の収量を予測した.4 つの地域ごとに各年の平均コメ収量を推計 して,各期間の 20 年間平均値および変動係数と,暖候期(5 月〜 10 月)平均気温の 1981 〜 2000 年に対する上昇度との関係を図 3 に示す.この図には,排出シナリオ,気候モデルおよび予測対象 期間の違いによる,全部で 34 ケースの推計結果がプロットされている. 図 3 気候変化がわが国の地域別のコメ収量に及ぼす影響.地域別平均収量(上図)および平均収量の変動係 数(下図)と,暖候期(5 月〜 10 月)平均気温の 1981 〜 2000 年に対する上昇度との関係を示す.横軸 は暖候期の平均気温の上昇度だけが示されているが,実際には日別の気温および日射の変動が含まれて いる.降水量の変化は考慮されていない.NJ:北海道・東北,EJ:関東・甲信越・北陸,CJ:東海・中 部・近畿,WJ:中国・四国・九州(横沢ら,2009). 北海道・東北地域では,気温上昇(気候変化)にともなってコメ収量は増加し,他の地域では収 量はおおむね減少すると推計された.ただし,3℃付近までの気温上昇では,コメ収量は全国で現 在と同程度か,やや増加する.北日本の収量増加は,おもに低温影響(冷害)による収量低下の発 生確率が減少することに起因する.地域平均コメ収量の年々変動の大きさを表す変動係数は,どの 地域においても気温上昇(気候変化)にともなって増大する傾向が見られる.とりわけ東海・中 部・近畿地域の変動係数の増加傾向が最も大きい.この原因は,開花期における平均日最高気温が 33℃以上になると高温による登熟障害が発生し,収穫係数の値が急激に(指数関数的に)減少する 機構をモデルに組み込んだためである.特に東海から中部,関西にかけて,気候変化にともなって 太平洋高気圧が強まり,その端がこの地域にかかることにより,夏季の高温傾向が強まることが気 候モデルの総観場として見られる.これは近年にも見られる特徴であり,将来,これが当該地域で の収量低下および変動の増幅のさらなる原因となる可能性がある.ただし,高温に対するコメ収量 60 4.日本およびアジアの主要生産地のコメ収量変動予測 の応答の不確実性はまだ大きく,作物生理的な側面から研究が進められている. 以上の推計結果は適応策の効果を含んでいないが,移植日の移動,および高温耐性品種の導入を 仮定したシミュレーションの結果によれば,各地域とも,気温上昇(気候変化)に対して適応策は 有効という結果が得られた.とりわけ,北・東日本では移植日の移動,西・南日本では高温耐性品 種の導入が最も効果的であることが示唆された(横沢ら,2009). ⑵ メコンデルタのコメ収量変動予測 ちゅうみつ IPCC AR4 では,南アジア,東アジアおよび東南アジアの人口が 稠 密なメガデルタ地帯は,海 からの洪水の増加によって,またいくつかのメガデルタでは河川の洪水によって,最大のリスクに 直面する,とされている.メコンデルタは,世界第 2 位のコメ輸出国であるベトナムのコメ生産の 約半分,輸出米の 90%を生産する穀倉地帯であるが,海抜 2m 以下の地域が多く,温暖化にともな そじょう う海面上昇などによって塩水遡上規模が拡大し,コメ生産や世界市場へのコメ供給に甚大な影響が かんがい 懸念される.メコンデルタにおけるコメ生産変動を評価するため,潅漑水の水理過程と水稲生育・ コメ収量推計のモデルを組み合わせた統合モデルを作成した.作付け暦は水田の水条件から生育可 能期間と播種可能期間を判定し,イネの生育や水田作業期間の配置を考慮して決定した.衛星デー タに基づいた生育期間推定結果と照合することにより,モデルの妥当性を検証した(Sakamoto et al., 2006) .水田水収支は,降雨や蒸発散量等に加え,イネ生育段階に応じた潅漑・排水の水管理の 影響を考慮して推定した.水路の塩分濃度が高い期間の潅漑や洪水期における高水位時の排水は行 えないとした.気候モデル MIROC3.2hires による SRES-A1B シナリオに基づく気候変化をこのモ デルに適用し,ベトナム領内のメコンデルタの小水域ごとに,作付け暦,収穫面積および収量の変 化を予測した.その結果,洪水年には上流部で,少雨(塩水遡上)年には沿岸部で,収穫可能面積 が大きく減少したが,同時にその他の地域では作付け可能面積が増加する相反関係が見られた.ま た,空間分布は変動するものの,デルタ全体の総収穫面積の減少は緩和されることがわかった.一 方,気温上昇による収量への影響は敏感であり,2020 年代後半におけるベトナム・メコンデルタ のコメ生産量は,現在よりも約 11%低下する可能性が示された(Kotera et al., 2010). また,気候変化による海面上昇が引き起こす塩水遡上がベトナム・メコンデルタのコメ生産へ及 ぼす影響を明らかにするため,メコン河の流量変化が引き起こす塩水遡上ならびに洪水の影響を同 時に評価する水理モデルを開発した.将来(2090 年代)の気候変化シナリオとして SRES-B2 シナ リオ(地域共存型社会シナリオ)に基づく気候変化を想定し,デルタ内の気温,降水量,日射量, 海面の上昇度,およびデルタ上流部におけるメコン河の流量変化を与え,2090 年代半ばにおける 12 月から翌年 6 月までの期間の流量と海水侵入の程度を推定した.その結果,基準年(1998 〜 2006 年)よりも将来(2090 年代)の方が,洪水の水位と浸水面積がはるかに大きいことがわかっ た.将来のコメ栽培期間の大幅な変化は,主として上流の流量増加にともなう洪水期間の長期化に よるものであり,三期作が可能な地域の面積が 31%から 5%に減少する一方で,一期のみ作付け可 能な地域の面積は 21%から 62%へ増加すると推定された(Khang et al., 2010,図 4). 第 4 章 気候変動と作物生産変動予測 61 図 4 メコンデルタの塩水遡上と洪水に対応してコメの作付けが可能な地域.左図は基準年(1998 〜 2006 年) ,右図は 2090 年代(Khang et al., 2010). ⑶ 東北タイの天水田地帯のコメ収量と水稲作付け面積の推定 世界のコメ生産面積の約 1/3 を占める天水田の稲作は,植え付け時期や栽培期間が降雨に強く依 存するため,今後予想される気候変化や水循環変動による影響が懸念されている.そこで,天水田 が多く分布する東北タイを対象に,降水量などの気象要素,作物の水・窒素利用から,天水田地帯 すう せい における水稲の地域収量の趨勢と変動を合理的に説明,予測するモデルを構築した.開発したモ デルは,降水パターンなどに依存する稲作暦を推定する部分と,水稲の成長・収量を予測する部 分からなる.稲作暦は,農家聞き取り調査に基づいて,3 ヶ月に及ぶ移植日の地域内分布(移植水 田割合)を降水パターンからモデル化し,移植日ごとの生育日数は日長と気温の関数で再現できた (Sawano et al., 2008,図 5) .水稲の収量予測については,作物の水利用に比例した乾物生産と収 穫指数の積からなる一般的な簡易収量モデルに,国際連合食糧農業機関(FAO)の統計資料から 容易に入手できる投入窒素の影響を取り入れたモデルを開発した.地域収量は,移植日ごとに計算 した収量を日々の移植水田割合で重み付けして求めた.このように地域内の移植日の分布を考慮す ることで,東北タイの県別収量の地理的,時間的変動をよく説明することができた.また,投入窒 実測値 推定値の幅 ( ) 6月1日からの積算降水量(mm) 移植日 図 5 東北タイにおける移植水田割合と 6 月 1 日からの積算降水量との関係(左図),および移植日と出穂まで の日数との関係(右図) .実測値は 2004 年と 2005 年に実施した農家聴取調査による.右図の推定値の幅 は,11 気象観測地点のデータと温度・日長の発育関数を用いて算出した. 62 4.日本およびアジアの主要生産地のコメ収量変動予測 素の影響を導入することにより,過去 25 年間 300 の県別収量の増加趨勢を再現できるようになっ Nong Khai た(図 6) .開発した地域収量予測モデルは, 200 気候シナリオと組み合わせることによって,将 ㊂ 乿(䋯䋛 来の収量変動の予測に利用できる. また,こ のモデルは潅漑地域においても適用可能である (Hasegawa et al., 2008) (平成 19 年度研究成 果情報,長谷川ら) . 気候変化にともなう地表面水循環過程の変化 ) が天水田地帯の水稲生産に与える影響を評価す 100 ⛔⸘୯ -2 RMSE1=79 g m ផቯ୯ Case1 -2 RMSE2=21 g m ផቯ୯ Case2 0 Khon Kaen 300 -2 RMSE1=79 g m -2 RMSE2=21 g m るため,一般的な気象データから東北タイの広 200 域水収支を算定するモデルを構築し,モデルか ら得られる地表面飽和面積を使用した水稲作付 100 け面積推定を行った.構築したモデルは 0.05°× 0.05°(約 5km × 5km)の解像度単位で水収支 0 を算定する分布型水文モデルであり,地形や土 1975 1980 1985 1990 1995 2000 ᐕ ᰴ 壌物理性の情報と,日別の気温,降水量,日射 量,湿度,風速から,蒸発散量や層別の土壌水 図 6 東北タイ収量の経年変化の推定(2 県の結果を 例示).Case 1 は移植日の幅を考慮せず,積算 降水量が一定値に達したときに移植されたと 仮定した場合の推定収量,Case2 は本モデル による推定収量.RMSE は標準推定誤差. 分量,地下水位等を計算する.その際,グリッ ド内の詳細な地形情報を指標化することによ り,グリッド内の土壌水分の不均一性や地表面 飽和領域の面積を算定する(図 7). 䂹䉫䊥䉾䊄䉕䉰䊑䉫䊥䉾䊄䈮ಽഀ䈜䉎 䊝䊂䊦䈮䉋䉎᳓ᡰ⸘▚䈱 ၮᧄන䈪䈅䉎䉫䊥䉾䊄䋨ฝ ࿑㤥ᨒ䋩䉕䇮䋶㬍䋶䈱䉰䊑䉫 䊥䉾䊄䋨ฝ࿑㕍ᨒ䋩䈮ಽഀ䈜䉎䇯 䉫䊥䉾䊄䋨㤥ᨒ䋩 䋺 㪇㪅㪇㪌㫰㬍㪇㪅㪇㪌㫰 䋨㪈㪏㪇㫊㪼㪺㬍㪈㪏㪇㫊㪼㪺䋩 䉰䊑䉫䊥䉾䊄䋨㕍ᨒ䋩 䋺 㪊㪇㫊㪼㪺㬍㪊㪇㫊㪼㪺 㪈㪍㪅㪇 㪈㪎㪅㪈 㪈㪍㪅㪌 㪈㪌㪅㪊 㪈㪌㪅㪌 㪈㪌㪅㪎 㘻㕙Ⓧ₸ (UCVߩផቯ 㪈㪋㪅㪏 㪈㪍㪅㪐 㪈㪌㪅㪍 㪈㪌㪅㪍 㪈㪌㪅㪋 㪈㪌㪅㪏 㪈㪌㪅㪊 㪈㪋㪅㪐 㪈㪌㪅㪌 㪈㪍㪅㪉 㪈㪍㪅㪉 㪈㪋㪅㪊 㪈㪌㪅㪊 㪈㪍㪅㪈 㪈㪋㪅㪌 㪈㪌㪅㪉 㪈㪌㪅㪎 㪈㪋㪅㪋 㪈㪋㪅㪐 㪈㪌㪅㪍 㪈㪋㪅㪋 㪈㪍㪅㪈 㪈㪎㪅㪇 㪈㪋㪅㪊 㪈㪋㪅㪐 㪈㪌㪅㪉 㪈㪋㪅㪊 㪈㪍㪅㪊 㪈㪌㪅㪏 㪈㪋㪅㪉 䂹ᮡ㜞䊂䊷䉺䈎䉌䇮ᒻ࿃ሶ㱗䉕⸘▚䈜䉎 ᮡ㜞䊂䊷䉺䉕↪䈚䈩䇮એਅ䈪ቯ⟵䈘䉏䉎ᒻ࿃ሶ 䋨㱗䋩䉕䉰䊑䉫䊥䉾䊄Ფ䈮᳞䉄䉎䋨࿑䈱ᢙ୯䋩䇯 O A ln l tan E A : ᵹၞ㕙Ⓧ l : ╬㜞✢㐳 E: ൨㈩ 䂹㘻㗔ၞ䉕䈜䉎 ᳓ᡰ⸘▚䈪ᓧ䉌䉏䉎䉫䊥䉾䊄ᐔဋਅ᳓䈎䉌䇮એਅ 䈱᧦ઙ䉕ḩ䈢䈜䉰䊑䉫䊥䉾䊄䉕㘻㗔ၞ䈫䈚䈩䈜䉎䇯 O t O f ZT O : 㱗䈱䉫䊥䉾䊄ᐔဋ ZT 0.6 ( m) 㪈㪍㪅㪇 㪈㪎㪅㪈 㪈㪋㪅㪏 㪈㪍㪅㪐 㪈㪌㪅㪍 㪈㪌㪅㪍 㪈㪍㪅㪌 㪈㪌㪅㪊 㪈㪌㪅㪋 㪈㪌㪅㪏 㪈㪌㪅㪊 㪈㪋㪅㪐 㪈㪌㪅㪌 㪈㪌㪅㪎 㪈㪌㪅㪌 㪈㪍㪅㪉 㪈㪍㪅㪉 㪈㪋㪅㪊 㪈㪌㪅㪊 㪈㪍㪅㪈 㪈㪋㪅㪌 㪈㪌㪅㪉 㪈㪌㪅㪎 㪈㪋㪅㪋 㪈㪋㪅㪐 㪈㪌㪅㪍 㪈㪋㪅㪋 㪈㪍㪅㪈 㪈㪎㪅㪇 㪈㪋㪅㪊 㪈㪋㪅㪐 㪈㪌㪅㪉 㪈㪋㪅㪊 㪈㪍㪅㪊 㪈㪌㪅㪏 㪈㪋㪅㪉 㪈㪍㪅㪇 㪈㪎㪅㪈 㪈㪋㪅㪏 㪈㪍㪅㪐 㪈㪌㪅㪍 㪈㪌㪅㪍 ZT 0.3 (m) 㪈㪍㪅㪌 㪈㪌㪅㪊 㪈㪌㪅㪋 㪈㪌㪅㪏 㪈㪌㪅㪊 㪈㪋㪅㪐 㪈㪌㪅㪌 㪈㪌㪅㪎 㪈㪌㪅㪌 㪈㪍㪅㪉 㪈㪍㪅㪉 㪈㪋㪅㪊 Fsat 0.28 㪈㪌㪅㪊 㪈㪍㪅㪈 㪈㪋㪅㪌 㪈㪌㪅㪉 㪈㪌㪅㪎 㪈㪋㪅㪋 㪈㪋㪅㪐 㪈㪌㪅㪍 㪈㪋㪅㪋 㪈㪍㪅㪈 㪈㪎㪅㪇 㪈㪋㪅㪊 㪈㪋㪅㪐 㪈㪌㪅㪉 㪈㪋㪅㪊 㪈㪍㪅㪊 㪈㪌㪅㪏 㪈㪋㪅㪉 㪈㪍㪅㪇 㪈㪎㪅㪈 㪈㪋㪅㪏 㪈㪍㪅㪐 㪈㪌㪅㪍 㪈㪌㪅㪍 ZT 㪈㪍㪅㪌 㪈㪌㪅㪊 㪈㪌㪅㪋 㪈㪌㪅㪏 㪈㪌㪅㪊 㪈㪋㪅㪐 㪈㪌㪅㪌 㪈㪌㪅㪎 㪈㪌㪅㪌 㪈㪍㪅㪉 㪈㪍㪅㪉 㪈㪋㪅㪊 Fsat 㪈㪌㪅㪊 㪈㪍㪅㪈 㪈㪋㪅㪌 㪈㪌㪅㪉 㪈㪌㪅㪎 㪈㪋㪅㪋 㪈㪋㪅㪐 㪈㪌㪅㪍 㪈㪋㪅㪋 㪈㪍㪅㪈 㪈㪎㪅㪇 㪈㪋㪅㪊 㪈㪋㪅㪐 㪈㪌㪅㪉 㪈㪋㪅㪊 㪈㪍㪅㪊 㪈㪌㪅㪏 㪈㪋㪅㪉 Fsat 0.08 0 ( m) 0.5 ZT : 䉫䊥䉾䊄ᐔဋਅ᳓ f : 䊌䊤䊜䊷䉺䋨=1.8䋩 図 7 水稲作付け面積推定のための地表面飽和領域の面積推定方法.地形因子λは,谷筋や盆地の底などの水 がたまりやすい場所ほど値が高くなる.モデルで計算されるグリッド内の地下水位が浅くなれば,飽和 しているサブグリッドの数が増加し,飽和領域が大きくなる. 第 4 章 気候変動と作物生産変動予測 63 1979 年から 2005 年までの期間について,このモデルで推定した東北タイの天水田地帯の飽和面 積を旬ごとに集計し,統計資料から得た各年の作付け面積と比較した結果,8 月中旬の飽和面積と 作付け面積の相関が最も高くなることがわかった.これは,この地域では日長感応性が強い品種 が作付けされており,水稲作付けは遅くとも 8 月末ごろまでに実施する必要があることを反映して いると考えられ,この時点での飽和面積から水稲作付け面積を適切に推定できる可能性が示され た(Ishigooka et al., 2009) (平成 22 年度研究成果情報,石郷岡ら).また,推定される水稲作付け 面積を上記の地域収量予測モデルと組み合わせることにより,この地域のコメ生産の予測が可能と なった. ⑷ 中国におけるコメ生産量変動の確率的リスク評価 気候変化の影響評価で問題となる,気候モデルと温室効果ガス排出シナリオの不確実さに対処す る一つの方法として,確率的なリスク評価手法を開発し,世界のコメの作付け面積の約 4 分の 1, 収量の 3 分の 1 以上を占める中国を対象に,気候変化がコメ収量に及ぼす影響を評価した.4 つの SRES 排出シナリオと 5 つの気候モデルの組み合わせによる,計 20 の気候変化シナリオの結果を 利用し,世界の平均気温が 1961 〜 1990 年レベルに比べて,それぞれ 1,2,3℃上昇する場合につ いて,各観測点の気候変化を確率的に予測し,イネの収量を予測した.潅漑と施肥は将来の気候に 適応させ,品種は現在の栽培種に固定した.中国の稲作地帯 6 地点(9 作期)での予測結果をまと めると,二酸化炭素の施肥効果を考慮しない場合は,世界の平均気温が 1℃上昇した場合でも 90% の確率で減収となり,減収率は 6.1 〜 18.6%で,蒸発散量と潅漑用水量は多くの場合で低下した. また,二酸化炭素の施肥効果を考慮すると減収率は小さくなるが,蒸発散量と潅漑用水量の低下は より高い確率で起こり,低下の程度も大きくなると予測された.このような確率的評価手法は,ど の程度の気候変化が食糧安全保障上重要かを判断するのに有効である(Tao et al., 2008). ⑸ 東アジア広域のコメ収量変動予測 東北タイの天水田用に開発した地域収量推定モデルに,作物による窒素の吸収および水田におけ る窒素の流れを明示的に取り扱うサブモデルを組み込んだ.この改良モデルを 20 世紀後半に収量 の著しい増加がみられた中国の 9 つの省に適用し,1990 年以降の収量や投入窒素量当たりの収量 の地域間差をよく再現できることを確認した.また,窒素肥料投入量,二酸化炭素濃度,気温の各 要因が 1961 年以降の長期的な増収に与えた影響を解析し,1961 年以降の増収のほとんどは窒素肥 料投入量の変化で説明できることを明らかにした(澤野ら,2009).現在,この地域収量推定モデ ルと,CMIP3 マルチモデルデータセット(SRES-A1B シナリオ)に基づく気候変化シナリオを, 中国東北部から東南アジアまでの 26 地点に適用し,2050 年までの収量予測を行っている.最新の 結果によれば,気候変化シナリオ間のばらつきは大きいが,全シナリオによる予測結果を地点ごと に平均した収量変化は, 1990 〜 1999 年と比べて−10 〜+10%であった.また,緯度帯別にみると, 低緯度地域および高緯度地域が増収傾向,中緯度地域では減少傾向を示し,既往の研究とは異なる 興味深い予測結果が得られている(Sawano et al,. 2009). 64 5. 農業水利用を考慮した大陸スケールの水循環モデルの開発 5. 農業水利用を考慮した大陸スケールの水循環モデルの開発 気候変動にともなう水循環の変動 は,世界の食糧生産に関わる大きな 問題を引き起こす可能性がある. そこで,乾燥地・半乾燥地を多く含 み,農業の水資源への依存度が大き いユーラシア大陸東部を対象に,広 域の気象情報から農業における水需 給の時間的・空間的特徴を明らかに できる,大陸スケールの水循環モデ ルを開発した.鉛直方向の水収支を 小流域ごとに計算するための「流域 流出モデル」と,各々の小流域から の流出量を流路網に従い下流へ流下 させる「河道流下モデル」を結合す ることによって,広域の水循環と河 道から農耕地への潅漑水の供給を含 む,農業的な水循環の再現が可能と なった(図 8) .各小流域を土地被 覆の種類によってサブ領域(水体, 潅漑水田,非潅漑水田,潅漑畑地, 非潅漑畑地,草地,森林,裸地)に 分割し,サブ領域ごとに人工衛星 データから得られる植生の季節変化 図 8 ユーラシア大陸東部を対象とした大陸スケールの水循環推 定モデルの概念図 (mm/mm) 2.0 1.5 1.0 0.5 を考慮して水収支を計算する.本モ デルでは,潅漑要水量(作物が水分 の不足によるストレスなしに生育で きるために必要な水の量)と,農耕 0 図 9 東アジアの小流域ごとの供給可能水資源量に対する灌漑要 水量の割合の分布.黒色の領域は農耕地以外の土地利用 地に供給可能な水資源量を計算でき る. 長期間の広域気象データを使用してモデルを実行し,潅漑要水量と供給可能水資源量の 30 年間 (1961 〜 1990 年)平均値を求めた.供給可能水資源量に対する潅漑要水量の割合は,農業におけ る水需給関係を表す指標となる(図 9) .この値が高い地域は,農耕地が集中して分布し降水量が 比較的少ない中央アジア,パキスタン,インド西部,中国華北などに分布している.この値が 1 を 超え,作物の生育に必要な最低限の水が,その地域に存在する水資源のみではまかなえない地域が 存在することが明らかになった.このモデルと将来の気候変化シナリオを用いて,温暖化などの気 候変化が農業の水需給関係に与える影響を評価することもできる.そのためには,空間的に高解像 度な気象データと,農耕地分布や作付け期間に関する正確な地理情報が必要である(平成 16 年度 研究成果情報,石郷岡ら) . 第 4 章 気候変動と作物生産変動予測 65 6. 今後の展望 作物生産変動予測の研究は,個人が構築した作物生育・収量モデルを用いて,環境要因に対する 感度解析を行う時代から,複数の専門家が協力して構築した結合モデルを用いて,現実的な気候変 化シナリオに基づいて,広域を対象に収量予測を行う時代に移行しつつある.これまで個別に行わ れてきた感のある作物生態反応,群落微気象,農地の水収支や窒素循環,病虫害予測などの研究 や,環境操作実験,モデル , モニタリング,データベースなどの研究手法・ツールが,気候変動に ともなう作物生産変動の予測という明確な目標に向けて連携を強めることは,農業環境研究で求め られる研究の連携・総合化のモデルケースとなるだろう.現在のところ,予測に不可欠な気候変化 シナリオに内在する不確実さは受け入れざるを得ず,予測対象領域の広域化にともなうモデル入力 データの収集の困難さなどのハードルも多いが,多くの専門家の英知を結集して,予測の不確実さ を少しでも低減することが求められている. 引用文献 1)Hasegawa, T., Sawano, S., Goto, S., Konghakote, P., Polthanee, A., Ishigooka, Y., Kuwagata, T., Toritani, H. and Furuya, J.(2008)A model driven by crop water use and nitrogen supply for simulating changes in the regional yield of rain-fed lowland rice in Northeast Thailand. Paddy and Water Environment, 6 (1) , 73-82 2)Iizumi, T., Nishimori, M. and Yokozawa, M.(2008)Combined equations for estimating global solar radiation: Projection of radiation field over Japan under global warming conditions by statistical downscaling. Journal of Agricultural Meteorology, 64(1),9-23 3)Iizumi, T., Yokozawa, M. and Nishimori, M.(2009)Parameter estimation and uncertainty analysis of a large-scale crop model for paddy rice: Application of a Bayesian approach. Agricultural and Forest Meteorology, 149, 333-348 4)飯泉仁之直,西森基貴,石郷岡康史,横沢正幸(2010) 統計的ダウンスケーリングによる気 候変化シナリオ作成入門.農業気象,66 (2) , 131-143 5)Ishigooka, Y., Kuwagata, T., Goto, S., Toritani, H., Ohno, H. and Urano, S.(2008)Modeling of continental-scale crop water requirement and available water resources. Paddy and Water Environment, 6 (1) , 55-71 6)Ishigooka, Y., Kuwagata, T., Goto, S., Toritani, H., Hasegawa, T., Sawano, S. and Ohno, H. (2010)Estimation of water saturated areas in Northeast Thailand using a large-scale water balance model. Journal of Agricultural Meteorology, 66(2),91-101 7)Khang, N.D., Kotera, A., Iizumi, T., Sakamoto, T. and Yokozawa, M.(2010)Variations in water resources in the Vietnamese Mekong Delta in response to climate change and their impacts on rice production. Journal of Agricultural Meteorology, 66(1),11-21 8)Kotera, A., Khang, N.D., Sakamoto, T., Iizumi, T. and Yokozawa, M.(2010)Modeling rice cropping schedules in the Vietnam Mekong Delta for adapting to changes in flooding, salinity intrusion and monsoon rains. Climata Adaptation Futures. Conference Handbook of the 2010 International Climate Change Adaptation Conference, held in Gold Coast, Australia, 29 June-1 July, 2010, 191 66 引用文献 9)西森基貴(2003)統計的ダウンスケーリングによる 10km スケールの降水量予測.文部科学省 科学技術振興調整費プロジェクト「21 世紀のアジア水資源変動予測」研究成果の詳細報告, 94-103 10)西森基貴(2007)気候モデルによる温暖化予測とそのダウンスケーリング ― 身近な地域への 影響を知るために ― .第 28 回農業環境シンポジウム,東京,2007 年 12 月,15-18 11)西森基貴,桑形恒男,石郷岡康史,村上雅則(2009)都市化の影響を考慮した近年の日本にお ける気温変化傾向とその地域的・季節的な特性について.農業気象,65(2),221-227 12)Okada, M., Iizumi, T., Nishimori, M. and Yokozawa, M.(2009)Mesh climate change data of Japan Ver. 2 for climate change impact assessments under IPCC SRES A1B and A2. Journal of Agricultural Meteorology, 65 (1) , 97-109 13)Sakamoto, T., Nguyen, N.V., Ohno, H., Ishitsuka, N. and Yokozawa, M.(2006)Spatio-temporal distribution of rice phenology and cropping systems in the Mekong Delta with special reference to the seasonal water flow of the Mekong and Bassac rivers. Remote Sensing of Environment, 100, 1-16 14)Sawano, S., Hasegawa, T., Goto, S., Konghakote, P., Polthanee, A., Ishigooka, Y., Kuwagata, T. and Toritani, H.(2008)Modeling the dependence of the crop calendar for rain-fed rice on precipitation in Northeast Thailand. Paddy and Water Environment, 6(1),83-90 15)澤野真治 , 長谷川利拡 , 石郷岡康史 , 桑形恒男 , 新藤純子(2009)気候変化と栽培技術要因を考 慮したイネの地域収量予測モデルの開発と 20 世紀後半の収量趨勢に関するモデル解析.日本 作物学会紀事別号,228, 228-229 16)Sawano, S., Hasegawa, T., Kuwagata, T., Ishigooka, Y. and Shindo, J.(2009)Evaluating the effect of uncertainty in the climate scenario on future prediction of rice production in Asia. In: Hasegawa, T. and Sakai, H.(eds.)Crop Production under Heat Stress: Monitoring, Impact Assessment and Adaptation. Proceedings of the MARCO Symposium 2009 held in Tsukuba, Japan, 5-9 October 2009, 77 17)Tao, F., Hayashi, Y., Zhang, Z., Sakamoto, T. and Yokozawa, M.(2008)Global warming, rice production, and water use in China: Developing a probabilistic assessment. Agricultural and Forest Meteorology, 148, 94-110 18)Yokozawa, M., Goto, S., Hayashi, Y. and Seino, H.(2003)Mesh climate change data for evaluating climate change impacts in Japan under gradually increasing atmospheric CO2 concentration. Journal of Agricultural Meteorology, 59, 117-130 19)横沢正幸,飯泉仁之直,岡田将誌(2009)気候変化がわが国におけるコメ収量変動に及ぼす影 響の広域評価.地球環境,14 (2) ,199-206 67 第 5 章 イネ・水田生態系の応答と適応戦略 1.はじめに 大気環境が水稲作に及ぼす要因として,地温・水温を含む気温,CO2 濃度,降水量および日射量 などがあり,これまで冷害や日射量不足への対応について研究が進められてきたが,近年,地球温 暖化にともなう気候変動の影響が水稲の収量・品質に及ぼしていることが確認されている.将来, CO2 濃度の上昇にともなう地球温暖化が進んだ場合,CO2 濃度や気温上昇や降雨量変動の増大など により,これまでに確立されてきた品種や栽培方法の転換を行うことが必要になると予想される. そのための,基礎的なデータ収集,影響解析およびモデルによる将来予測などが求められている. 2.温暖化影響の把握 ⑴ 2007 年の高温 2007 年 8 月には,熊谷,多治見で観測史上最高の 40.9℃を記録するなど,広い範囲で異常高温 に見舞われた.これまで多くの室内実験から,水稲の開花時の気温が 35℃を超えると,受精障害 もみ により不稔籾が多発することが知られている.2007 年夏に記録された異常高温は,これまで顕在 化していなかった高温不稔を誘発しうる温度域であり,被害発生が懸念された.そこで,記録的な 猛暑を観測した群馬県,埼玉県,茨城県,岐阜県,愛知県において,7 月下旬から 8 月下旬までに 出穂した 132 の水田を対象に不稔籾の発生を調査した. その結果,関東および東海の両地域で出穂・開花時期の最高気温が 35℃を越えた水田があり, 通常は約 5%程度の不稔率が 10%を超えた水田が認められた.しかし,調査対象田における不稔率 は,これまでの室内実験結果から予測される値よりも低い傾向にあった.これは,水稲の開花時間 帯(午前 10 〜 12 時頃)の穂温は,記録された最高気温よりも低かったと推定されること(図 1), また地域全体では出穂・開花の時期に高温に遭遇した水稲が少なかったことなどによると考えられ た(長谷川ら,2009) . (平成 20 年度研究成果情報,長谷川ら). 図 1 2007 年 8 月 16 日の最高気温の分布(左)と穂温推定モデルによる同日午前 10 〜 12 時 (開花時間帯)の推定穂温の分布(右) 68 3.温度,CO2 濃度上昇の水稲への影響解析 ⑵ 2009 年の低日照 2009 年の夏は,北日本日本海側地域の日照時間が 1946 年以降で最も少ないなど,ほぼ全国的に 日照の少ない夏であった.冷夏年だった 1980,1993,2003 年と異なり,2009 年のような冷夏をと もなわない全国的な低日照年の作況解析は,今後の気候変動の影響を評価する上でたいへん重要で ある.そこで当研究所が開発した「モデル結合型作物気象データベース:MeteoCrop DB」(平成 21 年度研究成果情報,桑形ら)を用いて, 2009 年の気象状況が水稲の作況に及ぼす影響を解析し, 過去の事例と比較した. その結果,2009 年は北海道北部で障害型冷害が発生する気温冷却量となったものの,全国的に はそれほどの冷却量ではなかったが,全天日射量は日本海側で少なかったことが明らかとなった (図 2) .また,日本海側では数年に一度の割合で低日照となるなど,気温と日照時間の対応関係は 低日照側に移動し,過去 30 年にわたり,徐々に「高温または低日照の夏」の傾向が進んでいるこ とがわかった(西森ら,2009) .このように,日本のコメ生産において,今後,日照不足による地 域的な作況低下が心配される(平成 21 年度研究成果情報,西森ら). 図 2 2009 年の水稲作況(左)および MeteoCrop を用いて推定した 2009 年(中)と 2003 年の 7 月日射量の平 年偏差の地域分布 3.温度,CO2 濃度上昇の水稲への影響解析 ⑴ 周年加温装置の開発 地球温暖化の影響は水稲の作付け期間である夏季だけでなく,冬季の非作付け期間の温度上昇に よる土壌水分や土壌微生物活性の変化を通じて年間の炭素・窒素収支を変え,長期的に夏季の水稲 生育環境や温室効果ガス動態に影響を及ぼす可能性がある.これらの長期的な応答を明らかにする ためには年間を通じて,屋外の水田生態系を温暖化できる装置が必要である.このため,周年開放 系温暖化実験システムを世界で初めて開発した. 農業環境技術研究所内の水田に,4 反復の温暖化区と対照区を設け(各 4 × 5m),夏季の水稲生 育期間は,電気温床線を畝間の水中に設置し,温暖化区の水温が対照区より 2℃高くなるように, 温度調節計を用いて自動的に制御した.水温について設定通りの制御ができ,水稲の根圏の地温も 加温できた.また,冬季の畑状態の非作付け期間は,電気温床線による制御が困難なため,赤外放 射反射シートを用いて夜間の放射冷却を抑制することにより夜間の地温を上昇させる装置を開発し た.シートの展張と巻き取りはモーター付き巻取機をデータロガーで操作することにより自動的に 第 5 章 イネ・水田生態系の応答と適応戦略 69 行う.この温暖化処理により,夜間 の地表面温度は対照区より 1 〜 4℃ 高く維持できた.特に,深さ 10cm では夜間のシート被覆による保温効 果が昼間まで残っており,土壌が安 定的に温暖化されていた. 開発された周年温暖化実験水田で 水稲の出穂日を調べたところ,温暖 化に対する出穂時期の反応に品種間 で違いがあることなど,水稲の生育 への温暖化影響について有益な成果 が得られつつある. (平成 19 年度研 究成果情報,吉本ら) ⑵ FACE(開放系大気 CO2 増加)実験による収量・ 品質への影響 図 3 夏季の水田温暖化実験システムと水温・地温の変化 温室やチャンバーを用いた従来 の実験結果から,高 CO2 濃度下では光合成速度が高まり水利用効率も上昇するため,農作物の生 長と収量は増加すると予想される.しかし,従来の実験ではチャンバー内の環境が現実のほ場と は異なるため,高 CO2 濃度に対する 植物の応答も異なる可能性が高い. そこで,水稲を対象とした世界最初 の FACE(開放系大気 CO2 増加)実 験により,岩手県雫石町の実際の農家 水田におけるイネの生長,収量や品質 に及ぼす CO2 濃度上昇の影響を明ら かにすることに取り組んだ(小林, 2001) .すなわち,1998 〜 2000 年の 3 年間,CO2 濃度 2 水準(外気,外気+ 200 〜 250ppm)×窒素肥料 3 水準(多 窒素,標準窒素,少窒素)で実験を行 い,水稲(あきたこまち)の生長,収 量,品質を調べた. 高 CO2 濃度下では水稲の乾物生長 が促進され,収穫時の全乾物重は高 CO2 濃度で 11 〜 13%多かった.米収 量は,多窒素と標準窒素では高 CO2 濃度により約 15%増えたが,少窒素 では 7%の増収に止まった(図 3) .高 CO2 濃度による増収は,面積当たりの モミ数が増加したためだったが,それ 図 4 大気 CO2 濃度上昇によるモミ収量(上),モミ数(中) および白米タンパク質含量の変化 70 3.温度,CO2 濃度上昇の水稲への影響解析 には幼穂形成期までの窒素吸収量が増えたことが関係していた(Kim et al., 2001).稔実率やモミ 千粒重は CO2 増加の影響を受けなかったが,収穫係数(穂重/全重)は高 CO2 濃度により 2 〜 3% 低下した.また,高 CO2 濃度は,どの窒素施肥量でも白米のタンパク含量を 6 〜 9g/kg(比率で は 7 〜 12%)低下させた(図 4) .タンパク含量が低いと食味が向上することが知られているため, 食味試験を行った結果,高 CO2 濃度で食味が向上する傾向を認めた.(平成 14 年度研究成果情報, 小林ら) ⑶ FACE 実験による水稲の水消費量への影響 FACE 実験によって,高 CO2 条件下における水稲の水消 費量の変化について実験を行った.その結果,大気 CO2 濃 表 1 大気 CO2 濃度上昇による水稲 生育期間中の水需要量の変化 度が上昇すると生育が促進されるので収量は増加し,高 CO2 要素 高 CO2 濃度による変化 濃度区の乾物重は現 CO2 濃度区より約 9%大きくなった.一 日中の平均葉温 + 0.27℃ ±0.14℃ 方,高 CO2 濃度により,葉の気孔が閉じ気味となって蒸散 収穫時の乾物重 + 9.1% 積算蒸散量 − 8.2% が抑制されるため,高 CO2 濃度区の葉温は現 CO2 濃度区よ り期間平均で約 0.3℃高くなった.このため,表 1 に示した ように,田植えから収穫までの水稲群落の蒸散量(水消費 乾物量 1g 生成に − 16% 要する水需要量 (葉温データは,平均値 ± 標準誤差) 量)は,高 CO2 濃度により約 8%(22mm)減少し,全生育 期間を通して 1g の乾物重を得るために植物が必要とする水需要量は,高 CO2 濃度よって約16%減 少することがわかった(Yoshimoto et al., 2005a) .他の主要な作物での高 CO2 濃度による水需要量 の減少率は 7 〜 41%と幅広いことが知られており,水稲の高 CO2 濃度による水節約の効果は世界 の主要作物の中ではそれほど顕著でないことが明らかとなった.(平成 18 年度研究成果情報,吉本 ら) ⑷ 人工気象室による水稲の窒素栄養 農業環境技術研究所内の自然光 人 工 気 象 室 を 用 い て, 水 稲( 品 種 「日本晴」 ) を 現 在 の 外 気 CO2 濃 度 (380ppm) と 高 CO2 濃度(680ppm) の 2 条件で栽培し,水稲群落の日中の 光合成量と夜間の呼吸量を算出し,水 稲の窒素栄養との関係を解析した. その結果,CO2 濃度の上昇による光 合成の促進率は生育初期に高く,生育 につれてしだいに低下するが,呼吸 速度は高 CO2 濃度によって生育期間 中促進されることが,群落レベルで 初めて明らかになった(Sakai et al., 2001) .また,高 CO2 濃度による水稲 の生長の促進は,生育にともない大き く低下した.さらに,図 5-5 に示した 図 5 3 年間の実験から得られた水稲葉の窒素濃度と群落光合 成の関係 ように高 CO2 濃度による群落光合成 の促進率については,葉窒素濃度が高い場合には高く,葉窒素濃度が低下すると低くなることがわ かった.葉窒素濃度は生育に伴い低下するが,高 CO2 濃度条件ではその程度が外気 CO2 濃度条件 第 5 章 イネ・水田生態系の応答と適応戦略 71 よりも大きいことも,群落光合成の促進率を低下させる要因であった(Sakai et al., 2006).これら の結果は,CO2 濃度上昇時の水稲生産を正確に予測するためには,CO2 濃度上昇下での水稲葉の窒 素濃度を的確に予測する必要があること,CO2 濃度上昇による生長の“施肥”効果を高めるために は,窒素管理が重要であることを示していた. (平成 18 年度研究成果情報,酒井ら) 4.影響把握・予測のためのモデル開発・データベースの構築 ⑴ 水稲穂温の推定 大気 CO2 濃度の上昇は,イネの成長促進と増収効果を持つ一方,気孔開度の減少,蒸散の抑制, 群落温度の上昇を通じて,温暖化や高温地域で問題となる開花期不稔を激化させる可能性がある. 高温,高 CO2 環境下における不稔発生を予測するためには,的確な穂温の予測が必要である.そ こで,穂をとりまく微気象環境が穂温に影響を及ぼすプロセスを考慮した穂温推定モデルを開発す るため,中国江蘇省無錫の水田 FACE 実験ほ場の[現 CO2 濃度区]と[高 CO2 濃度区(=現 CO2 濃度+ 200ppm) ]で,水稲(武香粳 14 号)の出穂・開花期に微気象観測を行った. その結果,高 CO2 濃度による気孔開度の減少は個葉の蒸散を抑制し,潜熱として葉面の熱が奪 われる冷却効果を減らすため, [高 CO2 濃度区]の葉温は[現 CO2 濃度区]に比べて 1 〜 2℃高 かったことが明らかとなった.また,群落内の気温は,[高 CO2 濃度区]の方が 0.5 〜 1℃高く, 相対湿度は 5 〜 8%低かった(図 6) .さらに,穂は表面に気孔がなく,表皮を通して蒸散している ため,高 CO2 濃度の穂の蒸散速度への直接的な影響はなかった.両 CO2 濃度区とも,穂の蒸散速 度は開花直後に高く,日数が経つ につれて減少する傾向を示した (Oue et al., 2005) . この微気象変化と穂の蒸散速度 の測定データを用いて,穂の温 度を推定する熱収支モデルを開 発した.このモデルによって高 CO2 濃度による穂温の上昇は 0.5 〜 1℃であり,高 CO2 濃度によっ て高温不稔を助長し得ること,ま た,高 CO2 濃度は穂温を上げる と同時に,群落内の湿度を下げる ため,穂の蒸散(脱水)を促進す ることが明らかにされた(平成 図 6 穂温推定モデルで計算された高 CO2 濃度による(a)穂温と(b) 穂の蒸散量の変化([高 CO2 濃度区]-[現 CO2 濃度区]の 較差で表示) 17 年度研究成果情報,吉本ら) (図 6) . ⑵ 水田の水温・地温の推定 今後の温暖化に対応した水稲の安定生産技術の確立や水田からの温室効果ガス発生量評価などの 目的で,水田の水温や地温の広域的な推定手法の確立が求められている.そこで,熱収支理論に基 づき,水田の水温・地温を計算するためのモデル(以下「水田水温モデル」)を新たに開発した. このモデルでは,気象データ(気温,湿度,風速,日射量,下向き長波放射量)と葉面積指数 LAI を入力データとして,水田の水温および地温(深さ 0 〜 5cm)の日平均値が計算できる.日 平均水温と深さ 0 〜 5cm の日平均地温はほぼ一致するため,モデルでは両者を等しいと仮定して いる.北海道から九州にかけての複数地点の水田で実測したデータを用いて,生育初期から出穂期 72 4.影響把握・予測のためのモデル開発・データベースの構築 までの測定値と計算値を比較 したところ,平均偏差(測定 値と計算値の差の平均値)が 概 ね 0.5 ℃ 以 内,2 乗 平 均 平 方根誤差(RMSE)は 1℃前 後であり,十分に実用的なレ ベルであった. ま た, 水 田 の 水 温・ 地 温 (日平均値)を広域推定する 手 法 を 開 発 し た.日本気象 協会の局地気象モデルによる 気温,湿度および風速などの 計算結果を,アメダスデータ や気象衛星 GMS データによ る日射量の推定値と組み合わ 図 7 モデルにより推定した東北地域における 1997 年 6 月 17 日の日射 量(左)と水田水温と気温の差(日平均)の 1km メッシュ分布. 右図の黒色は水田のないメッシュ. せることによって,水田の水温・地温の計算に必要な 1km メッシュ気象データを作成できる.本 手法によって推定した,東北地方における 1997 年 6 月 17 日の日射量および水田水温と気温の差の 1km メッシュ分布では,日射量の多い地域ほど水田水温と気温の差が大きな傾向にあり,ほぼ妥 当な水田水温の分布となっていた(平成 15 年度研究成果情報,桑形ら)(図 7). ⑶ 水稲穂温と水田水温の推定 温暖化による作物生産変動を評価・予測するため,アメダス地点における気象データからモデル によりアメダス地点の水稲穂温や水田水温を推定する「モデル結合型作物気象データベース」を開 発した. データベースには全国のアメダス地点(約 850 地点)における 1980 年以降の日別気象として, 気温,風速,降水量,日射量,湿度および蒸散要求量などの作物生産において重要な要素(モデル による算定値)を収納した.各地点における気象データには,地力保全基本調査による土壌デー タ(日本土壌協会)を付加した. データベース上のメニュー画面 もしくは Google Earth の地図上 から,任意のアメダス地点を選ぶ ことによって,気象データを容易 に取り出すことができる.また, データベース本体に「物理環境モ デル」と「生育モデル」が組み込 まれ,水稲の稔実や登熟に影響 を及ぼす水田水温(日別値)や 出穂・開花期における次期別穂 温(Yoshimoto et al., 2005b) ,主 要品種の生育ステージや LAI(葉 面積指数)などを推定することが できる(図 8) . 図 8 モデル結合型作物気象データベースの基本構造 第 5 章 イネ・水田生態系の応答と適応戦略 73 本データベースを既存の作物データベースや栽培試験データと組み合わせることによって,近年 の温暖化傾向や気象変動が水稲生産に及ぼしている影響を解析・解明するためのデータセットが容 易に得られる.これらの解析を通して,水稲生産の将来予測や収量低下のリスク評価,適応技術の 開発などに大きく貢献できる. (平成 19 年度研究成果情報,桑形ら) ⑷ 日射量の推定 将来の気候変動において,日射量の変化が水稲生育に影響を及ぼすことが懸念される.しかし, 日射量の一般に利用可能なデータは,全国で 67 地点の地上気象観測所(気象庁)における測定 データに限られる.そこで,日本全国のアメダス地点(約 850 地点)で測定されている毎時の日照 時間から時別日射量を高精度で推定する手法を新たに開発し,時別日射量のデータセットを作成し た. この手法において,時別日射量 (1 時間積算値)は,毎時の日照時 間,太陽高度角,時別降水量,積 雪の有無および日照時間(日積算 値と前後 1 時間の値)から推定し た(Masaki et al., 2009a,b) .時別日 射量の推定精度は 0.2MJ/m2/h 前後 で,従来の手法の推定精度以上で あった.また,時別日射量の推定値 から得られる日積算日射量の推定精 度は約 1.5MJ/m2/d となり,従来の 図 9 2004 〜 07 年の 67 地点での日射量の推定精度(左)と従来 法との比較(右) 日別日照時間による推定手法を用い た場合に比べて精度が向上した(図 9) . 今回新たに開発した手法を用いて,日本全国のアメダス地点における,1991 年以降の時別日射 量のデータセットを作成した.このデータと水田の物理環境モデルと組み合わせることで,水稲の 稔実や登熟に影響を及ぼす出穂・開花時刻の穂温を地域ごとに細かく評価することが可能で,高温 による生育障害の現況把握や対策技術の策定に役立つ.(平成 21 年度研究成果情報,桑形ら). ⑸ 国際観測ネットワークの構築 精密かつ簡便に微気象を測定でき る装置を用い,世界共通の手法で水 田微気象や高温障害の実態を把握す る国際ネットワークを構築した. 水田微気象の測定には,精密か つ簡便に水田内の気温と湿度を測 定できる自立型気象観測パッケー ジ MINCER(平成 22 年度研究成果 情報,福岡ら)を用い,国により 異なっていたイネの高温障害の調 査方法を統一した.2010 年現在の MINCERnet 参加国は,世界有数の コメ生産国であるインド,スリラン 図 10 MINCERnet 観測水田内および水田と最寄りの気象観測点 との気温差 74 5.温暖化に対するぜい弱性の予測 カ,ミャンマー,中国,フィリピン,台湾,アメリカおよび日本の 8 か国である.その結果,気候 の違いによって,イネの出穂期や登熟期における水田での湿度レベルや気温の日較差,夜温などが 大きく異なることが判明した.また,現地の最寄りの気象観測点は一般に水田地帯には設置されて いないため,水田上より気温が高く観測される場合が多いこと,イネの穂周辺の気温は水田上の 気温より低いことが多く,その差は地域により異なることが明らかとなった(図 10).このように MINCERnet は,各国共通の手法を用いることでイネの高温障害に直接関わるイネの穂周辺の気 温・湿度の正確な測定を可能とし,イネの高温不稔や登熟不良との関係を正確に把握できるため, 今後,高温障害の原因究明や温暖化影響の高精度評価に貢献することが期待される.(平成 22 年度 研究成果情報,吉本ら) 5.温暖化に対するぜい弱性の予測 地球温暖化が日本の水稲生産に 及ぼす影響については,単に登熟 期の高温による生産量の増減を評 価するだけでは不十分であり,病 害虫の発生や農業用水資源の不足 などの要因についても対象とする 必要がある.そこで,大気中の CO2 濃度が現状の約 2 倍となり, 日本付近の地上気温が約 2℃上昇 するとされる 2060 年代における 水稲生産量,害虫の世代交代速 度および降積雪量の変化を 10km メッシュ単位で推定し,温暖化に よって水稲生産が負の影響を受け る可能性のある地域,すなわちぜ い弱な地域を検出した. 水稲生育に関し,登熟期の気温 図 11 最大収量となる最適気温(現在気温との差).値が正(青系 色)であれば気温の上昇とともに収量は増大し,負(赤系 色)であれば収量は低下する を変数とする回帰推定式を用いて,最大収量が期待される適温値を推定し,温暖化時の水稲収量を 見積もった.その結果,図 11 に示したように北陸,南関東および九州北部では,現在の気候条件 においても適温値より高温の条件で栽培されているため,温暖化による高温ストレスの影響を受け やすいことが示された(西森ら,2001) . また,水資源について,温暖化時の気温の変化から降積雪量の変化量を推定すると,東北〜北陸 の日本海側では,降雪量が約 15%減少し,また融雪時期も現在より早まる.このため,これらの 地域では水稲の代かき・田植え期に河川水量が減少し,農業用水が不足する可能性が高くなると推 定した(井上ら,2001) . さらに,害虫影響では,有効積算温度則を用いてヒメトビウンカの発育速度を推定した結果,温 暖化時には発育速度が速まって世代数が増え,イネ縞葉枯病の多発危険地域が,東北〜北陸および 関東南部にまで広がると予測された(Yamamura et al., 2002). 以上の結果を総合すると温暖化時には,北陸地方は,気象,水資源および害虫の全要因の変化が 水稲生産に負の影響を及ぼす,すなわちぜい弱であると推定された.次にぜい弱な地域は,東北日 第 5 章 イネ・水田生態系の応答と適応戦略 75 本海側地域(水資源と害虫要因の変化にぜい弱)および南関東地域(気象と害虫要因の変化にぜい 弱)と考えられた. (平成 13 年度研究成果情報,西森ら) 引用文献 1)長谷川利拡,吉本真由美,桑形恒男,石郷岡康史,近藤始彦,石丸 努(2009)2007 年夏季 の水稲の高温不稔調査について.農業および園芸,84, 42-45 2)井上 聡,横山宏太郎,大野宏之,川島茂人(2001)地球環境変化時における降積雪の変動予 測(続報) ― 国内主要河川流域における降雪水量の変動.雪氷,63, 489-499 3)Kim,H.Y., Lieffering,M., Miura,S., Kobayashi,K. and Okada,M.(2001)Growth and nitrogen uptake of CO2-enriched rice under field conditions. New Phytologist, 150, 223-229 4)小林和彦(2001)FACE(開放系大気 CO2 増加)実験 . 日本作物学会紀事,70, 1-16. 5)Kobayashi,K., Lieffering,M. and Kim,H.Y.(2001)Growth and yield of paddy rice under free-air enrichment. In. Shiyomi,M. and Koizumi,H.(Eds), Structure and Function in Agroecosystem Design and Management, CRC Press, Boca Raton, FL, USA, p371-395 6)Masaki,Y., Kuwagata,T. and Ishigooka,Y.(2009a)Atmospheric turbidity estimation from hourly global solar radiation data under cloudless skies. Journal of the Meteorological Society of Japan, 87, 849-863 7)Masaki,Y., Kuwagata,T. and Ishigooka,Y.(2009b)Precise estimation of hourly global solar radiation for micrometeorological analysis by using data classification and hourly sunshine. Theoretical and Applied Climatology, 100, 283-297 8)西森基貴,横沢正幸(2001)気候変動・異常気象による日本の水稲単収変動の地域的変化.地 球環境,6, 149-158 9)西森基貴,長谷川利拡,桑形恒男,石郷岡康史,眞崎良光,飯泉仁之直,村上雅則(2009) 2009 年夏季の低日照と水稲作況の低下-中四国地域の事例を含めて-.中国四国の農業気象, 22, 68-69 10)Oue,H., Yoshimoto,M. and Kobayashi,K.(2005)Effects of free-air CO2 enrichment on leaf and panicle temperatures of rice at heading and flowering stage. Phyton, 45, 117-124 11)Sakai,H., Yagi,K., Kobayashi,K. and Kawashima,S.(2001)Rice carbon balance under elevated CO2. New Phytologist, 150, 241-249 12)Sakai,H., Hasegawa,T. and Kobayashi,K.(2006)Enhancement of rice canopy carbon gain by elevated CO2 is sensitive to growth stage and leaf nitrogen concentration. New Phytologist, 170, 321-332 13)Yamamura,K. and Yokozawa,M.(2002)Prediction of a geographical shift in the prevalence of rice stripe virus disease transmitted by the small brown planthopper, Laodelphax striatellus (Fallen)(Hemiptera: Delphacidae), under global warming. Applied Entomology and Zoology, 37, 181-190 14)Yoshimoto,M., Oue,H. and Kobayashi,K.(2005a)Energy balance and water use efficiency of rice canpies under free-air CO2 enrichment. Agricultural and Forest Meteorology,133,226-246 15)Yoshimoto,M., Oue,H., Takahashi,N. and Kobayashi,K.(2005b)The effects of FACE(FreeAir CO2 Enrichment)on temperatures and transpiration of rice panicles at flowering stage. Journal of Agricultural Meteorology, 60, 597-600 76 第 6 章 温室効果ガス発生抑制と土壌炭素蓄積 1.はじめに 急激な人間活動の拡大は地球規模での物質循環に影響を与え,大気中の温室効果ガス濃度を増加 させている.この問題に対し,気候変動に関する政府間パネル(IPCC)の最新の評価報告書は, 「我々を取り巻く気候システムの温暖化は決定的に明確であり,人類の活動が直接的に関与してい る」ことを明示し,あらゆる場面で温室効果ガスの排出を削減する緩和策を実行し,地球温暖化の 動きを遅らせ,さらには逆転させることの必要性を強く認識させている(IPCC, 2007a). 農業生産活動もその例外ではなく,農地からも主として土壌中の微生物活動に起因する温室効 果ガス,すなわち,二酸化炭素(CO2) ,メタン(CH4),および一酸化二窒素(亜酸化窒素:N2O) が排出される(図 1) .全球における農業分野からの温室効果ガス排出量は CO2 換算量で年間 51 〜 61 億トンと見積もられており,人為起源の 13.5%を占めている.このうち,最大の温室効果ガスで ある CO2 については,農地における発生と吸収の収支は全球でほぼバランスしていると考えられ ている.しかし,水田や家畜から発生する CH4 と,農地への窒素施肥や家畜排泄物からの N2O に ついて,農業生態系は,それぞれ,人為起源発生量の半分以上を占めており,重要な排出源となっ ている.これらに加えて,別に算定されている森林からの温室効果ガス排出(58 億トン)は,主 として,森林から農地への土地利用変化を原因とするものであることから,農業の影響は森林分 野にも及んでいると言える.これらを合計すると,農業の関与する温室効果ガス排出量は全人為 排出量の約 1/3 を占めることになり,地球温暖化に対する農業の影響はきわめて大きいと言える (IPCC, 2007b) . CO2 శวᚑ 䈫ๆ ფๆ N2O శวᚑ CH4 శวᚑ 䈫ๆ ൻቇ⢈ᢱ ᯏ‛ 䊥䉺䊷䉇ᱷᷲ 䈱ଏ⛎ ᓸ↢‛䈮 䉋䉎ಽ⸃ C ᯏ‛ N N2O ⎣ൻ䈫⣕⓸ C CH4 ფᯏ⚛ (SOC) 䊜䉺䊮↢ᚑ 図 1 農業生態系(農地)における温室効果ガスの発生と吸収 一方,農地管理の改良や家畜の飼料と排泄物管理の工夫により,温室効果ガス排出量を大きく削 減できる可能性がある(八木,2009) .実際,これまでの研究により農地からの CH4 および N2O 発 生削減技術の候補は多数提案され,多くについて現地試験等から削減効果が確認されている.加え て,農地への有機物投入量の増加や耕うん強度の低減により,農地土壌の炭素貯留量を増加する, 第 6 章 温室効果ガス発生抑制と土壌炭素蓄積 77 すなわち,農地を大気 CO2 の吸収源に変えることが可能である.これらの技術を活用し,農地か らの温室効果ガス排出量を大幅に減少させ,地球温暖化の緩和に寄与する方策を実行することが求 められている. 農業環境技術研究所では,1980 年代より,農地土壌からの N2O 発生および水田からの CH4 発生 の研究を行ってきた(野内,2005) .また,全国規模の土壌調査結果を一元的に収集してきた土壌 インベントリー(平成 15 年度研究成果情報,中井ら)から,農地土壌の CO2 吸収・排出を評価す ることが可能である.近年,地球温暖化対策がますます重要性を増し,温室効果ガスの発生量の評 価と排出削減への寄与が求められていることに対応して,農業環境技術研究所における温室効果ガ ス発生抑制と土壌炭素蓄積における研究は,過去の研究蓄積を活用し,さらなる展開を進めてき た.過去 10 年間における研究の焦点は,土壌中での温室効果ガス生成と炭素蓄積メカニズムの解 明,高精度かつ簡易なモニタリング手法の開発,広域推定のための発生量評価手法とモデル開発, および発生抑制技術の開発である. 2.温室効果ガス発生・土壌炭素蓄積メカニズムの解明 ⑴ 水 田 か ら の メ タ ン ( C H 4) 発 生 メ カ ニ ズ ム 水田ではかんがい水により土壌を湛水することから土壌中の酸化物質が徐々に還元され,嫌気的 な環境が発達した後,メタン生成菌と呼ばれる一群の絶対嫌気性古細菌の活動により有機物分解の 最終生成物として CH4 が生成される.CH4 生成は,嫌気条件下での物質代謝の最終ステップであ り,メタン生成菌は他の生物が複雑な有機物を分解して排出した低分子化合物から CH4 を生成す る.絶対嫌気性細菌であるメタン生成菌の特性から,土壌中での CH4 生成には,湛水にともなう 土壌の還元の発達が必要不可欠な条件となる.水田土壌では,湛水開始後,土壌中の酸化物質が 徐々に還元され,酸化還元電位(Eh)が−150mV 程度に低下した後,CH4 生成が開始される.土 壌中で生成された CH4 は,気泡として,田面水中を拡散して,または水稲を通って,のいずれか の経路で大気へと放出される.このうち,量的にもっとも重要なのは,水稲の通気組織を通って放 出される経路である.一方,水田土壌中には CH4 を酸化分解する別の一群の細菌(メタン酸化菌) が存在し,一部の CH4 はこれにより消費される(八木,2004). ⑵ 施 肥 窒 素 か ら の 一 酸 化 二 窒 素 ( N 2O ) 発 生 メ カ ニ ズ ム 作物生産に必要な化学肥料や有機物として農耕地に施用された窒素は,土壌中で微生物による形 態変化を受け,アンモニア態窒素(NH4-N)から硝酸態窒素(NO3-N)へ(硝化),NO3-N から分 子状窒素(N2)へ(脱窒)と変換される.N2O は土壌中での硝化および脱窒の両方の過程で副生 成物または中間生成物として生成され,大気へ放出される.同じガス態の窒素酸化物であり,光化 学スモッグや酸性雨の原因物質である一酸化窒素(NO)も同様にこれらの過程での副生成物とし て生成される(秋山ら,2011) . 硝化は好気的条件において起こる反応であり,NH4-N から亜硝酸態窒素(NO2-N)へ酸化される アンモニア酸化の過程と NO2-N から NO3-N へと酸化される亜硝酸酸化とに分かれる.前半のアン モニア酸化の過程は,従来はアンモニア酸化細菌のみによるものと考えられていたが,近年,アン モニア酸化古細菌が海洋や土壌中に広く分布し,アンモニア酸化細菌とともに反応を担っているこ とが明らかになってきている(Hayatsu et.al., 2008).後半の亜硝酸酸化の過程は亜硝酸酸化細菌 が担っている.一方,脱窒は嫌気的条件において有機物を電子供与体として,酸化的窒素化合物 を電子受容体として用いる呼吸の一種である.自然界に生息する多くの細菌がこのような脱窒能を 有している.従来は細菌のみが脱窒過程を担っていると考えられていたが,脱窒活性の測定にお 78 3.モニタリング手法の開発 いて,アセチレンを添加しなくても多量の N2O 発生する現象が認められ,既知の脱窒菌以外の微 生物による N2O 発生である可能性が示唆された(平成 16 年度研究成果情報,駒田ら)(駒田・竹 内,2003) .また,糸状菌(カビ)にも脱窒系を持つものがあることが明らかになり,このカビ脱 窒も土壌生態系で重要な役割を担っていると考えられるようになった(Hayatsu et.al., 2008).こ のように,さまざまな種類の微生物が硝化と脱窒を担い,これらの両方の過程において N2O が生 成される.さらに,硝化細菌のなかにも脱窒経路を持つものがあり,硝化菌による脱窒(nitrifier denitrification)も重要な N2O 発生源である可能性がある(Kool et al., 2010).このようなさまざ まな土壌微生物の群集構造と N2O の生成・発生メカニズムの関係について,近年急速に発達して いるメタゲノム解析等の分子生物学的手法を用いた研究が進みつつあり,わが国の転換畑(Chu et al., 2009)やインドネシアの農地土壌(Jumadi, 2008)でのアンモニア酸化細菌の群集構造が示さ れた. ⑶ 土 壌 炭 素 蓄 積 と 農 地 か ら の 二 酸 化 炭 素 ( C O 2) 発 生 ・ 吸 収 メ カ ニ ズ ム 農地における炭素循環は図 1 に示すように,植物を介した大気 CO2 と土壌有機物炭素の交換と 考えることができる.植物は光合成により大気中の CO2 を有機物として固定しているが,その一 部がリターフォールとして土壌に付加される.農地では,作物残渣と刈り株がこれに相当する.さ らに,土壌中では枯死根や根からの分泌有機物がこれに加わる.農地では,さらに,堆肥などの有 機物資材が加えられる.以上が土壌への炭素の入力量となる.これに対し,出力量としては,大気 への直接の CO2 放出である植物の呼吸と土壌有機物の分解がまずあげられる.土壌有機物の分解 は,土壌に生息するさまざまな微生物や動物の働きによるもので,従属栄養呼吸(heterotrophic respiration)とも呼ばれる.農地の CO2 収支はこれらの入力量と出力量のバランスから決定され, 農地土壌は大気 CO2 の吸収源とも発生源ともなりうる. このような農地における炭素循環と CO2 の吸収・発生量を評価するために,土壌有機物の生成 と分解の両過程に関する定量的理解が必要である.特に,わが国の黒ボク土には黒みの強い腐植酸 が異常に多量に蓄積されており,その生成・ 集積メカニズムについて議論が行われ,ススキ草原 を維持するために行われてきた野焼きによって生成されたススキの燃焼微粒炭が腐植酸の給源で あるとする学説が提案された(Shoji et al., 1993).この問題に対して,日本各地で収集した土壌か ら腐植酸を精製して炭素安定同位体比(δ13C 値)を測定した結果,ススキなどの C4 植物に由来 する炭素が腐植酸中の炭素に占める割合は 18 〜 52%であり,ススキ以外の植物も腐植酸の生成過 程で重要な給源であることを明らかにした(平成 15 年度研究成果情報,平舘ら)(Hiradate et al., 2004) . 3.モニタリング手法の開発 土壌から大気への温室効果ガス発生・吸収量を評価するためには,現場でのガスフラックス(流 束:単位面積,単位時間あたりのガス交換量)をモニタリングすることが欠かせない.その方法と しては,チャンバー法,ガス拡散理論の応用法,および微気象学的測定法がある.土壌から大気へ のフラックスを測定する場合にはチャンバー法が,植物群落上でのフラックス測定には微気象学的 測定法がよく使われている.微気象学的測定法については,第 12 章で研究の歩みが紹介される. チャンバー法は簡易で数多くの地点において計測できることから,発生要因の解明や発生抑制技術 の評価などに活用されている.しかし,チャンバー法でガスフラックスの空間・時間変動を精度良 く評価するためには,多点で,頻度の高い繰り返し計測が必要である. チャンバー法では,土壌面にチャンバー(底のない容器)を置き,一定時間ごとにチャンバー内 第 6 章 温室効果ガス発生抑制と土壌炭素蓄積 79 の空気サンプルを採取し,サンプル中のガス濃度を分析することによりフラックスを計算する. その際,チャンバー内に外部からの空気の入出流のないクローズドチャンバー法と,チャンバー 内の空気を常に交換できる流路を設置したダイナミックチャンバー法がある.水田からの CH4 や 施肥土壌からの N2O 計測では土壌空気とチャンバー内空気のガス濃度差が大きく,計測中のチャ ンバー内ガス濃度の増加は直線性が良いため,クローズドチャンバー法で十分計測可能である(八 木,1997) .しかし,土壌からの CO2 フラックスは計測中にガス濃度差が小さくなり,みかけのフ ラックスが減少するため,ガス濃度差を維持できるダイナミックチャンバー法,または計測時間を 数分に短縮したクローズドチャンバー法が用いられる. ほ場へのチャンバー設置やガスサンプリングを手動で行うクローズドチャンバー法は,労力がか かるが装置が簡単で安価なことから,土壌からの温室効果ガス発生量の計測に現在でも広く使われ ている.しかし,土壌から発生する温室効果ガスのフラックスは温度,降雨などの環境要因と施 肥,水管理,耕起などの作業により大きな時間変動を示すため,それを精度良く定量するために は,高頻度で測定することが望ましい.