JP 4270473 B2 2009.6.3 (57)【特許請求の範囲】 【請求項1】 C型肝炎ウイルス(HCV)E2抗原に対して免疫学的結合親和性を示す組換えヒトモノ クローナル抗体であって、該モノクローナル抗体が、ヒトγ1重鎖可変領域(VH)を含 むHCV E2抗原結合部分と、ヒトκ軽鎖可変領域(VL)のHCV E2結合部分と を含み、ここで、該VH領域が (i)図1A(配列番号1)、図1B(配列番号2)、図1C(配列番号3)、図1D( 配列番号4)、図1E(配列番号5)、図1F(配列番号6)、および図1G(配列番号 7)のいずれかひとつに示すアミノ酸の連続する配列、または (ii)(i)の配列のいずれかに少なくとも90%の同一性を有するアミノ酸の連続す 10 る配列、であり、そして VL領域が、 (a)図2A(配列番号8)、図2B(配列番号9)、図2C(配列番号10)、図2D (配列番号11)、図2E(配列番号12)、図2F(配列番号13)、および図2G( 配列番号14)のいずれかひとつに示される連続配列、または (b)(a)のいずれかひとつに少なくとも90の配列同一性を有するアミノ酸の連続配 列、 であり、ここで、 該モノクローナル抗体は、HCVゲノタイプ1aおよび1bからのE2抗原に対して交叉 反応性を有する、 20 (2) JP 4270473 B2 2009.6.3 モノクローナル抗体。 【請求項2】 前記Fab分子は、図1A(配列番号1)に示す重鎖可変領域(VH)アミノ酸配列を含 む、請求項1に記載のモノクローナル抗体。 【請求項3】 前記Fab分子は、図1B(配列番号2)に示す重鎖可変領域(VH)アミノ酸配列を含 む、請求項1に記載のモノクローナル抗体。 【請求項4】 前記Fab分子は、図1C(配列番号3)に示す重鎖可変領域(VH)アミノ酸配列を含 む、請求項1に記載のモノクローナル抗体。 10 【請求項5】 前記Fab分子は、図1D(配列番号4)に示す重鎖可変領域(VH)アミノ酸配列を含 む、請求項1に記載のモノクローナル抗体。 【請求項6】 前記Fab分子は、図2A(配列番号8)に示す軽鎖可変領域(VL)アミノ酸配列を含 む、請求項1に記載のモノクローナル抗体。 【請求項7】 前記Fab分子は、図2B(配列番号9)に示す軽鎖可変領域(VL)アミノ酸配列を含 む、請求項1に記載のモノクローナル抗体。 【請求項8】 20 前記Fab分子は、図2C(配列番号10)に示す軽鎖可変領域(VL)アミノ酸配列を 含む、請求項1に記載のモノクローナル抗体。 【請求項9】 前記Fab分子は、図2D(配列番号11)に示す軽鎖可変領域(VL)アミノ酸配列を 含む、請求項1に記載のモノクローナル抗体。 【請求項10】 前記Fab分子は、図1A(配列番号1)に示すアミノ酸の連続配列と、図2A(配列番 号8)に示す軽鎖可変領域(VL)アミノ酸配列とを含む、請求項1に記載のモノクロー ナル抗体。 【請求項11】 30 前記Fab分子は、図1B(配列番号2)に示すアミノ酸の連続配列と、図2B(配列番 号9)に示す軽鎖可変領域(VL)アミノ酸配列とを含む、請求項1に記載のモノクロー ナル抗体。 【請求項12】 前記Fab分子は、図1C(配列番号3)に示すアミノ酸の連続配列と、図2C(配列番 号10)に示す軽鎖可変領域(VL)アミノ酸配列とを含む、請求項1に記載のモノクロ ーナル抗体。 【請求項13】 前記Fab分子は、図1D(配列番号11)に示すアミノ酸の連続配列と、図2D(配列 番号11)に示す軽鎖可変領域(VL)アミノ酸配列とを含む、請求項1に記載のモノク 40 ローナル抗体。 【請求項14】 前記モノクローナル抗体は、C型肝炎ウイルス(HCV)E2抗原に結合する能力を有す るF(ab’)フラグメントを含む、請求項1に記載のモノクローナル抗体。 【請求項15】 前記モノクローナル抗体は、C型肝炎ウイルス(HCV)E2抗原に結合する能力を有す るFab分子である、請求項1に記載のモノクローナル抗体。 【請求項16】 前記モノクローナル抗体は、少なくとも約1.0×107の結合親和性を有することをさ らに特徴とする、請求項1∼15のいずれか1項に記載のモノクローナル抗体。 50 (3) JP 4270473 B2 2009.6.3 【請求項17】 該モノクローナル抗体は、ELISAアッセイによって決定されるC型肝炎ウイルス(H CV)E1/E2複合体にも結合することをさらに特徴とする、請求項16に記載のモノ クローナル抗体。 【請求項18】 請求項1∼17のいずれかに記載のモノクローナル抗体と、薬学的に受容可能なビヒクル とを含む、ワクチン組成物。 【請求項19】 請求項1∼17のいずれかに記載のモノクローナル抗体と、それに結合する検出可能な部 分とを含む、結合複合体。 10 【請求項20】 前記検出部分は、放射性同位体、蛍光剤、化学発光剤、酵素、酵素基質、酵素補因子、酵 素阻害剤、および色素からなる群より選択される、請求項19に記載の結合複合体。 【請求項21】 HCVを含むと予想されるサンプルにおいてHCV粒子の存在を検出する方法であって、 該方法は、 (a)請求項19または20のいずれかの結合複合体とともにサンプルをインキュベート する工程であって、該インキュベートは、抗体−抗原複合体の形成を可能にする条件下で 行われる、工程;および (b)前記抗体−抗原複合体の存在または不存在を検出する工程を包含する、方法。 20 【請求項22】 HCV感染した哺乳動物被験体の抗HCV治療処置の進行をモニターするための方法であ って、該方法は、該サンプルを、請求項19または20に記載の結合複合体とともにイン キュベートする工程であって、該インキュベートは、抗体−抗原複合体の形成を可能にす る条件下で行われる、工程;および 前記抗体−抗原複合体の存在または不存在を検出する工程を包含する、方法。 【請求項23】 請求項1∼17のいずれか1項に記載の組み換え抗体をコードする、単離された核酸分子 であって、ここで第一のヌクレオチド配列は、ヒトγ1重鎖可変領域(VH)HCV E 2抗原結合部分をコードし、そして第二のヌクレオチド配列は、ヒトκ軽鎖可変領域(V 30 L)のHCV E2抗原結合部分をコードする、核酸分子。 【請求項24】 請求項23に記載の単離された核酸分子であって、該分子は、ヒトFab分子をコードし 、前記ヒトγ1重鎖可変領域(VH)のHCV E2抗原結合部分をコードするヌクレオ チド配列は、 コンビナトリアルライブラリから得られるヒトFab分子のaγ重鎖可変領域(VH)の 結合部分をコードする第一のヌクレオチド配列を含み、ここで、該Fab分子は、C型肝 炎ウイルス(HCV)E2抗原に対する免疫学的結合親和性を示し、ここで、該第一のヌ クレオチド配列は、 (i)図4A(配列番号22)、図4B(配列番号23)、図4C(配列番号24)、図 40 4D(配列番号25)、図4E(配列番号26)、図4F(配列番号27)、および図4 G(配列番号28)のいずれかに示される連続配列;または (ii)(i)のいずれかに少なくとも90%の同一性を有する連続配列であり、 該Fab分子は、HCVゲノタイプ1aおよび1bからのE2抗原に対して交叉反応性を 有し、前記ヒトκ軽鎖可変領域(VL)のHCV E2抗原結合部分をコードするヌクレ オチド配列は、 コンビナトリアルライブラリから得られるヒトFab分子のaκ軽鎖可変領域(VH)の 結合部分をコードする第一のヌクレオチド配列を含み、ここで、該Fab分子は、C型肝 炎ウイルス(HCV)E2抗原に対する免疫学的結合親和性を示し、ここで、該第一のヌ クレオチド配列は、 50 (4) JP 4270473 B2 2009.6.3 (i)図3A(配列番号15)、図3B(配列番号16)、図3C(配列番号17)、図 3D(配列番号18)、図3E(配列番号19)、図3F(配列番号20)、および図3 G(配列番号21)のいずれかに示される連続配列;または (ii)(i)のいずれかに少なくとも90%の同一性を有する連続配列であり、 該Fab分子は、HCVゲノタイプ1aおよび1bからのE2抗原に対して交叉反応性を 有する、核酸分子。 【請求項25】 前記VHコード配列は、図4A(配列番号22)に示される、請求項24に記載の核酸分 子。 【請求項26】 10 前記VHコード配列は、図4B(配列番号23)に示される、請求項24に記載の核酸分 子。 【請求項27】 前記VHコード配列は、図4C(配列番号24)に示される、請求項24に記載の核酸分 子。 【請求項28】 前記VHコード配列は、図4D(配列番号25)に示される、請求項24に記載の核酸分 子。 【請求項29】 前記VHコード配列は、図4E(配列番号26)に示される、請求項24に記載の核酸分 20 子。 【請求項30】 前記VHコード配列は、図4F(配列番号27)に示される、請求項24に記載の核酸分 子。 【請求項31】 前記VHコード配列は、図4G(配列番号28)に示される、請求項24に記載の核酸分 子。 【請求項32】 前記VLコード配列は、図3A(配列番号15)に示される、請求項25に記載の核酸分 子。 30 【請求項33】 前記VLコード配列は、図3B(配列番号16)に示される、請求項25に記載の核酸分 子。 【請求項34】 前記VLコード配列は、図3C(配列番号17)に示される、請求項25に記載の核酸分 子。 【請求項35】 前記VLコード配列は、図3D(配列番号18)に示される、請求項25に記載の核酸分 子。 【請求項36】 40 前記VLコード配列は、図3E(配列番号19)に示される、請求項25に記載の核酸分 子。 【請求項37】 前記VLコード配列は、図3F(配列番号20)に示される、請求項25に記載の核酸分 子。 【請求項38】 前記VLコード配列は、図3G(配列番号21)に示される、請求項25に記載の核酸分 子。 【請求項39】 請求項23∼38のいずれかに記載の核酸分子であって、さらに、 50 (5) JP 4270473 B2 2009.6.3 第一のリーダー配列ペプチドをコードする第三のヌクレオチド配列であって、該第三のヌ クレオチド配列は、前記第一のヌクレオチド配列に作動可能に連結され、そして該コード された重鎖可変領域および該第一のリーダー配列ペプチドが発現されるとき、該コードさ れた重鎖可変領域の分泌を生じる能力を有する、第三のヌクレオチド配列;そして 第二のリーダー配列ペプチドをコードする第四のヌクレオチド配列であって、該第四のヌ クレオチド配列は、前記第二のヌクレオチド配列に作動可能に連結され、そして該コード された重鎖可変領域および該第二のリーダー配列ペプチドが発現されるとき、該コードさ れた重鎖可変領域の分泌を生じる能力を有する、第四のヌクレオチド配列 【請求項40】 前記HCV E2抗原に対する免疫学的結合親和性が少なくとも約1.0×107である 10 、請求項23∼39のいずれかに記載の核酸分子。 【請求項41】 前記第一のヌクレオチド配列および前記第二のヌクレオチド配列の転写を指向する制御配 列に作動可能に連結された請求項24∼40のいずれかに記載の核酸分子を含み、ここで 、該第一のヌクレオチド配列および該第二のヌクレオチド配列は、宿主細胞において転写 および翻訳されることができる、発現ベクター。 【請求項42】 前記制御配列は、原核生物宿主細胞において前記第一のヌクレオチド配列および前記第二 のヌクレオチド配列の転写を指向する能力を有する、請求項41に記載の発現ベクター。 【請求項43】 20 前記制御配列は、哺乳動物宿主細胞において前記第一のヌクレオチド配列および前記第二 のヌクレオチド配列の転写を指向する能力を有する、請求項41に記載の発現ベクター。 【請求項44】 請求項42に記載の発現ベクターを用いて形質転換された原核生物宿主細胞。 【請求項45】 請求項43に記載の発現ベクターを用いて形質転換された哺乳動物宿主細胞。 【請求項46】 組換え分子を生産する方法であって、該方法は: (a)請求項44または45に記載の形質転換された宿主細胞の集団を提供する工程;お よび 30 (b)前記発現ベクターから前記組換えFab分子を発現する工程、 を包含する、方法。 【請求項47】 請求項1∼17のいずれかに記載のモノクローナル抗体の、哺乳動物被験体のHCV感染 の処置または予防のためのワクチンの製造における使用。 【請求項48】 前記医薬が、α−インターフェロンまたはリビビリンとともに、HCV感染の処置のため のものである、請求項47に記載の使用。 【発明の詳細な説明】 技術分野 40 本発明は、C型肝炎ウイルス(HCV)に特異的なイムノグロブリン分子に由来する組成物 に関する。より詳細には、本発明は、HCV E2抗原に特異的に結合し得る組換えヒトモノク ローナル抗体に関する。 発明の背景 C型肝炎ウイルス(HCV)感染は、世界中で発生し、そして輸血に関連する肝炎の主要な 原因である。およそ150,000のHCV感染の新たな症例が、毎年合衆国において存在する。世 界中の血液ドナーにおける抗HCV抗体の血清罹患率は、0.02と1.23%との間で変化するこ とが示されており、なかには罹患率が約19%の高さの国もある。輸血誘導型肝炎の主要な 原因であることに加えて、HCVはまた、血液または血液製剤に曝露される個体における肝 炎の共通の原因である。従って、血液または血液製剤のレシピエント、静脈内薬剤使用者 50 (6) JP 4270473 B2 2009.6.3 、腎臓透析患者、および注射針で刺された犠牲者は、HCV感染の高リスク群を代表する。A lterら(1993)Infect Agents Dis 2:155-166。さらに、泌尿生殖器管を介するHCVの異性 間伝染、および母子感染は、かなり記録されている。Ohtoら(1994)N Engl J Med 330:7 44-750。HCV感染に関連する他の危険因子は、HCV感染個体との家族内または家庭内接触、 および血液および血液透析への職業的曝露を伴う医療事業を含む。Alterら(1990)JAMA 264:2231-2235。慢性肝炎は、感染の約62%において発症する。Alterら(1992)N Engl J Med 327:1899-1905。 HCVに関連する重篤な肝臓疾患のほとんどが、慢性的な、持続性の感染を引き起こす因子 の高い性向から生じる。肝硬変は、およそ20%の慢性の症例において起こり、そのうちの 20∼25%が肝不全を生じる。HCV感染に関連する別の重篤な続発症は、原発性肝細胞ガン 10 である。 HCVのウイルスゲノム配列は公知であり、その配列を得る方法も同様に公知である。例え ば、国際公開第WO 89/04669号;同第WO 90/11089号;および同第WO 90/14436号を参照の こと。HCVは、9.5kbの正方向の一本鎖RNAゲノムを有し、そしてウイルスのFlaviridae科 の一員である。現在、系統発生的な分析に基づいて同定されたHCVの異なるがしかし関連 する6つの遺伝子型が存在する(Simmondsら、J.Gen.Virol.(1993)74:2391-2399)。ウ イルスは、3000アミノ酸残基より多くを有する単一のポリペプチドをコードする(Chooら (1989)Science 244:359-362;Chooら(1991)Proc.Natl.Acad.Sci.USA 88:2451-2455;Ha nら(1991)Proc.Natl.Acad.Sci.USA 88:1711-1715)。ポリペプチドは、翻訳中および翻 訳後に構造タンパク質および非構造(NS)タンパク質の両方へとプロセッシングされる。 20 特に、N末端ヌクレオキャプシドタンパク質(「コア」と命名する)からなる3つの推定 HCV構造タンパク質、ならびに2つのエンベロープ糖タンパク質「E1」(Eとしても知ら れる)および「E2」(E2/NS1としても知られる)が存在する。(HCVタンパク質(E1およ びE2を含む)の議論については、Houghtonら(1991)Hepatology 14:381-388を参照のこ と)。E1は、32∼35kDa種として検出され、そして約18KDaの単一のエンドH感受性バンド に変換される。対照的に、E2は、複数の種の生成に一致して免疫沈降法の際に複合体パタ ーンを示す(Grakouiら(1993)J.Virol.67:1385-1395;Tomeiら(1993)J.Virol.67:4017 -4026)。HCVエンベロープ糖タンパク質E1およびE2は、共免疫沈降可能である安定な複合 体を形成する(Grakouiら(1993)J.Virol.67:1385-1395;Lanfordら(1993)Virology 19 7:225-235;Ralstonら(1993)J.Virol.67:6753-6761)。 30 慢性C型肝炎感染のための現在利用可能な唯一の処置は、αインターフェロン(α-IFN) 治療からなる。しかし、インターフェロン治療に対する長期の応答は、処置された個体の ほんの10%∼30%において起こり、異なるHCV株は、インターフェロン治療に対するその 応答性において非常に様々であり、I型ウイルスが最も屈折性であるという証拠が存在す る。さらに、インフルエンザ様の副作用が、インターフェロン治療に対して一般に起こり (処置される個体のおよそ60%∼80%において起こる)、さらに他のより一般的でない副 作用(例えば、吐き気、鬱、疲労、および血小板減少症)も起こる。インターフェロン治 療はまた、免疫無防備状態の個体には望ましくない。従って、慢性HCV感染の処置におけ る、より効果的な治療アプローチに対する必要性が存在する。この点に関して、いくつか の効果が、リビビリン、あるいはウルソジオールとα-IFNとの組合せ治療を用いて観察さ 40 れている。 特に、HCV E1およびE2タンパク質は、非常に興味深い。なぜなら、これらの分子に基づく 組換えワクチンは、霊長類研究におけるHCVでの実験的抗原投与に対して防御的であるこ とが示されているからである。(Chooら(1994)Proc.Natl.Acad.Sci.USA 91:1294-1298 )。ドナー試料から得られる抗HCV抗体分子の過免疫グロブリン組成物は、感染された個 体における、および高リスク群におけるHCV感染の防止における、HCVの処置について記載 されている。1991年9月25日に出願された欧州特許出願第447,984号。これらの組成物は 、ドナー血液製剤から作製されるので、例えば、ヒト免疫不全ウイルス(HIV)およびHCV のような感染性の可能な存在に起因して、内在的なリスクがそれらの使用に付随している 。従って、過免疫グロブリン調製物は、注意深くスクリーニングされなければならず、そ 50 (7) JP 4270473 B2 2009.6.3 してすべての感染性因子がヒト被験体に投与される前に不活化されていなければならない 。 正常な個体におけるHCVに対する免疫応答には、体液性および細胞媒介性成分の両方が含 まれることが知られている。Kozielら(1993)J Virol 67:7522-7532,Alterら(1989)N Engl J Med 321:1494-1500。さらに、いくつかの報告は、HCVに対して誘発された抗体が ウイルスの感染性を中和し得ることを示している。Shimizuら(1994)J Virol 68:1494-1 500,Farciら(1994)Proc Natl Acad Sci USA 91:7792-7796。このような結果は、有効な 抗体に基づく治療が開発され得るという希望を提供する。この点に関して、高度に反応性 の中和化HCV抗体調製物を、感染の危険にあるか、または最近因子に曝露された個体に投 与することにより、迅速な受動免疫がその個体に提供される。このような受動免疫は、正 10 常および免疫無防備状態の両方の被験体において、同様に首尾良いことが予想される。好 ましくは、中和化抗体は、異なるHCV株に対して広く交叉反応性であり、そしてインビボ で抗体の使用の効果を制御するためにモノクローナルである。 多くの実用上そして経済的な理由により、マウスモノクローナル抗体が、一般に研究およ び医学において使用されてきた。マウス抗体は、広範な種々の分子(例えば、HCV抗原) に対して惹起され得、そしてミエローマ細胞と融合されて、HCV抗原に対するモノクロー ナル抗体を産生するために培養物中で増殖され得るハイブリドーマを生じ得る。Kohlerら (1975)Nature 256:495-497。このようなモノクローナル抗体は、有意な治療的価値の抗 原結合特異性を有し得るが、ヒト疾患の処置におけるこのようなマウス抗体の使用は、こ れらの分子がヒト免疫系に対して免疫原性であるために制限されてきた。