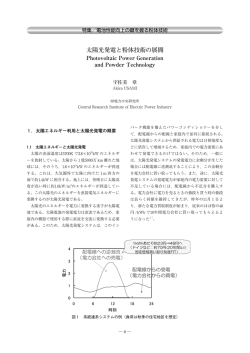
ISSN 0286−4754 栄 養 生 理 研 究 会 報 Proceedings of Japanese Society for Animal Nutrition and Metabolism Vol.57, No.2 家 畜 栄 養 生 理 研 究 会 Japanese Society for Animal Nutrition and Metabolism 2013 会 報 目 次 (Vol. 57, No. 2) カンショ焼酎粕濃縮液を利用した肉用牛肥育技術の開発 59 神谷 充 1・服部育男 1・佐藤健次 2・常石英作 1・神谷裕子 1・永濱誠一 3・ 日高明生 3・上村昌志 3・林 義朗 1 (1(独)農業・食品産業技術総合研究機構 九州沖縄農業研究センター; 2 (独)農業・食品産業技術総合研究機構 中央農業総合研究センター; 3 農業生産法人 錦江ファーム) 麹菌の栄養生理機能 67 大塚 彰 1・林 國興 2 (1 鹿児島大学農学部;2 源麹研究所) 周年放牧肥育牛肉の生産技術と肉質特性 79 中村好德 ((独)農業・食品産業技術総合研究機構 九州沖縄農業研究センター) Tissue distribution and blood circulation of agouti signaling protein (ASIP) in relationship with body composition in cattle 1 1 1 2 3 4 E. Albrecht , K. Komolka , K. Will , B. Kuhla , H. Sauerwein , T. Gotoh , S. Maak 89 1 (1Leibniz Institute for Farm Animal Biology, Institute for Muscle Biology and Growth, Dummerstorf, Germany; 2Leibniz Institute for Farm Animal Biology, Institute of Nutritional Physiology "Oskar Kellner", Dummerstorf, Germany; 3University of Bonn, Institute for Animal Science, Physiology and Hygiene Unit, Bonn, Germany; 4Kyushu University, Kuju Agricultural Research Center, Oita, Japan) The impact of retinoic acid on adipogenesis in 3T3-L1 cells 1 2 Katharina Stoecker , Steffen Sass , Michael W. Pfaffl 97 1 (1Physiology Weihenstephan, ZIEL Research Center for Nutrition and Food Sciences, Technische Universität München, Freising, Germany; 2Institute of Computational Biology, Helmholtz Center Munich, German Research Center for Environmental Health, Oberschleißheim, Germany) 家畜栄養生理研究会会則 107 栄養生理研究会報編集・投稿規定 108 家畜栄養生理研究会役員(平成 25 年度)名簿 109 Contents (Vol. 57, No. 2) Development of feeding systems using sweet-potato condensed distillers solubles in fattening Japanese Black steers 1 1 2 1 59 1 Mitsuru Kamiya , Ikuo Hattori , Kenji Sato , Eisaku Tsuneishi , Yuko Kamiya , Seiichi Nagahama3, Akio Hidaka3, Masashi Kamimura3, and Yoshirou Hayashi1 (1NARO Kyushu Okinawa Agricultural Research Center; 2NARO Agricultural Research Center, 3 Kinkou Farm) Nutritional and physiological function of Aspergillus awamori as a feed additive 67 Akira Ohtsuka1 and Kunioki Hayashi2 (1Department of Biochemical Science and Technology, Faculty of Agriculture, Kagoshima University; 2Biogenkoji Research Institute) A beef production system and meat quality using year-round outdoor grazing and self-sufficient feed 79 Yoshi-nori Nakamura (NARO Kyushu Okinawa Agricultural Research Center) Tissue distribution and blood circulation of agouti signaling protein (ASIP) in relationship with body composition in cattle 89 E. Albrecht1, K. Komolka1, K. Will1, B. Kuhla2, H. Sauerwein3, T. Gotoh4, S. Maak1 (1Leibniz Institute for Farm Animal Biology, Institute for Muscle Biology and Growth, Dummerstorf, Germany; 2Leibniz Institute for Farm Animal Biology, Institute of Nutritional Physiology "Oskar Kellner", Dummerstorf, Germany; 3University of Bonn, Institute for Animal Science, Physiology and Hygiene Unit, Bonn, Germany; 4Kyushu University, Kuju Agricultural Research Center, Oita, Japan) The impact of retinoic acid on adipogenesis in 3T3-L1 cells 97 Katharina Stoecker1, Steffen Sass2, Michael W. Pfaffl1 (1Physiology Weihenstephan, ZIEL Research Center for Nutrition and Food Sciences, Technische Universität München, Freising, Germany; 2Institute of Computational Biology, Helmholtz Center Munich, German Research Center for Environmental Health, Oberschleißheim, Germany) 平 成 25 年 度 家畜栄養生理研究会 秋季集談会開催のご案内 テ ー マ : 食 肉 生 産 と 栄 養 生 理 開 催 日 時 : 2013 年 11 月 9 日 ( 土 ) 11: 00∼ 11: 50 評 議 員 会 (3F: 研 修 室 1 ) 13: 00∼ 17: 00 集 談 会 ( 4F: 第 3 会 議 室 ) 18: 00∼ 20: 30 懇 親 会 会 場:熊本国際交流会館 ( 〒 860-0806 熊 本 市 中 央 区 花 畑 町 4 番 1 8 号 TEL: 096-359-2020 FAX: 096-359-5783 http://www.kumamoto-if.or.jp/kcic/) 懇親会・宿泊場所:平山温泉 恵荘 (熊本県山鹿市平山 5300 TEL:0968-44-0830 http://megumisou.com/index2.html) 会 費:集談会 ・・・・・・・・・ 一般会員 無料 非会員 2,000 円 懇親会 ・・・・・・・・・ 一般会員 14,000 円 (大学院生)10,000 円 懇親会のみ 7,000 円 連 絡 先:九州大学農学部附属農場・高原農業実験実習場 家畜栄養生理研究会(事務局庶務担当:後藤貴文) TEL:0974-76-1377 FAX:0974-76-1218 E-mail: [email protected] HP: http://www.agr.kyushu-u.ac.jp/lab/kuju_hp 平 成 25 年 度 家畜栄養生理研究会 秋季集談会予定時間 13: 00∼ 13: 40 農研機構・九州沖縄農業研究センター 神谷 充 「カンショ焼酎粕濃縮液を利用した肉用牛肥育技術の開発」 13: 40∼ 14: 20 鹿児島大学農学部 大塚 彰 「麹菌の栄養生理機能」 14: 20∼ 14: 40 休 憩 14: 40∼ 15: 20 農研機構・九州沖縄農業研究センター 中村 好徳 「周年放牧肥育牛肉の生産技術と肉質特性」 15: 20∼ 16: 00 ラ イ プ ニ ッ ツ 家 畜 生 物 学 研 究 所 Elke Albrecht 「Tissue distribution and blood circulation of agouti signaling protein (ASIP) in relationship with body composition in cattle」 16: 00∼ 16: 40 ミ ュ ン ヘ ン 工 科 大 学 生 理 学 研 究 所 Katharina Stoecker 「The impact of retinoic acid on adipogenesis in 3T3-L1 cells」 18:30∼20:30 懇親会 栄養生理研究会報 Vol. 57, No. 2 2013 カンショ焼酎粕濃縮液を利用した肉用牛肥育技術の開発 神谷 充 1・服部育男 1・佐藤健次 2・常石英作 1・神谷裕子 1・永濱誠一 3・日高明生 3・ 上村昌志 3・林 義朗 1 (1(独)農業・食品産業技術総合研究機構 九州沖縄農業研究センター、 2 (独)農業・食品産業技術総合研究機構 中央農業総合研究センター、 3 農業生産法人錦江ファーム) 1.はじめに TMR を活用した肉用牛肥育技術開発に関する試 焼酎粕など食品製造副産物は飼料費の低減 験を行った。 や食品循環資源の有効利用などを目的に飼料 利用が進められており、エコフィード認証制度 2.カンショ焼酎粕濃縮液の飼料成分 においても推進食品循環資源に位置づけられ、 1) 2006 年 5 月~2007 年 4 月の間に宮崎県の焼 有効活用が期待されている 。主に南九州で生 酎粕処理プラントから産出されたカンショ焼 産されている芋焼酎の製造副産物であるカン 酎粕濃縮液について、1 ヵ月に 1 回で合計 12 回 ショ焼酎粕については、一部で飼料として利用 サンプルを採取し、飼料成分を分析した 3)。カ されてきたが、焼酎粕の原液は水分が多く、保 ンショ焼酎粕濃縮液は、芋焼酎の廃液(水分約 存性が悪いのが難点であり、乾燥飼料化すると 95%)を固液分離後、液体部分をさらに遠心分 費用が掛かることから、焼酎粕の飼料利用は拡 離で固液分離し、その液体部分を加熱濃縮した 大しない状況にあった。しかし、最近では焼酎 ものである。カンショ焼酎粕濃縮液の乾物率は 粕を固液分離した後の液体部分を濃縮する焼 49%(変動係数 9.4%) 、粗蛋白質は 20%DM(変動 酎粕濃縮プラント 2) が設立され、水分含量が少 係数 7.6%)で、粗蛋白質の 95%が溶解性蛋白質 なく、保存性の高い焼酎粕濃縮液が生産される であった。固液分離の工程で、繊維は固体部分 ようになり、飼料利用の拡大が見込めるように として除去されるため、繊維成分はほとんど含 なった。 まれておらず、NFE は 65%DM(変動係数 4.0%) 、 一方、飼料生産調製作業の外部委託が進んで 粗灰分は 14%DM(変動係数 7.9%)と高い値であ いく中で、飼料自給率の向上や飼料費低減を目 った。推定 TDN は 77%(変動係数 1.5%)であり、 的に TMR(Total Mixed Ration:混合飼料)セン 肥育牛用飼料原料として期待できる飼料成分 ターが設立されるようになってきた。TMR セン 値であった。 ターでは食品製造副産物や国産飼料を利用し また、サイレージ発酵に影響を及ぼす成分に た発酵 TMR が生産されており、南九州では肉用 ついては、水分活性は 0.88(変動係数 3.2%) 、 牛向けの発酵 TMR の生産、 利用が始まっている。 pH は 3.9(変動係数 1.9%)、単少糖含量は 肉用牛における発酵 TMR の利用は繁殖牛や育成 22.5%DM(変動係数 10.6%)で問題ない数値であ 牛が主であるが、今後は飼料価格の高騰や飼料 ったが、緩衝能は 1358 mE/kg DM(変動係数 7.4%) 用米など国産飼料の推進事業を背景に、肥育牛 と高く発酵への悪影響が懸念される数値であ における発酵 TMR の利用拡大も見込まれる。 った。 そこで、カンショ焼酎粕濃縮液やその発酵 Development of feeding systems using sweet-potato condensed distillers solubles in fattening Japanese Black steers Mitsuru Kamiya1, Ikuo Hattori1, Kenji Sato2, Eisaku Tsuneishi1, Yuko Kamiya1, Seiichi Nagahama3, Akio Hidaka3, Masashi Kamimura3, and Yoshirou Hayashi1 (1NARO Kyushu Okinawa Agricultural Research Center, 2NARO Agricultural Research Center, 3Kinkou Farm) - 59 - 3.カンショ焼酎粕濃縮液の混合が発酵 TMR の 平均体重 575 kg)を用い、配合飼料にカンショ 発酵品質に及ぼす影響 焼酎粕濃縮液を添加して、 添加水準 0%DM, 10%DM, 発酵 TMR の原料としてカンショ焼酎粕濃縮液 20%DM, 30%DM の 4 区について1試験期 2 週間と を混合することが発酵 TMR の品質に及ぼす影響 する 4×4 ラテン方格試験で比較した 5)。飼料は を調べるため、夏季と冬季にカンショ焼酎粕濃 濃厚飼料を自由採食させ、粗飼料として稲わら 縮液を乾物あたり 0, 10, 20, 30%混合した TMR を 1 日 1 頭あたり原物重量で 1.5 kg を上限に を調製し、調製後 3 週間と 7 週間目に開封して 給与した。また、酸不溶性灰分(AIA)を指示 発酵品質と好気的変敗の生起の有無を調査し 物質としたインデックス法で糞中窒素排泄量 4) た 。発酵 TMR はビニール袋に 1 袋あたり 15 kg を算出し、クレアチニンを指示物質として尿中 詰め込んで、ビニール袋内の空気を脱気後、密 窒素排泄量を算出した。 封して保存した。 濃厚飼料摂取量(kg/日)は 10~20%区で最も 高値を示し(P < 0.05) 、粗飼料摂取量(kg/日) その結果、pH は 0%区と比較して 10%区が高く、 冬季では 10%区からカンショ焼酎粕濃縮液の混 に有意な影響は認められなかった。その結果、 合割合が高まるにつれて低下した。乳酸含量は TDN 摂取量(kg/日)は 10~20%区で最も高値を 0%区と比較してカンショ焼酎粕濃縮液を混合 示した(図 1、P < 0.05(二次)) 。また、CP 摂 することで低下し、乳酸発酵を抑制する方向に 取量(kg/日)もカンショ焼酎粕濃縮液給与に 作用したが、同時に C2+C3 も低下する傾向にあ よって高値を示した(図 2、P < 0.05(二次)) 。 ったため、フリーク評点は 90 点以上と発酵品 窒素出納に関しては、窒素摂取量はカンショ焼 質が優れたサイレージと評価された。したがっ 酎粕濃縮液給与によって高値を示し(P < 0.05 て、カンショ焼酎粕濃縮液を混合しても、混合 (二次))、尿中窒素排泄量は 10~20%区で最も 割合や調製時期に関わらず、優れた発酵品質が 高値を示した(P < 0.05(二次))が、窒素摂 得られることが分かった。 取量に対する糞中窒素排泄量、尿中窒素排泄量、 一方、発酵 TMR の開封後の好気的変敗による 蓄積窒素の割合には有意な影響は認められな TMR 温度の上昇は 0%区と他のカンショ焼酎粕濃 かった(表 1) 。 縮液添加区と同程度であった。これまでの実験 以上より、配合飼料にカンショ焼酎粕濃縮液 で、麦焼酎粕濃縮液や米焼酎粕濃縮液を混合し を添加した飼料を肥育期の黒毛和種去勢牛に た発酵 TMR では乳酸など有機酸含量が高まり、 給与したところ、配合飼料に 30%DM 添加しても 開封後の TMR 温度上昇を抑制する効果が確認さ 窒素利用に影響はなく、10~20%DM では濃厚飼 れていたが、カンショ焼酎粕濃縮液を混合した 料摂取量は増加する可能性があることが分か 発酵 TMR では乳酸など有機酸含量が比較的少な った。 く、開封後の TMR 温度上昇も 0%区と同程度であ ることから、カンショ焼酎粕濃縮液には好気的 5.黒毛和種肥育牛の仕上げ期におけるカンシ 変敗の抑制効果は期待できないことが分かっ ョ焼酎粕濃縮液の発酵 TMR 給与試験 た。 カンショ焼酎粕濃縮液を混合した発酵 TMR の 給与が肥育末期に仕上がるきめ・締まりなど肉 4.黒毛和種肥育牛の飼料摂取量と窒素出納に 質に及ぼす影響を検討するため、鹿児島県南さ 及ぼすカンショ焼酎粕濃縮液の添加割合の つま市の農業生産法人錦江ファーム金峰農場 影響 で仕上げ期 6 ヵ月間の肥育試験を行った 6)。黒 カンショ焼酎粕濃縮液の給与が肥育牛の飼 毛和種去勢牛 12 頭(23 ヵ月齢、770 kg)を用 料摂取量と窒素出納に及ぼす影響を検討する い、対照区 6 頭には慣行飼料(配合飼料と稲わ ため、黒毛和種去勢牛 4 頭(平均 16 ヵ月齢、 ら)、試験区 6 頭には発酵 TMR(カンショ焼酎粕 - 60 - るため、鹿児島県霧島市の農業生産法人錦江フ ァーム溝辺農場で肥育中後期 12 ヵ月間の肥育 試験を行った 7)。黒毛和種去勢牛 11 頭(17 ヵ 月齢、600 kg)を用い、対照区 5 頭には慣行飼 料(配合飼料と稲わら) 、試験区 6 頭には発酵 TMR(カンショ焼酎粕濃縮液 25%DM、 玄米 23%DM、 フスマ 21.5%DM、乾燥豆腐粕 11%DM、稲わら 14.1%DM、脂肪酸カルシウム 3%DM、ミネラル類 2.4%DM)で慣行飼料の 6 割程度を代替して給与 図 1.カンショ焼酎粕濃縮液の添加が TDN 摂取 した。 量に及ぼす影響 (日本畜産学会報からの転載) 飼料摂取量、日増体量、枝肉格付成績、胸最 長筋の粗脂肪含量に有意差は認められなかっ た(表 4、表 5、図 4)。胸最長筋の脂肪酸組成 では対照区と比較して試験区でステアリン酸 (C18:0)が有意に多かった(表 5)が、他の脂 肪酸には影響は認められなかった。また、胸最 長筋のαトコフェロール含量は対照区 0.52 mg/100 g、試験区 0.67 mg/100 g で高い傾向を 示した。さらに、配合飼料にカンショ焼酎粕濃 縮液を 20%DM 添加して仕上げ期の約 5 ヵ月間の 肥育試験を行ったところ、胸最長筋のαトコフ ェロール含量が 1.9 倍に増加したことを確認し 図 2.カンショ焼酎粕濃縮液の添加が CP 摂取 た(データ未発表)。したがって、カンショ焼 量に及ぼす影響 (日本畜産学会報からの転載) 酎粕濃縮液を混合した発酵 TMR の給与で飼養成 績に問題なく良好な枝肉成績が得られること が分かった。また、カンショ焼酎粕濃縮液を活 濃縮液 30%DM、玄米 30%DM、フスマ 26%DM、乾燥 用することで、飼料自給率に加えて肉質の向上 豆腐粕 11.5%DM、ミネラル類 2.5%DM)で慣行飼 も期待できると考えられた。 料の 6 割程度を代替して給与した。 飼料摂取量、日増体量、枝肉格付成績、胸最 7.まとめ 長筋の粗脂肪含量と脂肪酸組成に有意差は認 カンショ焼酎粕濃縮液の飼料成分値と年間 められなかった(表 2、表 3、図 3) 。カンショ 変動が明らかになり、発酵 TMR 原料としてはカ 焼酎粕濃縮液を混合した発酵 TMR の嗜好性は良 ンショ焼酎粕濃縮液による好気的変敗抑制効 好で、特に枝肉のきめ・締まりなど肉質に問題 果は期待できないが、30%DM 混合しても発酵品 は認められず、良好な枝肉成績が得られること 質には問題ないことが分かった。配合飼料にカ が分かった。 ンショ焼酎粕濃縮液を 30%DM 添加しても窒素利 用に問題なく、飼料摂取量も良好であることか 6.黒毛和種肥育牛の肥育中後期におけるカン ら、肥育牛用の飼料原料として利用できること ショ焼酎粕濃縮液の発酵 TMR 給与試験 が分かった。肥育牛の仕上げ期 5 ヵ月間または カンショ焼酎粕濃縮液を混合した発酵 TMR の 肥育中後期 12 ヵ月間に、カンショ焼酎粕濃縮 給与が飼養成績と肉質に及ぼす影響を検討す 液をそれぞれ 30%、25%配合した発酵 TMR で慣 - 61 - 表 1. 窒素出納に及ぼすカンショ焼酎粕濃縮液の添加割合の影響 (日本畜産学会報からの転載) 表 3. 仕上げ期におけるカンショ焼酎粕濃縮 表 2. 仕上げ期におけるカンショ焼酎粕濃縮 液の発酵 TMR 給与が胸最長筋の粗脂肪含量と 液の発酵 TMR 給与が飼養成績と枝肉格付成績 脂肪酸組成 に及ぼす影響 (日本暖地畜産学会報からの転載) (日本暖地畜産学会報からの転載) - 62 - 表 4. 肥育中後期におけるカンショ焼酎粕濃 表 5. 肥育中後期におけるカンショ焼酎粕濃 縮液の発酵 TMR 給与が飼養成績と枝肉格付成 縮液の発酵 TMR 給与が胸最長筋の粗脂肪含量 績に及ぼす影響 と脂肪酸組成 (日本暖地畜産学会報からの転載) (日本暖地畜産学会報からの転載) - 63 - 図 3.仕上げ期におけるカンショ焼酎粕濃縮液の発酵 TMR 給与試験のロース芯写真 第 6-7 胸椎間ロース芯の写真(上 6 枚:試験区、下 6 枚:対照区) 図 4.肥育中後期におけるカンショ焼酎粕濃縮液の発酵 TMR 給与試験のロース芯写真 第 6-7 胸椎間ロース芯の写真(上 6 枚:試験区、下 5 枚:対照区) 行飼料の 6 割程度代替したところ良好な枝肉成 ーム職員の皆様、九州沖縄農業研究センター業 績が得られ、カンショ焼酎粕濃縮液は、肥育牛 務第 1 科の皆様、三井造船株式会社の皆様に感 用の発酵 TMR 原料としても利用可能であること 謝いたします。 が分かった。 【引用文献】 謝 辞 1) エコフィード認証制度協議会.2010.エコ 本研究の一部は、農林水産省委託プロジェク フィード認証制度実施の手引き. 2.中央 ト(粗飼料多給による日本型家畜飼養技術の開 畜産会.東京. 発)および(自給飼料を基盤とした国産畜産物 2) 宮崎陽子・濱本 修・西岡俊一郎 2008. の高付加価値化技術の開発)で実施した。本研 省エネ・高付加価値型焼酎粕飼料化システ 究にご協力いただいた農業生産法人錦江ファ ムの構築.三井造船技報,193:30-35. - 64 - 3) 服部育男・佐藤健次・鈴木知之・神谷 充・ 加藤直樹 2010.原料および産出時期が焼 中窒素排泄量に及ぼす影響.日本畜産学会 報(印刷中). 酎粕濃縮液の飼料成分に及ぼす影響.日本 6) 神谷 充・服部育男・常石英作・上村昌志・ 暖地畜産学会報, 53:175-181. 日高明生・永濱誠一・中村好德・佐藤健次 4) 服部育男・神谷 充・鈴木知之・西村和志・ 2010.黒毛和種肥育牛の仕上げ期における 佐藤健次・加藤直樹 2012.焼酎粕濃縮液 玄米と食品残さの発酵TMR給与が飼養成績、 の混合割合および貯蔵季節,貯蔵期間が発 血漿中成分、枝肉成績、および肉質に及ぼ 酵TMRの発酵品質に及ぼす影響.日本草地 す影響.日本畜産学会報, 81:481-488. 学会誌, 58:173-182. 7) 神谷 充・服部育男・上村昌志・佐藤健次 5) 神谷 充・服部育男・神谷裕子・佐藤健次・ 林 義朗 2013.カンショ焼酎粕濃縮液の 2011.肥育牛の飼料用米と食品残さの発酵 TMR給与.日本暖地畜産学会報, 54:149-159. 給与が黒毛和種肥育牛における飼料摂取 量,消化率,血漿中尿素窒素濃度および尿 - 65 - 栄養生理研究会報 Vol. 57, No. 2 2013 麹菌の栄養生理機能 大塚 彰 1・林 國興 2 (1 鹿児島大学農学部、2 源麹研究所) 1.はじめに は酵母の代謝産物や化合物として焼酎粕中に 鹿児島を中心とした南九州地域では、甘藷 含有され、摂取した動物の生体内でそれらが生 (サツマイモ)焼酎が盛んに造られており、そ 理活性や機能性を発現すると考えられる。焼酎 の醸造工程において麹菌が必須となる。麹菌は 粕以外の食品残さ等を Asp.awamori の発酵熱を Aspergillus に属する糸状菌で、分生子(胞子) 利用して乾燥飼料化すると、成長促進等の機能 の色によって黄麹菌 Asp.oryzae、黒麹菌(泡盛 性が付与されることからも、この考えが支持さ 麹菌)Asp.awamori、白麹菌 Asp.kawachi に分 れる 14,15)。