また,作物の非栽培期間でも顕著な発生の見られる場合が あるため,通年で測定することが必要である.このような計測精度に関わる問題に対して,一般的 に用いられている手動のクローズドチャンバー法では多大な労力がかかるため,測定頻度や期間を 十分にとることは困難な状況であった.農業環境技術研究所には,自動開閉チャンバー,自動ガス サンプリング・分析システムを備えた温室効果ガス発生制御施設が設置されており,この施設にお いては高頻度・通年測定が可能である(Nishimura et al., 2005b).しかし,大型の固定式装置のた め,他のほ場に移動することはできない. このため,固定式の大型自動連続モニ タリング装置よりも小型で低コストな可 搬型ガスサンプリング装置を開発した 䉰䊮䊒䊥䊮䉫ⵝ⟎ (図 2) (平成 20 年度研究成果情報,秋 山ら) (Akiyama et al., 2009) .この装置 は自動開閉チャンバーとともに使用し, 制御部内のマイクロコンピューターにプ ログラムされた設定時刻になると,自動 開閉するチャンバーからのサンプルガス をシリンジポンプで採取し,装置内のカ セットに並べた真空バイアル瓶に自動で ⥄േ㐿㐽䉼䊞䊮䊋䊷 注入する.サンプリング済みバイアル瓶 はガスサンプル注入部の下のバイアル瓶 図 2 自動開閉チャンバーと可搬型自動サンプリング装置 受けに落下し,定期的に回収され,分析 に供される(図 3) .この装置を用いる と,大幅な労力削減となる.たとえば 6 つのチャンバーで毎日 1 回の割合でサンプリングを行った とすると,手動サンプリングでは 10 日間でのべ 10 時間以上必要であるのに対し,自動サンプリン グではバイアル瓶の設置と回収に要する約 1 時間のみとなり,1/10 以下の作業時間となる.また, 標準ガスを用いたテストの結果,本装置による測定精度は手動サンプリングと同等であることが確 認された(CO2: r2=0.998, CH4: r2=0.999, N2O: r2=1.00).本装置は可搬型(大きさ:390 × 590 ×高 さ 870 mm,重さ:約 50 kg(大きさ,重さともに設置用台および遮光版を含む))であるため,さ まざまなほ場に設置することができる. 80 3.モニタリング手法の開発 図 3 温室効果ガス自動サンプリング装置の概要図 一方,試料ガスを 1 回自動注入することによって,温室効果ガス 3 成分(CO2,CH4,N2O)を 同時に計測できる新たな分析計を開発した(平成 21 年度研究成果情報,須藤)(図 4).同時計測 は,CH4 および CO2 のピークを N2O のピークから完全に分離するための新たな充填剤の導入とそ れぞれのガスを検出するためのキャリヤーガスを共通化することで実現した.装置流路図(図 5) 中央に示すとおり,3 段階のガス分離の第 2 段に分離能の高い充填剤(Unibeads C,GL サイエン ス社製)を採用することで,これまで困難とされた 3 ガスの完全分離を実現した.また,CO2 およ び N2O の高感度検出のために,従来は,それぞれ,ヘリウムおよび 5% CH4 混合アルゴンがキャ リヤーガスとして用いられていたが,本装置では,ECD セル内でヘリウムに N2 と CH4 の混合ガ スをメークアップガスとして添加することで,従来法と同等の N2O の検出感度を得るとともに, N2O と CO2 の同時分析を可能にした.1 回の試料注入で 3 成分を同時に計測できるので,3 成分を 別々に測定する場合に比べて注入量による誤差が生じないこともメリットの 1 つである.3 成分の 繰り返し分析精度も従来法と同等で あった.試料注入については,既存の ヘッドスペースサンプラー(HSS-2B, 島津製作所)を改造し,高精度ガスタ イトシリンジ(容量 2ml,Valco 社) を装着した自動注入器を開発した.シ リンジの針を高気密性のブチルゴム栓 に貫通させるために,円錐形のガイド ラインを新たに製作した.この自動注 入器により,最大 40 試料を,1 試料 あたり 10 分で自動分析できた.本分 析計には,特定メーカーの検出器およ びガスクロマトグラフを使用している が,他社の製品を利用して製作する 図 4 温室効果ガス 3 成分同時分析計の外観 第 6 章 温室効果ガス発生抑制と土壌炭素蓄積 81 FC5 He (99.999%) FC6 '%&ᷝട↪⓸⚛ '%&ᷝട↪ࡔ࠲ࡦ ࠠࡖࡗࠟࠬ⚐ൻⵝ⟎ ࠟࠬ⚐ൻⵝ⟎ ࠟࠬ⚐ൻⵝ⟎ He He He FC1 MoleculerSieve 5A Porapak N ࠞࡓ FC3 FC2 ⹜ᢱᵈญ ࠞࡓ 第3段分離 vent ECD TCD 1 PorapakN ࠞࡓ 1 10 ⹜ᢱዉญ 9 ⹜ᢱ⸘㊂▤ SV1 8 7 6 1 Unibeads C 2 3 ࠞࡓ 4 第2段分離 SV2 5 5 第1段分離 Porapak N ࠞࡓ ഥΆ↪ⓨ᳇ 3 4 vent Simalite Q ⹜ᢱឃ᳇ญ (+&↪᳓⚛ 2 6 第3段分離 ឃ᳇ Porapak N He FC4 FID 1 ࠞࡓ TCD 2 FID 2 ࠞࡓ MoleculerSieve 5A ࠞࡓ SV:流路切り替えバルブ FC:流量調整弁 ഥΆ↪ⓨ᳇ (+&↪᳓⚛ 図 5 温室効果ガス 3 成分同時分析計の流路図 ことも可能である. 開発された可搬型ガスサンプリング装置と温室効果ガス 3 成分同時自動分析計を併用することに より,より効率的な温室効果ガスフラックスのモニタリングが可能になった.これにより,農地に おける温室効果ガスのモニタリングと発生抑制技術の開発に関する研究が大幅に進むことが期待さ れる. 4.発生量評価とモデル開発 ⑴ 水 田 か ら の メ タ ン ( C H 4) 発 生 1980 年代以降,世界の各地で水田からの CH4 発生の測定が行われ,発生量と気候や処理による その変動が報告された.これらの測定結果をまとめると,水稲栽培期間の 1 時間平均の CH4 フラッ クスは多くの場合 1m2 あたり数 mg 〜数十 mg,栽培期間全体の CH4 発生量は 1m2 あたり 1 g 〜 100 g の範囲にあり,測定地点や処理により CH4 発生量は大きく異なる.特に,有機物を多く施用 した場合,大きな CH4 発生が観察されている.世界各地の水田における CH4 発生量の変動は,温 度や降雨などの気候条件,土壌の理化学性,有機物や水管理などの耕作管理方法の違いなど,さま ざまな要因の寄与が明らかにされている(八木,2004). わが国においては,1992 〜 1994 年にかけて行われた,農耕地からの温室効果ガス発生に関する 全国的なモニタリングデータをもとに発生量評価が行われた(財団法人日本土壌協会,1996).こ の全国調査の結果は,水稲一作あたりの CH4 フラックスの平均値は,稲わらを秋に土壌還元した 処理区で 19.0 ± 12.5g/m2 であったことを報告した.さらに,このデータを土壌タイプごとに集計 し,有機物無施用や堆肥などの有機物施用実態とそれによる発生量の変化を考慮すると,わが国の 水田からの年間 CH4 発生量は 330Gg(33 万トン)と推定された(八木,2004). その後もわが国や他のアジア諸国の水田において CH4 発生モニタリングが継続され,特に, 82 4.発生量評価とモデル開発 新たな地点やさまざまな耕作体系におけるモニタリング,最新の計測技術を用いた国際共同研究 (Wassmann et al., 2003) ,および次節で述べる発生抑制技術の開発試験に焦点が当てられるように なった.農業環境技術研究所温室効果ガス発生制御施設を用いた研究では,水田を転換畑とする ことにより CH4 発生はなくなるが,N2O が増加するトレードオフ関係になることが明らかになっ た(Nishimura et al., 2004;2005a) .また,水田における開放系大気 CO2 増加(Rice-FACE)実験 や半閉鎖型自然光環境制御チャンバー(クライマトロン)実験から,大気 CO2 濃度の上昇が水田 からの CH4 発生を促進し(Inubushi et al., 2003),その程度は高夜温によって抑制されることを明 らかにした(平成 20 年度研究成果情報,長谷川ら)(Cheng et al., 2008).Rice-FACE 実験から, 水稲生育期間中の高 CO2(+200ppm) ,温暖化(+2℃)環境下では,水田からの CH4 発生が現在 よりも約 80%増加する可能性のあることを明らかにした.(平成 22 年度研究成果情報,長谷川ら) (Tokida et al., 2010) . 世界各国の温室効果ガス発生量の算定に用いられている 1996 年版 IPCC ガイドラインでは,水 田からの CH4 発生に関して,実測データの統計解析を行わずに排出係数を求めるなど大きな不確 実性が存在した.そこで,既往文献からアジア諸国における水田からの CH4 発生実測データを収 集し,8 カ国,103 地点,868 栽培期間データからなるデータベースを構築した.これらのデータ について,栽培期間の平均 CH4 フラックスと各発生制御要因の関係を解析し,算定の基準となる ベースライン排出係数(1.30kg CH4/ha/day)と農業生態系タイプ,水管理,有機物施用などの各 発生制御要因の寄与を表す拡大係数を求めた(Yan et al., 2005).このことから,水田から発生す る CH4 の算定方法の改訂案を提案した(平成 18 年度研究成果情報,八木ら).この算定方法の改 訂案は 2006 年に改訂された IPCC の新しいガイドラインに Tier 1 デフォルト方法論として採用さ れた(IPCC, 2006) .この方法で見積もられた 2000 年における一年間の水田からの CH4 発生量は 全世界で 25.6Tg(2560 万トン)であり,国別に見ると,中国とインドの合計が世界の約半分を占 め,アジア地域全体で世界の 93%を占めることが明らかになった(図 6).さらに,常時湛水のか んがい水田への間断かんがいの導入,および稲わらすき込み時期の改善により,それぞれ,年間 4.1Tg(410 万トン:CO2 換算量で 1.0 億トン)の削減ポテンシャルのあることが推定された(平成 21 年度研究成果情報,八木ら) (Yan et al., 2009). 上記の実測データをもとにした統計モデルによる発生量評価とは別に,水田における CH4 生 成・発生の各プロセスをコンピューターで予測できる数理モデルの開発が進められた.この研究 では DNDC(DeNitrification DeComposition)という既存の数理モデルを改良し,稲わら処理方法 や水管理の違いによる CH4 発生量の変化を定量的に計算して予測できるモデル(DNDC-Rice モデ ル)を開発した(Fumoto et al., 2008) .日本と中国の 3 地点の水田で行われたほ場実験によって, DNDC-Rice モデルが CH4 発生量を正しく予測できるかどうか検証したところ,従来の DNDC モ デルでは不十分だった稲わらの処理方法や肥料の種類の影響の予測精度が改善され,CH4 発生量 の観測値に対する平均誤差は 4gm-2 以下であった(平成 19 年度研究成果情報,麓ら)(Fumoto et al., 2009) (図 7) .さらに,タイの水田におけるデータを用いた検証も行われた(Smakgahn et al., 2009) .これらのことから,DNDC-Rice モデルを用いることで異なった環境の水田からの CH4 発 生量とその削減ポテンシャルを精緻に広域評価できる可能性が示され,現在,全国評価に向けた作 業が進められている. 第 6 章 温室効果ガス発生抑制と土壌炭素蓄積 83 図 6 世界の水田からのメタン発生量地理分布(モンスーンアジアのみを示す).各国,各地域について算定さ れた水田からの単位面積当たりの CH4 発生量について,緯度経度 5 分の解像度にて地理分布を表した. ផቯ㩷CH4 ឃ㊂㩷(kg C ha-1 ) 400 300 200 ᲧᏓ 100 ጊ 㦖䊱ፒ 0 0 100 200 300 ታ᷹㩷CH4 ឃ㊂㩷(kg C ha-1 ) 400 図 7 DNDC-Rice モデルの検証結果.3 地点において,そ れぞれ年次,水管理または有機物投入量を変えて測 定した水稲栽培期間 CH4 発生量を推定した. ⑵ 施 肥 窒 素 か ら の 一 酸 化 二 窒 素 ( N 2O ) 発 生 畑地や草地などの農地土壌では,窒素施肥,耕起および降雨にともなった特徴的な N2O 発生 パターンを示す.図 8 は,北海道東部の採草地において硫酸アンモニウムを施肥した場合の調査 結果である(甲田ら,2002) .2 年間の調査において,N2O フラックスは窒素施肥後および牧草収 84 4.発生量評価とモデル開発 2000ᐕ ㅊ⢈ -1 100 -2 N2 O Flux(㱘gNm hr ) 120 80 60 40 2001ᐕ ㅊ⢈ ⓠ ᣉ⢈ ᣉ⢈ ⓠ ⓠ ⓠ 20 0 -20 4/15 6/14 8/13 10/12 4/10 ᷹ቯᣣ 6/9 8/8 10/7 図 8 チモシー単播採草地における一酸化二窒素フラックスの推移.施肥時および追肥時に,それぞれ,213 お よび 107 kg N ha−1 相当量の硫酸アンモニウムを施肥. 穫後に高まることが明らかである(平成 13 年度研究成果情報,甲田ら).一般的な農地土壌から の N2O フラックスの変動パターンにおいては,施肥後 1 〜 2 週間程度の間に N2O フラックスが増 加し,施肥後 1 か月程度経過した後はバックグラウンドレベルに戻ることが多い(Akiyama and Tsuruta, 2003) .しかし,このような一般的な発生パターンとは異なり,排水性の悪い転換畑にお いて,施肥から 2 か月後の夏季に長期間にわたる発生が見られたり(Nishimura et al., 2005a),記 録的豪雨にともなって劇的な増加が起きたりした例もある(Akiyama et al., 2011). 一方,化学肥料だけでなく有機肥料も N2O の発生源である.有機肥料は種類が多岐にわたり, また成分のばらつきも大きいことから,有機肥料からの N2O の発生に関する研究は化学肥料に比 べて遅れていた.しかし,農業環境技術研究所温室効果ガス発生制御施設を用いて有機肥料から の N2O 発生量の高精度モニタリングが行われ,施用有機物の種類により N2O 発生量は大きく異な ることが示された.C/N 比が比較的低い有機物〔括弧内の数値は C/N 比〕〔発酵鶏ふん(9.7), 発 酵 豚 ぷ ん(10.7) ,菜 種油かす(9.6) ,魚かす (4.2) 〕 か ら の N2O 発 生 量は化学肥料(尿素)区 よ り も 大 き く, 逆 に, C/N 比 の 高 い 有 機 物 〔牛ふん堆肥(24.3) ,乾 燥 牛 ふ ん(15.9) 〕から の N2O 発 生 量 は 小 さ い ことが明らかにされた (図 9) .さらに,施肥量 等の統計データを用い て N2O の排出量を試算 し た 結 果, わ が 国 の 農 地 に お い て, 有 機 肥 料 の施用は化学肥料の施 図 9 有機物を施用した黒ボク土畑からの N2O 発生量.農業環境技術研究所 温室効果ガス発生制御施設での連続測定データのうち,施用後 30 日間 の発生量を示した.有機物区は右側より順に 2 種類ずつ 3 年間にわたっ て得られたデータである.対象区として化学肥料(尿素,3 年間の平均 値)を用いた.施用量は全窒素として各区 150kgN/ha であり,全量を 基肥とし,全面全層に施用した.作物はチンゲンサイである. 第 6 章 温室効果ガス発生抑制と土壌炭素蓄積 85 用に匹敵する N2O の主要な排出源のひとつであることが示された(平成 15 年度研究成果情報,秋 山ら) (Akiyama and Tsuruta, 2003) .また,家畜ふん堆肥の取り扱いを容易にし,広域流通を進 める方策として実用化されているペレット堆肥について,農地土壌に施用した場合の N2O 発生量 を計測した結果,通常の堆肥よりもペレット堆肥からの N2O 発生量の方が多いことを明らかにし た.その原因として,ペレット内部が嫌気的になることにより脱窒が促進されるためと考えられた (Hayakawa et al., 2009) . 熱帯アジアでは,人口の急増とともに熱帯林や泥炭湿地帯などの陸域生態系が消失して農耕地や プランテーションになるという土地利用変化が急速に起こっている.それにともなって,温室効果 ガスの発生・吸収量が大きく変化すると推測され,研究の重要性と緊急性が指摘されていた.この 問題に対して,スマトラ島の湿潤熱帯林とその周辺地域における現地調査から,土地利用変化によ り熱帯林が伐採・焼却されると,N2O 発生が急増すること,およびその後ゴムを植栽すると N2O 発生量は次第に減少し,約 2 年半後に伐採・焼却以前の値に戻ることが計測された(平成 13 年度 研究成果情報,鶴田ら) (Ishizuka et al., 2002;Nakajima et al., 2005). 施肥窒素由来の N2O 発生プロセスとして,農地土壌表面から直接大気へ発生する直接発生以外 に,農業地帯の地下水や河川水からの脱ガスによる間接発生が指摘されている.このプロセスにお ける N2O の生成過程や発生量については,十分明らかにはされていないが,その地球規模の発生 量は土壌からの直接発生量に匹敵する可能性が指摘され,重要な未解明の発生源とされている.農 業環境技術研究所温室効果ガス発生制御施設のライシメーターによる計測では,灰色低地土での陸 稲や大豆栽培において,土壌表面からの直接発生量の 50 〜 67%に相当する N2O が浸透水に溶存し て地下 1 m 以深に溶脱したことが定量された(Minamikawa et al., 2010). 農地からの N2O 発生量は,一般に,施用した窒素量に伴って増加するので,施用窒素量に対す る N2O-N 発生量の割合である排出係数が発生量の見積もりに用いられる.京都議定書第 1 約束期 間に使用される 1996 年版 IPCC ガイドラインでは,標準的な直接発生の排出係数(EF1)のデフォ ルト値として 1.25%が採用されていた(IPCC, 1996)(この値は 2006 年版では 1.0%に改訂された (IPCC, 2006) ) .しかし,水田からの排出係数(EF1-RICE)は畑や草地の排出係数と同じとされてい るが,湛水条件での N2O の発生量は畑条件とは大きく異なることや,溶脱による間接発生の排出 係数(EF5)が過大評価である可能性が高いことが指摘されていた.そこで,世界の水田から直 接発生する N2O の実測データ(29 地点,149 測定)を収集し統計解析した結果,EF1-RICE の平均 値(±標準偏差)は施肥窒素量の 0.31(± 0.31)%であり,IPCC のデフォルト値よりも著しく低 いことが明らかにされた(Akiyama et al., 2005).また,世界の農地から溶脱により間接発生する N2O 発生量のデータベースを作成し解析した結果,EF5 の構成要素のひとつである農地から系外に 排出された窒素が地下水や表面流去水において N2O を排出する際の係数(EF5g)は窒素溶脱量の 0.24%(95%信頼区間:0.18% ― 0.29%)と推定され,IPCC の推定値(1.5%)よりも著しく低かっ た(Sawamoto et al., 2005) .この値に,河川および沿岸域における既存の排出係数(それぞれ, EF5r = 0.75%および EF5e = 0.25%)を加えることから,溶脱による N2O 間接発生の排出係数(EF5) を 1.24%と見積もった(平成 17 年度研究成果情報,秋山ら).これら 2 つの排出係数は,2006 年版 IPCC ガイドラインにおいて,それぞれデフォルト値として採用された(IPCC, 2006). わが国の農地土壌からの N2O 排出量算定では,国独自の直接排出係数(EF1)を用いていたが, バックグラウンド排出量が差し引かれておらず,作物種によっては限られた数のデータから排出係 数が求められているなどの問題点が指摘されていた.そこで,わが国における観測データを検討し た結果,多くの場合,発生係数は IPCC ガイドラインのデフォルト値より低いが,茶園土壌等の一 86 4.発生量評価とモデル開発 部の例ではできわめて高い発生が見られることが明らかになった.これらを統計解析し,日本の 畑地と草地(茶園以外) ,水田,および茶園からの EF1 は,それぞれ 0.62%,0.31%および 2.9%と 算定された(Akiyama et al., 2006) .これらの結果と,間接排出についての解析結果をあわせ,わ が国の農地に関する新しい排出係数を提案した(平成 18 年度研究成果情報,秋山ら)(表 1).こ の改訂案は,日本国温室効果ガスインベントリ報告書に採用された(温室効果ガスインベントリオ フィス,2010) . 表 日本国温室効果ガスインベントリ報告書に採用された一酸化二窒素排出係数 排出源区分* 作物種 排出係数 不確実性 出典・根拠 (kg N2O−N/kg N)(kg N2O−N/kg N) 水稲 0.31% 茶 2.90% ± 1.8% その他の作物 0.62% ± 0.48% 作物残渣 1.25% 0.25%−6% IPCC デフォルト値 間接排出(大気沈降) 1.00% ± 0.5% IPCC デフォルト値 間接排出(溶脱・流出) 1.24% 0.6%−2.5% 文献 38,39) 合成肥料および 有機質肥料 ± 0.31% 文献 37,39) * 有機質土壌の耕起については,IPCC デフォルト値(排出係数:8kg N2O−N/ha /yr;不確実性:1−80kg N2O−N/ ha/yr)の使用を提案した。 さらに,これらの排出係数と 1985 〜 2005 年までの 5 年ごとに求めた農業生産に伴う窒素フロー データベースから,わが国の農業セクターからの N2O 発生量を都道府県別,排出源別に算定した 結果,全国合計の発生量は 1990 年の 25,749MgN から 2005 年には 18,907MgN に減少したことが示 された.1990 年と 2005 年に関して比較すると,農地への窒素施用量は減っていても発生する家畜 ふん尿窒素が増えている都道府県では,N2O 発生量が増加している傾向が明らかとなった.日本 全体で見た場合,畜産セクターから発生する N2O 量は耕種セクターより多く,家畜ふん尿の処理 に伴う N2O 発生が地域の発生量をより強く制御していると考えられた(Mishima et al., 2011). ⑶ 土 壌 炭 素 蓄 積 と 農 地 か ら の 二 酸 化 炭 素 ( C O 2) 発 生 ・ 吸 収 土壌圏の炭素収支は図 1 に示された土壌への炭素の入力量と出力量のバランスから決定され,土 壌は大気 CO2 の吸収源とも発生源ともなりうる.そして,その正味の発生・吸収量は主に有機物 として固定された土壌炭素蓄積量の変化に反映される.このことから,土壌炭素量を長期に追跡す ることにより,その間の農地からの CO2 発生・吸収量を推定可能である.わが国では,農林水産 省土壌保全対策事業の一環として,1979 〜 1998 年度の間,全国 20,000 地点近くで 5 年ごとに土壌 環境基礎調査(定点調査)が行われており,その中に土壌炭素の調査項目がある.この調査は, 1999 年度以降も調査地点数は約 5,700 地点に減少したものの,土壌機能モニタリング調査として継 続し,現在は土壌由来温室効果ガス計測・抑制技術実証普及事業として土壌炭素の全国モニタリン グを主な目的に行われている.これらの調査データと,1959 〜 1978 年度に実施された地力保全基 本調査の農耕地土壌図と 1992 年版基盤整備調査図等を用いて作成した 1992 年における全国版デジ タル農耕地土壌図(高田ら,2009)から,表層から深さ 30 cm までの農地土壌炭素賦存量の全国 分布図を 1 km メッシュで作成した.現在,国連気候変動枠組み条約の基準年である 1990 年時点 でのわが国の農地土壌の炭素蓄積量の評価と,基準年以降の農地土壌炭素蓄積量の変動解析が進め られている. 土壌炭素収支を定量的に変動評価する方法として,このようなモニタリング調査データの解析は 第 6 章 温室効果ガス発生抑制と土壌炭素蓄積 87 有用であるものの,さまざまな気候帯や農地管理において面的な評価を行うことは限界がある.ま た,将来予測を行うことも困難である.そのため,気象,土壌要因,農地管理の要因を含む土壌炭 素動態モデルを用いた研究が進められている.これに対し,広域評価に適用可能な,簡便かつ信頼 性が高い Rothamsted Carbon モデル(RothC モデル)について,わが国の農地土壌における適合 性を検証した.各地のほ場における有機物連用試験結果に RothC モデルを適用し,炭素蓄積量の 実測値とモデルの予測値とを比較した結果,非黒ボク土畑では予測値と実測値が一致した.しか し,黒ボク土畑および水田では,モデルの予測値は実測値を大きく下回った.黒ボク土は腐植が安 定な形で存在するために分解が遅いことを反映させるため,モデルにピロリン酸塩可溶アルミニウ ム含量(Alp)により腐植画分の分解率を変化させるファクター F(F = 2.50 Alp(%)+ 1.20)を 導入した.その結果,この式を使った改良 RothC モデルでは予測値と実測値が一致した(Shirato et al., 2004) (平成 13 年度研究成果情報,白戸ら)(図 10).一方,水田土壌については水稲耕作期 間の嫌気環境や土壌微生物組成の違いを考慮して,湛水期間および非湛水期間に個別の土壌有機物 分解率を低下させる係数を導入して改良した(Shirato and Yokozawa, 2005)(平成 22 年度研究成 果情報,白戸ら) .これらのことから,黒ボク土および非黒ボク土の畑用のモデルと水田用のモデ ルを併用することが可能になり,わが国の農地に適用可能になるとともに,田畑輪換が土壌炭素の 変動に及ぼす影響を同一のモデルを用いて評価できる. 図 10 (左図)非黒ボク土畑土壌における全炭素量の実測値と RothC モデルによる予測値の比較.(右図)黒 ボク土畑土壌における全炭素量の実測値と RothC モデルおよび改良 RothC モデルによる予測値の比較. 5.発生抑制技術の開発とその評価 ⑴ 水 田 か ら の メ タ ン ( C H 4) 発 生 抑 制 技 術 水田からの CH4 発生削減方策として,中干しや間断かんがいによる水管理,稲わらの堆肥化や 非湛水期間での分解を促進する有機物管理,肥料または資材の使用,土壌改良など,候補となる技 術が数多く提案されている(八木,2004) .特に,水管理については,中干し期間を長くして水田 土壌をより酸化的にすることによりメタン生成菌の活動を抑制し,水田からの CH4 発生削減に効 果のあることが明らかにされている.農業環境技術研究所温室効果ガス発生制御施設での計測か ら,水稲生育前期の常時湛水期間に増加した CH4 発生が,その後の中干し・間断かんがいに伴っ て大幅に減少することが明らかにされた(図 11).一方,水稲栽培期間における有意な N2O 発生 は,代かき直後,間断かんがい中の追肥直後,および最終落水後(水稲収穫直前)に一時的に観 測されただけであり,その量はこれまで報告された世界の水田での研究例の中でも最も少ないも 88 5.発生抑制技術の開発とその評価 図 11 水田からのメタン(CH4) ・一酸化二窒素(N2O)放出量の経日変化(上 段) ,および土壌中のアンモニア態窒素(NH4-N) ・硝酸態窒素(NO3-N)の 経日変化(下段) .斜線部は湛水期間を示す. のであった.これらの結果から,水田の中干しおよびそれに引き続く間断かんがいと 90kg N/ha 程度の窒素施肥は,CH4 発生量を大幅に減少させるだけでなく,N2O 発生の促進もほとんどな いことから,水田における温室効果ガス総発生量の削減技術として有効であることが実証された (Nishimura et al., 2004) (平成 14 年度研究成果情報,八木ら).また,水田を転換畑とすることに より CH4 発生はなくなるが,N2O 発生が増加するトレードオフが認められた.この場合,CH4 お よび N2O の年間発生量合計値は,CO2 等価量として,水稲区,陸稲単作区および大豆 ― 小麦二毛 作区で,それぞれ,100 〜 442,102 〜 110,および 79 〜 146g/m であり,畑転換による CH4 発生 の消失と N2O 発生量の増大が著しかった.しかし,ここで得られた水稲区からの CH4 年間発生量 は日本国温室効果ガスインベントリ報告書(温室効果ガスインベントリオフィス,2010)で採用さ れているわが国の平均値(19.0 ± 12.5g/m)よりも低かった.一方,転換畑からの N2O の年間発 生量は,わが国の測定値のなかでは,茶園を除けば比較的高い値であった.これらの結果から,温 室効果ガス発生削減の観点からは,水管理にともなう水田からの CH4 発生量の削減が重要であり, 排水不良や遊離酸化鉄含量の低い土壌など水田からの CH4 発生が多い場合は,畑転換は有効な温 室効果ガス発生抑制技術であることが示された(Nishimura et al., 2004;2005a)(平成 16 年度研 究成果情報,西村ら) .さらに,転換畑を再度水田とした復元田では,その水稲耕作初年の CH4 発 生量は著しく低下することが示された(Nishimura et al., 2011). 水管理による水田からの CH4 発生抑制技術は,現場への普及をめざして,農林水産省生産局事 業(平成 20 〜 21 年度)として全国 9 か所で実証試験が行われた.この試験では,中干しを慣行よ りも 1 週間程度前倒し・延長することにより,ほとんどの試験地で CH4 発生を効果的に削減でき ることが示された(図 12) .慣行水管理に比べた栽培期間全体の CH4 発生量の削減率は平均 30%程 第 6 章 温室効果ガス発生抑制と土壌炭素蓄積 89 度であった(Itoh et al., 2011) .水田 の排水にともなって発生する N2O は 無視できるほど小さかったことから, 日本の主要水田における温室効果ガス 発生の削減に対して,この技術が効果 的であることが明らかになった(平成 22 年度成果情報,須藤ら) .中干しの 強化により,慣行水管理区に比べて精 玄米収量が平均 4%低下する傾向がみ られたが,同時に登熟歩合の向上やた んぱく含量の減少など品質の向上が確 認された. 同様の水管理による水田からの CH4 発生削減効果はインドネシアの 図 12 中干しの強化による水田からのメタン発生削減.図中 の横線は湛水期間を示す. 水田でも確認されている(Hadi et al., 2010) .また,中国の水稲 - 小麦二毛作田では,小麦わらが直後の水稲作においてきわめて多量の CH4 発生を生じる原因となっているが,これを水稲栽培期間に土壌表面に適切に設置することによ り有効な発生削減技術となることを明らかにした(Ma et al., 2008). 一方,水稲耕作における温室効果ガス発生抑制技術の評価においては,土壌からの温室効果ガス 発生以外にも,耕作機械の使用と収穫物乾燥・運搬の際のエネルギー投入や資材または飼料の生産 と使用にともなう温室効果ガス発生などを含めた,システム全体の収支を取り扱うライフサイクル アセスメント(LCA)手法の導入が求められている.この問題に対し,東北地方の水田を対象に 研究がなされ,不耕起移植栽培は代かき移植栽培に比べ,CO2 に換算して約 1,800kg/ha の温室効 果ガス排出削減効果があることが試算された.これは主として CH4 発生量の低減に起因し,耕起, 専用田植機,除草剤など作業体系の影響は小さかった.同時に,無代かき移植栽培の地球温暖化へ の影響は,代かき栽培と同等であることも示された(Harada et al., 2007)(平成 17 年度研究成果 情報,原田ら) . ⑵ 施 肥 窒 素 か ら の 一 酸 化 二 窒 素 ( N 2O ) 発 生 抑 制 技 術 農地土壌から発生する N2O を制御するためには,まず,施肥窒素量を削減するなど,土壌中の アンモニウム態および硝酸態窒素プールをできるだけ小さくし,硝化や脱窒により変換される無機 態窒素量を少なくすることが考えられる.しかし,このことは同時に,作物が吸収できる窒素量を 制限することになる.したがって,より現実的には,作物による無機態窒素吸収効率を高め,無駄 に環境中へ放出される窒素の流れを制御することが望ましい.このことは,N2O や NO などのガ ス発生だけでなく,施肥窒素由来の別の重要な環境問題である地下水の硝酸汚染軽減にもつながる ものである. 作物による施肥窒素の吸収効率を高め,環境への窒素のロスを少なくするためには,作物が必要 なときに必要なだけ窒素を施用する必要がある.そのための技術としては,最適な窒素施肥量と分 施・局所施肥,適切な有機物施用など施用方法の改善設計が基礎となる.また,一般に,窒素肥料 投入量の増加に対して,作物収量はあるところまでは直線的に増加するが,一定量以上では頭打ち になる一方,環境負荷はどこまでも増加を続ける.作物の収量や品質と窒素肥料の投入量との関係 を作物や土壌タイプごとに検討し,土壌の環境容量を超えず,かつ高い収量が維持されるような食 90 5.発生抑制技術の開発とその評価 料生産と環境保全とを調和させるための適正な窒素肥料投入量を示し,広く普及させる努力も必要 であろう.さらに,土壌微生物による土壌中無機態窒素の固定化を促進するために,有機物施用を 促進することも効果的であろう. 別の方策として,肥料の種類を選択することによる N2O 発生の制御も可能である.N2O 発生率 は窒素肥料の形態により異なるが,N2O 発生率の高い無水アンモニアの使用や硝酸態窒素を水分 含量の高い土壌に施用することを避け,発生率の低い形態の肥料を使用することが勧められる.ま た,緩効性肥料や硝化抑制剤入り肥料など新しいタイプの肥料の使用が N2O 発生抑制に効果のあ ることが報告されている.硝化抑制剤には,DCD(dicyandiamide),AM(2-amino-4-chloro-6-methyl pyrimidine)など,さまざまな化合物があるが,これらは,もともと,硝化とそれにより生成され た硝酸態窒素の損失を軽減し,作物の窒素利用効率向上のために使用されてきた農業資材である. 黒ボク土のニンジンほ場における試験では,硝化抑制剤入り肥料を施用することにより,速効性肥 料で見られる施肥直後の多量の N2O 発生を抑制し,その後の発生も低くできることから,大きな N2O 発生削減効果のあることが示された(Akiyama et al., 2000).同様の結果は,中国東北地域の トウモロコシ畑での試験においても確認されている(Xu et al., 2007). このような世界各地で測定 された硝化抑制剤入り肥料に よ る N 2O 発 生 削 減 の デ ー タ ⎣ൻᛥ䉍⢈ᢱ ᐔဋ を集約し,統計解析を行うこ 䌄䌃䌄 とにより,平均的な削減効果 䊆䊃䊤䊏䊥䊮 の評価を行った.その結果, 䉦䊦䉲䉡䊛䉦䊷䊋䉟䊄 硝化抑制剤入り肥料の平均的 䌄䌍䌐䌐 な削減率は慣行肥料の 38%で 䉼䉥䉰䊦䊐䉢䊷䊃 あり,さまざまな環境のほ場 䊆䊷䊛 試験においても比較的安定し た削減効果がみられることが 明らかになった(Akiyama et al., 2010)( 平 成 21 年 度 研 究 䈱ᢙሼ䈲䊂䊷䉺ᢙ ᐔဋ୯䈫95%ା㗬㑆 -100 -80 -60 -40 -20 0 N2O䈱ᷫ₸(%) 図 13 硝化抑制剤による N2O 発生削減効果の統計解析結果 成果情報,秋山ら)(図 13). ⑶ 土壌炭素蓄積技術 農地土壌に炭素を蓄積する,すなわち,農地を大気 CO2 の吸収源に変える方法のひとつは作物 残渣や堆きゅう肥などの有機物を投入することである.この場合,投入された有機物の炭素の大部 分は微生物により分解され,大気へ帰って行くが,一部分は土壌中で腐植などの安定な有機物に変 換され,その結果,土壌からの CO2 発生量は緩和される.さらに,毎年の投入炭素量が分解炭素 量を上回れば,土壌有機物としての蓄積量が増加する.このような有機物管理による土壌炭素貯留 効果については,わが国全国各地における長期連用試験において実証されている.このほか,不耕 起・簡易耕起等,土壌耕起方法の改善,輪作やカバークロップの導入が土壌の炭素蓄積に有効な技 術であることが示されている. このような土壌の炭素蓄積に効果のある農業技術を導入した場合,わが国の農地土壌がどの程 度の炭素貯留効果,すなわち CO2 発生緩和効果を持つか定量評価するため,全国の農地と草地に おける土壌炭素賦存量の分布図を作成するとともに,栽培管理方法を変えた場合の土壌炭素貯留 量の将来予測をプロセスモデル(RothC モデル)を用いて行っている.その結果,全国の水田と 第 6 章 温室効果ガス発生抑制と土壌炭素蓄積 91 畑のすべてに堆肥を投入した場合,未投入に比べて,25 年間で 3200 万トンの炭素が,全国の水田 のすべてに二毛作を導入した場合は 1100 万トンの炭素が,それぞれ増加することを明らかにした (Yokozawa et al., 2010) (平成 21 年度研究成果情報,白戸ら)(図 14). 50 ᧂᛩ䉲䊅䊥䉥䈫䈱Ꮕ䋨M t C䋩 45 40 35 30 ၸ⢈+ੑᲫ 25 20 ၸ⢈ 15 10 ᳓↰ੑᲫ 5 0 1990 1995 2000 2005 2010 2015 図 14 全国の農地における土壌炭素量の変化予測.未投入との差を示す. 堆肥投入量は水田で 10 t/ha,畑で 15 t/ha. 6.今後の展望 過去 10 年間における研究の進展から,農地からの CH4 と N2O の発生の抑制と土壌炭素の蓄積 に関して,新たなモニタリング技術や広域評価のためのモデル開発が行われ,温室効果ガス間のト レードオフを考慮した総合的な温暖化緩和策の広域定量評価が可能となった.今後,これらの研究 蓄積を,より精緻で,より幅広い評価項目を含み,より広域に展開可能な研究へ発展させることが 期待される. 新たに得られた観測データと活動量データを加えたデータベースをさらに整備するとともに,モ デルや将来予測シナリオを改良することにより,わが国農地におけるさまざまな温暖化緩和策の定 量評価を精緻化することが期待される.また,温暖化緩和策のライフサイクルアセスメントを進め るともに,他の環境影響や生産性との総合評価を行うエコバランス評価手法を開発することが求め られている.多くの緩和策は,土壌肥沃度の増進,有機物資源のリサイクル推進,化学肥料施用量 の削減など,わが国で進められている環境保全型農業の推進方向と一致する技術であることから, 温暖化緩和を含めた総合的な農業生産環境評価へ発展させる新たな研究展開をめざすべきである. その際,緩和策のコスト評価等,社会経済評価との融合も必要である. 水田の水管理,硝化抑制剤,有機物施用による土壌炭素蓄積など,これまでに開発された緩和技 術はすでにわが国の農家に普及可能な段階にある.このような技術について,普及に向かうインセ ンティブを農家に与えるため,施策として検討されている行政による資金援助,カーボンフットプ リントによる認証,排出権取引といった制度の推進に寄与することを目的に,研究成果を地域や農 家レベルで利用できる方法の開発も必要であろう.たとえば,現場での普及を進めるための方法論 構築に関する研究,パソコン等でユーザーがその農地の温室効果ガス排出量と緩和可能量を計算で 92 引用文献 きる意志決定支援ツールの開発が期待される.その際,都道府県試験研究機関など地域の専門家, 普及組織,あるいは民間団体や企業とのこれまで以上の連携が求められる. さらに,発展途上国での温暖化緩和策の展開に対する潜在的可能性に着目すべきである.わが国 の温室効果ガス排出インベントリーに占める農業分野の割合は低いが,同じ水田耕作を基礎とする 農業体系を持つ熱帯アジア諸国では,農業分野の占める割合はきわめて大きく,農業分野からの温 室効果ガス排出抑制は優先度が高い.特に広大な農耕地を持ち,家畜頭数の多大な国では,農業技 術の適用による緩和策は大きな貢献が可能である.さらに,このような国においては,その適用に ついて,持続的開発政策と一致させることにより,削減の可能性をいっそう前進させると予測され る.京都議定書において設定されているクリーン開発メカニズム(CDM)は,新たな開発援助の ツールとして活用出来る可能性がある.農業環境技術研究所では,これまでもアジア諸国を中心 に各国の研究者と共同研究を展開すると同時に,IPCC,国際窒素イニシアティブ(INI),農業分 野の温室効果ガスに関するグローバル・リサーチ・アライアンス(GRA)などの国際研究ネット ワークに積極的に参加してきた.このような国際連携の強化がますます求められている. 引用文献 1)Akiyama, H., Tsuruta, H. and Watanabe, T.(2000)N2O and NO emissions from soils after the application of different chemical fertilizers. Chemosphere Global Change Science, 2, 313-320 2)Akiyama, H. and Tsuruta, H.(2003)Effect of organic matter application on N2O, NO and NO 2 fluxes from Andisol field. Global Biogeochemical Cycles, 17( 4), 1100, doi:10.1029/ 2002Gb002016 3)Akiyama, H., Yagi, K. and Yan, X.(2005)Direct N 2O emissions from rice paddy fields: summary of available data. Global Biogeochemical Cycles, 19, GB1005, doi:10.1029/ 2004GB002378 4)Akiyama, H., Yan, X. and Yagi, K.(2006)Estimations of emission factors for fertilizer-induced direct N2O emissions from agricultural soils in Japan: summary of available data. Soil Science and Plant Nutrition, 52, 774-787 5)Akiyama, H., Hayakawa, A., Sudo, S., Yonemura, S., Tanonaka, T. and Yagi, K.(2009)A portable automated sampling system for long-term monitoring of nitrous oxide and methane fluxes from soils. Soil Science and Plant Nutrition, 55, 435-440 6)Akiyama, H., Yan, X. and Yagi, K.(2010)Evaluation of effectiveness of enhanced-efficiency fertilizers as mitigation options for N2O and NO emissions from agricultural soils: metaanalysis. Global Change Biology, 16, 1837-1846 7)Akiyama, H., Morimoto, S., Hayatsu, M., Hayakawa, A., Sudo, S. and Yagi, K.(2011)Explosive N2O emission and abundance of ammonia-oxidising bacteria and archaea after fertilization. Journal of Environmental Quality, submitted 8)秋山博子,澤本卓治,八木一行(2011)窒素循環と土壌からの N2O 発生.化学と生物,印刷 中 9)Cheng, W., Sakai, H., Hartley, A., Yagi, K. and Hasegawa, T.(2008)Increased night temperature reduces the stimulatory effect of elevated carbon dioxide concentration on methane emission from rice paddy soil. Global Change Biology, 14, 1-13 第 6 章 温室効果ガス発生抑制と土壌炭素蓄積 93 10)Chu, H., Morimoto, S., Fujii, T., Yagi, K. and Nishimura, S.(2009)Soil ammonia-oxidizing bacteria in paddy rice fields as affected by upland conversion history. Soil Science Society of America Journal, 73, 2026-2031 11)Fumoto, T., Kobayashi, K., Li, C., Yagi, K. and Hasegawa, T.(2008)Revising a process-based biogeochemistry model DNDC to simulate methane emission from rice paddy fields under various residue managements. Global Change Biology, 14, 382-402 12)Fumoto, T., Yanagihara, T., Saito, T. and Yagi, K.(2009)Assessment of the methane mitigation potentials of alternative water regimes in rice fields using a process-based biogeochemistry model. Global Change Biology, doi: 10.1111/j. 1365-2486.2009.02050.x 13)Hadi, A., Inubushi, K. and Yagi, K.(2010)Effect of water management on greenhouse gas emissions and microbial properties of paddy soils in Japan and Indonesia. Paddy Water Environment, 8, 319-324 14)Harada, H., Kobayashi, H. and Shindo, H.(2007)Reduction in greenhouse gas emissions by no-tilling rice cultivation in Hachirogata polder, northern Japan: Life-cycle inventory analysis. Soil Science and Plant Nutrition, 53, 668-677 15)Hayakawa, A., Akiyama, H., Sudo, S. and Yagi, K.(2009)N2O and NO emissions from an Andisol field as influenced by pelleted poultry manure. Soil Biology and Biochemistry, 41, 521-529 16)Hayatsu, M., Tago, K. and Saito, M.(2008)Various players in the nitrogen cycle: Diversity and functions of the microorganisms involved in nitrification and denitrification. Soil Science and Plant Nutrition, 54, 33-45 17)Hiradate, S., Nakadai, T., Shindo, H. and Yoneyama, T.(2004)Carbon source of humic substances in some Japanese volcanic ash soils determined by carbon stable isotopic ratio, δ13C. Geoderma, 119, 133-141 18)Inubushi, K., Cheng, W., Aonuma, S., Hoque, M., Kobayashi, K., Miura, S., Kim, H. and Okada, M. (2003)Effects of free-air CO2 enrichment(FACE)on CH4 emission from a rice paddy field. Global Change Biology, 9, 1458-1464 19)IPCC(1996)Revised 1996 IPCC Guidelines for National Greenhouse Gas Inventories, IPCC 20)IPCC(2006)2006 IPCC Guidelines for National Greenhouse Gas Inventories. Prepared by the National Greenhouse Gas Inventories Programme, IGES, Japan 21)IPCC(2007a)Climate Change 2007: Synthesis Report. Contribution of Working Groups I, II and III to the Fourth Assessment Report of the Intergovernmental Panel on Climate Change. IPCC, Geneva, Switzerland, 104 pp. 22)IPCC(2007b)Climate Change 2007: Mitigation. Contribution of Working Group III to the Fourth Assessment Report of the Intergovernmental Panel on Climate Change. Cambridge University Press, Cambridge, United Kingdom and New York, NY, USA., 890 pp. 23)Ishizuka, S., Tsuruta, H. and Murdiyarso, D.(2002)An intensive field study on CO2, CH4, and N2O emissions from soils at four land-use types in Sumatra, Indonesia. Global Biogeochemical Cycles, 16, 1049, doi:10.1029/2001GB001614 24)Itoh, M., Sudo, S., Mori, S., Saito, H., Yoshida, T., Shiratori, Y., Suga, S., Yoshikawa, N., Suzue, Y., Mizukami, M., Mochida, T. and Yagi, K.(2011)Mitigation of methane emissions from 94 引用文献 paddy fields by prolonging mid-season drainage. Agriculture Ecosystem and Environment, submitted 25)Jumadi, O., Hala, Y., Anas, I., Ali, A., Sakamoto, K., Saigusa, M., Yagi, K. and Inubushi, K.(2008) Community structure of ammonia-oxidizing bacteria in acid tea soils and their potential to produce nitrous oxide and carbon dioxide. Geomicrobiology Journal, 25, 1-9 26)甲田裕幸,宝示戸雅之,三木直倫,三枝俊哉(2002)北海道東部の採草地における亜酸化窒素 の発生時期および発生量.北海道農業,2001, 268-269 27)駒田充生,竹内 誠(2003)野菜腐敗過程からの亜酸化窒素発生.日本土壌肥料学雑誌,74, 445-451 28)Kool, D.M., Wrage, N., Zechmeister-Boltenstern, S., Pfeffer, M., Brus, D., Oenema, O. and Van Groenigen, J.W.(2010)Nitrifier denitrification can be a source of N2O from soil: a revised approach to the dual-isotope labelling method. European Journal of Soil Science, 61, 759-772 29)Ma, J., Xu, H., Yagi, K. and Cai, Z.C.(2008)Methane emission from paddy soils as affected by wheat straw returning mode. Plant and Soil, 313, 167-174 30)Minamikawa, K., Nishimura, S., Sawamoto, T., Nakajima, Y. and Yagi, K.(2010)Annual emissions of dissolved CO2, CH4, and N2O in the subsurface drainage from three cropping systems. Global Change Biology, 16, 796-809 31)Mishima, S., Akiyama, H. and Yagi, K.(2011)Recent trends in agricultural nitrous oxide emission and its variation among Japanese prefectures and agricultural categories. Nutrient Cycling in Agroecosystems, submitted 32)Nakajima, Y., Ishizuka, S., Tsuruta, H., Iswandi, A. and Murdiyarso, D.(2005)Microbial processes responsible for nitrous oxide production from acid soils in different land-use patterns in Pasirmayang, central Sumatra, Indonesia. Nutrient Cycling in Agroecosystems, 71, 33-42 33)Nishimura, S., Sawamoto, T., Akiyama, H., Sudo, S. and Yagi, K.(2004)Methane and nitrous oxide emissions from a paddy field with Japanese conventional water management and fertilizer application. Global Biogeochemical Cycles, 18, GB2017, doi: 10.1029/2003GB002207 34)Nishimura, S., Sawamoto, T., Akiyama, H., Sudo, S., Cheng, W. and Yagi, K.(2005a) Continuous, automated nitrous oxide measurements from paddy soils converted to upland crops. Soil Science Society of America Journal, 69, 1977-1986 35)Nishimura, S., Sudo, S., Akiyama, H., Yonemura, S., Yagi, K. and Tsuruta, H.(2005b) Development of a system for simultaneous and continuous measurement of carbon dioxide, methane and nitrous oxide fluxes from cropfields based on the automated closed chamber method. Soil Science and Plant Nutrition, 51, 557-564 36)Nishimura, S., Akiyama, H., Sudo, S., Fumoto, T., Cheng, W. and Yagi, K.(2011)Combined emission of CH4 and N2O from paddies was reduced by preceding upland crop cultivation. Soil Science and Plant Nutrition, in press 37)野内 勇(2005)農業分野における温室効果ガスの放出削減.農業環境研究叢書第 16 号,農 業環境研究 20 年の歩み,独立行政法人農業環境技術研究所,p. 20-41 38)温室効果ガスインベントリオフィス(GIO)(2010)日本国温室効果ガスインベントリ報告 書.独立行政法人国立環境研究所,http://www-gio.nies.go.jp/index-j.html 第 6 章 温室効果ガス発生抑制と土壌炭素蓄積 95 39)Sawamoto, T., Nakajima, Y., Kasuya, M., Tsuruta, H. and Yagi, K.(2005)Evaluation of emission factors for indirect N2O emission due to nitrogen leaching in agro-ecosystems. Geophysical Research Letters, 32, L03403, doi: 10.1029/2004GL021625 40)Shirato, Y., Hakamata, T. and Taniyama, I.(2004)Modified Rothamsted carbon model for Andosols and its validation: Changing humus decomposition rate constant with pyrophosphate-extractable Al. Soil Science and Plant Nutrition, 50, 149-158 41)Shirato, Y. and Yokozawa, M.(2005)Applying the Rothamsted Carbon Model for long-term experiments on Japanese paddy soils and modifying it by simple tuning of the decomposition rate. Soil Science and Plant Nutrition, 51, 405-415 42)Shoji, S., Nanzyo, M. and Dahlgren, R.A.(1993)Volcanic Ash Soils. Elsevier Science Publishers, Amsterdam, 288 pp 43)Smakgahn, K., Fumoto, T. and Yagi, K.(2009)Validation of revised DNDC model for methane emissions from irrigated rice fields in Thailand and sensitivity analysis of key factors. Journal of Geophysical Research, 114, G02017, doi: 10.1029/2008JG000775 44)高田裕介,中井 信,小原 洋(2009)1992 年の農耕地分布に基づくデジタル農耕地土壌図 の作成.日本土壌肥料学雑誌,80, 502-503 45)Tokida, T., Fumoto, T., Cheng, W., Matsunami, T., Adachi, M., Katayanagi, N., Matsushima, M., Okawara, Y., Nakamura, H., Okada, M., Sameshima, R. and Hasegawa, T.(2010)Effects of free-air CO2 enrichment(FACE)and soil warming on CH4 emission from a rice paddy field: impact assessment and stoichiometric evaluation. Biogeosciences, 7, 2639-2653 46) Wassmann, R., Neue, H.-U., Lantin, R.S., Buendia, L.V. and Rennenberg, H. (2003) Characterization of Methane Emissions from Rice Fields in Asia. I. Comparison among Field Sites in Five Countries. Nutrient Cycling in Agroecosystems, 58, 1-12 47)Xu, H., Chen, G. and Yagi, K.(2007)Mitigation of N 2O emission from a maize field by applying commercially available fertilizer additives containing nitrification and urease inhibitors. Abstracts of Nitrogen 4th Conference, p. 258 48)八木一行(1997)温室効果ガス発生・吸収量.土壌環境分析法 , 博友社 , p. 129-138 49)八木一行(2004)大気メタンの動態と水田からのメタン発生.農業環境研究叢書第 15 号,農 業生態系における炭素と窒素の循環 , 独立行政法人農業環境技術研究所 , p. 23-50 50)八木一行(2009)農耕地からの温室効果ガス発生削減の可能性.シリーズ 21 世紀の農学,地 球温暖化問題への農学の挑戦,養賢堂,p. 127-148 51)Yan, X., Yagi, K., Akiyama, H. and Akimoto, H.(2005)Statistical analysis of the major variables controlling methane emission from rice fields. Global Change Biology, 11, 1131-1141 52)Yan, X., Akiyama, H., Yagi, K. and Akimoto, H.(2009)Global estimations of the inventory and mitigation potential of methane emissions from rice cultivation conducted using the 2006 IPCC Guidelines. Global Biogeochemical Cycles, doi: 10.1029/2008GB003299 53)Yokozawa, M., Shirato, Y., Sakamoto, T., Yonemura, S., Nakai, M. and Ohkura, T.(2010) Use of the RothC model to estimate the carbon sequestration potential of organic matter application in Japanese arable soils. Soil Science and Plant Nutrition, 56, 168-176 54)財団法人日本土壌協会(1996) ,土壌生成温室効果等ガス動態調査報告書(概要編) 96 Ⅲ.農業と生物多様性 ― 豊かな農業・農村環境をめざして 第 7 章 農業生態系における生物多様性の評価 1.はじめに 生命の誕生以来,数十億年の歳月を経て,様々な環境に適応した生物は多様に進化してきた.こ れらの生物は相互の関係をもって生存している.そして人類もまた生物であり,他の生き物との関 係の中で生きている.衣食住に必要な食料,繊維,木材,生存に必要な空気や水などは生物の恵み として得られるものであり,人類の生存にとって生物の多様性は不可欠である. 農業もまた,本来,自然と対立する形でなく,順応する形で自然に働きかけ,上手に利用し,循 環を促進することによってその恵みを享受する生産活動であり,生物多様性と自然の物質循環が健 全に維持されることで成り立つものである(農林水産省,2007).一方,農業は,それぞれの地域 の自然環境の中で長年にわたって活動を継続してきたことにより,原生自然とは異なる特有の農業 生態系を形成し,里山に代表される二次的自然を生み出してきた.赤とんぼやメダカなど身近な生 物はこのような二次的自然に適応してきたものであり,人間もその中で地域固有の文化を育んでき た.しかしながら,近年の集約的な農業の進展にともなう不適切な農薬・肥料の使用や効率性を優 先した過度の農地整備などは,農業生態系に環境負荷を与えるとともに,生物多様性に対しても負 の影響を及ぼしてきた.また,近年の生活様式の変化や急激なグローバリズムの進展,高齢化,農 村の過疎化などの社会的な変化を背景として農業活動が低下し,それが耕作放棄地の増加や里山の 利用低下を引き起こし,さらには農業生態系の身近な生き物の減少や深刻な鳥獣害の原因となって いる.2010 年に名古屋で開催された生物多様性条約第 10 回締約国会議(COP10)では,農業の生 物多様性に関する決議が採択されるとともに,二次的自然に着目し,人間と自然環境の持続可能な 関係の再構築を目指す SATOYAMA イニシアティブの国際パートナーシップが発足するなど,農 業生態系における生物多様性の重要性が国際的にも認識されつつある.われわれが今後,健全で健 康な生活を営むためには,持続可能な農業の発展が不可欠であるが,そのためには,農業生態系に おける生物多様性の実態の把握とその維持メカニズムの解明を通して,生物多様性の保全と調和し た農業活動の方策を提示することが必要である.農業環境技術研究所ではこれまで,農業生態系の 構造と機能の解明およびその評価に関する研究を実施してきた.