従って、マウス 20 モノクローナル抗体は、免疫診断において最も一般的に用いれらてきた。この点に関して 、推定HCV E2エンベロープポリペプチドに対するマウスモノクローナル抗体は、生物学的 試料中のHCVの検出における使用について記載されている。Mehtaらに対する米国特許第5, 308,750号。 従って、当該分野で、HCV E2抗原に対するヒトモノクローナル抗体を提供する必要性が残 存する。ここで、モノクローナルは、異種のHCV単離体と広く交叉反応性である。 発明の要旨 本発明は、HCV E2ポリペプチド抗原に対する免疫学的結合親和性を示し、そして異なるHC V株に対して交叉反応性であるヒトモノクローナル抗体分子の発見に基づく。モノクロー ナル抗体分子は、免疫されていないHCV感染した供給源から構築されたコンビナトリアル 30 ライブラリーから得られた。本発明の分子は、一般に、HCV E2抗原に対して免疫学的結合 親和性を示すヒト抗体Fab分子を含む。 従って、1つの実施態様において、本発明は、HCV E2抗原に対して免疫学的結合親和性を 示す組換えヒトモノクローナル抗体に関する。ここで、モノクローナル抗体は、コンビナ トリアル抗体ライブラリーから得られるヒト抗体Fab分子の結合部分に相同なアミノ酸配 列を含む。組換えモノクローナル抗体分子は、実質的に全イムノグロブリン分子の形態で あり得るか、または可溶性のFab分子、Fvフラグメント、もしくはsFv分子の形態であり得 る。ここで、各分子は、少なくともヒト抗体Fab分子の結合部分に相同なアミノ酸配列を 含む。 別の実施態様において、本発明は、HCV E2抗原に対して免疫学的結合親和性を示す、ヒト 40 Fab分子の重鎖または軽鎖可変領域(VHまたはVL)の結合部分に相同なポリペプチドにつ いての、ポリヌクレオチドコード配列を含む単離された核酸分子に関する。関連する実施 態様において、本発明は、第一のポリペプチドについてのポリヌクレオチドコード配列、 および第二のポリペプチドについてのポリヌクレオチドコード配列を含む単離された核酸 分子に関する。ここで、第一のポリペプチドは、HCV E2抗原に対して免疫学的結合親和性 を示すヒトFab分子の重鎖可変領域(VH)の結合部分に相同であり、第二のポリペプチド は、HCV E2抗原に対して免疫学的結合親和性を示すヒトFab分子の軽鎖可変領域(VL)の 結合部分に相同である。 他の実施態様において、本発明は、ベクターが宿主細胞内に存在するか、またはポリヌク レオチドコード配列の転写および翻訳に適切な条件下で存在する場合、ポリヌクレオチド 50 (8) JP 4270473 B2 2009.6.3 コード配列の転写を指向する制御配列に作動可能に連結した上記の核酸分子を含む発現ベ クターに関する。本発明のなおさらなる実施態様は、本発明のベクターで形質転換された 宿主細胞、および形質転換された宿主細胞を用いて組換えポリペプチドを産生するための 方法に関する。 別の実施態様において、本発明は、本発明の組換えモノクローナル抗体分子を含むワクチ ン組成物に関する。なおさらなる実施態様は、ワクチン組成物を使用する方法に関する。 ここで、ワクチンは、哺乳動物被験体においてHCVに対する抗体力価を提供するために、 および/またはワクチン化された被験体においてHCV感染に対する受動免疫を提供するため に用いられる。関連する実施態様において、ワクチン組成物は、公知の抗HCV治療法と組 み合わせて用いられる。 10 なおさらなる実施態様において、本発明の組換えモノクローナル抗体分子は、検出可能な 部分で標識された結合複合体を提供するために使用される。標識された結合複合体は、本 発明の関連する実施態様(例えば、HCVを含むと疑われる試料中にHCV粒子の存在を検出す るための特異的結合アッセイ法、およびHCV感染被験体の抗HCV処置の進行をモニターする ための特異的結合アッセイ)において用いられる。 本発明のこれらおよび他の実施態様は、本明細書中の開示を鑑みて当業者に容易に想到す る。 【図面の簡単な説明】 図1A∼1Gは、それぞれ、Fab分子クローン1:5(配列番号1)、1:7(配列番号2)、1:11 (配列番号3)、L3(配列番号4)、L1(配列番号5)、A8(配列番号6)、およびA12 20 (配列番号7)の推定されたγ1重鎖アミノ酸配列を示す。各重鎖のCDR領域は、図中の 「CDR1」、「CDR2」および「CDR3」として同定される。 図2A∼2Gは、それぞれ、Fab分子クローン1:5(配列番号8)、1:7(配列番号9)、1:11 (配列番号10)、L3(配列番号11)、L1(配列番号12)、A8(配列番号13)、およびA12 (配列番号14)の推定されたκ軽鎖アミノ酸配列を示す。各軽鎖のCDR領域(CDR1、CDR2 およびCDR3)もまた、上記図1A∼1Gに対して注記されたように同定される。 図3A∼3Gは、それぞれ、Fab分子クローン1:5(配列番号15)、1:7(配列番号16)、1:11 (配列番号17)、L3(配列番号18)、L1(配列番号19)、A8(配列番号20)、およびA12 (配列番号21)のκ軽鎖の核酸配列を示す。 図4A∼4Gは、それぞれ、Fab分子クローン1:5(配列番号22)、1:7(配列番号23)、1:11 30 (配列番号24)、L3(配列番号25)、L1(配列番号26)、A8(配列番号27)、およびA12 (配列番号28)のγ1重鎖の核酸配列を示す。 図5は、実施例12に記載のように行われた中和アッセイの結果を示す。本アッセイでは、 Fab分子クローン1:5、1:7、1:11およびL3より発現されたFab分子の、HCV E2ポリペプチド の標的細胞への結合をブロックする能力が評価された。 発明の詳細な説明 本発明の実施は、他で示されていない場合、当該分野における技術の範囲内のウイルス学 、微生物学、分子生物学、および組換えDNA技術の従来方法を使用する。そのような技術 は、文献に十分に説明される。例えば、Sambrookら,Molecular Cloning:A Laboratory M anual(第2版,1989);DNA Cloning:A Practical Approach,第I巻および第II巻(D.Glov 40 er編);Oligonucleotide Synthesis(N.Gait編,1984);Nucleic Acid Hybridization(B .HamesおよびS.Higgins編,1985);Transcription and Translation(B.HamesおよびS.Hi ggins編,1984);Animal Cell Culture(R.Freshney編,1986);Perbal,A Practical Gui de to Molecular Cloning(1984);Fundamental Virology,第2版,第I巻および第II巻( B.N.FieldsおよびD.M.Knipe編)を参照のこと。 本明細書中および添付された請求の範囲中に使用されるように、単数形の「a」、「an」 および「the」は、内容が明確に他を指示しない場合は、複数の対象物を含む。 A.定義 本発明の記載において、以下の用語が使用され、そして下記のように定義されることを意 図される。 50 (9) JP 4270473 B2 2009.6.3 本明細書中で使用されたように、用語「C型肝炎ウイルス」または「HCV」は、包括的な 様式でウイルスを記述し、そして従ってこの用語は、何ら特定のHCVのウイルス配列にも 単離株にも限定されない。この点について、系統学的な分析に基づいて同定された11の異 なるサブタイプを有する、6つの異なるHCVの遺伝子型が存在する(Houghton,M.(1996) 「Hepatitis C Viruses,」Fields Virology,第3版,Fieldsら編,Lippincott-Raven Pub lishers,Philadelphia PA.;Simmondsら,J.Gen.Virol.(1993)74:2391-2399)。さらに 、世界中の異なるHCV単離株由来のゲノムヌクレオチド配列の比較は、74の単離株間での 配列多様性の範囲内で、HCVが非常に不均一であることを立証する。それゆえ、用語「C 型肝炎ウイルス」および「HCV」は、本明細書中で使用されるように、そのような単離株 の全てを包括的に含む。 10 用語「E1ポリペプチドに由来する抗原」、「E1ポリペプチド抗原」および「HCV E1抗原」 は、本明細書中で交換可能に使用され、そしてHCV E1領域由来の分子を含む。用語「ポリ ペプチド」は本明細書中で使用されるように、アミノ酸のポリマーをいい、特定の長さの 産物をいうものではない;それゆえ、ポリペプチド、オリゴペプチド、およびタンパク質 は、ポリペプチドの定義に含まれる。この用語はまた、ポリペプチドの発現後修飾(例え ば、グリコシル化、アセチル化、リン酸化など)をいうものでもなく除外もしない。E1ポ リペプチド抗原は、HCV E1領域より物理的に誘導され得るか、または公知の配列に基づい て、組換え的または合成的に産生され得る。HCV1の成熟E1領域は、ポリタンパク質のほぼ アミノ酸192より開始し、ほぼアミノ酸383まで連続する。 指定されたHCV領域に「由来する」ポリペプチドまたはアミノ酸配列は、配列内にコード 20 されるポリペプチドのアミノ酸配列と同一のアミノ酸配列を有するポリペプチド、または その部分であって、少なくとも3∼5アミノ酸、好ましくは少なくとも4∼7アミノ酸、 より好ましくは少なくとも8∼10アミノ酸、そしてなおより好ましくは少なくとも11∼15 アミノ酸からなる部分、あるいは配列内にコードされるポリペプチドと免疫学的に同一視 し得るポリペプチドをいう。この用語法はまた、指定されたHCV領域から発現されるポリ ペプチドを含む。 用語「E2ポリペプチドに由来する抗原」、「E2ポリペプチド抗原」、および「HCV E2抗原 」は、本明細書中で交換可能に使用され、そしてHCV E2領域由来の分子を含む。そのよう な分子は、その領域より物理的に誘導され得るか、または公知の配列に基づいて、組換え 的または合成的に産生され得る。HCV1の成熟E2領域は、ほぼアミノ酸384∼385より開始す 30 ると考えられる。 本発明の目的のために、HCV E1およびE2ポリペプチドは、HCV1ゲノムによってコードされ るポリタンパク質の、アミノ酸番号(開始メチオニンが位置1と指定される)に関して決 定される。しかし、「E1ポリペプチド」または「E2ポリペプチド」由来の抗原は、正確な HCV1配列を有するポリペプチドに限定されないことに注意すべきである。実際に、HCVゲ ノムは、不断の不安定(flux)状態にあり、そして単離株間でが比較的高度な可変性を示 すいくつかの可変ドメインを含む。これらの用語が、Simmondsら,J Gen Virol(1993)7 4:2391-2399に記載の6つの遺伝子型のHCVのいずれかを有する単離株を含む、任意の種々 のHCV単離株由来のE1およびE2ポリペプチド由来の抗原を含むことは、容易に明らかであ る。このことについて、非相同のHCV単離株中の対応するE1またはE2領域は、その配列を 40 最大の整列(alignment)に導く様式で、2つの単離株由来の配列を整列させることによ って容易に決定し得る。これは、University of Virginia,Department of Biochemistry (Attn:Dr.William R.Pearson)より入手可能なALIGN 1.0のような、任意の多数のコンピ ューターソフトウエアパッケージで遂行され得る。Pearsonら,Proc Natl Acad Sci USA (1988)85:2444-2448を参照のこと。 さらに、用語「E1ポリペプチド抗原」および「E2ポリペプチド抗原」は、内部欠失、付加 および置換(天然で一般的に保存的な)のような、ネイティブな配列に対する改変を含む ポリペプチドを含む。これらの改変は、部位特異的変異誘発を通じてのように計画的であ り得るか、または天然に生じる変異事象を通じてのように偶発的であり得る。 「E1/E2複合体」は、上記E1およびE2ポリペプチドの複合体をいう。このような複合体に 50 (10) JP 4270473 B2 2009.6.3 おけるE1およびE2ポリペプチドの会合の様式は、重要でない。実際に、そのような複合体 は、個別に産生されたE1およびE2ポリペプチドを単に混合することにより、自然に形成し 得る。同様に、同時に発現または分泌された場合、E1およびE2ポリペプチドは、培地中で 自然に複合体を形成し得る。「E1/E2複合体」の形成は、ポリアクリルアミドゲル電気泳 動のような標準的なタンパク質検出技法および免疫沈降法のような免疫学的技法を用いる ことにより容易に検出される。 用語「抗体」は、モノクローナル抗体調製物、ならびにハイブリッド抗体、改変(altere d)抗体、F(ab’)2フラグメント、F(ab)分子、Fvフラグメント、単一ドメイン抗体、 キメラ抗体、および親抗体分子の免疫学的結合特性を示すその機能フラグメントを含む調 製物を含む。 10 本明細書中に使用されるように、用語「モノクローナル抗体」は、同種の抗体集団を有す る抗体組成物をいう。この用語は、モノクローナル抗体が作成される様式によって限定さ れない。この用語は、イムノグロブリン分子全体、ならびにFab分子、F(ab’)2フラグ メント、Fvフラグメント、および親モノクローナル抗体分子の免疫学的結合特性を示す他 の分子を含む。用語「組換えモノクローナル抗体」は、本明細書中では、組換えポリヌク レオチドの発現により産生されるモノクローナル抗体として定義される。 用語「抗原結合部位」または「結合部分」は、抗原結合に関与するイムノグロブリン分子 の一部分をいう。抗原結合部位は、重(「H」)鎖および軽(「L」)鎖のN末端可変( 「V」)領域のアミノ酸残基によって形成される。重鎖および軽鎖のV領域内の3つの非 常に相違した伸長部(stretch)は、「超可変領域」と呼ばれ、「骨格領域」または「FR 20 」として公知のより保存された隣接伸長部の間に介在する。それゆえ、用語「FR」は、イ ムノグロブリンの超可変領域間にこれに近接して天然に見出されるアミノ酸配列をいう。 抗体分子において、軽鎖の3つの超可変領域および重鎖の3つの超可変領域は、抗原結合 表面を形成する3次元空間に互いに相対して配置される。抗原結合表面は、結合抗原の3 次元表面に相補的であり、そして重鎖および軽鎖のそれぞれの3つの超可変領域は、「相 補性決定領域」または「CDR」といわれる。 本明細書中に使用されるように、用語「免疫学的結合」およ「免疫学的結合特性」は、イ ムノグロブリン分子とそれに対してこのイムノグロブリンが特異的である抗原との間に生 じるタイプの非共有結合性相互作用をいう。免疫学的結合相互作用の強度または親和性は 、その相互作用の解離定数(Kd)の面から表現され得るが、そのより小さなKdは、より強 30 い親和性を示す。選択されたポリペプチドの免疫学的結合特性は、当該分野で周知の方法 を用いて定量化され得る。そのような方法の一つは、抗原結合部位/抗原複合体の形成お よび解離の速度の計測を伴い、その速度は、複合体パートナーの濃度、相互作用の親和性 、および両方向の速度に均等に影響する幾何学的パラメーターに依存する。それゆえ、「 オン速度定数」(Kon)および「オフ速度定数」(Koff)は共に、濃度ならびに会合およ び解離の実際の速度を計算することにより決定され得る。Koff/Konの比率は、親和性とは 関係しないすべてのパラメーターの削除を可能にし、そしてそれゆえ解離定数Kdに等しい 。一般的には、Davisら,(1990)Annual Rev Biochem 59:439-473を参照のこと。 抗体分子の免疫学的結合特性を示し得る抗原結合部位を含む、多数の治療的に有用な分子 は、当該分野で公知である。そのような分子の一つは、インタクトな抗原結合部位を含む 40 ヘテロダイマーを含むFab分子である。酵素ペプシンは、両抗原結合部位を含む「F(ab’ )2」フラグメントを含むいくつかのフラグメントを提供するために、IgG分子を切断し得 る。「Fv」フラグメントは、IgM、およびごくまれにIgGまたはIgAイムノグロブリン分子 の選択的タンパク質分解性切断によって産生され得る。しかし、Fvフラグメントは、当該 分野で公知の組換え技術を用いて、より一般的に誘導される。Fvフラグメントは、ネイテ ィブな抗体分子の抗原認識能力および結合能力の大部分を保持する抗原結合部位を含む、 非共有結合性のVH::VLヘテロダイマーを含む。Inbarら,(1972)Proc.Nat.Acad.Sci USA 69:2659-2662;Hochmanら,(1976)Biochem 15:2706-2710;およびEhrlichら,(1980)B iochem 19:4091-4096。 指定されたFab分子またはFab核酸配列に「由来する」ポリペプチド分子またはアミノ酸配 50 (11) JP 4270473 B2 2009.6.3 列は、配列内にコードされるFabポリペプチドのアミノ酸配列と同一のアミノ酸配列を有 するポリペプチド、またはその部分であって、少なくとも3∼5アミノ酸、好ましくは少 なくとも4∼7アミノ酸、より好ましくは少なくとも8∼10アミノ酸、そしてなおより好 ましくは少なくとも11∼15アミノ酸からなる部分をいう。 単一鎖Fv(「sFv」)ポリペプチド分子は、ペプチドをコードするリンカーによって連鎖 されたVHをコードする遺伝子およびVLをコードする遺伝子を含む遺伝子融合物から発現さ れる、共有結合性のVH::VLヘテロダイマーである。Hustonら,(1988)Proc Nat Acad Sc i USA 85(16):5879-5883。自然に凝集されるが化学的に分離される抗体V領域由来の軽 および重ポリペプチド鎖を、抗原結合部位の構造と実質的に同様な3次元構造へ畳み込む sfv分子に変換するための化学構造を識別する多数の方法が記載される。例えば、Huston 10 らによる、米国特許第5,091,513号、および第5,132,405号;およびLadnerらによる、第4, 946,778号を参照のこと。 上記分子の各々は重鎖および軽鎖のCDRセットを含み、これらのセットは、CDRの支持を提 供しそしてCDRの互いに対する空間的関係を規定する重鎖および軽鎖のFRセットの間にそ れぞれ挿入される。本明細書中で使用される用語「CDRセット」とは、重鎖または軽鎖のV 領域の3つの超可変領域をいう。重鎖または軽鎖のN末端から始めて、3つの領域をそれ ぞれ「CDR1」、「CDR2」、および「CDR3」という。それ故、抗原結合部位は、6つのCDR (重鎖および軽鎖のV領域の各々に由来するCDRセットを含む)を含む。 本明細書中に使用される用語「FRセット」とは、重鎖または軽鎖のV領域のCDRセットのCD Rを構成する(frame)4つの隣接アミノ酸配列をいう。いくつかのFR残基は結合抗原と接 20 触し得る;しかし、FrはV領域を抗原結合部位(特に、CDRに直接隣接するFR残基)に畳み 込む(fold)ことを主に担う。 ポリペプチド配列またはヌクレオチド配列をいう場合、「精製された」および「単離され た」とは、示された分子が、同タイプの他の生物学的巨大分子の実質的非存在下において 存在することを意味する。本明細書中で使用される用語「精製された」および「単離され た」とは、好ましくは少なくとも75重量%、より好ましくは少なくとも85重量%、よりな お好ましくは少なくとも95重量%、そして最も好ましくは少なくとも98重量%の同タイプ の生物学的巨大分子が存在すること意味する。「特定のポリペプチドをコードする単離さ れた核酸分子」とは、目的のポリペプチドをコードしない他の核酸分子を実質的に有さな い核酸分子をいう;しかし、分子は、組成物の基礎的特徴に有害な影響を与えない、いく 30 つかのさらなる塩基または部分を含み得る。従って、例えば、抗体の特定の重鎖可変領域 の結合部分をコードする単離された核酸分子は、目的の結合部分についてのヌクレオチド コード配列から本質的に構成される(例えば、FRセットの間に挿入されたCDRセット)。 「相同性」とは、2つのポリヌクレオチドまたはポリペプチド部分間の同一性のパーセン トをいう。2つ以上の配列間の一致は、当該分野で公知の技術により決定され得る。例え ば、相同性は、2つのポリペプチド分子間の配列情報の直接比較により決定され得る。