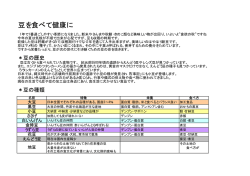
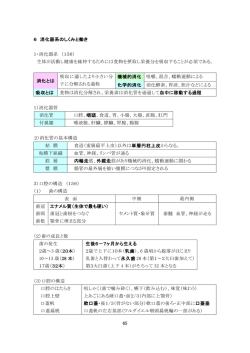
糖類[saccharide] 基本単位でそれ以上加水分解されない単糖類、数個の単糖が結合した少糖類、多数の単糖 が結合した多糖類の総称。また主要な炭水化物はすべて含まれるところから、実際には炭 水化物とほとんど同義に用いられる。なお、動物ではタンパク質、脂質とともに三大栄養 素の一つであり、ヒトでは摂取カロリーの半分以上を糖類に依存する。 少糖[ショウトウ oligosaccharide] オリゴ糖、寡糖ともいい、単糖が 2 個から 10 個程度まで結合した大きさの糖の総称。構 成単糖の分子数により二糖、三糖、四糖などに分かれる。 天然に遊離の形で存在するほか、多糖を化学的または酵素的に加水分解して得られる。溶 解性などの化学的性質は一般に単糖に類似し、多糖とは異なる。 同一単糖よりなるホモ少糖と、2 種以上の単糖よりなるヘテロ少糖に大別できる。天然 のホモ少糖にはトレハロース、マルトース、ヘテロ少糖にはショ糖(サッカロース)、乳 糖(ラクトース)、ラフィノースなどがある。多糖の加水分解では、種々の大きさと組成 のものが得られる。たとえば、アミロースの酸限定分解で二糖から七糖までの少糖が得ら れ、ヒアルロン酸の酵素分解では二糖のグルクロン酸‐アセチルグルコサミンが得られる。 結合様式から遊離還元基の有無で還元性少糖と非還元性少糖に分けることもあり、天然の 少糖には非還元性のものが多い。糖タンパク質や糖脂質に含まれる糖鎖部分は少糖である ことが多く、この場合、グルコースやガラクトースのほか、マンノース、フコース、キシ ロース、ノイラミン酸など各種の単糖が含まれる。 多糖[polysaccharide ] 多数の単糖がグリコシド結合で重合した高分子化合物の総称で、グリカン glycan ともい う。構成単糖は少なくとも 10 個程度以上で、通常、重合度は数十から数十万に達する。 直鎖状または枝分れした構造をもち、大きさはさまざまで、種類によって異なるとともに、 同じ種数でも大きさの異なった分子の混合物であることが多い。 タンパク質、脂質と並んで生体を構成する高分子の一つで、自然界の有機物質としてもっ とも多量に存在する。その機能は生物体の構造支持、皮膜物質、潤滑剤としての役割と、 生体エネルギーの貯蔵物質としての役割が主である。 貯蔵物質の例では、動物のグリコーゲンと植物のデンプン(アミロース、アミロペクチン) が代表的で、構造物質としては植物のセルロース、昆虫や甲殻類のキチン、動物の結合組 織に含まれるムコ多糖などがある。また細胞膜や細胞壁の多糖成分のように、細菌性抗原 多糖や血液型物質として細胞表面の特異性を担うものもある。 多糖は広く生物界に存在し、その種類も多様であるために統一的な分類はむずかしいが、 化学構造に基づいた分類のほかに、細菌、藻類、高等植物、脊椎(せきつい)動物といった 起源材料による分類や生物的機能に基づく分類が、実用上使われている。名称は構成単糖 の語幹にアン(-an)をつけたものが多いが、従来の慣用名も多く用いられている。 ホモ多糖は 1 種類の単糖よりなるもので、ペントサンは五炭糖多糖、ヘキソサンは六炭 糖多糖である。グルコースよりなるグルカンの場合、結合様式がαかβか、1・4結合か 1・6結合かなどで多数のものがある。 ヘテロ多糖は 2 種類以上の単糖から構成される。ムコ多糖はアミノ糖を含む多糖の一群で、 二糖の繰り返し構造をもつ。カルボニル基、硫酸基をもつ酸性のものが多い。複合多糖は 糖以外の脂質やタンパク質に糖鎖が共有結合したものである。 この場合、糖鎖部分は比較的短い。リポ多糖は大腸菌などの細胞壁成分で、リピドAとよ -1- ばれる脂質に結合した糖鎖が抗原特異性を担う。糖タンパク質は、糖鎖末端とタンパク質 中のアミノ酸との結合型からおもに 2 種に大別される。 多糖は分解酵素グリコシダーゼによって、生体内で加水分解される。構成糖の種類や結 合様式に特異性をもつ多数の酵素が知られる。肝臓の貯蔵多糖グリコーゲンは、ホスホリ ラーゼで加リン酸分解を受けて解糖系に入り、エネルギー源となる。多糖の性質は種類に よって異なるが、全体としては水酸基による親水性があり、塩溶液や緩衝液などの水性溶 媒に可溶性または分散されるが、不溶性のものもある。有機溶媒には溶けない。甘味はな く、フェーリング溶液を還元しない。希酸による加水分解で少糖を経て単糖を生じる。生 体組織中の多糖は、過ヨウ素酸シッフ反応で細胞化学的に検出される。 単糖 糖類のうちそれ以上加水分解できない糖の総称で、単糖類ともいう。おもなものに、グル コース、フルクトース、マンノース、ガラクトース、リボースなどがある。 一般に甘味をもつ無色結晶で、水に溶けるが、エタノール(エチルアルコール)には溶け にくく、エーテルには溶けない。 遊離形または少糖類、多糖類、配糖体などの構成糖として生物界に広く存在するが、遊離 形での存在は少ない。 