そのため、麹菌そのものをニワトリ 類される。黄麹菌は清酒、味噌、醤油、一部の に飼料投与することで、焼酎粕と同様の成長促 焼酎の製造に、黒麹菌および白麹菌は焼酎製造 進効果や腸内細菌叢に対するプロバイオティ に使われる 1) 。甘藷焼酎の製造に用いられる クス様効果が期待され、これが研究の発端でも Asp.awamori は、醸造工程の初期段階(製麹: あった。ところが、麹菌の栄養生理機能は当初 麹菌を蒸米に種付けして繁殖させ 麹 を作る の予想以上に多様であり、そのどれもが非常に 工程)より、原料デンプンの糖化を行う耐酸性 興味深い機能であることがわかってきた。麹菌 のアミラーゼをはじめとして、プロテアーゼ、 Asp.awamori の飼料投与の効果は 0.05%という リパーゼ、および非デンプン性多糖類分解酵素 非常に低いレベルでも再現良く現れ、飼料の利 など各種の多様な酵素を産生・放出し 2,3) 、同 用効率の向上をはじめとして、骨格筋タンパク 時にもろみの pH を低下させ雑菌の育成を抑え 質分解系への影響を介した成長促進および筋 ながら安全に発酵させるために重要なクエン 量増加効果、脂肪酸不飽和化の促進などの脂質 4) 酸を産生する 。加えて甘藷焼酎の香味に重要 代謝への影響、酵母との組合せによる相乗効果、 なゲラニオール等のモノテルペンアルコール ならびに生体抗酸化系の賦活効果が確認され、 の生成に麹菌のβ-グルコシダーゼが関与して それらの機能性を生かした新しい飼料添加物 いる 5,6) 。このように麹菌は醸造工程を通して 様々な重要な役割を担っている。 甘藷焼酎の製造に際して、大量の蒸留廃液 としての応用が期待される。本稿では、これま でに肉用鶏ブロイラーにおいて得られた知見 を中心に、麹菌 Asp.awamori の栄養生理機能に (焼酎粕)が排出されるが、95%を占める水分 ついて解説する。 を除去すれば CP 含量が 30%の比較的良質な飼 7,8) 。加えて、我々はこれまでに 2.Aspergillus awamori の飼料投与がブロイラ 焼酎粕、特にその遠心分離上清のエーテル抽出 ーの成長と骨格筋タンパク質代謝に及ぼす 画分中に、ニワトリの成長促進作用を見出し 影響(試験1) 料原料となる 9,10,11,12) 、その原因物質の一つがブトキシブチ ブロイラーにとって最も重要な生産物は鶏 ルアルコール(BBA)であることを明らかにし 肉−すなわち骨格筋である。ブロイラー生産に ている 13)。BBA 等の物質は、原料であるサツマ おいては、効率よく飼料を骨格筋へ変換させる イモやコメに由来するのではなく、麹菌あるい ことに力が注がれてきた。その研究過程におい Nutritional and physiological function of Aspergillus awamori as a feed additive Akira Ohtsuka1 and Kunioki Hayashi2 (1Department of Biochemical Science and Technology, Faculty of Agriculture, Kagoshima University, 2 Biogenkoji Research Institute) - 67 - 表1.Aspergillus awamori の飼料投与がブロイラーヒナの飼養成績、飼料の消化率および血漿 3-メチル ヒスチジン濃度に及ぼす影響(試験1) !"#Aspergillus awamori$%&'()*+,-./0$%1234%&$56789:;<3-=>?/@>ABCDEFGHIJKLM"N Aspergillus awamori !"#$% &'% 0.05% ()* (g/12 d) 691 !"+,* (g/12 d) 1197 !"-./ (gain/feed) 0123* (% body weight) 456789:/ (%) ;<=>?89:/ (%) ± a 36 ± 0.14 23.9 ± 1.1b ± 4.6 b ± b 64 (µmol/mL) 40b 1.7 59 @AB-CD=EFDG5HI ± 30.2 ± 4 1.5 808 1057 a 15a ± b 37 b 704 ± 33b 1098 ± 26ab 1.5 ± 0.05ab 1.3 ± 0.04 28.8 ± 0.4a 27.3 ± 1.0a ± 4.1 a 76 ± 4.4a ± a 73 ± 3a 18.7 ± 1.3ab 75 74 a 0.2% ± 15.8 ± 3 1.1 b JKLMNOPQRn=10STUVWXYZ[\]^Q_`RP < 0.05STRYa 18 b`cdefS て、焼酎粕の遠心分離上清エーテル抽出画分中 体して、臓器および組織を採取し、重量の測定 10,11) (後にこれは を行い、浅胸筋(M.pectoralis major)の一部 ヘキサンで効率よく抽出されることが分かっ を以下の分析に供した。全血を採取し、骨格筋 にブロイラーの成長促進因子 た 16) )が存在し、その作用機構として、骨格筋 タンパク質分解の指標である 3-メチルヒスチ タンパク質分解関連因子を遺伝子発現レベル で下方制御していることが示された ジンの血漿濃度を HPLC 法で測定した 16) 19) 。筋組 。麹菌 織 片 ( 100 mg) よ り 、 総 RNA の 抽 出 を 行 い Asp.awamori にも同様の成長促進効果が予測さ (RNeasy® Fibrous tissue mini, Qiagen)、続 れ、Asp.awamori そのものの飼料投与がブロイ けて cDNA の合成を行った[PrimeScript® RT ラーの成長および骨格筋タンパク質代謝に及 ぼす影響を調べた 1 17,18) reagent Kit (Takara)]。リアルタイム PCR 法 。 によって[7300 Real Time PCR system (Applied チャンキー種(Ross308)雄初生ヒナ 60 羽を Biosystems) + SYBR® Premix Ex Taq™ Perfect 保温環境下に置き、市販の肉用鶏肥育前・後期 Real Time(Takara)]、骨格筋タンパク質分解お 用配合飼料[日和産業、パワーチキンP(CP 22%、 よび合成関連因子の遺伝子発現量を調べた。 ME 3.0 Mcal/kg)]を自由摂取させ群飼育した。 増 体 量 は Asp.awamori 投 与 区 で 増 加 し 、 15 日齢時に体重のそろった 30 羽を選抜し、ト 0.05%が最大値(17%増、P<0.05)を示した。 ウモロコシおよび大豆粕を主原料とする自家 一方で飼料摂取量は Asp.awamori 投与区で減少 配合基礎飼料(CP 23%、ME 3.1 Mcal/kg、抗 し、その結果、飼料要求率が 0.05%投与区で改 生物質を含まない)のみを給与する対照区、な 善された(P<0.05)。浅胸筋重量も増体量と同 らびに同飼料に Asp.awamori 種麹を 0.05%およ 様の結果であった。タンパク質およびエネルギ 4 10 お ー消化率は共に Asp.awamori 投与区で改善され 10 個/g 飼料)添加して投与する二つ (P<0.05)、Asp.awamori 投与による飼料の利用 の試験区に分け(10 羽/区)、試験飼料の給与を 効率の上昇が確認された(表1)。飼料成分の 開始し、同時にアルミニウム製ケージ(幅 40 消化率の改善は、先に述べた麹菌が産生する酵 び 0.2%(胞子数としてはそれぞれ 25 よび 10 高さ 60 5 奥行き 50 cm)中での個別飼育に切 素群の影響が考えられる。摂取された り替えた。照明は常時点灯とした。試験中、増 Asp.awamori は 体量および飼料摂取量の測定を行い、飼料要求 トリの体温で保温されることで、胞子の発芽が 率を算出した。飼料のタンパク質およびエネル 起こり、酵素の産生・放出を開始する。産生さ ギー消化率の測定は、酸化クロムを用いた指示 れる酵素は耐酸性であるため、消化管上部の pH 物質法で行った。12 日間の試験の後、ヒナを解 でも働くことが可能である。また Asp.awamori - 68 - 嚢で水分を得ると同時にニワ 消化管下部に到達した後も効果を発揮できる。 麹菌はプロテアーゼを産生するが、抽出プロテ アーゼ単独投与では飼料の消化率へ影響を及 ぼさない 20) 。ところが麹菌由来のヘミセルラー ゼおよびペクチナーゼのブロイラーへの飼料 投与は、タンパク質消化率の改善を介して飼養 成績を向上させる 21,22)。それらの酵素による飼 料原料の細胞壁の消化によって、細胞内タンパ ク質と麹菌プロテアーゼとの接触が容易にな る。また、大豆中に含まれるトリプシンインヒ ビターは、麹菌による発酵処理で除去が可能で ある 23)。Asp.awamori による消化率の改善にも 上記のような複数の酵素が関与する機構が推 察された。 骨格筋タンパク質分解の指標である 3-メチ ルヒスチジンの血漿濃度は Asp.awamori 投与区 で減少し、0.05%区では対照区の半分程度であ った(P<0.05) (表1)。これより筋細胞内のタ ンパク質分解システムの抑制が考えられた。そ こで、ユビキチン−プロテアソーム系(UPS) 関連因子およびカルパインの遺伝子発現につ いて調べたところ、ユビキチン、プロテアソー ム C2 サブユニット、およびμ-カルパイン大サ ブユニットの各 mRNA の発現量が Asp.awamori ! .,/01/$/# 2345 6* "7 8"#$%"&9 $! , , #! "! ! %&! :%"$;-<"2; 2345 6* "7 8"#$%"&9 されることから、経口摂取した Asp.awamori は %&! !"#$%"& - $! '(')* '(+* , " , #! "! ! %&! !>!-&=-/# !-&=-/# 2345 6* "7 8"#$%"&9 !"#$%"& - '(')* $! '(+* , # , #! "! ! %(! !"#$%"& %'! ">58$/#- 2345 6* "7 8"#$%"&9 は嫌気状態でも生存可能であり、盲腸でも検出 '(')* , '(+* $ -, %&! - $! #! "! ! !"#$%"& '(')* '(+* !"#$%&'(()" *+*,-%' 投与区で有意(P<0.05)に減少していた(図1)。 カルパイン系は筋原線維の断片化に、UPS は断 図1.Aspergillus awamori のブロイラーヒナへの 片化線維の微細化に重要であり、両者の協調に 飼料投与が骨格筋タンパク質分解および合成関 より筋原線維の完全分解が行われている 24) 。タ 連因子の遺伝子発現量に及ぼす影響(試験1) ンパク質合成系に関しては、アクチン mRNA の 平均値 発現量のみが Asp.awamori 0.05%区で上昇して 差あり(P < 0.05).(文献 18 より改変引用) おり、影響は部分的であった。ニワトリにおい ては、骨格筋のタンパク質合成系よりむしろタ おいてもタンパク質分解系を抑制するが 11)、そ ンパク質分解系の方が、成長に対して大きな影 の作用様式より Akt-mTOR シグナル伝達系への 響を及ぼすため 25,26,27) 標準誤差(n=10).異なる文字間に有意 影響が示唆される 13)。 、より多くの骨格筋を生 産するためには分解系の制御が重要である。焼 酎粕中にはこれまでに二種類の成長促進物質 3.Aspergillus awamori の飼料投与が脂質代謝 [BBA およびジブトキシブタン(DBB)]が発見 および筋組織の脂肪酸組成に及ぼす影響 13) されている。 これらの物質は Asp.awamori の (試験2) 生成物と考えられ、ニワトリ初代培養筋細胞に - 69 - 麹菌はリパーゼを産生・分泌しており、原料 由来のトリグリセライドから遊離脂肪酸を生 が見出された(表2)。筋組織のカルニチンパ 成し、菌体内に取り込み、エネルギー源として ルミトイルトランスフェラーゼ(CPT1A)の mRNA 利用、あるいはトリグリセライドに再合成して 発現量が Asp.awamori 投与区では半減しており 貯蔵する 28) 。焼酎醸造における脂肪酸エチルの (P<0.05)(図2A)、脂肪酸β酸化系の抑制が 生成には、麹菌リパーゼが関与し、原料由来の 示唆された。一方で、脂肪酸合成に関わるアセ 脂肪からの遊離脂肪酸の生成と、その脂肪酸と チル CoA カルボキシラーゼ(ACC)および脂肪 エタノールから逆合成反応により脂肪酸エチ 酸合成酵素(FAS)の mRNA 発現量は増加してい ルが生成される 29) 。当初、ブロイラーにおいて た(図2B、C)。血漿のトリグリセライドおよ は、麹菌リパーゼによる脂肪の消化吸収率の向 び総コレステロールの各濃度は Asp.awamori 投 上が期待された。そこで、脂質代謝に対する 与区で低い値を示した(P<0.05)。対して、HDL Asp.awamori の飼料投与の影響を調べるための コレステロール濃度は Asp.awamori 投与区で上 実験を行った 30) 昇し(P<0.05)(表2)、Asp.awamori のコレス 。 試験1と同様の飼養試験を行い、血液および テロール合成系への影響が示唆された。 組織サンプルを得て分析に用いた。血液の脂質 Asp.oryzae はコレステロール生合成の律速酵 性状は、富士ドライケム 3500(Fuji Medical 素 3- ヒ ド ロ キ シ -3- メ チ ル グ ル タ リ ル -CoA Systems)を用いて調べた。浅胸筋組織より、 (HMG-CoA)レダクターゼの阻害物質を生成す Folch 法に従って脂質を抽出した 31) 。脂肪酸メ る 32) 。また、Penicillium citrinum から発見 チルエステルの調製は、脂質試料を水酸化ナト された HMG-CoA レダクターゼ阻害剤スタチンの リウム含有メタノール存在下でアルカリ加水 誘導体は高コレステロール血症の治療薬とし 分解した後、三ふっ化ほう素メタノール錯体を て臨床応用されている 用いて行った。脂肪酸メチルエステルをヘキサ Asp.awamori が HMG-CoA レダクターゼ阻害物質 ンで抽出し、ガスクロマトグラフ質量分析装置 を生成する可能性がある。 33 ) 。これより 2.5 mm 内 筋組織の脂肪酸は Asp.awamori の飼料投与 径 DB-coated, J & W Scientific) ] によっ によって、飽和脂肪酸パルミチン酸(C16)が て、脂肪酸の分離・同定を行った。浅胸筋は脂 減少し(P<0.05)、一方で不飽和脂肪酸オレイ 質代謝関連因子の遺伝子発現量の測定にも用 ン酸(C18:1)は 0.2%区で、リノール酸(C18:2) いた。 は両投与区で、リノレン酸(C18:3)は 0.05% [Thermo Fisher+分析カラム(30 m 興味深い事に、Asp.awamori の飼料投与によ 区で、アラキドン酸(C20:4)は 0.2%区でそれ って、腹腔内脂肪重量は減少し(P<0.05)、反 ぞれ増加し(P<0.05)、 全体的な不飽和脂肪酸 対に筋組織の総脂質が増加する(P<0.05)現象 の割合が増えていた(図2D∼H)。Aspergillus 表2.Aspergillus awamori の飼料投与がブロイラーヒナの腹腔脂肪重量、筋組織総脂質含量および血液 の脂質性状に及ぼす影響(試験2) Aspergillus awamori !"#$% &'% ()*+,- (% body weight) ./01*2 (g/100 g muscle) 1.5 3.0 0.05% 0.2 a ± 0.2 b a ± 34546789 (mg/dL) 29 ± 2 1:;<=>?@ (mg/dL) 145 ± 3a ± b HDL:;<=>?@ (mg/dL) 80 3 0.75 ± 0.09 ± 0.08b 1.2 7.0 ± 1.0a 17 ± 3 b 21 ± 4b 109 ± 2c 128 ± 7b ± a 95 ± 4a 90 3 a 0.8 ± 6.0 ABCDEFGHIn=10JKLMNOPQRSTUHVWIP < 0.05JKIPX 30 YWZ[\]J - 70 - 0.2% b !" ! % " ' # $ & 図2.Aspergillus awamori のブロイラーヒナへ の飼料投与が骨格筋の脂肪酸代謝関連酵素の遺 伝子発現量および脂肪酸組成に及ぼす影響(試 験2) 平均値 標準誤差(n=10).異なる文字間に有意 ( 差 あ り ( P < 0.05 ). CPT1A, Carnitine palmitoyltransferase 1A; FAS, Fatty acid synthase; ACC, Acetyl-CoA carboxylase.(文 献 30 より改変引用) 属はΔ9 およびΔ12 位脂肪酸不飽和化酵素を発 現しており 飽和化が起こり、末梢蓄積脂肪の脂肪酸組成へ 34,35) 、不飽和脂肪酸を高生産する Asp.oryzae 株も報告されている 影響を及ぼすと考えられる。以上の要因により、 36) 。その際、 脂肪酸の不飽和化が促進されると推察した。こ 菌体内に C16 やステアリン酸(C18)等の飽和 れらの腹腔内脂肪の減少や脂肪酸の不飽和化 脂肪酸が取り込まれ、鎖長延長および不飽和化 の促進等の脂質代謝に対する Asp.awamori の効 された後、トリグリセライドとなり、菌体外へ 果は、ラットにおいても類似の結果が得られて 分泌されることが示されている 37) おり 38)、ニワトリに限定されたものではない。 。そのためニ ワトリ消化管において飼料由来の脂肪酸の不 - 71 - 4.Aspergillus awamori の骨格筋タンパク質代 ーム C2 サブユニット、および m-カルパイン大 謝および脂質代謝への効果は酵母との同時 サブユニット)の mRNA 発現量は、全ての投与 投与によって増幅される(試験3) 区で減少した(図3A∼D)。反対に、ミオシン 焼酎の醸造工程では製麹に次いで、麹に水お およびアクチンの mRNA 発現量は、Asp.awamori よび酵母(Saccharomyces.cerevisiae )が混合 および Sacch.cerevisiae 同時投与によって相 され一次仕込みに移る。酵母が増殖したところ 乗的(P<0.05)に増加した(図3E、F)。腹腔 で主原料であるサツマイモが投入され、引き続 内脂肪重量および血漿トリグリセライド濃度 きデンプンの糖化・発酵が進行する(二次仕込 は Asp.awamori 単独あるいは Sacch.cerevisiae 1) み) 。焼酎醸造においては Sacch.cerevisiae との同時投与によって減少したが、 の中でもクエン酸耐性を持つ株(焼酎酵母)が Sacch.cerevisiae 単独では効果が無かった。一 4) 、Asp.awamori および焼酎酵 方で筋組織の脂質含量は、全ての投与区で増加 母はそれぞれの特性を生かして、あるいは相補 した(表3)。筋組織の脂肪酸組成は、飽和脂 しながら、効率よく原料デンプンを分解・発酵 肪酸含量(C16 および C18)は減少し、対して している。耐酸性株酵母は低 pH なニワトリ消 不飽和脂肪酸含量(C18:1、C18:2、C18:3、お 化管上部で減衰することなく、下部に到達し、 よ び C20:4 ) は 増 加 し 、 C18:2 を 除 い て 発酵を行える可能性がある。また、 Asp.awamori および Sacch.cerevisiae の相乗効 Sacch.cerevisiae はカビ毒アフラトキシンの 果(P<0.05)が確認された(表3)。この脂肪 影響を抑制する飼料添加物としても有望であ 酸組成の変化は、Δ6脂肪酸不飽和化酵素の遺 用いられており 39,40) 。以上のような理由より、Asp.awamori 伝子発現量の変化と部分的に関連していた(図 単独よりも Sacch.cerevisiae との組合せに相 3 G )。 以 上 よ り 、 Asp.awamori お よ び 乗効果が期待された。そこで Asp.awamori およ Sacch.cerevisiae の同時飼料投与は、骨格筋タ び Sacch.cerevisiae の同時飼料投与が、ブロ ンパク質代謝の促進を介して、飼養成績を相乗 イラーの成長ならびに骨格筋のタンパク質代 的に改善することが明らかとなった。加えて、 謝および脂肪酸組成の特性に及ぼす影響を調 Asp.awamori および Sacch.cerevisiae は筋組織 る べた 41) 。 の脂肪酸組成の特性に変化をもたらすことが 試験1および試験2と同様に、15 日齢のヒナ 示された。 を、対照区(基礎飼料)、同飼料に Asp.awamori を 5.Aspergillus awamori の抗酸化能と暑熱スト 0.05 % 添 加 し て 投 与 す る 区 、 Sacch.cerevisiae を 0.10%添加して投与する レスの抑制への応用(試験4) 区、および両者を同時投与する区に分け(7 羽/ 麹菌はフェルラ酸などの抗酸化物質を生成 区)、12 日間の飼養試験を行った。測定項目お し、発酵生産物に高い抗酸化能が付与される 42)。 よび方法等は試験1および試験2に準じた。 これまでに、Asp.awamori の飼料投与によって、 ヒナの成長は全ての投与区で促進され、増体 筋組織において、過酸化脂質より生成されるマ 量、飼料要求率、浅胸筋重量、および飼料のタ ロンジアルデヒドの減少、一方で抗酸化ビタミ ンパク質消化率において Asp.awamori および ンα-トコフェロールの増加が見出されている。 Sacch.cerevisiae 同 時 投 与 に よ る 相 乗 効 果 麹菌由来の抗酸化物質が、組織での過酸化脂質 (P<0.05)が観察された。血漿 3-メチルヒスチ の形成を抑制するためα-トコフェロールの消 ジン濃度は Asp.awamori と Sacch.cerevisiae 費が減少すると考えられる。 の同時投与により相乗的(P<0.01)に減少した ブロイラー生産における暑熱環境ストレス (表3)。筋組織におけるタンパク質分解関連 の影響は、増体量の抑制から熱死の発生にまで 因子(アトロジン-1、ユビキチン、プロテアソ 及び、生産性の低下を引き起こす 43,44)。南九州 - 72 - 表3.Aspergillus awamori および Saccharomyces cerevisiae の同時飼料投与がブロイラーヒナの飼養成 績、飼料の消化率、血液の脂質性状および 3-メチルヒスチジン濃度、ならびに筋組織の脂肪酸組成に及 !"#Aspergillus awamori $%& Saccharomyces cerevisiae '()*+,-./012345'*6789*+':;<9=>'?@AB$ ぼす影響(試験3) %& 3-CDE4FDGHIJ9KL&MNOP'?QRO7MSTUVWXYZ"[ !"# '(# *+, B[\ SC AA+SC AA! $%$& SC! AA)SC 742 1437 1.93 59 66 22.0 1.22 ± ± ± ± ± ± ± 10c 15a 0.08a 3c 3b 0.3b 0.09a 911 1362 1.49 69 77 24.4 0.96 ± ± ± ± ± ± ± 11b 12b 0.04b 3b 4a 0.4a 0.02b 895 1337 1.49 65 76 23.2 1.09 ± ± ± ± ± ± ± 9b 12b 0.03b 5ab 4ab 0.4ab 0.03ab 977 1286 1.31 73 79 26.3 0.71 ± ± ± ± ± ± ± 12a 13c 0.02c 5a 3a 0.6a 0.02c * * * * * * * * * * NS NS NS NS ** ** * * NS * ** JKLKMNOP (mg/dL) QRSTUV?= (mg/dL) HDL RSTUV?= (mg/dL) 3-WX=YTXZ5 (!mol/mL) 33 137 78 29 ± ± ± ± 1.2a 6a 5c 4a 29 105 91 21 ± ± ± ± 0.4b 5bc 7b 3b 30 118 83 19 ± ± ± ± 0.7ab 3b 2b 2b 26 100 94 16 ± ± ± ± 1.4c 3c 5a 2c * * * * NS * * * * * * ** QF8 (%) FG] (mg/100 mg fat) 16:0 18:0 18:1 (n-9) 18:2 (n-6) 18:3 (n-3) 20:4 (n-6) 2.4 ± 0.9b 5.7 ± 0.2a * * * * * * * * * * * * * * * * ** * NS ** * (g/12 d) (g/12 d) -.123 (gain/feed) 456789:3 (%) ;<=>?9:3 (%) @ABC, (% body weight) DEFGC, (% body weight) -./0, HI AA 1.41 0.132 0.90 1.0 0.041 0.51 ± ± ± ± ± ± 0.02a 0.04a 0.06c 0.1b 0.002c 0.03c 4.6 ± 0.3a 1.01 0.082 1.62 2.4 0.063 0.99 ± ± ± ± ± ± 0.04b 0.01b 0.10b 0.1a 0.004b 0.04b 4.1 ± 0.2a 1.19 0.089 1.51 2.2 0.067 1.22 ± ± ± ± ± ± 0.02b 0.02b 0.10b 0.2a 0.003ab 0.14ab 0.98 0.032 1.76 2.4 0.072 1.35 ± ± ± ± ± ± 0.04c 0.04c 0.10a 0.2a 0.004a 0.15a ^_`abcdefn=7ghijklmnopqrestfP < 0.05gh$%$&uNS, not significant ( P > 0.05); * P < 0.05; ** P < 0.01. AA, Aspergillus awamori (0.05%); SC, Saccharomyces cerevisiae (0.1%).fmv 41 wtxyz{g 3 地域はブロイラーの生産が盛んであるが、夏季 進を目的に飼料添加されている既存の抗生物 の酷暑時にはその生産性は大きく低下し、効果 質は、今後使用が厳しく規制されると予想され 的な対処法の開発が望まれている。