あ るいは、相同性は、相同な領域間で安定な二本鎖を形成する条件下でのポリヌクレオチド のハイブリダイゼーション(例えば、S1消化前に使用されるもの)、続いて一本鎖特異的 ヌクレアーゼ(単数または複数)での消化、続いて消化されたフラグメントのサイズ決定 により決定され得る。2つのDNAまたはポリペプチド配列は、分子の規定された長さにわ 40 たり、少なくとも約60%(好ましくは少なくとも約80%、そして最も好ましくは少なくと も約90%)のヌクレオチドまたはアミノ酸が適合する場合に、「実質的に相同性」である 。 用語「組換えDNA分子」または「組換え核酸分子」とは、その起源または操作に基づいて 、(1)天然で関連するポリヌクレオチドの全部または一部と関連しないか、(2)天然で 連鎖するポリヌクレオチド以外のポリヌクレオチドと連鎖するか、または(3)天然に存 在しない、ゲノム、cDNA、半合成、または合成起源のポリヌクレオチドをいうために本明 細書中で使用される。従って、この用語は、「合成的に由来する」核酸分子を包含する。 本明細書中で使用される用語「核酸分子」とは、リボヌクレオチドまたはデオキシリボヌ クレオチドのいずれかの、任意の長さのヌクレオチドのポリマー形態をいう。この用語は 50 (12) JP 4270473 B2 2009.6.3 、分子の一次構造のみをいい、従って二本鎖および一本鎖DNAおよびRNAを含む。これはま た、例えば、当該分野で公知の標識、メチル化、「キャップ」、アナログとの1つ以上の 天然に存在するヌクレオチドの置換、無荷電結合での修飾(例えば、メチルホスホネート 、ホスホトリエステル、ホスホラミダート(phosphoamidate)、カルバメートなど)およ び荷電結合での修飾(例えば、ホスホロチオエート、ホスホロジチオエートなど)ような ヌクレオチド間修飾、タンパク質のような付属物部分(例えば、ヌクレアーゼ、毒素、抗 体、シグナルペプチド、ポリ-L-リジンなど)を含む修飾、インターカレーター(例えば 、アクリジン、ソラーレンなど)での修飾、キレート剤(例えば、金属、放射活性金属、 ホウ素、酸化金属など)を含む修飾、アルキル剤を含む修飾、修飾された結合(例えば、 αアノマー核酸など)での修飾のような公知のタイプの修飾、ならびにポリヌクレオチド 10 の未修飾の形態を含む。 「コード配列」は、適切な調節配列の制御下に配置された場合に、通常はmRNAを介して、 ポリペプチドに翻訳される核酸分子である。コード配列の境界は、5’末端の翻訳開始コ ドンおよび3’末端の翻訳停止コドンにより決定され得る。コード配列には、CDNAおよび 組換えヌクレオチド配列が挙げられるが、これらには限定されない。 「制御配列」とは、それらが連結されたコード配列の発現をもたらすのに必要である核酸 配列をいう。このような制御配列の性質は、宿主生物に依存して異なる;原核生物におい ては、このような制御配列には、一般的に、プロモーター、リボソーム結合部位、および 転写末端配列が挙げられる;真核生物においては、このような制御配列には、一般的に、 プロモーターおよび転写終結配列が挙げられる。用語「制御配列」は、コード配列の発現 20 のためにその存在が必要である全ての成分を最小限含み、そしてその存在が有利であるさ らなる成分(例えば、リーダー配列および融合パートナー配列)を含み得る。 「作動可能に連結された」とは、そのように記載される成分がそれらが意図される様式で 機能することを可能にする関係にある近位をいう。コード配列に「作動可能に連結された 」制御配列は、コード配列の発現が制御配列と適合する条件下で達成されるような方法で 結合される。 本明細書中で使用される用語「発現カセット」とは、適切な宿主細胞におけるコード配列 のクローン化コピーの転写およびmRNAの翻訳のために必要な全てのヌクレオチド配列を含 む制御配列に作動可能に連結された少なくとも1つのコード配列を含む分子をいう。この ような発現カセットは、種々の宿主(例えば、細菌、ラン藻類、植物細胞、酵母細胞、昆 30 虫細胞、および動物細胞)において真核生物遺伝子を発現するために使用され得る。本発 明の下では、発現カセットには、クローニングベクター、特別に設計されたプラスミド、 ウイルスまたはウイルス粒子が挙げられ得るが、これらには限定されない。カセットはさ らに、宿主細胞における自立増殖のための複製起点、選択マーカー、種々の制限部位、高 コピー数についての可能性、および強力なプロモーターを含み得る。 「ベクター」とは、適切な制御エレメントに結合された場合に複製を可能にし、そして細 胞間で遺伝子配列を移送し得る、任意の遺伝的エレメント(例えば、プラスミド、ファー ジ、トランスポゾン、コスミド、染色体、ウイルスなど)を意味する。従って、この用語 は、クローニングビヒクルおよび発現ビヒクル、ならびにウイルスベクターを含む。 「組換え宿主細胞」、「宿主細胞」、「細胞」、「細胞培養物」、および他のこのような 40 用語は、例えば、組換えベクターまたは他の移送DNAのためのレシピエントとして使用さ れ得る(または、使用されている)微生物、昆虫細胞、および哺乳動物細胞を示し、そし て形質転換されている元の細胞の子孫を含む。単一の親細胞の子孫が、天然の、偶発的な 、または意図的な変異により、形態においてまたはゲノムもしく全DNA相補体においてそ の元の親と必ずしも完全に同一ではないかもしれない。哺乳動物宿主細胞についての例に は、チャイニーズハムスター卵巣(CHO)およびサル腎臓(COS)細胞が挙げられる。 詳細には、本明細書中で使用される「細胞株」とは、インビトロにおいて連続的なまたは 延長された増殖および分裂が可能である細胞の集団をいう。しばしば、細胞株は、単一の 祖先細胞に由来するクローン集団である。自発的なまたは誘導された変化は、このような クローン集団の保存または移送の間に核型において生じ得ることは、当該分野においてさ 50 (13) JP 4270473 B2 2009.6.3 らに公知である。それ故、言及される細胞株由来の細胞は、祖先の細胞または培養物と正 確に同一でないかもしれないし、そして言及される細胞株はこのような変異株を含む。用 語「細胞株」はまた、不死化細胞を含む。 本明細書中で使用される「形質転換」とは、挿入のために使用される方法(例えば、直接 取り込み、形質導入、f接合、またはエレクトロポレーション)とは無関係に、宿主細胞 への異種ポリヌクレオチドの挿入をいう。異種ポリヌクレオチドは、非組込み型(non-in tegrated)のベクター(例えば、プラスミド)として維持され得、あるいは宿主ゲノムへ 組み込まれ得る。 B.一般的方法 本発明は、HCV E2エンベロープ糖タンパク質に特異的である、新規な交叉遺伝子型反応性 10 ヒトモノクローナル抗体分子の生成に基づく。モノクローナル抗体は、非免疫化供給源か ら構築されたコンビナトリアル・抗体ライブラリーを使用して得られ、そして哺乳動物被 験体におけるHCV感染の予防、治療、および診断において有用である。より詳細には、モ ノクローナル抗体は、糸状DNAバクテリオファージの表面上でFab分子を発現するコンビナ トリアル・ライブラリーから抗原選択技術を使用して得られる。 コンビナトリアル・ライブラリーの調製 本発明の目的のためのコンビナトリアル・ライブラリーは、公知の技術(例えば、Chanoc kら、(1933)Infect Agents Dis 2:118-131およびBarbas,IIIら、(1995)Methods:Comp .Meth Enzymol 8:94-103により記載される技術)を使用して構築され得る。抗体産生細胞 は、非免疫化HCV感染供与者から、例えば、血漿、血清、脊髄液、リンパ液、呼吸器の外 20 部切片(external section)、消化管および尿生殖器管、涙液、唾液、乳液、白血球、お よびミエローマから得られ得る。好ましくは、抗体産生細胞供給源は、非免疫化被験体の 骨髄または末梢血試料から得られたリンパ球である。 リンパ球は、試料から得られ、そして全RNAが、公知の方法を使用して単離および抽出さ れ得る。例えば、Chomczynskiら、(1987)Anal Biochem 162:156-159を参照のこと。RNA は、オリゴ-dTプライミングを使用して第一鎖CDNAに逆転写され得る。イムノグロブリン 重鎖(Fd)および軽鎖のフラグメントをコードするDNAは、ポリメラーゼ連鎖反応(PCR) を使用して増幅されて、Fab抗原結合分子を産生するために必要な全ての遺伝物質を提供 し得る。Saikiら、(1986)Nature 324:163、Scharfら、(1986)Science 233:1076-1078 、ならびに米国特許第4,683,195号および同第4,683,202号。PCR増幅の実施においては、 30 多数の公知のプライマーがγ1重鎖およびκ軽鎖配列について選択するために好ましく使 用され得る。Perssonら、(1991)Proc Natl Acad Sci USA 88:2432-2436、Kangら、(19 91)Methods:Comp.Meth Enzymol 2:111-118。PCR産物は、重鎖および軽鎖DNA調製物に別 々にプールされ、次いで、例えば、ゲル電気泳動を使用して精製される。次いで、精製さ れた重鎖および軽鎖DNA分子は、適切な制限酵素で消化され、そして消化産物は精製され 、適切なファージミドベクター系に連結される。Yangら、(1995)J Mol Biol 254:392-4 03、Barbas,IIIら、(1995)Methods:Comp.Meth Enzymol 8:94-103、Barbas,IIIら、(19 91)Proc Natl Acad Sci USA 88:7978-7982。多数の適切なファージミドベクター系は、 当該分野において公知である;しかし、本明細書中の使用に特に好ましいベクターは、以 前に記載されているpComb3hベクターである。Barbas,IIIら、(1995)、前出。Pcomb3hフ 40 ァージミドベクターが使用される場合、重鎖DNAが、目的のファージミド中へ、ファージ ミドコートタンパク質III(cpIII)のC末端アンカードメインについての配列に隣接しか つその上流へクローン化される。cpiiiタンパク質は、主要膜タンパク質であり、それ故 、Fabアセンブリのための膜アンカーとして役立つ。 ベクターは、一般的に、当該分野で公知の選択マーカーを含む。例えば、Pcomb3hファー ジミドベクターは、細菌アンピシリン耐性遺伝子(βラクタマーゼ)を含む。ベクターは また、適切な第一および第二リーダー配列を含む。それらの配列は、重鎖および軽鎖をコ ードする配列のための挿入部位の上流にそれぞれに配置され、それによって、適切な宿主 細胞において産生される場合に、重鎖および軽鎖コード領域由来の発現産物は周辺質に標 的化される。Pcomb3hにおいて、これらのリーダー配列は、pelB配列、omp A配列、または 50 (14) JP 4270473 B2 2009.6.3 それらの組み合わせである。 次いで、ヒトイムノグロブリンDNAを含むファージミドベクター系は、(例えば、電気泳 動を使用して)適切な細菌宿主細胞に導入され、ここで、ファージミドは、重鎖cpiii融 合ポリペプチドおよび軽鎖ポリペプチド(これらの各々は、それらの関連するリーダー配 列により宿主細胞の周辺質に標的化される)を発現する。ファージミドベクターを含むト ランスフェクトされた細菌宿主細胞は、ファージミドベクターの選択マーカーに対応する 選択薬剤(例えば、アンピシリン)を含む適切な培地における増殖により選択される。 ファージミドDNAのレスキューは公知の技術を使用して実施される。特に、トランスフェ クトされた宿主細胞は、ファージミドDNAの組換えウイルス粒子中へのパッケージングの ためにトランスに必要な多数の発現産物をコードするヘルパーファージで感染される。フ 10 ァージミドDNAの一本鎖コピーはこのようにウイルス粒子中にパッケージングされ、これ は宿主細胞を離れるに際してファージcpVIII分子を取り込み、そして限定された数のファ ージcpiii分子によりキャップされ−−そのcpviiiおよびcpiii分子のいくつかはFab分子 に連結される。Fab分子を提示する組換えファージ粒子(「ファージ-Fab」と称する)は 、パッケージングされるゲノム内に対応する重鎖遺伝子および軽鎖遺伝子を含む。 上記の技術がファージミドの最初のライブラリーを使用して実施されると、レスキュープ ロセスはFab分子を提示する組換えファージのライブラリー(ファージディスプレイライ ブラリー)を生成する。レスキュープロセスはさらに最初のライブラリーの増幅を生じ、 その結果、複数のコピーの各組換えファージクローンが(イムノグロブリン重鎖および軽 鎖結合部分の各セットとともに)生成される。次いで、ファージディスプレイライブラリ 20 ーは、HCV E2抗原に対して「パニング(panned)」されて、その抗原に選択的に結合し得 るFab分子について選択される。より特定すると、パニング手順はファージディスプレイ ライブラリーを含有する懸濁液を、公知の方法に従ってプラスチック反応容器に固定化さ れたHCV E2抗原に適用することにより実施され得る。Burtonら、(1991)Proc Natl Acad Sci USA 88:10134-10137。適切な結合条件下でのインキュベーション後、非特異的に結 合したファージ粒子は、反復する洗浄により除去される。次いで、得られるHCV E2抗原特 異的ファージ-Fabは、低Phを使用して、または過剰の可溶性E2抗原の存在下で、不溶性抗 原から溶出される。パニング手順は数回反復され、ここで、細菌宿主細胞は、パニングの 各回の後に溶出されたファージにより感染されて、パニングの各々の次の回のためにファ ージ-Fabクローンが増殖される。Samuelssonら、(1995)Virology 207:495-502。 30 本発明において、パニング手順は、HCV E2抗原に特異的な、高度に強力な、交叉遺伝子型 反応性Fab分子について選択するために特定して開発された。特に、そこから抗体産生細 胞が得られた非免疫化、HCV感染ヒト被験体の血清HCVの遺伝子型は、公知の方法を使用し て決定された。Widellら、(1994)J Med Virol 44:272-279。株交叉反応性についての選 択は、実験的設計により提供された。ここで、パニング手順は、感染されたヒト供与者由 来のHCVの遺伝子型とは異なるHCV遺伝子型に由来するHCV E2抗原を使用して実施された。 さらに、パニング手順において使用されたE2抗原は、インビボにおいてその抗原について 予想されるコンフォメーションと実質的に同一のコンフォメーションのHCV E2抗原を提供 するように選択された。 2つの異なる組換えHCVエンベロープタンパク質調製物が、上記のパニング手順における 40 選択抗原を提供するために使用された(「コンフォメーショナル」CHO E2分子、およびCH O E1/E2複合体)。コンフォメーショナルE2分子は、Spaeteら、(1992)Virology 188:81 9-830に以前に記載のように、構築され、発現され、そして組換えCHO細胞から分泌され、 次いで公知の方法(Rosaら、(1996)Proc Natl Acad Sci USA 93:1759-1763)を使用し て精製された。組換え複合体化E1/E2調製物は、Spaeteら、(1992)(前出)に記載のよ うに、構築され、そして組換えCHO細胞から発現され、次いで公知の方法(Chooら、(199 4)Proc Natl Acad Sci USA 91:1294-1298)を使用して精製された。一旦精製されると、 選択抗原は、上記のように、プラスチック反応容器に固定化された。 選択抗原に対する優れた結合親和性を示す個々のクローンは、選択され、そして感染され た宿主細胞を、選択培地中で、適切な容量の細胞に達するまで増殖させることにより発現 50 (15) JP 4270473 B2 2009.6.3 された。細菌宿主細胞はペレット化され、次いで培地中に再懸濁された。適切なインキュ ベーション後、細胞はスピンダウンされ、そして周辺質内容物は凍結融解技術により遊離 された。細菌破片が遠心分離により除去された後、Fab含有上清は適切な容器に移され、 そして将来の使用のために貯蔵された。 一旦選択されたFabが発現されると、選択されたFab分子の結合特性が決定され得る。特に 、HCV E2抗原に対するFab分子の親和性は、阻害ELISA技術を使用して、本明細書において 決定された。例えば、Perssonら、(1991)Proc Natl Acad Sci USA 88:2432-2436、Rath ら、(1988)J Immun Methods 106:245-249を参照のこと。高い効力(例えば、阻害ELISA により決定したところ、少なくとも約1×107M-1、および好ましくは少なくとも約1.7×1 07M-1の親和性)のFab分子を発現したクローンが、配列決定のために同定された。 10 パニング選択プロセスにおいて高い効力の結合を示したクローン由来のファージ(プラス ミド)DNAが単離され、そして一本鎖DNAがイムノグロブリンクローニング領域の上流およ び下流にハイブリダイズするプライマー(その1つは、例えば、5’末端においてビオチ ン化されている)を使用するPCRにより得られた。PCR後、一本鎖DNAは、DNAをアルカリ性 条件下で変性し、そしてビオチン化DNA鎖を固体支持体に吸着させることにより得られた 。ジデオキシ配列決定反応は、公知の方法に従って(Sangerら、(1977)Proc Natl Acad Sci USA 74:5463-5467)、可変領域と定常領域との間の連結部の3’にハイブリダイズす る標識プライマーを使用して実施された。Kabatら、Sequences of Proteins of Immunolo gical Interest,第4版,(U.S.Dept.of Health and Human Services,U.S.Government Pr inting Office,1987)。反応手順は、自動化配列決定装置(例えば、Pharmacia Biotech 20 から入手可能であるA.L.F.)で行われた。このように得られた核酸配列情報は、選択され たモノクローナルFab分子の重鎖および軽鎖部分のコード配列を提供するために分析され た。同一のクローンの複数のコピーが、配列データの比較により同定された。さらに、推 定アミノ酸配列は、公知の方法を使用して得られた。 上記の核酸配列情報を使用して、Fab分子のコード配列はまた、公知の方法を使用して合 成的に産生され得る。ヌクレオチド配列は、所望の特定のアミノ酸配列に対する適切なコ ドンを用いて設計され得る。一般に、その中で配列が発現される意図される宿主について 好ましいコドンを選択する。全配列は、一般に、標準的方法により調製され、そして全コ ード配列に組立てられる重複するオリゴヌクレオチドから組立てられる。例えば、Edge( 1981)Nature 292:756;Nambairら、(1984)Science 223:1299;Jayら、(1984)J.Biol.C 30 hem.259:6311を参照のこと。 発現系 一旦Fab分子の重鎖および軽鎖部分のコード配列が単離または合成されると、それらは発 現のための任意の適切なベクターまたはレプリコン中にクローン化され得、例えば、細菌 、哺乳動物、酵母、およびウイルス発現系が使用され得る。多数のクローニングベクター が当業者に公知であり、そして以下に記載される。適切なクローニングベクターの選択は 好みの問題である。 i.細菌細胞における発現 細菌発現系が、Fab分子を産生するために使用され得る。細菌系における使用のための制 御エレメントとしては、プロモーター(必要に応じてオペレーター配列を含む)、および 40 リボソーム結合部位が挙げられる。有用なプロモーターとしては、糖代謝酵素(例えば、 ガラクトース、ラクトース(lac)、およびマルトース)由来の配列が挙げられる。さら なる例としては、生合成酵素(例えば、トリプトファン(trp))由来のプロモーター配 列、β-ラクタマーゼ(bla)プロモーター系、バクテリオファージλPL、およびT7が挙げ られる。さらに、tacプロモーターのような合成プロモーターが使用され得る。