天然物から抽出するか、少糖類、多糖類などを加水分解して得るが、化学的に合成するこ ともできる。最低 2 個の水酸基とこれに隣接するカルボニル基-CO-をもつ。 その多くは Cn(H2O)m という一般式で表すことができ、C(炭素)の数によってトリオー ス(三炭糖)、ペントース(五炭糖)、ヘキソース(六炭糖)などとよばれる。 またカルボニル炭素がアルデヒド基にあるかケトン基にあるかによって、アルドースまた はケトースとよぶ。 不整炭素原子をもち、多くの立体異性体があるので旋光性を示し、D、L二系列に分けら れる。もっとも簡単なアルドースであるグリセリンアルデヒドについて、右旋性(+)の ものをD型とし、これより誘導される糖をD系列とし、その対称体をL系列とする。 カルボニル基は通常遊離しておらず、五角または六角の環状構造をつくっており、それぞ れフラノース、ピラノースとよばれる。環形成の際にカルボニル炭素に生ずる水酸基に関 してα、βの異性体が生ずる。D系列ではより大きな右旋性をもつものをα型、他をβ型 とし、L系列ではより大きな左旋性をもつものをα型、他をβ型とする。 結晶の状態ではα、βのいずれかの型をとるが、溶液中では両者の間に平衡が成立し、時 間とともに旋光度が変化し最後に一定値に達する。この現象を変旋光という。 しかし、カルボニル炭素に生ずる水酸基がグリコシド結合によって他の単糖、アルコール、 フェノール性物質、リン酸などと結合している場合は、α、βのいずれかの形をとる。 グリコシダーゼにはα型に作用するものとβ型に作用するものとがあり、厳密に区別され る。単糖類はカルボニル基をもつので、アンモニア性銀液やフェーリング液を還元する。 また、フェニルヒドラジンとオサゾンをつくる。 アルコール基をもつため酸とエステルをつくる。酸性溶液中では脱水がおこり、アルカリ 性溶液中ではエピメル化がおこる。温和な酸化によっては酸のラクトンを生じ、強い酸化 ではジカルボン酸が生ずる。還元するとアルコールに変わる。 -2- 【炭水化物と生物】 炭水化物[carbohydrate] 地球上でもっとも多量に存在する有機化合物で、広義には糖類あるいは糖質ともいい、生 物にとってきわめて重要な物質である。 一般に炭素、水素、酸素の 3 種の元素からなり、一般式 Cm(H2O)n として書き表せるもの が多く、あたかも炭素と水の分子(H2O)からなっているようにみえるので炭水化物、古く は含水炭素とよばれた。なお、酸素原子の数が一般式より一つ少ないもの(デオキシリボ ースなど )、また硫黄(いおう)化合物を含むもの(コンドロイチン硫酸など)や窒素原子 を含むもの(アミノ糖など)も炭水化物のなかに含める。 化学的には、多価アルコールのアルデヒドまたはケトンおよびその誘導体の総称である。 【分類および種類】 炭水化物はそれを構成する単位糖の数により、単糖類、少糖類、多糖類に分類される。 (1)単糖類 アルデヒド型のものとケトン型のものがあり、それぞれアルドース(ブドウ糖など)お よびケトース(果糖など)とよぶ。 アルデヒド基またはα‐ケトール基をもつため還元性があり、フェーリング液を用いた還 元性の検出は古くからよく知られている。 また立体化学的には、不斉炭素をもつため旋光性があり、それぞれに 2 種の光学異性体が 存在する。単糖類は、1 分子のもつ炭素の数により、さらに三炭糖から七炭糖まで分類さ れる。ブドウ糖は六炭糖である。 (2)少糖類 単糖類が数個、グリコシド結合したもので、2 個結合したものをとくに二糖類といい、 ショ糖、麦芽糖などが含まれる。さらに、三糖類、四糖類などと分類される。少糖類でも アルデヒド基が結合に使われずに遊離の状態にあるものは還元性をもつが、もっともよく 知られた二糖類であるショ糖は還元基をもたない。 単糖類および少糖類はともに水に溶けやすく、エーテルには溶けない。いずれも甘味をも ち、とくに果糖は強い。 炭水化物は生物界に広く分布するが、その生物的機能は大きく分けて 〔1〕生物体の構成成分であることと、 〔2〕活動のエネルギー源となることである。 構造を保つ炭水化物はすべて多糖類であり、構造多糖ともよばれ、植物の細胞壁をつく るセルロース、昆虫や甲殻類などの外皮をつくるキチン、動物の軟骨や腱(けん)の成分で あるコンドロイチン硫酸、関節液や眼球硝子(しようし)体液に含まれるヒアルロン酸など がある。エネルギー源としての炭水化物は、脂質やタンパク質とともに生物体において重 要な役割を果たしている。 緑色植物は炭酸同化作用によって水と空気中の炭酸ガス(二酸化炭素)からブドウ糖を合 成し、これをデンプンとして貯蔵する。 動物は自ら炭水化物を合成できないので、これを植物から摂取し、グリコーゲンとして肝 臓に貯蔵する。 エネルギー源として生物体に蓄えられた多糖は栄養多糖(貯蔵多糖)ともよばれる。 これらの栄養多糖は、動植物においては解糖系やTCA回路、また酵母などでは発酵によ って代謝され、生命活動のエネルギー源や生合成に役だっている。 なお、栄養学的にはとくにデンプンとブドウ糖が重要である。 