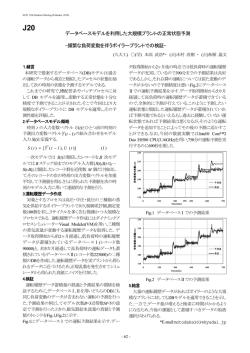
道衛研所報 Rep. Hokkaido Inst. Pub. Health, 57, 109-112(2007) 記憶喪失性貝毒成 ドウモイ酸によるマウスの自発的 替行動障害に 及ぼすニコチン性アセチルコリン受容体アゴニストの影響 Effects of Nicotinic Acetylcholine Receptor Agonists on Impairment of Spontaneous Alternation Behavior Induced by Domoic Acid, an Amnesic Shellfish Poison, in Mice 上野 一 Ken-ichi UENO ;domoic acid(ド ウ モ イ 酸);nicotinic Key words:amnesic shellfish poison(記憶喪失性貝毒) ;memory(記 憶) ;mouse acetylcholine receptor(ニコ チ ン 性 ア セ チ ル コ リ ン 受 容 体) (マウス) 1987年,カナダ東海岸のプリンス・エドワード島周辺 ことが知られている で養殖ムラサキイガイの摂食により胃腸障害及び神経障害 を主徴とする食中毒が発生した . ドウモイ酸はラットやマウスなどに記憶障害や海馬神経 .この食中毒により, 細胞死を引き起こすことが報告されている .これま 患者数 107名のうち4名が死亡し,12名に記憶障害の後 でに,我々はマウスのドウモイ酸誘発性自発的 替行動障 遺症が残ったことから,この中毒は記憶喪失性貝毒中毒と 害がカイニン酸型グルタミン酸受容体アンタゴニストや 呼ばれた.記憶喪失性貝毒中毒の原因物質として興奮性ア ミノ酸の一種であるドウモイ酸が同定され,ドウモイ酸は group 及び group 代謝調節型グルタミン酸受容体ア ゴニストにより予防されることや,ニコチンがドウモイ酸 記憶喪失性貝毒とも呼ばれるようになった. 誘発性自発的 ドウモイ酸は,その化学構造中に中枢神経の主要な神経 た 替行動障害を改善することを報告してき .しかしながら,ドウモイ酸誘発性自発的 替行 伝達物質であるグルタミン酸やかつて駆虫薬として利用さ 動障害に及ぼすサブタイプ選択的な nAChR アゴニストの れたカイニン酸と同様の骨格を有する.ドウモイ酸はカイ 作用については検討されていない. ニン酸よりも強力なカイニン酸型グルタミン酸受容体アゴ そこで,本研究はマウスのドウモイ酸誘発性自発的 替 ニストであり,カイニン酸と同様,海馬神経細胞の CA3 行動障害に及ぼすサブタイプである α4β2及び α7選択 領域を選択的に破壊することにより,ドウモイ酸中毒の特 的な nAChR アゴニストの作用を明らかにすることを目的 徴的な症状である記憶障害を引き起こすと とした. る えられてい . 方 グルタミン酸,N -methyl-D-aspartate 及びカイニン酸 などのグルタミン酸受容体アゴニストにより誘発される神 法 1.実験動物 経毒性や行動毒性は,ニコチン性アセチルコリン受容体 実 験 に は,6∼7 週 齢 の ddY 系 雄 性 マ ウ ス(日 本 (nAChR)アゴニストのニコチンにより抑制されることか ら,ニコチンによる神経保護作用が指摘されている . SLC)を 用した.動物は,入荷後約1週間,室温 22± 2℃の明暗サイクル(明期,6:00∼18:00)環境下で飼 また,ニコチン性アセチルコリン神経系は,注意力や記 育し, 及び水は自由に摂取させた. 憶・学習などの認知機能に密接に関連している 2.実験手順 .中枢 神経系の nAChR には,それぞれ5量体からなる α タイ プと αβ タイプがあり,さらに α タイプには α2,α5, 自発的 替行動の測定にはY字迷路(アーム長 40cm, 壁高 12cm,上部幅 10cm,床幅3cm)を 用した. α7,α8及び α9のサブタイプが,αβ タイプには α3 β4,α4β2及び α6β3のサブタイプがあることが知ら 自発的 替行動の測定は,Sarter et al.(1988)の方法 に準じて行った .マウスをY字迷路内に置き,8 間に れている .なかでも α4β2nAChR と α7nAChR は注 わたって迷路内を自由探索させ,マウスが選択したアーム 意力や作業記憶など認知機能に重要な役割を果たしている を順に記録した.マウスが測定時間内に各アームを選択し 109 た 回 数 を 記 録 し,こ れ を アーム 選 択 数(total arm )とした.