β-ラクタ マーゼおよびラクトースプロモーター系は、Changら、Nature(1978)275:615、およびGo eddelら、Nature(1979)281:544に記載されており;アルカリホスファターゼ、トリプト ファン(trp)プロモーター系は、Goeddelら、Nucleic Acids Res.(1980)8:4057および EP 36,776に記載されており、そしてtacプロモーターのようなハイブリッドプロモーター は、米国特許第4,551,433号およびdeBoerら、Proc.Natl.Acad.Sci.USA(1983)80:21-25 50 (16) JP 4270473 B2 2009.6.3 に記載されている。しかし、真核生物タンパク質の発現のために有用な他の公知の細菌プ ロモーターもまた適切である。当業者は、例えば、Siebenlistら、Cell(1980)20:269に 記載のように、任意の必要とされる制限部位を供給するためのリンカーまたはアダプター を使用して、このようなプロモーターをFab分子に作動可能に連結し得る。細菌系におけ る使用のためのプロモーターはまた、一般に、Fab分子をコードするDNAに作動可能に連結 されたシャイン-ダルガルノ(SD)配列を含む。天然のポリペプチドシグナル配列を認識 およびプロセシングしない原核生物宿主細胞については、シグナル配列が、例えば、アル カリホスファターゼ、ペニシリナーゼ、Ippまたは熱安定エンテロトキシンIIリーダーの 群から選択される原核生物シグナル配列により置換され得る。プラスミドpBR322由来の複 製起点は、大部分のグラム陰性細菌に適切である。 10 上記の系は、Escherichia coliに特に適合性である。しかし、とりわけ、Bacillus spp. 、Streptococcus spp.、Streptomyces spp.、Pseudomonas種(例えば、P.aeruginosa)、 Salmonella typhimurium、またはSerratia marcescansのようなグラム陰性またはグラム 陽性生物を含む細菌宿主における使用のための多数の他の系が存在する。外因性DNAをこ れらの宿主に導入するための方法としては、代表的には、CaCl2またはその他の薬剤(例 えば、二価カチオンおよびDMSO)の使用が挙げられる。DNAはまた、上記で引用されるSam brookら、(1989)に一般に記載されるように、エレクトロポレーション、核注入、また はプロトプラスト融合により細菌細胞中に導入され得る。これらの例は、例示であり限定 ではない。好ましくは、宿主細胞は、最小量のタンパク質分解酵素を分泌するべきである 。あるいは、クローニングのインビトロ方法(例えば、PCRまたはその他の核酸ポリメラ 20 ーゼ反応)が適切である。 本発明のFab分子を産生するために使用される原核生物細胞は、上記で引用されるSambroo kらに一般に記載されるように、適切な培地中で培養される。 ii.酵母細胞における発現 酵母発現系もまた、本発明のFab分子を産生するために使用され得る。発現および形質転 換ベクター(染色体外レプリコンまたは組み込みベクターのいずれか)は、多くの酵母中 への形質転換のために開発されている。例えば、発現ベクターが、とりわけ、以下の酵母 のために開発されている:Saccharomyces cerevisiae(Hinnenら、Proc.Natl.Acad.Sci.U SA(1978)75:1929;Itoら、J.Bacteriol.(1983)153:163に記載);Candida albicans(K urtzら、Mol.Cell.Biol.(1986)6:142に記載);Candida maltosa(Kunzeら、J.Basic Mi 30 crobiol.(1985)25:141に記載);Hansenula polymorpha(Gleesonら、J.Gen.Microbiol. (1986)132:3459、およびRoggenkampら、Mol.Gen.Genet.(1986)202:302に記載);Kluy veromyces fragilis(Dasら、J.Bacteriol.(1984)158:1165に記載);Kluyveromyces la ctis(De Louvencourtら、J.Bacteriol.(1983)154:737、およびVan den Bergら、Bio/T echnology(1990)8:135に記載);Pichia guillerimondii(Kunzeら、J.Basic Microbiol .(1985)25:141に記載);Pichia pastoris(Creggら、Mol.Cell.Biol.(1985)5:3376、 ならびに米国特許第4,837,148号、および同第4,929,555号に記載);Schizosaccharomyces pombe(BeachおよびNurse,Nature(1981)300:706に記載);ならびにYarrowia lipolyti ca(Davidowら、Curr.Genet.(1985)10:380、およびGaillardinら、Curr.Genet.(1985 )10:49に記載)、Aspergillus宿主(例えば、A.nidulans(Ballanceら、Biochem.Biophy 40 s.Res.Commun.(1983)112:284-289;Tilburnら、Gene(1983)26:205-221、およびYelton ら、Proc.Natl.Acad.Sci.USA(1984)81:1470-1474に記載)、ならびにA.niger(Kellyお よびHynes、EMBO J.(1985)4:475479に記載));Trichoderma reesia(EP 244,234に記 載)、ならびに糸状菌(例えば、Neurospora、Penicillium、Tolypocladium(WO 91/0035 7に記載))。 酵母ベクターのための制御配列は公知であり、そしてアルコールデヒドロゲナーゼ(ADH )(EP 284,044に記載)、エノラーゼ、グルコキナーゼ、グルコース-6-リン酸イソメラ ーゼ、グリセルアルデヒド-3-リン酸-デヒドロゲナーゼ(GAPまたはGAPDH)、ヘキソキナ ーゼ、ホスホフルクトキナーゼ、3-ホスホグリセリン酸ムターゼ、およびピルビン酸キナ ーゼ(Pyk)(EP 329,203に記載)のような遺伝子由来のプロモーター領域を含む。酵母P 50 (17) JP 4270473 B2 2009.6.3 HO5遺伝子(酸性ホスファターゼをコードする)もまた、Myanoharaら、Proc.Natl.Acad.S ci.USA(1983)80:1に記載のように、有用なプロモーター配列を提供する。酵母宿主での 使用のために適切な他のプロモーター配列としては、3-ホスホグリセリン酸キナーゼ(Hi tzemanら、J.Biol.Chem.(1980)255:2073に記載)、またはその他の解糖系酵素(例えば 、ピルビン酸デカルボキシラーゼ、トリオースリン酸イソメラーゼ、およびホスホグルコ ースイソメラーゼ(Hessら、J.Adv.Enzyme Reg.(1968)7:149、およびHollandら、Bioch emistry(1978)17:4900に記載))が挙げられる。増殖条件により制御される転写のさら なる利点を有する誘導性酵母プロモーターとしては、上記のリストからならびに、その他 (アルコールデヒドロゲナーゼ2、イソシトクロムC、酸性ホスファターゼ、窒素代謝に 関連する分解酵素、メタロチオネイン、グリセルアルデヒド-3-リン酸デヒドロゲナーゼ 10 、ならびにマルトースおよびガラクトース利用を担う酵素のプロモーター領域)が挙げら れる。酵母発現における使用のために適切なベクターおよびプロモーターは、Hitzeman、 EP 073,657にさらに記載されている。酵母エンハンサーもまた、酵母プロモーターととも に有利に使用される。さらに、天然に存在しない合成プロモーターもまた、酵母プロモー ターとして機能する。例えば、1つの酵母プロモーターの上流活性化配列(UAS)が、別 の酵母プロモーターの転写活性化領域と連結され得、合成ハイブリッドプロモーターを作 製し得る。そのようなハイブリッドプロモーターの例としては、米国特許第4,876,197号 および同第4,880,734号に記載のような、GAP転写活性化領域に連結されたADH調節配列が 挙げられる。ハイブリッドプロモーターの他の例としては、EP 164,556に記載のような、 解糖系酵素遺伝子(例えば、GAPまたはPyK)の転写活性化領域と組み合わされた、ADH2、 20 GAL4、GAL10、またはPHO5遺伝子のいずれかの調節配列からなるプロモーターが挙げられ る。さらに、酵母プロモーターは、酵母RNAポリメラーゼを結合し、そして転写を開始す る能力を有する非酵母起源の天然に存在するプロモーターを含み得る。 酵母発現ベクター中に含まれ得る他の制御エレメントは、ターミネーター(例えば、GAPD H遺伝子およびエノラーゼ遺伝子由来(Hollandら、J.Biol.Chem.(1981)256:1385に記載 ))、および分泌のためのシグナル配列をコードするリーダー配列である。適切なシグナ ル配列をコードするDNAは、分泌酵母タンパク質の遺伝子(例えば、酵母インベルターゼ 遺伝子(EP 012,873およびJP 62,096,086に記載)およびα因子遺伝子(米国特許第4,588 ,684号、同第4,546,083号、および同第4,870,008号;EP 324,274;およびWO 89/02463に 記載))に由来し得る。あるいは、非酵母起源のリーダー(例えば、インターフェロンリ 30 ーダー)もまた、EP 060,057に記載のように、酵母における分泌を提供する。 外因性DNAを酵母宿主に導入する方法は当該分野で周知であり、代表的には、アルカリカ チオンで処理したスフェロプラストまたは無傷酵母細胞のいずれかの形質転換を含む。酵 母への形質転換は、Van Solingenら、J.Bact.(1977)130:946およびHsiaoら、Proc.Natl .Acad.Sci.USA(1979)76:3829に記載の方法に従って実施され得る。しかし、DNAを細胞 に導入するための他の方法(例えば、核注入、エレクトロポレーション、またはプロトプ ラスト融合)もまた、Sambrookら(上記)に一般的に記載されるようにして用いられ得る 。 酵母分泌について、ネイティブなポリペプチドシグナル配列は、酵母インベルターゼリー ダー、α因子リーダー、または酸ホスファターゼリーダーにより置換され得る。2μプラ 40 スミド開始点由来の複製開始点が酵母に適切である。酵母における使用のための適切な選 択遺伝子は、Kingsmanら、Gene(1979)7:141またはTschemperら、Gene(1980)10:157に 記載の酵母プラスミドに存在するtrp1遺伝子である。trp1遺伝子は、トリプトファンで生 育する能力を欠損する酵母変異株に対する選択マーカーを提供する。同様に、Leu2欠損酵 母株(ATCC 20,622または38,626)は、Leu2遺伝子を有する公知のプラスミドにより相補 される。 酵母における本発明のポリペプチドの細胞内産生については、酵母タンパク質をコードす る配列が、発現の際に酵母細胞により細胞内で切断され得る融合タンパク質を産生するた めに、Fab分子のコード配列に連結され得る。このような酵母リーダー配列の一例は、酵 母ユビキチン遺伝子である。 50 (18) JP 4270473 B2 2009.6.3 iii.昆虫細胞における発現 Fab分子はまた、昆虫発現系においても産生され得る。例えば、バキュロウイルス発現ベ クター(BEV)は、発現されるべき外来性遺伝子のコード配列が、SmithおよびSummers、 米国特許第4,745,051号に記載のように、ウイルス遺伝子(例えば、ポリヘドリン)の代 わりに、バキュロウイルスプロモーターの後方に挿入されている組換え昆虫ウイルスであ る。 本明細書中の発現構築物は、昆虫細胞系の感染または形質転換のための中間物として有用 なDNAベクターを含み、このベクターは、一般に、バキュロウイルス転写プロモーターを コードするDNAを含み、必要に応じてであるが好ましくは、下流に、所望のタンパク質の 分泌を導き得る昆虫シグナルDNA配列、および外来性タンパク質をコードする外来性遺伝 10 子の挿入部位を含み、シグナルDNA配列および外来性遺伝子は、バキュロウイルスプロモ ーターの転写制御下に置かれ、本明細書中での外来性遺伝子は、Fab分子のコード配列で ある。 本明細書中で用いられるプロモーターは、一般に昆虫に感染する、500を超えるバキュロ ウイルスのいずれかに由来するバキュロウイルス転写プロモーター領域であり得る。例え ば、Lepidoptera、Diptera、Orthoptera、Coleoptera、およびHymenopteraの目が挙げら れる。例えば、Autographo californica MNPV、Bombyx mori NPV、rrichoplusia ni MNPV 、Rachlplusia ou MNPVまたはGalleria mellonella MNPV、Aedes aegypti、Drosophila m elanogaster、Spodoptera frugiperda、およびTrichoplusia niのウイルスDNAを含むが、 これらに限定されない。従って、バキュロウイルス転写プロモーターは、例えば、バキュ 20 ロウイルス前初期遺伝子IEIまたはIENプロモーター;39Kおよび後初期遺伝子を含むHindI IIフラグメントからなる群から選択されるバキュロウイルス後初期遺伝子と組み合わせた 前初期遺伝子プロモーター領域;またはバキュロウイルス後期遺伝子プロモーターであり 得る。前初期プロモーターまたは後初期プロモーターは、転写エンハンサーエレメントを 用いて増強され得る。 本明細書中における使用に特に適切なプロモーターは、バキュロウイルスの強ポリヘドリ ンプロモーター(高レベルのDNA挿入物の発現を導く、Friesenら、(1986)THE MOLECULA R BIOLOGY OF BACULOVIRUSES(W.Doerfler編)の「The Regulation of Baculovirus Gene Expression」;EP 127,839およびEP 155,476に記載);およびp10タンパク質をコードす る遺伝子由来のプロモーター(Vlakら、J.Gen.Virol.(1988)69:765-776に記載)である 30 。 本明細書中で用いられるプラスミドはまた、通常、ポリヘドリンポリアデニル化シグナル (Millerら、Ann.Rev.Microbiol.(1988)42:177に記載)、および原核生物アンピシリン 耐性(amp)遺伝子、およびE.coliにおける選択および増殖のための複製開始点を含む。 適切なシグナル配列をコードするDNAもまた含まれ得、そして一般に分泌される昆虫また はバキュロウイルスタンパク質の遺伝子(例えば、バキュロウイルスポリヘドリン遺伝子 )(Carbonellら、Gene(1988)73:409に記載)、ならびに哺乳動物シグナル配列(例え ば、ヒトαインターフェロン(Maedaら、Nature(1985)315:592-594に記載);ヒトガス トリン放出ペプチド(Lebacq-Verheydenら、Mol.Cell.Biol.(1988)8:3129に記載);ヒ トIL-2(Smithら、Proc.Natl.Acad.Sci.USA(1985)82:8404に記載;マウスIL-3(Miyaji 40 maら、Gene(1987)58:273に記載);およびヒトグルコセレブロシダーゼ(Martinら、DN A(1988)7:99に記載)をコードする遺伝子由来の配列に由来し得る。 多数のバキュロウイルス株および変異株、ならびに宿主由来の対応する許容昆虫宿主細胞 (例えば、Spodoptera frugiperda(毛虫)、Aedes aegypti(蚊)、Aedes albopictus( 蚊)、Drosophila melanogaster(ショウジョウバエ)、およびBombyx mori宿主細胞が同 定されており、そして本明細書中で用いられ得る。例えば、Luckowら、Bio/Technology( 1988)6:47-55、Millerら、GENETIC ENGINEERING(Setlow,J.K.ら編)第8巻(Plenum Pub lishing,1986)277∼279頁、およびMaedaら、Nature(1985)315:592-594を参照のこと。 種々のこのようなウイルス株は、公的に入手可能である。例えば、Autographa californi ca NPVのL-1変異株およびBombyx mori NPVのBm-5株が挙げられる。このようなウイルスが 50 (19) JP 4270473 B2 2009.6.3 、宿主細胞(例えば、Spodoptera frugiperda細胞)のトランスフェクションのためのウ イルスとして用いられ得る。 ポリヘドリンプロモーターに加えて、他のバキュロウイルス遺伝子がバキュロウイルス発 現系において用いられ得る。これらは、それらが発現されるウイルス感染段階に従って、 前初期(α)、後初期(β)、後期(γ)、極後期(δ)を含む。これらの遺伝子の発現 は、おそらく転写調節の「カスケード」機構の結果として、連続的に生じる。従って、前 初期遺伝子は、他のウイルス機能の非存在下で感染直後に発現され、そして得られた遺伝 子産物の1つ以上が後初期遺伝子の転写を誘導する。次いで、いくつかの後初期遺伝子産 物が、後期遺伝子の転写を誘導し、そして最終的に極後期遺伝子が、1つ以上のより早い クラスから既に発現された遺伝子産物の制御下で発現される。この調節カスケードの1つ 10 の比較的十分に規定された成分はIEIであり、これは、Autographo californica核ポリヘ ドロシスウイルス(AcMNPV)の好ましい前初期遺伝子である。IEIは、他のウイルス機能 の非存在下では抑制され、後初期クラスのいくつかの遺伝子(好ましい39K遺伝子(Guari noおよびSummers、J.Virol.(1986)57:563-571およびJ.Virol.(1987)61:2091-2099に 記載)を含む)ならびに後期遺伝子(GuannoおよびSummers、Virol.(1988)162:444-451 に記載)の転写を刺激する産物をコードする。 上記の前初期遺伝子は、後初期カテゴリーのバキュロウイルス遺伝子プロモーター領域と 組み合わせて用いられ得る。前初期遺伝子と異なり、このような後初期遺伝子は、他のウ イルス遺伝子または遺伝子産物(例えば、前初期遺伝子の遺伝子産物)の存在を必要とす る。前初期遺伝子は、いくつかの後初期遺伝子プロモーター領域(例えば、39Kまたはバ 20 キュロウイルスゲノムのHindIIIフラグメントに見出される後初期遺伝子プロモーターの 1つ)のいずれかと共に組み合わせられ得る。本発明の場合では、39Kプロモーター領域 が、発現されるべき外来性遺伝子に、その発現がIEIの存在によりさらに制御され得るよ うに連結され得る(L.A.GuarinoおよびSummers(1986a)上記;GuarinoおよびSummers(19 86b)J.Virol.(1986)60:215-223;ならびにGuarinoら、(1986c)J.Virol.(1986)60:2 24-229に記載)。 さらに、前初期遺伝子と後初期遺伝子プロモーター領域との組み合わせが用いられる場合 、異種遺伝子の発現の増強は、後初期遺伝子プロモーター領域と直接シス連結したエンハ ンサー配列の存在により実現され得る。このようなエンハンサー配列は、前初期遺伝子ま たはその産物が限られる状況で後初期遺伝子発現を増強することにより特徴づけられる。 30 例えば、hr5エンハンサー配列は、後初期遺伝子プロモーター領域である39Kに直接、シス で連結され得、それによりクローン化された異種DNAの発現を増強する(GuarinoおよびSu mmers(1986a)、(1986b)、およびGuarinoら(1986)に記載)。 ポリヘドリン遺伝子は、極後期遺伝子として分類される。従って、ポリヘドリンプロモー ターからの転写は、未知であるが、おそらく大多数の他のウイルスおよび細胞遺伝子産物 の事前の発現を必要とする。