そこで本章では,農業環境技術研 究所が独立行政法人に移行して以後の 10 年間に実施してきた農業生態系における生物多様性に関 する研究成果を中心に, (1)休耕田・耕作放棄地における生物多様性,(2)農業活動による生物多 様性の維持, (3)農業生態系の生物多様性に対する各種環境要因の影響,(4)長期・広域スケール における生物多様性の評価,に分けて紹介することとする. 2.休耕田・耕作放棄地における生物多様性 1970 年代から始まった減反政策や農家の高齢化の進展の結果,全国的に耕作放棄地や休耕田が 増加した.農地の管理が縮小または放棄されたこのような場所では,休耕・耕作放棄後の年数の経 過とともに植生遷移が進行するが,遷移の方向は水管理のやり方やそこの土壌の乾湿が大きく作用 することが指摘されている(清水,1998) .そこで,茨城県南部の休耕・耕作放棄水田において, 植物群落を調査して植生群落タイプを区分するとともに,土壌図,地質図,表層地質図のオーバー 第 7 章 農業生態系における生物多様性の評価 97 レイ解析に基づく自然立地区分,および地権者へのヒアリングに基づく管理履歴調査を行い,自然 立地区分,人為的条件,群落タイプ類型の対応関係を解析し整理した.その結果,形成される群 落タイプと自然立地区分に明確な対応関係が認められる場合があったが,明確ではない場合でも, 管理履歴を考慮することによって,群落タイプの成立要因を明らかにできることがわかった(図 1) .特に,休耕後の耕起の頻度と引水管理による土壌水分レベルが群落タイプの成立に大きく寄与 すること,植物種の多様性に富んだ良好な湿地性の植物群落は 3 年に 1 回程度の適度な耕起と水分 ポテンシャルを湿潤に保つことで維持できること,大規模な圃場整備によって乾田化した休耕・耕 作放棄水田であっても,適度な粗放的管理と引水管理により土壌水分レベルを常に湿潤に保つこと で種多様性に富む群落を創出できる可能性があることが明らかとなった(楠本ら,2005). 図 1 休耕・耕作放棄水田における植生タイプと管理履歴との対応模式図 休耕田や耕作放棄田は湿地植生の代替生息地として活用できる可能性が指摘されている(清水, ぐ 1998) .そこで,準絶滅危惧種であるタコノアシ Penthorum chinense Pursh. を例として,群落維持 に関係する環境条件を把握し,湿地植生の保全の指針に向けた検討を行った.本種は東アジアに自 生し,泥質の河川沿いや沼沢地に分布するが,休耕田や放棄水田にも分布することが知られるよ うになった(Ikeda et al., 2001) .タコノアシの発芽・成長特性や散布特性などの生態的特性と休耕 田・放棄水田の環境条件との関連を明らかにするため,茨城県つくば市北部の筑波稲敷台地の谷 底平野において,タコノアシの分布と生息水田の微細な形状を調査するとともに,光環境や土壌硬 度など種子発芽に関係する環境条件,およびタコノアシの潜在的分布を明らかにするための埋土種 子調査を行った.その結果,これらの放棄水田では,隣接する用水路や周辺の湧水等の水環境およ びほ場内の微細な起伏による水分条件の違いが,タコノアシの分布に影響していることが明らかと なった.また種子は調査水田内のほぼ全面に分布していたが,種子の発芽は本種がギャップ検出機 構を持つかく乱依存種であることを反映して,光環境の悪化やリターの堆積によって制限されてい ることが明らかとなった.このような知見に基づいて,休耕田・放棄水田におけるタコノアシの保 全のためには,水管理や草刈りによる競合種排除等の管理が場所によっては必要であると考えられ 98 3.農業活動による生物多様性の維持 た(大黒ら,2003) . また,水田や休耕田,河川の湿性植物に対して,水田で一般に使用されている除草剤がどのよう な影響を及ぼすかを明らかにするため,タコノアシを対象として水稲用除草剤ベンスルフロンメチ ルの影響を組み込んだ個体群動態モデルを作成した.標準量(処理濃度 115μg/L)のベンスルフ ロンメチルを連用すると仮定すると,水田では耕起と除草剤の影響のため個体群が急速に消失する が,不耕起で除草剤を全く使用しない休耕田でも,休耕を続けると自然遷移が進むことによりタコ ノアシ個体群は消失すると推測された.しかし,耕作と休耕を適度な間隔で転換することにより, 100 年後でも存続するようになることが判明した(図 2,3).この知見は,湿生植物を水田地帯で 保全する際の管理手法の指針として有効であると思われる(平成 22 年度研究成果情報,池田ら). 㪈㪇㪇ᐕᓟ䈱ᢙ䋨㪉㪇ᐕ㑆䈱ᐔဋ䋩 㩿㬍㪈㪇6㪆㫄2) 㪌 4 3 2 1 0 0 図 2 水田と休耕田での個体数変化 㪌 10 ㅪ⛯䈜䉎ભ⠹ᐕᢙ䋨ᐕ䋩 㪈㪌 図 3 耕作と休耕を転換する水田における休耕 期間の影響 3.農業活動による生物多様性の維持 かつての農村では,薪や炭の原料となる広葉樹の枝や幹,堆肥の原料となる落ち葉などを里山で 採取し,牛馬の餌となるまぐさを草地で刈っていた.四季折々,毎年繰り返されるこれらの活動が 二次的自然をつくり,赤とんぼやメダカなどの身近な生き物を育んできた.しかし近代農業の発展 とともに薪や炭は石油に,堆肥は化学肥料にとって代わられ,それにともなって人の手が入らなく なった里山や草地などでは植生遷移が進み,かつてそこに生息していた身近な生物の多くが減少し てきている.ところが,現在でも農業上の必要性から二次的自然が,単に保全されているだけでな く,農業上の必要性をもって残されている場所があることがわかってきた. ⑴ 裾刈り草地 かつて農村地域にはススキを主体とする多様な草原性植物から構成される半自然草地が広く見ら れたが,このような半自然草地は,農業形態や生活様式の変化により著しく減少したため,多くの 草原性の動植物が絶滅の危機に瀕している.一方,谷津田では田面が日陰になるのを防ぐため,水 田を取り囲む斜面林の下部で定期的に草刈りが行われる場所(裾刈り草地)(図 4)が現在でも見 られ,そこにはススキを主体とした草地が形成されている.そこで,筑波・稲敷台地のススキを主 体とする様々な草地の植生を調査し,多変量解析を用いることによって,主に谷津田を取り囲む斜 第 7 章 農業生態系における生物多様性の評価 99 面林周辺に分布するタイプ(谷津田型) ,主にアカマツ林の林床に分布するタイプ(松林型),台地 上平坦部にある畑作放棄地や造成跡地に分布するタイプ(平地型)の 3 つに分類した.さらに,そ れらを関東平野の過去(1970 〜 1980 年代)の半自然草地の種組成と比較したところ,谷津田型と 松林型では,過去の半自然草地と同様に在来多年性草本が豊富であるが,特に谷津田の裾刈り草地 には過去の半自然草地に生息していた多くの希少植物が見られることが明らかとなった.これらの ことから,水稲生産のために行われている谷津田を囲む斜面林の周辺の草刈りが,植物群落の多様 性を維持していることが判明した(小柳ら,2007)(平成 18 年度研究成果情報,楠本ら). ⵿ಿ䉍⨲ ⵿ಿ䉍⨲䈱┙ ⵿ಿ䉍⨲䈱ᬀ↢ 図 4 現存の裾刈り草地の例−茨城県阿見町 ⑵ 茶草場 東海地方の茶産地では,良質茶の栽培を目的として茶園にススキの敷草を施す農法が現在でも行 われており,その資材源としての茶草場が大面積で広がっている(図 5).このような茶草場の特 徴を明らかにするため,茶草場で 50 地点,比較対象として造成跡地等でススキが優占する 10 地点 において植生調査を実施し,種組成に基づいて分類した結果,ススキが優占する Gr.1,ネサザが 優占し,主に共有地として維持され,在来の草原性草本が豊富で,多様度指数(H')が高い Gr.2, 造成跡地にみられ,外来植物が侵入している Gr.3,水田跡地にみられ,外来植物が侵入している Gr.4,茶園脇に線状に存在し,多様度指数(H')の低いススキ草地の Gr.5 の 5 つの植物群落タイプ に分けられた.また,すべての調査地点で,土壌サンプリング,光環境の測定,斜面方位・角度お よび土壌水分の測定を行い,土地改変履歴や管理実態については,地権者へのヒアリングで確認し た結果,多様度の高い群落タイプ(Gr.1,Gr.2)が現れる場所は,土壌 pH が低く,一か所あたり の面積が広く,土地改変が行われていないという特徴があり,草原性の植物種群にダメージを与え る大きなかく乱をこれまで受けていないことが示唆された.以上のことから,茶草場における在来 植物の多様性には,土地改変や管理履歴等の歴史性が強く影響を及ぼしていることが明らかとなっ た(平成 21 年度研究成果情報,楠本ら) . 100 4.農業生態系の各種環境要因が生物多様性に及ぼす影響 図 5 茶草場の分布把握 茶園面積: 169.25ha 茶草場面積:110.97ha 4.農業生態系の各種環境要因が生物多様性に及ぼす影響 農業生態系における生物多様性を保全するためには,生物多様性の実態を把握するとともに,農 業生態系の景観構造や土地利用形態など各種環境要因が生物多様性にどのような影響を及ぼしてい るかを明らかにする必要がある.そこで,農業生態系における生物多様性の調査を効率化するため のシステムを構築し,その活用を図るとともに,個別の生物種群を対象とした調査を実施し,各種 環境要因がそれら生物種群の生息に及ぼす影響の解析を行った. ⑴ 農業景観調査情報システムの構築 農耕地およびその周辺に生育・生息する希少な生物や身近な生き物を保全するためには,それ らの状態をモニタリングすることと同時に周辺の環境変化との関連性を検討することが不可欠で ある.その際,調査によって把握される生物群集または生物種の分布の変化が,全体的傾向なの か,局所的減少なのかなどについて,的確な判断を可能とするようなモニタリングデータを集積す べきである.そこで,英国の Countryside Information System(CIS)および Countryside Survey (CS)の例を参考に,わが国の農業生態系における景観構造と生物多様性の変動に関する調査研 究を体系的・効果的に蓄積・利用するための農業景観調査情報システム RuLIS(Rural Landscape Information System)を構築した(井手ら,2005)(平成 17 年度研究成果情報,楠本ら). 本システムは,国土全体の生態系を対象に 3 次メッシュ(約 1km×1km)を単位として,気象, 土壌,地質,地形,植生,交通立地の因子にかかわる数値地図情報を多変量解析の手法を用いて客 観的に類型化し,約 37 万のすべての 3 次メッシュを 60 の景観タイプ(うち,農業生態系は 46 タ イプ)に区分し,さらに各景観タイプから選ばれたモニタリング地区において,種,生態系,景観 にわたる詳細なデータを調査し,そのデータを格納することで,国土全体にわたる生態系情報を体 系的,効果的に収集・蓄積・利用するための枠組みである(図 6).地域スケールから国土スケー ルに至る長期モニタリングのフレームとして活用できる. 農業環境技術研究所が自ら実施するモニタリングの対象地区として,利根川下流の農業生態系に 分布する 4 つの景観タイプのそれぞれからランダムに抽出した合計 32 メッシュにおいて,放棄水 田,畦畔,二次林,水田脇斜面の植生(1m×1m 方形区内の植物種ごとの高さと被度),鳥類分布 (繁殖期,越冬期)のデータを収集するとともに,空中写真の判読等により土地被覆図を作成し, 第 7 章 農業生態系における生物多様性の評価 101 ポリゴンデータとしてシステムに格納した. また,今後の全国的なデータ収集と情報提供の実現に向けて,WEB 版農業景観調査情報システ ム(RuLIS WEB)を開発した(平成 22 年度研究成果情報,山本ら).これを利用すると,国土全 体にわたる生物多様性観測情報を外部の研究者等の協力を得ることにより効率的に収集,蓄積でき るとともに,外部への情報提供を実現し,生態系構造に関する解析や評価のためのデータ利用を促 進することが可能となる. ᒻ䇮ფ䇮᳇⽎䇮ᬀ↢ ╬䈱⥄ὼ┙᧦ઙ䉇ㄘ ᬺ┙᧦ઙ䉕↪䈚䈢 ㄘᬺ↢ᘒ♽䈱㘃ဳൻ ㄘᬺ↢ᘒ♽䈱ಽ㘃 䋨䋳ᰴ䊜䉾䉲䊠න䋩 䊶ో࿖䈱ㄘᬺ↢ᘒ♽䉕60䈱䉪䊤䉴䈮ಽ㘃 䋪䈖䈱࿑䈲䊧䊔䊦䋳䈪␜ legend 64 66 67 68 ᩮᎹᵹၞ䈮䈍䈔䉎4䈧䈱䉪䊤䉴 䃂 6-e ౝ㒽ㇱ䈱᳓↰᥊ⷰ 䃂 6-f ਅᵹၞૐ᳓↰᥊ⷰ 䃂 6-g ਅᵹၞบ⼱ᵤ↰᥊ⷰ 䃂 6-h ਅᵹၞบᏒⴝ᥊ⷰ 図 6 農業景観調査情報システム RuLIS(Rural Landscape Information System)による全国の農村景観(農業 生態系)の類型化(60 タイプ) ⑵ 利根川流域の水田における植物群落の多様性と景観構造の関係 農業生態系における生物多様性を広範囲で評価するためには,個々の地点における生理・生態的 特徴や環境要因のみならず,土地利用などマクロスケールの情報との関係を把握しなければならな い.そこで,農業景観調査情報システムのモニタリング地区における調査データを利用して,植物 群落と景観構造の関係を解析した.全モニタリング地区における植物群落は 10 種類のタイプに分 類でき,そのうち 8 つのタイプは,単一もしくは少数の景観タイプとの関係が認められた.また, 土地被覆データ(住宅道路,荒地,ゴルフ場,畑,水田,放棄水田,二次草地,森林,水域)との 関係では,それぞれの群落タイプごとにいくつかの土地被覆形態と関係が認められたものの,どの ような土地被覆形態とも明確な関係が認められない植物群落タイプも少数認められた.それらはミ クロスケールにおける人間による管理状況の影響を受けていると推察された(楠本ら,2007). ⑶ チョウ類の生息ポテンシャルの評価 チョウ類の生息ポテンシャルを広範囲で評価するため,農業景観調査情報システムを用いた評価 手法を開発した.茨城県南部の谷津田周辺で,チョウ類のルートセンサスを行い,チョウの出現種 数とルート上の環境要素との関係を調べたところ,水田と森林の境界領域でチョウの出現種数多い 傾向があった.そこで,チョウの生息ポテンシャル評価の指標として水田と森林の境界線に注目 し,農業景観調査情報システムに格納されているモニタリング地区の 2 時期(1976 年と 1997 年) 102 4.農業生態系の各種環境要因が生物多様性に及ぼす影響 の土地被覆データから,水田と森林の境界線と土地利用面積の関係を解析した.その結果,水田と 森林の境界線は森林面積および水田面積との相関が高いことが判明し,チョウ類の生息ポテンシャ ルを農業景観調査情報システムに格納されている 3 次メッシュの土地利用データを利用して推定す ることが可能になった.また異なる時期の土地利用データを比較することにより,チョウの生息ポ テンシャルの変化を推定することができた(図 7)(楠本ら,2006;山本ら,2008). 1997ᐕ 1976ᐕ ಠ ᄢ ਛ ዊ 図 7 利根川流域におけるチョウ類生息ポテンシャル図(1976 年および 1997 年) ⑷ ため池の環境や配置がトンボ類に及ぼす影響 ため池は水生生物の重要な生息場所であり,わが国の水生植物やトンボの約半数の種がため池に 依存している.農業用水の供給方法の変化や都市化のために,ため池の数は急激に減少し,また環 境も悪化していると言われており,ため池に生息する生物には絶滅に瀕しているものが少なくな い.ため池の生物を保全するためには,その生息に好適な環境を知る必要がある.そこで,ため 池を代表する昆虫であるトンボ類を対象に茨城県筑波山南東麓地域にある 74 か所のため池におい て,そこに生息するトンボの種構成とため池の環境について調査を行い,ため池および周囲の環境 と生息するトンボの種構成との関係を多変量解析を用いて解析した.その結果,ため池に生息する トンボの種類は, (1)樹林に囲まれ水底に落ち葉などが多い池を好むグループ,(2)開放的で大き な池を好むグループ, (3)色々な環境の池に生息するグループなど,異なる 6 グループに分けられ ること(表 1) ,またトンボの保全のためには(1)と(2)両方の池の環境を維持することが重要 であることが判明した(図 8) (Hamasaki et al., 2011)(平成 18 年度研究成果情報,田中ら). 表 1 ため池のグループ分けとグループを指標するトンボ グループ グループ 1 グループ 2 グループ 3 グループ 4 グループ 5 グループ 6 平均種数 12.2 11.0 7.3 15.8 6.9 6.0 指標種 ノシメトンボ コヤマトンボ シオカラトンボ コシアキトンボ モノサシトンボ オオシオカラトンボ ヒガシカワトンボ クロスジギンヤンマ なし クロイトトンボ オオイトトンボ アオモンイトトンボ アジアイトトンボ コフキトンボ ショウジョウトンボ ウチワヤンマ ギンヤンマ オオヤマトンボ なし なし 第 7 章 農業生態系における生物多様性の評価 103 ᳓↢ᬀ‛ᄙ䈇 ╙䋲ゲ 4 1 2 ࿐䈏㐿 ᳰ㕙Ⓧᄢ䈐䈇 ࿐䈏᮸ᨋ ၸⓍ‛ᄙ䈇 ╙䋱ゲ ࠣ࡞ࡊ㧝 ࠣ࡞ࡊ㧠 ࠣ࡞ࡊ㧞 ࠣ࡞ࡊ㧡 ࠣ࡞ࡊ㧟 ࠣ࡞ࡊ㧢 䉮䊮䉪䊥䊷䊃⼔ጯ 図 8 トンボの生息環境からみたため池のグループ 表 1 で指標種に選ばれたグループは円で囲んである. 橙色の矢印と枠内の説明は環境の傾向を示す. また,最近の研究では,農業環境の変化にともなって,多くの昆虫種は好適な生息地を失うだけ でなく,生息地ネットワークが劣化することで,メタ個体群構造が崩れて存続できなくなってい る.そこで,トンボ類にとってのため池の配置の重要性を,池内の環境および池周辺の土地利用の 影響と比較して評価した.その結果,個々のトンボ種に注目すると,飛翔力のあまり強くない種で は,ため池の配置が最も重要であること,さらにトンボ類全体を対象にすると,ため池の配置は他 の影響と等しく重要であることがわかった(Hamasaki et al., 2009; Yamanaka et al., 2009)(平成 20 年度研究成果情報,山中ら) . ⑸ 水田地帯の鳥類の生息に及ぼす周辺環境の影響 水田とその周辺環境を含む水田地帯には様々な生物が生息している.近年,土地利用や農業活動 の変化がそこに生息する生物に対してマイナスの影響を及ぼすことが危ぐされているが,水田が主 体となる農村地域にどのような鳥類が生息しているかについては,その重要性がラムサール条約等 において国際的に指摘されているにもかかわらず,周辺環境を含め広域的に明らかにする調査は これまで行われていなかった.そこで,利根川流域の水田地帯において,夏季(6 月)と冬季(12 月)に調査したところ,夏季には 51 種,冬季には 73 種の鳥類が確認され,それらは生息地タイプ 別に 8 つのグループに分けられた.また 8 つのグループのうち,湿地性,草地性,樹林性の鳥類に 注目して,各調査地区で出現した種数と環境との関係を分析したところ,夏季には周囲 1km2 に水 田が多い地区では湿地性鳥類が,林地が多い地区では樹林性鳥類が,土地利用の多様度が高い(水 田,放棄田,樹林地等が偏りなく存在する)地区では草地性鳥類が,それぞれ多く確認された(図 9 左) .一方,冬季には,周囲 1km2 に水域が多い地区で湿地性鳥類が,林地が多い地区で樹林性鳥 類が,放棄田が多い地区で草地性鳥類が,それぞれ多く確認された(図 9 右).以上の結果から, 水田地帯では周辺土地利用によって出現する鳥類種群が大きく異なることが判明した(Amano et al., 2008) (平成 19 年度研究成果情報,天野ら) . 104 4.農業生態系の各種環境要因が生物多様性に及ぼす影響 1.0 0.0 ᫈↰㕙Ⓧ ᨋ㕙Ⓧ ᳓ၞ㕙Ⓧ ↪䈱 ᄙ᭽ᐲ ↪ߩᓇ㗀 ↪ߩᓇ㗀 0.5 ᳓↰㕙Ⓧ 1.0 A.ᄐቄ Ḩᕈ㠽㘃 ⨲ᕈ㠽㘃 ᮸ᨋᕈ㠽㘃 B.౻ቄ Ḩᕈ㠽㘃 ⨲ᕈ㠽㘃 ᮸ᨋᕈ㠽㘃 0.5 0.0 ᳓↰㕙Ⓧ ᫈↰㕙Ⓧ ᨋ㕙Ⓧ ᳓ၞ㕙Ⓧ ↪䈱 ᄙ᭽ᐲ -0.5 -0.5 ᄐቄߦߪ᳓↰ߩᄙߢߪḨᕈ㠽㘃߇㧘ᄙ᭽ߥ↪߇ ࠅߥߊሽߔࠆߢߪ⨲ᕈ㠽㘃㧘ᨋ߇ᄙߢߪ᮸ ᨋᕈ㠽㘃߇㧘ߘࠇߙࠇᄙߊ⏕ߐࠇ߹ߒߚ㧚 -1.0 ౻ቄߦߪ᳓ၞ߇ᄙߢḨᕈ㠽㘃߇㧘᫈↰߇ᄙߢ ߪ⨲ᕈ㠽㘃߇㧘ᨋ߇ᄙߢߪ᮸ᨋᕈ㠽㘃߇㧘ߘࠇߙࠇ ᄙߊ⏕ߐࠇ߹ߒߚ㧚 -1.0 図 9 利根川流域の水田環境で周辺環境が鳥類の出現種数に及ぼす影響 各土地利用の影響は−1 から+1 の値で示され,値が+1 に近いほど強い正の影響,−1 に近いほど強い 負の影響を示している. 「土地利用の多様度」は,水田,放棄田,樹林地等の土地利用が偏りなく存在す る程度を示す指標. ⑹ 水田の周辺環境が水生昆虫に対する環境保全型農業の効果に及ぼす影響 農地やその周辺に生息する生物を保全することなどを目的として環境に配慮した農業(環境保全 型農業)の普及が図られているが,このような農業が実際に生物の保全にどの程度効果があるのか についてはよくわかっていなかった.そこで,栃木県から福島県にわたる 6 地域において,農法の 異なる水田(有機栽培または殺虫剤を使わない減農薬栽培と慣行栽培)を選定し,水生昆虫(ゲ ンゴロウなどのコウチュウ類,アメンボなどのカメムシ類,トンボ類(ヤゴ))の種数と種構成, および周辺(5km×5km の範囲)の環境を調べた.その結果,有機・減農薬栽培水田では慣行栽 培水田に比べて多くの種類の水生昆虫がみられたが,その効果は地域によって差がみられた(図 10) .また水生昆虫の種構成または種数に対する農法と周辺環境の影響を解析したところ,周辺環 境の影響が大きく,特に周辺のため池の数が水生昆虫の種数に影響を及ぼしていることがわかっ た(図 11) .これらの結果は,周辺にため池などの生息場所が多く,水生昆虫の種が豊富な地域ほ ど,環境保全型農業の導入効果が高いことを示しており,環境保全型農業の効果的な普及に寄与す ると期待される(平成 22 年度研究成果情報,田中ら). 30 A D ᯏᷫㄘ⮎ᩱၭ ᘠⴕᩱၭ 25 C F Ⴃࠅߟ߱ߒ⸥ภ㧦ᯏᷫㄘ⮎ᩱၭ ⊕ᛮ߈⸥ภ 㧦ᘠⴕᩱၭ 30 ⒳ 25 20 ⒳ ᢙ ᢙ 15 ᯏᷫㄘ⮎ᩱၭ 20 15 10 10 ᘠⴕᩱၭ 5 5 0 B E 0 A B C D E F 図 10 農法が異なる水田における水生昆虫 の種数 有機・減農薬栽培水田では慣行栽培水 田より種数が多いが,種数の差の程度 は地域によって異なる. 0 10 20 30 40 ㄝߩߚᳰߩᢙ 図 11 水田周辺のため池の数と水生昆虫の種数の 関係 水田にいる水生昆虫の種数は周辺にため池 が多い地域ほど多く,この傾向は有機・減 農薬栽培水田の方が顕著である. 第 7 章 農業生態系における生物多様性の評価 105 ⑺ 野生動物の生息域拡大のしにくさを評価する手法の開発 近年,ニホンザルやシカ,クマ等の野生動物による農作物や人的被害が問題となっており,これ らの被害の防除や軽減のために,野生動物と人間の活動域を分離するための環境管理が必要とされ ている.これまで,土地利用タイプや植生に基づく生息地の評価は行われているが,その配置や面 積,形状等の空間構造を考慮した評価手法は確立していない.そこで,野生動物の生息域変化に及 ぼす空間構造の影響を定量的に評価するため,生息域の拡大のしにくさを示すコスト距離という概 念を用いた手法を開発した.房総半島におけるニホンザルの生息域変動を例として本手法を適用し た評価図を作成した(図 12) .その結果,房総半島のニホンザルの生息域拡大過程では,畑や水田 等の農耕地,ゴルフ場や草地等の開放的な空間や,住宅地が生息域の拡大を阻害することが明らか となった.本手法による評価図は,ニホンザルの生息域変動の予測に活用できるとともに他の野生 動物の生息域拡大のモデル化への適用も期待できる(岩崎ら,2005)(平成 17 年度研究成果情報, 岩崎ら) . (特許公開番号 : 2007−029046) a) 1970 ᐕઍߩ↢ᕷ⏕ὐ߆ࠄߩ⋥ ✢〒㔌࿑ b) 1970 ᐕઍߩ↢ᕷ⏕ὐ߆ࠄߩ⚥ Ⓧࠦࠬ࠻〒㔌࿑ 図 12 1970 年代のニホンザルの生息確認地点からの直線距離図とコスト距離図の比較 5.長期・広域スケールにおける生物多様性の評価 世界的に進行する生物多様性の損失を防ぐためには,特定の種だけではなく,種群もしくはグ ループ全体における変化を長期・広域スケールで把握し,評価することが必要であるが,データの 欠損や測定誤差などのため,これまでそのような試みは国内ではほとんど行われてこなかった.階 層ベイズモデルを適用することにより,そのようなデータでも,全体的な傾向を解析できる手法を 開発した. ⑴ 全国的なシギ・チドリ類の減少 生物多様性の状態を表す指標の開発が全世界で進められているが,特に生物個体数の変化を表 す個体数指数は,生物多様性の基本的な状態を表す指標の一つとして重要である.日本では,1970 年代から鳥類を中心に全国規模での生物の個体数調査が行われてきが,調査データの欠損や測定 誤差など様々な障壁のため,日本全体での個体数の変化を数値化する試みはほとんど行われてこ なかった.そこで,農業環境技術研究所はバース大学(英国),NPO 法人バードリサーチ,京都大 106 6.今後の展望 学,ケンブリッジ大学(英国)と共同で,日本国内で見られるシギ・チドリ類 42 種について,34 年に及ぶ全国調査の記録をもとに,個体数の変化を表す指数(個体数指数)を国内で初めて開発し た.解析の結果,42 種のうち 14 種が全国規模で顕著に減少していることが判明した(図 13).特 に減少していたのは,水田を生息地として利用する種や中国・韓国の黄海沿岸をよく利用する種で あった.この結果は,環境省が作成した「生物多様性総合評価」(2010)の資料としても活用され た(Amano et al., 2010b ) . ⑵ 英国における 250 年の記録から植物群集の開花時期指数を開発 気候変動は生物多様性に対する深刻な脅威の一つとして認識されており,その影響を指標とし て定量化し,観測していくことが求められている.英国には,全土で 1753 年から断片的に蓄積さ れてきた約 40 万件に及ぶ植物 405 種の開花日記録が整備されている.このデータに階層ベイズモ デルを適用することで,植物群集全体での「平均的な」開花時期変化を表す指数と種ごとの指数 を開発した.その結果,推定された 1753 〜 2009 年の開花時期指数によって,英国の植物群集は直 近の 25 年間に最も早い開花時期を示していることが明らかになった.また,2 〜 4 月の全国平均 気温が 1℃上昇するごとに開花時期は約 5 日早くなることが示された.この結果は,気候変動が生 物多様性に与える影響を表す指標の一つとして今後活用されることが期待される(Amano et al., 2010a) . ᢙᜰᢙ 図 13 1975 年から 2008 年までの日本全国におけるシギ・チドリ類の個体数指数の例(左:チュウシャクシ ギ,右:ムナグロ) 黒の実線は指数の推定値(中央値) ,灰色の幅はそのばらつき(95%信用区間)を示す.指数は 2008 年 の値を 100 とした相対値で示されており,指数の減少は個体数の減少を示している. 6.今後の展望 2010 年 10 月に名古屋で開催された生物多様性条約第 10 回締約国会議(COP10)では,「現在の 生物多様性の損失速度を 2010 年までに顕著に減少させる」という 2010 年目標が失敗したことを踏 まえ,それに代わるポスト 2010 年目標(愛知ターゲット)が採択された.2020 年までに達成され るべき 20 の個別別目標の中には, 「自然生息地の損失速度を少なくとも半減,可能な場合はゼロに 近づける」 , 「少なくとも陸域および内陸水域の 17%,沿岸域・海域の 10%を現状のまま保全する」 第 7 章 農業生態系における生物多様性の評価 107 などとともに, 「農業,養殖業,林業が行われる地域が生物多様性の保全を確保するように持続的 に管理される」が含まれている.また,農業生物多様性に関する決議には「食料安全保障や栄養 補給に対する農業生物多様性の重要性を認識する」および「農業生態系,特に水田およびオアシス の生態系について,生物多様性の保全および持続的な利用のために重要であることを認識し,(中 略) ,農業生態系によりもたらされる生態系サービスの評価に関する更なる研究に取り組むことを 要請する」など,農業生物多様性の重要性をうたった文言が少なからず含まれていることから,農 業生態系における生物多様性の保全に向けた研究は今後ますます重要になると思われる. これまでも,生物多様性に関する研究は,世界各国で精力的に実施されてきたが,そもそも生物 多様性はそれを構成する要素が膨大であり,しかも相互の関係が複雑であるため,われわれは依然 として生物多様性について十分には把握できていないというのが実情である.農業環境技術研究所 でも,農業生態系における景観構造や人間活動を含む各種要因が生態系の構造や生物多様性にどの ような影響を及ぼしているかについて調査・研究しているが,研究対象としてきた生物群は,技術 的および資源的な制約から,多くの場合,植物,昆虫類,鳥類等に含まれる特定の種あるいは群に 限定せざるを得なかった.今後は,これまで同様に特定の生物種・群に関する研究を進めることも もちろん重要であるが,農業生態系の全体像の把握に向け,より大きな単位である群集レベルを対 象とした研究やそのための手法開発に取り組む必要があろう.また,多様な生物の状態を効率的に 調査できる手法・システムの開発,生物多様性を簡易に評価するための指標の開発も不可欠であ る.さらに最終的な目標として,生物多様性の保全を実現するための管理技術についても,様々な アイデアを試しつつ開発に向けて努力する必要がある.また,地球温暖化は,生物に対する直接的 な影響だけでなく,農法の変化を通して間接的に生物に影響を与える可能性もあり,そのような観 点から農業と生物多様性の関係を考えることも重要である. 「生物多様性」という言葉は単に生物の多様性という生物学的な意味だけでなく,経済的,情緒 的,文化的な意味など様々なものが含まれているため,「生物多様性」のどの側面を問題とするか によって,研究の目標やアプローチの方法が異なる.農業生態系における生物多様性の研究におい ては,農業の持続可能性と身近な生物多様性の維持の両立が目標であるが,どちらに軸足を置くか によっても,研究の方向は異なる.今後の研究を進めるにあたっては,それらの問題を整理し,目 標を明確に見定めることが必要であろう. 引用文献 1)Amano, T., Kusumoto, Y., Tokuoka, Y., Yamada, S., Kim., E-Y. and Yamamoto, S.(2008) Spatial and temporal variations in the use of rice-paddy dominated landscapes by birds in Japan. Biological Conservation, 141, 1704-1716 2)Amano, T., Smithers, R.J., Sparks, T.H. and Sutherland, W.J.(2010a)A 250-year index of first flowering dates and its response to temperature changes. Proceedings of the Royal Society B: Biological Sciences, 277, 2451-2457 3)Amano, T., Székely, T., Koyama, K., Amano, H. and Sutherland, W. J.(2010b)A framework for monitoring the status of populations: an example from wader populations in the East Asian-Australasian flyway. Biological Conservation, 143, 2238-2247 4)Hamasaki, K., Yamanaka, T., Tanaka, K., Nakatani, Y., Iwasaki, N. and Sprague, D. S.(2009) Relative importance of within-habitant environment, land use and spatial autocorrelations for determinating odonate assemblages in rural reservoir ponds in Japan, Ecological Research, 108 引用文献 24(3) , 597-605 5)Hamasaki, K., Yamanaka, T., Tanaka, K., Nakatani, Y., Iwasaki, N. and Sprague, D. S.(2011) Environmental characteristics accounting for Odonate assemblages in rural reservoir ponds in Japan. JARQ, 45 (2) , 187-196(in press) 6)井手 任,大黒俊哉,楠本良延(2005)生物多様性保全のための景観・植生調査情報システ ム.インベントリー,4,20-23 7)Ikeda, H. and Itoh, K.(2001)Germination and water dispersal of seeds from a threatened plant species Penthorum chinense. Ecological Research, 16, 99-106 8)岩崎亘典,デイビッド スプレイグ(2005)房総半島のニホンザル生息拡大地域における土地 利用変遷に関する研究,農村計画論文集,7,1-6 9)小柳知代,楠本良延,山本勝利,大黒俊哉,井手 任,武内和彦(2007)関東地方平野部にお けるススキを主体とした二次草地の過去と現在の種組成の比較.ランドスケープ研究,70(5), 439-444 10)楠本良延,大黒俊哉,井手 任(2005)休耕・耕作放棄水田の植物群落タイプと管理履歴の関 係 ― 茨木県南部桜川・小貝川流域を事例にして ― .農村計画論文集,7,7-12 11)農林水産省(2007)農林水産省生物多様性戦略 12)大黒俊哉,白戸康人,伊藤一幸(2003)絶滅危惧植物タコノアシ個体群の維持にかかわる放棄 水田の環境特性 . ランドスケープ研究,66 (5),599-602 13)清水矩宏(1998)水田生態系における植物の生物多様性とは何か.農業環境技術研究所叢書, 第 10 号,82-126 14)山本勝利,井手 任,大黒俊哉,楠本良延,天野達也,徳岡良則,奥島修二,小出水規行,竹 村武士(2008)景観構造に関する調査・情報システムの開発とそれを活用した生物相の変動評 価.研究成果 462(農林水産省農林水産技術会議事務局),173-189 15)Yamanaka, T., Tanaka, K., Hamasaki, K., Nakatani, Y., Iwasaki, N., Sprague, D. S., Bjrnstad, O. N.(2009)Evaluating the relative importance of patch quality and connectivity in a damselfly metapopulation from a one-season survey. Oikos, 118, 67-76 109 第 8 章 外来生物のリスク評価 1. はじめに 外来生物とは,本来の分布域ではない地域に,何らかの目的をもって意図的に導入されたか,も しくは付着や混入によって非意図的に導入された生物である(角野,2010).わが国では明治以降 に外来生物の種数が増加し,経済のグローバル化による貿易量の拡大にともなって近年さらに大き く増加している.外来生物が生態系に侵入・定着すると,人の健康や農業生産に直接的な被害を与 えることが従来から問題となっていたが,最近では,地域固有の生態系や生物多様性に対する影響 が大きな問題として認識されるようになった.生物多様性国家戦略 2010(環境省,2010)におい ても,外来生物が生物多様性を脅かす 3 つの危機のひとつ(第 3 の危機:人間により持ち込まれた もの)として挙げられている.農作物に被害を与える可能性のある昆虫や動物に対しては,「植物 防疫法」に基づいた輸入植物の検疫が行われ,海外の農作物害虫の侵入を阻止するための措置がと られている.一方,その他の生物については,たとえ生態系に悪影響を及ぼす可能性のある生物で あっても,最近までそれらを取り締まる法律はなかった.2004 年に「特定外来生物による生態系 等に係る被害の防止に関する法律」 (外来生物法)が制定されたことにより,生態系,人の生命・ 身体または農林水産業に被害をもたらす特定外来生物の輸入,飼養,栽培,保管,運搬,譲渡等が 規制されるとともに,野外への放逐や植栽が禁止され,また特定外来生物による被害が生じている 場合は,必要に応じて国や地方公共団体等がそれらを防除することも定められた.しかし,特定外 来生物を選定する際の基準が必ずしも明確でない(藤井,2008),あるいは,防除の必要性の判断 基準が不明瞭である(西田,2010)といった問題点も指摘されている. 今後,特定外来生物の管理を実効あるものとするためには,特定外来生物の選定や防除の要否決 定に関する基準を科学的根拠に基づいて策定する必要がある.そのためには,既に国内に侵入して いる外来生物については,生息状況の実態を把握し,生態系への影響を評価することが必要であ る.また今後侵入が予想される,あるいは導入が検討されているものについては,侵入警戒措置も しくは導入の可否を決定するためのリスク評価法の開発が不可欠である.農業生態系においても, これまで多くの外来植物や昆虫,微生物などが侵入・定着し,あるものは農作物に大きな被害を与 え,またあるものは周囲の生態系に様々な影響を与えていると考えられる.農業環境技術研究所で は,農業生態系のかく乱と被害を防止するため,外来生物の生育・繁殖特性等の解明および外来生 物の生態系影響リスク評価法の開発など,農業生態系における外来生物問題に関する研究を実施し てきた.本章では,この 10 年間にわたる関連の研究成果を,(1)外来植物の定着・拡散,(2)外 来植物の雑草リスクの評価, (3)非意図的に導入された昆虫等に関する研究,(4)意図的に導入さ れた昆虫に関する研究, (5)外来性病害微生物に関する研究,(6)外来生物データベースの構築の 各項目に分けて紹介する. 2.外来植物の定着と拡散 まんえん 侵略的な外来植物の蔓延防止策を開発するには,それらの生態特性や拡散・定着過程を解明する ことが重要である.これまで農業環境技術研究所では,多くの外来植物の生態特性やアレロパシー 活性について研究を行ってきたが,ここでは,定着・拡散において在来種との交雑が重要な役割を き ぐ 果たしている事例,蔓延が危惧されている種に関する実態把握の例,および外来種の蔓延に及ぼす 110 2.外来植物の定着と拡散 土壌条件の影響に関する研究について紹介する. ⑴ タンポポ属植物の分布と遺伝特性 近年,セイヨウタンポポなどの帰化タンポポと日本産のタンポポが交雑可能であることが報告さ れたが(森田,1997) ,雑種個体は形態的に帰化タンポポに酷似していることから,形態形質以外 の特徴によって両者を識別・分離し,それぞれの分布域を再確認する必要がある(芝池,2005). そこで,分子生物学的手法を活用した簡便・迅速な雑種識別法を確立し,帰化タンポポと雑種個 体の全国レベルの分布域を把握することを試みた.DNA 含量および DNA の塩基配列などを指標 にしたセイヨウタンポポと日本産タンポポの雑種識別法を確立し,両者の雑種として 3 タイプを識 別した.また,環境省が行う「身近な生きもの調査」により採集された形態的にセイヨウタンポポ と見なされる 870 個体のサンプルについて,DNA 含量の測定と葉緑体 DNA マーカーを用いた解 析を行った結果,全体の約 85%にあたる 740 個体が雑種であることが明らかとなった(図 1)(山 野ら,2002) .その内訳は,4 倍体雑種が 54%の 470 個体,3 倍体雑種が 22%の 191 個体,雄核単 為生殖雑種が 9%の 79 個体であった(平成 13 年度研究成果情報,芝池ら).また,関東地方 1 都 6 県に注目すると,タンポポ属植物のうち,形態的にセイヨウタンポポと見なされる個体が 60%を 占め,その 71%は 4 倍体雑種であった.これら雑種性タンポポの分布拡大過程を解明するために は,特定の雑種個体の分布状況を把握することが有効と考え,雑種個体を遺伝的に識別し,それら の広がりを比較・解析したところ,環境省の「身近な生きもの調査(2001)」で収集された関東地 方 1 都 6 県のタンポポのうち,42%を占める 4 倍体雑種性タンポポの 94%は遺伝的に同一なクロー ンであり,しかも,その単一クローンの分布域は,関東全域に及んでいることが明らかになった (芝池ら,2005) (平成 16 年度研究成果情報,芝池ら). 図 1 セイヨウタンポポと雑種個体の全国分布 ⑵ 特定外来生物ナガエツルノゲイトウの水田への侵入拡散 特定外来生物ナガエツルノゲイトウ(図 2)は,南米原産の多年生草本植物で,水域から陸域ま で生育可能な範囲が広く,世界各地に侵入・定着している.日本国内での分布は現時点ではまだ一 部の湖沼や河川にほぼ限られているが,水田周辺では蔓延リスクが高いことが知られている.そこ 第 8 章 外来生物のリスク評価 111 で,本種の水田域における侵入,拡散の実態を明らかにするため,千葉県印旛沼周辺の水田を対象 に 3 年間にわたる現地調査を行った.その結果,本種は水田域で前年の分布範囲や用水路網の影響 けいはん を受けながら分布拡散していること,田面よりも畦畔で生育量が多いことが明らかになり,本種の 水田域への侵入・蔓延を防ぐには,用水路などを通じた侵入の防止と,畦畔を含む水田域全体での 防除が必要であると考えられた.ナガエツルノゲイトウによる農業生産や周囲の生態系への影響を 未然に防止するために,この知見が活用されることが期待される(平成 22 年度研究成果情報,山 本ら) . 図 2 田面のナガエツルノゲイトウ ⑶ 外来植物の生息と土壌条件 農地周辺に成立している草本群落における外来植物の蔓延と土壌の化学特性との関係を明らかに するため,北関東に広く分布している台地上黒ぼく土壌を中心に,畦畔,耕作放棄地,刈り取り草 地を対象として 122 地点で植生調査を実施したところ,農地周辺の草本植物群落は,セイタカアワ ダチソウなどの外来植物の比率が比較的高いタイプ I と,外来植物比率が低く,アズマネザサ,ア キカラマツ,ワレモコウ,ツリガネニンジンなどで特徴付けられるタイプ II の 2 つに大きく分け られた.タイプ I は土壌 pH が高いか,あるいは有効態リン酸が高い土壌で多く出現し,タイプ II は土壌 pH が低く,かつ有効態リン酸が低い土壌で出現した(図 3).また室内栽培実験でも,セ イタカアワダチソウやオオマツヨイグサなどの外来植物は酸性土壌で著しく生育が抑制されるのに 対し,ススキ,クズ,イヌビエなどの在来植物は酸性土壌でも生育障害を受けにくいことが判明し た.以上のことから,自然の黒ぼく土壌の化学的特性が維持されている場所ではタイプ II が出現 し,施肥や基盤整備等で土壌の化学的特性が大きく改変された場所ではタイプ I が出現しやすいと 考えられた.調査地周辺の農地ではリン酸の蓄積や土壌 pH の上昇が進行しており,これが農地周 辺や耕作放棄地での外来植物の蔓延を助長していると考えられた(平舘ら,2007;2010)(平成 20 年度研究成果情報,平舘ら) .今後,この結果を応用した外来植物の管理技術の開発が期待される. 112 3.外来植物の雑草リスクの評価 図 3 北関東地域における外来植物の侵入と土壌の化学特性の関係 土壌 pH および有効態リン酸が低い土壌では,外来植物がほとんど出現していない.また,このような土 壌の特性は,この地域において施肥等がなされていない表層土壌が示す典型的な化学特性の範囲内(図 中の点線の枠内)にあると言える. 3.外来植物の雑草リスクの評価 外来生物を意図的に導入しようとする場合,事前のリスク評価を行い,生態系影響が大きい可能 性があるものについては導入を中止することが,外来生物を管理する上で最も効率的かつ効果的な 方法である.また,非意図的に導入される可能性がある生物やすでに国内に侵入している生物につ いても,生態系影響に関するリスク評価に基づいて侵入の警戒レベルを設定,あるいは防除の優先 順位を決定することは,侵入防止対策および防除対策を進める上で有効である. ⑴ 雑草リスク評価法 外来雑草による被害を軽減するには,導入する前に雑草化の恐れが高い植物を予測し,導入を避 けることが最も効率的な防除法である.そこで,新たに日本へ導入される植物を対象に,日本で 雑草化する植物とそうでない植物を判別する手法を,国際的に定評のあるオーストラリア式雑草 リスク評価法を基に構築した.オーストラリア式雑草リスク評価法では 49 の質問に対する回答か ら雑草化しやすさを示す WRA スコアを算定する.日本にある 259 種の植物をあらかじめ専門家に 雑草とそうでないものに区分してもらい,WRA スコアでこの判断を再現できるか調べたところ, WRA スコアが 10 より大きい植物を雑草と判別する場合,雑草であるものを正しく雑草と判別で きる割合,および雑草でないものを正しく雑草でないと判別できる割合は,それぞれ 9 割弱と 8 割 弱となり,雑草とそうでないものを同程度に良く判別できることが分かった.この評価法を用い, WRA スコア 10 を超える植物の導入を避けることで,外来雑草による被害を未然に防ぐことが期 待できる(Nishida et al., 2009) (平成 20 年度研究成果情報,西田ら). 一方,国際連合食糧農業機構(FAO)は 2005 年に 13 因子で評価する簡易型の雑草リスク評価 法を発表した.そこで,この手法で使われている 13 因子に他感作用を加えた 14 因子を環境省が提 案している「要注意外来植物」に当てはめて,各因子の寄与率を相関係数で解析したところ,他感 作用の寄与率は高いが,寄生性等の 3 因子の寄与率が低いことが判明したため,それら 3 因子を省 き,また類似した 2 因子をひとつに合併することで,合計 10 因子で評価する簡易型雑草リスク評 第 8 章 外来生物のリスク評価 113 価法を開発した.この評価法を用いて「特定外来生物」および「要注意外来生物」を再評価したと ころ,現場での蔓延状況とかなり一致する結果がでたことから,今後のリスク予想にも活用できる 可能性がある(藤井,2008) (平成 19 年度研究成果情報,藤井ら). ⑵ 輸入穀物に種子が混入しやすい外来植物 日本は年間約 2700 万トンという膨大な量の穀物を輸入しているが,これらの輸入穀物には,割 合は少ないながらも様々な植物の種子が混入している.現在,国内の河川敷,飼料畑あるいは穀物 畑で問題となっている雑草の一部は,それら輸入穀物の混入種子が起源であると考えられており, 今後侵入してくる雑草に備えるためにも,どのような特徴をもった植物が輸入穀物に混入しやすい かを明らかにする必要がある.そこで,2006 年にカナダより輸入した小麦に混入していた植物種 子を調査したところ,13 科 42 種類の種子が混入しており,中でも特に多かったのが,オオムギ, カラスムギ等のイネ科とアブラナ科であった(図 4 右).この結果とカナダのコムギ畑における既 存の発生調査(図 4 左)を比較し,混入率との相関を分析したところ,「生産地の畑での発生量が 多い」 , 「背が高い」 , 「種子に冠毛が無い」 , 「作物種である」ことが混入率を高める要因であると推 定された(Shimono et al., 2008) (平成 19 年度研究成果情報,小沼ら). また,輸入穀物が栽培されている海外の穀倉地帯では,除草剤抵抗性雑草が多数発生しているこ とから,輸入穀物の混入雑草種子の中にも除草剤抵抗性を持つ個体が含まれている可能性がある. そこで,除草剤抵抗性のボウムギが問題になっている西オーストラリアから輸入された小麦を対象 にボウムギの混入種子を探しだし,その中に,現地の小麦畑で使用頻度の高い 4 種類の除草剤(ジ クロホップメチル,セトキシジム,クロロスルフロン,グリホサート)に対する抵抗性個体が含ま れているかどうかを調査したところ,グリホサート以外の 3 種類の除草剤について,抵抗性ボウム ギ種子の混入が確認された.グリホサート以外の除草剤はわが国では使用頻度が低いため,混入し ていたボウムギがすぐに難防除雑草になる可能性は低いが,他国からの輸入小麦も含めて今後とも 注意が必要である(平成 22 年度研究成果情報,小沼ら). ホウキギ セイヨウトゲアザミ アマ ソバカズラ グンバイナズナ オオムギ シロガラシ ホウキギ コハコベ カラスムギ カラスムギ アブラナ属 ソバカズラ エノコログサ 原産国のコムギ畑における発生量 ドクムギ属 エノコログサ ウマノチャヒキ イネ科 アブラナ科 タデ科 アカザ科 ナデシコ科 キク科 アマ科 その他 輸入コムギに混入していた種子 図 4 収穫時期のコムギ畑における雑草の発生量と輸入コムギ中に混入していた 種子の組成との比較 ⑶ 水田周辺における注意すべき外来植物 農業生産や生態系に深刻な影響を及ぼしている外来植物の被害を防止するには,適正なリスク評 価に基づいた対策を講ずる必要があるが,現在わが国に定着している外来植物の多くについては, その蔓延状況や被害実態が必ずしも明らかにされていない.そこで,利根川流域の水田周辺,農業 水路周辺の調査資料から平均および最大被度,立地別の出現率と特化度(立地との結びつきを表す 114 4.非意図的に導入された昆虫等に関する研究 指数)を用いて外来植物の蔓延状況を解析するとともに,多くの外来植物のうちどのような種に対 して優先的な対策が必要であるかを検討した.その結果,農業生産や生態系に影響が大きい外来種 の蔓延状況には,A)高被度で優占する,B)広範囲に生育する,C)特定の立地と結びつきが強 い,の 3 パターンがあることが判明した.そのうち特に,セイタカアワダチソウ(A,B),ナガエ ツルノゲイトウ(A,B) ,コカナダモ(A,C) ,ホテイアオイ(A),コセンダングサ(B,C), ヒメムカシヨモギ(B,C) ,アメリカアゼナ(B),アメリカセンダングサ(B),アメリカタカサ ブロウ(B) ,コバナキジムシロ(B) ,アメリカミズキンバイ(C),オオイヌノフグリ(C),シロ ツメクサ(C) ,ハルジオン(C) ,ヒメジョオン(C)の 15 種の外来植物が農業生産や生態系への 影響が大きく,優先的な対策を検討する必要があると考えられた(平成 19 年度研究成果情報,楠 本ら) . 4.非意図的に導入された昆虫等に関する研究 非意図的に国内に導入される昆虫や動物の数は近年の貿易量の拡大に伴って増加している.農業 環境技術研究所では,輸入物資への付着・混入による侵入を未然に防ぐための植物検疫におけるサ ンプリング方法の改善や侵入した生物を根絶するため初期防除に関する理論的研究を実施してい る.また国内に既に侵入し,分布を拡大しつつある昆虫等の生息実態の解明にも取り組んでいる. ⑴ 植物検疫のための理論的研究 病害虫が農作物や手荷物とともにわが国に侵入し,蔓延することを防止するため,輸入植物の検 疫が空港や港では実施されている.農業環境技術研究所では,植物防疫所との協力により,輸入農 産物等の荷口検査における具体的なサンプリング方法等,様々な検疫上の問題を解決するための理 論的な研究を行ってきた. 最近では,検疫強度を危険度分析に基づいて定めようとの国際的な動きがあるが,その理論的根 拠は必ずしも明確でないため,害虫の侵入確率をそれぞれの種の生態的特性に基づいて的確に推定 し,所定の安全レベルを達成するための検疫強度を理論的根拠に基づいて新たに算出することが必 要とされた.そこで,繁殖可能な害虫の国内への侵入確率の推定式,侵入確率を所定のレベル以下 に低下させるのに必要なサンプリング検査と検疫処理の組み合わせを決定するための式を導いた (図 5) (Yamamura et al., 1999) (平成 13 年度研究成果情報,山村ら). 図 5 メキシコミバエの荷口あたりの侵入確率 P1 をサンプル率と検疫処理後の生存率の関数として示す(荷口 内の果実数が 105 の場合) .この等高線から,所定の侵入確率を達成するために必要なサンプル率と検疫 処理の組み合わせを直ちに読み取ることができる. 第 8 章 外来生物のリスク評価 115 ⑵ 侵略的外来昆虫の初期防除に関するモデル解析 侵略的な外来昆虫がいったん侵入地で増殖し定着してしまうと,その対策に莫大な労力と費用が かかる上,しばしば根絶が不可能となる.そこで,侵入初期の低密度条件下で,どのような防除法 が最も効果的であるか,雄の交尾相手探索行動を定式化して組み込んだシミュレーションモデルを 開発し,様々な防除法の効果を検討した.雄除去法や不妊虫放飼法など,交尾を阻害する防除法で は,これまで使われてきた交尾探索行動を仮定しない害虫管理モデルの予測よりも,低密度条件で ははるかに簡単に害虫個体群を根絶できることがわかった(図 6).また 2 つの昆虫タイプ,すな わち甲虫型(長生きして何度も交尾できる)とガ型(短命でメスは生涯 1 回のみ交尾し産卵して死 ぬ)を仮定して各防除法の特徴を調べたところ,雄除去法は甲虫型よりガ型に効果的であるのに対 し(図 7),不妊虫放飼法では,たくさん放飼が可能であるときにガ型より甲虫型で効果が高いこと (図 8)等が判明した.実際の侵略的外来昆虫の防除を考えるモデルでは,交尾回数や寿命など,そ の害虫に合わせた生活史構造とパラメタの推定が必要となるが,交尾探索行動を組み込んだ今回の モデルは,そのような実際の防除プログラムにおいても,より現実的な防除の帰結を予測する強力 ࿑㧢 3 የត⚝ⴕേ䉕 ቯ䈚䈭䈇䊝䊂䊦䈱 ᩮ⛘น⢻ᢙ ೋᦼᢙߣ⟲Ⴧᱺ₸ 㒐㒰㐿ᆎᤨὐߩೋᦼᢙ߇ࠆޔ ৻ቯߩ୯ࠃࠅዊߐ႐วޔየ⋧ᚻ 2 ਇߩᓇ㗀ߦࠃߞߡ߿߇ߡ⛘Ṍߒ߹ ߔޕ࿁ߩࡕ࠺࡞ߢߪޔᚑ㐳₸߇㧝 1 ߣߥࠆᢙ㧔㧩ᩮ⛘น⢻ᢙޔ ࿑ਛߩ⿒ὐ㧕߇ޔየត⚝ⴕേࠍ ࿁䈱䊝䊂䊦䈱 ᩮ⛘น⢻ᢙ 0 ⟲ᚑ㐳₸ 4 なツールとなると期待される(Yamanaka et al., 2009a; 2009b)(平成 21 年度研究成果情報,山中). 0 10,000 20,000 30, 000 ቯߒߥࡕ࠺࡞ߩ੍᷹㧔࿑ਛ㤥ὐ㧕 ࠃࠅ߽ᄢ߈ߊޔᩮ⛘߇ኈᤃߢࠆߎ ߣࠍ␜ߒߡ߹ߔޕ 4X104 ࿑㧣 㓶㒰ᴺߩലᨐ ࡈࠚࡠࡕࡦߥߤߩ⺃ᒁࠍߞ ߡ㓶ࠍ⺃Ვߔࠆ㓶㒰ᴺߢߪޔ 䉧ဳ⯻ ↲⯻ဳ⯻ࠃࠅ߽ࠟဳ⯻ߩᣇ ߇ߡߓ✚ޔ㒐㒰ലᨐ߇㜞ߊޔᩮ 2X104 ᩮ⛘น⢻ᢙ ᩮ⛘ኈᤃ ଚೋᦼᢙ ⛘߇ኈᤃߢࠆߎߣ߇ࠊ߆ࠅ߹ ߒߚ ߫߃ޕ4000ߩ࠻࠶ ࡊ ↲⯻ဳ⯻ ࠍ⸳⟎ߒߚߣ߈㧔࿑ਛ⚦✢㧕ޔ ࠟဳߢߪ2ਁએߢ߽ᩮ⛘น⢻ ਁ2ජ⒟ᐲߢߔޔߪߢဳ⯻↲ޕ 0 ᩮ⛘࿎㔍 ߢࠆߩߦኻߒޔߪߢဳ⯻↲ޔ1 0 㒐㒰ዊ 4,000 㓶㒰↪䊃䊤䉾䊒ᢙ 8,000 㒐㒰ᄢ ₪ߐࠇߥ߆ߞߚ㓶߇㐳↢߈ߒ ߡޔᐲ߽የߔࠆߚലᨐ߇ ߇ࠄߥߣផኤߐࠇ߹ߔޕ 4X104 4.非意図的に導入された昆虫等に関する研究 ᩮ⛘ኈᤃ 116 ↲⯻ဳ⯻ ࿑㧤 ਇᅧ⯻㘺ᴺߩലᨐ ✢߿ ൻቇ ⮎ຠߥ ߤߢਇ ᅧ ߦ ߔਇ ᅧ⯻ 㘺ᴺ ߢߪ ޔ 2X104 ᩮ⛘น⢻ᢙ ൻ ߒߚ㓶 ᚑ⯻ ࠍޔᄢ ㊂ߦ㊁ ᄖ 㘺 ᢙ߇Ⴧ ടߔ ࠆߦߟ ࠇ⯻ ↲ޔ 䉧ဳ⯻ ဳ ߩᣇ߇ ߣဳ ࠟޔᲧ ߴߡ㒐 㒰 ല ᨐ߇㜞 ߎ ߣ߇ࠊ ߆ࠅ߹ ߒ ߚ ߪߢ ဳ⯻↲ ޔߪ ࠇߎޕਇ ᅧ 0 ᩮ⛘࿎㔍 ⯻ ߇㐳↢ ߈ߒ ߡޔ㊁ ᄖ㓽ߩ ᱜ Ᏹ 㓶ߣߩ የ ࠍࡉࡠ ࠶ࠢߔ ࠆ 0 㒐㒰ዊ 4,000 ਇᅧ⯻㘺ᢙ 8,000 㒐㒰ᄢ ߚ ߩ ߘޔല ᨐ߇㐳 ߊᜬ⛯ ߔ ࠆ߆ࠄߢߔޕ ⑶ ブタクサハムシの移動分散能力の解明 外来昆虫の分布拡大を予測するためには,飛翔による移動能力を定量化し,飛翔活動に影響する 要因を明らかにすることが重要である.ブタクサハムシは北米原産の外来昆虫で,日本では 1996 年に千葉県で初めて発見されたが,それ以来急速に分布を拡大し,現在(2011 年)では沖縄県を 除く全都道府県で見つかっている.初期には,年間 100km 以上の速度で分布を拡大し,外来昆虫 の中でも分布拡大速度が非常に大きい.一方,寄主植物であるブタクサを食い尽くすことがしば しば観察され,定着性が高いとも考えられた.そこで,本虫の飛翔活動に影響を及ぼす要因を明 らかにし,分布拡大能力を推定することとした.飛翔測定装置(フライトミル)を用いて,成虫 の平均飛翔時間と 1 分より長く飛翔した個体割合を指標として解析した結果,ブタクサハムシ成虫 の飛翔活動は羽化後 4 日後から増加し,7 日目以降は長期間高い値を維持すること,短日または休 眠が飛翔活動の低下を引き起こすこと,飛翔時間には個体間で大きな変異があり,飛翔活性の遺 伝変異が大きく,移動性の高い個体が分布拡大に寄与することが明らかとなった.また 1 日当たり 最大飛翔距離は 25km 以上と推定され,年 4 世代を経過することを考慮すると,少なくとも 1 年間 に 100km 以上移動可能であり,移動能力が高いと考えられた(Tanaka et al., 2009; Tanaka, 2009) (平成 17 年度研究成果情報,田中ら) . ⑷ カワヒバリガイの分布と移動分散 特定外来生物に指定されているカワヒバリガイは 1990 年代に西日本に侵入した中国原産の付着 性二枚貝で,侵入地域で在来生物群集の生息地を圧迫したり,利水施設の運用を妨げたりするなど の被害をもたらすことが知られている.2005 年には関東地域の一部で生息が確認されたが,霞ヶ 浦では生息分布の詳細や侵入時期については不明であった.そこで,霞ヶ浦全域でカワヒバリガイ の分布調査を 2006 年に行った結果,既に湖岸の約 2 分の 1 の範囲まで分布が広がっていることが 明らかになった.採集個体の大きさから,霞ヶ浦への侵入時期は 2004 年かそれ以前であると考え られた(平成 18 年度成果情報,伊藤) .その後,利根川河口から 180km までの範囲とその周辺の 河川・湖沼・水路を対象に分布調査したところ,河口から 120km 上流までの範囲と,小貝川・江 戸川・牛久沼などの周辺の河川・湖沼など広い範囲に生息していることが明らかとなった(伊藤, 2008) .利根川水系には,上流の群馬県大塩貯水池およびその周辺にもカワヒバリガイが分布する 第 8 章 外来生物のリスク評価 117 ことが知られているが,それを含めた関東の集団と西日本に生息する集団の遺伝的組成は,利根川 下流域の集団と西日本の集団の 2 つに大別されること,また群馬県の集団は西日本タイプであるこ とが判明した(Tominaga et al., 2009) .また利根川下流域の集団内には遺伝的違いはなく,地点間 で活発な遺伝的交流(移動分散)があることが示唆された.カワヒバリガイの分布は浮遊幼生期に おける水の流れに沿った受動的な移動の結果であると考えられるが,カワヒバリガイの密度分布を 考慮すると,移動の経路は自然河川だけでなく,導水路などの人工的な流れの関与も示唆された (図 9) (伊藤,2010) . 図 9 霞ヶ浦と小貝川を結ぶ霞ヶ浦用水の位置関係状況.矢印は流れる方向を示す. (伊藤,2010)より改変 5.意図的に導入された昆虫等に関する研究 導入天敵昆虫等(ダニ類を含む)は,非意図的に侵入してきた外来昆虫等の農業被害を防止する ため,意図的に導入されるものであるが,これらが野外に定着した場合,目的とする害虫種以外へ の捕食・寄生や土着近縁種との交雑・競合等を通じて,地域固有の生態系に影響を与える可能性が ある.そこで,導入しようとする生物の生態系影響を事前に評価できる手法を開発した.また,導 入後に土着近縁種との交雑が起こった事例について,遺伝子情報を利用した交雑個体の識別法を開 発した. ⑴ 導入天敵昆虫等の生態系影響評価 平成 11 年に環境省は「天敵農薬に係る環境影響評価ガイドライン」を策定し,導入天敵昆虫等 の生態系影響評価方法を示したが,この方法は定性的であり,海外の天敵昆虫等を導入する際に必 要な事前評価のための判断基準や導入判断基準について必ずしも明確ではなかった.そこで,ヨー 118 5.意図的に導入された昆虫等に関する研究 ロッパで提唱されている評価指標を使用する導入天敵昆虫等の定量的評価方法を参考に,わが国の 環境にも適用可能であり,客観的で透明性の高い新たな導入判断のための基準を開発した.ここ では,導入天敵昆虫等の生態系影響を,その可能性(L)と程度(M)に分割し,定着,分散性, 寄主範囲,希少種や地域固有種への直接的影響,近縁天敵との競争,近似種との交雑の 6 つの評価 項目について,それぞれ 5 段階の基準に基づいて評価し,それらの積和(L×M;損失期待値)を 導入判断基準の指標値として用いた.ただし,事前評価にかかる時間とコストを節約するため,文 献や標本調査あるいは閉鎖系での実験によって判定できる,定着,分散性,寄主範囲の 3 項目によ る 1 次基準(表 1)と,追加実験が必要な,希少種や地域固有種への直接的影響,近縁天敵との競 争,近似種との交雑の 3 項目からなる 2 次基準(表 2)の 2 段階で判定することとし,1 次基準で 一定の条件を満たさない種(生態影響が大きいと懸念される種)についてのみ 2 次基準による判定 を行い,1 次基準の指標値との合計によって総合判定することとした.過去のわが国および海外に おける導入天敵に関する生態系影響の事例を参考に計算したところ,1 次指標が 40 を超えるもの については 2 次基準による判定に進み,1 次および 2 次基準の合計による総合指標が 80 を超える 種については,導入を見送るか,指標値が 80 以下になるような方策をとる必要があると判断でき た(望月,2008) (平成 17 年度研究成果情報,望月ら). 表 1 導入天敵昆虫等の生態系影響評価 1 次基準 L:生態系影響の可能性 L 定着(可能性) 分散性(導入ステージの余命) 寄主範囲(天敵の摂取量≒体長) 1 越冬,越夏不能 1 週間未満 1mm 未満 2 年 1 化,休眠性なし 1 週間以上 2 週間未満 1mm 以上 3mm 未満 3 年 2 化以上,休眠性なし 2 週間以上 3 週間未満 3mm 以上 5mm 未満 4 年 1 化.休眠性有り 3 週間以上 4 週間未満 5mm 以上 10mm 未満 5 年 2 化以上.休眠性有り 1 ヶ月以上 10mm 以上 M:生態系影響の程度 M 定着 分散性(導入ステージの移動速度) 寄主範囲(標的種との関係) 1 ごく限られた地域だけ 10㎝ /s 未満(無風,以下同じ) 同属の種 2 10%未満の地域 10㎝ /s 以上 50㎝ /s 未満 同科の種 3 10 以上 25%未満の地域 50㎝ /s 以上 1m/s 未満 同目の種 4 25 以上 50%未満の地域 1m/s 以上 2m/s 未満 同網の種 5 50%以上の地域 2m/s 以上 門内あるいは門をまたがる 表 2 導入天敵昆虫等の生態系影響評価 2 次基準 L:生態系影響の可能性 L 稀少種や地域固有種への直接影響 近縁天敵との競争 近似種との交雑 1 生息場所・時期が異なる 寄主範囲が重ならない なし 2 生息場所・時期が重なることがある 寄主範囲が一部重複 種間交雑率が低い 3 生息場所・時期が常に一部重なる 寄主範囲が 50%程度重複 種間交雑率高,雑種性比が偏る 4 生息場所・時期が 5 割程度重なる 寄主範囲が大部分重複 種間交雑率高,雑種性比は 1:1 5 生息場所・時期が完全に一致する 寄主範囲がほぼ一致 種間雑種は,親より強勢 M:生態系影響の程度 M 稀少種や地域固有種への直接影響 近縁天敵との競争 近似種との交雑 1 稀少種等は寄主となりえない 常に勝敗に優劣なし / 劣勢 形態的な差が大きい 2 稀少種等の近縁種 +30 種以上が寄主 寄主の存在下で共存 雌雄交信信号に種間差がある 3 稀少種等の近縁種 +10 以上 30 種未満が寄主 1 ― 2 種に対しては勝利 近似種の雌が多回交尾可能 4 稀少種等の近縁種 +10 種未満が寄主 数種の相手に勝利 近似種の雌が 1 世代に 2 ― 3 回交尾 5 標的種より稀少種の近縁種を好む 広い範囲の相手に勝利可 近似種の雌が 1 世代に 1 回交尾 第 8 章 外来生物のリスク評価 119 ⑵ 遺伝子情報による外来天敵と近縁土着種の識別法の開発 化学農薬の使用を軽減し,環境に優しい農業を実現する手法のひとつとして,海外で開発された 有能な天敵を輸入し,国内で販売・利用されている例がいくつかあるが,これらの天敵が標的害虫 以外の昆虫を捕食するなど,農業生態系への影響を懸念する声もある.現在,土着天敵ヤマトクサ カゲロウ Chrysoperla nipponensis に近縁な外来種(C. carnea)が輸入され,生物農薬として利用さ れているが,両者は形態形質による外見上の区別が困難である.野外での分布状況や生態系影響を モニタリングするためには,両者を識別できる簡便な手法の開発が不可欠であることから,ミトコ ンドリア遺伝子の部分配列を利用して,この外来種と土着種を容易に識別できる方法を開発した (図 10) (Haruyama et al., 2008) (平成 20 年度研究成果情報,望月ら). また,1940 年以前に日本に侵入したクリ害虫クリタマバチを防除するため,1980 年頃にクリタ マバチの天敵昆虫チュウゴクオナガコバチが海外から導入され,日本各地に放飼された.チュウゴ クオナガコバチの放飼により,クリタマバチの被害は激減したが,チュウゴクオナガコバチと,そ れに近縁な日本土着種クリマモリオナガコバチとの中間的な形態を持つ個体がしばしば見られるよ うになった.両種間の交雑による生態影響が懸念されたが,交雑個体は外見だけでは親種との識別 が困難であることから,核リボソーム遺伝子の違いを利用した交雑個体の検出法を開発した.この 方法により,外来天敵チュウゴクオナガコバチと,それに近縁な土着天敵クリマモリオナガコバチ の交雑個体を容易に検出できるようになり,野外における交雑頻度を簡便かつ的確に把握できるよ うになった(Yara et al., 2009) (平成 21 年度研究成果情報,屋良). PF PR PCR PF ࡊࠗࡑ TACCTCCTTCATTAACTTTAT PR ࡊࠗࡑ PCR Ⴧ DNA(1018bp) TCAGGATAATCAGAATAACGC HpyCH4 V 㒢㉂⚛ಣℂ 㧔⼂㈩㧦TGCA㧕 㔚᳇ᵒേ 1.5㧑ࠕࠟࡠࠬࠥ࡞ 図 10 ミトコンドリア遺伝子の部分配列を用いた土着天敵ヤマトクサ カゲロウと外来近縁種の識別法の概要 6.外来性病害微生物に関する研究 米国産リンゴの火傷病に対する日本の植物検疫措置が科学的証拠に基づいていないとする米国の 主張の根拠となった論文中の侵入リスクの推定技法に関して,感染果実が日本に持ち込まれる確率 の推定法上にいくつもの問題点があったため,輸出元における感染確率のバラツキを考慮した新た 120 7.外来生物データベースの構築 な推定法を提案した.この方法で推定すると,感染果実の侵入リスクはより大きくなることが判明 した.本成果の内容を記載した論文は,日本側の主張の正しさを裏付ける証拠の一つとして WTO パネルに提出された(Yamamura et al., 2001) (平成 15 年度研究成果情報,山村ら). 青枯病菌は熱帯から温帯に広く分布し,5 つのレース(宿主植物に対する寄生性の違い),およ び炭水化物の利用性に基づく 6 つの生理型(biovar)にそれぞれ類別される多様な系統が存在す る.わが国では従来,40 数種の宿主植物が知られ,トマト,ナス等に寄生性のあるレース 1 とジャ ガイモに特異的なレース 3,および生理型 N2,3,4 が在来系統として報告があった.近年,高知 県において新たに 3 種のショウガ科植物(クルクマ,ショウガ,ミョウガ)に青枯病が認められ, 被害が拡大した.そこで,同病原細菌の同定と分子生物学的手法による外国産菌株との比較を行 い,新規系統の由来と伝搬経路について検討したところ,高知県において発生したショウガ科植物 の青枯病菌(Ralstonia solanacearum)は,レース 4 および生理型 4 であった.rep-PCR 解析の結果, タイまたは中国由来と考えられる 2 つの系統が,それぞれ異なる経路で高知県に侵入・伝搬したと 推察された(図 11) (土屋ら,1999) (平成 15 年度研究成果情報,土屋ら). 