ヒトは必要とするエネルギーの大半をデンプンとして摂取するが、これはほかのほとんど の多糖類を栄養とすることができないからである。 また、ブドウ糖は、動物においては血糖として全身の、とくに脳、中枢神経系、筋肉にお けるエネルギー源として重要な役割を果たしている。 (3)多糖類 単糖類が無数にグリコシド結合してつながったもので、分子量は数千から 100 万を超え るものもある。加水分解することにより、少糖類あるいは単糖類を生じる。単一の単糖類 から構成されるものを単一多糖といい、複数種の単糖類から構成されるものを複合多糖と いう。 グリコーゲン[glycogen] 動物体内に広く存在する多糖で,貯蔵物質として植物界におけるデンプンに相当する。 食物として摂取された単糖やグリセリンなどから生体内でウリジン二リン酸グルコースを 経て合成される。 肝臓や筋肉中にとくに多量に存在する。 肝臓グリコーゲンは体内のエネルギー保存の役割を果たし,筋肉グリコーゲンは筋収縮の エネルギー源として使われる(〓解糖)。カキ,エビの類にも存在する。 植物界では菌類,酵母や,トウモロコシの種子にも見出される。 無味,無臭,非結晶性白色の粉末で,アルコールおよびエーテルに不溶,水に溶けてコロ イド溶液をつくるが糊にはならない。 還元糖ではなく,ヨード反応は紫赤色から紫褐色。比旋光度=+ 180 ~+ 210(水溶液)。 化学的にはデンプンと非常によく似ており,酸で加水分解すれば定量的に D ‐グルコー スとなる。β‐アミラーゼで加水分解すればその構成成分の半量に相当する部分はマルト ースとして遊離する。 物理的方法(拡散,超遠心分析)で解析すると,グリコーゲンは扁平楕円状で,分子量は数 百万に達する。デンプンと同じくα 1 → 4 のグリコシド結合を主鎖とし,これに 12 ~ 18 個の D ‐グルコース単位からなる側鎖がデンプンの場合より多く 6 位の炭素原子に結合 して,全体としていちじるしく分枝した巨大分子(分子量はデンプンよりやや小)を構成し ていると考えられている。 -3- -4- 単糖類[monosaccharide] アルドース[aldose] 炭水化物の一種で,これより簡単な分子に加水分解されないものをいう。 一般式 CnH2nOn で n = 2,3,4,5,6,7,8,9 および 10 であるものを,それぞれジオース,トリオ ース,テトロース,ペントース,ヘキソース,ヘプトース,オクトース,ノノースおよび デコースという。 一般に鎖式多価アルコールのアルデヒドまたはケトンに相当するもので,前者を アルド ース,後者を ケトースという。 すべての炭水化物の基礎となる物質である。 n = 3 以上のものは 不斉炭素原子をもち,不斉炭素の数に応じ立体異性体が多数ありう るが,天然に知られているのはその一部である。 天然に存在するものはペントースとヘキソースが多い。 遊離して存在するほか, ピラノース型や フラノース型で多糖,配糖体,核酸,ヌクレオ チドなどに含まれる。 これらの生合成や分解過程ではリン酸エステルやヌクレオチド誘導体となって種々の酵素 作用を受ける。 単糖は概して中性,結晶性の物質で,水に溶けやすく,アルコールには溶けにくい。 エーテルにはほとんど溶けない。 多少の甘味をもち,また苦味を帯びるものもある。 結晶状で得られる単糖は安定なピラノース構造をとり,αとβの異性体が知られている。 水溶液中でもこれらの互変異性体が主であるが,ほかに遊離のカルボニル基をもつ直鎖構 造やフラノース構造もいくらか存在し,平衡混合物となる。 このため単糖の結晶を溶解すると 変旋光を示し,また,アルコール性ヒドロキシ基によ る反応のほか,ホルミル基またはカルボニル基の反応も行なう。 アルコール性ヒドロキシ基による反応では,各種の酸と糖エステルを生成する。 たとえばリン酸エステルや酢酸エステルなどが知られている。 ヨウ化メチルと酸化銀,またはジメチル硫酸とアルカリの作用ですべての遊離ヒドロキシ 基がメチル化される。 アセトンと脱水触媒の作用でアセトン糖を形成する。また過ヨウ素酸で酸化され分解する。 アルデヒドまたはケトンとしての性質があるため,単糖は強い還元性をもち,アンモニア 性硝酸銀溶液,フェーリング液,フェリシアン化カリウムなどを還元し,濃厚溶液はシッ フ試薬を着色する。 また,フェニルヒドラジンとはヒドラゾン,さらにオサゾンを生成する。 メチルアルコールと塩化水素とによりメチル配糖体を生成する。 アルドースを穏やかに酸化すればホルミル基だけが酸化されてアルドン酸となる。 ケトースの環は酸化に際してカルボニル基のところで開裂する。 さらに単糖は弱アルカリの作用で エピマー化を含む特異な分子内変化をおこし,また無 機酸と熱して脱水すればアルドペントースはフルフラールを,アルドヘキソースは 5 ‐ヒ ドロキシメチルフルフラールを与える。この反応はこれらの糖の検出に用いられる アルデヒド官能基(ホルミル基)をもつ単糖 をいう。 アルデヒドアルコールに属する。 アルドトリオース,アルドテトロース,ア ルドペントース,アルドヘキソースなどの 種類がある。 