ポリヘドリンプロモーターのこの遅延発現のために、標準的 なBEV(例えば、SmithおよびSummersにより例えば米国特許第4,745,051号に記載の例示の BEV)は、残りのウイルスゲノムからの遺伝子発現の結果としてのみ、かつウイルス感染 が十分に行われた後のみ、外来性遺伝子を発現する。このことは、現存のBEVの使用に対 する制限を表す。宿主細胞が新規に合成されたタンパク質をプロセスする能力は、バキュ 40 ロウイルス感染の進行につれて減少する。従って、ポリヘドリンプロモーターからの遺伝 子発現は、新規に合成されたタンパク質をプロセスする宿主細胞の能力が特定のタンパク 質(例えば、ヒト組織プラスミノーゲン活性化因子)について実質的に減少した時点で生 じる。結果として、BEV系における分泌糖タンパク質の発現は、クローン化遺伝子産物の 不完全な分泌により複雑化され、それにより不完全にプロセシングされた形態で細胞内の クローン化遺伝子産物を捕捉する。 昆虫シグナル配列が、切断されて成熟タンパク質を生じ得る外来性タンパク質を発現させ るために用いられ得ることが認識されているが、本発明はまた、哺乳動物シグナル配列を 用いても実施され得る。 本明細書中で適切な例示の昆虫シグナル配列は、Lepidopteran脂質動態ホルモン(AKH) 50 (20) JP 4270473 B2 2009.6.3 ペプチドをコードする配列である。AKHファミリーは、昆虫においてエネルギー基質動員 および代謝を調節する短いブロックされた神経ペプチドからなる。好ましい実施態様では 、Lepidopteran Manduca sexta AKHシグナルペプチドをコードするDNA配列が用いられ得 る。他の昆虫AKHシグナルペプチド(例えば、Orthoptera Schistocerca gregaria遺伝子 座からのAKHシグナルペプチド)もまた好都合に用いられ得る。別の例示の昆虫シグナル 配列は、Dorosophila角皮タンパク質をコードする配列である(例えば、CPI、CP2、CP3ま たはCP4)。 現在、本明細書中で外来性遺伝子をAcNPVに導入するために用いられ得る、最も通常に用 いられている移入ベクターは、pAc373である。当業者に公知の多くの他のベクターもまた 、本明細書中で使用され得る。バキュロウイルス/昆虫細胞発現系のための材料および方 10 法は、Invitrogen(San Diego CA)のような会社からキットの形態で市販されている(「 MaxBac」キット)。本明細書中で利用される技術は、当業者に一般に知られ、そしてSumm ersおよびSmith、A MANUAL OF METHODS FOR BACULOVIRUS VECTORS AND INSECT CELL CULT URE PROCEDURES、Texas Agricultural Experiment Station Bulletin No.1555、Texas A& M University(1987);Smithら、Mol.Cell.Biol.(1983)およびLuckowおよびSummers(1 989)に充分に記載されている。これらは、例えば、pVL985の使用を含む。これは、ポリ ヘドリン出発コドンをATGからATTに変え、そしてATTの下流にBamHIクローン化部位32塩基 対を導入している(LuckowおよびSummers、Virology(1989)17:31に記載)。 従って、例えば、本発明のポリペプチドの昆虫細胞発現について、所望のDNA配列が、公 知の技術を用いて、移入ベクター中に挿入され得る。昆虫細胞宿主は、挿入された所望の 20 DNAを野生型バキュロウイルスのゲノムDNAを共に含む移入ベクターで、通常同時トランス フェクションにより、同時形質転換され得る。ベクターおよびウイルスゲノムは組換えら れ得、容易に同定され精製され得る組換えウイルスを生じる。パッケージングされた組換 えウイルスは、昆虫宿主細胞を感染し、Fab分子を発現させるために用いられ得る。 本明細書中で適用可能な他の方法は、昆虫細胞培養の標準的な方法であり、同時トランス フェクションおよびプラスミドの調製は、SummersおよびSmith(1987)(上記)に記載さ れる。この参考文献はまた、遺伝子のAcMNPV移入ベクターへのクローン化、プラスミドDN A単離、遺伝子のAcmMNPVゲノムへの移入、ウイルスDNA精製、組換えタンパク質の放射性 標識、および昆虫細胞培養培地の調製の、標準的な方法にも関する。ウイルスおよび細胞 の培養のための手順は、VolkmanおよびSummers、J.Virol.(1975)19:820-832およびVolk 30 manら、J.Virol.(1976)19:820-832に記載される。 iv.哺乳動物細胞における発現 Fab分子を産生するために、哺乳動物発現系もまた用いられ得る。哺乳動物細胞発現のた めの代表的なプロモーターは、とりわけ、SV40初期プロモーター、CMVプロモーター、マ ウス乳ガンウイルスLTRプロモーター、アデノウイルス主要後期プロモーター(Ad MLP) 、および単純ヘルペスウイルスプロモーターを含む。他の非ウイルスプロモーター(例え ば、マウスメタロチオネイン遺伝子由来のプロモーター)もまた、哺乳動物構築物におい て利用されている。哺乳動物発現は、プロモーターに依存して、構成的または制御(誘導 的)のいずれかであり得る。代表的には、転写終結およびポリアデニル化配列もまた、翻 訳停止コドンの3’に位置して存在する。好ましくは、翻訳の開始を最適化するための配 40 列もまた、Fabコード配列の5’に位置して存在する。転写終結/ポリアデニル化シグナル の例は、Sambrookら、(1989)MOLECULAR CLONING:A LABORATORY MANUAL,第2版、(Cold Spring Harbor Press,Cold Spring Harbor,N.Y.)に記載のように、SV40由来のシグナル を含む。スプライスドナーおよびアクセプター部位を含むイントロンもまた、本発明の構 築物中に設計され得る。 本明細書において、エンハンサーエレメントもまた使用されて哺乳動物構築物の発現レベ ルを増大し得る。例としては、SV40初期遺伝子エンハンサー(Dijkemaら、EMBO J.(1985 )4:761に記載)、ならびにラウス肉腫ウイルス(Gormanら、Proc.Natl.Acad.Sci.USA(1 982b)79:6777に記載)、およびヒトサイトメガロウイルス(Boshartら、Cell(1985)41 :521に記載)の長末端反復(LTR)由来のエンハンサー/プロモーターを含む。哺乳動物 50 (21) JP 4270473 B2 2009.6.3 細胞における外来タンパク質の分泌を提供するシグナルペプチドをコードする配列を含む リーダー配列もまた、存在し得る。好ましくは、リーダーフラグメントと目的の遺伝子と の間にコードされるプロセシング部位が存在し、その結果リーダー配列はインビボまたは インビトロのいずれかで切断され得る。アデノウイルストリパータイトリーダーは、哺乳 動物細胞における外来タンパク質の分泌を提供するリーダー配列の例である。 Fab分子をコードするDNAの、哺乳動物における一過的な発現を提供する発現ベクターが存 在する。一般に、一過的な発現は発現ベクターの使用を含む。このベクターは、宿主細胞 において効率的に複製し得、その結果、宿主細胞は発現ベクターの多数のコピーを蓄積し 、次いで発現ベクターによってコードされる高レベルの所望のポリペプチドを合成する。 適切な発現ベクターおよび宿主細胞を含む一過的な発現系は、クローン化DNAによってコ 10 ードされるポリペプチドの好都合な陽性の同定、ならびに所望の生物学的または生理学的 特性についてのこのようなポリペプチドの迅速なスクリーニングを可能にする。一旦完成 すれば、哺乳動物発現ベクターは、任意のいくつかの哺乳動物細胞を形質転換するために 使用され得る。異種ポリヌクレオチドを哺乳動物細胞へ導入するための方法は当該分野で 公知であり、そしてデキストラン媒介トランスフェクション、リン酸カルシウム沈殿、ポ リブレン媒介トランスフェクション、プロトプラスト融合、エレクトロポレーション、リ ポソームへのポリヌクレオチド(単数または複数)のカプセル化、および核へのDNAの直 接的マイクロインジェクションを含む。哺乳動物細胞宿主系形質転換の一般的な局面は、 U.S.4,399,216によって記載されている。ポリヌクレオチドトランスフェクションに特に 有用な合成脂質は、N-[1(2,3-ジオレイルオキシ)プロピル]-N,N,N-トリメチル塩化アン 20 ▲R▼ モニウムである。これはLipofectin の名で市販されており(BRL,Gaithersburg,MD から入手可能)、そしてFelgnerら、Proc.Natl.Acad.Sci.USA(1987)84:7413によって記 載されている。 発現のための宿主として入手可能な哺乳動物細胞株もまた公知であり、そしてアメリカン タイプカルチャーコレクション(ATCC)から入手可能な多数の不死化細胞株を含む。これ らには、チャイニーズハムスター卵巣(CHO)細胞、HeLa細胞、ベビーハムスター腎臓(B HK)細胞、サル腎臓細胞(COS)、ヒト肝臓ガン細胞(例えば、Hep G2)、ヒト胚腎臓細 胞、ベビーハムスター腎臓細胞、マウスセルトリ細胞、イヌ腎臓細胞、バッファローラッ ト肝細胞、ヒト肺細胞、ヒト肝細胞、マウス乳ガン細胞などを含むが、これらに限定され ない。 30 本発明のFab分子を産生するための哺乳動物宿主細胞は、種々の培地中で培養され得る。 市販の培地(例えば、Ham F10(Sigma)、最少必須培地([MEM],Sigma)、RPMI-1640(Si gma)、およびダルベッコ改変イーグル培地([DMEM],Sigma))は宿主細胞を培養するた めに適切である。さらに、以下に記載される任意の培地が宿主細胞の培養培地として使用 され得る:HamおよびWallace,Meth.Enz.(1979)58:44、BarnesおよびSato,Anal.Biochem. (1980)102:255、米国特許第4,767,704号、同第4,657,866号、同第4,927,762号、または 同第4,560,655号、WO 90/103430、WO 87/00195、およびU.S.RE 30,985。これらの任意の 培地は、必要に応じて、ホルモンおよび/または他の増殖因子(例えば、インスリン、ト ランスフェリン、または上皮成長因子)、塩(例えば、塩化ナトリウム、カルシウム、マ グネシウム、およびリン酸塩)、緩衝液(例えば、HEPES)、ヌクレオシド(例えば、ア 40 デノシンおよびチミジン)、抗生物質(例えば、ゲンタマイシン(tm)M薬剤)、微量元 素(マイクロモルの範囲の最終濃度で通常存在する無機化合物として定義される)、およ びグルコースまたは等価のエネルギー供給源が補充され得る。適切な濃度で含まれ得る他 の任意の必要な補充物は、当業者に公知である。培養条件(温度、Phなど)は、発現のた めに選択された宿主細胞に以前から用いられているものであり、そして当業者には明らか である。 特異的結合分子の調製 上記の技術を用いて、HCV E2抗原に対して免疫学的結合親和性を示す、多数の特異的結合 分子が提供され得る。詳細には、選択された発現系および宿主に依存して、上記の発現ベ クターによって形質転換された増殖する宿主細胞により、重鎖および軽鎖部分が発現され 50 (22) JP 4270473 B2 2009.6.3 る条件下で、可溶性Fab特異的結合分子が容易に産生され得る。非共有結合した重鎖およ び軽鎖を含むヘテロダイマーは、宿主細胞から単離され、そして精製され得る。本発明は 、重鎖および軽鎖ポリペプチドの必要に応じた分泌もまた提供するので、Fabヘテロダイ マーは培地から直接的に精製され得る。適切な増殖条件および再生方法の選択は、当業者 の範囲内である。 さらに、本発明のFab分子は、確認されたアミノ酸配列に基づいて、タンパク質合成の従 来の方法を用いて産生され得る。一般に、これらの方法は、成長するペプチド鎖への1つ 以上のアミノ酸の連続的な付加を用いる。通常、第1のアミノ酸のアミノ基またはカルボ キシル基のいずれかは、適切な保護基によって保護される。次いで、保護または誘導体化 アミノ酸は、不活性固形支持体に結合されるか、または溶液中で利用され、アミド結合を 10 形成させる条件下で、適切に保護された相補性(アミノまたはカルボキシル)基を有する 次のアミノ酸に順番に付加される。次いで、保護基は、新たに付加されたアミノ酸残基か ら除去され、そして次に、次のアミノ酸(適切に保護された)が付加され、後も同様であ る。所望のアミノ酸が正しい配列で結合された後、任意の残りの保護基(および固相合成 技術が用いられた場合は、任意の固形支持体)は続いてまたは同時に除去され、最終的な ポリペプチドを生じる。この一般的手順の簡単な改変によって、成長する鎖に、1つ以上 のアミノ酸を一度に付加し得る。例えば、保護トリペプチドを適切に保護されたジペプチ ドに結合し(キラル中心をラセミ化しない条件下で)、脱保護化後、ペンタペプチドを形 成し得る。例えば、固相ペプチド合成技術については、J.M.StewartおよびJ.D.Young,Sol id Phase Peptide Synthesis,第2版、Pierce Chemical Co.,Rockford,IL(1984)ならび 20 にG.BaranyおよびR.B.Merrifield,The Peptides:Analysis,Synthesis,Biology,E.Grossお よびJ.Meienhofer編、第2巻、Academic Press,New York,(1980),3∼254頁;古典的な 溶液合成については、M.Bodansky,Principles of Peptide Synthesis,Springer-Verlag,B erlin(1984)ならびにE.GrossおよびJ.Meienhofer編,The Peptides:Analysis,Synthesi s,Biology、前出、第1巻を参照のこと。 組換えヒトモノクローナル抗体特異的結合分子が、公知の技術を用いて、Fab分子から調 製され得る。Benderら(1992)Hum Antibod Hybridomas 4:74。詳細には、選択されたFab クローンの重鎖部分のコード配列は、ヒトIg重鎖の定常ドメインのコード配列と共に、上 記の種々の組換え技術を用いて発現ベクターに挿入され得る。例えば、この目的のために 、哺乳動物発現ベクターpSG5(Greenら(1998)Nucleic Acids Res 16:369)が使用され 30 得る。 クローニングは、細菌リーダー配列(ファージミドベクター由来)を除去し、重鎖コード 配列のN末端をヒトコンセンサス配列に改変するための、重複PCRを含む。同様に、選択 されたFabクローンの軽鎖部分のコード配列は、ヒトコンセンサス配列を含むようにN末 端改変され、そしてPSG5のような発現ベクターにクローン化される。PSG5ベクターはSV40 複製起点を含み、その結果哺乳動物細胞(例えば、COS-7細胞)への重鎖および軽鎖ベク ターの同時トランスフェクトによって、機能的な抗体分子の産生が確認され得る。Burton ら(1994)Science 266:1024-1027。 重鎖および軽鎖は、引き続いて別々のクローニングベクターにクローン化され得、そして 重鎖または軽鎖コード配列のいずれかは他のベクターにサブクローン化され、コンビナト 40 リアルプラスミドを提供する。例えば、重鎖および軽鎖コード配列は、ヒトサイトメガロ ウイルスプロモーターおよびグルタミンシンセターゼ選択マーカーを含むpEE6およびpEE1 2ベクターにそれぞれ挿入され得る(Bebbingtonら(1992)Bio/Technology 10:169)。次 いで、重鎖は、PEE6ベクター由来の制御エレメントと共に、PEE12ベクターにサブクロー ン化され、コンビナトリアルプラスミドを提供し得る。コンビナトリアルプラスミドは、 CHO細胞発現系において発現され得る。十分なレベルの組換え抗体産生を提供するCHO発現 系由来のこれらのクローンは、拡大のために選択され得る。CHO系において発現された組 換え抗体は、公知の技術(例えば、プロテインAを用いるアフィニティクロマトグラフィ ー)を用いて精製され、そして組換え特異的結合分子の結合親和性は、上記のELISA阻害 アッセイを用いて評価され得る。 50 (23) JP 4270473 B2 2009.6.3 あるいは、Fabクローンのコード配列は、ベクターpcLCHCおよびpcIgG1にそれぞれ移入さ れ、次いで前記のように、CHO細胞中で完全なIgGとして発現され得る。Samuelssonら(19 96)Eur.J.Immunol.26:3029。 組換えF(ab’)2および組換えFv特異的結合分子もまた、公知の技術を用いて、ファージ 由来Fabクローンから調製され得る。Fv分子は、一般に、Fab分子の抗原結合部分を含み、 非共有結合した重鎖:軽鎖ヘテロダイマーを有し、そして天然の抗体分子のほとんどの抗 原認識および結合能力を保持する。Inbarら(1972)Proc.Nat.Acad.Sci.USA 69:2659-266 2;Hochmanら(1976)Biochem 15:2706-2710;およびEhrlichら(1980)Biochem 19:4091-4 096。代表的には、組換えモノクローナル抗体を構築するために使用した上記の組換え技 術は改変され、切断型の特異的結合分子を提供し得る。これらの分子もまた、上記のよう 10 にCHO発現系にクローン化され、精製され、そして特徴付けられ得る。 ファージ由来のFabクローンは、公知の技術を使用して一本鎖Fv(Sfv)分子を提供するた めにさらに使用され得る。これらのSfv分子は、ファージ由来Fab分子から得られた重鎖お よび軽鎖コード配列を含む遺伝子融合物から発現される、共有結合した重鎖:軽鎖ヘテロ ダイマーを有する。ここで鎖はペプチドコードリンカーにより結合される。Hustonら(19 88)Proc.Nat.Acad.Sci.USA 85(16):5879-5883。自然に凝集するが化学的に分離される 軽鎖および重鎖を、抗原結合部位の構造に実質的に類似した3次元構造に折り畳まれるSf v分子に変換するために、化学構造を識別するための多数の方法が記載されている。例え ば、Hustonらの米国特許第5,091,513号および同第5,132,405号、ならびにLadnerらの同第 4,946,778号を参照のこと。 20 本発明の実施に当たって、重鎖および軽鎖をSfv結合分子へと結合するために適切な化学 構造を開発するために、組換えDNA設計方法が使用される。設計基準は、1つの鎖のC末 端と別の鎖のN末端との間の距離を隔てる適切な長さの決定を含む。ここで、リンカーは 、一般に、コイル化または2次構造を形成しない傾向にある小さな親水性アミノ酸残基か ら形成される。このような方法は当該分野において記載されている。例えば、Hustonらの 米国特許第5,091,513号および同第5,132,405号、ならびにLadnerらの同第4,946,778号を 参照のこと。 リンカー設計の最初の一般的な工程は、結合されるべき妥当な部位の同定を含む。各々の 免疫グロブリン鎖上の適切な結合部位は、重鎖および軽鎖からの最少の残基の損失を生じ るものを含み、そして分子安定性の必要に一致する最少の残基の数を有するリンカーを必 30 要とする。一対の部位は、結合されるべき「ギャップ」を定義する。1つの鎖のC末端を 次のN末端に結合するリンカーは、一般に、生理学的溶液中で非構造的立体配置をとり、 そして好ましくは重鎖および軽鎖の正確な折り畳みを阻害し得る大きな側基を有する残基 を有さない、親水性アミノ酸を含む。従って、適切なリンカーは、グリシンおよびセリン 残基の交互の対のポリペプチド鎖を含み、そして可溶性を増強するために挿入されるグル タミン酸およびリジン残基を含み得る。本発明の実施に使用される1つの特別なリンカー は、アミノ酸配列[(Gly)4Ser]3を有する。別の特に好ましいリンカーは、[(Ser)4Gly ]の2または3反復を含むアミノ酸配列(例えば、[(Ser)4Gly]3)を有する。このよう なリンカー部分をコードするヌクレオチド配列は、当該分野で公知の種々のオリゴヌクレ オチド合成技術を用いて、容易に提供され得る。