次に,この中から連続して異なる3本 entries のアームを選択した組み合わせを調べ,この数を 替行動 3. 数とした. 替行動数を アーム選択数から2を引いた数 トの影響を検討した. で割り,その値に 100を掛けて求めた数値を 発性自発的 替行動障害に及ぼす選択的 nAChR アゴニス 替行動率 (alternation behavior(%) )とし,これを自発的 動の指標とした. 察 Y字迷路を用いてマウスの記憶喪失性貝毒ドウモイ酸誘 ニコチン性アセチルコリン神経系は,注意力や記憶・学 替行 習などの認知機能に密接に関連している .また,中枢 神 経 系 に お け る 主 要 な nAChR は α4β2nAChR と α7 3. 用薬物と投与方法 nAChR のサブタイプに大別されている. nAChR アゴニストのニコチンは,α4β2nAChR を介 ド ウ モ イ 酸(Domoic acid, Sigma 社 製) ,ABT-418 (3-methyl-5[(2S)-1-methyl-2-pyrrolidinyl]isoxazole して注意機能障害を改善することが報告されている . hydrochloride, Sigma-RBI 社 製), RJR-2403( N methyl-4-(3-pyridinyl)-3-butene-1-amine hemigalac- また,ニコチンは α7nAChR を介して学習・記憶障害を tarate, Sigma 社 製), anabasine((±)-anabasine, Sigma 社製)を 用した.これらの薬物はすべて生理食 られている海馬長期増強現象を増強すること 塩水に溶解した.ドウモイ酸は,自発的 改善すること や学習・記憶の電気生理学的基盤と え が報告さ れている. 替行動測定の 24時間前にマウスへ単回腹腔内投与した.ABT-418, RJR-2403,anabasine は,自発的 替行動測定の 20 前 に単回皮下投与した.対照群には 10mL/kg の 0.9%生理 食塩水を投与した. 4.統計学的処理 各実験において 用した動物数は1群あたり 15匹とし た. 替行動率及び アーム選択数は平 値±標準誤差で 表した.二群間の比較には t 検定を用い,三群以上の比較 には,一元配置 散 析を行い,有意差が認められたもの については,Dunnett の多重比較検定を行った.なお,こ れらの統計処理では,原則として危険率を5%とした. 結果及び 察 1.マウスのドウモイ酸誘発性自発的 替行動障害に及ぼ す選択的 α4β2nAChR アゴニストの影響 ドウモイ酸誘発性自発的 替行動障害に及ぼす選択的 α4β2nAChR アゴニストの影響を Fig.1 及び2に示す. ドウモイ酸(3mg/kg)は対照群と比較しマウスの 替行 動率を有意に低下させた(Fig.1A) .これに対し,マウス の アーム選択数はドウモイ酸による影響は認められな かった(Fig.1B).ドウモイ酸による 替行動率の低下は 選 択 的 α4β2nAChR ア ゴ ニ ス ト の ABT-418(0.01及 び 0.1mg/kg)及 び RJR-2403(3及 び 10mg/kg)に よ り用量依存的かつ有意に改善された(Fig.1A 及び 2A). また, アーム選択数は選択的 α4β2nAChR アゴニス トによる影響は認められなかった(Fig.1B 及び 2B) . 2.マウスのドウモイ酸誘発性自発的 替行動障害に及ぼ す選択的 α7nAChR アゴニストの影響 ドウモイ酸誘発性自発的 替行動障害に及ぼす選択的 Fig.1 Effects of ABT-418 on Domoic Acid-induced Impairment of Spontaneous Alternation (A) and Total Arm Entries (B) in Mice α7nAChR アゴニストの影響を Fig.3 に示す.ドウモイ 酸による 替行動率の低下は選択的 α7nAChR アゴニス Domoic acid (3 mg/kg,i.p.)was administered 24 hours before the test. ABT-418 (0.001-0.1 mg/kg, s.c.) was administered 20 min before the test.Data are expressed as the mean±S.E.M.