暑熱環境下 45,46) では、細胞内の活性酸素の産生が高まるととも また、脂肪酸不飽和化の促進およびコレステロ に、生体の抗酸化システムが十分に機能してい ール低減効果を応用して、脂質性状に特徴を持 ないと予測される。そこで、現在 Asp.awamori たせた高付加価値な鶏肉の作出が可能となる。 の抗酸化効果を暑熱ストレスの低減へ応用す 商品価値がほとんどない腹腔内脂肪が減少す ることを目指して、暑熱ストレスを負荷した雄 る事も Asp.awamori を使用するメリットとなる。 ブロイラーヒナの成長ならびに抗酸化状態に これら成長および肉質に対する効果は、酵母 及ぼす影響を調べている。その結果、 Sacch.cerevisiae との組合せによって増強さ Asp.awamori の飼料投与は、暑熱ストレスによ れる。そして、生体内の抗酸化系の賦活効果は、 って減弱した生体の抗酸化状態を改善するこ 夏季の暑熱環境ストレスによる生産性の低下 とを示すデータが得られつつある。 や鶏肉の品質劣化の防止に有効かもしれない。 以上より Asp.awamori の飼料投与によって、家 6.おわりに 畜の生産性の向上 および 食肉の品質向上が図 、Asp.awamori の代替利用が期待される。 Aspergillus awamori には多様な栄養生理機 られることが期待される。しかし、これらの栄 能が示され、新しい飼料添加物としての応用が 養生理機能に関する研究は端緒についたばか 期待される。飼料の消化率の向上および骨格筋 りで、原因物質はまだ2つしか特定されておら タンパク質分解系の抑制による成長促進ある ず、作用機序の詳細には多くの謎が残されてい いは筋量増加は飼料の節約につながる。成長促 る。今後は、理論的裏付けを行うための更なる - 73 - !" %&! &-.)12,34 #$%& '( )* +),-.)/0 $! #! "! " " " !" 5"2672-2, #$%& '( )* +),-.)/0 $! #! "! $! #! " %&! "! " =>=?@ #$%& '( )* +),-.)/0 8.)-9!:)#9 #$%& '( )* +),-.)/0 #! !" " $! ' " ! $! #! ! %#! $ ! " "! ! %&! ! %'! !" ! " + ! %(! % ! %&! ! "! ! %&! & %'! ;<):2, #$%& '( )* +),-.)/0 %(! # ! &+-2, #$%& '( )* +),-.)/0 ! ! %&! " ! " ! " " ! ! " " " ! " ( + (! )! ! %&! ! ) ! #3A!/B!2, #$%& '( )* +),-.)/0 " $! " !" #$#%&'( )" *+'+,(-(#+ " ! ! " " #! "! 図3.Aspergillus awamori および Saccharomyces ! !" #$#%&'( )" *+'+,(-(#+ ! ! " ! ! " cerevisiae のブロイラーヒナへの同時飼料投与が " " 骨格筋タンパク質分解および合成関連因子、なら びにΔ6位脂肪酸不飽和化酵素の遺伝子発現量 に及ぼす影響(試験3) 平均値 標準誤差(n=7).異なる文字間に有意差あり(P < 0.05). D6DES, Δ6-Fatty acid desaturase. (文献 41 より改変引用) 研究が必要である。 南九州という温暖な気候の地域での焼酎醸造 酒、味噌、醤油のような発酵食品は麹なしに に特化されてきた。今後は 焼酎 の枠を飛び はあり得ず、それらは古来より日本の食文化と 越え、 飼料 広く深く関わり、我々の食生活を豊かなものに 菌 Asp.awamori の活躍を期待したい。 してきた。現在利用されている醸造用麹菌は、 日本人が長年にわたり系統選抜を繰り返して 確立した栽培品種、すなわち微生物の家畜であ る 47) 。なかでも Asp.awamori はその特性より、 - 74 - という新たな広大な領域での麹 【引用文献】 A, Tomita Y. 1996b. Separation of growth 1) 小泉武夫 2012.発酵食品学.85-86.講 promoting factor of broiler chicken from 談社.東京. shochu distillery by-product. Jpn. J. 2) Amsal A, Takigami M, Ito H. 1999. Increased digestibility of raw starches Poult. Sci., 33: 96-103. 11) Mahfudz LD, Nakashima K, Ohtsuka A, by mutant strains of Aspergillus awamori. Hayashi K. 1997. Growth factors for a Food Sci. Technol. Res., 5: 153-155. primary chick muscle cell culture from 3) Hajati H. 2010. Effects of enzyme shochu distillery by-product. Biosci. supplementation on performance, carcass Biotechnol. Biochem., 11: 1844-1847. characteristics, carcass composition 12) Ohtsuka A, Otsuji Y, Hayashi K. 1998. and some blood parameters of broiler Plasma α-tocopherol of broiler chicken chicken. Am. J. Anim. Vet. Sci., 5: is increased by shochu distillery 221-227. by-product. Jpn. J. Poult. Sci., 35: 4) 高峯和則・瀬戸口真治・亀澤浩幸・神渡巧・ 緒方新一郎・尾ノ上国昭・濱崎幸男 1994. 132-137. 13) Kamizono T, Ohtsuka A, Hashimoto F, 焼酎酵母の分離に関する研究.鹿児島県工 Hayashi K. 2013. Dibutoxybutane 業技術センター報告, 8: 1-6. suppresses protein degradation and 5) 神渡巧・瀬戸口眞治・高峯和則・緒方新一 promotes growth in cultured chicken 郎 2005.ストレスを受けた焼酎原料サツ muscle cells. J. Poult. Sci., 50: 37-43. マイモのモノテルペンアルコール含量と 14) Yamamoto M, Saleh F, Ohtsuka A, Hayashi 芋焼酎の香気特性.日本醸造協会報, 100: K. 2005. New fermentation technique to 520-526. process fish waste. Anim. Sci. J., 76: 6) 高峯和則・吉﨑由美子・島田翔吾・髙屋総 245-248. 一郎・玉置尚徳・伊藤清・鮫島吉廣 2011. 15) Yamamoto M, Saleh F, Tahir M, Ohtsuka A, 芋焼酎に検出されたローズオキサイドの Hayashi K. 2007. The effect of Koji-feed 生成機構.日本醸造学会報, 106: 50-57. (fermented distillery by-product) on 7) 林國興・大谷敬亨・上野倫睦・濱元優・大 the growth performance and nutrient 塚彰・吉永和美・中川一朗 2003.乳牛飼 metabolizability in broiler. J. Poult. 料ならびにサイレージ添加物としての固 Sci., 44: 291-296. 液分離甘藷焼酎粕の利用.西日畜会報, 46: 16) Kamizono T, Nakashima K, Ohtsuka A, 35-37. Hayashi K. 2010. Effects of feeding 8) 林國興・前田真希・北原和弥・田子山徹・ hexane-extracts of a shochu distillery 大塚彰 2009.焼酎粕上清濃縮液給与によ by-product on skeletal muscle protein るブロイラーの生産性および肉質の改善. degradation in broiler chicken. Biosci. 日畜会報, 80: 35-39. Biotechnol. Biochem., 74: 92-95. 9) Mahfudz LD, Hayashi K, Ikeda M, Hamada 17) Saleh AA, Eid YZ, Ebeid TA, Kamizono T, K, Ohtsuka A, Tomita Y. 1996a. The Ohtsuka A, Hayashi K. 2011. Effects of effective use of shochu distillery feeding by-product as a source of broiler feed. Aspergillus niger on growth performance Jpn. J. Poult. Sci., 33: 1-7. and meat quality in broiler chickens. J. 10) Mahfudz LD, Hayashi K, Otsuji Y, Ohtsuka - 75 - Aspergillus awamori Poult. Sci., 48: 201-206. and 18) Saleh AA, Eid YZ, Ebeid TA, Ohtsuka A, Y, Inoue K, Hashizume T. 1985. The rate Yamamoto M, Hayashi K. 2012a. Feeding of degradation of myofibrillar proteins Aspergillus awamori reduces skeletal of skeletal muscle in broiler and layer muscle protein breakdown and stimulates chickens growth in broiler. Anim. Sci. J., 83: -methylhistidine in excreta. Br. J. 594-598. Nutr., 54: 157-163. 19) Hayashi K, Maeda Y, Toyomizu M, Tomita Y. 1987. Highperformance estimated by N τ 26) Nakashima K, Ishida A, Katsumata M. 2009. liquid Comparison of proteolytic-related gene chromatographic method for the analysis expression in the skeletal muscles of τ of N -methylhistidine in food, chicken layer and broiler chickens. Biosci. excreta, and rat urine. J. Nutr. Sci. Biotechnol. Biochem., 73: 1869-1871. Vitaminol., 33: 151-156. 27) Ohtsuka A, Kawatomi N, Nakashima K, 20) Saleh F, Ohtsuka A, Tanaka T, Hayashi K. Araki T, Hayashi K. 2011. Gene 2003. Effect of enzymes of microbial expression of muscle-specific ubiquitin origin on in vitro digestibilities of ligase, atrogin-1/MAFbx, positively dry matter and crude protein in maize. correlates J. Poult. Sci., 40: 274-281. proteolysis in food-deprived broiler 21) Tahir M, Saleh F, Ohtsuka A, Hayashi K. 2005. Synergistic effect of cellulase and hemicellulase on skeletal muscle chickens. J. Poult. Sci., 48: 92-96. 28) 戸井田仁一 2011.麹菌リパーゼ高生産株 nutrient の 育 種 と み そ の 試 験 醸 造 . 醸 協 , 106: utilization and performance in broilers fed a corn-soybean meal diet. Anim. Sci. with 457-461. 29) 工藤哲三・水谷政美・本部恭平・太田一良 J., 76: 559-565. 2001.Aspergillus kawachii の脂質代謝 22) Saleh F, Tahir M, Ohtsuka A, Hayashi K. 関連酵素に関する検宮崎県工業技術セン 2005. A mixture of pure cellulase, ター・宮崎県食品開発センター研究報, 46: hemicellulase and pectinase improves 159-164. broiler performance. Br. Poult. Sci., 30) Saleh AA, Eid YZ, Ebeid TA, Ohtsuka A, 45: 602-606. Hioki K, Yamamoto M, Hayashi K. 2012b. 23) Hong KJ, Lee CH, Kim SW. 2004. The modification of the muscle fatty Aspergillus oryzae GB-107 fermentation acid profile by dietary supplementation improves nutritional quality of food with Aspergillus awamori in broiler soybeans and feed soybean meals. J. Med. chickens. Br. J. Nutr., 108: 1596-1602. Food, 7: 430-435. 31) Folch J, Lees M, Stanley GHS. 1957. A 24) Sacheck JM, Ohtsuka A, MacLary CS, simple method for the isolation and Goldberg AL. 2004. IGF-1 stimulates purification of total lipides from muscle growth by suppressing protein animal tissues. J. Biol. Chem., 226: breakdown 497-509. and atrophy-related expression of ubiquitin-ligases, 32) Hajjaj H, Duboc P, Fay LB, Zbinden I, Mace 287: E591-E601. Aspergillus oryzae produces compounds 25) Hayashi K, Tomita Y, Maeda Y, Shinagawa - 76 - K, Niederberger P. 2005. atrogin-1 and MuRF1. Am. J. Physiol., inhibiting cholesterol biosynthesis downstream of dihydrolanosterol. FEMS Microbiol. Lett., 242: 155-159. feedstuffs. Poult. Sci., 92: 1655-1663. 41) Saleh AA, Hayashi K, Ohtsuka A. 2013b. 33) Endo A, Kuroda M, Tsujita Y. 1976. Synergistic effect of feeding ML-236A, ML-236B, and ML-236C, new Aspergillus awamori and Saccharomyces inhibitors cholesterogenesis cerevisiae on growth performance in produced by Penicillium citrinium. J. broiler chickens; promotion in protein Antibiot., 29: 1346-1348. metabolism and modification in fatty of 34) Wilson RA, Chang PK, Dobrzyn A, Ntambi acid profile in the muscle. J. Poult. JM, Zarnowski R, Keller NP. 2004a. Two D9-stearic acid desaturases Sci., 50: 242-250. are 42) 石原昌信・根路銘美穂・平良東紀・多和田 required for Aspergillus nidulans 真吉・小波本直忠 2002.泡盛醸造におけ growth and development. Fungal Genet. る原料米からのフェルラ酸の遊離.生物工 Biol., 41: 501-509. 学会誌, 80: 563-567. 35) Wilson RA, Calvo AM, Chang PK, Keller NP. 43) Yunianto VD, Hayashi K, Kaneda S, the Ohtsuka A, Tomita T. 1997. Effect of Aspergillus parasiticus D -desaturase environmental temperature on muscle gene: a role for lipid metabolism in the protein turnover and heat production in Aspergillus - seed interaction. tube-fed broiler chickens. Br. J. Nutr., Microbiol., 150: 2881-2888 77: 897-909. 2004b. Characterization of 12 36) 森 哲 也 ・ 佐 藤 望 ・ 角 田 潔 和 ・ 小 泉 武 夫 44) Azad KMA, Kikusato M, Hoque AM, Toyomizu 2004a.不飽和脂肪酸を高生産する糸状菌 M. 2010. Effect of chronic heat stress 〔Aspergillus oryzae〕の変異株取得とそ on performance and oxidative damage in の生産特性.日食品保蔵誌, 30: 77-80. different strains of chickens. J. Poult. 37) 森哲也・金内誠・進藤斉・角田潔和・吉澤 Sci., 47: 333-337. 淑・小泉武夫 2004b.中国ハム(金華火 45) Mølbak K, Baggesen DL, Aarestrup FM, 腿)脂質の不飽和化に関与する糸状菌の同 Ebbesen JM, Engberg J, Frydendahl K, 定と性質.日食品保蔵誌, 30: 9-15. Gerner-Smidt P, Petersen AM, Wegener HC. 38) Saleh AA, Ohtsuka A, Yamamoto M, Hayashi 1999. An outbreak of multidrug- K. 2013a. Aspergillus awamori feeding resistant, quinolone-resistant modifies lipid metabolism in rats. Salmonella enterica serotype Biomed Res. Int., online. typhimurium DT104. N. Engl. J. Med., 39) Stanley VG, Ojo R, Woldesenbet S, 341: 1420-1425. Hutchinson DH. 1993. The use of 46) Mølbak K. 2004. Spread of resistant Saccharomyces cerevisiae to suppress bacteria and resistance genes from the effects of aflatoxicosis in broiler animals to humans - The public health chicks. Poult. Sci., 72: 1867-1872. consequences. J. Vet. Med., B51: 40) Pizzolitto RP, Armando MR, Salvano MA, Dalcero AM, Rosa CA. 2012. Evaluation of Saccharomyces cerevisiae as 364-369. 47) 北本勝ひこ 2012.麹菌物語.生物工学, an antiaflatoxicogenic agent in broiler - 77 - 90: 424-42 栄養生理研究会報 Vol. 57, No. 2 2013 周年放牧肥育牛肉の生産技術と肉質特性 中村好德 ((独)農業・食品産業技術総合研究機構 九州沖縄農業研究センター) 1.はじめに 環境要因や放牧条件と、増体量に関する結果で わが国の肉用牛生産は、畜舎でウシを飼養し ある。積雪がほとんどない低標高の西南暖地で 海外からの穀物飼料に過度に依存するという は牧草の周年栽培が可能である。本研究は九州 特徴を有する。従って、本来は草食反芻動物で 沖縄農業研究センターの試験圃場(熊本県合志 あるウシが、少量の粗飼料と多量の穀物飼料に 市、緯度:32.88°、経度:130.74°、標高: よって生涯飼養される。近年のわが国の畜産業 85 m)で行われ、周年放牧肥育に要した放牧地 全体において、純国内産飼料自給率(可消化養 の面積(4 牧区の合計)は 38 a/頭であり、1 牧 分総量:TDN ベース)は 26%であり、大家畜経 区あたり 6〜12 a/頭(平均 9.5 a/頭)で飼養 営(肉用牛;肉専肥育)では、飼料自家生産の された(図 1)。また、本研究のような集約的な 割合および粗飼料給与率 (いずれも TDN ベース) 放牧飼養でも、補助飼料を効果的に用いること 1) は、それぞれ 1.7%および 10.8%である 。こ により草地荒廃を防ぎながら周年放牧肥育が のような脆弱な飼料生産基盤のもとに成り立 可能である。図 2 は補助飼料を飽食で維持させ つ肉用牛生産方法は、穀物需給や飼料価格の変 た場合の放牧草の草高(入牧時)と補助飼料の 動などの影響を受けやすいと考えられる。一方、 摂取量との関係を示したが、夏牧草(パリセー 生産される牛肉については、専ら筋肉中の脂肪 ドグラス)と冬牧草(イタリアンライグラス) 含量を高めることに改良の力点を置き、食品と の草高と補助飼料摂取量には負の相関 しての栄養学的な価値を高めることは追究さ (P<0.05)が見られた。 れていない。これは牛肉市場において枝肉単価 図 3 はイタリアンライグラス放牧期間中の飼 に及ぼす影響が最も大きい格付項目が、脂肪交 料構成と飼料摂取量を示した。その結果、生草 雑等級(BMS No.)であること 2)に起因するもの の乾物摂取量は、4.3〜9.1 kg/頭/日と変動が と考えられる。 激しかったが、平均 6.6 kg/頭/日を摂取した場 筆者らが提唱する“周年放牧肥育”とは、粗 合の生草からの粗蛋白質および可消化養分総 3) 飼料の利用性に優れる褐毛和種 を用いて、肥 量の摂取割合は、それぞれ 47.8%および 47.1% 育素牛で導入後、配合飼料無給与で周年放牧に であった。表 1 は給与飼料の中でも、特に牧草 トウモロコシサイレージなどの国産飼料を併 成分の季節変動と特徴について示したが、本研 給することにより、体重 700 kg 前後の出荷を 究で用いた各種牧草の栄養成分は著しい季節 可能にする新しい肉用牛生産方法である 4,5) 。 変動を示し、牧草間でも夏牧草であるバヒアグ 本稿では周年放牧肥育技術の概要と肉質特性 ラス、ヒエやパリセードグラス(粗蛋白質含 について述べる。 