例えば、Sambrook、およびManiatis、前 40 出を参照のこと。 一旦適切なリンカー配列が決定されると、Sfv分子をコードするヌクレオチド配列は、オ ーバーラップPCRアプローチを用いて結合され得る。例えば、Hortonら(1990)BioTechni ques 8:528-535を参照のこと。選択されたリンカー配列を介して結合されるべき軽鎖およ び重鎖の末端は、最初に鎖領域の末端配列、続いて所望のリンカー配列の全てまたはほと んどを含むプライマーを用いて、各鎖のPCR増幅により伸長される。この伸長工程の後、 軽鎖および重鎖は、重複する伸長部分(これは、一緒になって全リンカー配列を含み、そ して重複部分でアニールされ、そしてPCRで伸長され、当該分野で公知の方法を用いて完 全なSfv配列を入手し得る)を含む。次いで、発現カセット中に存在するsFv配列を含む遺 伝子は、適切な発現系において発現され、そしてこれから産生されたsFv分子は、上記の 50 (24) JP 4270473 B2 2009.6.3 ように精製されそして特徴付けられ得る。 ワクチン組成物 治療的および予防的ワクチン組成物が本明細書中に提供される。この組成物は、一般に、 1つ以上の上記の抗HCVモノクローナル抗体(Fab分子、Fvフラグメント、sFv分子、およ びそれらの組合せを含む)の混合物を含む。予防的ワクチンはHCV感染を予防するために 用いられ得、そして治療的ワクチンはHCV感染後の個体を処置するために用いられ得る。 予防的使用は、ワクチン接種した被験体における、HCVに対する増加した抗体力価の供給 を含む。この様式で、HCV感染に罹る高い危険性のある被験体(例えば、免疫無防備の個 体、臓器移植患者、血液または血液産物の輸血剤を得ている個体、およびHCV感染した個 体と密接な個人的接触をした個体)は、HCV因子に対する受動免疫が提供され得る。さら 10 に、本明細書中で産生されるモノクローナル抗体、Fab分子、およびsFv分子の交叉反応性 に起因して、防御のレベルは、多数の異種HCV単離株に対して提供される。本発明の抗HCV ワクチンについての他の予防的使用は、感染性因子に対する曝露の後の個体におけるHCV 疾患の予防を含む。本発明のワクチンの治療的使用は、感染した個体からの感染性因子の 減少および/または除去の両方、ならびに循環するHCVおよびこの疾患が伝播する可能性 の減少および/または除去を含む。 組成物は、付属的な免疫調節剤(例えば、IL-2、改変IL-2(cys125→ser125)、GM-CSF、 IL-12、γ-インターフェロン、IP-10、MIP1β、およびRANTESを含むがこれらに限定され ないサイトカイン、リンホカイン、およびケモカイン)と共に投与され得る。ワクチン組 成物が治療的ワクチンとして使用される場合、組成物は、公知の抗HCV療法(例えば、1 20 週間に3回の皮下での300万単位のα-IFNの投与を一般的に必要とする、α-インターフェ ロン(α-IFN)療法(Causseら(1991)Gastroenterology 101:497-502,Davisら(1989) N Engl J Med 321:1501-1506,Marcellinら(1991)Hepatology 13:393-397)、インター フェロンβ(β-IFN)療法(Omataら(1991)Lancet 338:914-915)、リビビリン療法(D i Bisceglieら(1992)Hepatology 16:649-654,Reichardら(1991)Lancet 337:1058-106 1)およびアンチセンス療法(Wakitaら(1994)J Biol Chem 269:14205-14210))と共に 投与され得る。本発明のモノクローナル抗体を含む治療的ワクチン組成物はまた、公知の 抗HCV組合せ療法(例えば、α-IFNとウルソジオール(ursodiol)との組合せ(Bottelli ら(1993)(要約)Gastroenterology 104:879,O’Brienら(1993)(要約)Gastroenter ology 104:966)、およびβ-IFNとリビビリンとの組合せ(Kakumuら(1993)Gastroenter 30 ology 105:507-512))と共に用いられ得る。 1つ以上の抗体、抗体フラグメント、sFv分子およびそれらの組合せを成分として含むワ クチン組成物の調製物は、一般に、当該分野で公知である。代表的には、このようなワク チンは、注射可能物質(例えば、液体溶液もしくは懸濁液、または注射前に液体中に溶解 もしくは懸濁するのに適切な固体形態のいずれか)として調製される。組成物はまた、一 般に、1つ以上の「薬学的に受容可能な賦形剤またはビヒクル」(例えば、水、生理食塩 水、デキストロース、グリセロール、エタノールなどまたはそれらの組合せ)を含む。さ らに、微量の補助的な物質(例えば、湿潤剤または乳化剤、pH緩衝化物質など)が、この ようなビヒクル中に存在し得る。ワクチン組成物は、乳化され得るか、または活性成分( モノクローナル抗体)は、リポソーム中にカプセル化され得る。 40 一旦処方されると、ワクチン組成物は、例えば、注射(皮下または筋肉内のいずれか)に より、従来通りに非経口投与される。他の様式の投与に適切なさらなる処方物は、経口お よび肺の処方物、坐剤、ならびに経皮適用を含む。坐剤のためには、伝統的なバインダー およびキャリアは、例えば、ポリアルキレングリコールまたはトリグリセリドを含み得る 。このような坐剤処方物は、0.5%∼10%、好ましくは1%∼2%の範囲の活性成分(単 数または複数)を含む混合物から提供され得る。経口処方物は、このような通常用いられ る賦形剤(例えば、薬学的グレードのマンニトール、ラクトース、デンプン、ステアリン 酸マグネシウム、サッカリンナトリウム、セルロース、炭酸マグネシウムなど)を含む。 ワクチン組成物は、投薬処方と適合性の様式でかつ予防的および/または治療的に有効で ある量で、処置される被験体に投与される。送達される組成物の量(一般に、1用量あた 50 (25) JP 4270473 B2 2009.6.3 り活性薬剤1∼500マイクログラムの範囲)は、処置される被験体、それ自身の免疫応答 をマウントするための被験体の免疫系能力、および所望される防御の程度に依存する。必 要な正確な量は、他の要因もあるが、処置される個体の年齢および全身状態、処置される 状態の重篤度、ならびに選択される特定の抗HCV剤およびその投与様式に依存して変化す る。適切な有効量は、当業者により容易に決定され得る。従って、組成物の「治療有効量 」は、HCV疾患の症状の処置または予防をもたらすために十分であり、そして日常的な治 験を通して決定され得る比較的広範な範囲内にある。 さらに、ワクチン組成物は、単回用量スケジュールで、または好ましくは多回用量スケジ ュールで与えられ得る。多回用量スケジュールは、最初のワクチン接種の過程が1∼10の 別々の用量であり得、その後他の用量が、組成物の作用を維持または補強するために必要 10 なその後の時間間隔で与えられるものである。従って、投薬養生法はまた、少なくとも部 分的に、処置される被験体の特定の必要性に基づいて決定され、そして当業者の判断に依 存する。 遺伝子治療 組換えモノクローナル抗体はまた、遺伝子治療に用いられ得る。これに関して、組換え抗 体をコードする遺伝子は、適切な哺乳動物宿主細胞に、発現または同時発現のために、哺 乳動物細胞への遺伝子移入のために開発されたウイルスに基づく多くの系を用いて導入さ れ得る。例えば、レトロウイルスは、遺伝子送達系の便利なプラットフォームを提供する 。VHおよび/またはVLドメインポリペプチドをコードする選択されたヌクレオチド配列 は、当該分野で公知の技術を用いて、ベクターに挿入され、そしてレトロウイルス粒子中 20 にパッケージングされ得る。次いで、組換えウイルスは単離され、そして被験体に送達さ れ得る。多くの適切なレトロウイルス系が記載されている(米国特許第5,219,740号;Mill erおよびRosman(1989)BioTechniques 7:980-990;Miller,A.D.(1990)Human Gene Ther apy-1:5-14;Scarpaら(1991)Virology 180:849-852;Burnsら,(1993)Proc.Natl.Acad. Sci.USA 90:8033-8037;およびBoris-LawrieおよびTemin(1993)Cur.Opin.Genet.Develop .3:102-109)。本明細書中の遺伝子治療のためのレトロウイルスベクターを産生および使 用するために特に好ましい方法は、例えば、1991年3月7日に公開された国際公開第WO 9 1/02805号、および「Eukarotic Layered Vector Initiation Systems」について1995年3 月15日に出願された米国特許出願第08/404,796号;「Recombinant α-Viral Vectors」に ついて1995年3月15日に出願された同第08/405,627号;および「Packaging Cells」につ 30 いて1993年11月23日に出願された同第08/156,789号に記載される。 多くの適切なアデノウイルスベクターがまた記載されている。宿主ゲノム中に組み込まれ るレトロウイルスとは異なり、アデノウイルスは染色体外で存続し、それゆえ挿入変異誘 発に関連する危険性を最小にする(Haj-AhmadおよびGraham(1986)J.Virol.57:267-274; Bettら(1993)J.Virol.67:5911-5921;Mitterederら(1994)Human Gene Therapy 5:717729;Sethら(1994)J.Virol.68:933-940;Barrら(1994)Gene Therapy 1:51-58;Berkner, K.L.(1988)BioTechniques 6:616-629;およびRichら(1993)Human Gene Therapy 4:461 -476)。 種々のアデノ随伴ウイルス(AAV)ベクター系は、最近、遺伝子送達のために、開発され ている。このような系は、制御配列(例えば、プロモーターおよびポリアデニル化部位) 40 ならびに選択マーカーまたはレポーター遺伝子、エンハンサー配列、および転写の誘導を 可能にする他の制御エレメントを含み得る。AAVベクターは、当該分野で周知の技術を用 いて容易に構築され得る。例えば、米国特許第5,173,414号および同第5,139,941号;国際 公開第WO 92/01070号(1992年1月23日公開)および同第WO 93/03769号(1993年3月4日 公開);Lebkowskiら(1988)Molec.Cell.Biol.8:3988-3996;Vincentら(1990)Vaccines 90(Cold Spring Harbor Laboratory Press);Carter,B.J.(1992)Current Opinion in Biotechnology 3:533-539;Muzyczka,N.(1992)Current Topics in Microbiol.and Immu nol.158:97-129;Kotin,R.M.(1994)Human Gene Therapy 5:793-801;ShellingおよびSmit h(1994)Gene Therapy 1:165-169;およびZhouら(1994)J.Exp.Med.179:1867-1875を参 照のこと。 50 (26) JP 4270473 B2 2009.6.3 Fab分子をコードする本発明の核酸分子を送達するための使用を見出すさらなるウイルス ベクターは、ポックスファミリーのウイルス(ワクシニアウイルスおよびニワトリポック スウイルスを含む)に由来するベクターを含む。例としては、遺伝子を発現するワクシニ アウイルス組換え体は、以下の通りに構築され得る。特定のFab分子をコードするDNAは、 まず、このDNAは、ワクシニアプロモーターおよび隣接するワクシニアDNA配列(例えば、 チミジンキナーゼ(TK)をコードする配列)に隣接するように、適切なベクター中に挿入 される。次いで、このベクターは、ワクシニアで同時に感染される細胞をトランスフェク トするために用いられる。相同組換えは、ワクシニアプロモーターおよびFab分子をコー ドする遺伝子をウイルスゲノム中に挿入するように作用する。得られるTK-組換え体は、 細胞を5-ブロモデオキシウリジンの存在下で培養し、そしてそれに耐性のウイルスプラー 10 クを拾うことにより選択され得る。 ワクシニアに基づく感染/トランスフェクション系は、宿主細胞におけるFabフラグメン トの誘導性で一過性の発現を提供するために都合良く用いられ得る。この系において、細 胞は、まず、インビトロで、バクテリオファージT7 RNAポリメラーゼをコードするワクシ ニアウイルス組換え体で感染される。このポリメラーゼは、これがT7プロモーターを有す るテンプレートのみを転写する点で、精巧な特異的を示す。感染後、細胞は、T7プロモー ターにより駆動される目的のポリヌクレオチドでトランスフェクトされる。ワクシニアウ イルス組換え体から細胞質中で発現されるポリメラーゼは、トランスフェクトされたDNA をRNAへと転写し、これは次いで宿主翻訳機構によりタンパク質に翻訳される。この方法 は、多量のRNAおよびその翻訳産物の、高レベルで一過性の細胞質産生を提供する。例え 20 ば、Elroy-SteinおよびMoss,Proc.Natl.Acad.Sci.USA(1990)87:6743-6747;Fuerstら,P roc.Natl.Acad.Sci.USA(1986)83:8122-8126を参照のこと。 あるいは、アビポックスウイルス(例えば、ニワトリポックスウイルスおよびカナリアポ ックスウイルス)はまた、Fabコードヌクレオチド配列を送達するために用いられ得る。 アビボックスベクターの使用は、ヒトおよび他の哺乳動物種において特に望ましい。なぜ なら、アビポックス属のメンバーは、感受性の鳥類種においてのみ生産的に複製し得、従 って、哺乳動物細胞において感染性でないからである。組換えアビポックスウイルスを産 生するための方法は、当該分野において公知であり、そしてワクシニアウイルスの産生に 関して上記のように、遺伝的組換えを使用する。例えば、国際公開第WO 91/12882号;198 9年4月20日に公開された同第WO 89/03429号;および1992年3月5日に公開された同第WO 30 92/03545号を参照のこと。 分子結合ベクター(例えば、MichaelらJ.Biol.Chem.(1993)268:6866-6869およびWagner らProc.Natl.Acad.Sci.USA(1992)89:6099-6103に記載されるアデノウイルスキメラベク ター)はまた、本発明の下で遺伝子送達のために用いられ得る。 アッセイ試薬および診断キット HCV粒子を含む試料と免疫学的に反応し得る上記の抗HCV結合分子(Fab分子、Fvフラグメ ント、およびsFv分子を含む組換えモノクローナル抗体)はまた、生物学的試料の特異的 結合アッセイにおけるHCVウイルス粒子および/またはウイルス抗原の存在を検出するた めに本明細書中で用いられ得る。特に、本発明の新規な特異的結合分子は、HCVを有する および/または感染した個体をスクリーニングおよび同定するため、ならびにHCVで汚染 40 した血液または血液産物をスクリーニングするための高感受性方法において用いられ得る 。本発明の結合分子はまた、処置された個体における抗HCV治療の進行をモニターするた め、およびHCV因子の研究および調査において用いられるHCV培養物の増殖速度をモニター するためのアッセイにおいて用いられ得る。 特異的結合アッセイの形式は、当該分野で周知である手順に従って多数の変更が施される 。例えば、特異的結合アッセイは、本発明に従って調製された組換えヒトモノクローナル 抗体(Fab分子、Fvフラグメント、ならびにsFv分子を含む)の1つまたはいくつかの混合 物を利用するためにフォーマットされ得る。アッセイの形式は、一般に、例えば、競合、 直接的結合反応、またはサンドイッチ型アッセイ技術に基づき得る。さらに、本アッセイ は、免疫沈降または他の技術を用いて実施されて、アッセイの間またはその開始後のアッ 50 (27) JP 4270473 B2 2009.6.3 セイ試薬を分離し得る。他のアッセイは、アッセイの開始前に不溶化されている特異的結 合分子を用いて実施され得る。これに関しては、多くの不溶化技術(免疫吸着剤などへの 吸着による不溶化、反応容器の壁との接触による吸着、不溶性マトリックスもしくは「固 相」基質への共有的架橋、イオン性もしくは疎水性相互作用を用いる固相基質への非共有 的結合、またはポリエチレングリコールのような沈澱剤もしくはグルタルアルデヒドのよ うな架橋剤を用いての凝集を含むが、これらに限定されない)が当該分野で周知である。 当業者により本アッセイにおける使用のために選択され得る多数の固相基体が存在する。 例えば、ラテックス粒子、微粒子、磁性ビーズ、常磁性ビーズ、もしくは非磁性ビーズ、 メンブレン、プラスチックチューブ、マイクロタイターウェルの壁、ガラスまたはシリコ ン粒子、ならびにヒツジ赤血球は、全て本明細書中での使用に適切である。 10 一般に、本アッセイの大部分は、検出可能な標識部分を有する特異的結合分子(組換えモ ノクローナル抗体、Fabフラグメント、Fvフラグメント、およびsFv分子)から形成される 標識化結合複合体の使用を含む。多くのこのような標識は当該分野で公知であり、そして (共有的または非共有的会合技術のいずれかを用いて)本発明の結合分子へ容易に結合さ れて、上記のアッセイ形式における使用のための結合複合体を提供し得る。適切な検出可 能な部分は、放射性同位元素、蛍光剤、発光性化合物(例えば、フルオレセインおよびロ ーダミン)、化学発光剤(例えば、アクリジニウム、フェナントリジニウム、およびジオ キセタン化合物)、酵素(例えば、アルカリホスファターゼ、西洋ワサビペルオキシダー ゼ、およびβ-ガラクトシダーゼ)、酵素基質、酵素補因子、酵素インヒビター、染料、 および金属イオンを含むがこれらに限定されない。これらの標識は、当該分野で公知の結 20 合技術を用いて結合分子と会合され得る。 例示的なアッセイ方法は一般に以下の工程を含む:(1)上記のような検出可能に標識さ れた結合複合体を調製する工程;(2)HCV粒子および/またはHCV抗原を含む疑いのある 試料を入手する工程;(3)特異的結合分子-抗原複合体(例えば、抗体-抗原複合体)の 形成を許容する条件下で、試料と標識された複合体とをインキュベートする工程;および (4)標識された結合分子-抗原複合体の存在または非存在を検出する工程。本明細書を 読んで当業者により理解されるように、このようなアッセイは、ヒト供与者の血液および 血清産物中のHIV感染の存在についてスクリーニングするため、診断的設定および/または 研究的設定におけるHCV培養物の増殖速度をモニターするため、個体におけるHCV感染を検 出するため、または感染被験体における抗HCV処置プロトコルの治療的効果をモニターす 30 るために使用され得る。アッセイが臨床的設定において(例えば、HCV感染を検出するた めまたは抗HCV治療をモニターするため)使用される場合、試料はヒト体液および動物体 液(例えば、全血、血清、血漿、脳脊髄液、尿など)から得られ得る。さらに、アッセイ は、当該分野に公知のような標準品または校正物質に対する参照を使用して、定量的情報 を提供するために容易に使用され得る。 本発明の1つの特定のアッセイ方法において、酵素結合免疫吸着アッセイ(ELISA)が、 試料中のHCV抗原濃度を定量するために使用され得る。その方法において、本発明の特異 的な結合分子は酵素に結合され、標識された結合複合体を提供する。ここで、アッセイは 定量的標識として結合した酵素を使用する。抗原を測定するために、選択されたHCV抗原 に特異的に結合可能な結合分子(例えば、抗体分子)は、固相基質(例えば、マイクロタ 40 イタープレートまたはプラスチックカップ)に固定化され、試験試料希釈物と共にインキ ュベートされ、洗浄され、そして本発明の結合分子-酵素複合体と共にインキュベートさ れ、次いで再度洗浄される。このことに関して、適切な酵素標識は一般に公知であり、こ れは例えば、西洋ワサビペルオキシダーゼを含む。