(vertical bars).The number of mice used are shown in the columns. * p< 0.001 vs. saline treated-group;♯♯ p<0.01;♯♯♯ p<0.001 vs. domoic acid alone. ト anabasine(0.3∼3mg/kg)で は 改 善 さ れ な かった (Fig.3A) .また, アーム選択数は anabasine による影 響は認められなかった(Fig.3B) . 110 そこで,ドウモイ酸誘発性自発的 替行動障害に及ぼす セチルコリン神経の神経終末に存在することが知られてい サブタイプ選択的な nAChR アゴニストの影響を検討した. る .ドウモイ酸により海馬グルタミン酸神経細胞が ドウモイ酸による 替行動率の低下は,anabasine では 改善されず,ABT-418 及び RJR-2403 により用量依存的 破壊されることから,海馬グルタミン酸神経の神経終末に かつ有意に改善されたことから,ドウモイ酸誘発性自発的 的 α7nAChR ア ゴ ニ ス ト の anabasine が 効 果 を 示 さ な 替行動障害には選択的 α4β2nAChR アゴニストが有 効であることが示唆された. かったものと推察される.この仮説は,ドウモイ酸誘発性 本研究において,選択的 α4β2nAChR アゴニストの ABT-418 及び RJR-2403 がドウモイ酸誘発性自発的 替 行 動 障 害 を 改 善 し,選 択 的 α7nAChR ア ゴ ニ ス ト の nAChR ではなく α4β2nAChR を介する と い う 報 告 からも支持される. anabasine が改善しなかったという結果から,次のような 仮説が えられる. 行動障害に及ぼすサブタイプ選択的な nAChR アゴニスト 海馬において α7nAChR はグルタミン酸神経の神経終 障害は選択的 α4β2nAChR アゴニストにより改善され 末に,α4β2nAChR は中隔から海馬へ投射しているア ることから,α4β2nAChR アゴニストにドウモイ酸中 存在する α7nAChR の機能が障害されたことにより選択 自発的 替行動障害に対するニコチンの改善作用が α7 以上より,マウスにおけるドウモイ酸誘発性自発的 替 の影響を明らかにした.ドウモイ酸誘発性自発的 替行動 Fig.2 Effects of RJR-2403 on Domoic Acid-induced Impairment of Spontaneous Alternation (A) and Total Arm Entries (B) in Mice Fig.3 Effects of Anabasine on Domoic Acid-induced Impairment of Spontaneous Alternation (A) and Total Arm Entries (B) in Mice Domoic acid (3 mg/kg,i.p.)was administered 24 hours before the test. RJR-2403 (1-10 mg/kg, s.c.)was administered 20 min before the test. Data are expressed as the mean±S.E.M . (vertical bars). The number of mice used are shown in the columns. * p<0.001 vs. saline treated-group;♯ p<0.05;♯♯♯ p<0.001 vs. domoic acid alone. Domoic acid (3 mg/kg,i.p.)was administered 24 hours before the test.Anabasine(0.3-3 mg/kg,s.c.)was administered 20 min before the test. Data are expressed as the mean±S.E.M . (vertical bars). The number of mice used are shown in the columns. * p<0.001 vs. saline treated-group. 111 毒による記憶障害に対する治療薬としての可能性が示唆さ れた. 