図 11 rep-PCR(BOX プライマー)による各国産ショウガ科植物青 枯病菌の DNA パターン 7.外来生物データベースの構築 近年,世界的な物流の拡大とともに侵略的外来生物が増加し,世界各国で農作物への直接被害や 固有の生物多様性に対する影響が問題となっている.外来生物の侵入・蔓延防止と防除は世界的な 緊急課題であり,わが国と地域的にも関係が深いアジア・太平洋諸国においても例外ではない.周 辺各国で外来生物の動態や分布状況,被害実態を互いに共有することは,外来生物の侵入防止対策 や防除対策を立てる上で有益であることから,2003 年に「アジア・太平洋諸国における侵入生物 の生態影響とデータベース構築」と題する国際セミナーを開催し,参加各国で侵入生物に関する 情報の交換を行うとともに「アジア・太平洋外来生物データベース(APASD)」を構築した(図 12) .その後 2004 年および 2007 年にも同様の国際セミナーを開催し,情報交換とデータベースの 充実に向けた検討を行った.本データベースには,外来生物の分類名,定着状況,分布拡大,経済 第 8 章 外来生物のリスク評価 121 的生態的被害,繁殖・生育特性,防除対策,文献,写真などの情報が格納されており,新たな情報 の入力や必要とする情報の検索・閲覧をインターネットを通じて行うことができる(Yamanaka et al., 2003) (平成 16 年度研究成果情報,山中ら) .2011 年 2 月現在で,種の登録は植物 771 種,昆 虫約 599 種,その他約 51 種であり,上記の詳細情報については,植物 73 種,昆虫 139 種,その他 13 種で閲覧することができる. 図 12 アジア・太平洋外来生物データベース(APASD)の種別詳細データの例 8.今後の展望 農業環境技術研究所では,これまで外来生物の生育・繁殖特性等の解明や生態系影響リスクの評 価法の開発などに取り組んできた.経済のグローバル化による貿易量の拡大は,非意図的な外来生 物の侵入を今後も引き起こすことは間違いないと思われる.生物多様性国家戦略 2010(環境省, 2010)では,外来生物の影響は,わが国の生物多様性を脅かす要因のひとつとしてあげられている が,外来生物の生物多様性に与える影響が不可逆的であることを考慮すると,長期的にみれば生息 環境の破壊よりその影響は深刻である(村上ら,2002)とも言われており,生物多様性の保全を図 122 引用文献 るためには,外来生物対策は今後も取り組むべき大きな問題である. 「外来生物法」により,特定外来生物の輸入や飼養,栽培等が規制され,野外への放逐等が禁止 されることとなったが,防除や警戒のための実施体制が法律的に確保されておらず,実効性に乏し いといった問題点も指摘されている(西田,2010).今後,外来生物の管理を実効あるものとする ためには,主要な外来生物について,生息実態や生態的特性の解明,分布拡散予測,生態系影響評 価等を実施することにより,防除優先順位を決定するとともに,効率的な防除法の開発が不可欠で ある. このほか,地域固有の生物多様性保全の観点から,国外からの外来種の移動のみならず,国内の 生物移動による「国内外来種」の問題や,近隣国で採取された日本在来種と同種の植物を土木工事 の際の法面緑化などに利用して問題となっている「外国産在来種」の問題など,取り組むべき課題 は多い.さらに,天敵や花粉媒介虫,緑化植物など,その有用性を目的に,意図的に導入される外 来生物については,生態系影響に対するリスクのみを問題にするのではなく,それらが本来持って いる有用機能による便益も考慮した上で,導入の可否を判断する必要があるとの意見も強い.生態 系への影響を便益と比較可能な形で評価することが難しく,リスクと便益を考慮した総合評価は現 時点では困難であるが,将来的には避けて通れない問題であろう. 引用文献 1)藤井義晴(2008)外来植物のリスクを評価し,その蔓延を防止する.日本農学会編/外来生物 のリスク管理と有効利用,養賢堂,19-59 2)Haruyama, N., Naka, H., Mochizuki, A. and Nomura, M.(2008)Mitochondrial phylogeny of cryptic species of the lacewing Chrysoperla nipponensis(Neuroptera: Chrysopidae)in Japan. Entomological Society of America, 101 (6) , 971-977 3)平舘俊太郎,楠本良延,吉武 啓,馬場友希(2010)土壌が支える生物多様性.根本正之編著 /身近な自然の保全生態学:生物の多様性を知る,培風館,131-148 4)平舘俊太郎,森田沙綾香,楠本良延(2008)土壌の化学的特性が外来植物と在来植物の住み分 けに与える影響.農業技術,63 (10) ,469-474 5)伊藤健二(2008)利根川水系におけるカワヒバリガイ Limnoperna fortunei の分布状況.日本ベ ントス学会誌,63,30-34 6)伊藤健二(2010)関東地域における特定外来生物カワヒバリガイの現状と侵入・拡大プロセ ス.Sessile Organisms,27 (1) ,17-23 7)角野康郎(2010)外来植物研究への期待−種生物学の課題.種生物学研究,33,11-23 8)環境省(2010)生物多様性国家戦略 2010. 9)望月 淳(2008)導入昆虫のリスク評価とリスク管理 ― 導入天敵のリスク評価と導入基準 ― .日本農学会編/外来生物のリスク管理と有効利用,養賢堂,171-185 10)森田竜義(1997)世界に分布を広げた盗賊種セイヨウタンポポ.山口裕文編/雑草の自然史, 北海道大学図書刊行会,192-208 11)村上興正,鷲谷いづみ(2002)外来種問題に対する国際的認識の高まり.日本生態学会編/外 来種ハンドブック,地人書館,5 12)西田智子(2010)外来生物法とこれからの外来種対策 ― 植物の場合 ― .種生物学研究,33, 345-365 13)Nishida, T., Yamashita, N., Asai, M., Kurosawa, S., Enomoto, T., Pheloung, P. C. and Groves, R. 第 8 章 外来生物のリスク評価 123 H.(2009)Developing a pre-entry weed risk assessment system for use in Japan. Biological Invasions, 11, 1319-1333 14)芝池博幸(2005)無融合生殖種と有性生殖種の出会い ― 日本に侵入したセイヨウタンポポ の場合 ― .生物科学,56,74-82 15)芝池博幸,植竹朋子,楠本良延,矢野初美,井手 任(2005)関東地方における雑種性タンポ ポの遺伝構造とクローン分布.環境情報科学論文集,19,279-284 16)Shimono, Y. and Konuma, A.(2008)Effects of human-mediated processes on weed species composition in internationally traded grain commodities. Weed Research, 48, 10-18 17)Tanaka, K.(2009)Genetic Variation in Flight Activity of Ophraella communa(Coleoptera: Chrysomelidae) : Heritability Estimated by Artificial Selection. Environmental Entomology, 38, 266-273 18)Tanaka, K. and Yamanaka, T.(2009)Factors Affecting Flight Activity of Ophraella communa (Coleoptera: Chrysomelidae) , an Exotic Insect in Japan. Environmental Entomology, 38, 235-241 19)Tominaga, A., Goka, K., Kimura, T. and Ito, K.(2009)Genetic structure of Japanese introduced populations of the golden mussel, Limnoperna fortunei, and the estimation of their range expansion process. Biodiversity, 10 (2&3),61-66 20)土屋健一,矢野和孝,堀田光生,森田泰彰,川田洋一,d'Usel C.M.(1999)わが国におけるショ ウガ青枯病(Bacterial wilt:新称)の初発生(講要).日植病報,65,363 21)Yamamura, K., Katsumata, H. and Watanabe, T.(1999)Estimation of the probability of insect pest introduction through imported commodities. Researches on Population Ecology, 41, 275-282 22)Yamamura, K., Katsumata, H. and Watanabe, T.(2001)Estimating invasion probabilities: a case study of fire blight disease and the importation of apple fruits. Biological Invasions, 3, 373-378 23)Yamanaka, T. and Liebhold, A. M.(2009a)Mate-location failure, the Allee effect, and the establishment of invading populations. Population Ecology, 51, 337-340 24)Yamanaka, T. and Liebhold, A. M.(2009b)Spatially implicit approaches to understand the manipulation of mating success for insect invasion management. Population Ecology, 51, 427-444 25)Yamanaka, T. and Matsui, M.(2003)Development and utilization of APASD(Asian-Pacific Alian Species Database) . Proceedings of the NIAES-FFTC Joint International Seminar on Biological Invasions, 155-176 26)山野美鈴,芝池博幸,浜口哲一,井手 任(2002)「身近な生きもの調査」を利用したタンポ ポ属植物の雑種分布に関する解析.環境情報科学,16,357-362 27)Yara, K., and Kunimi, Y.(2009)Discrimination of Torymus sinensis Kamijo(Hymenoptera: Torymidae)and T. beneficus Yasumatsu et Kamijo and their hybrids by allele-specific PCR. Applied Entomology and Zoology, 44 , 275-280 124 第 9 章 遺伝子組換え作物の生態系影響評価と共存のための管理手法の開発 1.はじめに 農業環境技術研究所は,わが国ではじめての隔離ほ場を保有し,1991 年に遺伝子組換えトマト の模擬的自然環境における環境影響評価を実施した . 一方,海外からナタネ,ダイズ,トウモロ コシを中心に除草剤耐性,害虫抵抗性の遺伝子組換え作物(GMO)がわが国に輸入されるように なり,食品としての安全性に関する国民の関心と平行して,環境への影響にも関心が高まりつつあ る . また,世界の遺伝子組換え作物の商業栽培面は,栽培が開始された 1994 年以来,年々増え続 け,2009 年の遺伝子組換え作物の作付け面積は,29 か国,のべ 1 億 3 千 4 百万ヘクタールに達し ている . このように遺伝子組換え作物の栽培面積が増加しつつあるなか,わが国においても GMO が輸入され,利用される機会が多くなっている . 欧州連合(EU)においても徐々にではあるが栽培が広がっている . これまで GMO に対しては モラトリアムなどにより非常にネガティブと見られていた EU であるが,一部の国では生産が開始 され,栽培面積が増加している . 欧州委員会は,2003 年 7 月,「遺伝子組換え作物と慣行・有機 農業との共存に関するガイドライン」 (2003/556/EC)を公表し,共存に関する基本的原則および 考慮すべき観点や具体的な交雑・混入を防止する手法を提示している(立川ら,2009). このような GMO に関する世界情勢にかんがみ,農環研では 2004 年のカルタヘナ議定書の批准 にともない,これまで行ってきた遺伝子組換え作物の環境影響評価そのものから,科学的な評価が 実施されるように,研究をシフトさせた . すなわち,遺伝子組換え作物の適正な評価手法の確立 と評価実施に必要な科学的知見を集積するとともに共存研究が推進されることになった . 2.ほ場条件下における遺伝子組換えダイズとツルマメとの自然交雑 ツルマメは,栽培ダイズと同じ Glycine 属,Soja 亜属に属し,ダイズの祖先種と考えられている つる性の一年草(図 1)で,朝鮮半島,中国,シベリア東部,日本では北海道から九州までに分布 さや する(大橋,1997). 種子と莢はともに栽培ダイズより小さく,種子は黒色である(図 2). 図 1 研究所(茨城県つくば市)内の 自生ツルマメ 図 2 遺伝子組換えダイズ(AG3701RR) およびツルマメの莢と種子 ダイズとツルマメは,人工授粉などによって,相互に交雑できるが(Karasawa, 1936; Kwon et al., 1972; Oka, 1983) ,ダイズは,株内混植においても,その他殖率が 2%以下の自殖性植物であり (Cutler,1934; Weber et al., 1961; Chiang et al., 1987; Ray et al., 2003),開花時にはすでに受粉して 第 9 章 遺伝子組換え作物の生態系影響評価と共存のための管理手法の開発 125 いる場合が多く(Free, 1970; 古谷 , 1980) ,ツルマメとの自然交雑はきわめて低いと考えられた . 一方,遺伝子組換えダイズ(以下組換えダイズ)は,そのほとんどが除草剤耐性品種であり,世 界のダイズ栽培面積の 77%を占めている(James,2009). 日本国内では,現在,試験栽培以外の 遺伝子組換え作物は栽培されていないが,現在最も普及している除草剤耐性の組換えダイズが,将 来,栽培された場合には,そのほ場周辺に自生するツルマメと交雑し,除草剤耐性のツルマメが発 生し,それが広がるのではないか? また,野生集団中の遺伝的多様性が損失するのではないか? と懸念する声がある.これまで,ダイズ同士の他殖率については,多くの報告(Woodworth, 1922; Cutler, 1934; Weber et al., 1961; Caviness, 1966; Chiang et al., 1987; Arent et al., 1994; Ray et al., 2003; Yoshimura et al., 2006)があり,ツルマメ同士の他殖率の報告(Fujita et al., 1997)もある が,ダイズとツルマメとの交雑に関する報告は少ない.Nakayama et al.(2002)は,野外でダイ ズとツルマメを 50cm 間隔で市松模様に配置して,0.73%の交雑率を報告している.また,Abe et al.(1999)は,全国から収集した 708 個体のツルマメの葉緑体 DNA の RFLP 解析から,日本の改 良品種由来の遺伝子を 1.8%の頻度で認めている.また,Dorokhov et al.(2004)は,除草剤耐性 ダイズとツルマメを 2 年間ほ場で栽培したが,交雑個体は得られなかったとし,人工授粉で得た F1 では,組換え DNA の断片が検出されたが,F2,F3 では,その断片がなくなったと報告してい る.このように,ダイズとツルマメの交雑についてはいくつかの報告はあるものの,組換えダイズ を使った試験では,ツルマメとの交雑個体は得られておらず,交雑個体の適応度,組換え遺伝子の 遺伝子浸透などについては,さらなる科学的知見の集積が必要であった. 「組換えダイズが,近縁野生種との交雑によって生物多様性に影響を与える」とした場合,その 最初のステップは, 「遺伝子組換えダイズほ場からその周辺に自生するツルマメへの花粉流動によ る交雑」となると考えられ,吉村らは,このステップに注目し,「ほ場規模で組換えダイズが栽培 された場合に,どのくらいの頻度で,あるいは,どのくらいの範囲までツルマメとの交雑が起こる のか?」について 2005 年から 2007 年の 3 年にわたって試験を行った. 両親の花が同時に咲いていることは,その自然交雑が起こるための必須の条件であり,同時に開 花する期間は,交雑頻度に与える重要な要因であると考えられるが,ツルマメの開花期は,ダイズ より遅く,関東地方では,1 か月ほどの差が見られた.そこで,本研究では,組換えダイズの播種 おくて 日を遅らせ,晩生の組換えダイズ品種を用いて,両種の開花期を近づける処理を行い,からみつく ほど近くで栽培して,より交雑しやすい条件を設定した.収穫したツルマメは閉鎖系温室内で播種 し,実生の葉(3-5 葉期)にグリホサートを散布し,耐性個体を選抜し,生き残った個体に耐性個 体固有のタンパク質と DNA 断片の存在を確認し,自然交雑率を評価した . 農業環境技術研究所 (茨城県つくば市)内の一般試験ほ場で栽培された組換えダイズは,除草剤グリホサート耐性組換 えダイズ(40-3-2 系統)で,食品安全性,飼料安全性が確認され,カルタヘナ法に基づいて第 1 種 使用規程が承認された組換えダイズであり,その栽培については,「第 1 種使用規程承認組換え作 物栽培実験指針」の交雑防止措置に従った. 2005 年は,除草剤グリホサート耐性組換えダイズ品種 AG3701RR およびツルマメ各 36 個体を 栽培し 7 月下旬から 9 月下旬に 1 〜 3 日ごとに開花数を計測した.また,組換えダイズとツルマメ (60 組)をからみつくほど近接して栽培したところ(図 3,4),遅く播種した遺伝子組換えダイズ ほどツルマメの開花のピークが近くなり,最も遅く播種(7 月 20 日)したダイズと近接して栽培 したツルマメ種子の中に 1 個体の交雑個体を得た(Mizuguti et al., 2009)(平成 18 年度研究成果情 報,吉村ら) .2006 年は,当研究所内の一般試験ほ場(20a)で,遺伝子組換えダイズの間にから みつくほど近づけてツルマメを網状の壁に沿わせて栽培する混植区および遺伝子組換えダイズから 126 2.ほ場条件下における遺伝子組換えダイズとツルマメとの自然交雑 䉿䊦䊙䊜 ⚵឵䈋䉻䉟䉵 図 3 2005 年のほ場配置 図 4 2005 年の栽培の様子 (8 月 31 日撮影) 2, 4, 6, 8, 10m 離してツルマメを網状の壁に沿わせて栽培する距離区を設けた(図 5,6). 組換えダイズには,開花期の異なる 4 品種(40-3-2 系統 : AG3602RR,AG3906RR,AG4801RR, AG5905RR)を栽培し,ツルマメは,網状の壁に沿わせて栽培した.収穫したツルマメ種子,計 112,469 個(うち 44,348 個が混植区)を検定し,自然交雑個体は得られなかった. 図 6 2006 年の栽培の様子 (7 月 27 日撮影) 図 5 2006, 2007 年の実験ほ場配置 2007 年は,2006 年度と同様のほ場配置で,開花期の遅い組換えダイズ 2 品種(AG5905RR およ び AG6702RR)を 6 月 20 日およびそれから 1 か月遅らせて 7 月 19 日に播種する区を設けた.組 換えダイズ,ツルマメの開花は,早いものから 6 月播種 AG5905RR,6 月播種 AG6702RR,7 月播 種 AG5905RR,7 月播種 AG6702RR,ツルマメの順であり,2007 年度は,2006 年度に比べ,両種 の開花ピークが近くなり,開花が重複する期間も長くなった.その結果,混植区のツルマメ種子, 第 9 章 遺伝子組換え作物の生態系影響評価と共存のための管理手法の開発 127 25,741 個を検定し,35 個体の交雑個体を得た.距離区(検定個体数 66,671)では,2,4,6m の区 で各 1 個体ずつ交雑個体を得た(Mizuguti et al., 2010). 結論として,組換えダイズとツルマメの開花期をある程度,人為的に重複させ,からみつくほど 近くで栽培すると,交雑は 1000 分の 1 程度生じる可能性があることを示したが,雑草管理をとも なう一般のダイズ栽培では,これよりさらに交雑頻度は低くなると考えられた.また,2006 年の 結果は,組換えダイズとツルマメの開花期がほとんど重複しなければ,自然交雑の可能性はほとん どないことも示しており,地域によって組換えダイズ品種を選び,開花期の重複を低下させると, ツルマメとダイズとの自然交雑率をゼロに近づけることができる.すなわち,品種の選択や除草方 法を含む栽培方法の選択によって,日本では,ツルマメへの生物多様性影響を防ぎながら組換えダ イズを栽培できると考えられた.これらの知見は,生物多様性評価情報として行政部局への貢献を 果した. 3.遺伝子組換えナタネのモニタリング ⑴ カナダのナタネ輸出港周辺における遺伝子組換えナタネの生育分布調査 日本はナタネの 99%以上を輸入しているが,そのうちの 80%はカナダからの輸入である . カナ ダでは,栽培面積の 8 割以上が遺伝子組換え除草剤耐性(GM)ナタネである(Yoshimura et al., 2006) ,わが国のいくつかのナタネ輸入港周辺で除草剤耐性遺伝子組換えナタネ(以下組換えナタ ネ)の生育が報告され(Saji et al., 2005; Aono et al., 2006),その分布拡大が懸念された. カナダでは,組換えナタネを含むセイヨウナタネは西部のプレーリー地帯で生産され,貨物列車 でバンクーバー港の穀物ターミナルへ輸送された後,そこから日本などへ船舶で輸出されている. その輸送中には,日本と同様に組換えナタネがこぼれ落ちによって野外へ拡散している可能性が あった.そこで,2005 年に吉村らは,カナダ農務省との共同研究を行い,輸出港周辺における組 換えナタネの分布状況から,育成地の拡大する可能性を予測できると考え,バンクーバーにおける 調査を実施した. 調査の結果,組換えナタネは,バンクーバー港や市街の線路と線路近くの道路に生育しており, 特に北側のバラード入江の穀物ターミナル付近で多く見られた(図 7).また,ナタネ群落の密度 およびその面積は穀物ターミナルにおいて特に大きい傾向があった(図 8). 図 7 バンクーバー港周辺の路線や道路周辺の除草剤 耐性遺伝子組換えおよび非組換えナタネの分布 ٨㧦 〝ㄝߩㆮવሶ㧘⚵឵߃࠽࠲ࡀណขὐ ع㧦 ✢〝ㄝߩㆮવሶ⚵឵߃࠽࠲ࡀណขὐ ਛⓨ߈ߩශߪߘࠇߙࠇߩ㕖ㆮવሶ⚵឵߃࠽࠲ࡀណข ὐ 128 3.遺伝子組換えナタネのモニタリング 図 8 セイヨウナタネの群落の個体密度および面積と,各群落と最も近い穀物ターミナルとの距離の 関係(2005 年) サンプリングした個体の 65%は組換えナタネであり,グリホサート耐性の方がグリホシネート 耐性の個体数より多く,特に,線路周辺で採取したナタネの各耐性の割合は,各耐性別栽培割合と よく似ていることから,線路周辺のこぼれ落ちナタネの割合は,栽培地域のナタネ品種の割合を反 映していると考えられた(表 1) . 表 1 グリホサート(RR),グリホシネート(LL)耐性ナタネおよび非組換えナタネの割合とカナダ産ナタネの耐性別栽培割合 調査地数 採取数 線路周辺 28 46 48 22 30 道路周辺 26 35 37 23 40 平均 27 40 43 22 35 48 22 30 (%) カナダ産ナタネの耐性別栽培割合(2003 年) RR 耐性ナタネ(%) LL 耐性ナタネ(%) 組換えナタネ(%) カナダでは組換えナタネを栽培し始めてすでに 10 年が経っており,現在のナタネ栽培面積の 80%以上を占めていたが,仮に遺伝子組換えナタネが他種を駆逐し,おう盛な繁殖力を持つのであ れば,すでに放棄地,河原などに大きな群落が見られると推測された.しかし,バンクーバー全域 にわたって調査を行ったが,その生育場所は鉄道線路やその道路周辺に限られていた . この調査 によって,こぼれ落ち種子からの組換えナタネを含むナタネの生育範囲は限定されていることが確 認され,輸入国であるわが国においても組換えナタネの分布が拡大する可能性は低いことを予測で きた(Yoshimura et al., 2006) (平成 18 年度研究成果情報,吉村ら). ⑵ 鹿島港で野生化した遺伝子組換えナタネの生育分布調査 これまで鹿島港を含む日本各地のナタネ輸入港では,野生化したセイヨウナタネの生育が報告さ れている(Saji et al., 2005; Aono et al., 2006) .セイヨウナタネ(遺伝子組換えセイヨウナタネを含 む)は,鹿島港周辺の主要な道路沿いに生育しており,主要物流道路以外にはほとんど生育しない こと等から,原材料用として輸入された種子の輸送時におけるこぼれ落ち由来のものであると推測 されている.原料として輸入された遺伝子組換えセイヨウナタネ(以下組換えナタネ)の生育が, 主要な輸入港である茨城県鹿島港周辺で確認されている.組換えナタネは,カルタヘナ法に基づく 第 9 章 遺伝子組換え作物の生態系影響評価と共存のための管理手法の開発 129 生物多様性影響評価により,野生化して生育する ことも前提として,遺伝子組換えでないセイヨウ ナタネ(以下非組換えナタネ)と差異がないこと から,生物多様性影響を生ずる恐れがないと判断 されている.この評価は畑やポット条件での調査 試験に基づくものであるため,松尾らは,実際に 輸入港周辺に生育した場合,組換えナタネが非組 換えナタネより広い範囲に優先して生育するかど うかを明らかにするため調査を行った. 茨城県鹿島港周辺の要道路沿いにおいて原材料 の陸揚げ地点を中心とした半径 5km 圏内でセイ ヨウナタネが生育していた 19 地点を調査地とし て設けた(図 9) . これらのセイヨウナタネの生 育地は,土壌が厚く生育できる面積が広い「空 図 9 鹿島港周辺の調査地点 あぜ き地や水田畦」 ,土壌は厚いが生育できる面積が 狭い「植栽帯や中央分離帯」 ,土壌も薄く面積も狭い「縁石沿い」に分類できた(表 2A).多くの 調査地点では,これら異なるタイプが複数存在した . また,生育していたナタネが組換えナタネ かどうかを調べるために,各調査地点で葉を採取した . 日本に多く輸入されている組換えナタネ は,除草剤グリホサート耐性と除草剤グルホシレート耐性の組換えナタネであることから,それぞ れが持つ特有のタンパク質の有無を調べた . その結果,組換えナタネは非組換えナタネと同様に 空き地や水田畦,植栽帯や中央分離帯,縁石沿いに生育し,多くの場合その個体数は非組換えナタ ネと同程度かあるいは少ないことが明らかとなった . 以上のことから,輸入港において,組換え ナタネは非組換えナタネの育成地にしか生育してないことが明らかとなり,組換えナタネが非組換 えナタネより広い範囲を優先して生育することはないと考えられた . この結果は,実際に日本国 内の輸入港において,野生化した組換えナタネの育成地の特性を示す唯一の知見であり,組換えナ タネの生物多様性影響評価を裏付ける知見であった . (平成 18 年度研究成果情報,松尾ら) 表 2 セイヨウナタネが生育する調査地点における生育地の特性(A)と組換えナタネ特有のタンパク質の検定結果(B) (A) 生育地 の特性 (B) 検定結果** 調査地番号* 15 16 14 空き地,水田畦(タイプⅠ) ○ ○ ○ 植栽帯,中央分離帯(タイプⅡ) ○ 縁石沿い(タイプⅢ) ○ 5 18 ○ ○ 1 2 3 4 6 9 10 19 21 11 12 13 17 20 ○ ○ ○ ○ ○ ○ ○ ○ ○ ○ ○ ○ ○ ○ ○ ○ ○ ○ ○ ○ ○ ○ ○ RUR(−), LL(−) 25 38 39 11 28 10 3 1 55 10 6 16 45 3 2 4 41 60 37 RUR(+), LL(−) 0 24 1 1 3 0 0 2 2 2 0 1 0 2 0 0 0 1 0 RUR(−), LL(+) 0 5 3 0 7 0 0 0 0 6 0 1 0 3 1 0 1 4 0 RUR(+), LL(+) 0 2 0 0 3 0 0 0 0 0 0 0 0 0 0 0 0 0 0 合 計 25 69 43 12 41 10 3 3 57 18 6 18 45 8 3 4 42 65 37 RUR (−), LL (−) : 非 GM セイヨウナタネ(→普通のセイヨウナタネ) RUR (+), LL (−) : 除草剤グリホサート耐性タンパク質の検出→グリホサート耐性 GM セイヨウナタネ RUR (−), LL (+) : 除草剤グルホシネート耐性タンパク質の検出→グルホシネート耐性 GM セイヨウナタネ RUR (+), LL (+) : 除草剤グリホサート耐性タンパクトおよびグルホシネート耐性タンパクの検出 →両除草剤に耐性を有する GM セイヨウナタネ 130 4.遺伝子組換え作物と一般栽培作物等との共存のための管理手法開発 さらに,セイヨウナタネが周辺群落に侵入し,他の植物を駆逐して生育域を拡大することがない かを確認する追加調査を実施した(追加調査:2006 年 7 月〜 2007 年 6 月).調査では,図 9 に示 した A 地点(No.6)と B 地点(No.14)を対象とした. 2006 年度からは,セイヨウナタネが周辺群落に侵入し,他の植物を駆逐して生育域を拡大する かどうかを確認するための追跡調査をした.図 9 の 19 調査地点のうち,セイヨウナタネ以外の植 物群落が広く存在し,セイヨウナタネとの競合が観察できる 2 地点(A 地点,B 地点)を調査した. その結果はそれぞれの地点において,他の植物群落が広く存在し競合が起こる空き地では,セイヨ ウナタネの生育は全く確認されなかった.搬送途中でこぼれ落ちたナタネの多くは,縁石沿いにた まった土砂の上や草刈りが行われる中央分離帯など,道路特有のかく乱の多い限られた場所には生 育したが,周辺群落へ積極的に侵入し広がっていく傾向は見られなかった.つまり,セイヨウナタ ネは周辺群落に侵入した場合でも,競合により他の植物を駆逐して生育域を拡大することはないと 考えられた.このような結果から,これまでの生物多様性影響の評価の考え方や内容は適切である と確認された(平成 22 年度研究成果情報,松尾ら). 4.遺伝子組換え作物と一般栽培作物等との共存のための管理手法開発 将来の遺伝子組換え作物の商業栽培に備え,関連の科学的データを蓄積するとともに,交雑・混 入の抑制技術を開発し,行政部局による栽培実験指針等の検討に資する知見を提供するため,研究 が進められてきた. ⑴ 風によるダイズ花粉の飛散に関する研究 現在,日本では遺伝子組換えダイズは一般栽培されていないが,今後組換えでないダイズを作る 農家との共存を実現するためのルールづくりが必要になると考えられる.ダイズは自殖性の作物で あり,0.7m 程度離して栽培した場合でも他殖率は 0.5%以下と低い.他殖は,虫媒によるものと考 えられているが,媒介する昆虫は特定されておらず,風媒による可能性も否定できない.そこで吉 村らは,ほ場および風洞施設においてダイズ花粉の飛散距離を調査し,その風媒の可能性について 検討した.長さ約 20m の風洞施設(図 10)を用いた花粉飛散実験では,風上に開花植物,風下に ワセリンを塗布したプレパラートを配置し,風速 2.0m/s で 15 分間空気を送風した後,プレパラー ト上に付着した花粉をカウントした. 図 10 風洞施設実験の様子 風上にダイズ個体を , 風下にワセリンを 塗布したプレパラートを乗せたスタンド を一定距離ごとに配置した. 落下花粉密度は,イネでは距離とともに減少し,風下 0.2m で 0.63 個 /cm2 であったのに対し, ダイズでは最も密度が高い風下 2.5m でも 0.08 個 /cm2 であった.また,ダイズ花粉の直径から計 算すると,本実験条件では,推定飛散距離は 15m 以上となったが,実際は 10m 地点でわずかに観 第 9 章 遺伝子組換え作物の生態系影響評価と共存のための管理手法の開発 131 察されたのみであり,15m 地点では観察されなかった(図 11).以上の結果に加え,めしべが花弁 の外に露出しないというダイズの特徴を考慮すると,ほ場外のダイズが風媒により受粉する可能性 はほとんどないと推察された(平成 22 年度研究成果情報,吉村ら).以上の結果は,今後遺伝子組 換えダイズを栽培する際,非組換えダイズとの交雑防止手段の開発に寄与するものと考えられる. 図 11 風洞施設におけるダイズおよびイネの落下花粉密 度と花粉直径から予測された花粉の最大飛散距離 予測されたダイズ花粉の最大飛散距離は , 15m 以上 だが,15m 地点では花粉は観察されなかった. ⑵ 遺伝子組換えトウモロコシの花粉飛散に関連する研究 Bt タンパク質遺伝子が導入された耐虫性遺伝子組換えトウモロコシ(以下,Bt 組換えトウモロ コシ)が米国で商業栽培されると,花粉がほ場内外に飛散し,オオカバマダラに影響を与える可能 性が報告された . わが国においても,栽培用 Bt 組換えトウモロコシの花粉が非標的鱗翅目昆虫に 及ぼす影響を評価する際に,Bt 花粉の毒性レベルを知る必要があり,そのために生物検定法の確 立が求められた . 松井らは,採集や入手が比較的容易なチョウ類やガ類からヤマトシジミを選定 した . 具体的には,ヤマトシジミは,Bt タンパク質(Cry1Ab)遺伝子を組換えたトウモロコシ の花粉に対して感受性が高く,Bt 花粉の非標的鱗翅目昆虫への毒性を推定するのに適していた . 検定品種の花粉密度を変えて生葉片上に載せ,ヤマトシジミ 1 齢幼虫に 5 日間連続摂食させ,その 毒性を死亡率でみられることを確認した(平成 13 年度研究成果情報,松井ら) . 遺伝子組換え作物を開放系で栽培した場合,同種 の非組換え作物と交雑することにより,花粉を介し た導入遺伝子の拡散が懸念される.このような導入 遺伝子の拡散を正確に,かつ簡便に定量する手法と して,松尾らは,トウモロコシの種子色に発現する キセニアを利用して交雑種子を判別することによ り,遺伝子組換えトウモロコシを栽培するほ場から 花粉が拡散する範囲を把握し,花粉源からの距離と 交雑率の関係を評価する手法を開発した(平成 14 年度研究成果情報,松尾ら) .また,川島らは,空 中を浮遊するトウモロコシ花粉を捕集し,それらを 光学的に検出することにより,花粉飛散量を簡易か 図 12 ダーラム法による花粉計測値と花粉モニ ター装置による花粉飛散量の関係 132 4.遺伝子組換え作物と一般栽培作物等との共存のための管理手法開発 つ連続的に自動計測できるモニター装置を開発した.この装置により検出された花粉飛散量は,空 中花粉の基準的測定法として広く用いられているダーラム法による花粉計測値と高い相関関係を示 したことから,トウモロコシ花粉を含めて大気中を拡散する微粒子のモニタリング装置としての活 用が期待される(図 12) (平成 15 年度研究成果情報,川島ら). 遺伝子組換え作物の商業栽培に際しては,同種の非組換え作物と交雑しないように,それぞれの ほ場を隔離することが有効である.2004 年 2 月に策定された「第 1 種使用規程承認組換え作物栽 培実験指針」によると,研究開発中の遺伝子組換えトウモロコシを栽培する際には,近隣の非組換 えトウモロコシほ場との間に 600m の隔離距離を確保することが求められている.このような隔離 距離と交雑率の関係を評価するためには,生物の拡散過程に関する理論的な研究が不可欠である. 村山は,ブラウン運動モデルを基に,花粉が飛散する際の 1 単位の長さを確率的に変動するように 改良することで,風などの環境のゆらぎによりランダムに拡散する花粉の飛散距離を高い精度で推 定する予測式を開発した(図 13) . ƦǕ ᤨ㑆㑆㓒ߩࡦ࠳ࡓ࠙ࠜࠢߦ߅ߡ㧘Ǖ ᤨὐߩ㧝ᱠߩ㐳ߐ ƦrǕ ߩ⏕₸ಽᏓߪ (1) ߎߎߦ m, v ߪቯᢙߢࠆ☳⧎ޕḮ߆ࠄ(x, y) 㔌ࠇߚὐߢߩ T ᤨ㑆ᓟߩ⏕₸ኒᐲߪᰴᑼ ߣߥࠆޕ (2) ߚߛߒ k = vT/ƦǕ㧘nj = 4m ߢࠆޕGx㧘Gy ߪߘࠇߙࠇ x, y ᣇะ߳ߩᵹࠇࠍ␜ߔቯᢙߢࠆޕ 図 13 環境のゆらぎを考慮した拡散予測式の概要 この予測式を用いて「第 1 種使用規程 承認組換え作物栽培実験指針」で示され るトウモロコシの隔離距離を評価した結 果,一般的な気象条件で 600m 地点にお ける交雑率は 0.125%程度と低く,隔離 距離がおおむね妥当であることを裏付け た(平成 16 年度研究成果情報,山村) . 上記指針には,除雄やネットの使用等 による隔離距離によらない交雑防止措置 も示されているが,適用条件の詳細につ いては明らかにされていない.そこで, 防風施設を利用した様々な交雑抑制策の 図 14 防風ネットおよび防風植生(ソルガム)を用いたト ウモロコシ 効果を比較し,交雑抑制効果の高い開花 期調整や防風施設の特性を明らかにした(図 14) . 第 9 章 遺伝子組換え作物の生態系影響評価と共存のための管理手法の開発 133 また,これらの効果を予測するためのシミュレーションモデルを構築した.また,防風網のメッ シュの大きさの違いによる比較を行い,メッシュの大きさによる交雑抑制効果も推定した.一方 で,これらの交雑抑制策は近距離交雑を効果的に抑制できる反面,低頻度で生じる遠距離交雑を抑 制する効果はあまり大きくないことが示された(牛山ら,2009a; Ushiyama et al. 2009b)(平成 22 年度研究成果情報,杜ら) . ⑶ 大規模ほ場におけるイネ花粉の長距離飛散と交雑の実態に関する研究 これまでの遺伝子組換え作物(GM 作物)の花粉飛散や交雑に関する研究は,GM 作物の開発に ともなう小規模な試験栽培を円滑に行うために必要な情報を収集することが主たる目的であった. しかし,GM 作物の商業栽培に備えて,GM 作物と同種の非 GM 作物との共存を図るための行動指 針を確立するためには,農家ほ場規模で発生する花粉飛散や交雑の実態を把握することが重要と考 えられる.ここでは水稲栽培農家のほ場を借り上げ,農家による一般的な水稲栽培を行った場合の 自然交雑の実態を把握した.茨城県つくばみらい市の農家ほ場において風上側に花粉親区(ウルチ 品種:約 0.2ha) ,風下側に種子親区(モチ品種:約 1.0ha)を設置し,種子親区中に 30 カ所の方形 区を設け 3,050 株の調査株を採集した.キセニアを発現した交雑種子を選別した後,DNA 鑑定に より想定される花粉親に由来する交雑であるかを確認した.その結果,花粉親区に最も近接した領 域における平均交雑率が最も高く,花粉親から遠ざかるにつれて平均交雑率は急激に低下すること が明らかとなった(図 15) .花粉源となるほ場の周辺では 0.005%程度の自然交雑率に落ち着くこ とが示された . (平成 22 年度研究成果情報,芝池ら). ���� ����� ����� ����� ����� ����� ����� ���� �������� ���� ���� 図 15 茨城県つくばみらい市の農家ほ場 で観察された水稲の自然交雑率と 95%信頼区間 (平成 20 年度は太実線,21 年度は 細実線で示した) ���� ���� ���� ���� � � �� �� �� �� ��� ����������� 遺伝子組換え作物を導入し,一般栽培作物との共存を実現しようとする場合には,導入を図る地 域全体で発生する交雑や混入の総量を把握し,それらを低減するための取り組みが必要になると考 えられる . ここでは,ある地域に存在するほ場の位置や面積,そこで作付けされるイネ品種や開 花期,気象条件などを含むほ場環境 GIS データベースを構築し,データベースから必要な情報を 取り出すことにより,ある地域内で発生する交雑や混入の総量を評価・予測するためのシミュレー ション・プログラムを作成した.秋田県大潟村,岩手県雫石町,茨城県つくばみらい市の 3 地域 (各 4km×3km)を対象としたほ場環境 GIS データベースを構築し,それぞれの地域内で発生する 134 5.今後の展望 交雑率を比較した(Yonemura et al., 2010) .その結果,大規模ほ場を多く含む地域ほど平均交雑 率が低くなる傾向が明らかとなった.このような結果は,多数の小規模ほ場が複雑に入り組んだ地 域ほどほ場間の接線長が長くなり,接線の近傍領域ほど交雑が発生しやすいことと関係していると 推察される.言い換えると,ある地域に遺伝子組換えイネを導入する際には,それらを作付けする ほ場を集団化することで,地域内で発生する平均交雑率を低減できると考えられる(図 16). ጤᚻ⎇㔑⍹↸ ⨙ၔ⋵䈧䈒䈳䉂䉌䈇Ꮢ Ȫȩȩ ȫȩȩ ȩ ɌɨɮɭɡȵȦȦȦȦȦȷɇɨɫɭɡ ɌɨɮɭɡȵȦȦȦȦȦȷɇɨɫɭɡ ɌɨɮɭɡȵȦȦȦȦȦȷșɇɨɫɭɡ ⑺↰⋵ᄢẟ ȩ ȩ Ȫȩȩ ȫȩȩ Ȫȩȩ ȫȩȩ Ȭȩȩ ȩ Ȫȩȩ ȫȩȩ Ȭȩȩ ȭȩȩ ȩ ɐɞɬɭȵȦȦȦȷȾɚɬɭ Ȫȩȩ ȫȩȩ Ȭȩȩ ɐɞɬɭȵȦȦȦȦȦȷȾɚɬɭ ȩ Ȫȩȩ ȫȩȩ Ȭȩȩ ɐɞɬɭȵȦȦȦȦȦȷȾɚɬɭ ȩȧȩȪ ȩȧȩȫ ȩȧȩȬ ȩȧȩȭ ȩȧȩȮ ȩȧȩȪ ȩȧȩȫ ȩȧȩȬ ȩȧȩȭ ȩȧȩȮ ȩȧȩȪ ȩȧȩȫ ȩȧȩȬ ȩȧȩȭ ȩȧȩȮ ȼɫɨɬɬȦɉɨɥɥɢɧɚɭɢɨɧșɋɚɭɞșȡȞȢ ȼɫɨɬɬȦɉɨɥɥɢɧɚɭɢɨɧșɋɚɭɞșȡȞȢ ȼɫɨɬɬȦɉɨɥɥɢɧɚɭɢɨɧșɋɚɭɞșȡȞȢ ᐔဋ㔀₸ 䋰䋮䋰䋲䋱䋦 ᐔဋ㔀₸ 䋰䋮䋰䋱䋷䋦 ᐔဋ㔀₸ 䋰䋮䋰䋱䋰䋦 図 16 3 地域のほ場配置と平均交雑率 5.今後の展望 生物多様性評価に関しては,今後,新たに開発される不良環境耐性遺伝子組換え作物等につい て,必要となる評価項目を従来の評価項目との比較において整理するとともに,栽培実験並びに各 種シミュレーションを実施して,このような遺伝子組換え作物の導入された場合の評価手法を研究 開発する必要がある.これらの研究成果に基づき管理手法の開発に必要な基本情報を蓄積 ・ 整理す る必要もあると考えられる.また,遺伝子組換え作物の自生化(volunteer),野生化(feral)によ る生態系へのリスク,および近縁野生種,野生化作物を介した導入遺伝子の拡散による間接的な生 物多様性への影響を評価する必要があろう. 共存研究については,4. で述べた研究成果を総合することで,食品や飼料としての安全性の確保 された遺伝子組換え作物を商業栽培する場合に,事前に一般栽培作物との交雑や混入を低減する対 策を取るだけでなく,交雑や混入が定められた水準以下に維持されていることを検証する順応的な 管理体制を確立することができると考えられる.また,本研究で得られた成果の一部は大気中の微 粒子のモニタリング(花粉モニター)や農耕地における外来生物の侵入リスクあるいは農薬の流出 リスクの評価・予測の基礎データ(ほ場環境 GIS データベース)に,さらに輸入穀物中の混入率 の推定(サンプル数決定式)など多岐にわたる分野での利活用が期待される.今後は収穫物の流通 段階における混入実態の把握をはじめとして,花粉の拡散を皆無なものとする生物的封じ込め,海 外から輸入される農産物に混入する未承認組換え作物の検知,海外で制定されている遺伝子組換え 作物の商業栽培に関する法的枠組みなど,関連の研究分野との連携を図りつつ,遺伝子組換え作物 と一般栽培作物との共存を図るための行動指針をより確固なものとする取り組みが望まれる. 第 9 章 遺伝子組換え作物の生態系影響評価と共存のための管理手法の開発 135 引用文献 1)Abe, J., Hasegawa, A., Fukushi, H., Mikami, T., Ohara, M. and Shimamoto, Y.(1999) Introgression between wild and cultivated soybeans of Japan revealed by RFLP analysis for chloroplast DNAs. Economic Botany, 53, 285-291 2)Aono, M., Wakiyama, S., Nagatsu, M., Nakajima, N., Tamaoki, M., Kubo, A. and Saji, H. (2006)Detection of feral transgenic oilseed rape with multiple-herbicide resistance in Japan. Environ. Biosafety Res., 5, 77-87 3)Arent, D. K. and Caviness, C. E.(1994)Natural cross-pollination of twelve soybean cultivars in Arkansas. Crop Sci., 34, 376-378 4)Caviness, C. E.(1966)Estimates of natural cross pollination in Jackson soybeans in Arkansas. Crop Sci., 6, 211-212 5)Chiang, Y. C. and Kiang, Y. T.(1987)Genometric position of genotypes, honeybee foraging patterns and outcrossing in soybean. Botanical Bulletin of Academia Sinica, 28, 1-11 6)Cutler, G. H.(1934)A simple method for making soybean hybrids. J. Am. Soc. Agron., 26, 252-254 7)Dorokhov, D.(2004)Biosafety of the herbicide resistant GM soybean 40-3-2, Experience in the Russian Federation. http://www.bsbanet.org/article/ps1_dorokhov_en.htm 8)Erickson, E. H.(1975)Variability of floral characteristics influence honeybee visitation to soybean flowers. Crop Sci., 15, 767-771 9)Free, J. B.(1970)Insect Pollination of Crops. Academic Press. London and NY 10)Fujita, R., Ohara, M., Okazaki, O. and Shimamoto, Y.(1997)The extent of natural crosspollination in wild soybean(Glycine soja) . J. Heredity, 88, 124-128 11)古谷義人(1980)ダイズ 農学大事典(第 2 版),養賢堂,東京,551-557 12)James, C.(2007)Global Status of Commercialized Biotech/GM Crops, 2007, ISAAA Brief No.37. ISAAA Ithaca NY. 13)Karasawa, K.(1936)Crossing experiments with Glycine soja and G. ussuriensis. Japan J. Bot., 8, 113-118 14)Kwon, S. H., Im, K. H. and Kim, J. R.(1972)Studies on diversity of seed weight in the Korean soybean. Korean J. Breed., 4, 70-74(in Korean) 15)Mizuguti, A., Ohigashi, K., Yoshimura, Y., Kaga, A., Kuroda, Y. and Matsuo, K.(2010) Hybridization between GM soybean(Glycine max(L.)Merr)and wild soybean(G. soja Sieb. Et Zucc.)under field conditions in Japan. Environ. Biosafety Res., 9, 1-11 16)Mizuguti, A., Yoshimura, Y. and Matsuo, K.(2009)Flowering phenologies and natural hybridization of genetically modified and wild soybeans under field conditions. Weed Biology and Management, 9, 93-96 17)Nakayama, Y. and Yamaguchi, H.(2002)Natural hybridization in wild soybean(Glycine max. ssp. soja)by pollen flow from cultivated soybean(G. max ssp. max)in a designed population Weed Biology and Management, 2, 25-30 18)大橋広好(1997)朝日百科植物の世界 4, 260-261 19)Oka, H.(1983)Genetic control of regenerating success in semi-natural conditions observed 136 引用文献 among lines derived from a cultivated×wild soybean hybrid. J. Appl. Ecol., 20, 937-949 20)Ray, J. D., Kilen, T. C., Abel, C. A. and Paris, R. L.(2003)Soybean natural cross-pollination rates under field conditions. Environ. Biosafety Res., 2, 133-138 21)Saji, H., Nakajima, N., Aono, M., Tamaoki, M., Kubo, A., Wakiyama, S., Hatase, Y., Nagatsu, M.(2005)Monitoring the escape of transgenic oilseed rape around Japanese ports and roadsides. Environ. Biosafety Res., 4, 217-222 22)立川雅司,畠山華子,菅野真美(2009)EU における遺伝子組換え作物の共存政策を巡る重層 的ガバナンスとその帰結.2009 年度日本農業経済学会報告論文集,402-408 23)牛山朋来 , 井上 聡 , 芝池博幸(2009)数値シミュレーションへの適用を目的とした風洞実験 と数値実験による防風ネットの防風特性調査.農業気象 , 65(3),273-281 24)Ushiyama, T., Du, M., Inoue, S., Shibaike, H., Yonemura, S., Kawashima, S. and Amano, K.(2009) Three-dimensional prediction of maize pollen dispersal and cross-pollination, and the effects of windbreaks. Environ. Biosafety Res., 8, 183-202 25)Weber, C. R. and Hanson, W. D.(1961)Natural hybridization with and without ionizing radiation in soybeans. Crop Sci., 1, 389-392 26)Woodworth, C. M.(1922)The extent of natural crosspollination in soybeans. J. Am. Soc. Agron., 14, 278-283 27)Yamamura, K.(2004)Dispersal distance of corn pollen under fluctuating diffusion coefficient. Popul. Ecol., 46, 87–101 28)Yonemura, S., Shibaike, H., Iwasaki, N., Kawashima, S., Ushiyama, T., and Du, M. A system model for estimating regional cross-pollination rates between GM and non-GM rice considering actual meteorological conditions and field distributions J. Agric. Meteorol., 印刷中 29)Yoshimura, Y., Beckie, H.J., Matsuo, K.(2006)Transgenic oilseed rape along transportation routes and port of Vancouver in western Canada. Environ. Biosafety Res., 5, 67-75 30)Yoshimura, Y., Matsuo, K. and Yasuda, K.(2006)Gene flow from GM glyphosate-tolerant to conventional soybeans under field conditions in Japan. Environ. Biosafety Res., 5, 169-173 31)農林水産省技術会議事務局技術安全課プレスリリース 2007 年 9 月 21 日 輸入港周辺における セイヨウナタネ個体群の調査結果(http://www.s.affrc.go.jp/docs/press/2007/0921.htm) 137 第 10 章 環境生物の情報化学物質と機能利用 1.はじめに 農業生産の対象となる植物は,もともと自然生態系の中で他の植物と養分や日光を奪い合う一方 で,微生物から昆虫,大型動物にいたる地球上のあらゆる生物の栄養源として絶えずその生存を脅 かされる存在であった.そのため,農業活動は,こうした農業対象植物の生存を脅かす因子からそ の植物を守り,その生育を促進する活動であるともいえる.人類は,作物の栄養となる肥料を工夫 し作物の品種改良を図る一方で,作物を害する微生物や昆虫を抑える化学農薬や,雑草の生育を抑 制する除草剤を開発して,農業生産を飛躍的に増大させてきた.しかし,1980 年代ころから,農 薬や化学肥料等の化学資材の過剰な使用による環境汚染が問題になるとともに,生物多様性や人 類の健康に及ぼす影響が懸念されるようになり,化学資材をふんだんに使う資材投入型の農業生 産から,環境に優しい低投入・持続的農業生産への移行が叫ばれるようになった.こうした傾向 は 1990 年代になるとさらに強まり,わが国でも,1992 年に農林水産省が公表した「新しい食料・ 農業・農村政策の方向」で,生産性向上を図りつつ環境への負荷軽減に配慮した持続的な農業,す なわち環境保全型農業が,今後の農業政策の重要課題として位置づけられた.これを受け,農林水 産省は,1994 年に全国環境保全型農業推進会議を設置し,1997 年には「環境保全型農業推進憲章」 を制定して,環境保全型農業運動の全国的展開を推進した.さらに,2000 年 7 月に「持続性の高 い農業生産方式の導入の促進に関する法律」が制定されると,この法律に基づき,持続性の高い農 業生産方式を導入している農家を,農業改良資金(環境保全型農業導入資金)の特例措置を受けら れるエコファーマーに認定する制度が導入されるなど,制度面でも環境保全型農業の推進がはから れている. こうした背景のもと,病害虫を適切に防除し農作物被害を防止するため,様々な防除手段を適切 に組み合わせることにより,環境負荷を低減しつつ病害虫・雑草の発生を経済的被害以下に低く管 理しようという総合的病害虫管理(Integrated Pest Management: IPM)が国際的に提唱され,わ が国においてもこれに向けた取組みが行われてきた.こうした IPM 推進のため,人工の化学物質 に頼らず,もともと作物が自然環境下で生存していく上で有利に働いていた生物機能や特定の生 物が持つ有用機能を,農薬や肥料の代替物として活用し減化学農薬・減化学肥料のために役立てた り,有害化学物質や資材による環境汚染を浄化したりするための研究が,日本全国の農業試験研究 機関・大学などで盛んに進められてきた.農業生態系では,多様な生物が様々な生理活性物質を介 してお互いに影響を及ぼしたり , 環境中に存在する化学物質を分解したりしている.それらの物質 を明らかにし,その機能を解明することは,農業生態系の維持向上や循環型社会の実現に資する 様々な技術開発につなげる上で重要である.そのため,農業環境技術研究所では,そのような技術 開発につながる可能性が高い生理活性物質の機能解明を進め,新技術の開発につなげる研究が行わ れてきた.たとえば害虫防除では,殺虫剤による害虫防除の代わりに害虫を誘引する性フェロモン を利用した害虫防除技術の開発に向けた研究が進められた.また,雑草防除ではアレロパシー作用 と呼ばれる植物機能を利用し雑草の生育を阻害する物質の研究が行われた.さらに,微生物の有す る各種分解酵素を用いた環境汚染浄化技術の研究も行われてきている.これらの研究は,いずれ も,自然界で生きてゆくために生物が自然界に放出する化学物質を利用する点で共通している. フェロモンは,他個体に自らの存在を伝えるため,また,アレロパシー物質は,自らの生存領域を 138 2. 昆虫の性フェロモンの利用に関わる研究 他の生物に示し守るため,さらに,分解酵素は,基質の存在を認識した微生物が栄養として取り込 むため,自然界に放出される化学物質である.農業環境技術研究所では,いずれも自然界で個々の 生物の生存戦略に関わる情報化学物質という観点から,これら情報化学物質の有効利用に関わる研 究を行ってきている.本章では,農業環境技術研究所が独立行政法人に移行してからの 10 年間に スポットをあて,その間, 「農業環境技術研究成果情報(第 18 〜 27 集)」に収載された主要成果を もとに,情報化学物質に関わる研究成果を,1)昆虫性フェロモンの利用に関わる研究,2)植物の アレロケミカルの利用に関わる研究,3)微生物の有用機能の利用に関わる研究に類別し,項目ご とに紹介する. 2.昆虫の性フェロモンの利用に関わる研究 生物が環境中に放出する化学物質の中でも,昆虫や動物が体外に分泌放出し,同じ種の別の個体 を誘引するといった特定の行動や生理反応を引き起こさせるフェロモン(pheromone)は,特に生 物が生産する情報化学物質として広く知られている.中でも,交尾行動や配偶行動に関与する性 フェロモンについては,古くは「ファーブルの昆虫記」にもその効果が記載されており,1956 年 にカイコガのメスからオスを興奮させる化学物質として,初めてその分子構造が明らかにされて以 来,今日まで数多くの昆虫の性フェロモンが構造決定されている. この性フェロモンは,きわめて微量で効果を発 揮し,対象となる害虫だけに効果があり,生態系 に存在する他の生物に対する影響も少なく毒性も ほとんどないことから,農業現場で早くから注目 され,害虫駆除の目的でその利用がはかられてき た.一つは,強い誘引力を有する性フェロモンを 塗布した粘着性の台に害虫を捕殺するフェロモン トラップ(図 1)を用いる方法で,メスのフェロ モンに誘引されるオスを捕殺することで,次世代 の害虫の密度を低下させることができる.また, フェロモントラップは原理的に対象害虫だけを捕 獲できることから,害虫の発生調査に広く使われ ており,この発生予察に基づいてその発生状況に 図 1 フェロモントラップ かくらん 応じた害虫対策が講じられている.その他の性フェロモンの利用法としては,交信撹乱がある.こ れは,性フェロモン剤を空気中に拡散させ,オスメス間の性フェロモンによる交信を文字通り撹乱 し,交尾率を下げることによってやはり次世代の害虫密度を低下させる方法である.これらフェロ モンを用いた,大量捕殺,発生予察,交信撹乱が,農業現場における性フェロモンの主な応用技術 である(野口,1999;フェロモン剤利用ガイド,2000). 農業環境技術研究所では,独立行政法人化前から精力的に性フェロモン研究が行われており,こ れまで様々な害虫から性フェロモンを抽出し構造決定を行う一方で,その利用法の開発と性フェロ モンの誘引特異性の解明を行うことで,わが国の害虫防除に多大の貢献をしている.以下,この 10 年間に得られた成果について紹介する. ⑴ 性フェロモン物質の探索とその利用 ハイマダラノメイガは,アブラナ科作物を加害し,コナガとともに東南アジアで重要な害虫と なっている.日本でも関西を中心にキャベツなどで被害が報告されていたが,近年関東でも被害が 第 10 章 環境生物の情報化学物質と機能利用 139 見られ大きな問題となっている.この害虫に対して性フェロモンを用いた防除法を適用しようと する際に,すでに報告されているハイマダラノメイガの性フェロモン,(E, E)−11,13−ヘキサデカ ジエナールでは誘引力が弱く,その誘引力の向上が求められていた.杉江は,性フェロモンの類縁 物質にこのフェロモンの誘引活性を強める活性があることを突きとめた(平成 14 年度研究成果情 報,杉江) (Sugie et al., 2003) .性フェロモン類縁物質を主成分と混合した数種の試料をろ紙に含 浸して粘着式の屋根型トラップ内に設置し,その活性を野外で検討した結果, (Z)−11−ヘキサデ カナールを混合した際に誘引活性が強くなることを見いだした.その混合物をフェロモントラップ の誘引源として利用を検討した結果,2 成分混合物の誘引力は処女メスに匹敵し,種特異的な誘引 源として用いることが可能であることが明らかとなった. 杉江らは,ナシの花芽や幼果を加害する重要害虫であるナシマダラメイガの発生予察を効率よく 行うための誘引剤の開発を目的として,ナシマダラメイガのメス成虫から性フェロモンを抽出し, その化学構造を明らかにした(平成 20 年度研究成果情報,杉江ら)(Tabata et al., 2009).ナシマ ダラメイガは,1 頭の幼虫が複数の果実を加害するため,個体数が少なくても大きな経済的被害を 与える害虫である.このガのメスから抽出した性フェロモン画分をガスクロマトグラフ ― 質量分 析計によって分析した結果,誘引活性を示す画分に(Z)−9−ペンタデセニルアセテート(Z9−15: OAc)とペンタデシルアセテート(15: OAc)が含まれていることがわかった(図 2).これらの成 分が,実際に野外のオス成虫に対する誘引性をもっているかどうか確認するため,化学的に合成し たそれぞれの成分を誘引源とするトラップを用いた野外調査を実施した結果,Z9−15: OAc に 15: OAc を 7%加えた混合物により,ナシマダラメイガのオス成虫を効率よく誘引できることが明らか となった.オス成虫を誘引するメスの性フェロモン成分解析から明らかとなった 2 成分の混合物を 誘引剤として利用することで,この害虫の発生予察を行うことが可能となった.なお,この成果に ついては,特許を出願している(特願 2008-205253). O O O O (Z)-9-䊕䊮䉺䊂䉶䊆䊦䉝䉶䊁䊷䊃 (Z9-15:OAc) 䊕䊮䉺䊂䉲䊦䉝䉶䊁䊷䊃 䋨15:OAc) 図 2 ナシマダラメイガの性フェロモン成分の化学構造 ⑵ 性フェロモンを利用した交信撹乱抵抗性出現の機構解明 性フェロモンによる交信撹乱には,比較的抵抗性が生じにくいとされているが,性フェロモン剤 も化学農薬と同様に,同じ剤を長く利用していると効果がおちることがある.その原因について は,メスの性フェロモンの変化,オスの感覚器官の麻痺からの回避,さらには種の変化など多くの 仮説が提唱されているが,環境に影響の少ない害虫防除法として重要な性フェロモン剤の普及を図 る上でこの原因の解明は重要である. 野口らは,交信撹乱剤処理により抵抗性が生じたチャノコカクモンハマキ抵抗性系統を確立し, その抵抗性が生じた原因を明らかにした(平成 15 年度研究成果情報,野口ら).チャノコカクモ ンハマキの性フェロモンは 4 成分からなる.そのうちの 1 成分だけを使った交信撹乱剤(ハマキコ ン L,成分:Z11−テトラデセニルアセテート)は 1983 年に農薬登録されたが,連年使用されて いた一部地域で 1995 年ころからその効果が低下し,世界で最初の撹乱剤に対する抵抗性現象とし て 1996 年に報告された.本研究では交信撹乱剤に対する抵抗性発現の要因の解明をめざし,抵抗 140 3. 植物のアレロケミカルの利用に関わる研究 性個体群の発生地から約 6,000 頭のチャノコカクモンハマキを採集,これを用いて交信撹乱剤処理 下で採卵・飼育を繰り返すことで交信撹乱処理を行っても約半数が交尾する交信撹乱抵抗性系統を 世界で初めて確立した.この抵抗性系統と交信撹乱剤感受性系統のメスとオスの交尾行動を調査し 比較した結果,抵抗性はオスに生じており,両系統間でオスの嗅覚能力に違いが生じていること, また,ハマキコン L 抵抗性系統でも 4 成分性フェロモン組成を持った剤を使用することで交信撹 乱をおこし,交尾は完全に阻害され抵抗性を示さなくなることを突きとめた.さらに,杉江らは, チャノコカクモンハマキの交信撹乱剤抵抗性の原因が,このハマキガのオスが主要な性フェロモン 成分である Z11−テトラデセニルアセテートに対する誘引性を失う一方で,他の性フェロモン成分 に誘引されるため,撹乱剤の影響を受けにくくなっていることを突きとめた(平成 18 年度研究成 果情報,杉江ら) (Tabata et al., 2007) . 一方,野口らは,上記チャノコカクモンハマキの性フェロモンについて,性フェロモンを構成す る 4 成分の比に地理的変異があり,地域によってフェロモン応答に差が出る可能性が示された(平 成 15 年度研究成果情報,野口ら) .性フェロモンは種特異性が高く,その 4 成分の比は種ごとに一 定と考えられていたが,日本各地で採集したチャノコカクモンハマキの性フェロモン組成を調べた ところ,関東,九州地方では主成分である Z11−テトラデセニルアセテートの比率が低く,東海, 近畿地方では高い傾向にあるなど地理的変異があることが明らかとなった.さらに同一個体群内で は性フェロモンの成分比は安定しており,各地のオスは,それぞれ該当する地域のメスが放出する 性フェロモンによく反応し,その変異に対応していることがわかった.以上の成果は,性フェロモ ンの誘引性に地域的な違いが生じる可能性を示しており,地域間での誘殺数の比較には注意が必要 であることが判明した. 3.