アルドースは酸化に対してケトースとは異 なった挙動を示し,水溶液中で臭素水によ っておだやかに酸化すればホルミル基だけ が酸化されて アルドン酸といわれるモノカ ルボン酸となる。 濃硝酸と加熱して強く酸化すると分子の両 端が酸化されて アルダル酸, ガラクタル 酸のようなジカルボン酸となる。 また,アルドースのホルミル基が還元され ると糖アルコールとなる。 これらを D ‐グルコースについて示すと次の ようである -5- D ‐系列のアルドースの 構造を示すと図のように なる。 これらのものの立体配置 は,相互に炭素数を減ら したり増やしたりして, あるいは還元して糖アル コールにしたり,酸化し てジカルボン酸にして, その構造の関係から知る ことができる。 -6- ケトース[ketose] ケトン官能基(カルボニル基)をもつ 単糖を いう。 ケトアルコールに属し,ケトトリオース, ケトテトロース ,ケトペントース,ケトヘ キソースなどの種類がある。 環化に伴いアセタールまたはケタール炭素は新たに不斉中心となって,アノマーを生じる。 水溶液中ではこれらの異性体は一定の平衡混合物となる。ハースが提唱した名称。 一般的には,ピラノースはもう 1 つの環状異性体であるフラノースより安定で,結晶性が よい。ピラノース環はシクロヘキサンと同じく,いす形配座と舟形配座とをとりうるが, いす形の方が安定であると考えられている。また,いす形配座には,C1,1C という 2 つの 配座がある。 単糖としての諸性質中で アルドースと対 照的なのは酸化のされ方である。 そのさいケトースの分子はカルボニル基の ところで切れ,2 個のカルボン酸を生ずる。 たとえば D ‐フルクトースは,次のように 分解する 図(左)の C1 配座は,D ‐グルコースのアノマーがとりうる最優位のピラノース環の配座 である。1C 配座(図右)は,C1 のピラノース環が反転したものであるが,置換基とくにヒ ドロキシ基とヒドロキシメチル基との間の立体障害が増大して,C1 より不安定となる。 しかし舟形よりは安定で,準優位の立体配座である。 いす形配座[chair conformation] シクロヘキサン C 〓 H 〓〓またはその誘導体が sp 〓(4 面体)混成の最適の結合角 109 〓 28 〓を保ったままでとりうる最も安定な立体配座。シクロヘキサンの場合は,室温では ほとんどすべてがこの配座をとっている。〓ニューマン投影式からわかるように,各 C ‐ C および C ‐ H 結合は互いに反発が最小のねじれ型(スタッガード型)の配置となって いる。シクロヘキサンの場合,室温では 1 つのいす形配座から他のいす形配座への速い相 互変化(活性化エネルギー約 10kcal/mol)をしている。 フラノース[furanose] ピラノース[pyranose] 単糖の環状異性体の 1 つ。 アルドースの C1 位またはケトースの C2 位のカルボニル炭素が, それぞれ C5 位または C6 位のヒドロキシ基との間に分子内ヘミア セタールまたはヘミケタールを形成して,テトラヒドロピラン環(ピ ラノース環)構造をとったとき,ピラノースまたはピラノシド型と いう。また,そのような糖がグリコシド結合をしているとき残基 の名前をピラノシド(pyranoside)という。 -7- 単糖の環状異性体の 1 つ。 アルドースの C1 位またはケトースの C2 位のカルボニル炭素 が,それぞれ C4 位または C5 位のヒドロキシ基との間に分子 内ヘミアセタールまたはヘミケタールを形成して,テトラヒ ドロフラン環構造をとったとき,フラノースという。 ピラノースと同様にアノマーの区別がある。 ハースによって提唱された名称。フラノースはピラノースの ようにα型(D ‐グルコースでは 1 位のヒドロキシ基が 5 員環 の下方)とβ型があるが,ピラノースにくらべて不安定で,多 くの場合は溶液中に,または配糖体の成分として存在する。 α‐またはβ‐ D ‐グルコフラノースのエチルグルコシドは結晶で知られ,そのほか数 種の結晶誘導体が知られている。スクロースはα‐ D ‐グルコピラノシル‐β‐〓‐フ ルクトフラノシドである。 -8- アノマー[anomer] 糖がピラノースまたはフラノース環構造をとると,直鎖構造の場合より不斉炭素原子が 1 個ふえる。そのために生じるジアステレオマーの対を互いにアノマーであるという。たと えばα‐グルコースとβ‐グルコースは互いにアノマーであり,1 位の OH がピラノース 環面の逆側に配置している。 ジアステレオマー[diastereoisomer] 不斉原子または〓分子不斉が 2 個以上存在する分子では,鏡像異性体を得るには両方の不 斉原子(または分子不斉の部分)を反転しなければならない。もし一方をそのままにして他 の一方だけを反転した場合,両者は鏡像異性体の関係にはなく,原子団間の相対的な空間 関係の異なった異性体となる。このような異性体をジアステレオマーという。 これは旋光度ばかりでなく,ほかの物理的,化学的性質も異なっている。