量:8.1〜17.9%、粗繊維含量:20.5〜39.0% ならびに可消化養分総量:46.9〜55.6%)は、 2.生産技術の特徴 冬牧草であり周年放牧肥育技術の基幹草種で 4,6〜8) あるイタリアンライグラス(12.2〜24.8%、 により、生産方法の体系化が行われた。図 1 は 14.7〜28.8%ならびに 55.1〜65.6%)と比べて 周年放牧肥育技術に関する一連の研究 A beef production system and meat quality using year-round outdoor grazing and self-sufficient feed Yoshi-nori Nakamura (NARO Kyushu Okinawa Agricultural Research Center) - 79 - 図2.放牧草の草高と補助飼料摂取量の関係 補助飼料は飽食を維持。入牧時の草高を測定し、 次の草高測定時までの補助飼料摂取量:乾物 kg/5頭合計/日を測定。 (文献 8 の図2を一部 改変) 図1.環境要因、放牧条件ならびに周年放牧 肥育牛(3頭平均値)の増体曲線 (文献 8 の図1を一部改変) 図3.冬期放牧90日間(2012 年 2 月から 4 月) の飼料構成 5頭平均値。CSはトウモロコシサイレージ、 麦液は麦焼酎粕濃縮液を示す。 (文献 8 の図3 を一部改変) - 80 - も低蛋白質・高繊維質であり、繊維の中でも低 する必要があるのかもしれない。 消化性繊維含量が高い傾向にあった。従って、 周年放牧肥育技術を用いて飼養された褐毛 播種時期や施肥条件などを改善しても牧草成 和種去勢雄牛の牛枝肉格付評価および産肉量 分の季節変動は避けられず、周年放牧肥育で年 を表 3 に示した。(公)日本食肉格付協会によ 間を通じて高い増体量を維持させるためには る枝肉評価の結果、周年放牧肥育牛の肉質等級 何らかの補助飼料が必要であると考えられた。 は 2 等級のみであった。特に周年放牧肥育牛の 周年放牧肥育では、牧草成分の季節変動を補 枝肉評価の特徴として、BMS No.が低く、BCS No. う目的として国産飼料に限定して選定した結 と BFS No.が高いことが示唆された。 果、トウモロコシサイレージを用いた。しかし、 基幹草種であるイタリアンライグラスとの組 3.肉質の特徴 み合わせの効果を期待できるのは、冬期の一部 周年放牧肥育牛肉の肉質の特徴について表 4 の期間(特に粗蛋白質含量が高い 12 月、2 月〜 〜6 に示した。表 4 および表 5 の項目について 4 月) であることが明らかになったため (表 1) 、 は、日本食品標準成分表 201014)および五訂増補 九州地方で特有の食品副産物である麦焼酎粕 日本食品標準成分表脂肪酸成分表編 200515)に 濃縮液 9,10) (原材料は大麦)を補助飼料として 記載されている和牛肉(黒毛和種去勢雄牛の A3 追加した。トウモロコシサイレージと麦焼酎粕 および A4 等級の格付のもの)、乳用肥育牛肉(20 濃縮液を原物重量比で 2:1(飼料成分は乾物% ヵ月齢まで肥育したホルスタイン種去勢雄牛 で、粗蛋白質含量が 23%ならびに可消化養分総 の B2 等級の格付のもの)ならびに輸入牛肉(オ 量が 71%に設定)に混合させて給与した。 ーストラリアおよびニュージーランドで穀物 周年放牧期間中の褐毛和種去勢雄牛の血液 を主体として飼養された牛の標準的な品質の 性状の変化(代謝プロファイルテスト)を表 2 に示した。代謝プロファイルテスト もの)と比較した。 11) とは、飼 主要な部分肉 8 部位の五成分(水分、たんぱ 料から摂取した栄養成分の過不足を血液性状 く質、脂質、炭水化物ならびに灰分)およびエ から客観的に診断・評価するものであるが、放 ネルギー含量の結果であるが、特に脂質含量か 牧牛における報告は少なく、特に周年放牧肥育 ら判断すると周年放牧肥育牛肉の 8 部位中 7 部 牛については報告されていない。しかし、前報 位(かた、かたロース、サーロイン、もも、そ 4,7,12) と本研究の結果から判断する限り、周年 ともも、ランプならびにヒレ)が、和牛肉と乳 放牧肥育牛の血液性状は季節による変動はあ 用肥育牛肉の間に位置した。また、エネルギー るが、概ね正常値範囲内 13) の推移であると考え については 8 部位とも和牛肉より低い傾向にあ られ、健康状態に異常はなかったと判断した。 った(表 4)。リブロースの無機質(13 項目) 、 これは周年放牧肥育における一連の研究にお ビタミン(14 項目)、脂肪酸(4 項目) 、コレス いて、周年放牧肥育牛の代謝疾患の発症がほと テロール含量ならびに食塩相当量について調 んど見られなかったことからも裏付けられた。 査した結果を表 5 に示した。その結果、日本食 一方、増体の指標として経時的モニタリングを 品標準成分表 201014)に記載される和牛肉、乳用 行った血漿中尿素窒素濃度は、6.7〜22.8 mg/dL 肥育牛肉ならびに輸入牛肉と比べて、大差が見 と変動が激しく(表 2) 、目標値(文献 4 および られた項目は無機質(セレン) 、ビタミン(β- 12 より設定)である 15 mg/dL を下回る時期も カロテン、B12)、脂肪酸(n-6/n-3 比)ならびに 見られた(表 2) 。周年放牧肥育牛の蛋白質やエ コレステロール含量であった。特に周年放牧肥 ネルギー摂取量の季節変動については未解明 育牛肉のβ-カロテン含量は、和牛肉などが 0 の部分が多いため、今後、牧草成分の季節変動 〜4 μg/100 g であるのに対して、11 μg/100 g に合わせた補助飼料の種類や給与方法を検討 と高い傾向にあった。 β-カロテンはサイレー - 81 - 表1.給与飼料成分の特徴(文献 7 の表 2 を一部改変) - 82 - 表2.周年放牧期間中の褐毛和種去勢雄牛の血液性状の変化(文献 7 の表 3 を一部改変) 表3.周年放牧肥育牛の牛枝肉格付評価および産肉量 (文献 7 の表 4 および文献 8 の表 3 を一部改変) ジや配合飼料よりも生草に多く含まれる 4)ため 26 万頭)結果から選定]ならびにホルスタイン に、放牧を取り入れた肉用牛生産方法では筋肉 種去勢牛で肉質等級 2 等級のデータ(n=40)[乳 8) 用牛去勢 2 等級割合 93.4%(最多) ;平成 23 年 へのβ-カロテンの蓄積による外観の黄色化 は特徴の一つになると考えられた。 度 1 月〜12 月集計(約 22 万頭)結果から選定] 日本農林規格登録認定機関である(株)日本 を用いた。一方、輸入牛肉は豪州産牛肉のデー 認証サービスが保有する肉質データベースを タ(n=60)を用いたが品種、性別や飼養条件な 用いて、本研究で生産した周年放牧肥育牛肉 どは不明であった。その結果、周年放牧肥育牛 (胸最長筋:リブロース部分)の品質について、 肉とその他の牛肉の相違点は次のようにまと 和牛肉、乳用肥育牛肉ならびに輸入牛肉に対す められた。①周年放牧肥育牛肉は和牛肉と比べ る対比評価を行った(表 6) 。比較対照とする牛 て、遊離アミノ酸総量やペプチド構成アミノ酸 16) 肉は農畜産業振興機構編(2011) 、日本食品 総量などの旨味成分含量が多く、味わいのある 14) 標準成分表 2010 ならびに(公)日本食肉格付 肉質であり、食感も軟らかく噛み応えも低い。 協会の牛枝肉格付結果(集計表)を参考にして しかし、加熱損失率が高く、圧搾肉汁率が低い 選定し、和牛肉は黒毛和種去勢牛で肉質等級 4 ため、和牛肉よりも肉汁を失いやすい肉質であ 等級のデータ(n=40)[和牛去勢 4 等級割合 る。一方、和牛肉よりも脂質含量は低いが、脂 38.4%(最多) ;平成 23 年度 1 月〜12 月集計(約 肪融点は高く、加えて脂が黄色く特徴的な色を - 83 - している。②周年放牧肥育牛肉は乳用肥育牛肉 ミノ酸総量などの旨味成分含量が少ないが、食 と比べて、遊離アミノ酸総量やペプチド構成ア 感は軟らかく噛み応えも低い。また、乳用肥育 表4.周年放牧肥育により生産された褐毛和種去勢雄牛の肉質の特徴 (文献 7 の表 6 を一部改変) - 84 - 表5.周年放牧肥育により生産された褐毛和種去勢雄牛の肉質の特徴 (文献 7 の表 7 を一部改変) - 85 - 表6.周年放牧肥育により生産された牛肉の特徴(文献 8 の表 10 を一部改変) - 86 - 牛肉よりも加熱損失率が低いため肉汁を失い の比較.九州農業試験場年報, 63-67. にくい肉質である。一方、脂質含量は乳用肥育 4) 中村好德・平野 清・中西雄二・神谷 充・ 牛肉よりも高い。③周年放牧肥育牛肉は輸入牛 常石英作・加藤直樹・林 義朗・山田明央 肉と比べて、遊離アミノ酸総量が少ないが、ペ 2010.放牧と自給飼料を組み合わせた新し プチド構成アミノ酸総量は同程度である。食感 い肉用牛生産方法“草地育成・肥育技術” は輸入牛肉よりも軟らかく噛み応えも低い。ま の可能性と肉質の特徴.日本暖地畜産学会 た、輸入牛肉よりも加熱損失率が低く、圧搾肉 報, 53(1):41-49. 汁率が高いためジューシーな肉質である。一方、 5) (独)農研機構 プレスリリース.2011. 脂肪融点は輸入牛肉よりも低いため脂の口溶 「周年放牧にトウモロコシサイレージ給与 けは良好であるが、脂質含量は非常に高い。 を組み入れた飼料自給率100%の肥育技術」 http://www.naro.affrc.go.jp/publicity 4.おわりに _report/press/laboratory/karc/018328. 本研究の結果は、日本においても配合飼料か html ら脱却し、放牧と国産飼料のみで牛肉を生産で 6) 中村好德・守田 智・金子 真・加藤直樹・ きる可能性を示唆している。今後、出荷頭数を 林 義朗・常石英作・山田明央 2011.慣 増やしてデータの蓄積を行い、増体量の個体差 行肥育と草地肥育により生産された褐毛 や肉質の季節変化などについても検討する必 和種去勢雄牛の半腱様筋における肉質比 要がある。しかし、周年放牧肥育牛肉は現在わ 較. 日本暖地畜産学会報, 54(1):49-60. が国で流通しているその他の牛肉と比べて、そ 7) 中村好德・金子 真・林 義朗・莟 博行・ れぞれ異なる特徴を有することが示唆された 山田明央 2012.草地肥育により生産され ため、牛肉市場に新しい食品素材としての価値 た褐毛和種去勢雄牛の産肉性と肉質に関 を創造できる可能性は十分にあると考えられ する調査.日本暖地畜産学会報, 55(2): た。 181-194. 8) 中村好德・金子 真・林 義朗・莟 博行・ 謝 辞 山田明央 2013.褐毛和種および黒毛和種 本研究を遂行するにあたり、九州沖縄農業研 去勢雄牛を用いた周年放牧肥育事例.日本 究センターの皆様から多大なご支援を賜りま 暖地畜産学会報, 56(1):79-96. した。ここに付記し深甚な謝意を表します。 9) 服部育男・佐藤健次・鈴木知之・神谷 充・ 加藤直樹 【引用文献】 焼酎粕濃縮液の飼料成分に及ぼす影響.日 1) 農林水産省編.2013.飼料をめぐる情勢. pp1,6.農林水産省.東京. 2010a.原料および産出時期が 本暖地畜産学会報, 53:175-181. 10) 服部育男・鈴木知之・神谷 充・佐藤健次・ 2) 西岡輝美・石塚 譲・安松谷恵子・入江正 加藤直樹 2010b.焼酎粕濃縮液の混合が 和 2008.市場および小売店における牛肉 発酵TMRの発酵品質と乾物回収率に及ぼす 脂肪の嗜好性と理化学的特性との関連.日 影響.日本草地学会誌, 55:297-301. 本畜産学会報, 79:391-401. 11) (社)全国家畜畜産物衛生指導協会編. 3) 滝本勇治・黒肥地一郎・中西雄二・美濃貞 2004.生産獣医療を目指して・テキストシ 治郎 1975.若齢牛の代償性成長に関する リーズ③ 研究 第3報 草利用による若齢肥育法と濃 pp183-191. 厚飼料飽食肥育法との増体および産肉性 生産獣医療システム 肉牛編. 12) Nakamura Y-N,Hirano K,Kato N,Tsuneishi - 87 - E,Kamiya M,Yamada A,Nakanishi Y.2009. 獣医臨床病理学.pp220, 223, 228, 232, Possibility of new beef production 234-238.近代出版.東京. system using grazing and 14) 資源調査分科会編.2010.日本食品標準成 self-sufficient feed for Japanese Black and Japanese Brown (Kumamoto) steers in southern Japan: A 分表2010.文部科学省.東京. 15) 資源調査分科会編.2005.五訂増補日本食 preliminary 品標準成分表 脂肪酸成分表編2005.文部 investigation of growth performance, carcass characteristics and meat 科学省.東京. 16) (独)農畜産業振興機構編.2011.畜産の qualities.食農と環境.6:79-91. 情報 13) 小野憲一郎・太田亨二・鈴木直義編.1998. 別冊統計資料.pp16-18,62.農畜 産業振興機構 調査情報部.東京. - 88 - 栄養生理研究会報 Vol. 57, No. 2 2013 Tissue distribution and blood circulation of agouti signaling protein (ASIP) in relationship with body composition in cattle E. Albrecht1, *, K. Komolka1, K. Will1, B. Kuhla2, H. Sauerwein3, T. Gotoh4, S. Maak1 1 Leibniz Institute for Farm Animal Biology, Institute for Muscle Biology and Growth, Wilhelm-Stahl- Allee 2, 18146 Dummerstorf, Germany; 2 Leibniz Institute for Farm Animal Biology, Institute of Nutritional Physiology "Oskar Kellner", Wilhelm-Stahl-Allee 2, 18146 Dummerstorf, Germany; 3 University of Bonn, Institute for Animal Science, Physiology and Hygiene Unit, Katzenburgweg 7-9, 53115 Bonn, Germany; 4Kyushu University, Kuju Agricultural Research Center, Kuju 4045-4, Takeda 878-020, Oita, Japan SUMMARY body fat traits. In contrast, plasma leptin The agouti signaling protein (ASIP) has been concentrations were not correlated with body fat previously related to obesity in mice and humans, traits. Localization of ASIP and agouti related beside its role in melanogenesis. Different auto- protein and paracrine effects of ASIP were demonstrated, hypothalamus may be indicative for competitive but also endocrine functions are possible while action of ASIP and AgRP in receptor binding. circulating in blood. Binding to receptors Our results indicate that ASIP is widespread in involved the bovine tissues and is circulating in blood. A potentially crucial role in adipocyte development hitherto unknown endocrine function can be makes it a tempting candidate for economically assumed with possible involvement in fat relevant, fat related traits in cattle. Therefore, we deposition investigated the protein abundance of ASIP in regulation in cattle. in appetite regulation and (AgRP) and in the same interference cells with in appetite different tissues and blood of cattle and potential relationships with fat deposition. A custom made, 1. INTRODUCTION bovine-specific antibody was used for detection The agouti signaling protein (ASIP) was of ASIP in tissue, plasma, and differentiating originally known for its role in melanogenesis primary bovine stromal vascular (sv) cells by and determination of coat color in different western blot or immunohistochemistry. Blood species [1,2]. It has been related to obesity since samples of 215 Charolais (CH) x Holstein (HS) identification of a fusion transcript of ASIP and F2 crossbred bulls were taken at 250 days of age a non-coding exon of an adjacent gene (RALY, and fat related traits were determined after hnRNP protein associated with lethal yellow) as slaughter at 18 months of age. ASIP was cause for embryonic lethality in mice which are detected in all investigated tissues with higher homozygous for this aberration and for obesity, concentrations in adipose tissues and liver. These insulin tissues may be a main source of the circulating susceptibility protein. Significant, positive relationships were heterozygous, obese mice display an ectopic detected between plasma ASIP abundance and expression of ASIP [4,5]. Therefore, the agouti resistance, in and heterozygotes Proceedings of Japanese Society for Animal Nutrition and Metabolism Vol. 57, No. 2, 2013. * Corresponding author: [email protected] - 89 - increased [3]. tumor These locus became known as the first obesity gene to ASIP in different tissues and blood of cattle and be cloned in mice [6,7]. While ASIP expression possible relationships between ASIP and body in mice is restricted to hair follicles under fat deposition. physiological conditions, it is expressed in different tissues in humans and cattle without 2. MATERIALS AND METHODS pathological changes [8,9]. Investigations on Animals and sampling possible ASIP function in development of Tissue samples from longissimus muscle (LM), metabolic disorders like obesity and type II intermuscular and subcutaneous fat (SCF), lung, diabetes led to several hypotheses regarding the liver, heart, and skin were harvested from 5 role of ASIP in adipocyte differentiation and Japanese Black crossbred bulls during regular metabolism. ASIP actions via melanocortin slaughter for protein extraction and subsequent receptors (MCR) as well as MCR independent western effects were proposed [10]. ASIP binds primarily histochemistry as described elsewhere [15]. to MC1R and MC4R, but also to MC3R with Blood samples were taken at 250 days of age weaker affinity [11,12]. These receptors exist in from 215 Charolais (CH) x Holstein (HS) F2 different tissues. In the skin during regulation of crossbred bulls. Details of the experiment are melanogenesis, ASIP binds competitively to described elsewhere [16]. More than 100 MC1R and thus precludes alpha-melanocyte phenotypic traits were assessed for each bull stimulating hormone (α-MSH) initiated signaling after slaughter at 18 months of age. Twenty bulls which blocks production of cAMP [1,2]. MC3R with the highest or lowest amount of carcass fat and MC4R are expressed in the central nervous were assigned to a high carcass fat (HCF) and a system, where they are involved in appetite low carcass fat (LCF) group, respectively, for regulation. These receptors are strongly bound separate by the agouti related protein (AgRP). Since hypothalamus of one cow from the experiment ASIP and AgRP are antagonists of MC1R and described MC4R immunohistochemistry. with inverse activity [13,14], an blot analysis analyses. elsewhere and for immuno- Additionally, [7] was used the for association of ASIP with appetite regulation is also conceivable. Alternatively, ASIP could be Protein analyses involved in appetite