固相に結合した酵素活性は、特異的な 酵素基質を添加し、そして生成物形成または基質利用性を比色的に測定することにより測 定される。固相基質に結合した酵素活性は、試料中に存在する抗原量の直接的関数である 。 本発明の別の特定のアッセイ方法において、生物学的試料中のHCVの存在(例えば、HCV感 染の指標として)は、従来のウェスタンブロッティングおよびドットブロッティング技術 (例えば、RIBA▲R▼(Chiron Corp.,Emeryville,CA)試験)を組合せた当該分野に公知 50 (28) JP 4270473 B2 2009.6.3 の技術のようなストリップ免疫ブロットアッセイ(SIA)技術を使用して検出され得る。 これらのアッセイにおいて、1つ以上の特異的な結合分子(組換えモノクローナル抗体( Fab分子を含む))は、膜状の支持体試験ストリップ上に個々の分離したバンドとして固 定化される。生物学的試料中に存在するHCV粒子との反応の可視化は、比色酵素基質と組 み合わせて標識された抗体結合体を用いるサンドイッチ結合技術を使用して達成される。 内部コントロールはまたストリップ上に存在し得る。このアッセイは、手動で行われ得る かまたは自動化された型式で使用され得る。 さらに、本発明に従って調製された組換えヒトモノクローナル抗体(Fab分子、Fvフラグ メント、およびsFv分子を含む)は、生物学的試料中のHCVの存在を検出するために、アフ ィニティークロマトグラフィー技術において使用され得る。このような方法は当該分野に 10 周知である。 任意の上記のアッセイおよびアフィニティークロマトグラフィー技術の実施における使用 に適切な、そして適切な標識された結合分子複合体試薬を含むキットはまた、本発明の実 施に従って提供され得る。アッセイキットは、アッセイ説明書の適切なセットとともに、 適切な容器中の、アッセイを行うために必要な全ての試薬および物質を含む、適切な材料 を包装することにより組み立てられる。 以下は、本発明を行うための特定の実施態様の実施例である。実施例は例証の目的のみの ために提供され、そしていかにしても本発明の範囲を限定するように意図されない。 使用される数値(例えば、量、温度など)の点で正確さを保証するために努力をされてい るが、いくらかの実験的誤差および偏差はもちろん許容されるべきである。 20 実施例 実施例1 ライブラリー供与者の特徴付け 骨髄を、献血に関連する通常のスクリーニングの間にHCVポジティブであることが見出さ れた60才の無症候性、男性血液供与者から得た。感染の原因は不明であった。供与者は免 疫化されておらず、そして約3mlの量の骨髄採取の以前に、HCV感染のための処置は受け ていなかった。骨髄提供の時点の供与者の血清中のHCVの遺伝子型を、刊行された方法を 使用して測定し、そしてHCV 2bであることを見出した。Widellら、(1994)J Med Viro1 44:272-279。 詳細には、供与者からのHCV単離物のE2遺伝子の超可変領域1(HVR1)のヌクレオチド配 30 列を、以下のように得た。1本鎖テンプレートDNAを得、そして製造者の説明書に従って 、標識されたヌクレオチドを用いるサイクル配列決定反応(PCRサイクル配列決定キット ,Perkin-Elmer)において、新たに設計したプライマー(A.Widellら、投稿準備中)を使 用して、配列決定反応を行った。Allanderら、(1994)J Med Virol 43:415-419。反応産 物を自動化シークエンサー(Applied Biosystems,CA)で泳動し、そしてデータをMacMoll yソフトウェア(Soft Gene,Berlin,Germanyより入手可能)を使用して編集および解析し た。供与者単離物のHVR-E2領域の推定アミノ酸配列は以下のように決定された:VAGVDAST YTTGGQSGRTTYGIVGLFSLGPSQKLSLINTNGSWHINR(配列番号29)。 実施例2 ファージ提示ライブラリーの構築 40 リンパ球を、Ficoll-Paque(Pharmacia Biotech,Uppsala,Sweden)を使用して実施例1に おいて得た骨髄試料から単離した。全RNAを酸性フェノール抽出方法(Chomczynskiら、( 1987)Anal Biochem 162:156-159)により抽出し、10μgのRNAのオリゴdTプライミングを 利用する、第1鎖cDNA合成を行い(cDNA合成キット、Pharmacia Biotech)、そして重鎖 (Fd)DNAおよび軽鎖DNAを、γ1重鎖およびκ軽鎖のための以前に公開された配列の5’ビ オチン化プライマー(5’プライマー:VH1a、VH1f、VH2f、VH3a、VH3f、VH4f、VH6a、VH6 f、Vk1a、Vk2a、Vk3a;3’プライマー:CG1zおよびCK1a) を用いてPCR増幅した。例えば、Perssonら、(1991)Proc Natl Acad Sci USA 88:2432-2 50 (29) JP 4270473 B2 2009.6.3 436 Kangら、(1991)Methods:Comp.Meth Enzymol 2:111-118を参照のこと。 PCRを、サーマルサイクラー4800(Perkin-Elmer)を使用して、94℃で5分、次いで94℃ で60秒、52℃で30秒、および72℃で180秒を35サイクル行った。35サイクルの後、72℃で1 0分の伸長工程によりPCR手順を完了した。PCR産物を1.5%アガロースゲルにおいて各々の 画分を泳動することにより分析し、重鎖DNAおよび軽鎖DNAを別々にプールし、フェノール -クロロホルム抽出し、次いでエタノール沈澱した。 重鎖DNAおよび軽鎖DNAの各々12μgを2.5%アガロースゲルでゲル精製し、電気溶出し(Sc hleicher&Schuell,Germany)、そしてそれぞれ制限エンドヌクレアーゼXhoI/SpeIおよび SacI/XbaI(Life Technologies,Gaithersburg,MDより入手可能)により消化した。消化し たPCR産物を、続いてゲル精製し、電気溶出により回収し、そして類似したベクター系に 10 ついて以前に記載したように(Yangら、(1995)J Mol Biol 254:392-403,Barbas,IIIら 、(1991)Proc Natl Acad Sci USA 88:7978-7982)対応する制限酵素により消化し、そ してゲル精製/電気的溶出した後、ベクターpComb3H(Barbas,IIIら、(1995)Methods:Co mp.Meth Enzymol 8:94-103)に連結した。連結には、T4リガーゼ(Life Technologies) を10μlの反応容量あたり0.5ユニットを、約16℃で一晩使用した。報告されるように、コ ンビナトリアルライブラリーをファージで発現させた(ファージの回収を含む)。Samuel ssonら、(1995)Virology 207:495-502。 pComb3Hへの軽鎖遺伝子の連結は、2×107メンバーのライブラリーを与えた。続く、この ライブラリーへのFd遺伝子の連結は、2×106メンバーのγ1/κライブラリーを生じた。 実施例3 20 HCV E2選択抗原の調製 アミノ酸383ala∼715lys(Chooら、(1991)Proc Natl Acad Sci USA 88:2451-2455の命 名法を使用)を含むHCV E2分子の短縮分泌形態を、構築した。以下のようにE2分子をチャ イニーズハムスター卵巣細胞/ジヒドロ葉酸レダクターゼ(CHO/DHFR)発現系を使用して 発現させ、「コンフォメーショナルHCV E2抗原」を提供した。HCV1のアミノ酸383∼アミ ノ酸715のHCV E2のDNAフラグメントをPCRにより生成し、次いでマウスサイトメガロウイ ルス(MCMV)前初期プロモーター/エンハンサー(Dorsch-Haslerら、(1985)Proc Natl Acad Sci USA 82:8325-8329)および選択可能なdhfr遺伝子マーカーを有するプラスミド ベクターに連結した。次いで、生じるプラスミドをdhfr-CHO細胞に安定にトランスフェク トし、コンフォメーショナルHCV E2抗原を分泌する安定な組換えCHO細胞株を生成した、 30 コンフォメーショナルE2抗原を、公知の方法(Rosaら、(1996)Proc Natl Acad Sci USA 93:1759-1763)を用いて以下のように精製した。CHO細胞からの馴化培地を限外濾過によ り15倍に濃縮し、続いて75%飽和の硫酸アンモニウム沈澱によりさらに10倍容量濃縮し、 そして25mM Trisクロライド/1mM EDTA、pH7.5に再溶解した。モノクローナル抗体5E5/H7 (HeLa E1/E2に対して惹起した)を精製のために使用した。抗体カラムを25mM Trisクロ ライド/0.15M NaCl、pH7.5で平衡化した。硫酸アンモニウム沈澱したE2を25mM Trisクロ ライド/1mM EDTA、pH7.5に溶解し、そしてカラムにロードした。カラムをリン酸緩衝化 生理食塩水(PBS)/1M NaClで洗浄し、次いでActisep(Sterogene,Arcadia,CA)の3∼ 4カラム容量で溶出した。黄色に着色したActisep含有画分を全てプールし、撹拌セル限 外濾過機において濃縮し、そしてPBS緩衝液中にダイアフィルターした。 40 組換えHCV E1/E2複合体抗原を、以下のように、構築し、そしてチャイニーズハムスター 卵巣細胞/ジヒドロ葉酸レダクターゼ(CHO/DHFR)発現系を使用して発現した。HCV1ゲノ ムのMet1∼Gly917にわたる917アミノ酸ポリペプチドとしてHCV構造領域をコードするプラ スミドpMCMV-HC5p(Spaeteら、(1992)Virology 188:819-830)を、pGEM-4 blue-HC5p-1 8由来の2813塩基対(bp)のHCV StuIフラグメントを哺乳動物細胞発現ベクターpMCVAdhf r(Spaeteら、(1990)J Virol 64:2922-2931)の唯一のSalI部位へクローニングするこ とにより生成した。pMCV Adhfrベクターは、MCMV前初期プロモーター/エンハンサーによ り駆動されそしてSV40ポリアデニル化配列により終結される発現遺伝子の転写を伴う、選 択可能なdhfr遺伝子をコードする。Klenowフラグメント(Boehringer-Mannheim Biochemi cals,Indianapolis,IN)を使用し、StuIフラグメントの連結の前にSalI部位を埋めた。pM 50 (30) JP 4270473 B2 2009.6.3 CMV-HC5pプラスミドをdhfr-CHO細胞に安定にトランスフェクトし、組換えHCV E1/E2複合 体抗原を発現する組換えCHO細胞株(#62)を生成した。 組換えE1/E2複合体抗原を公知の方法(Chooら、(1994)Proc Natl Acad Sci USA 91:129 4-1298)を使用して以下のように精製した。CHO細胞株#62をペレット化することにより 回収しそして凍結した。低浸透圧性緩衝液中でのDounceホモジナイズ化による溶解後、ペ レットを2%のTriton X-100▲R▼中でホモジナイズすることにより抽出し、そしてE1( 33kD)およびE2(72kD)を、アガロース結合Gralanthus Nivalis-レクチン(Vector Labo ratories)およびファーストフローS-Sepharoseカチオンイクスチェンジャー(Pharmacia )での連続的なクロマトグラフィーにより、選択的に同時精製した。 実施例4 10 コンビナトリアル・ライブラリーのパニング 特異的に結合するFab分子の抗原性選択を、公知の技術の変形を使用して行った。Burton ら(1991)Proc Natl Acad Sci USA 88:10134-10137。詳細には、マイクロタイタープレ ート(Costar 3690,Cambrige,MA)の4つのウェルを、CHO細胞で発現させた、精製した組 換えコンフォメーショナルHCV E2抗原または精製した組換えE1/E2複合体抗原(実施例3 において調製した)のいずれかの50μl(2.5μg/ml)を用いて、4℃にて一晩コートした 。Spaeteら(1992)Virology 188:819-830。ブロッキングを、ウェルをPBS中の5%脱脂 粉乳で22℃にて1時間完全に満たすことによって達成した。50μlのファージライブラリ ー(5×1010cfu)を、各ウェルに添加し、そしてプレートを37℃にて2時間インキュベー トした。ファージを除去し、そして各ウェルを、PBSと0.5% Tween 20の溶液で5分間完 20 全に満たすことによって洗浄し、次いで洗浄溶液を完全に除去した。洗浄を下記のように 1∼10回行った。1ウェル当たり50μlの溶出緩衝液(固体グリシンによってpH2.2に調整 した0.1M HCl)を添加し、そして室温にて10分間インキュベートすることによってファー ジを溶出した。溶出緩衝液を除去し、そして50μlの溶出緩衝液当たり3μlの2M Trisベ ースで中和した。E.coli XL-1 blue細胞(Barbas,IIIら(1991)Methods:Comp.Meth Enzy mol 2:l19-124)を、溶出させたファージによって感染させ、アリコートをプレーティン グし、そしてパニングの各ラウンド後のファージの増殖を、記載されているように達成し た。Samuelssonら(1995)Virology 207:495-502。 パニングの第1の群(パニングシリーズI)において、洗浄の数を3回のラウンドについ て各後続のパニングで増加させ(それぞれ1、3、および10回目の洗浄)、一方パニング 30 の第2の群(パニングシリーズII)においては、10回の洗浄を伴う単一のパニングを行っ た。表Iに示す結果によって理解され得るように、溶出されたファージで100倍の上昇が 、パニングシリーズIにおいて記録された。 40 シリーズIおよびIIの両方のパニングにおいて使用した抗原は、組換えコンフォメーショ ナルE2抗原であった。パニングの第3の群(パニングシリーズIII)を、シリーズIのよう 50 (31) JP 4270473 B2 2009.6.3 に行った;しかし、組換えE1/E2複合体抗原を使用して、陽性クローンを選択した。 実施例5 Fab分子の発現 Fab分子を、アンピシリン耐性E.coli XL-1 blue細胞コロニー((a)不溶性Fabフラグメ ントを提供するためにインタクトであるか、または(b)可溶性Fabフラグメントを提供 するためにSpeIおよびNheIでの消化(これらの酵素での消化は、適合性の粘着末端を提供 する;従って、生じるDNAフラグメントを欠失しているgIIIフラグメントはゲルで精製さ れ得、そして自己連結され得る)での消化によって欠失されたgIII遺伝子(cpIIIアンカ ータンパク質をコードする)を有するFabプラスミドを含有する)を、50μg/mlアンピシ リンおよび1%グルコースを含有するSB培地(スーパーブロス;1リットル当たり30gト 10 リプトン、20gのイーストエキストラクト、および10gのMOPS(pH7))(Burtonら(1991 )Proc Natl Acad Sci USA 88:10134-10137)中で、約1.0のOD600nmに達するまで増殖さ せることによって発現させた。細菌宿主細胞を遠心分離によってペレット化し、そして培 地を1mM IPTGおよび20mM MgCl2を有するSB培地に交換し、そして細胞を再懸濁した。生 じた培養物を、約290rpmに設定された振盪プラットホーム上で室温にてインキュベートし 、そして一晩放置した。次の日、細胞をスピンダウンし、上清を捨て、PBSを(もとの培 養物容量の2∼4%の間)で添加し、そして細菌細胞のペリプラズムの内容物を3サイク ルの凍結融解によって遊離させた。細菌細片を遠心分離によってペレット化し、そしてFa b分子含有上清を新たなバイアルに等分した。Fab分子を使用するまで-20℃で維持した。 実施例6 20 Fabクローンの発現レベル Fab分子(実施例5で得た)の発現レベルを、公知のELISA技術を使用して確認した。Samu elssonら(1995)Virology 207:495-502。詳細には、ヤギ抗ヒトF(ab’)2(Pierce,USA )またはヤギ抗ヒトFd(The Binding Site,UK)を0.1M炭酸-重炭酸緩衝液(pH9.6)で1:1 000に希釈し、そして4℃にて一晩のインキュベーションすることによってマイクロタイ ターウェルをコートした。コーティング溶液を捨て、そしてウェルを室温にて1時間PBS 中の5%の粉乳でブロックし、その後、ブロッキング溶液を除去し、そしてPBS-Tでの適 切な希釈でFab試料(実施例5から)を添加した。室温での1時間のインキュベーション 後、プレートを洗浄し、そして1:500の希釈でのALP-ヤギ抗ヒトF(ab’)2を添加した。 1時間のインキュベーション、および続く5回の洗浄後、標識基質溶液である0.1Mジエタ 30 ーノルアミン(pH9.8)中のp-ニトロフェニルホスフェート(Sigma,USA)を添加した。吸 光度を405nmにてマイクロプレートリーダー(Dynaster,MA)で測定した。ほとんどのFab クローンが、1Lの培養物当たり0.2∼2.0mgのFabを産生することが見出された。これは、 ペリプラスミック調製物中の10∼100μg/mlに対応する。発現させたFabクローンを、E2反 応性についてスクリーニングし、そして同一のもとのクローンの複数のコピーを同定する ためにすぐに配列決定した。 実施例7 重鎖および軽鎖発現についてのウェスタンブロット 両鎖の正確な発現を試験するために、いくつかのFab分子を、ヒトFdおよび軽鎖に対する 抗血清を使用するウェスタンブロットで分析した。詳細には、10μlのペリプラスミックF 40 ab分子調製物(実施例5で調整した)を、予め形成させた12%Tris-グリシンポリアクリ ルアミドゲルで分離し、そしてXcell Mini-cell装置(Novex Experimental Technology,S an Diego,CA)を使用するエレクトロブロッティングによってニトロセルロースメンブレ ンに移した。メンブレンを5%粉乳中で一晩ブロックし、そして5%粉乳および0.05%Tw een 20(PBS-MT)で1:1000に希釈したアルカリホスファターゼ結合抗ヒトFab抗血清(Pie rceから入手可能)とともに、一定の振盪で22℃にて3時間インキュベートした。被験体 抗ヒトFab抗血清を選択した。なぜならば、それが主に軽鎖に対して反応性であることが 公知であるからである。重鎖(Fd)発現産物を検出するために、ストリップを、まず、1: 1000に希釈されたヒツジ抗ヒトFd血清(Binding Site,U.K.)とともにインキュベートし (上記のように)、洗浄し、次いで1:500の希釈で第2次抗体、AP-抗ヤギIgG(Sigma,St. 50 (32) JP 4270473 B2 2009.6.3 Louis,MO)とともに再びインキュベートした。 22℃での1時間の最後のインキュベーションに続いて、メンブレンをPBS-T中で3回洗浄 し、そして発色を2mlのBCIP/NBT溶液(Sigam,St Louis,MO)を用いて7分間行った。メ ンブレンを水でリンスし、そして乾燥させた。予め染色した分子量マーカー(Amersham,U .K.)を各ブロットにおいて使用した。 試験した全てのクローンについて、両鎖の発現はほぼ等しかった。重鎖(短縮したgIIIタ ンパク質との融合ポリペプチドとして発現させた)は、70kDのおおよその分子量を示した 。 実施例8 Fabクローンの配列決定 10 E.coli XL-1 blue細胞培養物(実施例5)中で増殖させた各Fab分子クローンからのプラ スミドDNAを、WizardミニプレップDNA精製試薬システム(Promega)を使用して単離した 。一本鎖DNAをpComb3Hベクター(pC3H-2488S:5’-CAA CGC AAT TAA TGT GAG TTA G(配列 番号30);G-back:5’-GCC CCC TTA TTA GCG TTT GCC ATC(配列番号31))中のクローニ ング領域の上流および下流にハイブリダイズさせたプライマーを使用するPCRによって得 た。