たとえば 2 個 の不斉炭素原子が直接結合した abcC ‐ Cdef の構造をもつ分子では,図のフィッシャー投 影式で示した 4 種の立体配置があり,このうちⅠとⅡ,ⅢとⅣは互いに鏡像異性体であっ て,I とⅢ,ⅡとⅣ,ⅡとⅢ,ⅠとⅣはそれぞれジアステレオマーである。 アレン型の有機化合物でも,たとえば図Ⅲに示すような種々のアレンが鏡像異性体に光学 分割されている。メチルシクロヘキシリデン酢酸(図Ⅳ)のようなものも鏡像異性体に分割 されており,さらにⅤのようなジフェニル誘導体も分割されている。 また非平面縮合環芳香族化合物のヘキサヘリセン(図Ⅵ,ヘリセン)も分割されている。 エピマー[epimer] 分子不斉[molecular asymmetry] 光学活性体の旋光性が分子(またはイオン)固有の特性による場合,分子内に不斉炭素原子 が存在するためのものとそうでないものとがある。後者の場合,分子の構造上旋光性の原 因となる何らかの非対称性があると考え,これを分子不斉という。分子が第 2 種の対称要 素を欠くこと,すなわち対称心,鏡映面,回反軸をもたないことがその必要条件である。 ウェルナーは金属錯体の構造に関して初めてこの説明を提出した。たとえば,トリス(エ チレンジアミン)コバルト(Ⅲ)イオン(エチレンジアミン錯体)は,図Ⅰ,Ⅱのような互い に鏡像異性体となる構造をとるため分子不斉となる。 -9- 多くの不斉炭素原子の存在による異性体のうち,1 つの不斉炭素原子についてだけの 2 つ のジアステレオマーを互いにエピマーであるという。糖について用いられる語で,狭義に は糖の 2 位の炭素原子につく OH,NH2 あるいはそれらの誘導体の向きが異なった 2 つのジ アステレオマーをいう。たとえば D ‐グルコースの狭義のエピマーは D ‐マンノースで あるが,広義には D ‐ガラクトースも含まれる。 エピマーの相互変換を触媒する酵素をエピメラーゼという。 エピマー化[epimerization] エピマーの 1 つをほかのものに変える反応。とくにアルドースおよびその一部の誘導体の 2 位(α位)の炭素原子に関するエピマーについての転位をさすことが多い。α転位(α‐ inversion)ともいう。これらの化合物をアルカリ金属またはアルカリ土類金属の水酸化物 のうすい水溶液か,ピリジンまたはキノリン中で熱すると,エピマーの 1 つは一部ほかの ものに転化して,両者の間に平衡が成立する。たとえば,D ‐グルコン酸をピリジン中で 140 ℃に熱すると,一部分 D ‐マンノン酸に転化し,糖酸など 2 塩基酸は各カルボキシ基 の隣の炭素原子においてエピマー化する。遊離のアルドースをうすいアルカリ液で処理す ると,ある程度エピマー化されるが,副反応もおこる。 なお,一般に分子中の不斉炭素原子の 1 つだけの立体配置が反対のものに変化して別のジ アステレオマーとなる反応もエピマー化という。 - 10 - エピメラーゼ[epimerase] イソメラーゼ(異性化酵素)の 1 分類をなす酵素の総称。イソメラーゼのうち 2 つ以上の光 学活性中心をもつ化合物の一方の立体構造を反転する酵素をいう。たとえば,ヒドロキシ プロリンエピメラーゼは次のように L ‐ヒドロキシプロリンの 2 位の炭素の構造を反転 し D ‐アロヒドロキシプロリンを生ずる。 二糖[disaccharide] 加水分解により 2 分子の単糖を生ずる糖の一群をいう。天然の二糖を構成する単糖は 2 分 子ともヘキソースである場合がふつうであるが,ヘキソースとペントースからなるもの(た とえばプリムベロース)もある。構成単糖の結合形式により 2 種類に分類される。 1)トレハロース型二糖。トレハロース,スクロースなど,両構成単糖の還元基をもつ炭素 原子がエーテル結合しているもの。還元糖ではなく,変旋光を示さない。 2)マルトース型二糖。マルトース,セロビオース,ゲンチオビオース,ラクトースなど, 第 1 の構成単糖の還元基をもつ炭素原子と第 2 の単糖の還元基をもたない炭素原子との間 でエーテル結合しているもの。還元糖であり,変旋光を示す。 還元糖[reducing sugar] 糖分子のアルデヒド官能基またはケトン官能基が遊離またはヘミアセタール型で存在する 単糖およびオリゴ糖。アノマーが存在するが,水溶液中では平衡混合物となるため変旋光 を示す。還元性をもち,アルカリ性重金属塩(〓フェーリング液やアンモニア性硝酸銀溶 液など)を還元する。またオサゾン,ヒドラゾンなどの誘導体を生成しうる。グルコース, フルクトース,マルトースなどがその例である。 三糖[trisaccharide] 加水分解により 3 分子の単糖を生じる糖の一群をいう。還元性の有無,構成単糖の種類お よび結合型式により分類される。植物界にはラフィノース,ウンベリフェロースなど遊離 の三糖のほか,多糖の部分加水分解物が存在するが,動物由来の 3 種は乳汁中のラクトト リオース類を除けば,複合糖質の糖鎖として存在する。 四糖[tetrasaccharide] 加水分解により 4 分子の単糖を生ずる糖の一群。スタキオース(stachyose)が知られている。 