regulation via autocrine Total protein was extracted from all tissues using action on leptin in adipocytes [7]. ASIP CelLytic MT lyses reagent (Sigma-Aldrich, circulating in blood as a prerequisite for an Munich, Germany) with protease inhibitor endocrine function has neither been described in according to manufacturer’s instructions. Protein human nor in mice. However, ASIP is extract, 40 μg, was separated on a 15% SDS- circulating in bovine blood with hitherto PAGE 10x10 cm mini gel and transferred to a unknown (Albrecht, polyvinylidene difluoride (PVDF) membrane as unpublished data). The potentially crucial role in described elsewhere [15]. The amount of ASIP adipocyte development and potential endocrine in bovine plasma was quantified by western blot functions makes it a tempting candidate for analysis after albumin removal with Aurum Affi- economically relevant traits in farm animals like Gel Blue (Bio-Rad, Munich, Germany) at least body composition and fat deposition. Therefore, in duplicates. Twenty μg denatured plasma in this study, we investigated the abundance of protein was separated on Criterion TGX 12% endocrine function - 90 - gels (Bio-Rad) and transferred to PVDF Immunofluorescence was visualized with a membranes. A custom-made, polyclonal bovine Nikon Microphot SA fluorescence microscope ASIP antibody [15] was used to detect ASIP in (Nikon, Duesseldorf, Germany) and an image tissues and plasma. The primary antibody analysis system equipped with CELL^F software (1:10,000) was incubated at 4°C overnight and and a CC-12 high resolution color camera (OSIS, the Muenster, Germany). secondary HRP conjugated antibody (1:50,000, rabbit IgG TrueBlot, eBioscience, Frankfurt, Germany) at room temperature for 1 Cell culture hour. with Primary stromal vascular cells were isolated Immunstar from bovine intermuscular fat and grown for 9 Antibody label chemiluminescence was detected substrate Western C (Bio-Rad) and a Chemocam HR-16 days imager Germany). differentiated for 13 days in Insulin–Transferrin– LabImage 1D software (Kapelan Bio-Imaging, Selenium (ITS) medium and fixed with 4% Leipzig, Germany) was used to quantify paraformaldehyde. Cells were immunostained intensity of specific bands. for (INTAS, Goettingen, The amount of leptin in plasma samples in growth detection of medium. ASIP as Cells described were for cryosections. was determined by a bovine-specific ELISA as described elsewhere [19]. Statistical analysis Statistical analysis was performed using the SAS Immunohistochemistry statistical software (Version 9.2, SAS Inst. Inc., Samples of different tissues were cryo-sectioned Cary, USA). Data were analyzed by ANOVA (8 µm thick) using a Leica CM3050 S (Leica, using the GLM procedure with fixed factor Bensheim, microtome. group (high and low carcass fat). Values of Sections were immunostained to localize ASIP different blots for the same animal were included as described in [15]. Cryo-sections of the right as repeated measurements. The t-test was used as hypothalamic hemisphere were cut, fixed, and post-hoc test with P 0.05 as threshold for incubated with a guinea pig-anti-AgRP antibody significant differences. Germany) cryostat (1:200, GP-029-50, Novus Biologicals, USA) as Relationships between traits were described by Börner et al. [17]. For double calculated as Pearson’s-correlation coefficients labeling with ASIP, both antibodies were using the CORR procedure of SAS. incubated simultaneously. Specific binding of the primary antibodies was detected using Alexa 3. RESULTS AND DISCUSSION Fluor 568 labeled goat-anti-guinea pig IgG In contrast to mice, where physiological ASIP (1:250, 949212, Invitrogen, USA) and MFP 488 expression is restricted to hair follicles [1], a labeled goat-anti-rabbit IgG (1:500, MoBiTec, widespread expression of ASIP was detected in Goettingen, Germany) secondary antibodies, bovine tissues by western immunoblotting respectively. Disturbing auto-fluorescence was (Figure 1) and/or immunohistochemistry [15]. reduced by including a step of Sudan black Comparing the band intensity of equal amounts (0.1% in 70% Ethanol) staining for 30 min. of protein extracts from different tissues, highest Slices were counterstained with 1 µg/ml Hoechst amounts of ASIP were measured in liver and 33258 adipose tissues suggesting an association with (Sigma-Aldrich) to display nuclei. - 91 - effects on the surrounding muscle cells. Also non-adipose cells in adipose tissue may secrete ASIP and affect the metabolism of adipocytes and other cells or adipocytes are affected by endogenous ASIP production in a more autocrine manner as suggested by Smith et al. [19]. To clarify whether adipocytes are the exclusive source of ASIP in adipose tissue, primary sv cells were isolated from Figure 1. ASIP protein expression in different intermuscular adipose tissue and differentiated to bovine tissues. Chemiluminescence detection of adipocytes. ASIP was mainly detected in the ASIP by western blotting of 40 µg protein of the cytoplasm of cells containing several lipid respective tissues using a bovine specific antibody droplets (Albrecht, unpublished data). This (1:10,000). Lanes 1 and 10: molecular weight marker, implies that ASIP is expressed early during 2: subcutaneous fat, 3: intermuscular fat, 4: M. adipogenic differentiation before maturation of longissimus, 5: heart, 6: liver, 7: lung, 8: skin (white), the adipocyte and may be secreted either upon a 9: skin (black). The specific bands are framed certain stimulus or spontaneously. Additionally, (Reproduced from Ref. 15, Figure S2a, with the a nuclear signal was detected in single cells permission). without lipid droplets as described for cells in tissue sections [15]. We can only speculate that this is a sign of paracrine or autocrine action of body fat deposition. When the band intensity of ASIP with unknown consequence. SCF (Figure 1, lane 2) was set to 100%, Western blot results indicated that liver intermuscular fat had 111% and liver 325% of and adipose tissues were the tissues with the the signal strength. White and black skin, lung highest content of ASIP in cattle. If this protein and heart contained much less amounts of ASIP is mainly secreted, it could be expected to (7.1, 6.4, 4.6, and 1.9%, respectively). In skeletal circulate in the blood. Indeed, ASIP was detected muscle, however, ASIP was not detectable with in bovine plasma by western blot. In contrast to western immuno- several unspecific bands additionally observed in histochemical ASIP detection were described in bovine tissues, a single band was detected in [15] and revealed that also few cells in skeletal plasma at ~20 kDa. This band was shown to be muscle, intramuscular adipocytes and other cells specific by blocking with the respective peptide located in connective tissue, showed a specific [15]. The size is larger than the theoretical size signal. ASIP signals appeared in two distinct of ASIP which may be due to glycosylation. forms, either cytoplasmic, as expected for a Circulation of ASIP implies a hitherto unknown secreted protein, or associated to nuclei. endocrine function with possible involvement in Immunofluorescence signals were never attached fat deposition in cattle. A prerequisite for such a to muscle fibers confirming that myocytes did function is the expression of respective targets. It not express ASIP. This does not exclude that was early shown that ASIP is a potent antagonist ASIP, secreted from IMF inclusions, may have of the MC1R explaining its role in melanogene- blotting. Results of - 92 - Figure 2. Carcass fat and plasma concentrations of ASIP and leptin in cattle differing in body composition. Normalized ASIP levels were determined by western blot relative to albumin and leptin concentration by ELISA. Crossbred bulls were assigned to low (LCF) or high fat (HCF) group according to their amount of carcass fat (n = 20 per group). *significant differences between groups (P < 0.05). sis [1]. Furthermore, MC4R was antagonized in deposition, we analyzed plasma samples from a similar manner, thus indicating a potential role 215 F2 crossbred bulls taken at 250 days of age of ASIP outside melanogenesis [1]. In our earlier and correlated ASIP and leptin concentrations to study [15], we demonstrated that MC1R and phenotypic data generated at slaughter with 18 MC4R are expressed in SCF and thus, may be months of age. Earlier studies have shown that putative targets for autocrine action of ASIP in leptin serves as an indicator of body fat content this tissue. [23,24]. If ASIP can be considered as an The melanocortine receptors 3 and 4 are adipokine like leptin, we hypothesized that ASIP expressed in the central nervous system where concentrations in bovine plasma during growth they are involved in appetite regulation [20]. could be indicative for body composition traits at These receptors are mainly controlled by AgRP slaughter. Comparing ASIP plasma levels of 20 [12]. Studies in mice revealed that ASIP can bulls with lowest carcass fat (LCF) and 20 bulls mimic AgRP signaling through the MC4R in the with highest carcass fat (HCF), relative amounts brain [21,22]. As a first attempt to confirm a of ASIP in plasma was significantly higher in the possible action of ASIP in bovine hypothalamus, HCF we tried to co-localize ASIP and AgRP in this concentrations were not different between the tissue. The co-immunohistochemistry revealed groups (Figure 2). Furthermore, our results that both proteins were detectable in the same revealed structures of the brain but with different staining correlations between body fat traits and plasma pattern (Albrecht, unpublished data). This result ASIP abundance (carcass fat - ASIP: r = 0.2, n = might indicate that ASIP influences appetite 215, P = 0.002). In contrast, leptin plasma regulation via competition of MCR binding with concentrations AgRP. correlation with any of the fat related traits nor To investigate possible associations group, whereas weak, but exhibited plasma significant, neither leptin positive significant with ASIP plasma levels. between plasma ASIP abundance and body fat Our results indicate that ASIP is expressed - 93 - by adipocytes and further cells in many bovine 5. Duhl DM, Vrieling H, Miller KA, Wolff GL, tissues. Adipose tissue and liver may be main Barsh sources of the circulating protein. Our results mutations in obese yellow mice. Nat. Genet. provide evidence for an association between 8: 59-65. circulating ASIP and body fat deposition in GS. 1994. Neomorphic agouti 6. Bultman SJ, Russell LB, Gutierrez-Espeleta cattle and suggests hitherto unknown endocrine GA, functions for ASIP. Different MCRs as putative characterization of targets are expressed in diverse tissues and may associated with mutations at the agouti locus be involved in paracrine and endocrine actions of in the mouse. Proc. Natl. Acad. Sci. USA 88: ASIP. 8062-8066. Whether ASIP influences body Woychik RP. 1991. Molecular a region of DNA composition via interference with appetite 7. Claycombe KJ, Xue BZ, Mynatt RL, Zemel regulation as competitor to AgRP or via MB, Moustaid-Moussa N. 2000. Regulation regulation of leptin as indicated in [7] or distinct of leptin by agouti. Physiol. Genomics 2: pathways, needs further investigations. 101-105. 