植物のアレロケミカルの利用に関わる研究 アレロパシーとは「他感作用」ともよばれ,ある植物が放出する情報化学物質(アレロケミカ ル)が他の植物の生長を抑えたり,あるいは動物や微生物の接近を防いだり,あるいは引き寄せた りする効果を及ぼす作用のことで,植物のニッチ拡大に関与している.ギリシャ語の allelon(互 いに)と pathos(一方が他に障害を与える)を合成した合成語である.1937 年にドイツの植物学 者であるハンス・モーリッシュにより提唱された.これまでアレロパシーは植物遷移や連作障害の 要因として研究が進められてきたが,近年,雑草・病害虫防除への利用が注目され,他の植物の生 育を阻害するアレロパシー作用を示す植物の選抜とそのアレロパシー効果およびその元となる化学 物質の同定研究が盛んに進められてきた.現在では,植物生育阻害物質は,環境中で速やかに代 謝・失活されやすいことから,合成除草剤の代替資材として期待されている.農業環境技術研究所 では,1983 年に,アレロパシーの農業利用の研究を行う他感物質研究室が設置されて以来,長年 にわたり研究が行われてきた.その間,アレロパシー現象を評価する新たな生物検定法を複数開発 して,約 4,000 種の植物を検定し,ムクナ,ヘアリーベッチなど,強いアレロパシー作用を持つ植 物をスクリーニングするとともに,農業現場でもこれら植物に雑草抑制作用があることを明らかに した.また,これらの植物が放出するアレロケミカルを分析・同定し,9 種の新規生理活性物質を 発見,特許を取得している.以下に,この 10 年間に得られた成果を紹介する. ⑴ アレロケミカル活性測定法の開発 しん しゅつ これまで農業環境技術研究所では,アレロパシーの 3 つの作用経路(根からの滲出,茎葉や残 さからの溶脱,揮発性物質による揮散)に特異的な検定法として,それぞれプラントボックス法 (平成 3 年度研究成果情報) ,サンドイッチ法(平成 9 年度研究成果情報),揮発性物質検定法(平 第 10 章 環境生物の情報化学物質と機能利用 141 成 11 年度研究成果情報)を独 3ml 䉝䉥䉮Ớ❗ᶧ 自に開発してきた.しかし,植 40ml 物の根から土壌に放出された 3ml CT ၭ 物質が土壌中で示す活性を評 価する手法は,まだ開発されて い な か っ た. そ こで,藤井ら + ㆙ᔃಽ㔌 3000rpm, 30min ኻᾖ䈱⚕ ⹜ᢱ䋨䊥䊷䊐䊂䉞䉴䉪䋩 30 / 25 ͠ ( /ᥧ) 15 ᣣ㑆 ⮣⮺䋨䉝䉥䉮䋩 䈱䈦䈢 ኙᄤᐔ᧼ は,他感物質の濃度が最も高い 根圏土壌を材料とし,土壌中に おけるアレロケミカルの活性を 䋷ᣣ㑆ၭ㙃 評価するための検定法(根圏土 壌法)を開発した(平成 15 年 度研究成果情報,藤井義晴ら) (Parvez et al., 2003; Iqbal et al., ↢⢒㒖ᱛ ↢⢒㒖ᱛ䈱⋥ᓘ䈪ᵴᕈ䉕䈅䉌䉒䈜 䍲䍭䍑䍷䍚 Rhus trichocarpa 図 3 リーフディスク法 2004-1) .本法は,根から放出 されたアレロケミカルの,土壌中での活性を検定する方法で,具体的には,根圏土壌を含む寒天上 に,検定層としてさらに寒天を重層しレタス等の検定植物を置床する.暗黒下,恒温装置中で 3 日 間培養後,幼根長と下胚軸長を測定することにより,土壌中に含まれているアレロケミカルの影響 を容易に検出できる.本法を用いていろいろな植物の活性を測定した結果,アレロパシー作用の報 告があるヘアリーベッチや,大型侵入植物であるクワモドキの根圏土壌から,強い阻害作用を検出 することができた. また,藤井らは,アオコを構成するラン藻類を用いて,アオコの増殖を阻害する作用を生物検定 するための手法を開発した(平成 17 年度研究成果情報,藤井義晴ら)(Tsuda et al., 2005).ため ろ か 池や湖で異常増殖したラン藻類はアオコを形成し,異臭の発生や浄水処理時の濾過障害,有毒物質 ミクロキスチンの生成という問題を引き起こしている.一方,樹木の落ち葉によるアオコの発生抑 制が観察されており,ため池周辺に生育する植物の葉に含まれる成分によるラン藻類の増殖阻害作 用を調べるため,ディスクパンチで落ち葉を打ち抜いてリーフディスクを作成し,これを一定濃度 のアオコを含む培地上にのせ,アオコの生育阻止円の直径を測定することにより,葉の有するアオ コ増殖阻害作用を調べる手法を開発した.本法( 「リーフディスク法」)は,落ち葉をそのままリー フディスクとして用いるもので,検体から溶媒で抽出した物質の活性を吸光度で測定する既存の手 法より簡便であり,かつ,現場での活性を反映していると考えられる(図 3).この「リーフディ スク法」を用いて,ため池周辺に生育する 50 種の樹木の落ち葉を検定した結果,アカメガシワ, ユキヤナギの落ち葉が,ラン藻類に対する強い増殖阻害作用を示すことが明らかとなった. ⑵ アレロケミカルの同定 農業環境技術研究所で独自に開発されたアレロケミカル活性測定法を駆使し,これまで様々な植 物から種々のアレロケミカルが同定されている.藤井らは,休耕水田や遊休農地の植生・雑草管理 に導入が推奨されているヘアリーベッチが示す植物生育阻害作用の本体がシアナミドであることを 明らかにした(平成 13 年度研究成果情報,藤井義晴ら).ヘアリーベッチには,雑草等の生育を阻 害する成分が存在することが以前から報告されていたが,作用成分は不明であった.本研究では, ヘアリーベッチの茎葉部より得た粗抽出液から,レタス伸長阻害活性を指標とし,各種のクロマト グラフィーにより植物生育阻害物質を単離し,その化学構造を解析した結果,その活性本体がシア ナミド(cyanamide)であると同定した.シアナミドは,窒素肥料である石灰窒素の成分として殺 142 3. 植物のアレロケミカルの利用に関わる研究 虫・殺菌・除草効果のあることが知られているが,生物の体内成分としては新発見である.本成果 は,これまでに知られているヘアリーベッチの雑草抑制作用や耐虫・耐病性を,シアナミド合成と 関連して解明する大きな手がかりとなった.この研究に関連して,平舘らは,ガスクロマトグラフ 質量分析計(GC/MS)を用いたシアナミドの高感度定量法も開発している(平成 17 年度研究成果 情報,平舘ら) (Hiradate et al., 2005-1) .1994 年にヘアリーベッチによる雑草防除制御技術を提唱 して以来(平成 6 年度研究成果情報,藤井義晴ら),こうした一連のヘアリーベッチに関わる業績 により藤井は,平成 16 年度の日本土壌肥料学会賞を受賞している. その他,藤井らは,日本在来の被覆植物リュウノヒゲからアレロケミカルとしてサリチル酸を同 定している(平成 16 年度研究成果情報,藤井義晴ら)(Iqbal et al., 2004-1; Iqbal et al., 2004-2). 本研究では,まず,サンドイッチ法とプラントボックス法により日本在来の被覆植物リュウノヒゲ が強い他感作用を示す被覆植物として選抜された.植物生育阻害物質として,リュウノヒゲからβ ― シトステロール,p ― ヒドロキシ安息香酸,サリチル酸が同定され,含有量と比活性から,サリ チル酸の寄与が大きいことが明らかにされた. また,平舘らは,ユキヤナギに含まれる植物生育阻害物質を分離・精製して,その化学構造を明 らかにした結果,植物生育阻害活性の本体がシス−ケイ皮酸グルコシドであることを明らかにした (平成 16 年度研究成果情報,平舘ら) (Hiradate et al., 2004; Hiradate et al., 2005-2).この物質の植 物生育阻害活性は,天然の植物生育阻害物質としては最も強い部類に属しており,天然由来の除草 剤のリード化合物として注目されている.なお,この成果については,特許登録がされている(特 許 4415106) . さらに,藤井らは,ソバが有する植物生長阻害物質の中で,特に,フェノール性物質の一種で あるルチンがアレロパシー活性の原因物質であることを突きとめた(平成 18 年度研究成果情報, 藤井義晴ら) .藤井らは,戦後に導入された外来種,ナギナタガヤ(Vulpia myuros C.C.Gmel)がヘ アリーベッチに匹敵する強い阻害活性を示すことを明らかにし,2 種のアレロケミカル,(−)− 3−ヒドロキシ−β−イオノン ((−) −3−hydroxy−β−ionone)と(+)−3−オキソ−α−イオ ノール ( (+) −3−oxo−α−ionol)を検出した(平成 21 年度研究成果情報,藤井義晴ら)(Kato Noguchi, et al., 2010) . 以上は,いずれも除草剤のリード化合物となる植物生育阻害活性を有するアレロケミカルの研究 を紹介したが,藤井らは,逆に植物の生育促進物質を発見している(平成 16 年度研究成果情報, 藤井義晴ら) (Nakajima et al., 2005) .循環型社会を実現するための一方策として,林業由来の間 伐材および刈り払いされた下草などの有効利用が注目されている.本研究では,これらを原料とし て得られる精油成分混合物の産業利用について評価・検討する一端として,本資材が作物の生育や 収量等に及ぼす影響を評価するとともにその作用成分を分析し,農業利用の可能性について検討 した.国内で生産される 35 種類の間伐材等から精油成分混合物を抽出し,レタス幼根伸長に及ぼ す影響を評価した結果,その中から植物の生育促進物質として,1, 2−プロパンジオールを同定し た.本成分は,精油成分混合物中に 0.4%含まれ,0.01ppm の低濃度でも,植物の生育を促進する ことが明らかとなった.この物質は,天然物としては,イチゴの香気成分の前駆体として報告され ているが,植物の生育促進活性の報告は新たな発見である. ⑶ アレロケミカルの作用解析 各種アレロケミカル活性測定法でスクリーニングした物質でも,環境中でその生理活性を発現で きなければ農業現場では利用できない.そのため,選抜したアレロケミカルについて,環境中での 活性の確認や様々な植物における感受性の違いを明らかにすることは,そのアレロケミカルの除 第 10 章 環境生物の情報化学物質と機能利用 143 草剤としての効果を検証する上で重要となる.藤井らは,マメ科のムクナが生産する強力な植物 生育阻害剤である L−ドーパ(L−3, 4−dihydroxyphenylalanine)が,ムクナの根から栽培環境中 に放出されることを高感度分析により確認し,周辺植物の生育阻害要因となることを検証した(平 成 16 年度研究成果情報,藤井義晴ら) (Fujii et al., 2003; Nishihara et al., 2004).また,様々な植 物種を供試して,L−ドーパの植物生育阻害活性に対する感受性を検定し,植物種間における感受 性の違いならびにその原因を解毒・耐性機構の面から解明した.さらに,ムクナの生育地周辺に おける L−DOPA の,他の植物に及ぼす影響を評価するために,平舘らは,L−DOPA が土壌環 境中で受ける化学反応およびその特性を解明するとともに,これらの反応にともなう L−DOPA の生理活性の変化を明らかにした(平成 17 年度研究成果情報,平舘ら)(Hiradate et al., 2005-3; Furubayashi et al., 2005) . 一方,藤井らは,遊休農地や法面などで雑草管理をするために有望な被服植物を,プラントボッ クス法を用いて評価したアレロパシー活性を指標に選抜 した結果,ソバ属植物の植物生育阻害活性が強いことが 見いだされた(平成 15 年度研究成果情報,藤井義晴ら) (Iqbal et al.,2002; Iqbal et al., 2003; Fujii et al., 2004). そこで,ソバの雑草抑制効果を現地栽培試験によって確 認した結果,雑草放任区ではおう盛な雑草の発生が認め られたのに対し,雑草発生前にソバを播種した区では, 雑草の発生は顕著に抑制されることを見いだし,ソバ のりめん は,休耕地や法面等における雑草防除と景観向上を目的 とした実用的な被覆植物となる可能性があることを示し た(図 4) .なお,前述したようにソバのアレロパシー 活性の原因物質は,フェノール性物質の一種であるルチ ンであった. 図 4 ソバの雑草抑制効果 ソバの生育している下には雑草がほ とんど生えない. 4.微生物の有用機能の利用に関わる研究 農耕地に生息する微生物の中には,肥料成分を作物と奪い合ったり病気を引き起こしたりするな ど,作物の生育を阻害するものが存在する一方で,根に共生して土壌中のリンや空気中の窒素を作 物に供給することで,作物の生育をよくするものがいるなど,微生物は農業生産に大きな影響を及 ぼしている.そのため,農耕地土壌の土壌微生物をコントロールし,その機能の有効利用を図るこ とはきわめて重要な課題といえよう.そのような課題達成のためには,土の中にどのような微生物 が生息し,どのような機能を発揮しているかを正確に把握することが必須となる.微生物は,自然 環境下で様々な環境シグナル物質を感知・認識し,自らの生育環境を有利な状態にするために種々 の分解酵素や抗生物質などの生理活性物質を放出することで環境の変化に応答している.農業環境 技術研究所では,農業環境における微生物や小動物のコミュニティーを解析する一方で,微生物の 認識と応答に関わる酵素やシグナル物質などの情報化学物質を明らかにし,その有効利用に関する 研究を長年行ってきた. ⑴ 土壌微生物相の解析 農耕地土壌の土壌微生物相を把握しコントロールするためには,まず,農耕地に生息する生物相 を正確に把握することが必須となる.その場合,線虫などの土壌小動物の場合には,顕微鏡等で比 較的観察しやすいため,目視による観察が一般的に行われている.荒城は,土壌線虫を指標生物と 144 4. 微生物の有用機能の利用に関わる研究 して,耕起という撹乱の影響を比較,評価した.不耕起栽培畑などから土壌線虫を分離して,属レ ベルで分類・計数して,多様度指数や Maturity Index を計算することで,不耕起栽培の方が慣行 栽培より土壌中の線虫多様性が増すことを明らかにした(平成 13 年度研究成果情報,荒城).この 成果は黒ボク土畑における耕起の影響の評価指標として利用できる.また,荒城らは,クロルピク リン,臭化メチルなどによる土壌くん蒸処理が,標的である病害虫以外の土壌小動物に及ぼす影響 を調査した(平成 16 年度研究成果情報,荒城ら).その結果,クロルピクリン,1, 3−ジクロロプ ロペン(D−D) ,あるいは臭化メチルによる土壌くん蒸処理をした場合,約 12 か月で土壌線虫の 密度は処理前の水準に回復するが,土壌線虫の群集構造が変化し,微生物食性で増殖率が高い線虫 が優占種になることが明らかになった.本成果は,土壌くん蒸剤処理などの薬剤処理が農耕地の非 標的生物へ及ぼす影響を評価するための基礎データとなる. 線虫のようにある程度の大きさを持ち,比較的その存在が把握しやすい場合には,上記のよう に,土壌中の生物を直接観察し解析することができるが,細菌やカビなどの糸状菌は,従来,寒天 培地等で培養してその存在を知ることしかできなかった.人間が培地で培養できるのは,自然界に 生息する微生物のせいぜい数%程度であるといわれており(Amann et al.,1995),培養法に頼っ て土壌微生物を解析していた時代には,土壌の微生物のコミュニティー全体を把握することは, 実質的に不可能であった.しかし,近年,土壌から直接 DNA を抽出する手法が開発され,土壌微 生物由来の遺伝子や DNA 配列から土壌の微生物相を解析しようとする研究が主流となってきてい る.このような,土壌環境から直接抽出した DNA は,土壌微生物コミュニティーを構成する個々 の微生物ゲノムの総和となることからメタゲノムと呼ばれている.このメタゲノムという言葉は, 「高次」という意味の“メタ”と“ゲノム”から作られた造語で,1998 年にウィスコンシン大学の Handelsman 教授らにより命名された(Handelsman et al.,1998).近年の大量シークエンシング 技術のめざましい進歩により,土壌のメタゲノム解析は,今後ますます盛んになることが予想され る.農業環境技術研究所では,それまで難しかった黒ボク土からの DNA 抽出法を開発して以来, 土壌をはじめ様々な環境サンプルから抽出した DNA や RNA を用いて,その土壌に生息する微生 物相や特定の機能に関わる遺伝子の消長をモニターする研究が行われてきている. 星野らは,農業環境技術研究所におけるメタゲノム解析の先駆けとなった研究で,これまで DNA を強く吸着するため土壌微生物 DNA の直接抽出が困難であった黒ボクから DNA を抽出す る際に,スキムミルクを添加することにより,土壌微生物群集の解析に使用可能な DNA を抽出す ることができることを示した(図 5) (平成 14 年度研究成果情報,星野ら)(星野,2005; Hoshino ㄘⅣ⎇ ㄘⅣ⎇ 㕍 ፉ (⇌) (ሽᨋ) (ᨐ᮸) (⇌) kb 23.1 9.4 6.7 M ፉ ᾢᧄ ᾢᧄ (⇌) 䋨᳓↰䋩 䋨⇌䋩 -+ -+ -+ -+ -+ -+ -+ 䉭䊉䊛 DNA 2.3 2.0 図 5 スキムミルク添加,無添加における日本各地の黒ボク土からの DNA 抽出 +:スキムミルク添加 , −:スキムミルク無添加 , M:サイズマーカー 第 10 章 環境生物の情報化学物質と機能利用 145 et al., 2004; Hoshino et al., 2005) .この研究成果により,日本の様々な土壌から DNA を抽出でき るようになり,日本の土壌メタゲノム解析は飛躍的に進展することとなった. この成果を受け,星野らは,土壌くん蒸処理の土壌微生物相に及ぼす影響を,土壌から直接抽出 した DNA を PCR−DGGE 解析により評価している(平成 16 年度研究成果情報,星野ら).臭化 メチルの廃止にともない,代替薬剤クロルピクリンや D−D の使用量の増大が予想されており,こ れらが非標的生物に及ぼす影響が懸念されている.そこで,ホウレンソウ栽培ほ場において年 1 回 上記薬剤でくん蒸処理を行い,土壌から直接抽出した DNA を用いて行うポリメラーゼチェーン反 応 ― 変性剤濃度勾配ゲル電気泳動(PCR−DGGE)解析によって,培養困難な微生物を含む微生 物群集の変動を追跡し土壌くん蒸剤の影響を評価したところ,これら処理が培養困難な微生物を含 む土壌微生物の群集構造に大きな変化をもたらすこと,また,ほ場におけるクロルピクリンの影響 は 1 年では完全に回復しないことを明らかにした. また,同様のアプローチとして,森本らは,培養法に頼らず土壌中のクロロ安息香酸分解菌群を 直接検出することをめざし,土壌から直接抽出した DNA を鋳型に分解遺伝子を PCR で増幅し, PCR−DGGE 法で解析することにより,培養法では見落とされていた土壌中のクロロ安息香酸分 解菌を効率的に探索できることを示した(平成 15 年度研究成果情報,森本ら)(Morimoto et al., 2005) .また本研究により,従来の液体集積培養などの培養手法では,土壌中の培養困難な微生 物の大半は見逃されていたことが明らかとなった.森本らは ,さらにこの方法を用いて,クロロ 安息香酸添加土壌において増加する DGGE バンドと同じ塩基配列を持つ菌を選抜することによっ て,土壌環境に適した分解菌を効率的に探索できることを示している(Morimoto et al., 2008). また,DGGE バンドとして得られた DNA 断片の塩基配列を用いてメタゲノムから取得する新手法 もあみ出している(Morimoto et al., 2009) こうした成果を受け,PCR−DGGE 法を用いたメタゲノム解析を農耕地土壌で大規模に行おう という農林水産省の委託プロジェクト「土壌微生物相の解明による土壌生物性の解析技術の開発」 が開始された.このプロジェクトは,通称「eDNA プロジェクト」と呼ばれ,2006 年から 2010 年 度までの 5 年間,国や地方の農業関連試験機関や大学の研究者が参加して行われた.本プロジェ クトでは,メタゲノムという言葉の代わりに,土壌から培養過程を経ず得た DNA という意味で eDNA(environmental DNA)という言葉を用いているが,基本的には同義である.このプロジェ クトは,農耕地における微生物多様性を調査する手法等を開発し,土壌生物相の機能と構造を土 壌から抽出した DNA に基づき解析するとともに,作物生産性と土壌生物相との関連性を解析する ことを目的としている.森本らは,このプロジェクトで PCR−DGGE 法を用いて農耕地土壌の微 生物相の構造を解析するための標準法を開発した(平成 19 年度研究成果情報,森本ら)(森本ら, 2008;大場ら,2008) .本研究では,農耕地土壌の生物相(細菌・糸状菌・線虫)を,DNA 分析 (PCR−DGGE)によって解析するための手順を,対象生物群ごとに最適化し,マニュアルを作成 した(図 6) . 「eDNA プロジェクト」では,この手法を用いて日本各地の土壌から抽出した DNA を PCR−DGGE 法を用いて解析し,その画像データと土壌の物理化学的性質や作付け/肥培管理 情報とともにデータベース化している.今後,本マニュアルに基づいて得られた,相互に比較可能 な土壌生物相情報が蓄積されることによって,農業生産活動にともなう様々な現象(作物の生育, 病害の発生や抑止,温室効果ガス発生等)と土壌生物の関連が,より明らかになっていくことが期 待される.この土壌生物相解析マニュアルは,現在,論文や Web 等で広く公開されている(森本 ら,2008;大場ら,2008) . 146 4. 微生物の有用機能の利用に関わる研究 ⚦⩶ ♻⁁⩶ ✢⯻ 図 6 土壌生物相解析マニュアルに従って土壌生物相を PCR−DGGE 法で解析した例 ⑵ 芳香族塩素化合物分解酵素の解析 ポリ塩化ビフェニル(PCB)等の POPs 類や有機溶剤等の難分解性芳香族塩素化合物による環境 汚染が問題となっており,これを微生物の能力により除去・防止する環境浄化(バイオレメディ エーション)技術の開発が期待されている.クロロカテコール類は,PCBs 類,クロロ安息香酸,2, 4−ジクロロフェノキシ酢酸(2, 4−D)等のクロロフェノキシ酢酸類,クロロベンゼン類等の様々 な芳香族塩素化合物の分解過程で生じる主要な中間代謝産物であり,これを分解するクロロカテ コール分解経路は,多くの芳香族塩素化合物の完全分解に共通である.農業環境技術研究所では, 微生物を用いたバイオレメディエーション技術の開発をめざし,クロロ安息香酸等の芳香族塩素化 合物分解菌を多数選抜し,長年分解遺伝子群発現の制御機構を解析してきた. 小川らは,土壌から分離した 2 種類のクロロ安息香酸分解菌を用いて,それぞれの分解菌が有す る分解遺伝子群の発現の制御機構を解析し,クロロ安息香酸ジオキシゲナーゼやクロロカテコール オルソ開裂経路の遺伝子群の発現が,クロロ安息香酸類やクロロカテコール類により活性化される LysR−type や AraC/XylS−type の転写調節因子によって制御されることを明らかにした(平成 13 年度研究成果情報,小川ら) (Francisco et al., 2001). また,小川らは,クロロカテコール 1, 2−ジオキシゲナーゼの基質特異性に関与する部位を明ら かにすることで,本酵素の基質特異性の改良を通してバイオレメディエーション技術の発展に資す るために,アミノ酸配列を変化させた変異型酵素を作成して比較解析を行い,基質特異性に関与す る部位を決定した(平成 16 年度研究成果情報,小川ら)(Ogawa et al., 2004).その結果,本酵素 のクロロカテコール類に対する基質特異性は,48, 52, 73 番目にあるアミノ酸によって決定される ことを明らかにした.本研究成果は,クロロカテコール 1, 2−ジオキシゲナーゼの基質特異性の改 良や分解能の向上を介した分解菌の機能改変に活用できる. 自然界から分離した芳香族塩素化合物分解遺伝子は,分解菌のゲノムではなくプラスミドと呼ば れる細胞中でゲノム DNA とは独立に複製,分離する小型 DNA にコードされていることが多い. このプラスミドの中には,細菌から細菌へ移るものがあり,将来自然環境下における分解微生物を 用いたバイオレメディエーションを行う場合に,土壌微生物相におけるプラスミドの動態を把握 しておく必要があるが,野外環境における分解遺伝子の多様性および遺伝子の動態については十 分に解明されていないのが現状である.酒井らは,日本国内 8 地点の水田土壌から分離された 2, 4 −D 分解菌を解析し,分離された 14 株のうち 8 株は,塩基配列が同じ分解遺伝子を,大きさが同 一のメガプラスミド上に持つことを明らかにし,2, 4−D 分解遺伝子が,異なる微生物間をプラス 第 10 章 環境生物の情報化学物質と機能利用 147 ミドを介して移動していることが示唆された(平成 17 年度研究成果情報,酒井ら)(Sakai et al., 2007) .本成果で得られた 2, 4−D 分解遺伝子を有するメガプラスミドの構造や機能を解析するこ とにより,土壌微生物における遺伝子の伝播を予測し,遺伝子組換え微生物の安全使用技術を開発 するための基礎的知見が得られる. また,自然環境下でバイオレメディエーションを行う場合には,分解遺伝子が実際に発現してい るのか確認したり,あるいはその発現条件を検討したりする必要がある,そのためには,自然環境 下でその分解遺伝子の発現を直接検出する必要がある.しかし,遺伝子発現を解析するために必要 な RNA を土壌から抽出することが難しいこともあり,これまで土壌環境下における微生物の遺伝 子発現を解析した例はきわめて少なかった.前述した「eDNA プロジェクト」では,土壌から高 純度の RNA を抽出する方法を開発している(Wang et al., 2008; Wang et al., 2009).王らは,この 抽出法を用いて,PCB 分解菌を滅菌土壌で生育させたモデル土壌から RNA を抽出し,PCB の分解 に必要となる遺伝子の発現を,定量 PCR 法で検出した(平成 19 年度研究成果情報,王ら)(Wang et al., 2008) .その結果,この遺伝子の発現を誘導する PCB 類縁物質,ビフェニルを土壌に添加し た場合に,PCB 分解遺伝子の発現が誘導されることを確認することができた.この研究成果は, 今後,肥料の溶脱や有害物質の浄化,大地からの温室効果ガスの放出など,土壌微生物が土壌中で その機能をどのように発揮するかを直接観察・解析するための大きな突破口を開くものである. ⑶ キチン分解酵素の利用研究 昆虫,甲殻類の外殻や作物病原糸状菌の細胞壁の構成成分であるキチンは,セルロースに次いで 多量に自然界に存在するバイオマスである.キチンを分解する酵素キチナーゼは,自然界の物質循 環で大きな役割を演じるとともに,農業的には作物病害抑制因子としてその利用が期待されてい る.農業環境技術研究所では,放線菌キチナーゼの有効利用技術の開発に資するため,多様なキチ ナーゼを生産する代表的な土壌細菌,放線菌に着目し,その酵素化学的解析や遺伝子発現制御機構 を明らかにする研究を行ってきた.以下に,この 10 年間に得られた成果を紹介する. 藤井らは,キトオリゴ糖放射能標識を導入する手法を開発し,放線菌のキチナーゼ生産を誘導す るキトオリゴ糖と特異的に相互作用する細胞表層の受容体(レセプター)を検出するアッセイ法を 開発した(平成 14 年度研究成果情報,藤井毅ら).このアッセイ法を用いることにより,キチナー ゼ生産の誘導物質であるキトビオースの細胞への取り込みは,キトビオースによって誘導されるこ と,また,このような誘導条件下では,細胞の膜画分中にキトビオースと結合する因子が蓄積して くることが明らかとなった.また,このアッセイ法を利用して,放線菌の膜画分からキトビオース 結合たんぱく質を精製することができた.さらに,藤井らは,誘導物質キトビオース添加後の放線 菌細胞内で起こるキチン分解関連遺伝子の遺伝子発現パターンの変化を解析し,遺伝子破棄等の技 術を駆使することにより,放線菌が環境中に存在するキチンを認識して,その情報を細胞内に伝 達し,キチナーゼ遺伝子群およびキトオリゴ糖の取り込みに関わるトランスポーターたんぱく質 群を誘導する一連の反応機構を明らかにした(平成 17 年度研究成果情報,藤井毅ら)(Fujii et al., 2005; Saito et al., 2006; Saito et al., 2007) . 以上は,土壌細菌におけるキチナーゼの発現制御に関わる研究を紹介したが,藤井らは,キチ ナーゼの遺伝子を導入した組換え根圏細菌を作出し,作物病害防除への利用を検討している(平 成 17 年度研究成果情報,藤井毅ら) (Ohno et al., 2011).作物病原糸状菌はその細胞壁にキチン質 を含んでおり,このキチン質を分解する酵素キチナーゼは,作物病害菌を溶菌させることにより作 物病害の抑止効果が期待される. そこで,キュウリ根圏に定着性が高い根圏細菌にキチナーゼ遺 伝子を導入した組換え根圏細菌を作出し,この組換え根圏細菌の培養液に発芽したキュウリ種子を 148 4. 微生物の有用機能の利用に関わる研究 浸した後,キュウリ苗立枯病汚染土壌に播種したところ,キュウリ苗立枯病の抑制効果が認められ た.また,この組換え根圏細菌の接種は根圏の土壌微生物の群集構造に影響を与えなかった.た だ,組換え生物の農耕地での利用については,まず,その安全性についてパブリックアクセプタン スを得ることが課題となっており,本研究もすべて閉鎖系温室内で実施された. ⑷ 生分解性プラスチック分解菌のスクリーニングと分解酵素の解析 農業用プラスチックフィルムの廃棄量と農作業軽減のために,現在,農業用資材などに生分解性 プラスチック(生プラ)の導入が勧められている.しかし,生産現場では使用後の分解が遅いなど 問題があり,生プラ資材の分解を促進する技術の開発が農業現場から求められている.そこで,北 本らは,生プラの構造と葉の表面構造が似ていることに着目して,独自に開発した効率よい生プラ 分解菌の選抜法を用いて,各種の葉の表面から生プラを分解する酵母を多数選抜した(平成 19 年 度研究成果情報,北本ら) .中でも Pseudozyma 属酵母の生プラ分解酵素は,各種生プラと,植物由 来のポリ乳酸を分解した(図 7) .本研究では特許を出願している(特願 2008−023030 号).さら に,小板橋らは,イネ科作物の葉の表面から強力な生プラ分解能を持つカビを分離し,このカビ がこれまで見つかった生プラ分解微生物の中でも特に高い分機能を有し,培養液中に高純度で,生 プラ分解酵素のみを生産する優れた特性を持っていることを突きとめた(平成 20 年度研究成果情 報,小板橋ら) .本研究では,特許を出願している(特願 2008−250869 号).以上二つの成果で得 られた分解菌と分解酵素は,生プラ製農業資材の分解促進技術の開発に有望であり,現在,農業現 場での生プラ資材分解技術の開発に使われている.なお,この一連の成果は,平成 20 年度農林水 産研究成果 10 大トピックスに選定された. 図 7 イネから分離した酵母による各種プラスチック膜の分解 PBSA: ポリブチルサクシネートアジペート,PBS: ポリブチルサクシネート ⑸ バイオエタノール生産のための固体発酵法の開発 石油資源の枯渇が心配される中,セルロース系バイオマスを原料としたエタノール生産技術の開 発が求められているが,その実用化のためには,原料の貯蔵・糖化・蒸留コスト・廃液処理といっ た克服すべき課題が多くある.北本らは,飼料作物を収穫後,サイレージ発酵している間に酵素 (セルラーゼ+グルコアミラーゼ)と微生物(酵母と乳酸菌)を添加し,糖化とエタノール発酵を 同時に行う,いままでにないバイオエタノール「固体発酵法」を開発した(平成 21 年度研究成果 情報,北本ら) .この方法では,通常バイオマスからエタノールを生産する際に問題となる有害な 廃液や廃棄物が出ないため,この方法で生み出されたバイオエタノールで農機具を動かし,発酵残 さを家畜の飼料として利用する一方で,家畜排せつ物を肥料として農地に還元すれば,地域内で消 費する化石エネルギー消費量を減らし資源を循環利用できる.この過程で,北本らは,様々なバイ オマスについて,バイオエタノール生産原料としての評価,原料の糖化・エタノール発酵に最適な 第 10 章 環境生物の情報化学物質と機能利用 149 酵素の種類と発酵微生物の選定,および実証規模の発酵条件の検討を簡便かつ効率的に行うことが できる方法を開発した(平成 21 年度研究成果情報,北本ら).なお,本研究では特許を出願してい る(特開 2009−213440) ⑹ 植物病害防除因子に関する研究 生物の相互作用には,ある種の情報化学物質が原因となって複雑な連鎖系で構成され,植物,微 生物,昆虫など複数の生物種間を越えて発現するものがある.たとえば,植物の葉面に常在する微 生物の中には,抗微生物活性物質を生産・分泌して他の微生物を抑制するものがおり,植物の病 害発症を抑え,植物の生育に好影響をもたらすことが知られている.平舘らは,葉面バクテリア Bacillus amyloliquefaciens が,抗微生物活性物質を分泌することにより他の葉面微生物の生育を抑 制していることを突きとめ,抗微生物活性物質として 7 種類の化合物を単離した(平成 14 年度研 究成果情報,平舘ら) (Hiradate et al., 2002; Yoshida et al., 2001; Yoshida et al., 2002).これら物 質の化学構造を解析した結果,いずれも Iturin と呼ばれる環状ペプチド化合物であることを特定 した.この Iturin は,各種植物病原微生物に対して幅広い抗微生物スペクトルを有することが明 らかとなり,農業資材として有望である. もん ぱ また,松本らは,紫紋羽病菌および白紋羽病菌のいくつかの菌株が,菌類ウイルスに由来する 病原力低下因子(dsRNA)を保有していることを明らかにした(平成 14 年度研究成果情報,松本 かんちゅう ら) .果樹類の土壌伝染性病害である紋羽病の防除は困難で,農薬の潅注,くん蒸剤による土壌消 毒,あるいは表土のはく離など,化学的あるいは物理的方法による方法のみが実用的な対策であ る.しかし,慣行防除は環境負荷が大きいにもかかわらず,土壌中の病原菌は完全に死滅するわけ ではないので,頻繁な対応が不可欠である.病原力低下因子を含む菌株は,紋羽病菌の病原力の低 下を招くので,紋羽病の生物防除への利用が期待できる. 5.今後の展望 以上,この 10 年間に農業環境技術研究所で行われた環境生物の情報化学物質に関わる研究成果 を紹介してきたが,そのいずれも農薬等の化学資材の投入量を減らし環境負荷の低減に役立つ農業 技術の開発に貢献する研究である.こうした環境負荷低減農業技術が開発される背景として,2003 年 12 月に取りまとめられた「農林水産環境政策の基本方針」(農林水産省循環型社会構築・地球温 暖化対策推進本部決定)で,環境保全に向けて農業者の主体的な努力を促すため,適切な肥料,農 薬の使用等による環境負荷の低減とたい肥を利用した土づくりによる物質循環を促進する方針が策 定された.さらに,2005 年 3 月に閣議決定された新たな「食料・農業・農村基本計画」でも,「環 境保全を重視した施策の展開」を図ることが農業の改革に当たっての基本的視点として位置づけら れている.こうした流れは,2010 年 3 月 30 日に閣議決定された新たな「食料・農業・農村基本計 画」でも,気候変動をはじめとする地球環境問題の進行に対する対応方向として,環境に配慮し た持続可能な経済社会への転換に向けて,農業・農村や食品産業においても,環境に配慮した生産 活動を適切に推進することがうたわれている.このように,農薬や化学肥料の使用を低減するため に,農業環境に生息する生物が生産する情報化学物質を介した有用機能を利用しようとする研究に 対するニーズは,ますます強くなっているといえる. 昆虫の性フェロモンに関わる研究は,大量捕殺や交信撹乱,害虫の発生予察のための技術として すでに広く農業現場で利用されており,2000 年 7 月に制定された「持続性の高い農業生産方式の 導入の促進に関する法律」の中で,フェロモン剤利用技術が「持続性の高い農業生産方式」として 公式に認められている.今後は,有効なフェロモン剤のない害虫に対する新規フェロモン剤の開発 150 引用文献 や,既存のフェロモン剤に対するより高い誘引性の付与,誘引耐性の発生機構の解明などの研究 ニーズに答えてゆく必要があるだろう.また,植物が生産するアレロケミカルについても,「持続 性の高い農業生産方式」の中に,対抗植物利用技術(土壌中の有害動植物を駆除し,又はそのまん 延を防止する効果を有する植物を栽培する技術)という表現でアレロパシー作用を持つ植物の有効 利用技術として言及されており,こうした技術に利用できる有用植物やアレロケミカルの探索に対 するニーズはきわめて高い.現在,農業環境技術研究所では,生物系特定産業技術研究支援セン ターの「イノベーション創出基礎的研究推進事業」で 2008 年に採択された「アレロケミカルの探 索と新規生理活性物質の開発」の研究が,2012 年度までの予定で進行しており,新しい除草剤や 農薬のリード化合物となるアレロケミカルと,新しい有用な被服植物として利用が期待されるアレ ロパシー活性の強い植物の探索が精力的に進められている.微生物の有用機能酵素や生理活性物質 については,これまで農業現場よりも食品加工や医療現場で主にその利用が図られてきたが,作物 きっこう 病原菌に対する拮抗微生物や作物の生育を促進したり植物の生体防御機能を活性化させたりする微 生物の研究は,文字通り環境負荷の少ない農業技術として,その利用が期待される.最近,発展の めざましいメタゲノム解析技術は,これまで培養できないが故に手つかずであった未知の微生物資 源から有用機能を有する酵素や生理活性を作り出す可能性を秘めており,飛躍的な技術革新を引き 起こす技術として特に期待される. 引用文献 1)Amann, R. I., Ludwig, W. and Schleifer, K. H.(1995)Phylogenetic identification and in situ detection of individual microbial cells without cultivation. Microbiological reviews, 59, 143-169 2)Arakawa, M., Nakamura, H., Uetake, Y. and Matsumoto, N.(2002). Presence and distribution of double-stranded RNA elements in the white root rot fungus, Rosellinia necatrix. Mycoscience, 43, 21-262 3)大場広輔,岡田浩明(2008)PCR−DGGE 法による土壌生物群集解析法(2)土壌線虫相の解 析 . 土と微生物 , 62, 69-73 4)Fujii, T., Miyashita, K., Ohtomo, R. and Saito, A.(2005)DNA-binding protein involved in the regulation of chitinase production in Streptomyces lividans. Bioscience, Biotechnology, and Biochemistry, 69, 790-799 5)Fujii, Y.(2003)Allelopathy in the natural and agricultural ecosystems and isolation of potent allelochemicals from Velvet bean(Mucuna pruriens)and Hairy Vetch(Vicia villosa).Biological Science in Space, 17, 6-13 6)Fujii, Y. Shibuya, T., Nakatani, K., Itani, T., Hiradate, S. and Parvez, M. M.(2004)Assessment method for allelopathic effect from leaf litter leachates. Weed Biology and Management, 4, 19-23 7)Francisco, P. B. Jr., Ogawa, N., Suzuki, K. and Miyashita, K.(2001)The chlorobenzoate dioxygenase genes of Burkholderia sp. strain NK8 involved in the catabolism of chlorobenzoates, Microbiology, 147, 121-133 8)Furubayashi, A., Hiradate, S. and Fujii, Y.(2005)Adsorption and transformation reactions of L-DOPA in soils. Soil Science and Plant Nutrition, 51, 819-825 9)Handelsman, J., Rondon, M. R., Brady, S. F., Clardy, J. and Goodman, R. M.(1998)Molecular 第 10 章 環境生物の情報化学物質と機能利用 151 biological access to the chemistry of unknown soil microbes: a new frontier for natural products. Chemistry & Biology, l5, 245-249 10)Hiradate, S., Yoshida, S., Sugie, H., Yada, H. and Fujii, Y.(2002)Mulberry anthracnose antagonists(Iturins)produced by Bacillus amyloliquefaciens RC-2. Phytochemistry, 61, 693-698 11)Hiradate, S., Morita, S., Sugie, H., Fujii, Y. and Harada, J.(2004)Phytotoxic cis-cinnamoyl glucosides from spiraea thunbergii. Phytochemistry, 65, 731-739 12)Hiradate, S., Kamo, T., Nakajima, E., Kato, K. and Fujii, Y.(2005-1)Direct quantitative determination of cyanamide by stable isotope dilution gas chromatography-mass spectrometry. Journal of Chromatography, A 1098, 138-143 13)Hiradate, S., Morita, S., Furubayashi, A., Fujii, Y. and Harada, J.(2005-2)Plant growth inhibition by cis-cinnamoyl glucosides and cis-cinnamic acid. Journal of Chemical Ecology, 31, 591-601 14)Hiradate, S., Furubayashi, A. and Fujii, Y.(2005-3)Changes in chemical structure and biological activity of L-DOPA as influenced by an andosol and its components. Soil Science and Plant Nutrition, 51, 477-484 15)Hoshino, Y. T. and Matsumoto, N.(2004)An improved DNA extraction methods using skimmed milk from soils that strongly adsorb DNA. Microbes and Environments, 19, 13-19 16)Hoshino, Y. T. and Matsumoto, N.(2005)Skim milk drastically improves the efficacy of DNA extraction from andisol, a volcanic ash soil, JARQ, 39, 247-252 17)星野(高田)裕子(2005) スキムミルクを用いた土壌からの DNA 抽出改良法とその活用に ついて . 農業技術 , 60, 127-130 18)Iqbal, Z., Iqbal, Hiradate, S., Noda, A., Isojima, S. and Fujii, Y.(2002)Allelopathy of buckwheat: Assessment of allelopathic potential of extract of aerial parts of buckwheat and identification of fagomine and other related alkaloids as allelochemicals. Weed Biology and Management,2,110-115 19)Iqbal, Z., Hiradate, S., Noda, A., Isojima, S. and Fujii, Y.(2003)Allelopathic activity of buckwheat: isolation and characterization of phenolics. Weed Science, 51, 657-662 20)Iqbal, Z., Furubayashi, A. and Fujii, Y.(2004-1)Allelopathic effect of leaf debris, leaf aqueous extract and rhizosphere soil of Ophiopogon japonicus Ker-Gawler on the growth of plants. Weed Biology and Management, 4, 43-48 21)Iqbal, Z., Hiradate, S., Araya, H. and Fujii, Y.(2004-2)Plant growth inhibitory activity of Ophiopogon japonicus Ker-Gawler and role of phenolic acids and their analogues: a comparative study. Plant Growth Regulation, 43, 245-250 22)Kato-Noguchi, H., Yamamoto, M., Tamura, K., Teruya, T., Suenaga, K. and Fujii, Y.(2010) Isolation and identification of potent allelopathic substances in rattail fescue. Plant Growth Regulation, 60, 127-131 23)野口 浩(1999)性フェロモン利用による害虫防除の現状と課題 . 植物防疫,53, 398-402 24)Morimoto, S., Togami, K., Ogawa, N., Hasebe, A. and Fujii, T.(2005)Analysis of a bacterial community in 3-chlorobenzoate-contaminated soil by PCR-DGGE targeting 16S rDNA and the benzoate 1,2-dioxygenase gene(benA). Microbes and Environments, 20, 151-159 25) Morimoto, S., Ogawa, N., Hasebe, A. and Fujii, T. (2008) Isolation of effective 152 引用文献 3-chlorobenzoate-degraders in soil using community analyses by PCR-DGGE. Microbes and Environments, 23, 285-292 26)森本 晶,星野(高田)裕子(2008)PCR-DGGE 法による土壌生物群集解析法(1)一般細 菌・糸状菌の解析 . 土と微生物 , 62, 63-68 27)Morimoto, S. and Fujii, T.(2009)A new approach to retrieve full lengths of functional genes from soil by PCR-DGGE and metagenome walking. Applied Microbiology and Biotechnology, 83, 389-396 28)Nakajima, E., Iqbal, Z., Araya, H., Hiradate, S. and Fujii, Y.(2005)Isolation and identification of a plant growth promotive substance from mixture of essential oils. Plant Growth Regulation, 45, 47-51 29)Nishihara, E., Parvez, M. M., Araya, H. and Fujii, Y.(2004)Germination growth response of different plant species to the allelochemical L - 3, 4 - dihydroxyphenylalanine(L-DOPA). Plant Growth Regulation, 42, 181-189 30)Ogawa, N., Chakrabarty, A. M. and Zaborina, O.(2004)Degradative plasmids, p.341-376. In E. Funnell and G. J. Philips(ed.) , Plasmid biology. ASM Press, Washington, D.C. 31)Ohno, M., Kataoka, S., Numata, S., Yamamoto - Tamura, K., Fujii, T., Nakajima, M., Akutsu K. and Hasebe, A.(2011)Biological control of Rhizoctonia damping-off of cucumber by a transformed Pseudomonas putida strain expressing a chitinase from a marine bacterium. JARQ, 45,91-98 32)Parvez, S.S., Parvez, M. M., Fujii, Y. and Gemma, H.(2003)Allelopathic competence of Tamarindus indica L. root involved in plant growth regulation. Plant Growth Regulation,41, 139-148 33)Saito, A., Kaku, H., Minami, E., Fujii, T., Ando, A., Nagata, Y., Schrempf, H. and Miyashitra, K. (2006)An enzyme-linked immunosorbent assay(ELISA)to determine the specificity of the sugar-binding protein NgcE, a component of the ABC transporter for N-acetylglucosamine in Streptomyces olivaceoviridis. Bioscience, Biotechnology, and Biochemistry, 70, 237-242 34)Saito, A., Shinya, T., Miyamoto, K., Yokoyama T., Kaku H., Minami, E., Shibuya, N., Tsujibo, H., Nagata, Y., Ando, A., Fujii, T. and Miyashita, K.(2007)The dasABC gene cluster adjacent 1 to dasR encodes a novel ABC transporter for the uptake of N,N’-diacetylchitobiose in Streptomyces coelicolor A3(2) . Applied Environmental Microbiology, 73, 3000-3008 35)Sakai, Y., Ogawa, N., Fujii, T., Sugahara, K., Miyashita, K. and Hasebe, A.(2007)2,4-Dichro lophenoxyacetic acid-degrading genes from bacteria isolated from soil in Japan: Spread of Burkholderia cepacia RASK-type degrading genes harbored on Large Plasmids. Microbes and Environments, 22, 145-156 36)Sugie H., Yase, J., Futai, K. and Shirai, Y.(2003)A sex attractant of the cabbage webworm, Hellula undalis Fabricius(Lepidoptera: Pyralidae).Applied Entomology and Zoology, 38, 45-48 37)Tabata, J., Minamishima, M., Sugie, H., Fukumoto, T., Mochizuki, F. and Yoshiyasu, Y.(2009) Sex pheromone components of the pear fruit moth, Acrobasis pyrivorella(Matsumura). Journal of Chemical Ecology, 35, 243-249 38)Tabata, J., Noguchi, H., Kainoh, Y., Mochizuki, F. and Sugie, H.(2007)Sex pheromone production and perception in the mating disruption-resistant strain of the smaller tea 第 10 章 環境生物の情報化学物質と機能利用 153 leafroller moth, Adoxophyes honmai. Entomologia Experimentalis et Applicata, 122, 145-153 39)Tsuda, K., Takamura, N., Matsuyama, M. and Fujii, Y.(2005)Assessment method for leaf litters allelopathic effect on Cyanobacteria. Journal of Aquatic Plant Management, 43, 43-46 40)Wang, Y., Shimodaira, J., Miyasaka, T., Morimoto, S., Oomori, T., Ogawa, N., Fukuda, M. and Fujii, T.(2008)Detection of bphAa gene expression of Rhodococcus sp. strain RHA1 in soil using a new method for RNA preparation from soil. Bioscience, Biotechnology, and Biochemistry, 72, 694-701 41)Wang, Y., Morimoto, S., Ogawa, N., Oomori, T. and Fujii, T.(2009)An improved method to extract RNA from soil with efficient removal of humic acids. Journal of Applied Microbiology, 107, 1168-1177 42)Yoshida, S., Hiradate, S., Tsukamoto, T., Hatakeda, K. and Shirata, A.(2001)Antimicrobial activity of culture filtrate of Bacillus amyloliquefaciens RC-2 isolated from mulberry leaves. Phytopathology, 91, 181-187. 43)Yoshida, S., Shirata, A. and Hiradate, S.(2002)Ecological characteristics and biological control of mulberry anthracnose. JARQ, 36, 89-95 44) (社) 日本植物防疫協会編(2000)フェロモン剤利用ガイド (コラム 1)薬剤耐性獲得機構の解明 イネいもち病防除薬剤として全国で広く使用されるシタロン脱水酵素阻害剤(MBI−D 剤)に, 最近西日本でいもち病菌の耐性菌が出現し問題となっている.この薬剤耐性はシタロン脱水酵素遺 伝子のコドン 75 における GTG(推定アミノ酸バリン)から ATG(同メチオニン)への 1 塩基変 異によって起こることが知られている.石井は,PCR−Luminex 法でシタロン脱水酵素遺伝子の 1 塩基変異を解析することにより,イネいもち病菌のシタロン脱水酵素阻害剤(MBI−D 剤)耐性菌 を,多検体の中から迅速に検出する技術を開発した(平成 15 年度研究成果情報,石井).この方法 は従来法に比べて,制限酵素処理や電気泳動,エチルブロマイドによる電気泳動ゲルの染色などの 煩雑な操作を必要とせず,簡便で作業の安全性や快適性が高い.また,正確かつ短時間に結果を判 定することができるほか,96 穴のマイクロプレートを使用するので多検体処理(ハイスループッ ト)が可能である. 農薬に対する耐性出現は,害虫でも同様で,化学合成殺虫剤の連用による殺虫剤抵抗性の発達が 問題となっている.殺虫剤抵抗性は遺伝するため,発達の実態把握や動態予測のためには,抵抗性 に関与する遺伝子を持つ個体の割合を正確に知る必要がある.ワタアブラムシは多種の農作物を加 害する重要害虫で,近年各種殺虫剤に対して抵抗性を発達させてきている.鈴木は,ワタアブラム シの有機リン剤抵抗性の原因となる解毒酵素(カルボキシルエステラーゼ)遺伝子の塩基配列には 抵抗性系統と感受性系統の間に違いがあることを見いだし,この違いに基づいて設計したプライ マー対を用いた PCR によって抵抗性系統と感受性系統を明確に識別できることを示した(平成 16 年度研究成果情報,鈴木) .これまで,ワタアブラムシの有機リン剤抵抗性の検定は,労力や時間 がかかり,その上得られるデータの信頼性が低かったが,本法は,ゲノム DNA を鋳型として明確 に識別可能であり,RNA の精製や逆転写反応等の煩雑な過程を要さないため,多検体を調査する 必要のある抵抗性のモニタリング等に活用できる. (コラム 2)植物の生体防御機能の活性化機構の研究 植物の生体防御機能を活性化して各種病害に対する複合抵抗性を発現させる全身抵抗性誘導の分 154 子機構は,草本植物を中心に解明されつつある.しかし,殺菌剤使用回数が多いナシなど永年性 作物における抵抗性誘導の研究事例は少ない.石井は,病害抵抗性誘導剤であるアシベンゾラル S メチルによってナシの感受性品種‘幸水’に黒星病抵抗性が誘導される過程で,ポリガラクツロ ナーゼ阻害たんぱく質(PGIP)遺伝子の転写量が抵抗性品種と同様増大することを見いだし,抵 抗性発現には PGIP が密接に関与することを明らかにした(平成 15 年度研究成果情報,石井).ま た,農薬の環境影響軽減に果たすアシベンゾラル S メチルの可能性を探るために,作物の葉面糸 状菌相に及ぼすアシベンゾラル S メチルの影響を数種の殺菌剤と比較した(平成 16 年度研究成果 情報,石井) .その結果,ジチオカーバメート系などの殺菌剤散布は,キュウリやナシの葉面糸状 菌相を単純化させ糸状菌数を減少させるが,非殺菌性の病害抵抗性誘導化合物,アシベンゾラル S メチルは葉面糸状菌相に影響しないことが明らかとなった. 155 Ⅳ.農業環境の把握と情報の活用 ― 農業環境を支える 第 11 章 リモートセンシング・地理情報システムの利用 1.はじめに 農業環境研究においては,様々な空間解像度において研究を展開する必要がある.すなわち,圃 場レベルから,国・地球レベルまでの範囲が研究対象となる.そのとき,国・地球レベルの範囲に 対して,現地における観測・調査のみで広域にわたって信頼性のおける情報を得ることは不可能で ある.リモートセンシングは,その言葉の定義のとおり,人工衛星や航空機など地上から離れた場 所から観測を行う技術であり,とくに広域にわたる範囲の情報を迅速に得る技術として有効であ る.そのため,農業環境技術研究所では,1988 年の研究所発足時から,リモートセンシング研究 を本格的に開始している. リモートセンシング技術は,人工衛星からの広域的な観測のみでなく,圃場レベルでの観測にお いても,対象に非接触で迅速な観測を行う技術として開発が進んでいる. 地理情報システム(Geographic Information System, GIS)は,地図情報を電子化して解析を行う システムである.GIS で扱う情報は,紙の地図に記載された境界線などにとどまらず,位置情報を もつ様々な属性データを含めることができる.農業環境に関する研究においては,たとえば,地球 温暖化問題,生物多様性問題,リスク評価問題など,いずれにおいても,地理的な位置関係が複雑 に関与している.すなわち,農業環境研究において GIS 研究は必須であるといえる.農業環境技 術研究所においては,とくに独立行政法人化とともに,GIS 研究が実施されている. 2.リモートセンシング技術 リモートセンシングにおいては,太陽から照射され対象物によって反射した電磁波,あるいはセ ンサー自体から照射され対象物によって反射した電磁波の強度が観測される.そのとき,得られる リモートセンシング画像は,センサーの持つ(1)空間解像度,(2)波長解像度,(3)時間解像度, の 3 つの側面から影響を受ける.リモートセンシングにおいては各種のセンサーが利用されている ため,それぞれの特徴を考慮した解析が行われている.衛星画像を利用したリモートセンシングで は,空間解像度は制限されるものの,広域での土地利用把握が可能である.農業環境技術研究所に おいても,人工衛星搭載のセンサーの特徴を生かしたデータ解析手法が開発されている. 圃場レベルでの計測においては,高い空間解像度で対象の観測を行うことができる.詳細な植物 生育状態の計測を行うとともに,衛星搭載のセンサーへの基礎情報を提供するものである. ⑴ 衛星リモートセンシングによる広域的土地利用の把握 1 衛星マイクロ波レーダによる雲に影響されない水稲作付け面積の推定 広域的な水稲作付け面積の推定に関して,現在行われている標本調査法では多大な労力を必要と するため,リモートセンシングを利用した省力化・迅速化が求められていた.そこで,衛星搭載の 合成開口レーダ(SAR)画像を用いて水稲作付け地を精確に,また省力的に推定する手法が開発さ れた(平成 16 年度研究成果情報 , 大野ら) (石塚ら , 2003). SAR は衛星からマイクロ波を照射し,地表面での散乱後に戻ってくる電磁波(後方散乱)の強 度を観測する.可視光より波長の長いマイクロ波は雲を透過するため,天候にかかわらず地表面を 156 2.リモートセンシング技術 観測することが可能である.田植期の水田では鏡面反射し,水稲生長期には植物体により後方に強 く散乱することを利用して,二つの時期の画像を比較することにより天候によらず確実に水稲作付 け地を検出することができる.この解析により,調整水田(雑草対策のため湛水のみを実施する減 反水田)も抽出可能である(図1a) .さらに本研究では,地理情報システムを利用して非水田地域 を除去することにより精度を高めている.佐賀平野の 26 市町村において現行の統計的手法と比較 ᳓㕙ߢߩ㏜㕙 ᳓Ⓑ߇↢㐳ߒߚ᳓↰ ᧄᛛⴚߦࠃࠆઃ㕙Ⓧផቯ㧔ha㧕 したところ,現行と同等の精度が得られている(図1b). 3000 2500 ᐔဋੑਸ਼⺋Ꮕ㧦85ha 2000 1500 1000 500 0 0 500 1000 1500 2000 2500 3000 ⴕߩ⛔⸘⺞ᩏߦࠃࠆઃ㕙Ⓧ㧔ha㧕 図 1a 二時期 SAR 画像による水稲作付地の検出 田植期と生長期におけるマイクロ波散乱の違い により水稲作付け地を検出する. 図 1b 現行統計調査法との比較 現行の方法と同等な精度で水稲作付け面 積の推定が可能(佐賀平野 26 市町村). 2 高頻度観測衛星(MODIS/Terra)を活用した湛水域と水稲生育ステージの把握 高頻度観測衛星 MODIS/Terra 画像は,空間解像度は高くないものの(500m) ,ほぼ毎日観測さ れている.このことを利用して,短期的な雲の影響を取り除くことが可能である.MODIS/Terra の時系列的に観測される画像データから,地上の湛水状態や,水稲の生育ステージを動的に把握 する手法が開発された(平成 18 年度研究成果情報,坂本ら)(Sakamoto et al., 2005, 2006).メコ ンデルタの水稲栽培地帯において,植生指数の季節変化曲線から推定した水稲作付パターンを図 2 図 2 ベトナムメコンデルタの推定水稲作付パターン 右側のグラフは,2002 年から 2003 年にかけて乾期二期作から三期作に変化し た場所(図中矢印)の植生指数の時系列変化を示している. 第 11 章 リモートセンシング・地理情報システムの利用 157 に示す.二期作や三期作の空間的な広がりや,年次間での変動が把握できる.今後,広域的な農業 的土地利用の季節および年次変動のモニタリングへの利用が期待される. 3 衛 星 画 像 による東南アジア山岳焼畑地帯 の 炭 素 蓄 積 量 の 実 態 把 握 と シ ナ リ オ の 提 示 焼畑は現在においても東南アジア山岳地帯の広範な地域で重要な食糧生産システムである.しか し,面積の拡大と休閑期間の短縮が急速に進み,土地生産性・労働生産性の低下だけでなく,森林 資源の劣化と CO2 の放出,土壌浸食,生物多様性の損耗等が懸念されている.そこで衛星画像デー タの解析により,焼畑農業の現状把握と管理シナリオを提示する研究が行われた(平成 19 年度研 究成果情報 , 井上) (Inoue et al., 2007) . ラオスにおける 30 年間以上にわたる Landsat 衛星画像と最新の高解像度衛星(QuickBird/ IKONOS)画像を用い,年々移動する焼畑利用地の区画を高精度で抽出することによって,焼畑面 積・休閑期間・樹齢などの空間的分布と時間変化が明らかにされた.さらに,様々な管理シナリオ を比較することによって,収益を維持あるいは向上させつつ炭素蓄積量も増加できることが示され ている(図3) .今後,途上国における食料確保と環境保全を両立させるための方策へ役立てること が期待される. 