動物界では人乳中のラクトース系四糖のほかに,ガングリオテトラオースおよびグロボテ トラオースとして,糖脂質の糖鎖成分をなす。植物界ではスタキオースのように,スクロ ースを含むことが多い。 - 11 - 多糖[polysaccharide] 広義には二糖,三糖,四糖などのオリゴ糖を含めて,加水分解によって 2 分子以上の単糖 を生じるすべての炭水化物をさす。狭義には広義の多糖のうち,溶解度,味,化学的性質 が単糖と非常に異なる高分子化合物(ふつう 7 分子以上の単糖が縮合したもの)をさす。 同一の単糖分子からなるものを単一多糖(simple polysaccharide)といい,種々の単糖あるい はその誘導体(アミノ糖,ウロン酸など)からなるものを複合多糖(complex polysaccharide) という(ムコ多糖)。 多糖のうち,デンプン,グリコーゲン,イヌリン,リケニンなどは貯蔵物質として,また セルロース,キチン,ヘミセルロース,ペクチン,植物ゴム(たとえばアラビアゴム)など は皮膜物質,支柱物質として自然界に広く存在する。 一般に前者は水中で膨潤してコロイド溶液となる。生体内では酵素によって分解され,エ ネルギー源となる。これに反し後者は水に溶けにくい。カビなど限られた微生物がこれを 酵素分解して利用するが,一般に分解されにくい。希酸により加水分解すれば,三糖,二 糖などの中間生成物をへて,ついに単糖に分解される。 ムコ多糖[mucopolysaccharide] プロテオグリカン( proteoglycan)ともいう。ヘキソサミンを構成成分とする多糖。天然で はムコタンパク質を形成していて,アルカリ処理などで容易にタンパク質を除ける。糖タ ンパク質の糖部分との相異は厳密ではないが,ふつうタンパク質を含まないものとして分 子量 1 万~ 2 万以上のものをいう。ヘパリン,ヘパリチン硫酸,コンドロイチン硫酸,ヒ アルロン酸,ケラト硫酸,キチンが代表的で,単糖または二糖が重合した構造をもつ。 デンプン[starch] 澱粉,殿粉。植物の最も重要な貯蔵栄養素で,細胞内に粒状に存在する。 日中に炭酸ガスと水とから光合成されたデンプン(同化デンプン)は夜間に可溶性の糖に加 水分解してそれぞれの貯蔵場所に運ばれ,そこで粒状の貯蔵デンプンに変えられるので, 種子,果実,根,根茎に多い。デンプン粒は原料植物(ジャガイモ,トウモロコシ,コム ギなど)を冷水と粉砕し,固形物を除去した溶液から静置沈降させて得られる。 デンプンは分子が規則正しく配列して成長生成した一種の結晶とみなされる粒子として存 在する。顕微鏡で観察すると原料によって形や大きさを異にし,凹みをもつもの,それを 中心に層状構造をとるものなどがある。大きいもの(ジャガイモのデンプン粒など)は 径 0.1mm 前後,小さいもの(コメのデンプン粒など)は数μ m である。 吸湿性が強く風乾状態で 20 %の水分を含み,湿った空気中では 35 %の水分を吸収する。 冷水には不溶であるが,熱水中ではミセルが崩れて分子間隔が広がり,ついには溶けて青 みがかったゾルとなる。濃度が高いときは冷えると固まり,白みがかったゲルとなる。ま たこのような溶液からアルコールまたは塩基性酢酸鉛により沈殿する。デンプンをそのま ま熱するとまず水を失い,約 200 ℃で一部分はデキストリンに変わる。うすい無機酸と長 く煮沸すると中間生成物として可溶性デンプン,デキストリン,マルトースを経てほとん ど定量的にグルコースに加水分解される。デンプンは冷時ヨウ素溶液で青紫色に呈色する (〓ヨウ素デンプン反応)。 - 12 - デンプンの結晶構造に関しては,アミロースは顕微鏡的にも結晶性であって X 線回折 図形もその形態に従って A 形,B 形,または V 形を与えるのに反して,アミロペクチン は X 線的にも非晶形の図形しか与えない。 一般に A および B 形は生のデンプン粒(β‐デンプン)または冷水に溶けない老化したア ミロースについて得られ,そのアミロース分子は鎖状であることを示す。 これに対して,一度糊化したデンプン(α‐デンプンまたはデンプン糊という)または溶液 からコンプレクス形成試薬の存在下に老化させて得た結晶アミロースは V 形を与える。 この場合にはアミロース分子の鎖は 6 個または 7 個のグルコース残基を単位として環状に 巻き,全体として,らせん構造をとるものと考えられている。 【デンプン粒】 天然のデンプン粒は微結晶構造の部分があり、特有のX線回折図形(A、B、C形に分類 される)を示す。デンプン粒に水を加えて加熱すると糊化(こか)し、粒が壊れるとともに 微結晶構造も壊れる。オランダのカッツ J。 R。 Katz は、糊化したデンプンをα‐デン プン、もとのデンプンをβ(ベータ)‐デンプンとよんだ。α‐デンプンを水分を含んだ状 態(水分含有量 30 ~ 60 %が最適)で放置すると、部分的にもとの微結晶に近い状態に戻 る。この過程を老化というが、もとのデンプン粒のような粒に戻るのではない。 