8. Kwon HY, Bultman SJ, Loffler C, Chen WJ, ACKNOWLEDGEMENTS Furdon PJ et al. 1994. Molecular structure The authors wish to thank Ines Rothe, Karola and chromosomal mapping of the human Marquardt, Elke Schwitulla, and Birgit Mielenz homolog of the agouti gene. Proc. Natl. Acad. for excellent technical assistance. Sci. USA 91: 9760-9764. 9. Wilson BD, Ollmann MM, Kang L, Stoffel REFERENCES M, Bell GI et al. 1995. Structure and 1. Lu D, Willard D, Patel IR, Kadwell S, function of ASP, the human homolog of the Overton L, et al. 1994. Agouti protein is an mouse agouti gene. Hum. Mol. Genet. 4: antagonist of the melanocyte-stimulating- 223-230. hormone receptor. Nature 371: 799-802. 10. Mynatt RL, Stephens JM. 2001. Agouti 2. Cone RD, Lu D, Koppula S, Vage DI, regulates adipocyte transcription factors. Am. Klungland H, et al. 1996. The melanocortin J. Physiol. Cell Physiol. 280: C954-C961. receptors: agonists, antagonists, and the 11. Yang YK, Ollmann MM, Wilson BD, hormonal control of pigmentation. Recent. Dickinson C, Yamada T, et al. 1997. Effects Prog. Horm. Res. 51: 287–317. of recombinant agouti-signaling protein on 3. Michaud EJ, Bultman SJ, Stubbs LJ, melanocortin action. Mol. Endocrinol. 11: Woychik RP. 1993. The embryonic lethality 274-280. of homozygous lethal yellow mice (Ay/Ay) 12. McNulty JC, Jackson PJ, Thompson DA, is associated with the disruption of a novel Chai B, Gantz I, Barsh GS, Dawson PE, RNA-binding protein. Genes Dev. 7: 1203- Millhauser GL. 2005. Structures of the 1213. Agouti Signaling Protein. J. Mol. Biol. 346: 4. Yen TT, Gill AM, Frigeri LG, Barsh GS, 1059-1070. Wolff GL. 1994. Obesity, diabetes, and 13. Chai BX, Neubig RR, Millhauser GL, neoplasia in yellow A(vy)/- mice: ectopic Thompson DA, Jackson PJ, Barsh GS, expression of the agouti gene. FASEB J. 8: Dickinson CJ, Li JY, Lai YM, Gantz I. 2003. 479-488. - 94 - Inverse agonist activity of agouti and agouti- Nguyen T, Murrell A et al. 2003. Agouti related protein. Peptides 24: 603-609. expression in functional consequences 14. Nix MA, Kaelin C, Ta T, Weis A, Morton human adipose and tissue: increased GJ, Barsh GS, Millhauser GL. 2013. expression in type 2 diabetes. Diabetes 52: Molecular and functional analysis of human 2914-2922. β-defensin 3 action at melanocortin receptors. 20. Rossi M, Kim MS, Morgan DG, Small CJ, Chem. Biol. 20: 784-795. Edwards CM, Sunter D, Abusnana S, 15. Albrecht E, Komolka K, Kuzinski J, Gotoh T, Goldstone AP, Russell SH, Stanley SA, Maak S. 2012. Agouti Revisited: Transcript Smith DM, Yagaloff K, Ghatei MA, and Quantification of the ASIP Gene in Bovine Bloom SR. 1998. A C-terminal fragment of Tissues Related to Protein Expression and Agouti-related protein increases feeding and Localization. PlosOne 7: e35282. antagonizes the effect of alpha-melanocyte 16. Kühn C, Bellmann O, Voigt J, Wegner J, stimulating hormone in vivo. Endocrinology Guiard V, Ender K. 2002. An experimental 139: 4428-4431. approach for studying the genetic and physiological background of 21. Wilson BD, Ollmann MM, Barsh GS. 1999. nutrient The role of agouti-related protein in transformation in cattle with respect to regulating body weight. Mol. Med. Today 5: nutrient secretion and accretion type. Arch. 250-256. Tierz. 45: 317-330. 22. Ollmann MM, Wilson BD, Yang YK, Kerns 17. Börner S, Albrecht E, Schäff C, Hacke S, JA, Chen Y, Gantz I, Barsh GS. 1997. Kautzsch U, Derno M, Hammon HM, Antagonism Röntgen M, Sauerwein H, Kuhla B. 2013. receptors in vitro and in vivo by agouti- Reduced related protein. Science 278: 135-138. AgRP activation in the of central melanocortin hypothalamus of cows with high extent of fat 23. Bunger L, Nicolson M, Hill WG. 1999. mobilization after parturition. Gen. Comp. Leptin levels in lines of mice developed by Endocr. Doi: 10.1016/j.ygcen.2013.08.002 long-term divergent selection on fat content. 18. Sauerwein H, Heintges U, Hennies M, Genet. Res. 73: 37-44. Selhorst T, Daxenberger A. 2004. Growth 24. Delavaud C, Bocquier F, Chilliard Y, Keisler hormone induced alterations of leptin serum DH, Gertler A. 2000. concentrations in dairy cows as measured by determination in ruminants: a novel enzyme immunoassay. Livest. Prod. nutritional status and body fatness on plasma Sci. 87: 189-195. leptin concentration assessed by a specific 19. Smith SR, Gawronska-Kozak B, Janderova L, - 95 - Plasma leptin effect RIA in sheep. J. Endocrinol. 165: 519-526. of 栄養生理研究会報 Vol. 57, No. 2 2013 The impact of retinoic acid on adipogenesis in 3T3-L1 cells Katharina Stoecker1, *, Steffen Sass2, Michael W. Pfaffl1 1 Physiology Weihenstephan, ZIEL Research Center for Nutrition and Food Sciences, Technische Universität München, Freising, Germany; 2Institute of Computational Biology, Helmholtz Center Munich, German Research Center for Environmental Health, Ingolstädter Landstraße 1, 85764 Oberschleißheim, Germany SUMMARY System (GEPS) as well as a multiple linear Adipogenesis, the development of fat cells is one regression model combined with the target of the most studied issues in the field of cell predictions of Targetscan Mouse 5.2 differentiation. All-trans-retinoic-acid (ATRA) performed a joint analysis of our microRNA and inhibits fat cell differentiation almost completely mRNA expression data. We could show that by suppressing peroxisome proliferator-activated ATRA downregulates the caveolin-1 signaling as receptor (Ppar) expression, the key regulator well as the actin cytoskeleton rearrangement in of adipogenesis and therefore only few mature 3T3-L1 cells. The inhibition of these signaling adipocytes are built. The inhibition of fat cell cascades may prevent the conversion from differentiation is subjected to transcriptional and fibroblast-like to the adipocyte phenotype. post-transcriptional by Moreover we could show that the fatty acid microRNAs. To examine the effects of ATRA anabolism in 3T3-L1 cells is also inhibited, on adipocyte differentiation, murine 3T3-L1 which is in accordance with the ATRA mediated preadipocytes were cultured in presence and phenotype, the loss of lipid accumulation. changes mediated we absence of 3 µM ATRA and total RNA was isolated at 0, 2, 7, 10, 24, 48, 96 hours and 12 1. INTRODUCTION days after induction. Using high-throughput Juiciness, tenderness and flavor, the most analysis tools like microarray technology and important quality features of beef, are addicted reverse transcription quantitative polymerase to the intramuscular fat content, which is also chain reaction (RT-qPCR), new ATRA target termed as marbling. Because the accumulation of genes could be detected. GATA binding protein intramuscular fat is influenced by endogenous 6 (Gata6), is induced by ATRA and the function and exogenous factors like genetic disposition, was analysed using small interfering RNAs nutrition and breed, cattle breeds with a high (siRNAs) knockdown knockdown of Gata6 technology. The genetic marbling linked to innovative nutritional prevents lipid feeding strategies are examined in nowadays accumulation even in absence of ATRA. Due to [1,2] . this effect, Gata6 may be a proadipogenic player, The intramuscular fat content increases which mediates compensatory effects to ATRA continuously during growth, therefore nutritional in 3T3-L1 cells. Using Genomatix Pathway effects of feed have to be examined in more Proceedings of Japanese Society for Animal Nutrition and Metabolism Vol. 57, No. 2, 2013. * Corresponding author: [email protected] - 97 - detail, because the knowledge of the involved small interfering RNAs (siRNAs) are small non- molecular mechanism is far from complete. coding RNA species. These non-coding RNA Vitamin A, a nutrient can be found in many food types mediate RNA interference, the inhibition e.g. meat, milk, carrot and also in the grazing or degradation of specific mRNA [16]. RNA lands. Many of the non-visual functions of interference is a natural mechanism in living vitamin A are mediated by all-trans-retinoic-acid cells and is important for the protection of cells (ATRA), a derivate of vitamin A [3]. The effect against external nucleotide sequences e.g. of of ATRA signaling depends on concentration parasites, viruses, and transposons as well as in and is a fine-tuned process [4]. Therefore, ATRA controlling differentiation and gene expression is a potent modulator of differentiation [5]. It [17,18]. induces the differentiation of osteoblasts [6] and siRNAs can be used in experimental biology to myoblasts [7] and on the other hand it inhibits examine the function of genes in vitro. the development of mature adipocytes [8]. Furthermore, synthetical generated This study examines the inhibitory effects The development of mature adipocytes is of ATRA on fat cell differentiation. Our findings a fine-tuned and optimized intracellular process indicates that ATRA modulates microRNA as and can be induced by treating the cells with the well as mRNA expression in 3T3-L1 cells. We prodifferentiative agents dexamethasone, 3- identified GATA binding protein 6 (Gata6) as isobutyl-1-methylxanthine and insulin (DMI, the new ATRA target gene and a comprehensive in differentiation cocktail). The effects of ATRA on silico based analysis of these real mRNA and fat cell differentiation can be studied using cell microRNA culture models e.g. the preadipocyte cell line rearrangement of the cytoskeleton is affected by 3T3-L1 [9]. For terminal differentiation of ATRA. data demonstrates that the growth arrested and with DMI induced 3T3-L1 cells it is necessary to renter the cell cycle and 2. MATERIALS AND METHODS undergo mitotic clonal expansion, whereby the Cell culture cell number increases about 4 times [8,10,11]. Murine 3T3-L1 cells were purchased from the After stimulation, a complex signaling cascade is American Type Culture Collection (ATCC) and activated, including the activation of the differentiated as described by the supplier. transcriptional key regulator the peroxisome Differentiation (D0) was started by using proliferator-activated receptor (Ppar) [12]. In differentiation medium (DMI) containing 10% conclusion, the expression of adipocyte specific fetal bovine serum (FBS), 1% Penicillin-Strepto- genes and the activation of metabolic enzymes mycin, combined with the rearrangement of the actin (IBMX), 1 µM dexamethasone (DEX) and 1 cytoskeleton results in the formation of mature µg/ml insulin in 500 ml Dulbecco’s modified adipocytes [13-15]. Through this intracellular Eagle’s interplay the phenotype is modified from preadipocytes were cultivated with DMI in fibroblast-like to an adipose cell type. presence and absence of 3 µM ATRA. After 96 The regulation of differentiation is related to transcriptional and 0.5 mM medium 3-isobutyl-1-methylxanthin (DMEM). 