100 䋱ᐕઃ+䋵ᐕભ㑄 ↢ᘒ♽⚛㊂ (t/ha) 90 䋲ᐕઃ+10 ᐕભ㑄 80 㐳ᦼᐔဋ 62.1 t/ha 70 60 㐳ᦼᐔဋ 51.9 t/ha 50 40 㐳ᦼᐔဋ 34.9 t/ha 30 20 10 䋱ᐕઃ+䋲ᐕભ㑄 0 0 5 10 15 20 ᐕ 25 30 35 40 ᢙ 図 3 焼畑 - 休閑期間の違いによる生態系炭素量の長期変化 「2 年作付+10 年休閑」パターンの場合,炭素量(35 年間平均)は,「1 年作付+2 年休閑」に 比べ,約 27t/ha 多くなると推定される. ⑵ 圃場における計測システムの開発 1 多周波マイクロ波リモートセンシングによる群落特性の計測 圃場に設置した計測器から,異なる周波数のマイクロ波を照射し,植物体からの後方散乱を計測 するシステムが開発された(平成 15 年度研究成果情報 , 井上)(Inoue et al., 2002).たとえば生育 した水稲を観測すると,周波数の違いによりマイクロ波の散乱の様子が異なることが分かる(図4 a) .この性質を利用して,葉面積指数・バイオマス・茎数密度・穂重などの形質を推定することが できる.図4b に Ku バンド(周波数 15.95GHz)による穂重の推定値と実測値の関係を示す. この結果は,水田や畑の精密な管理に利用したり,将来の衛星リモートセンシングへの基礎的な 情報提供として利用したりすることが可能である. 2.リモートセンシング技術 㽳 㽶 㽵 1.2 㽲 1.0 丏丱丈ฬ ᵄᢙ ᢔੂ䊒䊨䉶䉴 (GHz) 㽲 㽳 㽴 㽵 㽶 Ka 35.25 Ku 15.95 X 9.60 C 5.75 L 1.26 1:1 㽴 ታ᷹ߒߚⓄ㊀ (kg m-2) 158 2 r =0.998 0.8 0.6 0.4 ⷺ 35° ု⋥ᵄ 0.2 0.2 0.4 0.6 0.8 1.0 1.2 Ku ࡃࡦ࠼ߦࠃࠆⓄ㊀ߩផቯ୯ (kg m-2) 図 4a 周波数の異なるマイクロ波による様々な散 乱プロセス 図 4b Ku バンドによる穂重推定 2 リモートセンシングと生態プロセスモデルの協働による評価手法 土壌 ― 植物 ― 大気系の熱・水収支や 光合成過程を記述する生態プロセスモデ ᬀ‛䊌䊤䊜䊷䉺 ფ䊌䊤䊜䊷䉺 ᳇⽎䊂䊷䉺 ルは,入力パラメータが完全に得られる ↢ᘒ䊒䊨䉶䉴䊝䊂䊦 場合には生長や蒸発散等を的確に予測す ることができる.しかし実際には,プロ セスモデルが必要とするパラメータを広 域的に入手することは困難であり,プロ することは難しい.そこで,リモートセ ンシング信号を出力するプロセスモデル ᳇ሹ䉮䊮䉻䉪䉺䊮䉴 శวᚑㅦᐲ ⟲⪭శวᚑㆊ⒟ ᳓䉴䊃䊧䉴 ⪲㕙Ⓧᜰᢙ 䊋䉟䉥䊙䉴↢㐳 ⪲㕙Ⓧ↢㐳 ᾲᡰㆊ⒟ ᳓ᡰㆊ⒟ ᓸ↢‛ๆ を構築し,実際のリモートセンシングに よる観測値と対比することによって,プ ロセスモデル内の未知パラメータを自動 的にチューニングする協働手法が開発さ れた(平成 16 年度研究成果情報 , 井上) (Inoue et al., 2005) .図5 に協働システ ムの概要を示す.たとえば土壌水分デー タが得られない場合でも,リモートセン シングによる植生指数の実測値を本協働 หൻ㊂ 䊥䊝䊷䊃䉶䊮䉲䊮䉫 ାภ‛ℂ䊝䊂䊦 䉲䊚䊠䊧䊷䉲䊢䊮୯ >㕙᷷ᐲ >ಽశ₸ 両丆个丐丨丠丶䷾䶺一两丶上丱䷯ セスモデル単独では精度と汎用性を確保 ᳇ሹᔕ╵ㆊ⒟ ⪲శวᚑㆊ⒟ ⺋Ꮕ䈱 ᦨዊൻ 䊥䊝䊷䊃䉶䊮䉲䊮䉫ታ᷹୯ >㕙᷷ᐲ >ಽశ₸ システムに適用することにより,ほぼ実 測値に近い土壌水分を推定でき,バイオ マス生長および生態系 CO2 フラックス を動的かつ高精度で予測できることが確 ᬀⵍ䈱↢ℂ↢ᘒᄌ㊂䈱േ⊛੍᷹ > ᬀ↢ [⪲㕙Ⓧᜰᢙ,䊋䉟䉥䊙䉴,⫳ᢔ,శวᚑ] > ფ [᳓ಽ, ⫳⊒ᢔ] > 䊐䊤䉾䉪䉴 [䉣䊈䊦䉩, H2O, CO2] 認された. 図 5 生態プロセスモデルとリモートセンシングの協働シス テム 第 11 章 リモートセンシング・地理情報システムの利用 159 3 ハイパースペクトル計測を利用した群落光合成パラメータの評価 地球規模の炭素循環における陸域植生の機能や作物生産力の広域評価のため,光吸収率などの群 落光合成パラメータをリモートセンシングで計測する方法の開発が強く期待されている.そこで, ハイパースペクトル計測を利用して,重要な群落光合成パラメータを精度よく推定するための評価 指数が開発された(平成 19 年度研究成果情報 , 井上)(Inoue et al., 2008).水稲群落において,ハ イパースペクトルデータ(波長範囲 400 〜 2400nm, 解像度 1nm)を 3 か年にわたって計測し,群 落光合成に関連するパラメータを推定するための正規化分光反射指数 NDSI [Ri, Rj] =(Ri−Rj) (Ri+Rj) / Ri と Rj は波長 i nm と j nm の分光反射率 を探索した結果,光吸収率 fAPAR と高い相関をもつ指数として NDSI [420, 720] が見いだされた (図6a) .この新規指数は,従来用いられてきた植生指数 NDVI=NDSI [660, 830] に比べて全生育期 間を通じて一貫性が高く,高精度で安定した推定が可能である(図6b).この成果は,リモートセ ンシングによって作物群落や陸域生態系のバイオマス生産量・CO2 フラックスを広域的に評価する ための基礎として多方面での活用が期待される. 図 6a 全波長組合せによる NDSI の推定力 (群落光吸収率 fAPAR と NDSI の相関係数) 図 6b 新規スペクトル指数による群落光吸収率 の推定 4 作 物 か ら の分光反射率画像を長期間自動 的 に 観 測 し 生 育 状 態 を 追 跡 す る シ ス テ ム 水田や畑で育つ農作物は常に生長を続けており,その生育量や栄養状態は時々刻々と変化してい る.このような農作物の生育状態の変化を経常的に把握することは,各種の農業研究の現場で必要 とされるだけでなく,衛星リモートセンシングの地上検証データとしても重要である.しかし,こ れまで行われていた草丈・乾物・葉面積などの生育調査法は,多大な労力を要し,そのためサンプ リング回数も限られていた.そこで,作物の生育状態を長期間自動的に追跡するシステムが開発さ れた(平成 21 年度研究成果情報 , 芝山ら) (Shibayama et al., 2009; 高田ら , 2009).この装置は, 次のような特徴をもっている(図7a) . (1)圃場の上空数メートルに設置し,継続的に撮影する. (2)完全防水であり,天候にかかわらず観測を続けることができる.(3)近赤外などを含めた波長 フィルタを簡単に交換できるので,目的に応じた特性値を観測できる.(4)入射光に対する反射光 の比率(反射率)を計算するため,撮影時の明るさに依存しない安定した値が得られる. 160 3.地理情報システムを利用した農業生態系の解析 本装置の利用例として,近赤外バンド反射率の日平均値,撮影方向,植え付け方向などを説明変 数とする回帰式を生育期間中の葉面積指数に適用したところ,精度よく省力的に推定できることが 確認された(図7b) . 図 7a 全天候反射率画像撮影装置 カメラヘッドは 2 ないし 3 波長バンドの分光 反射率画像を捉えることがでる. 図 7b 撮影装置による葉面積の推定 3.地理情報システムを利用した農業生態系の解析 リモートセンシングデータについては,各種衛星や航空写真による観測が開始された時点からの データの利用が可能であるものの,時間的にさかのぼって農業環境を調べるには限界がある.一 方,地図として残されているデータには,100 年以上前までさかのぼることのできるものも存在す る.紙として残された地図データも,電子化することによって,農業環境の時間的な変化を解析す ることが可能である. ⑴ 迅速測図を用いた過去 120 年間の農業的土地利用変化の解析 明治初期(1880 年代)に日本陸軍により作成された迅速測図は,日本で初めて近代的な手法に よって関東一円にわたって測量された地図であり,世界的にみても 19 世紀にこれだけ広範囲の地 形図を整備した例はまれである.迅速測図には,地形と土地利用が描かれており,120 年前のわが 国の土地利用状況を知る上で貴重な資料となっている.しかし,迅速測図には地図座標が無く, 緯度・経度も付されていないため,現在の地形図との重ね合わせが困難であった.そこで,2 段階 の幾何補正によって迅速測図の GIS 化を行う手法が開発された(平成 15 年度研究成果情報 , スプ レイグら) (Sprague et al., 2007; スプレイグら , 2007).最初に基準点を用いたアフィン変換を行い (1 次補正) ,さらにラバーシーティングによる詳細な補正(2 次補正)を行うことによって高い精 度で現在の地図と重ね合わせることが可能となった.図8a)に迅速測図から GIS 化した茨城県牛久 地方の 1880 年代の土地利用を示す. さらに,図法や縮尺の異なる地形図や植生図を GIS に取り込み,共通の座標系で重ね合わせる ことにより,長期間にわたる農業的土地利用変化の詳細な解析が可能となる.茨城県牛久地方で は,急激な都市化が進むとともに,14%を占めていた草地が 120 年の間にほとんど消失しているこ とが分かる(図8) . 第 11 章 リモートセンシング・地理情報システムの利用 161 a) 1880 ᐕઍ b) 1900 ᐕઍ c) 1920 ᐕઍ d) 1980 ᐕઍ e) 2000 ᐕઍ 図 8 茨城県牛久地域における 120 年間の土地利用変化 a)迅速測図,b)5 万分の 1 地形図,c)2 万 5 千分の 1 地形図 , d)環境省現存植生図,e)現存植生図を航空写真にて修正 ⑵ コスト距離法を用いた野生動物生息域の評価・予測手法の開発 ニホンザルやシカ,クマ等の野生動物による農作物や人的被害が問題となっており,これらの被 害の防除や軽減のために,野生動物と人間の活動域を分離するための環境管理が必要とされてい る.これまで,土地利用地目や植生に基づく生息地の評価は行われているが,その配置や面積,形 状等の空間構造を考慮した評価手法は確立していなかった.そこで,野生動物の生息域変化に及ぼ す空間構造の影響を定量的に評価するため,生息域の拡大のしにくさを示すコスト距離という概念 を用いた手法が開発された(平成 17 年度研究成果情報 , 岩崎ら).コスト距離とは,土地利用ごと に設定されるコスト値と実際の距離との積で示され,コスト値が大きいほど生息域拡大時に通過し にくい土地利用といえる.本研究では,生息域拡大の有無を目的変数,生息域が拡大したコスト距 離を説明変数とするロジスティック回帰分析によりコスト値が推定されている. 1970 年代から 1980 年代半ばにかけての房総半島におけるニホンザルの生息確認データから推定 されたコスト値を表 1 に示す.コスト値は一定値ではなく,樹林地で低く,畑や水田等の農耕地, ゴルフ場や草地等の開放的な空間および住宅地で高い傾向が認められる.直線距離とコスト距離と の比較を図9 に示す.対象地域の南部には直線距離が近いにもかかわらず生息域拡大が認められな いが,この地域は水田や住宅地が密集し累積コスト距離が遠いためと考えられる. 表 1 各種土地利用に対する推定コスト値 土地利用 単純距離 コスト値 落葉広葉樹 1 2 常緑広葉樹 1 1 常緑針葉樹 1 1 笹 1 1 竹 1 6 草地・荒地・ゴルフ場 1 28 果樹園等 1 1 畑 1 8 水田 1 9 土手・崖 1 2 湿地 1 1 住宅地・空地 1 12 162 4.時空間情報のデータベース化とインターネットでの利用 a) 直線距離図 b) コスト距離図 図 9 1970 年代のニホンザルの生息確認地点からの直線距離図とコスト距離図の比較 4.時空間情報のデータベース化とインターネットでの利用 農業生態系に関するリモートセンシングデータや地理情報データは,空間的に広域に広がるだけ でなく,時間的にも大きく変動し,そのデータの規模は極めて大きなものとなる.このような農業 生態系データを有効に利用するため,効率的に設計された形でデータベース化が行われるととも に,その利用法が開発されている. ⑴ 日本の全流域圏・農耕地を網羅する農業生態系空間情報解析システム わが国の全流域圏・全農耕地をカバーし,農業生態系情報の各種解析とシミュレーションを行 うことのできる汎用的プラットフォームとして「農業生態系空間情報解析システム」(GSAS: GeoSpatial Agro-ecosystem Simulator)が開発されている(平成 18 年度研究成果情報 , 井上). GSAS は全国全流域圏の全農耕地をカバーし,圃場境界に沿った 1 区画数 ha の農地区画(セル) を単位として,自然立地データ(標高・傾斜・気温・降水量・土壌等),圃場基盤データ(用排水 条件・田畑面積等) ,統計調査データ等を一元的に集積・管理している.このシステムにより,任 意の流域界・行政界や任意の形・サイズの領域を選定し,解析やシミュレーションを行うことがで きる.さらに,衛星画像情報やシミュレーション結果をセルごとに追加し自己増殖することが可能 である(図10) . このシステムは,農耕地の作物生産機能や生態的役割,環境負荷など,農業と環境に関わる諸問 題を空間的に把握・分析・予測に有用であり,それによる意思決定や改善シナリオ策定への貢献が 期待されている. 第 11 章 リモートセンシング・地理情報システムの利用 163 図 10 農業生態系空間情報システム GSAS ⑵ 広域的な栽培暦情報を集積・共有するための Web データベース 栽培暦は,農作物の生育を栽培法などと共に時間を追って記録したもので,栽培試験結果のとり まとめ,新しい栽培技術の普及,地域研究の一環など,様々な目的で作成されてきた.しかし栽培 暦は,それぞれの目的に合うように多様な書式で記載され,それぞれの使命を終えた後は保存に も特段の配慮がされてこなかったため,有用な情報が活用されていなかった.そこで,インター ネット技術を利用することによって,栽培暦を広く収集・保存し,利用しやすい形で提供する栽 培暦データベースシステムが構築された(平成 19 年度研究成果情報 , 大野ら).このデータベース では,栽培暦を,作物名や年次などを記載した「概要」,記載事項や書式などを定めた「数値化規 約」 ,および,それに基づいて記される「数値化された暦」に分離して保存する(図11).この方法 により,多様な栽培暦を統一的に記録することが可能である. 栽培暦データベースはインターネット上に構築され(http://dccw.dc.affrc.go.jp),広く一般に公 開されるとともに,インターネットから新たな栽培暦情報を登録する機能をそなえている. ޟⷐޠ ᥲID ⷙ⚂ID ᤨೞ䈱න ‛ ၞ ᐕᰴ 䊶䊶䊶 ᳓Ⓑ 㚅Ꮉ 2003 䊶䊶䊶 (ᐕ䈱ಽഀᢙ) 236 ⒖ᬀ 4 5 ⓠ 6 7 8 9 10 1 36 ޟᢙ୯ൻⷙ⚂ޠ ޟᢙ୯ൻߐࠇߚᥲޠ ⷙ⚂ID 㗄ฬ 㗄ID ᥲID 㗄ID ᤨೞ 1 ⒳䈱㐿ᆎ 1 236 1 12 1 ⒳䈱⚳ੌ 2 236 2 17 1 ⓠ䈱㐿ᆎ 3 236 3 23 1 ⓠ䈱⚳ੌ 4 236 4 29 図 11 栽培暦の記録方式の模式図 栽培暦は,「概要」 , 「数値化規約」 , 「数値化された暦」の 3 つのテーブルに分割して記録される. 164 5.今後の展望 中国黒竜江省における栽培暦と,高頻度衛星センサー AMSR-E 画像から計算される NDPI36 と 呼ばれる指数の変化とを重ね合わせると,NDPI36 の特徴的な変化が移植時期と収穫時期に対応し ていることが分かる(図12) .この結果を利用すると,これまで困難とされていた水稲の移植や収 穫時期の把握が衛星画像で可能となることが期待される. NDVI / LSWI ⒖ᬀ ⒳ 1.0 ಽ⯌ ಽ䈕䈧 Ⓞ ᚑᾫ 0.08 NDVI 0.8 LSWI LSWV 0.6 NDPI36 0.06 0.04 0.4 NDPI36 ⊒⧘ ᩱၭᥲ 0.02 0.2 0.0 0.00 0 60 120 180 ᣣઃ (ㅢ▚ᣣ) 240 300 360 図 12 衛星データから導出された指数と栽培暦の重ね合せ 水分条件に反応する NDPI36 指数は,移植の時期に顕著に増加した後低い値 となり,収穫期より再び増加を示す. ⑶ 歴史的農業環境閲覧システム 第 3 節で説明したように,関東地方を対象とした「迅速測図」は土地利用の地目ごとに彩色さ れており,樹種等も書かれているので,120 年前の土地利用や景観を知るための貴重な資料とな る.しかし,迅速測図は全部で約 900 枚の紙に印刷されており,その広域的な地図情報の有効利 用は困難であった.そこで,この関東地方を対象とする迅速測図をインターネット上に公開する ための「歴史的農業環境閲覧システム」 (Historical Agro-Environmental Browsing System,略称 : HABS)が構築された(平成 20 年度研究成果情報 , 岩崎ら)(岩崎ら , 2009).まず約 900 枚の地図 をデジタル化し,一枚の大きな画像に合成するとともに,GIS ソフトウェアを用いて位置情報を与 え,電子的地図が作成された.インターネットでの公開には,無料で入手でき,改良が可能なオー プンソース GIS ソフトが使用されている(図13) . このシステムは,過去の土地利用と現在の生物多様性との関係の研究や,農耕地の特性評価の基 礎的資料としての活用が期待されている. 5.今後の展望 これまで,日本の農業現場でのリモートセンシングの応用に関しては, ・空間解像度(日本の小規模な農地を観測できない) ・利用可能な波長帯(限られた数の波長帯しか観測できない) ・観測頻度(年に数回の観測のため雲の影響を受ける) など様々な要因により,実践的な利用が進んでいるとは言えなかった.また,外国の商業衛星に よる画像が高価なことも普及の妨げになっていた.農業環境技術研究所では,これらの問題に対し ても,利用できる最大限の資源を使用し新しい技術を開発してきた.たとえば,空間解像度は低い ものの高頻度で観測され無償でデータを利用できる MODIS 画像を利用した生育ステージの推定手 法や,雲を透過するマイクロ波を利用した水田作付け地の推定技術などを紹介した. 第 11 章 リモートセンシング・地理情報システムの利用 165 図 13 歴史的農業環境閲覧システム(茨城県土浦市周辺) 現在の道路と水涯線を,それぞれ赤線と水色線で表示. しかし近年,衛星に搭載される観測センサーの高機能化が急速に進んでいる.また,画像を比 較的安価に利用できる国産衛星の打ち上げも行われている.2006 年に打ち上げられた国産衛星 「ALOS(だいち) 」は,地上解像度 2.5m のパンクロマチックセンサー PRISM や地上解像度 10m の多偏波 SAR センサーを搭載しており,農業環境技術研究所でも,その画像の解析技術の開発が 進行している.なお,現在の ALOS の後継衛星 ALOS-2 の打ち上げも計画されている.今後,農 業・環境問題へのリモートセンシング技術の実践的な利用が加速されると予測される. 農業環境研究における地理情報活用の必要性は,本章において説明したとおりであり,現在,急 速に地理情報システム(GIS)の利用が拡大している.GIS 利用の拡大には,コンピュータのハー ドウェアとソフトウェアの両面での発展の貢献が大きい.まず,ハードウェアに関しては,記憶装 置の大容量化と計算速度の向上により,従来は計算不可能であった広域のデータ処理を実施する ことが可能となっている.また,ソフトウェアの開発に関しては,有償の商業的プログラムのほ かに,フリー(無償)で利用でき,オープンな(プログラムが公開されている)解析プログラム の開発が進んでいる.これらのフリーなプログラムには最新の GIS 解析手法が素早く組み込まれ るため,利用者数は拡大している.また,野外での調査を多用する農業環境研究においては,GPS (Global Positioning System,全地球測位システム)の利用も重要である.今後,農業環境技術研 究所においては,各個別の研究分野において GIS の利用が必須となる. リモートセンシングデータや,地図情報などの空間情報データは,時間的な変化も考慮すると大 量な情報量となる.しかし上述したように,コンピュータのハードウェアの向上により,大量な データも電子化して蓄積することが可能となっている.今後は,これらの空間情報データを,イン ターネットを通じて広く共有し,様々な研究分野で有効に利用していくことが重要である. 166 引用文献 引用文献 1)Inoue, Y., Kurosu, K., Maeno, H., Uratsuka, S., Kozu, T., Dabrowska-Zielinska, K. and Qi, J. (2002)Season-long daily measurements of multi-frequency(Ka, Ku, X, C, and L)and fullpolarization backscatter signatures over paddy-rice field and their relationship with biological variables. Remote Sensing of Environment, 81, 194-204 2)Inoue, Y. and Olioso, A.(2005)Synergy of remote sensing and biophysical process-based modeling for estimating dynamics of CO2 flux in agro-ecosystems. Journal of Agricultural Meteorology, 60, 561-564 3)Inoue, Y., Douangsavanh, L., Qi, J., Olioso, A., Kiyono, Y., Horie, T., Asai, H., Saito, K., Ochiai, Y. and Shiraiwa, T.(2007)Traceability of slash-and-burn land-use history using optical satellite sensor imagery: a basis for chronosequential assessment of ecosystem carbon stock in Laos. International Journal of Remote Sensing, 28, 5641-5647 4)Inoue, Y., Penuelas, J., Miyata, A. and Mano, M.(2008)Normalized difference spectral indices for estimating photosynthetic efficiency and capacity at a canopy scale derived from hyperspectral and CO2 flux measurements in rice. Remote Sensing of Environment, 112, 156-172 5)石塚直樹 , 斎藤元也 , 村上拓彦 , 小川茂男 , 岡本勝男(2003)RADARSAT データによる水稲作 付面積算出手法の開発 . 日本リモートセンシング学会誌 , 23(5),458-472 6)岩崎亘典 , デイビッド S. スプレイグ , 小柳知代 , 古橋大地 , 山本勝利(2009)FOSS4G を用いた 歴史的農業環境閲覧システムの構築.GIS ― 理論と応用,17(1),83-92 7)Sakamoto, T., Yokozawa. M., Toritani, H., Shibayama, M., Ishitsuka, N. and Ohno, H.(2005) A crop phenology detection method using time-series MODIS data. Remote Sensing of Environment, 96, 366-374 8)Sakamoto, T., Nguyen, N. V., Ohno, H., Ishitsuka, N. and Yokozawa, M.(2006)Spatio-temporal distribution of rice phenology and cropping systems in the Mekong Delta with special reference to the seasonal water flow of the Mekong and Bassac rivers. Remote Sensing of Environment, 100, 1-16 9)Shibayama, M., Sakamoto, T., Takada, E., Inoue, A., Morita, K., Takahashi, W. and Kimura, A. (2009)Continuous monitoring of visible and near-infrared band reflectance from a rice paddy for determining nitrogen uptake using digital cameras. Plant Production Science, 12, 293-306 10)Sprague, D., Iwasaki, N. and Takahashi, S.(2007)Measuring rice paddy persistence spanning a century with Japan's oldest topographic maps: georeferencing the Rapid Survey Maps for GIS analysis. International Journal of Geographic Information Science, 21, 83-95 11)スプレイグ・デイビッド , 岩崎亘典(2007)迅速測図に見る農業的土地利用の変遷・歴史 GIS の構築と活用.システム農学 , 23 (1) , 33-40 12)スプレイグ・デイビッド , 岩崎亘典(2009)迅速測図をはじめとする各種地図の GIS 解析によ る茨城県南部の農業的土地利用の時系列変化の研究.ランドスケープ研究,72(5),623-626 13)高田英治 , 井上晃宏 , 芝山道郎 , 坂本利弘 , 守田和弘 , 木村昭彦 , 高橋 渉(2009)定点カメラ 画像による水稲の生育状況の推定.システム農学 , 25(1),27-34 167 第 12 章 農業環境のモニタリングと手法の開発 1.物理環境・ガスフラックス変動のモニタリング ⑴ はじめに 本節では,農業環境のモニタリングのなかで,ガス以外の大気質の放出・拡散過程に関する研究 として花粉と黄砂,地球環境変化に関わる物理環境のモニタリングとして,農業水資源に関わる降 雪水量の観測手法と,温暖化影響の早期出現が予想されるチベット高原での気象のモニタリング, ガスフラックス変動のモニタリングとして,農耕地に関わりの深い二酸化炭素とメタンのフラック スの長期観測研究を紹介する.このうち,花粉に関わる研究は,農業環境技術研究所では大気生物 学(Aerobiology)分野の重要課題として,1990 年代から研究が実施されてきた.2000 年代に入っ ほじょう てからは,遺伝子組換え植物の圃場試験における花粉の飛散に関連する研究や,花粉症が社会問題 化したスギ花粉に関する研究などが行われた.ガスフラックスのモニタリングは,1990 年代に行 われた微気象学の原理に基づく大気微量ガスのフラックスの測定法に関する研究の成果を受けて, 2000 年代には農耕地や自然生態系でモニタリングが展開された.なかでも,茨城県つくば市真瀬 の水田では,10 年以上にわたる長期観測が現在でも継続されている.この研究は,ガスフラック スモニタリング手法の高度化に関する研究も含めて,陸域生態系の炭素・水循環の解明を目的とし た国際観測ネットワーク(FLUXNET)や,その地域ネットワークである AsiaFlux との連携のも とで行われた. ⑵ 大気質(花粉,黄砂)のモニタリング 1 トウモロコシ花粉飛散量自動モニター装置の開発 遺伝子組換え植物の花粉の飛散による同種間交雑の問題に対処するには,作物花粉の拡散動態 を把握し,気象条件等との関係を明らかにする ことが重要である.ダーラム(Durham)法な Cశቇ♽ ᄢ᳇ どの既存の花粉計測法は,顕微鏡での花粉計数 ೨ᣇᢔੂᬌེ శ᧤ࡦ࠭ ⓨ᳇ᵹ〝 ඨዉࠩ ᢛᵹ▤ に多大の労力を必要とするうえに,日単位での 計測しかできない.そこで,代表的な風媒花植 物であり,花粉による交雑が問題となっている (平面図) トウモロコシについて,大粒径花粉の光学的お (立面図) よび空気力学的特性を考慮して,花粉飛散量を 簡易に,かつ連続的に測定できる花粉モニター 装置を開発した(Kawashima et al., 2005) .こ の装置では,スギ花粉のおよそ 3 倍の粒径,27 倍の重さに相当するトウモロコシ花粉の光学特 性と運動特性にあわせて,既存の粒子カウン ターの光学系および気流系を改良することによ D᳇ᵹ♽ ⺞శࠪࡦ࠳ࡦ࠭ ᣇᢔੂᬌེ ᄢ᳇ๆᒁญ ガラス製ロート ⓨ᳇ឃญ ᵹ㊂⸘ ⍾Ⴒỹㆊⵝ⟎ ࡃ࡞ࡉ ࡈࠖ࡞࠲ 毎分 4.1 φᓴⅣ᳇ᵹ శቇ♽ ←エアージャケット方式 ポンプ り,花粉を効率よく計測部に導き,該当粒径の 球状粒子だけを検出することを可能にした(図 1) .この装置で計測した花粉飛散量の日積算値 は,空中花粉の基準的測定法として広く用いら ࡈࠖ࡞࠲ ✭ⴣ࠲ࡦࠢ 図 1 トウモロコシ花粉モニター.破線内が既存の パーティクルカウンターに改良をくわえた部分. 168 1.物理環境・ガスフラックス変動のモニタリング れているダーラム法による花粉計測値との相関がきわめて高かった.また,計測の時間分解能が向 上し,午前中にピークがあり,正午にはピーク濃度の半分以下に減少するという花粉飛散の日変動 を計測することができた(平成 15 年度研究成果情報,川島ら). 2 積雪地域におけるスギ花粉の飛散開始日の推定 スギ花粉の飛散開始日は,花粉症予防薬の投 与開始時期に関係する重要な情報であり,1990 140 年代からさまざまな研究が行われてきた(川 120 2.5 㪉㒠㔐ᷓ 䉴䉩⧎☳㘧ᢔ㐿ᆎផቯᣣ 㪈䋬㪉ᐔဋ᳇᷷ 2 が,雪を考慮した飛散開始日の推定手法はな かった.そこで,月平均気温と月降雪深を用い る統計的手法によって,飛散開始日を推定する 手法を開発した(Inoue et al., 2002) .1983 年 から 1998 年までの飛散開始日と 1,2 月平均気 温および 2 月の降雪深を用いて,山形市におけ るスギ花粉の飛散開始日を推定する重回帰式を 決定した.積雪の影響を表す変数としては,積 雪深(地表面に堆積した雪の深さ)ではなく, 100 1.5 80 1 60 0.5 40 0 20 -0.5 0 㪈䋬㪉ᐔဋ᳇᷷䋨㷄䋩 散開始日の推定精度が悪いことが知られている 㪉㒠㔐ᷓ㩿㪺㫄㪀㪃 䉴䉩⧎☳㘧ᢔ㐿ᆎᣣ㩿㪛㪦㪰㪀 島・高橋,2002) .積雪地域では,多雪年に飛 -1 1981-2000 2031-2050 ⛔⸘ᦼ㑆㩿ᐕ䋩 2081-2100 図 2 山形市におけるスギ花粉の飛散開始推定日の 気候学的推移. 樹体への着雪量との関係が深い月降雪深(1日 に降り積もった雪の深さの月積算値)を用いた.この結果,1,2 月平均気温のみを用いた従来法 よりも飛散開始日の推定精度が向上し,5 日以内の誤差で推定が可能になった.また,気象庁気候 統一シナリオ第 2 版(排出シナリオ SRES−A2(多元化社会))から農環研が作成したメッシュ気 候値を用いて,将来の山形市でのスギ花粉の飛散開始日を予測したところ,平均気温の上昇と降雪 量の減少により,2081 〜 2100 年には飛散開始日が現在よりも 3 週間早まると予想された(図2). この飛散開始日の推定法は,スギ花粉の飛散開始日の統計があれば,他の積雪地域でも活用でき る.気温上昇,降雪深の減少は他の地域でも共通して予測されていることから,山形市と同様に飛 散開始日の早まりが予想される(平成 17 年度研究成果情報,井上ら). 3 風送ダストの発生機構 黄砂,すなわちアジア大陸の乾燥・半乾燥地域から風によって大気中に舞い上がる風送ダスト は,発生域の農業生産や生活環境に影響を与えるばかりでなく,自由大気に鉱物質エアロゾルとし て浮遊し,日射の散乱・吸収および赤外放射の吸収過程や雲・降水過程を通じて,グローバルな気 象・気候に影響を及ぼしていると考えられる.わが国では,中国や韓国ほど深刻な経済的被害や健 康被害は報告されていないが,今後,発生源地域の乾燥化の進行によっては直接的被害が顕在化す る可能性がある.農業環境技術研究所は,日中科学技術協定に基づく日中共同研究プロジェクトに 参画し,地表面状態が風送ダストに及ぼす影響を明らかにするため,黄砂の主要な発生域・通過経 れき 路にあたる甘粛省敦煌において,ゴビ(礫沙漠)とオアシス農地でダストの比較観測を実施した. かん がい オアシス農地には防風林があるため,ゴビに比べて風速が 40%低く,しかも潅漑により土壌水分 も高く保たれている.しかし,表層に 5μm 以下の細かな粒子が多く流動性が高いため,ダストが 舞い上がる臨界風速は,細砂(72 〜 450μm)も含めて,オアシス農地の方がゴビに比べて小さい こううん ことがわかった(杜ら,2003) .また,春季のオアシス農地の耕耘は臨界風速を低下させるととも に,同じ風速でもダストの発生量を増加させることが明らかになった(Du et al., 2005). 第 12 章 農業環境のモニタリングと手法の開発 169 ⑶ 地球環境変化に関わる物理環境のモニタリング 1 積雪地帯厳冬期における新型降水量計の動作特性 地球温暖化の進行により,積雪水資源量の変動が予想されている.積雪地帯の水資源のモニタ リングのためには降雪水量(降雪を水に換算した量)の計測が重要である.しかし,従来の降水 量計は,測器の周囲の風の乱れによる雪の捕捉損失が大きく実測値が過小となる,降水継続時間 が正確に計測できない,冬季も含めて定期的な保守が不可欠なため河川源流域等の無人地では観 測できない,という問題がある.近年開発された,レーザー光やマイクロ波を用いて大気中の降 水粒子濃度を非破壊で遠隔計測する降水量計は,これらの問題を回避できる可能性がある.そこ で,北海道芽室町でこれらの新型降水量計 2 機種を含む 4 種類の降水量計の比較試験を行った. 平均気温−10℃,最低気温−27℃の低温条件下で約 2 か月間の比較を行った結果から,レーザー 光を用いて大気を直接計測する降水量計は,補正なしでも日降水量を推定誤差 0.8mm 以下という 高い精度で測定でき,また弱い降水にも反応するため,降水継続時間も従来の手法より正確に実 測できることが明らかになった(Inoue et al., 2009) .非降雪期間を含む通年観測でも,半年に 1 回,センサー部の動作確認と清掃を行うだけで,良好に動作することを確認した.このように, レーザー光を用いる新型降水量計は降雪水量の評価に有効なことが示され,積雪地帯の降水量観 測網の整備に役立つ知見が得られた(平成 20 年度研究成果情報,井上ら) . 2 チベット高原における草線の気候条件 北極,南極に次ぐ地球の第三極と呼ばれるチベット高原では,温暖化に対する生態系の影響が早 期にかつ急激に現れる可能性が指摘されており,畜産に対する温暖化の影響がすでに報告されてい る(Du, et al., 2004).しかし,チベット高原の 34%の面積を占める標高 4800m 以上の地域では,気 象観測と生態調査はほとんど実施されておらず,生態系への温暖化影響の実態は未解明である.そ こで,チベット高原内の二つの地点で,山ろくの高原部から山頂に至るトランゼクト気象観測を実 施した.両観測地点には草線(草本類による被度がほぼ 100%の限界線.さらに標高が高い場所でも 植生は見られるが,被度が急激に低下する)がみられる.両観測地点の草線標高は 1000m もの差が あるが,5 年間のモニタリングの結果から,7 〜 8 月の平均気温が約 5℃,日平均気温 5℃以上の日数 が約 40 日など,両地点の草線高度における夏季の気候条件の類似性が明らかになった.観測地点の 一つである当雄の山ろくにある気象官署の過去の観測値と,草線付近で観測した気象データから, 現在の草線高度における過去の気温の変動を推定した結果,過去 47 年間で 7 月の平均気温は 0.8℃上 昇し,日平均気温 5℃以上の日数は 15 日以上も増加したと推定された.この気温上昇は 100m 以上 の標高差に相当し,この期間に草線の移動をもたらした可能性が示唆された.この結果の直接的な 裏付けとなる画像やリモートセンシングデータなどは,まだ得られていないが,モニタリングによ る温暖化影響の検知につながる知見といえる(平成 22 年度成果情報,杜ら)(杜ら,2009). ⑷ 温室効果ガスフラックスのモニタリング 1 アラスカの湿地ツンドラの二酸化炭素とメタンフラックス 地球規模での炭素循環や温室効果ガス収支における陸域生態系の役割を評価するためには,世界 の土壌炭素の 3 分の 1 を蓄積しているとされる高緯度湿地帯の炭素収支の解明が重要である.なか でも,北極域に広く分布するツンドラは地球温暖化に対する脆弱性が指摘されているが,温室効 果ガス収支の現状やその年次変動を評価するために必要な観測データは少なく,特に北極域の代 表的なツンドラである湿地ツンドラで,二酸化炭素とメタンのフラックスを同時に通年観測した 例はなかった.そこで,日米科学技術協定(日米地球環境リエゾン会合)に基づく共同研究とし て,アメリカ合衆国アラスカ州バロー(Barrow)の北極海沿岸の湿地ツンドラで長期連続観測を 170 1.物理環境・ガスフラックス変動のモニタリング 行った.3 年間の観測の結果から,夏季 4 か月 間の二酸化炭素の吸収量は冬季 8 か月間の放出 量を上回り,湿地ツンドラは年間収支としては 二酸化炭素を吸収していること,夏季に湛水状 態となるため,他の高緯度地域の湿地に比べて 純生産量が大きいこと,地球温暖化係数を用い て換算すると,メタンの年間発生量は二酸化炭 素の年間吸収量に比べて少なく,現在の湿地ツ ンドラは地球温暖化の抑制に寄与していること がわかった.また,両ガスのフラックスの年次 変動は大きく,1999 年のように夏季に多照, 高温で総光合成量が多い年には,純生産量は平 年の約 2 倍となり,メタン発生量も増加した. 3 年間にわたる観測により,二酸化炭素やメタ 図 3 米国アラスカ州バローの湿地ツンドラにおけ る二酸化炭素収支とメタン発生量 ンの年間収支や,気象の年次変動にともなう変 動幅が明らかになった(平成 14 年度研究成果情報,宮田ら)(間野ら,2003; Harazono et al., 2003; Harazono et al., 2006) . 2 イネ単作田の二酸化炭素フラックスのモニタリングと炭素収支 茨城県つくば市真瀬のイネ単作田(図4)でも, 1999 年以降,二酸化炭素とメタンのフラックスのモ ニタリングが行われている(Saito et al., 2005; Han et al., 2007) .農耕地ではガス態での炭素移動に加えて, 収穫,作物残さの処理や有機質資材の投入などの圃場 管理も炭素移動をともなう.ガスフラックスに非ガス 態での炭素移動量を加えて,イネ単作田の年間炭素収 支を定量化した.その結果,単作田の炭素収支の主要 項は,植物の光合成・呼吸および土壌有機物分解にと もなう水田 ― 大気間の二酸化炭素の交換と,収穫・ 作物残さ処理であり,作付期間の純生産量(NEP) のおよそ半分が収穫時に持ち出され,残りの半分 に相当する炭素量が休耕期間に有機物分解により 大気中に放出されることがわかった(図5) .この 結果,年間炭素収支は 8 年間の平均でほぼ均衡し 図 4 茨城県つくば市真瀬のガスフラックス モニタリングサイト +4.3±0.5 (ઃᦼ㑆䈱NEP) +8.7±0.4 శวᚑ -4.4±0.3 ๆ ಽ⸃ -0.1 䊜䉺䊮 ていた.また,炭素収支の年次間変動の主な要因 は,作付期間の NEP と不定期に行われた有機質 資材投入量が多かった年は,単年の炭素収支が 流入となった(平成 22 年度研究成果情報,宮田 ら) .以上の結果は既往の研究と大きく異なるも のではないが,二酸化炭素フラックスの標準的 -0.3±0.4 ‛ᱷᷲළ -2.2±0.1 ಽ⸃䋨ભ⠹ᦼ㑆䈱NEP䋩 +2.6±0.5 ᩮㇱ䊶‛ᱷ ᷲ㍁ㄟ䉂 資材(堆肥)の投入であり,7 月が多照で光合成 による二酸化炭素吸収量が多かった年や,有機質 න䋺t C ha-1 y-1 -2.3±0.2 ⓠ‛ᜬ䈤䈚 +0.3±0.5 ᯏ⾰⾗᧚ ᵹ -0.1 ઃᦼ㑆 ભ⠹ᦼ㑆 図 5 イネ単作田の年間炭素収支構造 真瀬観測点の 2002 〜 2009 年の平均値±標 準偏差(t C ha−1 y−1)で,正値は水田への 流入を表す. 第 12 章 農業環境のモニタリングと手法の開発 171 測定法とされる渦相関法によるモニタリング結果に基づいて,炭素収支を定量化した点に意義があ る. 真瀬観測点の観測データは,休耕期間を含む通年の二酸化炭素フラックスや熱収支諸項,気象要 素の 30 分値が,一定の基準による品質管理を施され,農業環境技術研究所フラックス・微気象観 測データベース(Ecosystem Database) (http://ecomdb.niaes.affrc.go.jp/)で公開されている(平 成 15 年度研究成果情報,宮田) .また,AsiaFlux や FLUXNET 等の国際的な研究ネットワークの データベースにも登録されており,サイト間比較,モデル構築,リモートセンシングの地上検証等 に活用されている.真瀬観測点は,2011 年には日本生態系長期研究ネットワーク(JaLTER)の準 サイトとして登録され,今後も分野間連携の拠点としての役割を果たすことが期待されている. ⑸ ガスフラックスモニタリング手法の高度化 1 オープンパス型とクローズドパス型渦相関法の比較 渦相関法は,二酸化炭素フラックスのモニタリングに標準的に用いられる,代表的な微気象学的 ガスフラックス測定法(微気象学的手法)である.渦相関法は,ガス分析計のタイプにより,オー プンパス型とクローズドパス型に分けられる.この二種類の渦相関法にはそれぞれ長所と短所があ り,世界中に両方の観測システムが混在しているのが現状である.観測データの標準化のために は,二種類の渦相関法による観測値の系統差の有無を明らかにする必要がある.このため,両者を 併用している真瀬観測点のデータを利用した研究が行われた.まず,水田の休耕期間(地表面は裸 地)や積雪期間の森林のように植物の光合成がない期間に,オープンパス型渦相関法を用いて計測 された二酸化炭素フラックスに不自然な吸収フラックスがみられることがある.その原因の究明の ため,真瀬サイトのデータを詳細に解析した結果,超音波風速温度計で計測された温度変動を用い る一般的なオープンパス型渦相関法では,赤外線分析計の測定光路上に生ずる温度変動の影響を正 しく補正(密度変動補正)できず,水田の休耕期間や積雪期間の森林のように,二酸化炭素フラッ クスが小さく顕熱フラックスが大きい条件では,オープンパス型渦相関法の精度が低下し,場合に よっては不自然な吸収フラックスが計測されることが明らかになった(Ono et al., 2008).次に, 両システムを併用している真瀬観測点と苫小牧観測点(森林)のデータを用いて,二種類の渦相関 法による NEP を比較し,オープンパス型はクローズドパス型に比べて年間の NEP を 10%(真瀬) 〜 16%(苫小牧) ,大きく評価することを明らかにした.両手法間の系統差は密度変動補正の大き さとともに増加するため,両観測点の熱収支構造の違いにより,密度変動補正の大きい苫小牧観測 点の方が,真瀬観測点よりも系統差が大きいことがわかった(小野ら,2007). 2 渦相関法のデータ処理ソフトウェアの開発と公開 近年,微気象学の専門家だけでなく,その周辺分野でも渦相関法の利用者が増加しつつある. 渦相関法による観測で得られる乱流生データの統計処理や,計算値に含まれる異常値(降雨等の観 測に適さない気象条件や,人為的な影響を受けた観測値)の判別には,接地境界層の乱流輸送に関 する専門知識が必要とされるが,公開されている処理プログラムは少ない.この問題は渦相関法の 普及の障害になっており,観測データの標準化を妨げる要因ともなっている.そこで,最新の研究 成果を取り入れた,渦相関法による観測データの処理プログラムを,専門外の者でも利用可能なプ ログラムとして公開,提供した.処理すべき乱流生データのほかに,処理法の選択や測器に関する 情報が入力されたファイルを指定のフォーマットに従って準備し,プログラムを実行することによ り,乱流生データの各種統計量,品質検査情報,補正適用前後のフラックス値,偶然誤差などが得 られる.利用者は,品質検査情報を参照することにより,フラックス値の信頼性や誤差の程度を判 断でき,異常値を容易に除去して実用的なフラックスのデータセットを作成できる(間野ら,2007) 172 引用文献 (平成 16 年度研究成果情報,間野ら).このプログラムは国内の研究機関や大学等に提供され,渦 相関法によるガスフラックスのモニタリングに活用されている.さらに,最新の知見に基づいて改 良したプログラムを,Web 上で実行できるシステムとして公開した.Web 公開することにより, 将来のプログラムの追加・修正も,利用者に迅速に周知することが可能となった.近年利用者が増 加しているアジア各国を中心に幅広い普及が期待される(平成 22 年度研究成果情報,間野ら). 3 農地の蒸発散量と二酸化炭素吸収量のオンライン監視用ソフトウェア 環境の変化にともなって時々刻々変化する農地の蒸発散量や二酸化炭素吸収量の情報をオンライ ンで収集できれば,潅漑水の管理や作物生育の監視などの適切な圃場管理に活用できる.渦相関法 は圃場規模での蒸発散量や二酸化炭素吸収量を計測するのに適した技術であるが,圃場管理に適用 するためには,回収データの事後処理として行っていた観測値の精度評価や異常値の判別,欠損 データの補完などの煩雑な作業を,オンラインで処理する必要があった.そこで,乱流データの誤 差解析の結果(Kim et al., 2009)を利用して,品質判定と欠測値補完を簡易・迅速化し,蒸発散量 と二酸化炭素吸収量のオンライン監視用のソフトウェア(FluxPro)を開発した(平成 21 年度研 究成果情報,金ら) .さらに,観測データを用いて生態系のガス交換モデルのパラメータを推定す るデータ同化機能を追加した.これにより,迅速なフラックスの算定と短期予測が可能になり,渦 相関法を用いた潅漑水管理や作物生育の監視が,一歩,実用化に近づいた.現在,水田および樹園 地で本システムの実用化に向けたテストが行われている. ⑹ 今後の展望 微気象学的手法による二酸化炭素やメタンなどのガスフラックスの計測手法は,その長期観測へ の適用も含めて,おおむね完成の域に近づいており,同時にその限界も明らかになりつつある.今 後は,この計測手法や,それによって得られた詳細かつ膨大なデータを,モニタリングを含めてど んな研究に利用していくのかという戦略が重要となる.生態系のモデル研究やリモートセンシング と連携したガスフラックスの広域評価は,その一つの方向だろう.本節では紹介しなかったが,ア ンモニアやオゾン,エアロゾルなどの粒子状物質のフラックスのモニタリングも実施されている. 将来,ガス分析計の制約が解決できれば,同位体を含むさまざまな大気微量ガスのフラックスの計 測への適用も考えられる.なお,ガスを含めた大気質の研究は,地表面でのフラックスの計測だけ ではなく,花粉や黄砂にみられるように,大気中での拡散過程の研究も重要であることを忘れては ならない. 物理環境やガスフラックスに限らず,モニタリングは長期にわたる研究となるので,研究者自身 がモニタリングの実務を担当せざるを得ないわが国では,計測の多くが自動化されたとしても,担 当者の負担は決して軽くはない.また,気象官署での観測とは異なり,モニタリングを継続するこ とに十分な社会的承認が得られているとは言えない状況で,モニタリングを研究として継続するこ とは,じつは容易ではない.研究としてのモニタリングの継続のためには,研究者が所属する組織 としての支援の在り方も十分に検討する必要がある. 引用文献 1)杜 明遠,米村正一郎,真木太一,山田 豊,沈 志宝,申 彦波,汪 万福,川島茂人, 井上 聡(2003)中国敦煌のオアシスにおけるダストの舞い上がりの特徴.地球環境 , 7(2), 187-195 2)Du, M., Kawashima, S., Yonemura, S., Zhang, X. and Chen, S.(2004)Mutual influence between the human activities and the climatic change in Tibetan Plateau during recent years. Global 第 12 章 農業環境のモニタリングと手法の開発 173 and Planetary Change, 41 (3-4) , 241-249 3)Du, M., Yonemura, S., Shen, Z., Shen, Y., Wang, W., Yamada, Y., Maki, T., Kawashima, S. and Inoue, S.(2005)Tillage effects on Aeolian dust emission in bare agricultural fields at Dunhuang, China. Journal of Agricultural Meteorology, 60(5),503-506 4)杜 明遠,米村正一郎,張 憲州,李 英年,趙 亮,張 法偉,劉 景時,唐 艶鴻,川島 茂人(2009)チベット高原における高山草原生存極限(草線)の気候とその変動.日本農業気 象学会 2009 年全国大会講演要旨 , 37 5)Han, G.H., Yoshikoshi, H., Nagai, H., Yamada, T., Ono, K., Mano, M. and Miyata, A.(2007) Isotopic disequilibrium between carbon assimilated and respired in a rice paddy as influenced by methanogenesis from CO2. Journal of Geophysical Research, 112, G02016, doi:10.1029/ 2006JG000219 6)Harazono, Y., Mano, M., Miyata, A., Zulueta, R.C. and Oechel, W.C.(2003)Inter-annual carbon dioxide uptake of a wet-sedge tundra ecosystem in the Arctic. Tellus, 55B, 215-231 7)Harazono, Y., Mano, M., Miyata, A., Yoshimoto, M., Zulueta, R.C., Vourlitis, G.L., Kwon, H. and Oechel, W.C.(2006)Temporal and spatial differences of methane flux at arctic tundra in Alaska. Memoirs of National Institute of Polar Research, Spec. Issue, 59, 79-95 8)Inoue, S., Kawashima, S. and Takahashi, Y.(2002)Estimating the beginning day of Japanese cedar pollen release under global climate change. Global Change Biology, 8, 1165-1168 9)Inoue, S., Hirota, T., Iwata, Y., Suzuki, K. and Nemoto, M.(2009)Comparison of four instruments for measuring solid precipitation below the freezing point condition. Journal of Agricultural Meteorology, 65 (1) , 77-82 10)川島茂人,高橋裕一(2002)スギ花粉の放出と拡散過程に関する研究.天気,49(11),873-885 11)Kawashima, S., Fujita, T., Matsuo, K., Shibaike, H., Du, M., Inoue, S. and Yonemura, S.(2005) Variation feature of airborne corn pollen number obtained by the automatic pollen monitor. Journal of Agricultural Meteorology, 60 (5) , 1033-1036 12)Kim, W., Komori, D. and Cho, J.(2009)The characteristic of relative error in eddy covariance measurements and its application to data quality control in rainfed paddy field. Journal of Agricultural Meteorology, 65 (2) , 201-207 13)間野正美,原薗芳信,宮田 明,Zulueta, R.C. and Oechel, W.C. (2003) Alaska 州 Barrow の湛水ツンドラにおける 2000 年度植物生育期間の CO2 フラックスの季節変化と CO2 収支.農 業気象,59 (2) , 141-154 14)間野正美 , 宮田 明 , 安田幸生 , 永井秀幸 , 山田智康 , 小野圭介 , 齊藤 誠 , 小林義和(2007)オー プンパス型渦相関法データの品質管理.農業気象,63(3),125-138 15)小野圭介 , 平田竜一 , 間野正美 , 宮田 明 , 三枝信子 , 井上吉雄(2007)オープンパス型とクロー ズドパス型の渦相関法による CO2 フラックスの系統的差異と密度変動補正の影響.農業気象 , 63 (3) , 139-155 16)Ono, K., Miyata, A. and Yamada, T.(2008)Apparent downward CO2 flux observed with openpath eddy covariance over a non-vegetated surface. Theoretical and Applied Climatology, 92, 195-208 17)Saito, M., Miyata, A., Nagai, H. and Yamada, T.(2005)Seasonal variation of carbon dioxide exchange in rice paddy field in Japan. Agricultural and Forest Meteorology, 135, 93-109 174 2.農業環境中の放射性物質のモニタリング ⑴ 人工放射性核種の長期モニタリングと農業環境中の動態解析 1 はじめに 近年,大気中の二酸化炭素の増加抑制,石油資源の節約,エネルギーセキュリティー対策等の観 点から原子力利用が見直され,原子力開発計画や発電所等の建設が世界各国で増加傾向にある.特 に,わが国周辺の後発の東アジアの国々においても原子力開発が大幅に推進されようとしている. また,原子力発電に用いるウランの資源量の不足を補い核燃料廃棄物のプルトニウムの再利用を図 るために,わが国においても試験用高速増殖炉「もんじゅ」の運転や青森県六ヶ所村における大規 模核燃料再処理施設の試験稼働が進められようとしており,国民の間に賛否両論を巻き起こしてい る.このように,原子力利用が拡大する世界的趨勢にあって,エネルギー供給面では多くのメリッ トが期待される反面,原子力関係施設の事故等にともなって放射能による環境汚染が起こるリスク は今後高まることが懸念される. 一方,1960 年代まで盛んに行われていた核実験は,現在に近づくにつれ急激に影を潜め,新た な人工放射性核種による環境汚染は減少している.しかし,世界的な政治情勢から見て核拡散に歯 止めがかかるかどうかは疑問で,我々は依然として核の脅威にさらされていると言える.また,国 際放射線防護委員会(ICRP)等により絶えず見直しが図られている被ばくリスク評価について, 人間活動によって高められた天然放射性核種(NORM)の寄与が最近注目されており,農業資材 中の 40K やウラン系列核種も議論の対象となっている. このような状況の中で,農産物の安全を確保し国民を放射能汚染の不安から守るために,環境中 の放射性核種のレベルについて常時,監視をしておくことは大変重要である.また,核実験や原子 力発電所の事故等により突発的に周辺環境が放射能に汚染された場合,監視体制を整えておけば, いち早く対応することもできる. 以上の基本的認識の基に,農業環境技術研究所は独立行政法人農業・食品産業技術総合研究機 構畜産草地研究所と連携協力して昭和 30 年代より文部科学省放射能調査研究事業に参画し,農作 物・農林地及び牛乳を対象とする継続的なモニタリングを実施することにより,わが国の環境及び 農産物からの人工放射性核種による被ばくリスク評価に資する基礎データを提供してきた. また,放射能の農業環境への影響を予測するには,放射能モニタリングによるバックグラウンド データの整備に加え,土壌から作物,あるいは飼料から畜産物への放射性核種の吸収・移行過程と それを支配する要因を解明して農畜産物を取り巻く農業環境中における放射性核種の動態を把握す る必要があり,現在,そのための研究も様々な課題を設定して実施している. 2 90 Sr と 137 Cs の長期モニタリングとデータのインターネット上への公開 農業環境技術研究所では,1959 年から現在に至るまで農林水産省傘下の独立行政法人や農業関 係試験研究機関に設置した全国十数カ所の定点観測地点から毎年作物体・土壌試料の提供を受け, ストロンチウム 90(90Sr)とセシウム 137(137Cs)の分析を行っている.提供を受けている試料は, 水田圃場あるいは畑圃場の作土,およびそれらの圃場で栽培したコメ(白米,玄米),小麦(玄麦) であるが,その他に農業環境技術研究所内で野菜(ほうれんそう等)も栽培し放射性核種のレベル を調査している. そ の 中 で, 玄 麦 中 の 137Cs の レ ベ ル は, 大 気 圏 核 実 験 が 盛 ん に 行 わ れ て い た 1963 年 に 43,600mBq/kg と最大値を記録したが,その後急激に減少し 1985 年には 45mBq/kg まで低下した. しかし,1986 年のチェルノブイリ原子炉事故により,その年度のみ測定値が一気に 5,690 mBq/kg 第 12 章 農業環境のモニタリングと手法の開発 175 䉼䉢䊦䊉䊑䉟䊥 ⢻䋨㫄㪙㫈㪆㫂㪾䋩 100000 Ⓝ㘃 10000 1000 䂓䃂 100 10 137 ⊕☨䇭 Cs ⊕☨䇭 90 Sr ₵㤈䇭 137 Cs ₵㤈䇭 90 Sr 1 ᣣᧄᶏ 䂓 䂓䃂 䂓 0.1 䂓 䃂 ფ 10000 䂓 ᳓↰ 䃂 ⇌ 䂓 ⢻䋨㪤㪙㫈㪆㫂㫄2䋩 䂓 䂓䃂 䃂 䂓䃂 䂓 䂓 1000 䂓䃂 䂓 䂓 ᄥᐔᵗ ᳓↰䇭137 Cs ᳓↰䇭90 Sr ⇌䇭䇭137 Cs ⇌䇭䇭90 Sr 100 10 1960 1970 䋨⺋Ꮕ▸࿐䈲䈱䉂␜䇯䋩 1980 1990 2000 ᐕᐲ 図 1 水田と畑での作物(白米,玄麦)と土壌の放射性同位体濃度の推移(左)と調査地点(右) まで上昇した.そして,その後また減衰する傾向がもどり,2009 年度には 13±19mBq/kg となっ ている.一方,玄麦中の 90Sr レベルは,1963 年に最大値 12,300mBq/kg を記録したが,チェルノ ブイリ原子炉事故の影響は見られず,徐々に低下して行く傾向が現在まで続いて,2009 年度には 61±34mBq/kg であった.この 1960 年代からの 137Cs と 90Sr の経時的推移と近年の低レベルの状 態は,コメに関しても 1986 年の 137Cs を除いてほぼ同様の傾向である. これらの作物と栽培土壌に関する 90Sr と 137Cs の分析値については,2009 年からインターネット 上に公開し,一般にも閲覧できるようになっている(平成 21 年度研究成果情報,木方ら).公開し ている項目は, ・白米,玄米,玄麦および小麦粉中の 90Sr,137Cs 濃度(1959 〜 2006 年):単位 mBq/kg ・水田作土および畑作土中の全 90Sr と置換態 90Sr 並びに全 137Cs と置換態 137Cs 濃度(1959 〜 2006 年) :単位 Bq/kg ・採取地点の土壌群名と作土の土性 などである.整理したデータのデータベース化は,パッケージソフトウェア "V/GAI−AGRIP (ヴィジブルインフォメーションセンター社) " を用いている.これらのデータは,2009 年 12 月 から Internet Explorer(Microsoft 社)や Mozilla Firefox(Mozilla Foundation)などの標準的な Web プラウザを使用して,http://psv92.niaes3.affrc.go.jp/vgai_agrip/ で,閲覧可能となっている. さらに,土壌中の 90Sr と 137Cs のデータで現在公開しているのは,作土の分析値のみであるが, これらの核種は放射壊変だけでなく雨水等による下層土への浸透によっても分布が変化する.その 土壌下層への浸透移動は土壌の性質や土地利用の違いによって異なるが,これまで定点調査圃場で は圃場周辺の未耕地土壌を対比して深さ別の土壌のサンプリングと分析も行っており,放射性核種 の深度分布の調査データも保有しているので,今後,それらをデータベース化し,Web 公開をす る予定にしている. 3 北朝鮮核実験に対する放射能緊急時調査 2009 年 5 月 25 日 9 時 54 分(推定) ,北朝鮮で地下核実験が行われた.農業環境技術研究所では, 同日から所内環境放射能観測圃場で栽培しているホウレンソウのサンプリングの他,水盤による大 176 2.農業環境中の放射性物質のモニタリング 気降下物サンプリング,大気浮遊塵サンプリング,空間線量率連続測定を開始した.ホウレンソウ のサンプリングは 6 月 1 日まで毎日,大気降下物・浮遊塵調査は 5 月 28 日まで実施したが,いず れも異常値は検出されず,周辺環境と農作物への影響のないことが確認された. このように,農業環境技術研究所では,不測の事態によりわが国の農業環境の放射能汚染の可能 性が高まった場合には,各種の監視プログラムを緊急に実施して農業環境への影響を調査している. 4 農作物可食部中のウラン濃度の調査 天然物に含まれる放射性物質の危険性の再評価も人工放射性核種とともに国際的に行われている が,ウランも対象の一つとなっている.農業においては,肥料原料に不純物としてウランが含まれ ているので,食の安全の観点から正しく評価をしておく必要がある. そこで,作物,特に可食部のウラン濃度と,ウランが土壌から移行する割合(移行係数)を主要 な農作物について明らかにすることを目的に調査を行った(Yamaguchi et al., 2009)(平成 19 年度 研究成果情報,木方ら) .日本各地から集めた主要穀類の可食部中ウラン濃度を測定したところ, 0.1μg/kgDW 以下と極めて低い値であり,作物可食部中のウラン濃度 / 土壌作土中のウラン濃度 の移行係数も 1/20,000 以下となった.また,約 30 年間畑として施肥をしている農業環境技術研究 所内の黒ボク土圃場では,表層 10cm のウラン濃度が,隣接する未耕地よりも明らかに高くなって いるが,容易に植物に吸収されると考えられる酸可溶態画分は全ウランの 1.6%でしかなかった. この畑圃場で栽培した 34 作目の可食部ウラン濃度およびウラン移行係数を計算したところ,全体 として可食部ウラン濃度で 10μg/kgDW,移行係数で 0.005 を超えるものはなく,ウランは可食部 に移行しにくい元素であることが証明された. また,これらのデータに日本人の平均的作物摂取量(国民栄養調査報告)を乗じて作物のウラン 摂取による内部被ばく線量を概算しところ,植物性食品(果物,キノコ,海藻類を除く)から摂取 されるウランの放射能量は,年間 0.25Bq となり,また,核種による人体への影響度を加味したウ ランの実効線量は年間 0.01μSv と,わが国の農作物に含まれるウランの放射能面からの安全性が 示された. 5 農作物の放射能汚染経路の解析 農作物の放射能汚染経路としては,作物地上 90Sr 部を経由する直接汚染と汚染土壌を経由して根 れらの汚染経路の寄与率を小麦の移行係数(作 物 体 の 90Sr+137Cs 含 量 / 土 壌 の 90Sr+137Cs 含 量)をもとに計算すると,1963 年頃の放射能 ⋥ធᳪᨴ ᳪᨴ⚻〝ഀว䋨䋦䋩 からの吸収による間接汚染が知られている.そ 㑆ធᳪᨴ 多量降下期の両核種と 1986 年のチェルノブイ リ原子炉事故年の 137Cs ではともに直接汚染が 137Cs 大部分を占めていたが,最近では両核種とも新 量が間接汚染に起因していると考えられる(平 成 13 年度研究成果情報,駒村ら) . 1986 年のチェルノブイリ原子炉事故におい ては,前述のように玄麦の 137Cs のピークが観 測されたが,その原因としては,チェルノブイ リ起源の降下物が集中した 1986 年 5 月の時期 ⋥ធᳪᨴ ᳪᨴ⚻〝ഀว䋨䋦䋩 たな降下がほとんど観測されないため,ほぼ全 䉼䉢䊦䊉䊑 䉟䊥 㑆ធᳪᨴ 䉼䉢䊦䊉䊑 䉟䊥 図 2 玄麦中 Sr と Cs 含量の汚染経路別割合の推移 第 12 章 農業環境のモニタリングと手法の開発 177 がちょうど小麦の出穂開花期に当たり,直接汚染により地上部(特に穂)に付着した 137Cs が効率 よく体内に取り込まれたためと推察された(駒村ら,2001).水稲の場合には,生育時期が小麦よ り遅いために,直接の影響を免れた. 白米,玄麦における 137Cs や 90Sr の濃度とそれぞれの放射性核種の降下量の関係は,y=aXb(y: 子実中の核種含量,X:核種の地上への降下量,a:間接汚染相当分)の関係式で表され,不測 の核事故等による子実中の放射性核種の濃度の推定に用いることができると考えられた.なお, 137 Cs,90Sr とも,コメでは白米より玄米,麦では小麦粉よりも玄麦で濃度が高く,それぞれ,ヌカ とフスマに集積しやすいことがわかっている(駒村ら,2002). 6 畑土壌中の 90 Sr の下層への溶脱と支配要因 畑土壌に吸着された放射性ストロンチウム 90Sr の減少速度が,どのような環境要因によって決 まるのかを明らかにすることを目的として 1950 年代から 45 年間収集してきた土壌の放射能モニタ リングデータを解析した(平成 18 年度研究成果情報 , 山口ら)(Yamaguchi et al.,2006). 畑土壌の作土層では,30 〜 80%の 90Sr が交換態として存在しているが,交換態の 90Sr の長期モ ニタリングデータをもとにその減少速度から土壌中の滞留半減時間を計算したところ,90Sr は,放 射壊変による半減期(28.8 年)よりも速く作土から減少していた.それは,畑土壌の作土層からの 90 Sr の減少が,放射壊変以外に水浸透に伴う下層への溶脱や小麦吸収による収奪にも起因するため と考えられた. 90 Sr の減少速度は降水量などとは直接的な関係がなかったが,土壌陽イオン交換容量(CEC)と 正の相関関係が認められた.さらに,CEC が大きい土壌ほど 90Sr の小麦により吸収される割合 (CR)が低い傾向にあったことから,90Sr は土壌の陽イオン交換サイトに保持されることで畑土壌 から失われにくくなったと考えられた.