デンプン粒を構成する主成分はα 1 → 4 結合の D ‐グルカンであるアミロース (amylose)と,これにα 1 → 6 結合の側鎖が加わったアミロペクチン(amylopectin)である。 両者の割合は原料によって異なる。一般にアミロペクチン含量は 70 ~ 80 %であるが,あ る種の穀類では 98 %に達するもの,30 %以下のものなど例外もある。 デンプン粒を 70 ℃の温水で膨潤させるとアミロースだけが溶解抽出される。 これを低温に長く放置すると再び不溶性になり沈殿する(老化,retrograding)。 アミロースは 250 ないし 300 個(光散乱によれば 6000 個)の D ‐グルコピラノース単位が α 1 → 4 グルコシド結合した直鎖状分子からなり,これがらせん状に巻く傾向がある。 比旋光度[α]D =+ 220 °(水溶液)。ヨウ素と包接化合物をつくり,青色を呈する。 β‐アミラーゼの作用で,ほとんど完全にマルトースに加水分解される。 アミロペクチンはα‐ D ‐グルコピラノースの 1 → 4 結合連鎖にα 1 → 6 分岐した構造 の分子で,分子量は数百万に達する。 各連鎖は 20 ~ 26 個のグルコース残基からなる。 [α]D =+ 150 °(1N 水酸化ナトリウム溶液)。ヨウ素溶液により赤紫色に呈色する。 β‐アミラーゼの作用で分岐点より糖 4 残基手前まで加水分解され,全グルコース単位の 約 55 %に相当するマルトースを遊離し,β‐限界デキストリンとなる。 アミロペクチンにα‐アミラーゼを長時間作用させるとマルトトリオース,マルトテトラ オースとともにイソマルトースを生ずる。 アミロース,アミロペクチンなどは,無機リン酸の存在でホスホリラーゼの作用を受けて 加リン酸分解されグルコース 1 ‐リン酸を生じる。 一方,生体内では ADP グルコースまたは UDP グルコースからデンプンシンターゼの作用 で合成される。 - 13 - 【糊化デンプン】 デンプン粒を水の存在下で加熱するか、アルカリ水溶液などの水素結合を破壊する媒体中 に置くと、粒は不可逆的に膨満(または溶解)して糊化(こか)する。 糊化とともに粒は微結晶性、複屈折性を失い、粘度が上昇し、酵素(アミラーゼ)や酸な ど化学薬品に対する反応性が急激に大きくなる。 糊化はα化ともいい、糊化デンプンをα‐デンプン、元のデンプン粒をβ‐デンプンとよ ぶこともある。 糊化デンプンを水とともに放置すると、微生物の夾雑(きようざつ)がなくても粘度は低下 し、ときには沈殿を生じ、酵素や酸に対する抵抗性が増し、微結晶性を一部回復する。 この現象を老化といい、老化したデンプンを老化デンプンとよんでいる。 老化をβ化ともいうが、厳密には老化デンプンは元のデンプン粒(β‐デンプン)とは異 なり、β´化というべきである。 可溶性デンプン[soluble starch] デンプン糊に酸を作用させると最初に生ずる分解生成物で,主としてアミロデキストリン からなる。その水溶液はデンプン糊にくらべはるかに透明で,硫酸アンモニウムの半飽和 では徐々に,飽和ではただちに沈殿する。ヨウ素反応は青色で,還元性はない。 - 14 - セルロース[cellulose] (C6H10O5) n 繊維素ともいう。植物細胞壁の主成分をなす多糖。自然界に産出する有機物 中もっとも多量に存在する。藻類にも存在する。動物ではホヤの被嚢をつくっているツニ シンがセルロースの一種とされている。木綿の繊維は自然界で得られるもっとも純粋なセ ルロースである。ふつうの植物繊維はリグニン,ヘミセルロースなどを含むので,水酸化 ナトリウム,亜硫酸水素ナトリウム,次亜塩素酸ナトリウムなどで処理して精製する(〓 パルプ)。セルロース分子は D ‐グルコピラノースがβ 1 → 4 結合で連なった構造で側鎖 はない。もっとも自然界の状態に近い形で得られる木綿のセルロース分子では,約 1 万個 までの D ‐グルコース単位が結合し,長さ 1.57 μ m に達する細い繊維状分子(断面 0.4nm × 0.8nm)を形成している。セルロース繊維は分子が一定の配列をした結晶部分と,乱雑 に集合した非結晶部分とからなり,両者の適当な配合によって繊維に強度,たわみやすさ, 弾力性,染色性,吸湿性 などが生ずるものと考え られている。セルロース の X 線回折によれば,そ の結晶部分の構造は分子 内および分子間水素結合 により図のような鎖状分 子が互いに平行に規則正しく配列したものである。 純セルロースは白色の物質で,熱および電気の不良導体,比抵抗 10 〓〓 cm。水,アルコ ール,エーテルなど通常の溶媒に溶けず,シュワイツァー試薬(アンモニア性水酸化銅溶 液)や塩化亜鉛,酒石酸ナトリウム鉄とは可溶性の錯体を形成し,濃塩酸,硫酸,リン酸 などに溶ける。アルカリ溶液中で膨潤させたものはビスコースレーヨンやベンジルセルロ ース製造の中間体である。希酸と煮沸すると加水分解してついには D ‐グルコースとな るが,この分解は他の多糖にくらべて容易でない。無水酢酸と硫酸の作用で分解するとセ ロビオースのオクタ酢酸エステルが得られる。