3T3-L1 hours DMI was replaced by growth medium post-transcriptional containing 10% FBS, 1% Penicillin/ changes in gene expression communicated by Streptomycin and 1 µg/ml insulin in 500 ml small non-coding RNA species. MicroRNAs and DMEM. Medium was renewed every two days. - 98 - Cells were harvested at the time point 0, 2, 4, 7, curves. Standard dilutions 10, 24, 48, 96 hours and after 12 days. concentrations of 10 –10 molecules were used 2 for absolute series with 7 quantification. Results were expressed as copy numbers per μl. RNA extraction During manifold cell culture experiments, total The concentrations of gene primers for RNA of developing 3T3-L1 preadipocytes were Ppar (5´-ACGTGAAGCCCATCGAG-3´ and isolated from 6-well plates with the miRNeasy 5´-AGCACCTTGGCGAACAG-3´), the mono- Mini Kit (Qiagen) as described by the supplier. cyte to macrophage differentiation-associated After RNA extraction, RNA concentration and (Mmd; purity the GT-3´ and 5´-TGGTGTGGGCCACCAGGAGT- spectrophotometer NanoDrop1000. All, later 3´), the dehydrogenase/reductase (SDR family) analyzed samples occupy a 260/280 ratio with a member 3 (Dhrs3; 5´-GGCACAGAGTGCCA range between 1.8-2.1. The RNA quality was CTACTTCA-3´ and 5´-CAGCTTTGGCCATCT confirmed GGTACA-3´) was determined by using by the using Agilent 2100 5´-GGGCAAAGGCATGTAG´CGAA and for Gata6 (5´-TGCA Bioanalyzer randomly. Only samples with a CTCCGCCTCCAGGATAGA-3´ and 5´- RNA Integrity Number (RIN) greater 8 were CCCGACCACCGTGCACATTT-3´) were 200 used for further analysis. nM for each primer and 1 µl of cDNA was added to each RT-qPCR reaction in a total volume of mRNA and microRNA profiling 10 µl. The cycling conditions of the supplier of mRNA expression was quantified by using high SsoFast EvaGreen (Bio-Rad) were used. throughput transcriptomics technology (Affymetrix) transcription quantitative like microarray With the nCounter mouse microRNA reverse expression assay (NanoString, Seattle, United chain States) more than 600 murine and murine reaction (RT-qPCR). Gene expression analysis associated viral microRNAs can be detected. was performed using the Gene 1.0 ST Array This assay was used for microRNA profiling. and polymerase System of Affymetrix. With this Affymetrix GeneChip 28 853 well annotated genes can be RNA interference detected. For the validation of the microarray To examine the function of genes in vitro, the results a moloney murine leukemia virus reverse technology of siRNAs was used. We used transcriptase (M-MLV RT [H–] (Promega, purchasable ready to use siRNAs of Dharmacon. Germany)) was used for cDNA synthesis. The A final siRNA concentration of 25 nM was used reaction mix was incubated for 20 min at 21°C, for each transfection and the general protocol of then the cDNA synthesis step was performed at Thermo Scientific DharmaFECT transfection 120 min at 48°C and the reaction was stopped by was used as described by the supplier. To 2 min incubation at 90°C. The SsoFast EvaGreen determine undesired cellular responses in RNAi Supermix (Bio-Rad, Germany) was used for the experiments, a pool of 4 non-targeting siRNAs RT-qPCR were used as negative control. performance. quantification of gene For the absolute expression external calibration curves were generated. Templates Pathway analysis derived from RT-PCR with known nucleic acid Only transcripts with an 2-fold up- or down- content were used for the preparation of standard regulation induced by ATRA were analyzed with - 99 - the software Genomatix Pathway System induction of terminal differentiation [11]. After (GEPS) to generate and discover biological induction of differentiation, the confluent and networks based on literature data [19]. growth-arrested 3T3-L1 cells normally reenter the cell cycle and undergo mitotic clonal In silico microRNA target predictions expansion followed by the expression of For microRNA-mRNA Data integration, a adipocyte specific genes e.g. Ppar[8]. The multiple linear regression model in combination morphology was determined by microscopic with target predictions of Targetscan Mouse 5.2 analysis. 12 days post induction 3T3-L1 cells was used [20,21]. 189 KEGG Pathways were were stained with oil-red-O. In Fig. 1B mature analysed. For Visualization Cytoscape 2.8.3 was adipocytes are shown. The cells have an adipose- utilized [22,23]. like phenotype: the cell shape is rounded and lipid is accumulated. ATRA inhibits lipid Oil-red-O-staining For differentiation accumulation in 3T3-L1 preadipocytes. Mature control, the lipid adipocytes were only built in patches and accumulation was quantified at day 12 by oil- therefore in most 3T3-L1 cells no conversion of red-O-staining. The degree of staining is the phenotype was performed (Fig. 1C). The proportional to the extent of cell differentiation morphology of ATRA treated cells is still [24]. fibroblast-like and only in few cells lipid is accumulated (Fig. 1C). The inhibition of Quantity of cells phenotype conversion is associated with the gene At each point in time, cells were quantified by expression counting the DAPI-stained nuclei of 3T3-L1 Pparexpression cells image concentration-dependent manner. On day 4 after processing program of the National Institute of induction the Pparexpression was quantified Health [25]. For this, six independent pictures by qRT-PCR. The Ppar expression is 3-fold per 6-well/time point were generated and downregulated by 1 µM ATRA (Fig. 1D). using ImageJ, a Java-based of Ppar. of ATRA 3T3-L1 inhibits cells in a analyzed. High throuput transcriptomic studies for 3. RESULTS AND DISCUSSION identification of new ATRA targets ATRA has no effect on cell proliferation, but Using microarray technology we were able to inhibits adipocyte conversion identify 755 genes, with more than 2-fold up- or In early studies it was shown that ATRA has no down-regulation by ATRA. Out of 755 genes 13 effect on the proliferation of cells [26]. We transcription factors are induced by ATRA and confirmed these findings: On day 12 after 22 transcription factors are repressed by ATRA initiation of differentiation the cell number is (Stoecker et al., unpublished data). For fat cell nearly the same in presence or absence of 3 µM differentiation a sequential induction of the ATRA. Independent of the ATRA treatment, the transcription factors Cebp, Cebp and Ppar is quantity of cells on day 12 is 4 fold higher necessary. ATRA inhibits adipogenesis by relative to 0 hours (Fig. 1A). Therefore, ATRA diminishing Cebp signaling, which cause a has no effects on the clonal expansion phase in significant lower expression of Ppar and Cebp 3T3-L1 cells, which is important for the [27,28]. For the validation of our microarray - 100 - Figure 1. Phenotype and Genotype of ATRA treated 3T3-L1 cells. (A) DAPI-stained nuclei of 3T3-L1 cells were counted, using ImageJ. ATRA has no effect on cell proliferation. The cell quantity on day 12 is 4 fold higher relative to 0 hours in both, ATRA treated and untreated cells. The standard deviations for replicates of 3 independent experiments are shown as error bars. (B, C) Oil-red-O staining of ATRA untreated and treated 3T3-L1 cells 12 days after induction. For microscopic image generation a magnification factor of 5-fold was used. (B) Lipid accumulation in mature adipocytes. (C) Lipid accumulation is inhibited in ATRA treated cells. (D) ATRA inhibits Ppar expression of 3T3-L1 cells in a concentration dependent manner. Gene expression was detected on day 4 after induction by RT-qPCR. Standard deviations are shown as error bars. The experiment was repeated 4 times. - 101 - Figure 2. Identification of new ATRA target genes in 3T3-L1 cells. Expression profiles of Dhrs3 (A), Mmd (B) and Gata6 (C) in untreated (DMI) and ATRA-treated (DMI + 3 µM ATRA) 3T3-L1 cells. The expression was detected at the indicated times and the results were validated by 3 independent microarray experiments (data not shown) as well as by qRT-PCR. Standard deviation is shown as error bars of duplicates. results, we examined the expression of these knockdown as a method. For all siRNA transcription factors with RT-qPCR and the data experiments we used transient transfection. The confirm the significance of our array data. siRNA knockdown effects of siDhrs3, siMmd (Stoecker et al., unpublished data). In addition, and siGata6 on gene expression were detected by out of the 755 regulated gene transcripts we RT-qPCR (Fig. 3A, B, C) and oil-red-O staining selected those which were immediately early was used for phenotyping (Fig. 3D). The induced by ATRA e.g. Dhrs3, Mmd and Gata6 knockdown efficiency of siDhrs3, siMmd and (Fig. 2A, B, C). Genes, which are upregulated by siGata6 are shown in Figure 3A, B and C. We ATRA possibly are involved in ATRA signaling utilized untransfected 3T3-L1 cell and 3T3-L1 and are of utmost interest for us. Our hypothesis cells transfected with non-targeting siRNAs as was, the knockdown of ATRA induced genes controls. All controls were also cultured in DMI should abrogate the inhibitory effects of ATRA. and DMI + 3 µM ATRA. Subjected to the To verify our hypothesis we used siRNA knockdowns the expression of Dhrs3, Mmd and - 102 - Figure 3. siRNA knockdown effects of new ATRA target genes in 3T3-L1 cells. siRNA knockdown of Dhrs3 (A), Mmd (B) and Gata6 (C) expression in untreated (DMI) and ATRA-treated (DMI + 3 µM ATRA) 3T3L1 cells. (D) Oil-red-O staining on day 11 of siRNA transfected and induced 3T3-L1 cells. The knockdown of Gata6 inhibits lipid accumulation in ATRA untreated cells. - 103 - Gata6 is extremely reduced relative to the in the ATRA-dependent regulation of molecular controls. The Dhrs3 expression was more than processes in fat cells. ATRA inhibits the 2.5-fold downregulated by the corresponding caveolin-1 signaling pathway (p-value 8.56e-5) knockdown in DMI treated cells as well as in by suppression of caveolin-1 and caveolin-2 DMI + 3µM ATRA treated cells. The Mmd expression. Caveolin-1 is a integral membrane expression is 18-fold downregulated by siMmd protein and interacts with cholesterol as well as in DMI treated cells and more than 21-fold with long chain unsaturated fatty acids [29]. downregulated by siMmd in DMI +3 µM ATRA Furthermore caveolin-1 is co-located with treated cells and the Gata6 expression is more filamin. Kanzaki et al. showed that 3T3-L1 cells than 3-fold downregulated by siGata6 in DMI have an unique organized F-actin filament treated cells and more than 6-fold downregulated structure arising from the caveolin-enriched lipid by siGata6 in DMI + 3 µM ATRA treated 3T3- raft at the plasma membrane [30]. These findings L1 cells. Against our first assumption, the point out, that ATRA influences the cell shape knockdown of Gata6 resulted in a dramatically via the structure of the cytoskeleton. loss in lipid accumulation, even in absence of ATRA (Fig. 3D). Furthermore, the Ppar A comprehensive analysis of microRNA and expression was also inhibited by the Gata6 mRNA expression data. knockdown (Stoecker et al., unpublished data), To whereas the Mmd and Dhrs3 knockdown have regulatory networks in preadipocytes, it is very no effects on lipid accumulation (Fig. 3D) as important to get a better understanding of well as on Ppar gene expression (Stoecker et al., microRNA-mRNA target regulation. For this, we unpublished data). Our results indicate, that on carried out an integrative analysis of mRNA and the one hand basic gene expression of Gata6 is microRNA expression in ATRA-treated 3T3-L1 needed for adipogenesis (Fig. 3D). On the other cells. The microRNA expression in ATRA hand, Gata6 is an immediate early response gene treated and untreated cells were analyzed in two of ATRA (Fig. 2C). Maybe, the induction of independent Gata6 does not serve as mediator of the ATRA Technologies). Out of 600 detectable murine and inhibitory effects in 3T3-L1 cells. The induction murine of Gata6 is much more a counter action of the microRNAs were found to be more than 2-fold cell to ATRA as a kind of protection