以上から,畑土壌の 90Sr の長期的な減少を支配する多く の環境要因の中で,土壌の CEC が最も重要な支配要因であることが示された.核実験や原子力施 設事故などが発生した場合,90Sr の農作物への影響の程度は,継続的に行っている放射能濃度のモ ニタリングから推定できるが,さらに,事故後長期にわたり土壌と農作物に 90Sr の影響がどの程 度残るかを予測する上でこの知見は有用である. 7 農業環境中における 137 Cs と 210 Pb の動態の解析 放射性核種の濃度変動は多くの場合,水移動によって引き起こされ,核種が地表土壌から河川水 へ移動する形態は溶存態および懸濁態に大別される.137Cs およびウラン系列核種の 210Pb について は懸濁態が移動の主要形態と考えられる.農業環境技術研究所では,これらの核種の農業環境中で の動態を解明し,土地利用ごとの水系への流亡ポテンシャルを見積もることを目的として,農業環 境中における土地利用ごとの 137Cs および 210Pb 存在量の調査と水系への流亡フラックスの観測を 行っている.これらの核種の土地利用ごとの深度別存在量および水文観測地点における平水時のフ ラックスについて調査をした結果,小流域内の畑地および水田における,137Cs および 210Pb の濃度 は,同流域の林地と比較して低い値を示し,土地利用形態がこれらの核種の挙動に強く影響してい ることが示された.また,農業環境中における 137Cs と 210Pb の動態には少なからず違いがあるこ とが明らかになったが,詳しい解析には今後,更にデータを収集する必要がある.また,現在,平 水時のみならず大雨などのイベント時の流出実態などについても調査研究を進めている. 8 農耕地土壌におけるヨウ素の挙動の解明 放射性ヨウ素(129I)は半減期が約 1600 万年と非常に長く,ヒトの甲状腺に蓄積することにより 放射線障害をもたらす最も危険な放射性核種の一つである.この 129I は核燃料再処理施設の運転に 伴い,ごく微量ではあるが環境中へ放出される.また,ヨウ素の別の放射性同位体 131I は半減期が 178 2.農業環境中の放射性物質のモニタリング 8 日と短いが,チェルノブイリ原子炉事故の際に大量に環境中に放出された.放射性廃棄物処分施 設の稼働等をひかえ,これらの放射性ヨウ素の継続的な監視を行って,原子力施設の安全評価,お よび事故等緊急時の対応策に活用できる基礎的・基盤的な知見を蓄積するとともに,放射性ヨウ素 の農業環境中における動態を解明して,ヒトの内部被曝量を予測するための基礎データを得る必要 がある. 放射性ヨウ素は非放射性ヨ ウ素と物理化学的性質や環境 中での挙動が実質的に同じと 考えられることから,放射性 ヨウ素の動態解明には非放射 性ヨウ素の解析結果が適用で きる.そこで,環境中の濃度 が非常に低い放射性ヨウ素の 代わりに非放射性ヨウ素につ いて挙動を解析した.環境中 のヨウ素は,ヨウ素酸イオン (IO3−, +5 価 ) ,ヨウ化物イ オ ン(I−,−1 価 ) ,分子状 ヨウ素(I2,0 価) ,有機態ヨ ウ素など複数の形態をとりう る.このうち,ヨウ化物イオ ンは土壌に保持されにくいた め,地下水や河川水などに流 出しやすい形態である.近接 する畑,森林土壌,水田土壌 を比較すると,水田土壌のヨ ウ素濃度は特異的に低い.さ 図 3 XANES の概要と適用例 らに水田では土壌が還元状態 にある 8 月ごろに,土壌水中の濃度が上昇するが,畑や森林の土壌水ではこのような季節変動は観 察されない(Yuita et.al., 2005, 2006) .土地利用により,なぜヨウ素の挙動が異なるのかを明らか にするためには,土壌固相中でヨウ素がどのような形態で存在するのかを調べる必要がある.とこ ろが,一般的な抽出操作によりヨウ素の分析をしようとすると,化学形態が変化してしまう可能性 がある.シンクロトロン放射光源 X 線吸収スペクトル近傍構造(XANES)を適用すると,固体の ままで土壌中ヨウ素の非破壊形態分析が可能であり,従来法で問題となるその抽出過程の形態変化 を回避できる(平成 17 年度研究成果情報 , 山口ら)(Yamaguchi et al., 2006, 2010). XANES による非破壊形態分析により,IO3−は湛水条件下で微生物活動に伴う酸化還元電位の低 下によって,I2 あるいは有機態ヨウ素となることがわかった.また,I −まで還元されたヨウ素は土 壌に保持されにくいため,土壌溶液に溶出することが灌漑期の水田土壌からのヨウ素溶脱の主要因 と考えられた.さらに,還元により生じた I2 の一部は,土壌固相に保持されると推察された.腐 植物質に富む黒ぼく土では畑条件下でも IO3−が分子状ヨウ素への還元を経て,有機態ヨウ素とし て土壌に蓄積すること明らかになった(Yamaguchi et al., 2010). 第 12 章 農業環境のモニタリングと手法の開発 179 ⑵ 放射性核種等の同位体比を利用した解析技術の開発 1 はじめに 放射性核種やその壊変生成物の空間的 ・ 時間的分布は,地質年代的な岩石,土壌等の生成過程に 加え,有史以来の人間活動等の影響も受けて,複雑な各種要因の基に成立している.特に,人工放 射性核種の影響は 20 世紀以降の最近の人間活動によって生じたものである.その複雑な要因を解 きほぐして現在の状況を説明することは困難であるが,逆に,それぞれの地域に結果として生じて いる特定の元素同位体の特徴的な分布を利用して,近年起こっている地球環境(あるいは地域環 境)の変化や物質の起源来歴等を解明するツールとして活用することが可能である.農業環境技術 研究所は,この点に着目して様々な評価手法の開発を行ってきた. 2 天然放射性核種 210 Pb を用いての中国砂漠化域の風食の評価 近年,中国,モンゴルなどの東アジアでは,農耕地での土壌侵食や,砂漠化にともなう土地荒廃 が進み,それにともなう黄砂の発生が深刻な問題となっている.これらの問題は,黄砂の飛来等に より,わが国へ及ぼす影響も大きいことから,できるだけ早く適切な対策を施すことが必要であ る.そこで,砂漠化域を対象に,過放牧等による植生の退行とともに風食の初期段階で起こる土壌 中の微細粒子の移動を検知するための指標として,空中から降下する微細粒子に吸着している天然 放射性核種 210Pb の利用が検討された(平成 15 年度研究成果情報,藤原ら). 210 Pb は,核爆発由来の 137Cs とは異なり,天然由来の空中降下物であるため, (1)侵食により一 度表層の微細粒子が失われても,再び表層に集積する,(2)半減期が約 22 年であり 10 年オーダー の変化を追跡できる等の特徴があるため,土壌侵食評価に利点をもつ.また,210Pb は水に不溶で, 通常,土壌の表層に集積されていることから,表層土壌の濃度比較で風食の程度を推定できる. 1992 年以降,中国内モンゴル自治区奈曼の試験地において,1996 年までは放牧,1997 年以降は 全て禁牧の処理で放牧試験を実施し,210Pb を用いた風食の推定法の有効性について検討した. 1998 年 8 月に地表から深さ 5cm までの土壌を採取して,210Pb 放射能濃度を測定したところ, 210 Pb 濃度は,放牧圧が高く,裸地化していた重〜中放牧区の場所ほど低く,また各放牧区の 210Pb 濃度平均値は,禁牧区から重放牧区へかけて,植被率が小さくなるほど,小さくなる傾向が認めら れた.この結果により,210Pb 濃度分布が,植被の退行にともなう土壌表層部の風食を表す指標と して有効に活用できることが明らかとなった. 3 黄砂現象にともなう 137 Cs 大気降下量の変動の解析 2002 年 3 月には,青森や新潟など北日本や日本海側の複数地点で,チェルノブイリ原発事故以 来最大となる 137Cs 大気降下量が記録された.この時期に浮遊粒子状物質(SPM)濃度の上昇も観 測されており,砂塵の飛来が降下量増大の原因であったと推定された.また,同時期に中国大陸の 草原域において顕著な砂塵の発生が観測されたが,大陸の草原では最近,気候変動や人為による砂 漠化が進行しており,2001 年から 2002 年にかけての中国北部の深刻な干ばつにより植生被覆が脆 弱化していたことがその原因と考えられた. そこで中国北部の草原地域を対象に現地調査を実施したところ,深さ 2cm までの表土から比較 的高濃度の 137Cs が検出された.こうした 137Cs の分布は,少ない降水量による下方への浸透の遅 れと,草原の植生被覆による表土侵食の抑制の二つの複合的要因により 137Cs の減損が少なかった ということで説明できるが,土壌中 137Cs 蓄積量と年間降水量の間に正の相関が認められたことか ら,この 137Cs は特定の核実験場や施設からの放出物ではなく,1980 年代以前のグローバルフォー ルアウト(地球規模の放射性降下物)に由来することが判明した(平成 19 年度研究成果情報,藤 原ら) (Fujiwara et al., 2007) . 180 引用文献 この研究により,わが国における 137Cs 大気降下量の変動に,黄砂現象が強く関与しており, 137 Cs を含む砂塵の主要な起源は大陸の草原域であることが証明された. 4 ストロンチウム同位体比による野菜の産地判別技術の開発 海外からの農産物の輸入量が急増し,国内産品の価格よりも安価に流通している状況下で原産国 表示の徹底が求められているが,その表示を確認する手法は未だ確立されていない.これまで提案 されている手法は,DNA の塩基配列の違いにより判別する方法,元素組成の違いにより判別する 方法,軽元素の同位体比の違いにより判別する方法等様々であるが,農業環境技術研究所ではスト ロンチウム同位体比(87Sr/86Sr)に着目し中国産ネギの 3 大産地である山東省,上海,福建省産ネ ギと国内産ネギの判別法を開発した(平成 15 年度研究成果情報,織田ら). すなわち,87Rb の壊変により生成する 87Sr と安定同位体の 86Sr の比は放射年代測定法の一つと して利用されているが,中国産ネギについてこのストロンチウム同位体比をとってみると,山東省 産(18 点)が 0.710 〜 0.712 の範囲,上海産(13 点)が 0.710 のごく近傍,福建省産(6 点)が 0.706 〜 0.709 の範囲にあるのに対し,国内産(67 点)のストロンチウム同位体比(0.704 〜 0.710)は, 中国の山東省及び上海産より低い.また,中国福建省産のストロンチウム同位体比は国内産と同 水準にあるが,福建省産のストロンチウム濃度(平均 37mg kg−1)は国内産のストロンチウム濃度 (平均 14mg kg−1)より概して高かった.そこで,中国産ネギと国内産ネギのストロンチウム同位 体比及び濃度から線形判別関数を求め,産地判別試験用のフローチャートを作成して店頭買い取り 品を用いた判別試験を行った結果,中国産 36 点中 35 点,国内産 6 点中 5 点が正しく判別された. これらのネギのストロンチウム同位体比は生産された産地土壌の値と一致しており,他の農産物に 対しても同じ手法が適用できると考えられるので,現在は,ネギ以外の各種の農産物に関してもス トロンチウムをはじめとする重金属元素の同位体比の地域特異性を利用して産地判別が盛んに試み られている(川崎 ・ 織田 ,2005) . ࿖ౝ↥ ᶏ 0.713 ጊ᧲⋭ ᑪ⋭ 0.712 0.711 87 86 Sr/ Sr 0.710 0.709 図 4 ネギの Sr 同位体比と濃度 図中のラインは線形判別関数 式 1:0.154x+2547y−1809=0 式 2:0.364x−550y+380=0 x は Sr 濃度,y は Sr 同位体 0.708 0.707 0.706 0.705 0.704 0 10 20 30 40 50 㪪㫉Ớᐲ㩷㪲㫄㪾㩷㫂㪾-1] 60 70 引用文献 1)Fujiwara, H., Fukuyama, T., Shirato, Y., Ohkuro, T., Taniyama, I. and Zhang, T. H.(2007) Deposition of atmospheric 137 Cs in Japan associated with the Asian dust event of March 2002. Science of the Total Environment, 384, 306-315 第 12 章 農業環境のモニタリングと手法の開発 181 2)川崎 晃 , 織田久男(2005)水田土壌と米のストロンチウム同位体比の関係 . 日本土壌肥料学 雑誌 , 76, 579-585 3)駒村美佐子 , 津村昭人 , 小平 潔(2001)我が国での 90Sr と 137Cs による白米の汚染:1959 年以 来 7 年間の長期観測とその解析 . Radioisotopes, 50, 80-93 4)駒村美佐子 , 津村昭人 , 木方展治 , 小平 潔(2002)国産小麦の 90Sr と 137Cs 汚染に関する長期 観測と解析:1959 年以来チェルノブイリ事故を含む 37 年間 . Radioisotopes, 51, 345-363 5)Yamaguchi, N., Seki, K., Komamura, M. and Kurishima, K.(2006)Long-term mobility of fallout 90 Sr in ploughed soil, and 90 Sr uptake by wheat grain. Science of the Total Environment, 372, 595-604 6)Yamaguchi, N., Nakano, M., Tanida, H., Fujiwara, H. and Kihou, N.(2006)Redox reaction of iodine in paddy soil investigated by field observation and I K-edge XANES spectroscopy. Journal of Environmental Radioactivity, 86 (2),212-226 7)Yamaguchi, N., Kawasaki, A. and Iiyama, I.(2009)Distribution of uranium in soil components of agricultural fields after long-term application of phosphate fertilizers. Science of the Total Environment, 407, 1383-1390 8)Oda, H., Kawasaki, A. and Hirata, T.(2002)Determination of the geographic origin of brownrice with isotope ratios of 11B/10B and 87Sr/86Sr. Analytical Sciences, 17, 1627-1630 9)Yamaguchi, N., Takamatsu, R., Nakano, M. and Tanida, H. (2010) Inorganic iodine incorporation into soil organic matter: Evidence from iodine K-edge X-ray absorption nearedge structure. Journal of Environmental Radioactivity, 101, 451-457 10)Yamaguchi, N., Watanabe, Y., Kawasaki, A. and Inoue, C.(2007)Uptake of uranium by spinach grown in Andosols accumulating trace amounts of fertilizer-derived uranium. Environmental Radiochemical Analysis, 52-57 RSC Publishing ISBN. 978-0-85404-263-0 11)Yuita, K., Kihou, N., Yabusaki, S., Takahashi, Y., Saitoh, T., Tsumura, A. and Ichihashi, H. (2005)Behavior of iodine in a forest plot, an upland field and a paddy field in the upland area of Tsukuba, Japan. Iodine concentration in precipitation, irrigation water, ponding water and soil water to a depth of 2.5 m. Soil Science and Plant Nutrition , 51, 1011-1021 12)Yuita, K., Kihou, N., Ichihasi, H., Yabusaki, S., Fujiwara, H., Kurishima, K. and Noda, T.(2006) Behavior of iodine in a forest plot, an upland field and a paddy field in the upland area of Tsukuba, Japan: Seasonal variations in iodine concentration in precipitation and soil water and estimation of the annual iodine accumulative amount in soil horizons. Soil Science and Plant Nutrition, 52, 122-132 182 第 13 章 農業環境情報の活用 1.はじめに 農業環境問題を対象とする研究においては,農業環境を構成する土壌,水,大気,昆虫,微生 物,動物,植物に加え,これら環境構成要素と農業活動などによって投入される肥料や農薬との相 互の関係を把握することが重要である.このためには,これら要素の調査・観測・分析などによる データの蓄積と,同時にそれらを収集・分析するための手法の開発が必要である.古くから保存 されている標本や,分類・特性・機能・動態などの知見,予測手法など多くの情報が求められてい る. 2001 年に発足した農業環境インベントリーセンターは,これら農業環境の構成要素に関する情 報を収集し,研究活動や行政・農業生態管理に役立ててもらうための利用法を開発することを目的 として設立されたセンターである.インベントリーとは,一般には財産や在庫品の目録を意味する が,自然資源の目録,目録の作成,さらには目録に記された物品の意味でもある.最近では「温暖 化ガスインベントリー」のように,自然科学系でもよく使われるようになっている. 農業環境インベントリーでは, (1)標本の作製と保存・管理,(2)分類,同定および診断,(3) 農業環境の要素間の相互作用を含む特性の解明, (4)各種のデータベースの開発と公開,を行って いる.その結果,10 年間で,多数の標本や調査データの蓄積と,それら情報の公開を行うことが できた. ここでは,この 10 年間の主要な研究成果を紹介する. 2.分類 ⑴ 土壌 1 包括的土壌分類第 1 次試案の作成 包括的土壌分類第 1 次試案が策定された.わが国では,土地利用に左右されず土壌の種類を判定 することができる土壌分類法の整備が遅れ,林野と農耕地の境界域では,2 種類以上の分類を使用 する必要があった.こうした背景から,日本ペドロジー学会では,2003 年に「日本の統一的土壌 分類体系 ― 第二次案(2002) 」を発行した(図 1)が,この分類案も完全なものではなく,「大縮 尺土壌図に活用するためには下位カテゴリーを設定する必要がある」(日本学術会議答申 2004) とされた.そこで,農業環境技術研究所では,第 2 期中期計画において,(1)国土全域をカバー し, (2)実用的な土壌図に対応可能な下位カテゴリー区分を設定する,ことを主眼に「包括的土壌 分類第 1 次試案」の作成を開始した.この作成には,土壌の地域的な分布,多様な土地利用等をカ バーする必要性があるため,アドバイザーとして大学・関係独法から専門家に意見を求めた.こう して,2011 年 3 月に農業環境技術研究所報告として,「包括的土壌分類第 1 次試案」を公表した. 「包括的土壌分類第 1 次試案」では,上位分類群から順に土壌大群,土壌群,土壌亜群,土壌統 群から構成され,まずはじめに,対象となる土壌の主たる生成作用と発達程度の差異などによっ て, 「造成土大群」 , 「有機質土大群」 , 「ポドゾル大群」,「黒ボク土大群」,「暗赤色土大群」,「低地 土大群」 , 「赤黄色土大群」 , 「停滞水成土大群」 , 「褐色森林土大群」,「未熟土大群」の 10 土壌大群 ふ かん に分けた.これは,日本の土壌を大きくグループ分けして俯瞰する際に効果的である(表 1).ま た,土壌大群を,地下水位などの水分条件,土壌母材などによって,さらに土壌群として分類し 第 13 章 農業環境情報の活用 183 た.次に,各土壌群間で中間的な性質をもつ土壌からその土壌群の典型的な性質をもつ土壌までを れき 土壌亜群としていくつかに分類し,その後さらに,各土壌亜群を粘土含量の違いや礫層の有無など によって最下位分類群である土壌統群に分類した. 土壌の種類と名前は,現地調査といくつかの分析データを組み合わせた検索表を用いて検索・同 定することができる. 図 1 近年の日本土壌分類の現状 表 1 土壌大群と土壌群,亜群 包括的土壌分類 第 1 次試案 群 亜群 A. 【造成土大群】 土壌大群 2 5 統群 B. 【有機質土大群】 1 4 C. 【ポドゾル大群】 1 5 15 D. 【黒ボク土大群】 6 26 102 E. 【暗赤色土大群】 3 6 18 F. 【低地土大群】 5 24 78 G. 【赤黄色土】 2 15 60 H. 【停滞水成土大群】 2 9 36 I. 【褐色森林土大群】 1 9 36 J. 【未熟土大群】 4 13 21 合 計 10 27 116 381 農耕地土壌分類,第3次改訂版 24 77 204 日本の統一的土壌分類,第二次案 10 31 116 15 184 2.分類 ⑵ 昆虫 国内外から持ち込まれた昆虫の同定依頼に多数対応するととともに,昆虫の簡易な識別法などを 開発して広く公開してきた.そうした取り組みの中で,新たな害虫を発見して,単に同定だけでな く,現場での防除対策にも役立つ成果を上げてきた.こうした新発見の背景には,農業環境技術研 究所に保存されている多くの標本情報が役立っていることを特に明記したい. また,成果の一部は,同定依頼された都道府県でいち早く対策として役立てられている.とく に,北海道で発生した新害虫ヘリキスジノメイガ(下記参照)については飛来源の検討が必要との ことで,同定した研究者が政府の要請でロシアにわたり現地の調査を行うなど,迅速な対応を行っ ている. 下記には,害虫の同定や識別法の開発についての成果を項目ごとに紹介する. 1 分類,同定の成果 平成 14 年度には,3 種のヤガ科新害虫の九州以北における発生を確認した.ゴルフ場芝草地に おけるクシナシスジキリヨトウの初発生と,わが国では害虫とは考えられていなかったイラクサギ ンウワバの多種作物での発生を確認した.また,シイタケ菌床栽培におけるムラサキアツバの害虫 化を確認し,これら害虫の発生状況を明らかにした.いずれも,九州以北でも最初の確認となっ た. このシイタケ菌床栽培の研究を続ける中で,平成 18 年度には新たな害虫化を明らかにした.菌 床シイタケの菌糸を食害するチョウ目ヤガ科幼虫の飼育羽化成虫の形態を詳しく検討したところ, これら成虫がこれまで害虫としては記録のなかったナミグルマアツバであることが明らかになっ た.同時に,この害虫に関しては,専門家以外でも識別可能にするために,それまで雌成虫での識 別が難しかった近縁なヒメナミグルマアツバとの識別法も開発した. これらの国内の害虫だけでなく,侵入害虫についても大きな成果が得られた. まず,侵入害虫クロテンオオメンコガ(新称)の国内における広範囲での発生の確認がある.ク ロテンオオメンコガ(新称)はチョウ目ヒロズコガ科に属し,海外では観賞用植物,バナナ,サツ マイモ,トウモロコシなどの害虫として知られているが,これまで,わが国では小笠原での採集記 録のみであった.研究の結果,これら害虫が本州から沖縄に至る広い範囲で発生していることを新 たに確認した(平成 16 年度研究成果情報,吉松ら). また,北海道内の広域で発生した新害虫がヘリキスジノメイガであることをいち早く同定した. 平成 20 年夏に北海道内の広域においてダイズやニンジンなど多種類の作物を加害する見慣れない チョウ目幼虫が多発生した.そこで,羽化させた成虫を詳細に調べ,これまでわが国では害虫とし ての記録がないヘリキスジノメイガであることを明らかにした(平成 20 年度研究成果情報,吉松 ら) .この成果は,北海道の防除対策にいち早く活用された.また,政府の要請により,飛来源と 考えられるロシアに研究員が行って情報収集をするなど,ヘリキスジノメイガの国内での被害拡大 を抑制する対策に大きな貢献をしている. さらに,平成 22 年には,南西諸島で多発生した新害虫をアフリカシロナヨトウと同定した.平 成 20 年夏以降に,沖縄県多良間島・西表島,鹿児島県喜界島・奄美大島でイネ科牧草やサトウキ ビを加害する見慣れないヨトウ類幼虫が多発生した.そこで,依頼を受けて,羽化した成虫を詳細 に調べた結果,これまでわが国では害虫としての記録がないアフリカシロナヨトウと同定した(図 2) (平成 22 年研究成果情報,吉松ら) .この成果も現地で対策に役立てられている. 第 13 章 農業環境情報の活用 185 図 2 アフリカシロナヨトウ成虫 左:多良間島産の羽化成虫, 右:農業環境技術研究所所蔵の杉繁郎コレクションの標本 (沖縄本島で 1955 年 6 月 12 日に採集された個体) 2 識別法の開発 これまでの多くの知見を基に,ヤガ科害虫 4 グループ類似種の幼生期の識別法を開発することが できた.チョウ目ヤガ科主要害虫のうち,特に幼生期の類似した 4 グループ(タバコガ類 4 種,キ さなぎ ヨトウ類 2 種,ネキリムシ類 3 種,ヨトウ類 2 種)合計 11 種について,終齢幼虫および蛹を記載 し,検索表を作成して,幼生期の識別法を確立した(平成 13 年度研究成果情報,吉松). ⑶ 微生物 微生物に関しては,従来から農業環境技術研究所が国内の中心となって進めてきた植物病原菌の 標本の収集・保存や病原菌の分類・同定依頼への対応を行うとともに,第 1 期,2 期の中期計画の 目玉の一つである農業環境中に生息する「微生物インベントリーの構築」の中で収集された微生物 の分類・同定が行われた.こうした取り組みの結果は,作物に生息する微生物の特徴を明らかにす るとともに,そうした研究成果を基に,作物生育促進菌や農業廃棄物(イナワラや生分解性プラス チックなど)や汚染物質(アレルゲンとなるカビやカビ毒など)の浄化に役立つ機能を有する微生 物情報の収集にも役立っている.また,こうして集められた情報はインベントリー「microForce」 (データベースの項参照)で公開してきた. 以下の項目ごとに紹介する. 1 分類・同定:植物病原菌の同定 国内のサトウキビ生産地では,サトウキビ白すじ病が出ているが,健全な種苗を生産するために は,類似症状を示すサトウキビとの簡易な識別法の開発が必要となり,その開発の依頼があった. 解決作の一つとして遺伝子診断が考えられたので,細菌が生存するために必須の 3 種類の遺伝子の 塩基配列を比較して,世界のサトウキビ白すじ病菌を分類したところ,本病原細菌は,5 つのタイ プ(A 〜 E)が存在し,国内(沖縄県)にはそのうち B タイプのみが存在することが明らかになっ た(平成 19 年度研究成果情報,對馬ら) .この結果は,国内の菌の識別には B 型のみをターゲッ トにした遺伝子診断を行うことによって,同定・診断が可能であることを明らかにした. 2 分類・同定:植物生息微生物の分類・同定 植物生息微生物として,健全イネでの常在が明らかになった Pseudomonas huttiensis の再分離が ある.Pseudomonas huttiensis は,もともと水から分離された細菌で,環境中での生存の報告はな かったが,健全イネ(品種コシヒカリ)から高頻度に分離され,はじめて植物体に生息しているこ とを明らかにした.また,本菌は遺伝子の相同性と細菌学的性質の詳細な検討から,1986 年に新 設された Herbaspirillum 属の細菌と位置づけられることを報告した(平成 13 年度研究成果情報, 篠原ら) .その他,栽培イネ,コムギなど 8 種の植物に生息する多数の細菌の定性・定量的解析を 186 3.収集・保存 行い(微生物の収集の項目参照) ,その解析の結果,有用機能を有する細菌を分離し,属レベルの 推定を行った. さらに,これら分類・同定の情報に加えて,植物上の微生物数も記録されている.この結果か ら,たとえば,温室栽培トマトと露地栽培トマトの葉面に生息する細菌相が顕著に異なるだけでな く,露地栽培トマトの菌量は温室栽培トマトの約 100 倍で,優占する細菌群も異なることも明らか にした.こうした研究は,微生物の管理を通して植物の栽培管理に役立つと考える. 3 微生物の活用法 農業廃棄物の浄化に関連して,バイオエタノール生産に用いるバイオマス原料および発酵条件の 簡便な評価方法の開発があげられる.平成 21 年度には,様々なバイオマスについて,バイオエタ ノール生産原料としての評価,原料の糖化・エタノール発酵に最適な酵素の種類と発酵微生物の選 定,および実証規模の発酵条件の検討を簡便かつ効率的に行うことができる方法を開発した.赤か び毒デオキシニバレノールを分解する細菌 Nocardioides sp. WSN05-02 株をムギ栽培圃場から分離 した(平成 22 年度研究成果情報,對馬ら) . また,栽培イネ「コシヒカリ」の葉鞘から世界で初めて窒素固定細菌を発見するとともに,ト マトに生息する細菌の研究の結果,トマトが生産する毒素α- トマチンの分解菌を報告した.さ らに,ムギ類赤かび病菌が生産するかび毒デオキシニバレノール(DON)の分解細菌も発見し報 告するとともに,特許出願を行った.DON は世界中でムギ類やトウモロコシなどの生産地域で 問題になっている人畜に毒性を示す物質であり,国内においてもムギ圃場,サイレージ,食品中 の DON の低減技術の開発が求められている.微生物インベントリーの成果や微生物情報を基に, DON を分解する細菌の分離をめざした結果,DON を分解するグラム陽性細菌 Nocardioides sp. WSN05-02 株をムギ栽培土壌から分離することに成功した.グラム陽性細菌の DON 分解菌の報 告は世界初である.DON を処理したムギ穀粒に本分解細菌を処理したところ,7 日間で DON が 90%減少することを確認した. 3.収集・保存 土壌分類案の提案や,新害虫の同定,新機の微生物を発見することは,直接成果として役立つも のであるが,それらを効率よく,どこの機関よりも早く成果にするためには,土壌,昆虫,微生物 の標本やその情報のデータベース化など,これらの一見地味で単調で,必ずしもすぐには業績にな らない作業の積み重ねがきわめて重要である.農業環境インベントリーでは,これらの収集,保存 にも積極的に取り組んできた. ⑴ 土壌の収集・保存 1 土壌標本 土壌情報の核となる,土壌モノリスの作成(図 3),全国土壌試料,深層土壌の調査(採取装置) に勢力的に取り組み,10 年間で土壌モノリス 60 点を作成した.平成 20 年度から始まった農林水 産省土壌炭素調査事業では,毎年全国から送付された土壌試料(年間約 6,000 点)を保管してお り,必要に応じての県の分析結果の検証等を行っている.深層土壌の調査資料 12 点(地表から最 大 5m までの層別の調査データ)の収集を行った.一部については,公開データベース(全国土壌 情報閲覧システムなど;後述)や包括的土壌分類第 1 次試案(前述)の作成において,役立ててい る.分類案の作成では,実際に土壌モノリスを用いて分類案の検証を行っている.また,インベン トリー展示館に毎年多くの見学者を受け入れ,全国の土壌モノリスの断面試料,データ,写真など を用いて全国の土壌に関して分かりやすく紹介して,国民への普及に努めた. 第 13 章 農業環境情報の活用 187 2 試料分析のための機器開発 こうした土壌収集に取り組む一方で,収集した土壌の分析をより迅速,簡便,効率的に行うた め,環境試料の粉砕器の開発を行った.土壌や植物体などの環境試料を分析に供試する前処理にお ける微粉砕を効率よく行うことのできるもので,装置の構造は単純でかつ操作が容易なため,維持 管理コストも大幅に削減できる.商品化も行った. ᚒ䈏࿖䈱⠹ფ ᚒ䈏࿖䈱⠹ფ䈱⺞ᩏ䈲䇮䈜䈪䈮ᴦ15ᐕ(1882)䈎䉌ᆎ䉄䉌䉏䈩䈇 䉁䈚䈢䇯╙2ᰴᄢᚢᓟ䈲ᣂ䈚䈇⺞ᩏᴺ䈮ၮ䈨䈇䈩ფ⺞ᩏ䈏ో࿖⊛䈮ታ ᣉ䈘䉏䇮䈇䉐䈇䉐䈭⒳㘃䈱ფ࿑䈏䉌䉏䇮ᣉ⢈ᡷༀ䇮ജో䈍䉋䈶ㆡ ್ቯ䈭䈬䈮ᓎ┙䈩䉌䉏䈩䈐䉁䈚䈢䇯 㤥䊗䉪䈠䈱ઁ 3% ⶊ⦡ᨋ 2% 3% 䃂⠹ფ䈱ფ⟲ 䈱㕙Ⓧഀว 䈠䈱ઁ 12% 㤛⦡ 2% ⶊ⦡ૐ 4% Ἧ⦡ૐ 21% ᮸ 40.3ਁha ⇌ 180.0ਁha ᳓↰ 288.5ਁha 䉫䊤䉟 18% 㤥䊗䉪 17% 䈠䈱ઁ 6% ᄙḨ㤥䊗䉪 6% 䃂ᒻ䈫ფಽᏓ 㤛⦡ 3% ⶊ⦡ᨋ ⿒⦡ 㤛⦡ ⶊ⦡ૐ 3% 㤥䊗䉪 ⶊ⦡ૐ Ἧ⦡ૐ 䉫䊤䉟 図 3 土壌モノリスを活用して全国の土壌を分かりやすく紹介している ᵆ 188 4.データベース ⑵ 昆虫の収集・保存 1 ジーンバンク事業 10 年間で,14 系統を登録した.この登録系統の中には,抵抗性イネ品種を加害するトビイロウ ンカのバイオタイプ 4 系統や,薬剤感受性でかつ抵抗性品種を加害できない系統などが含まれる. これらは,新しい抵抗性品種の育成や,抵抗性機構の解明に役立つと考える. また,研究成果としては,ヒメカメノコテントウの飼育を容易にするために,スジコナマダラメ イガの凍結卵を代替餌とする簡易飼育法を開発した. 2 昆虫標本の収集 10 年間で,寄贈 18 万点も含めて,昆虫標本 20 万点以上を新たに収集している.こうした努力 により,昆虫標本館には,現在(平成 22 年 12 月現在)合計約 130 万点の標本を保管している.な お,これらの標本のうち,タイプ標本,一部のグループ昆虫についてはデータベース化して,Web で公開した(後述) . ⑶ 微生物の収集・保存 1 ジーンバンク事業 10 年間で,細菌と糸状菌合わせて合計 5,590 菌株を登録した.これらの中には,トマト青かび病 などの新病害の病原菌(MAFF306723 株)が含まれる.さらに,沖縄県のサトウキビ白すじ病菌 の遺伝子型の決定(前述)に役だっている. 2 微生物インベントリー この 10 年間で主要作物の表面に生息する植物生息微生物の収集,解析を行い,細菌の定性・定 量的解析とデータベースの構築を行った.栽培イネ,コムギ,トマト,イチゴ,チンゲンサイなど 8 種の植物に生息する細菌約 25,000 株の定性・定量的解析を行い,それらの情報をデータベース化 した.収集した細菌の一部は,微生物インベントリー「microForce」(下記データベースの項参照) で公開した. 3 微生物の保存技術 凍結保存細菌の反復利用効率を高めるための分散媒の改良を行った.凍結保存した細菌を融解し て一部を取り出し,残りを再び凍結すると,生き残る菌数が減少する.その程度は菌種によって異 なる.既知の分散媒にトレハロースを添加すると,凍結・融解の処理を多数回反復した後でも十分 な生存菌数が得られることを明らかにし,特許出願を行った(平成 15 年度研究成果情報,西山ら). 4.データベース 農業環境インベントリーセンターの中心課題の一つとして,土壌,昆虫,微生物など様々な農業 環境情報を統合したデータベースの構築があり,センターではこの統合データベースの開発を目指 した.その結果,平成 20 年度に農業環境インベントリーシステムを開発し,公開した.本システ ムでは,昆虫,微生物,土壌や植物標本情報の登録・編集,検索・閲覧および採集地点を地図上に 表示する機能をもつデータベースシステムである.本システムにより,分野の異なる標本の時間 的・空間的情報を地図上で同時に把握することができる. ⑴ 土壌,メッシュデータ関係 まず,特筆すべき成果として,土壌情報閲覧システムの開発,公開をあげたい.各農家や地域の 田畑にどのような土壌が分布しているのかを,Web 上で調べることができるシステムを公開した (図4) .このシステムでは,土壌図と土壌の種類ごとの写真やその性質などを,だれでも見ること ができ,全国の農耕地土壌を調べることができる.平成 22 年 4 月にはプレスリリースを行い普及 第 13 章 農業環境情報の活用 189 図 4 土壌情報閲覧システム 土壌情報閲覧システムは最新の農耕地土壌図(左)と農耕地土壌図閲覧ページ(右) 土壌図から土壌解説資料が見られるため,だれでも全国各地の田畑に分布する土壌 を調べることができる. に努めた結果,平成 22 年 10 月時点で 16 万件のアクセスがあった.加えて,民間,普及関係者か ら成果の利用依頼や講演依頼が多数あり,土壌情報の利活用に関し関心の高い人がきわめて多いこ とや,この成果へのニーズがきわめて高いことが明らかになった. また,平成 15 年度には,土壌情報の一元的収集システムの開発を行った.これまで,全国規模 の土壌調査結果は,ファイルを転送していたが,Web GIS の機能を使って全国規模の土壌調査結 果(現地情報,断面記載,理化学分析値,土壌図)を地形図上に表示し,調査現場においてオンラ インで新たな調査結果を入力や修正できるシステムを初めて開発した.これにより,現行の行政調 図 5 農業統計情報メッシュデータ閲覧システムによるデータの表示 図では,茨城県つくば市周辺の地図.農地面積の大きさを青〜茶色に色分けして表示してい る.各メッシュをクリックすると,その 1km メッシュ内の作物ごとの栽培面積が見られる. 190 4.データベース 査事業における結果を効率的かつ一元的に収集できようになった. さらに,農業統計情報メッシュデータ閲覧システムも様々なニーズに対応できる成果と考える. 本システムでは,農業集落単位で集計された耕地面積や家畜飼養頭数といった農業統計データを農 業環境研究で利用しやすい 1km メッシュ単位のデータに変換し,データフォーマットが CSV 形式 のファイルを作成した.本成果は,21 年度には Web 公開し,だれもが全国マップ上の 1km メッ シュの単位で農業統計データを閲覧できるようにした(図 5.).この成果の一部は,職務発明プロ グラム(平成 21 年度)として申請するとともに,プレスリリースも行い,多くの新聞に掲載され た結果,民間,行政などから情報の利用申請が来ている. ⑵ 昆虫関係 1 農業環境技術研究所が所蔵する昆虫標本のデータベース化 まず,平成 15 年度には,農業環境技術研究所で所蔵している昆虫タイプ標本 508 種の一覧表と 279 種の画像を含む標本情報を Web 上で公開した.外部から,タイプ標本の所蔵状況の確認およ び標本の形態情報の入手が容易になる.平成 21 年度には,さらに杉 繁郎コレクション(ガ類), 寺山 守コレクション(ハチ目) ,田中和夫コレクション(オサムシ科)など,最近寄贈されたタ イプ標本 379 点を Web 公開した(図 6) . 図 6 タイプ標本室で保管している杉氏のガ類タイプ標本 タイプ標本以外では,土生コレクション標本情報閲覧システムの開発がある.故土生昶申博士が 収集した標本コレクションのうち,アオゴミムシ族,マルクビゴミムシ亜科の標本情報が閲覧でき るシステムを開発した.標本ラベルの閲覧だけでなく,採集地の地図投影,データダウンロードも 可能なため,様々な研究に利用できる.また,所蔵標本の情報に関しては,それらが登録,検索, 閲覧ができる昆虫インベントリーシステムを構築した. 2 三橋ノート画像データベース 国内屈指の古い文献の情報として利用価値の高い三橋ノートの情報を何回かに分けてデータベー 第 13 章 農業環境情報の活用 191 ス化して公開してきた.平成 22 年度には全情報を公開することができた.三橋ノートは,明治時 代から昭和 20 年代後半までに国内で出版された主要な昆虫関連図書や雑誌に現れた昆虫名とその 文献の書誌情報を記録したものである.以下,公開の順に紹介する. 平成 17 年度には,トンボ目とチョウ目(約 20,000 ページ)の画像データベースを作成し,Web 上に公開した.21 年度には,昆虫の中で最大のグループであり,農林学的にも重要なコウチュウ 目に関係する 135 冊,19,992 ページの画像をデータベース化し公開した.さらに,平成 22 年度に は,全 483 冊のうち,作業がまだであったハチ目等全 111 冊の画像を公開し,さらに,全冊子につ いてページ裏面に記載されている昆虫の分布等に関するメモの情報も公開した(図 7). 図 7 三橋ノート画像データベースのトップページ(左)と検索で表示された三橋ノート(右) 右図は, ”アオムシヤドリコマユバチ”をキーワードとして検索されたページを示す 3 各種昆虫の検索情報 まず,わが国の農林水産試験研究独立行政法人において飼育されている昆虫・ダニ類の譲渡,飼 育情報の交換等に役立たせるため,これらに関する情報を収集し,飼育昆虫・ダニ類データベース を作成するとともに,平成 14 年度成果として Web 上で公開した.また,日本産ヒョウタンカス ミカメ族 17 種の図説検索表をウェブサイト上に公開した(平成 18 年度農環研成果情報,中谷). 鱗毛の形態など識別点となる形質を図示したことで,高度の専門知識がなくとも容易に種が同定で きる. ⑶ 微生物関係 1 微生物インベントリー「microForce」の公開 分散型データベースにより,微生物インベントリーシステムを構築し,農業環境技術研究所が 所蔵する微生物標本,除草剤 2,4-D 分解菌および人畜植物共通の病原菌 Burkholderia 属細菌などの データベースを Web 上で公開した(図 8) .種名などによるキーワードで複数データベースから情 報が取得できる.なお,microForce は商標登録して,微生物情報の利用促進を目指している.平成 17 年度公開後毎年約 20,000 件のアクセスがある. この microForce の中で,農業環境技術研究所所蔵の標本情報や,各種の微生物データベースの 公開も行っている.以下に紹介する. 文献からの情報を基にした日本野生植物寄生・共生菌類目録を平成 14 年度に報告し,Web で公 192 4.データベース 開した.日本で報告された 95 科 1,626 種の野生草本植物に寄生あるいは共生する 312 属 1,302 種の 菌類の学名,異名,文献等を初めて目録化した.これを菌名および植物名から検索可能なデータ ベースとして,48 菌種の画像および標本情報を含む糸状菌類図鑑とともに Web 上で公開した.作 物の寄生菌の目録はすでにあるが,野生植物の寄生菌,共生菌の目録では国内最大である.また, 糸状菌類図鑑は,教育やマスコミなどでも利用されており,一般国民の微生物への理解を深めるた めにも役立っている. 特質すべき成果としては,130 年にわたって採集され,農業環境技術研究所に保存されていた 微生物さく葉標本の目録の作成と公開がある.平成 17 年度に,農業環境技術研究所に保存されて いる 1876 年以降に採集された微生物さく葉標本 7,204 点の目録を作成した.タイプ標本,菌学者 Sydow 氏らの標本,一般さく葉標本など菌種 365 属 1,477 種,寄生植物 621 属 1,322 種の標本情報 が収録されている. 図 8 微生物インベントリー「microForce」のトップ画面(左)と農業環境技術研究所所蔵微生物さく葉標本 データベースのトップ画面(右) microForce(左)で目的の微生物データベースを選ぶことができる.たとえば,微生物さく葉標本データ ベースでは,130 年間にわたって集められた貴重な微生物標本の情報が得られる. 2 農耕地 eDNA データベース(eDDASs) 平成 18 年から開始された農林水産省委託プロジェクトにおいて得られた,全国の農耕地土 壌の理化学性・生物性情報を蓄積し,だれでも簡単に利用できる農耕地 eDNA データベース (eDDASs)を開発し,平成 23 年 3 月に公開した.eDDASs では,これまで解析が困難とされてき た土壌の生物性を,DNA 情報としてみることができる.全国 18 か所で集めた 3,000 以上の土壌サ ンプルの情報(栽培条件,土壌理化学性や eDNA 情報など必須入力項目 38 件を含む最大で 68 件 の情報)を蓄積している.生物性では,細菌,糸状菌,線虫の情報が eDNA 情報として入力され ており,土壌情報,生物情報をこれほど統一的に管理しているデータベースは世界的にもない. ⑷ 線虫 平成 19 年度には,土壌線虫の簡易同定に役立つ画像付形質一覧表の公開を行った.関東北部の 畑の代表的な土壌線虫(62 属)の分類・同定に有用な形質項目を一覧表に取りまとめ公開したも のである.このうち,29 属の線虫については,特徴を捉えた鮮明な画像を見ることができる. 一方,平成 20 年度には,国内における線虫学関連文献目録を公開した.1997 年から 2006 年にわ が国で発表された線虫学に関係する文献データベースである.日本人が海外の雑誌に発表した文献 なども採録し,この期間のわが国の線虫学に関わる文献情報のほとんどを閲覧することができる. 第 13 章 農業環境情報の活用 193 5.今後の展望 農業環境インベントリーの任務は,冒頭で紹介したように,(1)標本の作製と保存・管理,(2) 分類,同定および診断, (3)農業環境の要素間の相互作用を含む特性の解明,(4)各種のデータ ベースの開発と公開である.こうした研究は,農業環境に関する研究の基盤となる研究であり,長 期的視点に立って着実に取り組むことが重要である.したがって,今後は,これらの情報を確実に 増やし,規模を拡大していく必要がある.しかし,これらの維持にはそれなりの予算,人員が投入 されていることは間違いなく,常に,蓄積した情報を広く国民に還元して,役立ててもらうことが きわめて重要であると考える. このため,これまでの 10 年間でもある程度のデータベースの構築と公開をしてきたが,その情 報の公開にあたっては,今まで以上に利用しやすく,多くのユーザーに活用してもらうシステムの 構築も重要と考えている.以上の点を踏まえ,土壌,昆虫,微生物ならびに情報発信システムの将 来展望を以下に記す. ⑴ 土壌 土壌分野では,包括的土壌分類第 1 次試案を用いた包括的な土壌情報(土壌調査データ,土壌図 など)の整備を進めるなど,土壌インベントリーを充実させていきたい.土壌データベースに関し ては,すでに土壌図データの提供を行っているが,土壌統群レベルの理化学性データの提供など, 内容の充実を図るとともに,アジア諸国の土壌データベースとの連携を行い,日本,アジアにおけ る土壌情報の発信センターとして活動していきたい. ⑵ 昆虫 昆虫分野では,今後も,既存の標本データベースを拡充するとともに新たな標本データベースを 作成し,昆虫インベントリーシステムなどを介して公開していく.形態学的な分類学的研究に加え て,DNA 分析用標本を蓄積し,解析することで,日本における当分野のセンターとして活動して いきたい. ⑶ 微生物 すでに,微生物情報を提供するサイト(microForce:商標登録済み)を構築しているが,さらに 利用者を増やすための工夫をしたい.新しい機能が明らかになった植物生息微生物などが確実に増 えてきているので,これら植物生息微生物の機能に関するデータベースの充実を推進し,国内でも ユニークな植物生息微生物情報発信基地にしたい. ⑷ 新しい情報の発信システム インベントリーの利用においては,個別インベントリーを横断的に利用できるシステムを構築 し,利便性を高めるとともに,それらを活用した研究をすすめたい.システムには所内,国内に限 定せず利用可能な情報資源を積極的に取り込み,インベントリー構築から利用までを一貫させた研 究を行うことで,特定分野に限定しないインベントリー利用研究の中心にしたい. 引用文献 (土壌) 1)神山和則(2009)1km メッシュ単位で推定した農業統計のデータベース化.農業技術,64, 299-304 2)中井 信,小原 洋,戸上和樹(2006)土壌モノリスの収集目録及びデータ集.農業環境技術 研究所資料,第 29 号 194 引用文献 3) 「土壌を愛し,土壌を守る」― 日本の土壌,ペドロジー学会 50 年の集大成 ― .日本ペドロジー 学会編. (編集者として中井 信,著者として神山和則,前島勇治,中井 信,小原 洋,谷 山一郎) pp.396,博友社,東京.2007. 4)小原 洋,大倉利明,高田裕介,神山和則,前島勇治,浜崎忠雄(2011)包括的土壌分類 第 1 次試案.農業環境技術研究所報告,??:? ― ?.(予定) 5)高田裕介,小原 洋,中井 信,神山和則(印刷中)1973 年から 2001 年までの地目改変に伴 う土壌群分布面積の変動特性の解析.日本土壌肥料学雑誌 6)高田裕介,中井 信,小原 洋(2009)1992 年の農耕地分布に基づくデジタル農耕地土壌図の 作成.日本土壌肥料学会誌,80,502-505 (昆虫) 1)馬場友希,吉武 啓,平舘俊太郎,楠本良延,栗原 隆,吉松慎一(2010)茨城県南部の農地 で採集されたクモ標本目録.Kishidaia,97,84-104 2)Hamasaki, K., Yamanaka, T., Tanaka, K., Nakatani, Y., Iwasaki, N. and Sprague, D. S. (2009) Relative importance of within-habitat environment, land use and spatial autocorrelations for determining odonate assemblages in rural reservoir ponds in Japan. Ecological Research, 24, 597-605 3)平舘俊太郎 , 楠本良延 , 吉武 啓 , 馬場友希(2010)土壌が支える生物多様性 , 身近な自然の保 全生態学 生物多様性を知る.根本正之編著 , 培風館 , 131-148 4)Huang, J., Yoshitake, H., Zhang, R. and Ito, M.(2008)Two new species of Hainokisaruzo Yoshitake and Colonnelli from China, with a checklist of the soecies of the genus(Coleoptera: Curculionidae: Ceutorhynchinae) , Zootaxa, 1776, 59-68 5)Huang, J., Yoshitake, H., Zhang, R. and Ito, M.(2008)A new species of Augustinus Korotyaev from China, with a key to the species groups of the genus(Coleoptera: Curculionidae: Ceutorhynchinae) . Zootaxa, 1871, 63-68 6)Kato, T., Bonet, A., Yoshitake, H., Romero-Napoles, J., Jinbo, U., Ito, M. and Shimada, M.(2010) Evolution of host utilization patterns in the seed beetle genus Mimosestes Bridwell(Coleoptera: Chrysomelidae: Bruchinae) . Molecular Phylogenetics and Evolution, 55, 816-832 7)栗原 隆,吉武 啓,中谷至伸,吉松慎一(2010)農業環境技術研究所所蔵日本産タマムシ科 (昆虫綱:コウチュウ目)標本目録.農業環境技術研究所報告 ,(27),141-160 8)栗原 隆 , 吉武 啓 , 中谷至伸 , 吉松慎一(2010)農業環境技術研究所所蔵日本産タマムシ科 (昆虫網:コウチュウ目)標本目録.農業環境技術研究所報告 ,(27),141-159 9)栗原 隆 , 吉武 啓 , 吉松慎一(2010), 農業環境技術研究所・標本シリーズ 6:堀川正美コレ クション(カミキリムシ類) .インベントリー ,(8),48-49 10)三宅規文,吉松慎一,中尾弘志(2009) : 2008 年に北海道で発生したヘリキスジノメイガの発 生地域と被害状況.北日本病害虫研究会報,60,223-226 11)中谷至伸,石井 実(2002)農薬施用の異なる水田の畦畔におけるカメムシ群集の多様性.日 本応用動物昆虫学会誌,46,92-96 12)Nakatani, Y.(2001)New distributional records of deraeocorine plant bugs(Heteroptera, Miridae)from Japan. Rostria, 50, 27-29 13)Nakatani, Y.(2001)Cimicicapsus Poppius a separate genus, with description of new species 第 13 章 農業環境情報の活用 195 (Heteroptera, Miridae). Tijdschrift voor Entomologie, 144, 253-259 14)中谷至伸(2006)昆虫類,363-448(執筆分担),滋賀県で大切にすべき野生生物 滋賀県レッド データブック 2005 年度版,滋賀県生きもの総合調査委員会編,pp. 563,サンライズ出版 15)中谷至伸(2005)殺虫剤・除草剤を使用しない水田畦畔の昆虫の多様性.(財)日本自然保護 協会編:生態学から見た里山の自然と保護,p186-188,講談社 16)Nakatani, Y.: Predators, Hayashi, T., Nakamura, S., Visarathanonth, P. Uraihuen, J. and Kengkanpanich, R. eds.(2004)Stored rice insect pests and their natural enemies in Thailand. JIRCAS International Agriculture Series, No.13, 73pp 17)中牟田潔,X, Chen,北島 博,中西友章,吉松慎一(2007)日本産ボクトウガ科 Cossus 属 3 種の生態.森林防疫,56, 5-9 18)那須義次,玉嶋勝範,柴尾 学,吉松慎一,内藤尚之(2010)ニッポンシロシンクイ(新称) の再発見とモモシンクイガの合成性フェロモントラップに誘殺されるシンクイガ類(鱗翅目: シンクイガ科) ,日本応用動物昆虫学会誌,54(3),115-126 19)Nishi, A., Takahashi, K., Koyama, K., Yoshimatsu, S. and Christensen, M.(2002)Effect of N-formylloline on the survival of Sawtoothed Grain Beetle, Oryzaephilus surinamensis (Linnaeus) (Coleoptera: Silvanidae) . Entomological Science, 5, 287-289 20)Tanaka, A., Yasuda, K. and Wakamura, S.(2001)Sex Pheromone of Acrolepiopsis nagaimo Yasuda(Lepidoptera: Acrolepiidae) , a predominant leafminer on Chinese yam in Tottori Prefecture: field evaluation and activity of the third component. Applied Entomology and Zoology, 36, 203-207 21) 田 中 篤, 安 田 耕 司, 若 村 定 男(2001) ナ ガ イ モ コ ガ の 発 生 生 態 と 防 除. 植 物 防 疫,54, 514-517 22)Wakamura, S., Tanaka, A., Yasuda, K. and Yasuda, T.(2001)Sex Pheromone of Acrolepiopsis nagaimo Yasuda(Lepidoptera: Acrolepiidae), a predominant leafminer on Chinese yam in Tottori Prefecture: identification and field attraction. Applied Entomology and Zoology, 36, 71-76 23)Yago, M., Yoshitake, H., Ohshima, Y., Katsuyama, R., Sivaramakrishnan, S., Murugan, K. and Ito, M.(2010)Butterflies collected in Coimbatore, Tamil Nadu, South India, with comments on its conservation significance. Butterflies(Teinopalpus),54, 45-53 24)Yamanaka, T., Tanaka, K., Hamasaki, K., Nakatani, Y., Iwasaki, N., Sprague, D. S. and Bjørnstad, O. N. (2009)Evaluating the relative importance of patch quality and connectivity in a damselfly metapopulation from a one-season survey. OIKOS, 118, 67-76 25)Yamada, K., Yasunaga, T., Nakatani, Y. and Hirowatari, T.(2006)The minute pirate-bug genus Xylocoris Dufour(Hemiptera: Heteroptera: Anthocoridae)from rice mills in Thailand. Peoceedings of the Entomological Society of Washington, 180(3),525-533 26)Yasuda, K., Yoshimatsu, S. and Nakatani, Y.(2005) Construction of Insect Inventory and its Utilization in NIAES of Japan. Material circulation through agro-ecosystem in East Asia and assessment of its environmental inpact(Niaes Series 5),207-213 27)安田耕司(2005)トコロミコガ(鱗翅目:アトヒゲコガ科)の多化性の確認と幼虫の寄主摂食 部位.昆蟲(ニューシリーズ) , 8, 7-8 28)安田耕司(2002)ナガイモの害虫ナガイモコガとその近縁種.植物防疫 , 56, 522-524 196 引用文献 29)安田耕司(2002)サヤムシガ類,農林水産研究文献解題 大豆害虫.p471-477,農林水産技術会 議事務局 30)安田耕司(2002)マメシンクイ,農林水産研究文献解題 大豆害虫.p463-471,農林水産技術会 議事務局 31)安永智秀,高井幹夫,中谷至伸:安永智秀,高井幹夫,澤哲夫編(2001)日本原色カメムシ図 鑑第 2 巻,350pp,全国農村教育協会 32)廣森 創,合田光太郎,吉松慎一,廿日出正美(2003)芝草地に発生するクシナシスジキリヨ トウの形態および生態学的特徴に関する研究.芝草研究,31, 128-131 33)吉松慎一(2010)日本周辺における長距離移動性鱗翅類の研究.やどりが,227, 16-20 34)吉松慎一(2010)広渡俊哉,第 10 部 26 クロテンオオメンコガ,「昆虫の低温耐性」.302-303, 岡山大学出版会,岡山 35)吉松慎一,栗原 隆(2010)農業環境技術研究所に最近寄贈された杉繁郎コレクションの蛾類 原模式標本(primary types)リスト,蛾類通信,256, 139-148 36)吉松慎一(2009)いまどきデータベース 1:標本館所蔵タイプ標本 ― 昆虫標本の宝庫 ― .日本 農学図書館協議会誌 , 154, 14-16 37)吉松慎一(2009)侵入害虫クロテンオオメンコガの発生状況.農業技術 , 64, 2, 80-83 38)吉松慎一(2009)農業環境技術研究所における昆虫インベントリー研究.植物防疫 , 63, 1, 49-52 39)吉松慎一(2007)ギンスジアカヤガの日本から 2 頭目の記録と雌雄交尾器形態.蛾類通信 , 245, 365-366 40)Yoshimatsu, S.(2007)A European-Mediterranean species, Mythimna albipuncta( [Denis and Schiffermüller] ) (Lepidoptera, Noctuidae, Hadeninae)new to the East Asian fauna. Transactions of the Lepidopterorological Society of Japan, 58, 202-204 41)吉松慎一(2007)未知であったスソミダレアツバ雄の形態.蛾類通信,242, 301-302 42)吉松慎一(2007)作物被害を引き起こす新たな害虫.農業技術 , 62, 10, 442-446 43)吉松慎一(2006)アマミノコメセダカヨトウ5 ― 6 月の記録と斑紋変異.蛾類通信,241, 285-286 44)Yoshimatsu, S. and Nakata, Y.(2006)Fungivory of Anatatha lignea, an interesting habit in Noctuidae(Lepidoptera) . Entomological Science, 9, 319-325 45)吉松慎一(2006)ドラセナ・サンデリアーナ・害虫 / クロテンオオメンコガ.農業総覧花卉病 害虫診断防除編追録,2 号 5 巻,p2-5,農山漁村文化協会 46)吉松慎一,櫛下町鉦敏,行徳直久,上和田秀美,佐藤嘉一,坂巻祥孝(2005)日本においてタ ケ・ササ類タケノコを加害する鱗翅類.昆蟲(ニューシリーズ)8, 91-97 47)吉松慎一,宮本泰行,広渡俊哉,安田耕司(2004)クロテンオオメンコガ(新称)Opogona sacchari(Bojer)の日本における発生状況.日本応用動物昆虫学会誌,48, 135-139 48)Yoshimatsu, S.(2004) A synopsis of Mythimna(Sablia)griseofasciata(Moore) (Lepidoptera, Noctuidae, Hadeninae)and a close relative with a new replacement name. Transactions of the Lepidopterological Society of Japan, 55, 307-314 49)Yoshimatsu, S. and Takeuchi, K.(2004)Description of Helicoverpa sugii sp. nov.(Lepidoptera, Noctuidae, Heliothinae)from the Ogasawara(Bonin)Islands, Japan, with brief biological notes. Transactions of the Lepidopterological Society of Japan, 55, 34-38 50)吉松慎一,仲田幸樹(2003)シイタケの害虫としてのムラサキアツバ(鱗翅目: ヤガ科).昆蟲 第 13 章 農業環境情報の活用 197 (ニューシリーズ) ,5, 101-102 51)Yoshimatsu, S.(2003)A remarkable new species of the genus Mythimna from Papua New Guinea(Lepidoptera, Noctuidae, Hadeninae). Transactions of the Lepidopterological Society of Japan, 54, 5-10 52) 吉 松 慎 一(2003) チ ョ ウ・ ガ 類( 昆 虫 綱・ チ ョ ウ 目 ). 日 本 農 業 害 虫 大 事 典 第 II 部, p1080-1090,全国農村教育協会 53)吉松慎一(2003)ヤガ科新害虫の発生確認と発生生態.今月の農業,8 月号,13-16 54)Yoshimatsu, S.(2002)Discovery of Mythimna lineatipes(Moore)from Japan and Thailand, with taxonomic study of its close relatives from Asia(Lepidoptera, Noctuidae, Hadeninae). Transactions of the Lepidopterological Society of Japan, 53, 55-62 55)吉松慎一(2002)マガタマリンガの雌と最も古い採集記録.蛾類通信 , 217,326 56)Yoshimatsu, S. and Legrain, A.(2001)Review of the genus Mythimna Ochschenheimer (Lepidoptera, Noctuidae, Hadeninae)in Taiwan, with description of a new species and checklist. Entomological Science, 4, 431-437 57)吉松慎一(2001)植物防疫基礎講座:ヤガ類の見分け方(4)ヨトウ類の識別法 ― ヨトウガ・ シロシタヨトウ ― .植物防疫 , 55, 277-280 58)吉松慎一(2001)植物防疫基礎講座:ヤガ類の見分け方(3)ネキリムシ類の識別法 ― カブラ ヤガ・タマナヤガ・シロモンヤガ ― .植物防疫 55, 176-179 59)吉松慎一(2001)植物防疫基礎講座:ヤガ類の見分け方(2)アワヨトウとクサシロキヨトウ の識別法.植物防疫 , 55, 130-133 60)吉松慎一(2001)植物防疫基礎講座:ヤガ類の見分け方(1)タバコガ類の識別法 ― タバコ ガ・オオタバコガ・ツメクサガ・キタバコガ ― .植物防疫 , 55, 83-86 61)吉松慎一(2001) 「肉食幼虫についての記録 ― 他種幼虫だけでなく牛肉も食べた風変わりな性 格のオオタバコガ幼虫 ― 林悦子著」に対するコメント.サイアス , 5, 54-55 62)吉武 啓,栗原 隆,吉松慎一,中谷至伸,安田耕司(2011)農業環境技術研究所所蔵土生コ レクションオサムシ科標本目録(昆虫綱:コウチュウ目).農業環境技術研究所報告,第 28 号,1-327 63)Yoshitake, H. and Niisato, T.