セルロースを分解する酵素セルラーゼはカ ビ,細菌,発芽種子,カタツムリなどに存在し,セルロースをセロデキストリン,セロヘ キサオース,セロテトラオース,セロトリオース,セロビオースなどに分解する。さらに β‐ D ‐グルコシダーゼ(セロビアーゼともいう)が共存すれば D ‐グルコースにまで分 解する。このほか人工のセルロース誘導体にはアルキルセルロース,ニトロセルロース, アセチルセルロース,セルロースキサントゲン酸塩など繊維工業で重要な誘導体や各種の イオン交換セルロースがあり,多くの用途がある イオン交換セルロース[ion ‐ exchange cellulose] セルロース粉末に各種の解離性置換基を導入しイオン交換樹脂様の性質をもたせたセルロ ース誘導体。ソーバーとピーターソンが開発した(1954)。通常のイオン交換樹脂にくらべ て親水性が大きいためタンパク質や核酸,多糖のような親水性生体高分子も容易に交換基 まで拡散し,急速に吸着平衡に達する。 また比較的ゆるやかな条件でこれら高分子の吸着や溶離が行なえるので,生化学の分野で イオン交換クロマトグラフ用交換体として広く使われている。 リグニン[lignin] 木材中にセルロースに伴って 20 ~ 30 %存在する,高分子のヒドロキシフェニルプロパン を基本単位とする重合化合物.細胞膜と細胞膜の間の中間層を構成し,一部は細胞膜にも 存在する.植物体の木化の原因となる。 - 15 - 【構造】セルロースはD‐(+)‐グルコースのみより構成される多糖類であり、グルコ ース単位は六員環グルコピラノース環となり、隣接するグルコース単位との間で水 1 分子 を失った形のβ‐1・4‐グルコシド結合により直鎖状につながった構造をとっている。 セルロースとデンプンは立体配置が異なっている。セルロースの分子量は数十万またはそ れ以上のこともあり、1 分子につき存在するグルコース単位の数(分子式でのnの数)は 3000 ~ 6000 個、セルロース分子の長さは 2 万~ 3 万オングストローム(Å)といわれて いる。これら鎖状セルロース分子が数多くの水酸基間の水素結合によって結び合って平行 に規則正しく配列し束をつくり、束の両端は房状となり、他の房と網状につながりセルロ ース繊維となっていると考えられている。実際に天然セルロースにおいては微結晶部分と 無定形部分とが認められ、このような構造が繊維の強さ、弾力性、染色性、吸湿性を生ん でいる。セルロースを溶解したのち再生すると結晶構造は変わる。木材ではセルロース分 子の束がリグニンの中に埋め込まれ、さらに強化された構造となっている。 【性質】 セルロースは吸湿性の強い無味・無臭の白色固体で、普通のセルロース試料は 10 %程度 の水を含んでいる。水、エタノール(エチルアルコール )、エーテルなど通常の溶媒には 溶けず、シュワイツァー試薬(アンモニア性水酸化銅溶液 )、硫酸、塩酸、リン酸に溶け る。セルロースを希酸と煮沸すると加水分解されD‐グルコースが得られるが、デンプン など他の多糖類と比べると加水分解されにくく、アルカリでは加水分解されない。セルロ ースのグルコース単位はそれぞれ 3 個の水酸基をもち、反応はこの水酸基でおこる。すな わち、セルロースはアルコールと同様にエステル、エーテルを生成する。硝酸、硫酸の混 合酸を作用させると硝酸エステルであるニトロセルロースが、クロロスルホン酸を作用さ せると硫酸セルロースが得られる。また無水酢酸と少量の硫酸で処理するとトリアセテー トとなり、これを部分加水分解すると、鎖の開裂、アセチル基の部分脱離がおこり、 200 ~ 300 単位のジアセテートすなわちアセチルセルロースが生成される。アルカリで処理し て得たアルカリセルロースに二硫化炭素、ハロゲン化アルキル、クロロ酢酸塩を作用する とセルロースキサントゲン酸塩やメチルセルロース、CMCなどのセルロースエーテルが 得られる。これらセルロースの反応は、安価で入手しやすい既製の高分子の性質を改良す るため行われ、工業的に重要な意味をもっている。 【用途】 セルロースは紡織繊維、紙として大量に用いられるほか、セルロース誘導体は綿火薬、プ ラスチック、フィルム、ビスコースレーヨン、消泡剤、分散剤、保護コロイドなどとして 用いられ、イオン交換セルロースは生化学分野での用途が多い。かつて多く製造されたセ ルロイド、セロファンもセルロース誘導体である。 【人体との関係】 人間の消化管には繊維素分解酵素(セルラーゼ)は分泌されないから、セルロースは消化 吸収されないで排泄(はいせつ)される。しかし大腸に生育する微生物のあるものはセルロ ースを分解する。セルロースは食品分析における「粗繊維」の主成分である。また腸の蠕 動(ぜんどう)運動を促進し、便秘を防ぐ。 - 16 -
© Copyright 2026 Paperzz

![【FdData中間期末:中学理科2年:消化】 [消化液の種類] [問題](1学期](http://s3.paperzz.com/store/data/005516166_1-b9b055c5cacd6c3d00557930dd989014-250x500.png)