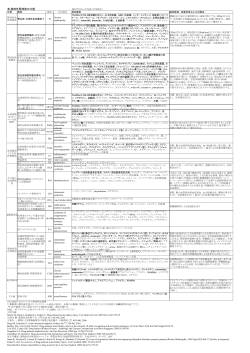
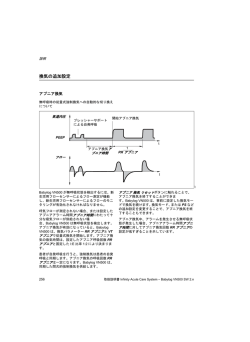
VOL.34 NO.2 2010 一般社団法人 145 Gout and Nucleic Acid Metabolism Vol.34 No.2(2010) 総説1 運動後急性腎不全(ALPE) 石川 勲 要 旨 腎性低尿酸血症は,この運動後急性腎不全 運動後急性腎不全とは,「短距離を全力疾走す (ALPE)の発生リスクとして最も重要なもので るなど無酸素運動後に,強い背腰痛を伴って発 ある.発生機序についてはまだ解明されていな 症する非ミオグロビン尿性の急性腎不全」をい い.しかし著者らは,delayed CT(造影24-48時 う.したがって,従来から知られている,マラ 後の単純CT)で,両腎に楔形の造影剤残存がみ ソンなど有酸素運動後に発生するミオグロビン られることから,無酸素運動によって筋肉から 尿性急性腎不全(Exertional rhabdomyolysis with 何らかの腎血管攣縮因子が発生し作用するから acute renal failure)とは異なるものである.また という仮説を考えている.一方,腎性低尿酸血 この「運動後急性腎不全」は症状の特徴から 症患者に起こりやすい理由としては,急性尿酸 ALPE(Acute renal failure with severe Loin pain and 腎症が起こるから,あるいは低尿酸血症で活性 Patchy renal ischemia after anaerobic Exercise)とも 酸素消去系がうまく働かないから,さらには尿 呼ばれるので,著者らは「運動後急性腎不全 酸トランスポートの異常からなど,種々の説が (ALPE)」と記載することを提唱している. ある. 運動後急性腎不全(ALPE)は患者の98%が男 性で,年齢の中央値は19歳(IQR:16∼26)と, はじめに 主に若い男性に発生する.典型例は,運動会や 最近,運動後急性腎不全が特に腎性低尿酸血 体育祭で200m走を複数回全力疾走し,その数時 症患者に多く報告されるようになり注目されて 間後に,強い背腰痛,嘔気・嘔吐を訴えて,夜 いる1).そこで,運動後急性腎不全2,3)とはどのよ 間救急外来を受診するというものである.しか うな疾患か述べたい. し強い痛みから尿路結石と診断されることが多 く,血清クレアチニンを測定しないと,急性腎 (1)運動と血清尿酸値,血清尿酸値と急性腎不 不全の診断がつかない.また多くは非乏尿性急 全 性腎不全で,尿の色に変化はなく,褐色尿も認 1)運動と血清尿酸値 めない.血清CK値は基準値内か高くても9倍以 長時間に及ぶ激しい運動,マラソン・ジョギン 内である.221例の集計によると受診時の血清ク グ・水泳などの有酸素運動では,脱水状態がな レアチニン(中央値)4.0mg/dl,最高値(中央 い限り血清尿酸値は上昇しない.これに比べ, 値)5.6mg/dlで,大部分は保存的治療で回復し 短い時間に筋肉を激しく使い,瞬発力が要求さ ているが,23%の症例では血液透析が必要になる. れる無酸素運動では脱水の有無にかかわらず血 また報告例の58%が腎性低尿酸血症患者であり, 清尿酸値が上昇する4). 浅ノ川総合病院 腎臓内科 Isao Ishikawa Key Words:急性腎不全,無酸素運動,背骨痛,腎虚血,尿酸トランスポータURAT1 146 痛風と核酸代謝 第34巻 第 2 号 (平成22年) 表2 運動後急性腎不全(ALPE)と運動性横 2)血清尿酸値と急性腎不全 紋筋融解による急性腎不全の鑑別診断3) Ejazら5)は総説の中で,尿酸と急性腎不全の発 生について述べている.それによると,高尿酸 血症で尿酸の結晶が尿細管を閉塞すると急性尿 酸腎症が起こることはもちろんだが,尿細管を 閉塞しなくても,高尿酸血症が長期間持続する と,腎血管の攣縮,抗血管新生作用,炎症促進 作用,活性酸素作用,腎自動調節能異常によっ て,急性腎不全の発生が促進されるとしている. このことから考えると,尿酸結晶による閉塞 は別として,腎性低尿酸血症患者では急性腎不 全が起こりにくいように思われるが,実際には, 運動後急性腎不全(ALPE)に限り起こりやすく1), 他の原因によっても虚血性急性腎不全が起こり 6) やすいということはない . (2)運動誘発性急性腎不全の2タイプ 運動誘発性急性腎不全には2つのタイプがあ る(表1).1つはミオグロビン尿性急性腎不全 の運動性横紋筋融解による急性腎不全7)と非ミオ グロビン尿性急性腎不全の運動後急性腎不全 (ALPE) 2,3)である.