各反応において、2つのPCRプライマーのうち1つを5’末端でビオチン化した。PCR 増幅の35サイクルの後、一本鎖DNAを、アルカリ条件下でDNAを変性させ、そしてビオチン 化したDNA鎖をストレプトアビジンでコートしたビーズ(Dynal,Oslo,Norway)に公知の技 術を使用して吸着させることによって得た。Hultmanら(1989)Nucl Acid Res 17:4937-4 945。 20 Sangerら(1977)Proc Natl Acad Sci USA 74:5463-5467の方法に従ってジデオキシ配列 決定反応を、可変Ig領域と定常Ig領域との間の接合部の3’または重鎖遺伝子および軽鎖 遺伝子の開始点の5’にハイブリダイズするFITC標識したプライマーを利用して行った。 詳細には、SEQKb:5’-ATA GAA GTT GTT CAG CAG GCA(配列番号32)およびomp-seq:5’-A AG ACA GCT ATC GCG ATT GCA G(配列番号33)を、κ軽鎖について使用した。SEQGb:5’GTC GTT GAC CAG GCA GCC CAG(配列番号34)およびpel-seq:5’-ACC TAT TGC CTA CGG C AG CCG(配列番号35)を、γ重鎖について使用した。反応産物を自動化シークエンサー( A.L.F.,Pharmacia Biotech)で泳動して翻訳し、そしてMacMollyソフトウェア(Softgene ,Berlin,Germany)を使用して整列させた。 実施例4で行ったパニングの第1のシリーズ(パニングシリーズI)由来の、発現につい 30 てアッセイした(実施例6および7に記載のように)10個のFab分子クローン(Fab分子ク ローンL1-L10として同定した)を上記の配列決定法を使用して配列決定した。これらの10 のクローンは、それらの重鎖(H3領域)において極めて類似のCDR3配列を有することが見 出された。これは、それらが全て同一のB細胞クローン由来であったことを示す。Litwin ら(1990)J Exp Med 171:293-297。しかし、VDJ接合部およびH3領域の長さは、同一であ ったが、多くの異なる点変異がそれらの重鎖において同定され、そして各重鎖を異なる軽 鎖と組み合わせた。2つのクローン(Fab分子クロンL1およびL3として同定された)をさ らなる試験のために選択した。 実施例4で行ったパニングの第2のシリーズ(パニングシリーズII)由来の、発現につい てアッセイした(実施例6および7に記載のように)20個のFab分子クローン(Fab分子ク 40 ローン1:1∼1:20として同定した)を配列決定した。このラウンドの配列決定から、6つ のFabクローンが、不十分なレベルのFabを産生することが見出され、そして4つのFabク ローンが、特異的結合アッセイにおいてコントロール抗原に対して交叉反応性を示すこと が見出された。残りの10個のFabクローンから得られた配列決定情報から、7個のFabクロ ーンが、パニングシリーズI(L1-10Fab分子クローン)において見出されたクローンに関 連する重鎖を有することが見出された。しかし、7個のFabクローンの内3個は、パニン グシリーズIのクローンとは明らかに異なるH3領域を有し、そしてまた相互に独特に相関 していた。これらのクローンをさらに試験するために選択し、そして本明細書中でFab分 子クローン1:5、1:7、1:11として同定した。 実施例4において行ったパニングの第3のシリーズ(パニングシリーズIII)由来の、30 50 (33) JP 4270473 B2 2009.6.3 個のFab分子クローンを発現についてアッセイし(実施例6および7に記載のように)、 そして16個クローンが両E1/E2複合体抗原に対しておよびE2抗原のみに対して反応性であ ることが見出された。これらの16個のクローンを上記のように配列決定した。配列決定し た16個のクローンの内12個がクローンL1-L10に類似のH3配列を有することが見出されたが 、一方残りの4つは独特のH3配列を有することが見出された。独特のH3配列を有する4つ のクローンの内2つをさらに試験するために選択し、そして本明細書中でfab分子クロー ンA8およびA12として同定した。 以下のFab分子クローン:1:5(配列番号15);1:7(配列番号16);1:11(配列番号17) ;L3(配列番号18);L1(配列番号19);A8(配列番号20);およびA12(配列番号21) のκ軽鎖核酸配列を、各々、図3A∼図3Gに示す。 10 以下のFab分子クローン:1:5(配列番号22);1:7(配列番号23);1:11(配列番号24) ;L3(配列番号25);L1(配列番号26);A8(配列番号27);およびA12(配列番号28) のγ1重鎖核酸配列を、それぞれ、図4A∼図4Gに示す。 以下のFab分子クローン:1:5(配列番号1);1:7(配列番号2);1:11(配列番号3) ;L3(配列番号4);L1(配列番号5);A8(配列番号6);およびA12(配列番号7) の推定のγ1重鎖アミノ酸配列を、それぞれ、図1A∼図1Gに示す。各鎖由来のCDR領域(CD R1、CDR2、およびCDR3)もまた図中に同定した。 以下のFab分子クローン:(1:5(配列番号8);1:7(配列番号9);1:11(配列番号10 );L3(配列番号11);L1(配列番号12);A8(配列番号13);およびA12(配列番号14 )の推定のκ軽鎖アミノ酸酸配列を、それぞれ、図2A∼図2Gに示す。各鎖由来のCDR領域 20 (CDR1、CDR2、およびCDR3)もまた上記のように同定した。 要約すれば、3つのパニングシリーズから得た50クローンの内、36クローンは、E2に特異 的であることが見出され、そしてそれらの36のE2特異的クローンのうちの29は関連する重 鎖を共有する。 実施例9 HCV E2抗原反応性についてのELISAアッセイ Fab分子クローン1:5、1:7、1:11およびL3を、HCV E2抗原反応性について以下のようにス クリーニングした。組換えコンフォメーショナルHCV E2抗原、または組換えHCV E1/E2複 合体抗原(実施例3に記載のように調製した)のいずれかを、0.05M、pH9.6の炭酸−重炭 酸緩衝液中の0.25μg/mlに希釈し、4℃にて1晩、マイクロタイターウェル(Costar #36 30 90;Life Technologies)にコーティングした。結合していない抗原を捨て、そしてウェル をPBS中の5%脱脂乾燥ミルクで60分間室温にてブロッキングした。ブロッキング溶液を 捨てた後、試験されるFab分子を含む溶液を1:2、1:10、および1:100希釈(希釈物:0.1% NP-40を有するPBS)で添加した。プレートを室温で2時間インキュベートし、0.05% Twe en 20を有するPBS(PBS-T)で5回洗浄し、そしてALP-ヤギ抗ヒトF(ab’)2(Pierce、R ocherford,IL)を1:1000希釈で添加した。60分後、そして続く洗浄後に、基質溶液(p-ニ トロフェニルホスフェート)(SIGMA,St.Louis,MO)を添加し、そして吸光度を405nmにて マイクロプレートリーダー(Dynastar,MA)において測定した。 ポジティブな読み取りについてのカットオフ値は、ネガティブコントロール試料について 得られるOD値の4倍に設定した。この試料は、等濃度の抗HIV Fabを含んだ。Barbas III ら(1991)Proc Natl Acad Sci USA 88:7978-7982。コントロールの目的のために、ウシ 血清アルブミン(BSA)(SIGMA)、HIV gp120LAI(Intracell,Cambridge,MA)、および破 傷風トキソイド(TT)(SBLワクチン、Solna,Sweden)でそれぞれ5、1、および1μg/m lで、コートしたものを、対応するELISAにおいて、非特異的な反応性についてのコントロ ールとして使用した。 ELISAからの結果を、以下の表IIに示す。理解され得るように、1:5、1:7、1:11、およびL 3クローンから発現されるFab分子は、各々、コンフォメーショナルHCV E2抗原およびHCV E1/E2複合体抗原の両方と強力に反応する一方で、コントロール抗原(BSA、HIVgp120、お よびTT)とは交叉反応を示さなかった。 40 (34) JP 4270473 B2 2009.6.3 10 実施例10 HCV E2抗原反応性についてのウェスタンブロットアッセイ 20 Fab分子クローン1:5、1:7、1:11、L1、L3、A8、およびA12をHCV E2抗原反応性について、 以下の技術を用いてスクリーニングした。ウェスタンブロットを、1μgの組換えコンフ ォメーショナルHCV E2糖タンパク質(実施例3において得た)を使用して実施した。この タンパク質は、Laemmli緩衝液中で98℃にて5分間加熱することにより変性させ、8∼16 %ポリアクリルアミド勾配ゲル(Novex Experimental Technologies)上で分離し、そし て上記のようにブロッキングしたニトロセルロースメンブレンに移し、そして小片に切断 した。続いて、各々の小片を、PBS-MT中で1:20に希釈したFab調製物(1:5、1:7、1:11、L 3、L1、A8、およびA12クローンから発現した)とともに22℃にて2時間、一定に振盪しな がらインキュベートした。この小片をPBS-T中で3回洗浄し、そしてPBS-MT中で1:1000に 希釈したアルカリホスファターゼ結合ヤギ抗ヒトFab血清(Pierce)を添加した。22℃で 30 の1時間のインキュベーションに続いて、小片を再び0.05% Tween 20を有するPBS中で3 回洗浄し、そして発色を、2mlのBCIP/NBT溶液(Sigma、St Louis,MO)で10分間で行った 。ポジティブコントロールとして、Fab調製物の代わりにヒト抗HCVポジティブ血清を、1 つの小片とともにインキュベートした。 試験したFab分子(クローン1:5、1:7、1:11、L3、L1、A8、およびA12由来)のいずれもウ ェスタンブロットにおいて変性HCV E2抗原とは反応せず、このことは、各々のFab分子が 、HCV E2抗原のコンフォメーショナルエピトープに結合することを示している。しかし、 ポジティブコントロール(ヒト抗HCVポジティブ血清)は、ウェスタンブロットにおいて 変性E2抗原と反応した。 上記のアッセイを、非変性条件下においてゲル分離したHCV E2抗原の使用という単一の変 40 化以外は同一条件下で繰り返した。このさらなるアッセイにおいて試験した両方のクロー ン(1:7およびA8)は非変性E2抗原と結合することが見い出された。 実施例11 親和性決定についての阻害ELISAアッセイ HCV E2抗原へのFab分子(クローン1:5、1:7、1:11、L3、L1、A8、およびA12由来)の親和 性を、阻害ELISA法を以前に記載したように用いて評価した。Perssonら(1991)Proc Nat l Acad Sci USA 88:2432-2436、Rathら(1988)J Immun Methods 106:245-249。試験され る試料を、まず濃度を階層化するために10倍希釈で力価測定した。ここで、濃度が10倍減 少すると、HCV E2 ELISAにおいて検出された結合において実質的な減少が生じた。親和性 測定のために、マイクロタイターウェルのHCV E2抗原(HCV遺伝子型1a)でのコーティン 50 (35) JP 4270473 B2 2009.6.3 グおよび続くブロッキングを、実施例9において実施したELISAについて上記のように行 った。Fab試料の適切な希釈物を、可溶性HCV E2抗原(最終濃度5μg/ml)を添加するか または添加しないで、ウェルに添加し、そして室温で3時間インキュベートした。プレー トを、PBS-Tで4回均一に洗浄し、そして実施例9において上記のようにAP-抗Fab、基質 、および吸光光度計リーダーを用いて発色した。可溶性HCV E2抗原の存在下でのODの減少 を計算し、そして外挿法により50%減少に必要な濃度を見積もった。 以下の表IIIで示すように、組換えコンフォメーショナルHCV E2抗原(HCV遺伝子型1a)に 対するFab分子(クローン1:5、1:7、1:11、L3、L1、A8、およびA12由来)のおおよその親 和性は、1×107と2×108M-1の間で変化した。 10 20 異なるHCV E2抗原(HCV遺伝子型1b)についてのFab分子(クローン1:7、A8、およびA12由 来)の親和性もまた、上記の阻害ELISAを用いて見積もった。試験した分子の各々におけ る遺伝子型1bのE2抗原についての親和性は、遺伝子型1a E2抗原について上記で報告した 30 もの(表III)と同様であることが見い出された。 さらに、以下のFab分子クローン(L1;L3;1:5;1:7;および1:11)から調製した全組換 えIgG分子を、HCV遺伝子型1a E2抗原を用いる上記の阻害ELISAを使用して見積もった。観 察された親和性は、Fab分子クローンについて上記で報告したもの(表III)と同様であっ た。 実施例12 HCV E2結合の阻害 Fab分子(クローン1:5、1:7、1:11、L3、L1、A8、およびA12由来)が標的細胞に対するHC V E2の結合をブロッキングする能力を、Rosaら(1996)Proc Natl Acad Sci USA 93:1759 -1763の結合の中和(NOB)方法を用いて決定した。より詳細には、精製コンフォメーショ 40 ナルHCV E2抗原(遺伝子型HCV 1aおよびHCV 1bの両方由来、そして実施例3に記載のよう に調製した)を間接的免疫蛍光実験に用いて、2つの別々のバッチの細菌で発現したFab 分子クローンが、インビトロでのヒト細胞に対するHCV E2ポリペプチドの結合を中和する 能力を評価した。 アッセイにおいて、20μlの精製したコンフォメーショナルHCV E2抗原(PBS中にて0.5μg /ml)を種々の希釈のFabクローンと混合した。4℃での1時間のインキュベーション後、 MOLT-4細胞(Shimizuら(1992)Proc Natl Acad Sci USA 89:5477-5481に記載のようにイ ンビトロでの低レベルのHCV複製を可能にすると報告されているヒト細胞株)のペレット を添加し、そして生じる反応混合物を4℃にて1時間インキュベートした。結合していな いHCV抗原および抗体を、PBS中4℃での5分間の200×gでの2回の遠心分離により除去し 50 (36) JP 4270473 B2 2009.6.3 た。次いで、細胞を、ヒト抗HCV E2反応性血清と4℃にて30分間インキュベートした。次 いで、細胞をPBS中で二回洗浄し、そしてフルオレセインイソチオシアネート結合ヒトFab 特異的抗血清とともに30分間インキュベートした。細胞を、PBS中で4℃にて再び洗浄し 、そして100μlのPBSに再懸濁した。細胞に結合した蛍光を、Lysis IIソフトウェア(Bec ton Dickinson)を用いてフローサイトメーター(FACScan、Becton Dickinson)で分析し た。種々のFab調製物とともにインキュベートした細胞集団の平均蛍光強度を計算し、そ して抗体なしか、またはE2抗原なしでインキュベートした細胞の平均蛍光強度と比較した 。 結果を以下の表IVに示す。理解され得るように、7つの試験したFabクローンすべてが、 コンフォメーショナルHCV E2抗原(遺伝子型HCV 1aおよびHCV 1bの両方)によりMOLT-4細 10 胞結合を効率的に阻害した。クローンA8、1:7、L1、およびL3は、アッセイにおいて非常 に高い中和活性を有していた。50%減少力価を、試験したクローンすべてについて表IVに 示し、そしてクローン中の4つについてのアッセイの完全な結果を図5に示す。HIV-1エ ンベロープ糖タンパク質gp120(クローンb12およびb14)に指向させた以外は上記と同様 の様式で調製した2つのネガティブコントロールのFabクローンは、アッセイにおいて中 和活性を有さなかった。真核生物細胞において発現したFabクローン、およびFabクローン から由来する組換え体の完全なIgG分子は、同様なNOBアッセイにおいてネガティブである ことが見い出された。 20 30 40 HCVウイルスとその宿主との第一の接触が、ウイルスエンベロープの細胞表面レセプター に対する結合を介して生じるため、本発明のFab分子がこの相互作用を中和する能力は、H CVに対する受動免疫を提供するためのワクチン接種においてそれらの分子を使用する有効 性を確立する。 従って、HCV E2抗原に対する新規なヒトモノクローナル抗体が開示される。本発明の好ま しい実施態様を、ある程度詳細に記載しているが、添付の請求の範囲により定義される本 発明の精神および範囲から逸脱することなく明らかな改変が行われ得ることが理解される 。 50 (37) 【図1A】 【図1B】 【図1C】 【図1D】 JP 4270473 B2 2009.6.3 (38) 【図1E】 【図1F】 【図1G】 【図2A】 JP 4270473 B2 2009.6.3 (39) 【図2B】 【図2C】 【図2D】 【図2E】 JP 4270473 B2 2009.6.3 (40) 【図2F】 【図2G】 【図3A】 【図3B】 JP 4270473 B2 2009.6.3 (41) 【図3C】 【図3D】 【図3E】 【図3F】 JP 4270473 B2 2009.6.3 (42) 【図3G】 【図4A】 【図4B】 【図4C】 JP 4270473 B2 2009.6.3 (43) 【図4D】 【図4E】 【図4F】 【図4G】 JP 4270473 B2 2009.6.3 (44) 【図5】 JP 4270473 B2 2009.6.3 (45) JP 4270473 B2 2009.6.3 フロントページの続き (51)Int.Cl. FI A61K 39/395 (2006.01) A61K 39/395 S C07K 16/10 (2006.01) C07K 16/10 C12N 1/15 (2006.01) C12N 1/15 C12N 1/19 (2006.01) C12N 1/19 C12N 1/21 (2006.01) C12N 1/21 A61K 38/21 (2006.01) A61K 37/66 G G01N 33/53 (2006.01) G01N 33/53 10 (72)発明者 アランデル,トビアス エリック スウェーデン国 エス―171 76 ストックホルム,(番地なし),デパートメント オブ メディシン エム3:00,カロリンスカ ホスピタル内 審査官 森井 隆信 (56)参考文献 国際公開第93/002190(WO,A1) Princess Takamatsu Symp., 米国, 1995年, Vol.25, 129-137 Virology, 米国, 1994年, Vol.205, 141-150 20 J. Virol., 米国, 1994年, Vol.68, No.10, 6147-6160 Proc. Natl. Acad. Sci. USA., 米国, 1991年, Vol.88, No.18, 7978-7982 Proc. Natl. Acad. Sci. USA., 米国, 1993年, Vol.90, No.9, 4141-4145 Immunotechnology, 1995年, Vol.1, No.1, 21-28 (58)調査した分野(Int.Cl.,DB名) C12N 15/00 A61K 39/395 A61K 38/21 A61P 31/18 A61P 37/04 C07K 16/10 C12N 1/00 C12N 5/00 G01N 33/53 BIOSIS/MEDLINE/WPIDS(STN) GenBank/EMBL/DDBJ/GeneSeq UniProt/GeneSeq PubMed 30
© Copyright 2026 Paperzz




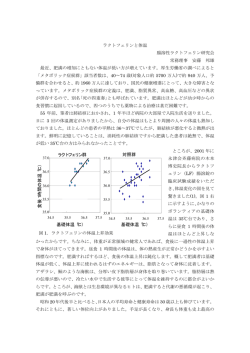


![[別紙様式1]見本 機械類輸入承認申請明細書 受付年月日 申請者名](http://s3.paperzz.com/store/data/005619819_1-da8b1199cabee6bf7e4ad850ce040e83-250x500.png)