(2010)A new Euryarthrum(Coleoptera, Cerambycidae)from Sabah, East Malaysia. Elytra, 38, 3-8 64)Yoshitake, H. and Niisato, T.(2009)A redescription of Euryarthrum hastigerum Holzschuh (Coleoptera, Cerambycidae) , with description of its new relative from Kalimantan, Indonesia. Elytra, 37, 155-163 65)Yoshitake, H., Kato, T., Jinbo, U. and Ito, M.(2008)A new Wagnerinus(Coleoptera: Curculionidae)from northern Japan: description including a DNA barcode. Zootaxa, 1740, 15-27 66)Yoshitake, H.,(2010)A biological note on Bagous spiculatus O’Brien et Morimoto(Coleoptera, Curculionidae) . Elytra, 38, 1-2 67)Yoshitake, H., Kurihara, T. and Niisato, T.(2010)New record of Euryarthrum elegans Hayashi (Coleoptera, Cerambycidae)from East Kalimantan, Indonesia. Elytra, 38, 41-42 68)吉武 啓 , 馬場友希 , 田中幸一(2010)農林水産省委託プロジェクト研究「農業に有用な生物 多様性の指標及び評価手法の開発」における分類学者の関わり.昆蟲(ニューシリーズ), 13, 198 引用文献 93-103 69)吉武 啓 , 上地奈美 , 平野幸彦(2010)台湾からの移入種ゲットウトゲムネサルゾウムシ(コ ウチュウ目:ゾウムシ科)の沖縄島における分布状況について.甲虫ニュース ,(171),1-3 70)吉武 啓 , 馬場友希 , 栗原 隆 , 吉松慎一 , 中谷至伸(2010)農業環境技術研究所・標本シリー ズ 5:昆虫・クモ類の DNA バーコーディングに関連する標本コレクションおよびデータベー スの構築.インベントリー ,(8) , 44-47 71)吉武 啓 , 細谷忠嗣 , 大林延夫 , 宇津木望 , 加藤俊英(2009)甲虫類の DNA バーコード化へ向 けた取り組み.昆虫と自然 ,(株)ニュー・サイエンス社 , 44(3),9-13 72)Yoshitake, H. and Niisato, T. (2009) Two new Euryarthrum blanchard (Coleoptera, Cerambycidae)from Kalimantan, Indonesia, with a checklist of the species. Special Bulletin of The Japanese Society of Coleopterology,(7),193-204 73)神保宇嗣,伊藤元己,広渡俊哉,中西明徳,矢田 脩,吉松慎一,上田恭一郎(2009)日本に おける鱗翅類コレクション画像データベースの構築.昆虫と自然,44(13),18 74)神保宇嗣 , 吉武 啓 , 伊藤元己(2008)DNA バーコーディングにいよる同定支援システムと JBOLI 構想 , 日本生態学会誌 , 58, 123-130 (微生物) 1)包 智華,對馬誠也(2009)土壌微生物生態研究への回帰木解析の応用.土と微生物,63, 39-44 2)Enya, J., Shinohara, H., Yoshida, S., Tsukiboshi, T., Negishi, H., Suyama, K., and Tsushima, S.(2007)Culturable Leaf-Associated Bacteria on Tomato Plants and Their Potential as Biological Control Agent. Microbial Ecology, 53, 524-536 3)Enya, J., Koitabashi, M., Shinohara, H., Yoshida, S., Tsukiboshi, T., Negishi, H., Suyama, K., and Tsushima, S.(2007)Phylogenetic diversities of dominant culturable Bacillus, Pseudomonas, and Pantoea species on tomato leaves and their possibility as biological control agents. Journal of Phytopathology, 155, 446-453 4)Ikunaga, Y., Sato, I., Grond, S., Numaziri, N., Yoshida, S., Yamaya, H., Hiradate, S., Hasegawa, M., Toshima, H., Koitabashi, M., Ito, M., Karlovsky, P. and Tsushima, S.(2011) Nocardioides sp. strain WSN05-2, isolated from a wheat field, degrades deoxynivalenol, producing the novel intermediate 3-epi-deoxynivalenol. Applied Microbiology and Biotechnology, 89, 419-427 5)月星隆雄,吉田重信,篠原弘亮,對馬誠也(2002)日本野生植物寄生・共生菌類目録 . 農業環 境技術研究所資料,26, 1-169 6)月星隆雄,吉田重信,篠原弘亮,對馬誠也(2007)農業環境技術研究所所蔵微生物さく葉標本 目録.農業環境技術研究所研究資料,30 7)月星隆雄,小板橋基夫,吉田重信,田村季実子,篠原弘亮,對馬誠也(2007)130 年間収集さ れた農業環境技術研究所所蔵微生物標本のデータベース作成.日本菌学会報,48(1),7-11 8)Tsushima, S., Shinohara, H., Nakazato, T., Ando, S., Sugisawa, T., and Tabei, Y.(2009) Phylogenetic analysis of Xanthomonas albilineans strains from Okinawa, Japan, through a comparison of the gyrB and rpoD genes in geographically distinct strains. Journal of Phytopathology, 154, 683-687 9)對馬誠也,小板橋基夫(2009) 微生物インベントリー.植物防疫,63, 42-46 第 13 章 農業環境情報の活用 199 10)包 智華,對馬誠也(2009)土壌微生物生態研究への回帰木解析の応用.土と微生物,63, 39-44 11)對馬誠也(2010) eDNA による農耕地土壌の生物性解析・評価手法の開発.土と微生物,64, 64-69 201 農業環境技術研究所 研究成果 平成 13 年度(農業環境研究成果情報 第 18 集) 殷 熙洙ら 野菜類におけるダイオキシン類の汚染軽減方策 芝野 和夫ら ダイオキシン類の水田からの流出・流下特性 川崎 晃ら ダイズのカドミウム吸収特性解明のための 113Cd トレーサー試験法 加藤 英孝 カラム浸潤法による黒ボク土の硝酸イオン吸着容量の測定 小川 直人 Burkholderia 属細菌のクロロ安息香酸分解遺伝子群発現の調節因子による制御 松井 正春ら 耐虫性遺伝子組換えトウモロコシの花粉に含まれる Bt トキシンの生物検定法 山村 光司ら 害虫の侵入定着を防止し所定の安全レベルを達成するための植物検疫強度の推定法 伊藤 一幸ら 水生植物の奇形葉による水田からの除草剤流出の検出法 平舘俊太郎ら 火山灰土壌による 2, 4 - D の吸着メカニズムの解明 藤井 義晴ら ヘアリーベッチに含まれる植物生長阻害物質シアナミドの発見 芝池 博幸ら DNA からみたタンポポ属植物の雑種個体の識別と全国分布 荒城 雅昭 土壌線虫を指標とした黒ボク土畑における耕起の影響評価 西森 基貴ら 生育阻害要因を考慮した日本の水稲生産の温暖化に対するぜい弱性の評価 小原 裕三ら 被覆資材による土壌くん蒸用臭化メチルの大気放出量削減技術の開発 鶴田 治雄ら 湿潤熱帯林の土地利用変化に伴う土壌からの温室効果ガス発生・吸収量の変動 白戸 康人ら 日本の畑土壌に適用可能な土壌炭素循環モデル(改良 RothC モデル) 駒村美佐子ら わが国における小麦の放射能汚染の長期観測 井上 吉雄ら 航空機搭載多目的デジタル分光画像計測システム 井上 吉雄ら 短波長赤外反射特性に基づいた作物収穫残渣の土壌被覆率計測手法 吉松 慎一 ヤガ科害虫4グループ類似種の幼生期の識別法 篠原 弘亮ら 健全イネでの常在が明らかになった Pseudomonas huttiensis の再分類 平成 14 年度(農業環境研究成果情報 第 19 集) 桑原 雅彦ら イネは土壌からダイオキシン類を吸収しない 清家 伸康ら 化学資材による土壌中ダイオキシン類の分解 小原 裕三ら 土壌表層中ダイオキシン類の光分解反応実験装置の開発 牧野 知之ら 連続遠心分離法を用いた土壌懸濁物質の粒径別ダイオキシン類濃度の測定 谷山 一郎ら 湖沼底質中ダイオキシン類の流域土地利用別寄与率の推定 板橋 直ら 流域単位で河川への懸濁物質流出負荷量を算定するモデルの作成 櫻井 泰弘ら 土壌溶液中カドミウム濃度による玄米中カドミウム濃度の推定 織田 久男ら 113 阿江 教治ら ダイズの子実カドミウム低蓄積系統における根の役割 江口 定夫 浅層地下水位変動の解析による水と硝酸イオンの移動経路・フラックスの推定 竹内 誠ら 水質モニタリングデータの図形表示・データ集約システムの作製 高木 和広ら 分解菌集積木質炭化素材を用いたシマジンの現地におけるバイオレメディエーション 高木 和広ら モミガラ成形炭粉末を用いた水稲用除草剤の系外流出削減技術 堀尾 剛ら メダカを使って化学物質の内分泌かく乱作用を簡易に検出する 松尾 和人ら キセニアを利用したトウモロコシ交雑率の簡易調査法 松本 直幸ら 紋羽病菌に見出された菌類ウィルス由来の病原力低下因子の発見 Cd 標識肥料を用いた肥料由来カドミウムの土壌負荷量の推定 202 平成 15 年度(農業環境研究成果情報 第 20 集) 星野(高田)裕子ら スキムミルクを用いた黒ボク土からの DNA 抽出法 藤井 毅ら キトオリゴ糖と特異的に作用するレセプターの検出法の開発 杉江 元 分布を拡大しているハイマダラノメイガの雄成虫を誘引する物質の発見 平舘俊太郎ら 葉面バクテリアが生産する抗微生物活性物質の化学構造の解明 池田 浩明ら 湿性植生による土壌懸濁物質の捕捉機能の評価 小林 和彦ら FACE(開放系大気 CO2 増加)圃場では米収量が増え,タンパク含量が減る 宮田 明ら 湿地ツンドラにおける二酸化炭素とメタンの年間収支とその年次変動 八木 一行ら 水田の間断灌漑水管理・慣行施肥管理は水稲栽培期間中のメタンと亜酸化窒素の発生を 共に抑制する 吉松 慎一ら 3種のヤガ科新害虫の九州以北における発生の確認と同定 月星 隆雄ら 日本野生植物寄生・共生菌類目録の作成と Web 上での公開 望月 淳ら 飼育昆虫・ダニ類データベースの作成と Web 上での公開 平成 15 年度(農業環境研究成果情報 第 20 集) 清家 伸康ら 水田土壌中のダイオキシン類起源の推移 牧野 知之ら 凝集剤による水田からのダイオキシン類の流出防止法 織田 久男ら ストロンチウム同位体比を利用したネギの産地国判別 村上 政治ら 畑条件で栽培するイネはカドミウム汚染水田の修復に最適である 杉山 恵ら 子実カドミウム蓄積性が高いダイズ品種は幼植物の段階で簡易に検定できる 江口 定夫 硝酸性窒素の浅層地下水および第二帯水層への到達時間 三島慎一郎ら 農業生産に伴う養分収支を都道府県・市町村単位で算出するシステム 森本 晶ら PCR-DGGE 法による土壌中のクロロ安息香酸分解菌群の検出 石原 悟ら 日本産珪藻および藍藻を用いた OECD 藻類生長阻害試験法の改良 石井 英夫 PCR-Luminex 法を用いてイネいもち病菌の MBI-D 剤耐性菌を遺伝子診断する 石井 英夫 アシベンゾラル S メチルによるナシの黒星病抵抗性誘導には PGIP が密接に関与する 山村 光司ら リンゴ火傷病が日本に侵入するリスクの推定法 土屋 健一ら ショウガ科植物に寄生する外来性青枯病菌系統の侵入と伝搬 澤田 宏之ら マメ科植物根粒菌のモニタリング手法開発のための 16S rDNA 情報 野口 浩ら チャノコカクモンハマキの交信撹乱剤抵抗性系統の作出と抵抗性の要因 野口 浩ら チャノコカクモンハマキの性フェロモン構成成分比の地理的変異 池田 浩明ら イヌホタルイのスルホニルウレア系除草剤の抵抗性遺伝子は集団中に急速に広まる 藤井 義晴ら 土壌を用いた他感作用の検定手法の開発 藤井 義晴ら ソバ属植物のアレロパシーとソバを利用した植生管理 平舘俊太郎ら 土壌中の腐植酸を構成する炭素に占めるススキ由来炭素の割合 鳥谷 均ら 衛星画像を用いたモンスーンアジアでの主要穀物の栽培期間の推定 桑形 恒男ら 局地気象モデルを活用した水田の水温・地温の広域的な推定手法の開発 宮田 明 水稲単作田の熱収支と CO2 フラックスの通年データセットの構築 秋山 博子ら 農耕地への有機物施用は亜酸化窒素の主要な排出源のひとつである 川島 茂人ら トウモロコシ花粉飛散量自動モニター装置の開発 織田健次郎 わが国の食料供給システムにおける 1980 年代以降の窒素収支の変遷 デイビッド・スプレイグら 迅速測図を用いて過去 100 年間の土地利用変化を定量的に計測する 藤原 英司ら 天然放射性核種 210Pb は乾燥地草原での風食を示す指標として有効である 農業環境技術研究所 研究成果 203 馬場 浩司ら 玄米に含まれるカドミウムのレーザーを利用した直接定量法の開発 井上 吉雄 多周波マイクロ波は全天候下で作物群落特性のリモートセンシングを可能にする 井上 吉雄 熱赤外リモートセンシングによる表面温度は土壌面 CO2 フラックスの広域評価に 有効である 三中 信宏ら 遺伝子情報に基づく巨大系統樹推定プログラムの開発 中井 信ら 土壌情報の一元的収集システムの開発 中谷 至伸ら 農業環境技術研究所が所蔵する昆虫タイプ標本一覧表ならびに画像の Web 上での公開 對馬 誠也ら 分散型データベースによる「微生物インベントリー」の構築と Web 上での公開 西山 幸司 凍結保存細菌の反復利用効率を高めるための分散媒の改良 平成 16 年度(農業環境研究成果情報 第 21 集) 上垣 隆一ら 水田土壌におけるダイオキシン類の年間収支の推定 菅原 和夫ら ダイズ子実のカドミウム濃度を土壌属性から予測するための手法 牧野 知之ら カドミウムで汚染された水田を修復するための土壌洗浄法 川崎 晃ら 水耕栽培したダイズにおけるカドミウムの移行特性 石川 覚ら 玄米のカドミウム濃度に係わる遺伝子座の検索 鈴木 克拓ら 亀裂の発達した粘土質転換畑の暗渠からのリンの流出 駒田 充生ら 畑土壌からの嫌気的な亜酸化窒素発生に脱窒菌以外の微生物が関わっている 中島 泰弘ら 台地畑における浅層地下水中の亜酸化窒素の由来 竹内 誠ら 畑ほ場における窒素動態・収支シミュレーションシステム 小川 直人ら クロロカテコール分解酵素の基質特異性は3つのアミノ酸によって決定される 髙木 和広ら シマジン分解細菌群 CD7 の構成種とその機能 大津 和久 農薬生態毒性評価のためのコガタシマトビケラの室内人工飼育法 石井 英夫 アシベンゾラルSメチルは葉面糸状菌相に影響を及ぼさない 鈴木 健 解毒酵素遺伝子塩基配列の違いを利用した有機リン剤抵抗性ワタアブラムシの識別法 山村 光司 環境のゆらぎを考慮した花粉拡散距離の推定法 松井 正春ら アジア・太平洋外来生物データベースシステムの構築 星野(高田)裕子ら 土壌抽出 DNA を用いた土壌くん蒸処理の微生物群集への影響評価 土屋 健一ら 農業環境に由来するセパシア菌群(Burkholderia cepacia complex)の遺伝子型 平舘俊太郎ら ユキヤナギに含まれる高活性植物生育阻害物質の発見 藤井 義晴ら 日本在来の被覆植物リュウノヒゲの他感作用と他感物質としてのサリチル酸の発見 藤井 義晴ら ムクナの根から放出される L−ドーパに対する植物の抵抗性機構 藤井 義晴ら 外来植物ニセアカシアのアレロパシー活性と作用物質 藤井 義晴ら 間伐材から抽出された植物生育促進物質 1, 2 - プロパンジオール 芝池 博幸ら 関東地方でみられるタンポポの4割は単一のクローンである 荒城 雅昭ら くん蒸処理が畑の土壌線虫の種数および密度に及ぼす影響と回復の内容 石郷岡康史ら 農業水利用を考慮した新しい大陸スケールの水循環モデルの開発 間野 正美ら 渦相関法による CO2 フラックス計算用の実用的プログラム 西村 誠一ら メタン発生の多い水田での畑転換は有効な温室効果ガス発生抑制技術である 渡辺 栄喜ら 農作物中の殺虫剤イミダクロプリドは簡易かつ迅速に分析できる 大野 宏之ら 水稲作付地を衛星搭載レーダと地理情報を用いて確実に精度良く検出する技術 井上 吉雄 リモートセンシングと生態プロセスモデルの協働による CO2 フラックスの動的 評価手法 204 平成 18 年度(研究成果情報 第 23 集) 吉松 慎一ら 侵入害虫クロテンオオメンコガ(新称)の国内における広範囲での発生確認 平成 17 年度(農業環境研究成果情報 第 22 集) 坂西 研二ら 中規模河川における懸濁性水質サンプルの採取位置による濃度差は小さい 酒井 順子ら 水田土壌から分離した 2, 4 - D 分解菌の分解遺伝子はメガプラスミドに存在する 藤井 毅ら キトビオースが誘導する放線菌の効率よいキチン分解機構 高木 和広ら クロロ及びメチルチオトリアジン系農薬を完全分解できる複合微生物系 石原 悟ら 珪藻の除草剤感受性を群集レベルの生育速度により簡易に検定する 松尾 和人ら 雑草群落に混生する野生化したアブラナ属植物の種間交雑種子の検出 藤井 毅ら キュウリ苗立枯病を抑える組換え微生物の土壌微生物相への影響 望月 淳ら 導入天敵昆虫等の新たな生態系影響評価方法と導入判断基準 田中 幸一ら 外来昆虫ブタクサハムシの飛翔による分布拡大能力 池田 浩明ら 浮遊シダ植物のサンショウモはスルホニルウレア系除草剤に対して感受性が高い 平舘俊太郎ら 土壌中における L-DOPA の植物生育阻害活性の消長 平舘俊太郎ら 固体 NMR によるアロフェン・イモゴライトの検出と Al 配位数解析 平舘俊太郎ら 13 量体 Al は根圏環境中で不安定なため生育阻害要因となりえない 平舘俊太郎ら 高感度・高精度・迅速なシアナミド定量法 藤井 義晴ら アオコの増殖抑制植物を検定する「リーフディスク法」の開発 楠本 良延ら 生物多様性保全のための景観・植生調査解析システム 吉本真由美ら 大気 CO2 濃度の上昇と群落微気象要因を考慮したイネの穂温推定モデル 秋山 博子ら 農耕地からの亜酸化窒素の排出係数は現在の IPCC デフォルト値よりも低い 井上 聡ら 積雪地域におけるスギ花粉の飛散開始日の推定 新藤 純子ら 食料生産・消費拡大がもたらす東・東南アジアの水質変動の予測 岩崎 亘典ら 野生動物の生息域拡大のしにくさを評価する手法の開発 山口 紀子ら XANES を用いた水田土壌中のヨウ素の非破壊形態分析とその溶脱機構 芝山 道郎ら 反射光の偏光特性を利用した作物個体群情報の計測方法 安田 耕司ら 昆虫文献目録「三橋ノート」の画像データベースの作成と公開 對馬 誠也ら 130 年にわたって採集された微生物さく葉標本の目録の作成と公開 對馬 誠也ら 植物生息細菌の定性・定量的解析とデータベースの構築 平成 18 年度(研究成果情報 第 23 集) (普及に移しうる成果) 大谷 卓ら ディルドリンを吸収しにくいカボチャ台木を用いてキュウリ果実中の残留濃度を低減 阿部 薫ら イムノクロマトアッセイを用いた玄米等のカドミウム濃度簡易測定法 八木 一行ら 2006 年版 IPCC ガイドラインに採用された水田から発生するメタンの新しい算定方法 秋山 博子ら 京都議定書第一約束期間の開始を前に,農耕地から発生する亜酸化窒素の新しい 排出係数を算定 齋藤 雅典ら 「水環境保全のための農業環境モニタリングマニュアル改訂版」の発行 (主要研究成果) 藤井 義晴ら コンフリーのアレロパシーと植物生育阻害物質の同定 藤井 義晴ら 小笠原父島に生育する樹木葉のアレロパシー活性の検定 伊藤 健二 特定外来生物カワヒバリガイ,霞ヶ浦に定着し分布域を拡大中 農業環境技術研究所 研究成果 205 吉村 泰幸ら カナダのナタネ輸出港周辺において遺伝子組換えナタネの分布は拡大しない 吉村 泰幸ら ほ場で遺伝子組換えダイズとツルマメが交雑する可能性は低い 松尾 和人ら 輸入港周辺の遺伝子組換えナタネは,従来のナタネ生育地にしか生育していない 田中 幸一ら トンボの生息環境としてのため池の特徴 楠本 良延ら 谷津田を囲む斜面林周辺の草刈りが生物多様性を高める 藤井 義晴ら ソバ粗抽出液に含まれる主要な植物生長阻害成分はルチン 杉江 元ら チャノコカクモンハマキの交信撹乱剤に対する抵抗性発現要因 酒井 英光ら 大気 CO2 増加による水稲の群落光合成の促進は葉窒素濃度に依存する 吉本真由美ら 大気 CO2 濃度の上昇は水田の水需要を減らす 岸本 文紅ら 土壌炭素動態の温暖化応答解明のための高精度土壌加温システム 山口 紀子ら 畑土壌における交換態放射性ストロンチウム減少速度は土壌の陽イオン交換能が 支配する 馬場 浩司ら 土壌及び作物中のフェニル置換ヒ素化合物の定量法 坂本 利弘ら 高頻度観測衛星センサー(MODIS/Terra)を活用してメコンデルタの洪水と稲作を 動的に捉える 井上 吉雄 日本の全流域圏・農耕地を網羅する農業生態系空間情報解析システム 吉松 慎一ら 菌床シイタケの新害虫をヤガ科ナミグルマアツバと同定 中谷 至伸 日本産ヒョウタンカスミカメ族(カメムシ目)の Web 図説検索表 平成 19 年度(研究成果情報 第 24 集) (普及に移しうる成果) 横山 淳史ら コガタシマトビケラ1齢幼虫を用いた農薬の急性毒性試験法マニュアル 小原 裕三ら 環境への負荷がより小さい低濃度エタノールを用いた低コストの新規土壌消毒法 高木 和広ら ゼロエミッションを目指したもみ殻ガス化残渣の有効利用 荒尾 知人ら スズメノナスビを台木としてナス果実中カドミウム濃度を低減 森本 晶ら 土壌 DNA による土壌生物相解析マニュアル 北本 宏子ら 葉の表面に棲む生分解性プラスチック分解酵母 桑形 恒男ら 水稲の温暖化影響評価のための「モデル結合型作物気象データベース」 須藤 重人 温室効果ガス3成分自動同時分析計の開発 大野 宏之ら 広域的な栽培暦情報を集積し,共有するための Web データベース (主要研究成果) 井上 恒久ら 渥美半島地域を対象とした農耕地土壌分類第3次改訂版に基づくデジタル農耕地土壌図 の作成 楠本 良延ら 水田周辺ではどのような外来植物に注意すべきか 小沼 明弘ら 輸入穀物に種子が混入しやすい外来植物の特徴 藤井 義晴ら 要注意外来植物ハリエンジュにもシアナミドが含まれる 山谷 紘子ら 外来植物アカギに含まれる植物生育阻害物質はL−酒石酸 藤井 義晴ら 簡易型雑草リスク評価法の提案とこれによるリスク評価 木方 展治ら 作物可食部のウラン濃度が低いことをほ場試験で確認 川崎 晃ら 有機性廃棄物の施用に伴うカドミウムの農地負荷量と作物中カドミウム濃度への影響 荒尾 知人ら カドミウム非汚染水田における年間カドミウム収支 林 健太郎ら 窒素追肥によりイネがアンモニアを放出する 神山 和則ら 1㎞メッシュ単位で推定した農業統計のデータベース化 206 平成 20 年度(研究成果情報 第 25 集) 西森 基貴ら 日本で使用された農薬のゆくえを明らかにできる地球規模の挙動予測モデル 王 勇ら 土壌中で生育している微生物の遺伝子の発現を検出した 麓 多門ら 水田からのメタン発生量を推定する数理モデル・DNDC Rice の開発 對馬 誠也ら 日本に生息するサトウキビ白すじ病菌の遺伝子型 對馬 誠也ら 温室栽培トマトと露地栽培トマトの葉面生息細菌相は著しく異なる 天野 達也ら 水田地帯では周辺環境によって生息する鳥類が異なる 江口 定夫ら 黒ボク土畑下層における不均一な水移動の定量化 江口 定夫ら 台地に隣接した水田下の浅層地下水中における脱窒の定量化 加藤 英孝ら 土壌特性・土層構成に基づく硝酸性窒素の地下水到達時間の面的予測 荒城 雅昭 土壌線虫の簡易同定に役立つ画像付形質一覧表の公開 吉本真由美ら 未来の水田生態系を予測する周年開放系温暖化実験システム 長谷川利拡ら 天水田地帯の水稲収量を推定する簡易モデル 藤原 英司 黄砂とともに飛来する放射性セシウム(137Cs) 井上 吉雄ら 群落光吸収率・光利用効率の遠隔評価 のための新反射スペクトル指数 井上 吉雄 空間情報技術による山岳アジア焼畑生態系の土地利用履歴と炭素ストックの広域評価 平成 20 年度(研究成果情報 第 25 集) (普及に移しうる成果) 清家 伸康ら キュウリのディルドリン残留濃度を予測できる土壌抽出法 大谷 卓ら カボチャのヘプタクロル汚染対策技術 小板橋基夫ら 植物の葉から採れたカビが生分解性プラスチックを強力に分解 杉江 元ら 性フェロモンを利用したナシマダラメイガの発生予察用誘引剤 秋山 博子ら 温室効果ガスの可搬型自動サンプリング装置の開発 岩崎 亘典ら 明治時代初期の関東地方の土地利用をインターネットで閲覧できるシステムを開発 吉松 慎一ら 北海道内の広域で発生した新害虫はヘリキスジノメイガだった (主要研究成果) 岩船 敬 河川における水稲用除草剤の分解物の消長と藻類に対する急性毒性 高木 和広ら ヘキサクロロベンゼン(HCB)を好気的に分解・無機化する細菌の単離・同定と 主要代謝経路の解明 大谷 卓ら ズッキーニを利用した土壌残留ディルドリンのファイトレメディエーション 川崎 晃ら 安定同位体を用いて植物に吸収されうるカドミウムの最大量を評価する 石川 覚ら イネ品種間差を利用して,玄米のカドミウム汚染を低減 西田 智子ら 日本における導入前雑草リスク評価法の構築 平舘俊太郎ら 外来植物の侵入は土壌 pH と有効態リン酸に関連している 望月 淳ら ミトコンドリア遺伝子の部分配列による土着天敵ヤマトクサカゲロウと外来近縁種との 識別法 山中 武彦ら ため池の多様なトンボ類を守るためには,池の環境だけではなく,池の配置も重要です 長谷川利拡ら 2007 年夏季異常高温下での水稲不稔率の増加を確認 長谷川利拡ら 水田からのメタン放出に対する大気二酸化炭素濃度と夜温上昇の相互作用 新藤 純子ら 東南アジアにおけるバイオ燃料増産による環境への窒素負荷の評価 井上 聡ら 極低温条件でも降水の量と継続時間を正確に長期間保守なしで実測できる測器の検証 石塚 直樹ら 東アジアの土地被覆・土地利用の時間的変化を広範囲で捉えるための植生指数データ セット 農業環境技術研究所 研究成果 207 上田 義治ら 農業環境インベントリーシステムの Web 公開 荒城 雅昭ら 国内における線虫学関連文献目録の公開 平成 21 年度(研究成果情報 第 26 集) (普及に移しうる成果) 殷 熙洙ら 水浄化を目的とした微粉末活性炭タブレットの開発 村上 政治ら カドミウム高吸収イネ品種によるカドミウム汚染水田の浄化技術 (ファイトレメディエーション) 福岡 峰彦ら 低コストで高精度の気温測定を可能にする強制通風筒 桑形 恒男ら 時別日射量の高精度推定法の開発と日本全域におけるデータセットの作成 八木 一行ら 世界の水田からのメタン発生量とその削減可能量の推定 木方 展治ら 主要穀類および農耕地土壌の人工放射性核種(90Sr,137Cs)分析データの インターネット上への公開 殷 熙洙ら 水中の有害な有機性化学物質を対象とした分析マニュアル 高田 裕介ら 農耕地土壌に関する情報を WEB 上で閲覧するシステムの公開 (主要成果) 髙木 和広ら ディルドリンを好気的に分解する糸状菌の単離 髙木 和広ら メラミン分解細菌を土壌中から発見 永井 孝志 農薬による環境リスクの大きさを計算する確率論的評価法 横山 淳史ら 殺虫剤の分解物は河川で水生節足動物に影響を及ぼしているのか? 石川 覚ら 玄米カドミウム濃度を制御する新規の遺伝子座の同定 石川 覚ら 稲やナスのカドミウム集積は導管にカドミウムを輸送する能力に支配される 屋良佳緒利ら 侵入害虫クリタマバチ防除のために導入された外来天敵昆虫とそれに近縁な 日本在来昆虫の交雑個体の検出法 藤井 義晴ら ナギナタガヤのアレロパシーとそのアレロケミカルの同定 山中 武彦 侵略的外来昆虫は,密度の低い侵入初期に根絶が容易 ― 交尾探索行動を仮定した新しいモデルによる解析 ― 藤井 義晴ら ナガミヒナゲシはアレロパシー活性が強く,雑草化リスクが大きいので, 広がらないようにする必要があります 楠本 良延ら 茶生産のために維持される茶草場は貴重な二次的自然の宝庫です 北本 宏子ら 飼料作物から牛の餌とバイオエタノールを作る 西森 基貴ら 2009 年夏の低日照が特に西日本の水稲作況低下に影響したことを 農環研データベース MeteoCrop で解明 秋山 博子ら 農耕地から発生する亜酸化窒素の削減効果を定量的に評価 ― 硝化抑制剤入り肥料の平均的な削減率は−38% ― 白戸 康人ら 有機物施用が及ぼす農地土壌への炭素蓄積効果を全国推定 坂西 研二ら 露地野菜畑における土壌流亡や栄養塩流出を解析するためのモニタリングシステム 金 元植ら 耕地の水消費と CO2 吸収のオンライン監視用ソフトウェア 芝山 道郎ら 作物からの分光反射率画像を長期間自動的に記録し生育状態を追跡するシステム 三島慎一郎ら 農耕地での窒素・リン酸収支を都道府県単位で算出したデータベース 吉松 慎一ら 農業環境技術研究所に寄贈された昆虫タイプ標本 379 点の公開 堀田 光生ら バイオエタノール生産に用いるバイオマス原料および発酵条件の簡便な評価方法 吉武 啓ら 昆虫文献目録「三橋ノート」コウチュウ目 135 冊の画像公開 208 平成 22 年度(研究成果情報 第 27 集) 平成 22 年度(研究成果情報 第 27 集) (普及に移しうる成果) 牧野 知之ら 山本 勝利ら 塩化鉄によるカドミウム汚染水田の実用的土壌洗浄技術 生物多様性に関する情報を収集,蓄積,提供するための WEB 版農業景観調査情報システム (RuLIS WEB) 福岡 峰彦ら イネ群落内の微気象を捉える自立型気象観測パッケージ「MINCER」 桑形 恒男ら 気 象, 土 壌, 農 地 利 用, 温 室 効 果 ガ ス に 関 す る 情 報 を ま と め て 取 得 で き る web シ ス テ ム (gamsDB) 阿部 薫ら イムノクロマトキットを用いた農産物のカドミウム濃度の簡易測定法 吉松 慎一ら 南西諸島で多発生した害虫は目本初発生のアフリカシロナヨトウである 吉武 啓ら 昆虫データベース統合インベントリーシステム 小原 洋ら 全国土を詳細に区分できる包括的土壌分類第 1 次試案 (主要成果) 清家 伸康ら 稲生 圭哉ら ディルドリン汚染地では,ウリ科作物,根菜類,イモ類の栽培を避けるのが適切 水田で使用する農薬の河川における濃度変化を予測し地図上に表示するシミュレーションモデ ルの開発 前島 勇治ら 土壌中におけるジフェニルアルシン酸の化学形態変化とイネヘの移行 山口 紀子ら スズメノナスビの根によるカドミウム吸収抑制を可視化 荒尾 知人ら 農作物中のカドミウム低減対策技術集 小沼 明弘ら 西オーストラリアからの輸入小麦に混入していた除草剤抵抗性雑草種子 加茂 綱嗣ら 天敵昆虫ナミテントウの幼虫に対する農地周辺植生に生息するアブラムシの餌適性を解明 山本 勝利ら 特定外来生物ナガエツルノゲイトウは水田に侵入し拡散する 吉村 泰幸 ダイズ花粉の空中飛散量は少なく風による飛散距離も短い 松尾 和人ら 道路沿いのセイヨウナタネは草刈りなどの撹乱が多い環境に生育しやすい 芝池 博幸ら 大規模圃場で水稲の自然交雑率を実測しました 杜 明遠ら 防風ネット・防風植生による交雑抑制効果を評価する数値モデル 田中 幸一ら 有機・減農薬栽培は水田の水生昆虫にプラスの効果があり,その効果は周辺の環境によって違 う 池田 浩明ら 水田,休耕田,河川における湿生多年草タコノアシの個体群動態に及ぼす除草剤の影響 藤井 義晴ら 目本原産のシランのアレロパシーとその原因物質の同定 釘宮 聡一ら コナガの天敵である寄生蜂コナガサムライコマユバチが寄主探索で利用する植物揮発性成分 吉本真由美ら 国際観測ネットワークでイネの高温障害を解明する 石郷岡康史ら 広域水収支モデルにより天水田地帯の水稲作付面積を推定する 長谷川利拡ら 須藤 重人ら イネ生育期間中の CO2・温度上昇は水田からのメタン発生量を大幅に増加させる 「三次メッシュ気象値ファイル」の WEB ブラウジング化プログラム 白戸 康人ら 日本の水田と黒ボク士畑に適合する改良 RothC モデル 林 健太郎 化学肥料の施肥由来のアンモニア発生インベントリー 江口 定夫ら 日本の農地土壌の物理的性質データベース So1phyJ の作成 中島 泰弘ら 窒素・酸素安定同位体自然存在比を用いた地下水中の硝酸イオンの起源推定と脱窒過程の評価 板橋 直ら 浅層地下水を通じた環境負荷物質の混合・減衰と濃度分布を面的に予測するための GIS モデル MacT の開発 板橋 直ら 農地由来の硝酸性窒素による地下水汚染リスク評価システム RealN の開発 宮田 明ら ガスフラックスのモニタリングにもとづくイネ単作田の年間炭素収支とその年次間変動 農業環境技術研究所 研究成果 209 米村正一郎ら 温度が土壌からの温室効果ガスの発生・吸収に及ぼす影響を自動連続測定できる実験装置 杜 明遠ら チベット高原における草原生存極限(草線)の気候条件 間野 正美ら WEB 上で手軽に実行できる渦相関法データ処理システム デイビッド・スプレイグ 對馬 誠也ら 「谷津田」に代表される枝流水田の分布状況の空間指標としての役割 赤かび毒デオキシニバレノールを分解する細菌物 Nocardioides sp. WSN05 - 2 株をムギ栽培圃場 から分離 吉松 慎一ら 昆虫文献目録「三橋ノート」全 476 冊の画像の完全公開 吉武 啓ら 土生コレクション・オサムシ科の標本目録 大澤 剛士ら オサムシ科標本情報閲覧システム 大倉 利明ら 微地形区分による農地の精密土壌図の作成 211 農業環境問題と研究の流れ 年 国 際 国 内 農業環境技術研究所 2011 (平 23) ・世界人口 70 億人(予測) ・国連生物多様性の 10 年 (2011 ― 2020)の開始 ・コメのカドミウム基準値 「0.4 mg/kg 以下」の施行(2 月) ・衛星情報・空間情報の先進的利 用に関する MARCO 国際ワーク ショップ(3 月) ・土・水研究会「温暖化緩和策と 土・水圏の物質循環研究の接点」 (2 月) 2010 (平 22) ・国連気候変動締約国会議 (COP16,COP/MOP6)( メ キシコ・カンクン) (12 月) ・生物多様性条約第 10 回締約 国会議(COP10)(10 月) 遺伝資源へのアクセスと 利 益 配 分(ABS) に 関 す る名古屋議定書 ポスト 2010 年目標(愛知 目標) SATOYAMA イ ニ シ ア ティブ国際パートナー シップ創設 ・カルタヘナ議定書第 5 回締 約国会議(MOP5)(10 月) 「責任と救済」に関する名 古屋・クアラルンプール 補足議定書 ・ロシア西部での熱波・干ば つ(6-8 月)により小麦が不 作・輸出停止へ ・国連「国際生物多様性年」 ・行政刷新会議事業仕分けで 大型研究開発事業のほか科 学技術振興や研究者育成の 予算に厳しい判定(11 月) ・ 平 成 21 年 度 の 食 料 自 給 率 (カロリーベース)が 40%に (8 月) ・夏の高温によるコメの不 作・品質低下など農業への 影響が問題化(7-9 月) ・農用地土壌汚染防止法に基 づくカドミウム汚染農地の 指定要件を改正(6 月) ・食品衛生法に基づくコメの カ ド ミ ウ ム 基 準 値 を「1.0 mg/kg 未満」から「0.4 mg/ kg 以下」に改正(4 月) ・生物多様性国家戦略 2010 を 決定(3 月) ・食料・農業・農村基本計画 を決定(3 月) 2020 年度食料自給率 50% を目標 ・農業環境技術公開セミナー in 熊 本(12 月) ・研究成果発表会 2010「未来につ なげよう安全・安心な農業と環 境」(11 月) ・アジア太平洋地域における持続可 能な食料生産のための機能的生物 多様性の増強に関する MARCOFFTC 国際セミナー(11 月) ・研究会「農薬の作物残留と大気中 挙動 −規制と研究−」(10 月) ・農業環境シンポジウム「農業から みた生物多様性,生物多様性から みた農業」(9 月) ・水田管理と温室効果ガス発生・吸 収 に 関 す る MARCO/GRA 合 同 ワークショップ(9 月) ・土壌洗浄法によるカドミウム汚染 水田の低コスト浄化技術(8 月) ・つくばみらい水田 FACE 実験を 開始 ・農業環境シンポジウム「21 世紀 の農業と環境問題を考える」(5 月) ・農耕地土壌に関する情報をイン ターネットで公開(4 月) ・土・水研究会「食の安全,農業環 境問題におけるトレードオフ」(2 月) ・農業統計情報メッシュデータ閲覧 システムを公開(2 月) 2009 (平 21) ・国連気候変動締約国会議 (COP15,COP/MOP5)( デ ンマーク・コペンハーゲン) (12 月) ・POPs 条約の第 4 回締約国会 議(COP4)開催 残留性有機汚染物質 (POPs) と し て 9 物 質 を 追加 ・ 国 連 環 境 計 画(UNEP) が 報 告 書「 グ ロ ー バ ル・ グ リーン・ニューディール: 政策の概要」を公表(3 月) ・オーストラリアで記録的な 干ばつ・熱波と森林火災(1 月) ・京都議定書約束期間(2008 ― 2012)の初年度(2008 年) の総排出量(基準年比 1.9% 増)の公表(11 月) ・2009 年夏の低日照により水 稲作況が全国的に低下 ・国連気候変動サミットで, 日本の温室効果ガス排出 量 を 2020 年 ま で に 90 年 比 25%削減することを表明(9 月) ・ 平 成 20 年 度 の 食 料 自 給 率 (カロリーベース)が 41%に (8 月) ・微粉末活性炭タブレット(錠剤) による水浄化技術(11 月) ・農業環境技術公開セミナー in 滋 賀(11 月) ・国際セミナー「農薬および POPs の土壌残留と食の安全」 (10 月) ・MARCO シ ン ポ ジ ウ ム「 モ ン スーンアジアにおける農業環境問 題と研究の挑戦」 (10 月) ・国際会議「東アジアの陸域生態系 における炭素窒素循環とその環境 影響」 (9 月) ・カドミウム高吸収イネ品種による 汚染水田の浄化技術(8 月) ・窒素循環モデルで環境への窒素流 出と水質汚染の変化を推定(8 月) 212 農業環境問題と研究の流れ 年 国 際 国 内 農業環境技術研究所 2009 (平 21) ・「アメリカ復興・再投資法」 が成立(2 月) 省エネ対策や再生可能エ ネ ル ギ ー 促 進, 水 質・ 土 壌汚染対策など推進 ・米国オバマ大統領が就任 (1 月) ・温室効果ガスの排出量削減 中期目標(2005 年比 15%減) の発表(6 月) ・バイオマス活用推進基本法 成立(6 月) ・「 温 暖 化 影 響 総 合 予 測 プ ロ ジェクト」成果発表(5 月) ・農林水産分野における省 CO2 効果の表示の指針(4 月) ・温室効果ガス観測技術衛星 「いぶき」GOSAT 打ち上げ (1 月) ・複合微生物系による有害物質メラ ミンの分解・無害化技術(3 月) ・セルロース系バイオマスに対する バイオエタノール固体発酵技術 (3 月) ・イネの生育を推定できる気象デー タベース「MeteoCrop DB」を公 開(3 月) ・「情報:農業と環境」訪問者の累 計 100 万人に(3 月) ・気象環境研究会「開放系大気 CO2 増加(FACE)実験 ― 過去 , 現在 , 未来 ― 」 (2 月) 2008 (平 20) ・国連気候変動締約国会 議(COP14,COP/MOP4) ( ポ ー ラ ン ド・ ボ ズ ナ ン ) (12 月) ・ラムサール条約第 10 回締約 国会議(10 月) ・湿地システムとしての水田 における生物多様性の向上 (水田決議) ・国際連合環境計画(UNEP) 「 グ ロ ー バ ル・ グ リ ー ン・ ニューディール」イニシア チブ発表(10 月) ・「 グ リ ー ン・ ニ ュ ー デ ィ ー ル」報告書発表(7 月) ・G 8 北海道洞爺湖サミット (7 月) ・ ロ ー マ で 食 料 サ ミ ッ ト(7 月) ・化学肥料の価格が急上昇 ・主要穀物やダイズの価格が 上昇,過去最高に ・農林水産省が食料自給率引 き上げの工程表を発表(12 月) ・農林漁業バイオ燃料法 施行 (10 月) ・事故米の不正転売・食用流 通が発覚(9 月) ・北海道で新害虫ヘリキスジ ノメイガが大量発生(8 月) ・ 平 成 19 年 度 の 食 料 自 給 率 (カロリーベース)が 40%に 増加(8 月) ・環境省報告書「気候変動へ の賢い適応」 (6 月) ・生物多様性基本法 成立(5 月) ・「 温 暖 化 影 響 総 合 予 測 プ ロ ジェクト」成果発表(5 月) ・農林水産省が食料安全保障 課を設置(4 月) ・京都議定書目標達成計画改 定(3 月) ・「 遺 伝 子 組 換 え 農 作 物 等 の 研究開発の進め方に関する 検討会」最終取りまとめ(1 月) ・京都議定書第一約束期間の 開始(1 月) ・「イネ科作物の葉の表面などに生 息する微生物が生分解性プラス チックを効率よく分解」が農林研 究成果 10 大トピックスに(12 月) ・公開セミナー「農業分野における メタゲノム解析技術の応用の可能 性」 (12 月) ・カドミニウム低吸収品種による 玄米カドミウム汚染低減技術(12 月) ・研究成果発表会 2008(11 月) ・国際セミナー「アジア・太平洋地 域における農業環境で問題となる 有害植物・昆虫」 (11 月) ・生分解性プラスチックを強力に分 解するカビを発見(10 月) ・MARCO ワ ー ク シ ョ ッ プ「 ア ジ アにおける自然資源管理のための 土壌情報システムの新しい取り組 み(10 月) ・農業環境技術公開セミナー in 福 島(9 月) ・シンポジウム「穀物の争奪戦が食 卓を襲う」(6 月) ・シンポジウム「温室効果ガス排出 をどう削減できるのか」(5 月) ・「歴史的農業環境閲覧システム」 を公開(4 月) ・2007 年夏季異常高温下での水稲 不稔率の調査結果を発表(3 月) ・生分解性プラスチックを強力に分 解する酵母菌を発見(3 月) ・気象環境研究会「2007 年夏季異 常高温が水稲生産に及ぼした影響 を検証する」(2 月) ・シンポジウム「農業・農村におけ る身近な生物多様性の保全と活用 をめざして」(2 月) ・谷津田における植物多様性の調査 結果を発表(2 月) 農業環境問題と研究の流れ 213 年 国 際 国 内 農業環境技術研究所 2007 (平 19) ・世界の陸上平均気温が観測 史上最高に(12 月) ・国連気候変動締約国会議 (COP13)( イ ン ド ネ シ ア・ バリ) (12 月) ・IPCC 第4次評価報告書 統 合報告書(11 月) ・アル・ゴア氏と IPCC がノー ベル平和賞を受賞(10 月) ・IPCC 第4次評価報告書 第 3作業部会報告書(気候変 動の緩和策) (5 月) ・IPCC 第4次評価報告書 第 2作業部会報告書(影響・ 適応・脆弱性) (4 月) ・IPCC 第4次評価報告書 第 1作業部会報告書(自然科 学的根拠) (2 月) ・米国でバイオエタノール生 産が本格化(1 月) ・中国産の冷凍食品から高濃 度 の 農 薬 を 検 出(12 月 〜 2008 年 2 月) ・「独立行政法人整理合理化計 画」閣議決定(12 月) 農 環 研, 生 資 研, 種 苗 管 理センターは平成 23 年 4 月に統合 ・「 第 三 次 生 物 多 様 性 国 家 戦 略」閣議決定(11 月) ・日本の最高気温記録を更新 する異常高温(8 月) ・農林水産省生物多様性戦略 (7 月) ・「21 世紀環境立国戦略」閣議 決定(6 月) ・農林水産省「地球温暖化対 策総合戦略」を策定(6 月) ・農林水産省「農地・水・環 境保全向上対策」を開始(4 月) ・農林水産省「有機農業の推 進に関する基本的な方針」 を公表 ・研究独法バイオ燃料研究推 進協議会 設立(4 月) ・農林水産研究基本計画 改定 (3 月) ・農林水産省「国産バイオ燃 料の大幅な生産拡大に向け た工程表」を発表(2 月) ・「環境への負荷がより少ない低濃 度エタノールを用いた低コストの 新規土壌消毒法の開発」が農林 研究成果 10 大トピックスに選定 (12 月) ・シンポジウム「温暖化によって何 が起こり,どう対応できるのか」 (12 月) ・低濃度エタノールによる新規土壌 消毒技術を発表(11 月) ・研究成果発表会「食と環境の安全 を求めて:農林水産生態系におけ る有害化学物質」 (11 月) ・公開セミナー「遺伝子組換え作物 の栽培と生態影響評価」 (11 月) ・農林水産地球温暖化対策研究連絡 協議会が発足(11 月) ・農地土壌炭素の変動実態の調査結 果を発表(11 月) ・国際シンポジウム「モンスーンア ジア農業生態系における侵略的外 来生物の実態と制御」 (10 月) ・ シ ン ポ ジ ウ ム「 食 料 vs エ ネ ル ギー ― 穀物の争奪戦が始まった ― 」(5 月) レスター・ブラウン氏基調講演 「バイオ燃料が食卓を脅かす」 ・特定外来生物カワヒバリガイの 霞ヶ浦での分布を発表(4 月) ・カボチャ台木によるキュウリ残留 農薬低減技術(1 月) 2006 (平 18) ・映画「不都合な真実」が公 開 ・南極のオゾンホールが過去 10 年間で最大規模に拡大(9 月) ・コメ(精米)のカドミウム の国際基準値が 0.4ppm に決 定(7 月) ・世界の遺伝子組換え作物の 作付面積が1億ヘクタール を超える ・国連「砂漠と砂漠化に関す る国際年」 ・有機農業推進法施行(12 月) ・カエルのツボカビ病を国内 で発見(12 月) ・宍道湖産シジミで残留基準 値を超える除草剤(チオベ ンカルブ)を検出(11 月) ・農林水産省が地球温暖化・ 森林吸収源対策推進本部を 設置(10 月) ・特定外来生物「セイヨウオ オマルハナバチ」規制開始 (9 月) ・国産カボチャのヘプタクロ ル残留が問題化(9 月) ・農産物の残留農薬に関する ポジティブリスト制度施行 (5 月) ・第三次環境基本計画(4 月) ・バイオマス・ニッポン総合 戦略改定(3 月) ・第3期科学技術基本計画(3 月) ・国産陸域観測技術衛星 ALOS-1( 愛 称「 だ い ち 」) 打ち上げ(1 月) ・「農耕地から発生する温室効果ガ スである亜酸化窒素の発生量を正 しく推定」が農林水産技術会議 10 大研究成果に選定(12 月) ・国際シンポジウム「モンスーンア ジアにおける持続的農業のための 農業資源の評価と有効利用」 (12 月) ・研究成果発表会 2006(9 月) ・豊橋技術科学大学と連携推進協定 を締結(9 月) ・国際ワークショップ「侵入生物に 対するアジア・太平洋外来生物 データベースの構築」 (9 月) ・「昆虫タイプ標本画像情報」公開 (6 月) ・「情報:農業と環境」への訪問者 の累計 50 万人(6 月) ・インベントリー展示館に肥料・煙 害展示室を新設(4 月) ・「三橋ノート画像データベース」 を公開(4 月) ・東京大学と教育研究協定を締結 (4 月) 214 農業環境問題と研究の流れ 年 国 際 2006 (平 18) 国 内 農業環境技術研究所 ・北海道遺伝子組換え作物交 雑防止条例 施行(1 月) ・研究所の理念・行動憲章・環境憲 章を制定(4 月) ・研究所組織の変更(研究領域, リサーチプロジェクトの設置) (4 月) ・第2期中期計画の開始(4 月) ・「水環境保全のための農業環境モ ニタリングマニュアル 改訂版」 発行(3 月) ・日韓共同国際ワークショップ「農 業生態系における窒素負荷と河 川・地下水への流出」 (3 月) ・国際ワークショップ「モンスーン アジア農業生態系における温室効 果ガス発生」 (3 月) 2005 (平 17) ・国連気候変動会議(COP11 & COP/MOP1)( モ ン ト リ オール) (11 月 ― 12 月) ・ 大 型 の ハ リ ケ ー ン( カ ト リーナ)により米国南東部 に甚大な被害(8 月) 原油価格の急上昇とエタ ノール生産の増大 ・国連ミレニアム生態系アセ スメント総合報告書(3 月) ・京都議定書 発効(2 月) ・食品の安全性に関するリス ク管理の標準手順書(6 月) ・POPs 条約に基づく国内実施 計画(6 月) ・湖沼水質保全特別措置法 改 正(6 月) ・窒素の面源負荷対策を強化 ・外来生物被害防止法施行(6 月) ・京都議定書目標達成計画(4 月) ・環境と調和のとれた農業生 産活動規範(農業環境規範) (3 月) ・新 農林水産研究基本計画(3 月) ・新 食料・農業・農村基本計 画(3 月) ・「カドミウムで汚染された水田の 土壌洗浄法による修復」が農林水 産技術会議事務局 10 大研究成果 に選定(12 月) ・初めての環境報告書公表(12 月) ・成果発表会「食と環境の安全を求 めて:有害化学物質のリスク評価 と低減技術」 (11 月) ・シンポジウム「モンスーン・ア ジアの農業とフード・セキュリ ティー」 (11 月) レスター・ブラウン氏基調講演 ・シンポジウム「農業環境における リスク研究に果たすインベント リーの役割と課題」 (10 月) ・農業環境インベントリー展示館を 公開(4 月) ・「農業環境研究 20 年の歩み」,「農 業環境研究の最前線」,「散策と思 索」を刊行(3 月) ・気候変動に関する日中韓国際シン ポジウム(2 月) 2004 (平 16) ・EU,組換え食品・作物の輸 入規制を解除(5 月) ・POPs 条約(残留性有機汚染 物質に関するストックホル ム条約)発効(5 月) ・国連「国際コメ年」 ・世界イネ研究会議東京シン ポジウム(11 月) ・カルタヘナ法 施行(2 月) 未承認遺伝子組換え生物 の 使 用( 栽 培, 飼 育。 販 売等)を禁止 ・農林水産省「第1種使用規 程承認組換え作物栽培実験 指針」 (2 月) ・高病原性鳥インフルエンザ の発生(1 月) ・「ダイズのカドミウム吸収抑制の ための対策技術」公開(12 月) ・アジア・太平洋外来生物データ ベ ー ス(APASD) の 公 開(11 月) ・国際ワークショップ「アジア太平 洋地域における侵入生物に対する データベース構築」 (11 月) ・日中ワークショップ「農業生態系 における窒素循環とその環境負 荷」 (10 月) ・環境研究機関連絡会成果発表会 「持続可能な社会をめざして」 (9 月) ・「情報:農業と環境」訪問者の累 計 20 万人に(4 月) ・ 研 究 成 果 発 表 会 2004 ― 安 心・ 安全な農業環境をめざして ― (4 月) 農業環境問題と研究の流れ 215 年 国 際 国 内 2004 (平 16) 農業環境技術研究所 ・国際ワークショップ「地球温暖化 に伴う東アジアの食料生産変動予 測」 (3 月) ・ボン大学開発研究センターとの MOU(3 月) ・研究所内より旧農事試験場におけ る肥料依頼分析の記録を発見 2003 (平 15) ・ペンタゴン・リポート発表 (10 月) ・バイオセイフティに関する カルタヘナ議定書が発効(9 月) ・第 3 回世界水フォーラム(3 月) ・内閣府に食品安全委員会設 置(7 月) ・農林水産省に消費・安全局 設置(7 月) ・食品安全基本法(7 月) ・カルタヘナ法(遺伝子組換 え生物等の使用等の規制に よる生物の多様性の確保に 関する法律)成立(6 月) ・農薬取締法改正(6 月) ・特定防除資材(特定農薬) の指定(3 月) ・茨城県神栖町の井戸水から 高 濃 度 の 有 機 ヒ 素 検 出(3 月) ・自然再生推進法施行(1 月) ・国際シンポジウム「農産物から重 金属汚染を低減するための農業戦 略とテクノロジー ― カドミウム 汚染リスクを減らす ― 」 (11 月) ・国際共同セミナー「アジア・太平 洋諸国における侵入生物による環 境影響とデータベース構築」 (11 月) ・環境研究機関連絡会成果発表会 「環境研究の連携をめざして」 (7 月) ・被害植物標本など明治から戦前期 の「煙害調査試料」を研究所内か ら発見 ・日中韓国際ワークショップ「東ア ジアの農業生態系における物質循 環と環境影響評価」 (3 月) 2002 (平 14) ・持続可能な開発ヨハネスブ ルグ世界サミット (WSSD2002) (8 ― 9 月) ・バイオマス・ニッポン総合 戦略(12 月) ・バイオテクノロジー戦略大 綱(12 月) ・農薬取締法改正(12 月) ・米政策改革大綱(12 月) ・中央環境審議会 遺伝子改変 生物の生物多様性影響防止 に関する中間答申(9 月) ・国内無登録農薬の輸入,使 用が問題化(8 月) ・京都議定書批准(6 月) ・地球温暖化対策推進法改正 (6 月) ・土壌汚染対策法(5 月) ・農水省「食」と「農」の再 生プラン(4 月) ・生物多様性国家戦略改訂(3 月) ・地球温暖化対策推進大綱改 訂(3 月) ・「水稲のカドミウム吸収抑制のた めの対策技術」を公開(10 月) ・中国科学院土壌科学研究所との MOU(7 月) ・農業環境技術研究所研究成果発表 会(4 月) ・農業環境技術研究所「友の会」発 足(4 月) 2001 (平 13) ・COP6 再 開 会 合(7 月)〜 COP7 開 催, 議 定 書 実 施 ル ー ル 合 意 (10 ― 11 月) ・国連ミレニアム生態系アセ スメントの開始(6 月) ・POPs 条約(残留性有機汚染 物質に関するストックホル ム条約)採択(5 月) ・IPCC 地球温暖化第3次評価 報告書 ・BSE 感染牛を初めて確認(9 月) ・食品リサイクル法 施行(4 月) ・新科学技術基本計画(3 月) ・中央省庁再編(1 月) 環境省 発足 内閣府に総合科学技術会 議 設置 ・環境研究機関連絡会発足(10 月) ・韓国農村振興庁農業科学技術院と の MOU(10 月) ・特定独立行政法人 農業環境技術 研究所 設立(4 月) ・農林水産省農業環境技術研究所 廃止(3 月) 216 農業環境問題と研究の流れ 年 国 際 国 内 農業環境技術研究所 2000 (平 12) ・CODEX バ イ テ ク 応 用 食 品 特別部会設置 ・バイオセイフティに関する カ ル タ ヘ ナ 議 定 書 採 択(1 月) ・WTO 農業交渉で農業の多面 的機能の論議 ・環境ホルモン戦略計画 SPEED'98 改定(11 月) ・加工食品への未認可 GM 混 入(10 ― 11 月) ・ダイオキシン類環境基準(7 月) ・循環型社会形成推進基本法 (6 月) ・食料・農業・農村基本計画 (3 月) ・有機 JAS マーク制定(3 月) ・農水省 遺伝子組換え農作物 環境安全性検討専門委員会 設置(2 月) ・環境研究三所連絡会発足(12 月) ・環境化学物質分析施設完成 ・公式ウェブサイトに Web マガジ ン「情報:農業と環境」を掲載開 始(5 月) 1999 (平 11) ・CODEX オ ー ガ ニ ッ ク 農 業 基準合意 ・ベルギー ダイオキシン汚染 事件 ・Bt トウモロコシのオオカバ マダラへの影響の論文 ・世界人口 60 億人 ・OECD が農業の多面的機能 の検討作業開始 ・農林水産研究基本目標(11 月) ・東海村 JCO 臨界事故(9 月) ・ 農 業 環 境 3 法( 持 続 農 業 法,家畜排泄物法,肥料取 締法改正) (7 月) ・ダイオキシン類対策特別措 置法(7 月) ・食料・農業・農村基本法(7 月) ・ 中 央 省 庁 等 改 革 関 連 法(7 月) ・天敵農薬に係る環境影響評 価ガイドライン(3 月) ・農作物ダイオキシン汚染報 道(2 月) ・独立行政法人農業環境技術研究所 準備委員会(11 月) ・農作物中放射性物質の緊急調査 (10 月) ・農業環境技術研究所推進方策委員 会(3 月) ・農業環境技術研究所将来方向検討 会(1 月) 1998 (平 10) ・WHO ダ イ オ キ シ ン TDI 見 直し ・COP4 ブエノスアイレス行 動計画 ・地球温暖化対策推進法(10 月) ・食料・農業・農村基本問題 調査会 答申 ・農政改革大綱 ・地球温暖化対策推進大綱(6 月) ・ 中 央 省 庁 等 改 革 基 本 法(6 月) ・環境ホルモン戦略計画 SPEED'98(5 月) ・イネ FACE 実験の開始(岩手県 雫石町) 1997 (平 9) ・ 温 暖 化 防 止 京 都 会 議 (COP3) 京都議定書採択 ・OECD 農業環境指標の枠組 みできる ・環境保全型農業推進憲章 ・環境影響評価法 ・地力増進基本指針 改正 ・環境ホルモン 中間報告 ・地下水の水質汚濁に係る環 境基準 1996 (平 8) ・コルボーンら「奪われし未 来」 ・世界食料サミット ・遺伝子組換え作物の商業生 産開始 ・APN(アジア太平洋地球変 動研究ネットワーク)発足 ・有機農産物・特別栽培農産 ・OECD 農業環境評価指標策定に 物表示ガイドライン(12 月) 向けたプロジェクト研究を開始 ・水質汚濁防止法 改正 ・研究基本計画 策定 ・大気汚染防止法 改正 ・農水省研究基本目標 ・科学技術基本計画 農業環境問題と研究の流れ 217 年 国 際 国 内 1995 (平 7) ・先進国のフロン,臭化メチ ル全廃 ・IPCC 第2次評価報告 ・世界貿易機構(WTO)発足 ・環境分野に初のノーベル賞 ・環境保全型農業総合推進事 業 ・農業基本法研究会 ・食糧管理法廃止,新食糧法 施行 ・科学技術基本法施行 ・生物多様性国家戦略 1994 (平 6) ・気候変動枠組み条約発効 ・砂漠化対処条約 ・環境基本計画決定 ・環境保全型農業推進本部 設 置 1993 (平 5) ・OECD「農業と環境」合同作 業部会設置,農業環境指標 検討開始 ・ウルグアイ・ラウンド農業 合意 ・アジェンダ 21 行動計画 ・環境基本法 ・大冷害,米輸入部分開放 ・有機農産物表示制度 ・特定農山村法 ・窒素,リンの水質汚濁防止 法排出基準 ・生物多様性条約批准 1992 (平 4) ・リオデジャネイロ地球環境 サミット(国連環境開発会 議) アジェンダ 21 採択,リオ 宣言 森林保全の原則声明 生物多様性条約 気候変動枠組条約 ・農水省環境保全型農業対策 室 ・農林水産省新政策公表(環 境保全型) 1991 (平 3) 農業環境技術研究所 ・研究所公式ウェブサイト開設 ・土壌の汚染に係る環境基準 ・地球環境モニタリング計画 ・レッドデータブック ・通産省 公害資源研究所を資 源環境技術総合研究所に改 組 ・遺伝子組換え植物隔離ほ場を日本 で初めて設置 1990 (平 2) ・IPCC 第1次評価報告書 ・シャロン・ローン 「オゾン・クライシス」 ・農林水産基本目標 ・環境庁 国立公害研究所を国 立環境研究所に改組 ・地球温暖化防止行動計画 ・地球科学技術基本計画 ・地球環境研究チーム設置 ・研究基本計画 策定 1989 (平元) ・CGIAR「持続可能な農業生 産」 ・有害廃棄物規制バーゼル条 約 ・北極にオゾンホール ・特定フロン全廃ヘルシンキ 宣言 ・農水省有機農業対策室 ・農林水産分野組換え体利用 指針 ・遺伝子組換え植物(トマト)の非 閉鎖系実験を日本で初めて開始 1988 (昭 63) ・環境と農業 EC 委員会 ・IPCC(気候変動政府間パネ ル)設置,IGBP 発足 ・窒素酸化物排出抑制 ソフィ ア議定書 ・米 LISA 研究プロジェクト 開始 ・オゾン層保護法 ・情報システム研究室と生物情報計 測研究室を新設 1987 (昭 62) ・WCED 報告「地球の未来を 守るために」 ・絶滅野生動植物譲渡規制法 ・遺伝子組換え体安全性評価研究を 開始 218 農業環境問題と研究の流れ 年 国 際 国 内 1987 (昭 62) ・オゾン層破壊物質モントリ オール議定書 ・世界人口 50 億人 ・特別栽培米制度 ・農林省が組換え体安全性評 価のプロジェクト開始 ・科学技術会議答申「国立試 験研究機関の中長期的あり 方について」 1986 (昭 61) ・チェルノブイリ原発事故 ・チェルノブイリ放射能汚染緊急調 査 1985 (昭 60) ・FAO 熱帯林行動計画 ・オゾン層保護ウイーン条約 ・ヘルシンキ議定書 ・研究基本計画 策定 1984 (昭 59) ・環境と開発に関する委員会 (WCED)発足 1983 (昭 58) 1982 (昭 57) ・湖沼水質保全特別措置法 ・世界湖沼環境会議開催 ・農水省バイテク室設置 ・地力増進法 ・農林水産研究基本目標 ・農業技術研究所 廃止 ・UNEP特別会議ナイロビ 宣言 ・南極でオゾンホール発見 ・遺伝子組換え植物の作成 ・緑資源の維持・培養と環境 保全の論議 1980 (昭 55) ・米政府調査報告「西暦 2000 年の地球」(地球温暖化・種 の消滅を警告) ・過疎地域振興特別措置法 ・ワシントン条約,ラムサー ル条約加入 ・農業技術研究所ほか 11 試験 研究機関が筑波農林研究団 地に移転 1979 (昭 54) ・スリーマイル島原発事故 ・世界気候会議(WMO)が温 室効果による温暖化を警告 ・科学技術庁が組換え DNA 実 験指針 1981 (昭 56) 1978 (昭 53) 農業環境技術研究所 ・農林省が農林水産省に改称 1977 (昭 52) ・砂漠化防止行動計画 ・気象衛星ひまわり1号打ち 上げ 1976 (昭 51) ・セベソダイオキシン汚染事 件 1975 (昭 50) ・ラムサール条約発効 ・アシロマ会議で遺伝子組換 え実験自主規制の合意 ・母乳から残留農薬検出 ・有吉佐和子 「複合汚染」 (PCB・水銀等汚染問題深 刻化) 1974 (昭 49) ・世界人口会議 ・世界食糧会議 ・ローランドら フロンによる オゾン層破壊の可能性を発表 ・国連砂漠化防止会議 ・世界人口 40 億人 ・環境庁 国立公害研究所設置 ・生産緑地法 ・農林水産省 農業環境技術研究所 設立(12 月) 農業環境問題と研究の流れ 219 年 国 際 国 内 1973 (昭 48) ・遺伝子組換え基礎技術の開 発 ・化審法(化学物質の審査及 び製造等の規制に関する法 律)制定 1972 (昭 47) ・ストックホルム人間環境宣 言 ・ローマ・クラブ「成長の限 界」 ・ 国 連 環 境 計 画(UNEP) 設 立 ・地球観測衛星ランドサット 1号打ち上げ ・自然環境保全法 ・PCB 生産中止 1971 (昭 46) ・ラムサール条約採択 ・水質汚濁に係る環境基準 ・環境庁発足 ・BHC・DDT 販売禁止 ・PCB 環境汚染問題化 1970 (昭 45) ・OECD 環境委員会設立 ・水質汚濁防止法 ・海洋汚染防止法 ・農用地土壌汚染防止法 ・( 光 化 学 ス モ ッ グ 被 害 深 刻 化) 1960 年代 (昭 35 〜 44) ・レイチェル・カーソン 「沈黙の春」 (1962) ・世界人口 30 億人(1960) ( 緑 の 革 命(1960 〜 70 年 代)) ・稲作転換対策開始(1969) ・大気汚染防止法(1968) ・カネミ油症事件(1968) ・公害対策基本法(1967) ・園芸・畜産拡大(1960 年代 前半) ・OECD 正式加入(1964) ・農業基本法(1961) 1950 年代 (昭 25 〜 34) ・カイコガの性フェロモン (ボンビコール)発見 (1059) ・降下放射性核種分析研究開 始(1957) ・PCB 生産開始(1954) ・水俣病発生(1953) ・PCB 輸入開始(1952) ・農林省 農業技術研究所 設置 (1950) 1940 年代 (昭 15 〜 24) ・BHC 農薬登録(1949) ・DDT 農薬登録(1948) 農業環境技術研究所 執筆者一覧 佐藤 洋平 独立行政法人農業環境技術研究所理事長 西尾 隆 独立行政法人農業環境技術研究所土壌環境研究領域長 與語 靖洋 独立行政法人農業環境技術研究所有機化学物質研究領域長 新藤 純子 独立行政法人農業環境技術研究所物質循環研究領域長 宮田 明 独立行政法人農業環境技術研究所大気環境研究領域長 谷山 一郎 独立行政法人農業環境技術研究所研究コーディネータ 八木 一行 独立行政法人農業環境技術研究所研究コーディネータ 安田 耕司 独立行政法人農業環境技術研究所生物多様性研究領域長 田中 宥司 独立行政法人農業環境技術研究所研究コーディネータ 藤井 毅 独立行政法人農業環境技術研究所生物生態機能研究領域長 三輪 哲久 独立行政法人農業環境技術研究所生態系計測研究領域長 對馬 誠也 独立行政法人農業環境技術研究所農業環境インベントリーセンター長 廉澤 敏弘 独立行政法人農業環境技術研究所広報情報室長 農業環境研究叢書 第18号 平成23年3月 発行 農業環境研究 2001 ― 2010 編 者 発行者 独 立 行 政 法 人 農 業 環 境 技 術 研 究 所 〒305 - 8604 茨城県つくば市観音台3 - 1 - 3 制作 筑波印刷情報サービスセンター協同組合 農業環境研究叢書刊行一覧 和文叢書 第 1号 農・林・水生態系へのアプローチ(昭和 61 年 3 月発行) 第 2号 環境中の物質循環(昭和 62 年 3 月発行) 第 3号 農林水産業における環境影響評価(昭和 63 年 3 月発行) 第 4号 農業環境を構成する生物群の相互作用とその利用技術(平成元年 3 月発行) 第 5号 微量元素・化学物質と農業生態系(平成 2 年 3 月発行) 第 6号 環境インパクトと農林生態系(平成 2 年 3 月発行) 第 7号 地球環境と農林業(平成 3 年 2 月発行) 第 8号 農村環境とビオトープ(平成 5 年 7 月発行) 第 9号 農林水産業と環境保全 −持続的発展を目指して−(平成 7 年 10 月発行) 第10号 水田生態系における生物多様性(平成 10 年 3 月発行) 第11号 21 世紀の食料確保と農業環境(平成 10 年 7 月発行) 第12号 農業におけるライフサイクルアセスメント(平成 12 年 3 月発行) 第13号 農業を軸とした有機性資源の循環利用の展望(平成 12 年 12 月発行) 第14号 遺伝子組換え作物の生態系への影響評価(平成 15 年 3 月発行) 第15号 農業生態系における炭素と窒素の循環(平成 16 年 2 月発行) 第 16号 農業環境研究 20 年の歩み(平成 17 年 3 月発行) 第17号 農業生態系の保全に向けた生物機能の活用(平成 18 年 1 月発行) 英文叢書(NIAES Series) No.1 Ecological Processes in Agro-Ecosystems, 1992 No.2 CH4 and N2O : Global Emissions and Controls from Rice Fields and Other Agricultural and Industrial Sources, 1994 No.3 Biological Invasions of Ecosystem by Pests and Beneficial Organisms, 1999 No.4 Plant Nutrient Acquisition : New Perspectives, 2001 No.5 Material Circulation through Agro-Ecosystems in East Asia and Assessment of its Environmental Impact, 2005
© Copyright 2026 Paperzz