文 献 1) Perl TM ,Bedard L,Kosatsky T,Hockin JC,Todd ECD, Remis RS:N. Engl. J. Med., 322, 1775-1780 (1990) 2) Teitelbaum JS, Zatorre RJ, Carpenter S, Gendron D, Evans AC, Gjedde A, Cashman NR:N. Engl. J. Med., 322, 1781-1787 (1990) 3) 篠崎温彦:Clinical Neuroscience, 16, 1006-1009 (1998) 4) 篠崎温彦:日薬理誌,116,125-131(2000) 5) Akaike A,Tamura Y,Yokota T,Shimohama S,Kimura J:Brain Res., 644, 181-187 (1994) 6) Borlongan CV, Shytle RD, Ross SD, Shimizu T, Freeman TB, Cahill DW, Sanberg PR:Exp. Neurol., 136, 261-265 (1995) 7) O Neill AB, Morgan SJ, Brioni JD:Neurobiol. Learn. M em., 69, 46-64 (1998) 8) Kim HC, Jhoo WK, Ko KH, Kim WK, Bing G, Kwon M S, Shin EJ, Suh JH, Lee YG, Lee DW :Life Sci., 66, 317-326 (2000) 9) Levin ED, Simon BB:Psychopharmacology, 138, 217230 (1998) 10) Rezvani AH, Levin ED:Biol. Psychiatry, 49, 258-267 (2001) 11) Cordero-Erausquin M,M arubio LM,Klink R,Changeux J-P:Trends Pharmacol. Sci., 21, 211-217 (2000) 12) Felix R, Levin ED:Neuroscience, 81, 1009 -1017 (1997) 13) Levin ED, Bettegowda C, Blosser J, Gordon J:Behav. 112 Pharmacol., 10, 675-680 (1999) 14) Bancroft A, Levin ED:Neuropharmacology, 39, 27702778 (2000) 15) Blondel A, Sanger DJ, M oser PC:Psychopharmacology, 149, 293-305 (2000) 16) Grottick AJ, Higgins GA:Behav. Brain Res., 117, 197208 (2000) 17) Ueno K, Togashi H, M atsumoto M , Ohashi S, Saito H, Yoshioka M :J. Pharmacol. Exp. Ther., 302, 95-100 (2002) 18) Sutherland RJ, Hoesing JM, Whishaw IQ:Neurosci. Lett., 120, 221-223 (1990) 19) Kuhlmann AC, Guilarte TR:Brain Res., 751, 281-288 (1997) 20) Clayton EC, Peng Y-G,M eans LW,Ramsdell JS:Toxicon, 37, 1025-1039 (1999) 21) 上野 一,田沢悌二郎,石下真通,堀 義宏:道衛研所報, 51,87-90(2001) 22) 上野 一:道衛研所報,54,69-72(2004) 23) 上野 一:道衛研所報,56,53-56(2006) 24) Sarter M , Bodewitz G, Stephens DN:Psychopharmacology, 94, 491-495 (1988) 25) Hunter BE, de Fiebre CM , Papke RL,Kem WR,M eyer EM :Neurosci. Lett., 168, 130-134 (1994) 26) Gray R, Rajan AS, Radcliffe KA, Yakehiro M , Dani JA:Nature, 383, 713-716 (1996) 27)Wonnacott S:Trends Neurosci., 20, 92-98 (1997) 28) Buccafusco JJ, Letchworth SR, Bencherif M, Lippiello PM :Trends Pharmacol. Sci., 26, 352-360 (2005)
© Copyright 2026 Paperzz