concerning upregulated the homeostasis of the adipogenic process. (Stoecker et al., unpublished data). In earlier Therefore, the proadipogenic function of Gata6 studies, researchers focused on discovery of is a well-balanced activity. To clarify this novel players in biological processes rather than hypothesis much more experiments will be generating needed such as overexpression of Gata6 in 3T3- transcriptional networks. The regulation of L1 cells. metabolic generate a comprehensive experiments associated or viral networks microRNAs mechanisms by of (NanoString downregulated global picture by e.g. 23 ATRA. post- microRNA-mRNA interaction was analyzed using a multiple linear GEPS Analysis: the caveolin-1 signaling regression model. Using this method, we were pathway is influenced by ATRA able to show, that the rearrangement of the actin We next used the GEPS pathway analysis cytoskeleton [31] as well as the fatty acid software tool of Genomatix to get novel insights anabolism is downregulated by ATRA (Stoecker - 104 - et al., unpublished data), which is in accordance 6. Wan DCD, Shi Y-YY, Nacamuli RPR, with the ATRA mediated phenotype, the Quarto NN, Lyons KMK et al. 2006. profound reduction of lipid accumulation (Fig. Osteogenic differentiation of mouse adipose- 1C). This study is the first report concerning an derived adult stromal cells requires retinoic in silico based joint analysis of real microRNA acid and mRNA expression data in ATRA treated receptor type IB signaling. Proc. Natl. Acad. 3T3-L1 cells. The results indicate that ATRA Sci. USA 103: 12335-12340. and bone morphogenetic protein mediates fine-tuned changes in microRNA 7. Alric SS, Froeschlé AA, Piquemal DD, expression, which in turn modulates the mRNA Carnac GG, Bonnieu AA. 1998. Functional expression in an elaborated manner. specificity of the two retinoic acid receptor RAR and RXR families in myogenesis. ACKNOWLEDGEMENTS Oncogene 16: 273-282. The authors gratefully acknowledge the support 8. Gregoire FM, Smas CM, Sul HS. 1998. by the Faculty Graduate Center Weihenstephan Understanding of TUM Graduate Physiol. Rev. 78: 783-809. School at Technische adipocyte differentiation. Universität München, Germany. The authors 9. Green H, Meuth M. 1974. An established also thank NanoString Technologies for free of pre-adipose cell line and its differentiation in charge nCounter mouse microRNA expression culture. Cell 3: 127-133. assay processing. 10. Otto TC, Lane MD. 2005. Adipose development: from stem cell to adipocyte. REFERENCES Critical Rev. Biochem. Mol. Biol. 40: 229- 1. Albrecht E, Teuscher F, Ender K, Wegner J. 242. 2006. Growth- and breed-related changes of 11. Tang Q-Q, Otto TC, Lane MD. 2003. Mitotic marbling characteristics in cattle. J. Anim. clonal expansion: a synchronous process Sci. 84: 1067-1075. required for adipogenesis. Proc. Natl. Acad. 2. Grant AC, Ortiz-Colon G, Doumit ME, Buskirk DD. 2007. Optimization of in vitro Sci. USA 100: 44-49. 12. Rosen ED, Spiegelman BM. 2000. conditions for bovine subcutaneous and Molecular regulation of adipogenesis. Annu. intramuscular preadipocyte differentiation. J. Rev. Cell. Dev. Biol. 16: 145-171. Anim. Sci. 86: 73-82. 13. Yeh WC, Cao Z, Classon M, McKnight SL. 3. Frey SK, Vogel S. 2011. Vitamin A 1995. Cascade regulation of terminal metabolism and adipose tissue biology. adipocyte differentiation by three members Nutrients 3: 27-39. of the C/EBP family of leucine zipper 4. Safonova II, Darimont CC, Amri EZE, Grimaldi PP, Ailhaud GG, et al. 1994. proteins. Genes Dev. 9: 168-181. 14. Spiegelman BM, cell differentiation. Mol. Cell. Endocrinol. expression 104: 201-211. differentiation of 3T3 adipocytes. Cell 29: A in reproduction and development. Nutrients 3: 385-428. - 105 - prior and to actin 1982. Decreases 53-60. tubulin SR. Retinoids are positive effectors of adipose 5. Clagett-Dame M, Knutson D. 2011. Vitamin in Farmer gene morphological 15. Jump DB, Botolin D, Wang Y, Xu J, Kuri-Harcuch W. 1992. Quantitation of Christian B. 2006. Fatty acids and gene adipose conversion and triglycerides by transcription. Food Nutri. Res. 50: 5-12. staining intracytoplasmic lipids with Oil red 16. Carthew RW, Sontheimer EJ. 2009. Origins and mechanisms of miRNAs and siRNAs. O. Histochem. 97: 493-497. 25. Schneider CA, Rasband WS, Eliceiri KW. Cell 136: 642-655. 2012. NIH Image to ImageJ: 25 years of 17. Khvorova A, Reynolds A, Jayasena SD. 2003. Functional siRNAs and miRNAs image analysis. Nat. Methods 9: 671-675. 26. Stone exhibit strand bias. Cell 115: 209-216. endogenous siRNAs Bernlohr DA. 1990. The molecular basis for inhibition of adipose 18. Sontheimer EJ, Carthew RW. 2005. Silence from within: RL, conversion of murine 3T3-L1 cells by and miRNAs. Cell 122: 9-12. retinoic acid. Differentiation 45: 119-127. 27. Schwarz EJE, Reginato MJM, Shao DD, 19. Schaefer CF, Anthony K, Krupa S, Buchoff J, Krakow SLS, Lazar MAM. 1997. Retinoic Day M et al. 2009. PID: the pathway acid blocks adipogenesis by inhibiting interaction database. Nucleic Acids Res. 37: C/EBPbeta-mediated transcription. Mol. Cell D674-D679. Biol. 17: 1552-1561. 20. Friedman RC, Farh KKH, Burge CB, Bartel 28. Xue JC, Schwarz EJ, Chawla A, Lazar MA. DP. 2008. Most mammalian mRNAs are 1996. Distinct conserved targets of microRNAs. Genome revealed Res. 19: 92-105.. differentiation by stages in retinoid after adipogenesis inhibition induction of of 21. Muniategui A, Pey J, Planes FJ, Rubio A. PPARgamma. Mol. Cell Biol. 16: 1567-1575. 2012. Joint analysis of miRNA and mRNA 29. Liu P. 2002. Multiple functions of caveolin-1. expression data. Brief Bioinform. 14: 263278. J. Biol. Chem. 277: 41295-41298. 30. Kanzaki M, Pessin JE. 2002. Caveolin- 22. Smoot ME, Ono K, Ruscheinski J, Wang P- associated filamentous actin (Cav-actin) L, Ideker T. 2011. Cytoscape 2.8: new defines features for data integration and network adipocytes. J. Biol. Chem. 277: 25867-25869. visualization. Bioinform. 27: 431-432. a novel F-actin structure in 31. Takenouchi T, Miyashita N, Ozutsumi K, 23. Cline MS, Smoot M, Cerami E, Kuchinsky Rose M, Aso H. 2004. Role of caveolin-1 A, Landys N et al. 2007. Integration of and cytoskeletal proteins, actin and vimentin, biological networks and gene expression data in adipogenesis of bovine intramuscular using Cytoscape. Nat. Protoc. 2: 2366-2382. preadipocyte cells. Cell. Biol. Int. 28: 615- 24. Ramírez-Zacarías JL, Castro-Muñozledo F, - 106 - 623. 家 畜 栄 養 生 理 研 究 会 会 則 (総 則) 第一条 本会は家畜栄養生理研究会と称する。 第二条 本会は家畜栄養生理に関する研究及びその成果の応用を促進する事を目的とする。 第三条 前条の目的を達成するため下記の事業を行う。 一.家畜栄養生理の研究に関する討議(春季・秋季集談会) 二.家畜栄養生理に関する研究情報、文献の蒐集配布並びに交換 三.家畜栄養生理に関する共同研究の促進及び研究相互の連絡 四.家畜栄養に関する研究成果及び技術につき必要と認めた場合、印刷物刊行等により普及を図ること 五.その他本会の目的達成のために必要な事業 (会 則) 第四条 会員は正会員と賛助会員及び名誉会員とする。 第五条 本会の正会員になろうとするものは、会員の推薦により事務局に申し込むものとする。 2.事務局は前項の申し込みに対して可否を決定する。但し、名誉会員は評議員の推薦により、総会において決定する。 3.会員は総会及び集談会に参加し発言することができる。但し、非会員も別途定める参加費を納入すれば、集談会に参加 し、発言することができる。 4.名誉会員は評議員会に出席し、助言することができる。 第六条 会員は会費を納入するものとする。但し、名誉会員は会費を免除する。 2.正会員会費及び賛助会費は総会に於いてこれを定め、徴収方法は事務局が決定する。非会員の集談会参加費は正会員会 費と同額とする。 第七条 会員は下記の事項に該当する時は会員たる資格を失う。 一.本人の意志による退会 二.長期の会費未納の場合 三.会員として不適当と事務局が認め、総会がこれを承認した場合 但し、第一項により退会しようとする時は、事務局宛に届け出るものとする。 (役 員) 第八条 本会に役員として会長 1 名、事務局員若干名、会計監査 2 名、評議員若干名、及び編集委員若干名を置く。役員の任期 は 2 年とし、重任を妨げない。 第九条 役員は総会において選定する。 第十条 会長及び事務局員は事務局を、編集委員は編集委員会を組織する。 2.事務局は総会で議決された方針に従って会務を運営する。 3.会長は本会を代表し、事務局員・編集委員の業務を統轄する。 4.事務局員は庶務、会計その他の業務を分担する。 5.編集委員は評議員の推薦による話題提供者の論文等を査読し、編集業務を担当する。 第十一条 評議員は本会の重要な事項につき会長の諮問に応じる。 (会 議) 第十二条 会議は総会及び評議員会とする。 第十三条 総会を分けて通常総会及び臨時総会とする。 2.通常総会は毎年1回開催する。 3.総会は評議員会の議を得て会長がこれを召集する。 4.総会の議長はその都度選出するものとする。 第十四条 総会は本会の経理、人事及び事業の全般に亘る主要事項を審議決定する。 第十五条 評議員会は役員を以て構成する。 2.評議員会は会長がこれを召集し、その議長となる。 3.評議員会は原則として年 2 回開催する。 第十六条 評議員会は本会運営の基本方針を審議し、主要事項について総会へ提出し、その審議を求める。 (昭和 59 年 4 月 6 日改正) (平成 12 年 5 月 13 日改正) (平成 15 年 5 月 10 日改正) (平成 17 年 11 月 12 日一部改正) - 107 - 栄養生理研究会報編集・投稿規程 1.本会報は、家畜栄養生理に関する論文および研究会資料を掲載する。 2.掲載論文の著作権は家畜栄養生理研究会に属する。 3.評議員会は、春季集談会(自由課題形式)および秋季集談会(シンポジュウム形式)の話題を決定し、その話題 提供者を推薦する。 4.編集委員長より査読を委託された編集委員は、話題提供者等からの論文を査読し、著者に修正を求め、本研究会 報への掲載可否を判断する。 5.原稿はコンピュータソフト(MS-Word、一太郎など)を用いて作成し、ファイル(E-mail 添付ファイル、FDま たは CD-R)とA4 サイズの原稿一部と共に事務局へ提出する。図表は本文とは別ファイルとして作成し、本文中 には埋め込まない。図はモノトーンとする。電気泳動写真、顕微鏡写真などの原板は、鮮明なものを添付する。 図表を挿入する位置を本文左側の余白に朱書きで指示する。本文の句読点は “。 “、 を用いる。本文中に数 値および英文を記載する場合は半角で入力し、コンマ(,)、セミコロン(;)、コロン(:)、ピリオド(.)を適当 な位置に挿入する。 イ)評議員より推薦を受け、本研究会報への原稿執筆を了承した話題提供者は、編集委員長が指定した期日まで に原稿を事務局に提出する。 ロ)原稿は原則として返却しない。 ハ)原稿は簡易書留便で郵送する。 二)原稿提出時に著者略歴(氏名、生年月日、学歴、職歴、専門分野)を添付する。 6.原稿本文は原則として和文で書き、当用漢字を用いる。原稿はA4 サイズの用紙を用い、上下左右とも 2.5 cm の 余白を設け、35 字 25 行の横書きとする。図表はA4 紙 1 枚に 1 表、1 図を作成する。図の説明は、番号順に別 紙にまとめて記載する。一論文の原稿枚数は、図表、図の説明を含めて 25 枚以内(約 1500 字が刷り上り 1 ペー ジとなる)とする。本文には連続したページ番号を見やすい位置に記入する。行番号をページごとに付け、左側 の余白に記入する。 イ)原稿には表題、著者名、所属機関名を和文および英文で明記する。ただし英文は 1 ページ目の脚注に記す。 ロ)特殊文字は用いず、ベ夕打ちとし、原稿中には特殊文字などを赤字で明記する。 ハ)引用文献は、本文中の関連箇所に肩つきで引用順に一連番号をつけ、番号はアラビア数字を用い片カッコで 囲む。受理済みおよび印刷中のものを除き、投稿中の論文は引用文献として用いない。 二)引用文献が雑誌の場合の書き方は、著者名、年号、表題、雑誌名、巻:頁一頁の順とする。 (例) 甫立京子・浜田龍夫・前田昭二 1995. 銅とビタミン E のアマニ油含有飼料への添加が子豚臓器中の銅, ビタミン E と過酸化脂質に与える影響.日畜会報, 66:142-148. (例) Tomonaga S, Kaneko K, Kaji Y, Kido Y, Denbow DM, Furuse M. 2006. Dietary β-alanine enhances brain, but not muscle, carnosine and anserine concentrations in broilers. Anim. Sci. J., 77:79-86. ホ)単行本の記載は、著者名、発行年、書名、引用頁、発行所、発行地の順とする。 分担執筆の場合は、所収の表題、編集または監修者名を加える。 (例) 糸川嘉則 1995. マグネシウム. 33-48. 光生館. 東京. (例) 板橋久雄 2006. ルーメンにおける栄養素の代謝. ルミノロジーの基礎と応用 (小原嘉昭編), 32-50. 農文協. 東京. 7.初校は著者が行い、文章、図表の改変や追加は原則として認めない。校正の時に著しい改変や追加によって生じ た費用は著者の負担とする。 8.刷りは著者に対し 50 部までを無料贈呈とし、それ以上については著者の実費負担とする。 9.栄養生理研究会報サイズはA4 版とする。 (平成 7 年 10 月 21 日 評議員会審議) (平成 8 年 4 月 20 日 評議員会承認) (平成 11 年 10 月 30 日 評議員会承認) (平成 13 年 5 月 19 日 評議員会承認) (平成 15 年 5 月 10 日 評議員会承認) (平成 18 年 10 月 7 日 評議員会承認) (平成 19 年 3 月 28 日 評議員会承認) (平成 19 年 11 月 30 日 評議員会承認) - 108 - 家畜栄養生理研究会役員(平成 25 年度)名簿 (名誉会員)(9 名) 安保 佳一(元岩手大学農学部) 石橋 晃(日本科学飼料協会) 川島 良治(元京都大学農学部) 佐々木康之(元帯広畜産大学) 柴田 章夫(元日本大学農獣医学部) 新林 浜田 本好 渡邉 恒一(元家畜衛生試験場) 龍夫(元東京農業大学農学部) 茂一(全国家畜畜産物衛生指導協会) 泰邦(元信州大学農学部) (評議員)(40 名) 秋葉 征夫(東北生活文化大学) 佐野 宏明(岩手大学農学部) 朝井 洋(日本中央競馬会日高育成牧場) 柴田 正貴(元畜産草地研究所) 麻生 久(東北大学大学院農学研究科) 菅原 邦生(宇都宮大学農学部) 阿部 亮(畜産・飼料調査所 御影庵) 高橋 和昭(山形県立米沢女子短期大学) 新井 敏郎(日本獣医生命科学大学) 谷口 幸三(広島大学大学院生物圏科学研究科) 板橋 久雄(日本獣医生命科学大学) 寺田 文典(九州沖縄農業研究センター) 小形 芳美(NOSAI 山形) 豊水 正昭(東北大学大学院農学研究科) 奥村 純市(元名古屋大学農学部) 中川 二郎(豊橋飼料株式会社) 小堤 恭平(元畜産草地研究所) 中村 修(日本配合飼料KK) 小原 嘉昭(明治飼糧水戸研究牧場) 林 國興(元鹿児島大学農学部) △加藤 和雄(東北大学大学院農学研究科) 左 久(環境リサイクル肉牛協議会) 加藤 清雄(酪農学園大学獣医学部) 藤原 勉(元島根大学生物資源科学部) 喜多 一美(岩手大学農学部) ◎古瀬 充宏(九州大学大学院農学研究院) △木村 信熙(元日本獣医生命科学大学) 甫立 孝一(北里大学獣医学部) 久米 新一(京都大学大学院農学研究科) 松井 徹(京都大学大学院農学研究科) 小林 泰男(北海道大学大学院農学研究院) 松本 光人(農林水産・食品産業技術振興協会) 坂口 英(岡山大学農学部) 村井 篤嗣(名古屋大学大学院生命農学研究科) 佐々木晋一(元信州大学農学部) 矢野 秀雄(元京都大学大学院農学研究科) 佐藤 幹(東京農工大学) 和田 賢二(NOSAI 山形) 祐森 誠司(東京農業大学) (編集委員)(10 名) 麻生 久(東北大学大学院農学研究科) 小林 泰男(北海道大学大学院農学研究院) 加藤 和雄(東北大学大学院農学研究科) 豊水 正昭(東北大学大学院農学研究科) 喜多 一美(岩手大学農学部) ○辰巳 隆一(九州大学大学院農学研究院) 黒瀬 陽平(北里大学獣医学部) 松井 徹(京都大学大学院農学研究科) 後藤 貴文(九州大学大学院農学研究院) 谷口 幸三(広島大学大学院生物圏科学研究科) (平成 25 年 10 月 1 日修正) ◎会長 ○編集委員長 △会計監事 - 109 - 賛 助 会 員 全国酪農業協同組合連合会購買部 豊橋飼料(株)テクニカルセンター 理 研 科 学(株) (五十音順) 栄 養 生 理 研 究 会 報 第 57 巻 第 2 号 平成 25 年 10 月発行(会員頌布:年会費 4,000 円) 発行者 古瀬充宏 発行所 家畜栄養生理研究会 〒878-0201 大分県竹田市久住町大字久住字鶴が笹 4045-4 九州大学農学部附属農場・高原農業実験実習場 家畜栄養生理研究会事務局 TEL: 0974−76−1377 FAX: 0974−76−1218 E-mail: [email protected] 郵便振替:口座番号 01060−8−33558 加入者名:家畜栄養生理研究会 銀行口座:ゆうちょ銀行 店番 818 口座番号 0207245 口座開設者名:家畜栄養生理研究会 印刷所 一正堂 〒812-0053 福岡市東区箱崎 6 丁目 14-17 TEL:092−651−3771 FAX:092−651−3771
© Copyright 2026 Paperzz