表2に鑑別点が解説してあ る. (3)ミオグロビン尿性の急性腎不全:運動性横 紋筋融解による急性腎不全 ミオグロビン尿性の急性腎不全 7-9)は,マラソ ンなど長時間の激しい運動後に脱水をともない 発生し,乏尿,赤褐色のミオグロビン尿,使用 筋肉の疼痛・圧痛(compartmental syndrome)を みる.筋肉細胞の破壊により,血清クレアチニ ン上昇,血清Creatine Kinase(CK) 高値(基準 値の20倍以上),血清ミオグロビン高値,血清電 解質異常として,高K・P・尿酸血症,著しい低 Ca血症を伴う.また回復時にはしばしば,高Ca 血症を来す. 脱水,高温多湿での運動,慣れない運動,発 症前の低K血症は発症のリスクとなる10). 表1 運動で生じる急性腎不全 (運動誘発性急性腎不全) 1)ミオグロビン尿性 運動性横紋筋融解による急性腎不全 2)非ミオグロビン尿性 =運動後急性腎不全(ALPE) a)腎性低尿酸血症を伴わない運動後急性 腎不全(ALPE) b)腎性低尿酸血症を伴う運動後急性腎不 全(ALPE) 治療は,乏尿性腎不全であるので,透析療法 が必要となることが多い.脱水の治療を行いつ つ,透析療法で尿毒症,電解質異常の補正を行 い,腎機能の改善を待つ. 腎性低尿酸血症患者が運動性横紋筋融解によ る急性腎不全を起こしやすいという報告はない. (4)非ミオグロビン尿性の急性腎不全:運動後 急性腎不全(ALPE) 運動後急性腎不全(ALPE)は,1982年著者ら によって背腰痛と腎の血管攣縮を伴う急性腎不 全として提唱され11,12),2002年ALPE(Acute renal failure with severe Loin pain and Patchy renal ischemia after anaerobic Exercise)13)としてまとめ 147 Gout and Nucleic Acid Metabolism Vol.34 No.2(2010) られた. 環として明治時代に始まり,現在も年間行事と 現在,非ミオグロビン尿性の運動誘発性急性 腎不全としては,非典型例 2,3) はあっても,運動 して小学校から大学,職場,町内会などで行わ れるが,中でも徒競走は必ず行われる種目であ 後急性腎不全(ALPE)しか知られておらず,1 り,発症の機会も多いことがあげられる. つの疾患と考えられる. 表3 有酸素運動と無酸素運動3) その根拠は,横紋筋融解症にはdelayed CTによ る楔形の造影剤残存がみられないが2,3),運動後急 性腎不全(ALPE)の症例では,血清クレアチニ ンが6mg/dl以下であれば(よくみられるのは 1.5-3.0mg/dlの間),楔形の造影剤残存がみられ るからである2,3). Delayed CTによって楔形の造影剤残存を見ると きには,患者のインフォームドコンセントを得 た後に,患者に脱水がないことを確認のうえ, 少量(40ml)の造影剤を使用し,施行する.造 2)背腰痛 影剤の投与がためらわれる場合には,感度はや 痛みは運動後3-12時間後(1-48時間後)に発症 や落ちるものの,MDP骨スキャンや14),MRI画像 し,背腰痛として訴えることが最も多い 13) が, 2,3,15) ときには腹痛,腰痛として訴えることもある. と一致する所見を得ることが出来る. ただし,激しく使用した筋肉の痛みではなく, ,造影超音波検査16)でも,楔形の造影剤残存 おそらく腎血管の攣縮による腎性狭心症(renal (5)運動後急性腎不全(ALPE)の臨床的特徴 angina)としての痛みと著者はとらえている13). 典型例の病歴と特徴を述べる2,3).若い男性が運 背腰痛は平均4日間持続する17).腹痛として訴 動会で複数回200mを全力疾走し,その夜激しい え,嘔気を伴うとしばしば急性胃腸炎と診断さ 背腰痛で夜間外来を受診する.救急診察医は腎 れることがある. 尿路結石を疑う.しかし翌日内科を受診すると, 最近Goldsteinら18)は,acute kidney injury (AKI) 血清クレアチニンが上昇しており急性腎不全と の早期マーカーについて,腎の血管攣縮(腎の 診断される.血清CK値は基準値以内か,基準値 虚血)をrenal anginaと表現し,「狭心症の早期マ をわずかに超えているぐらいである. ーカーとしてトロポニンがあるように,renal anginaにもトロポニンに準じるAKIの早期マーカ 1)運動の種類 ーを探す必要がある」と報告している.運動後 運動には有酸素運動と無酸素運動があるが 急性腎不全(ALPE)でみられる背腰痛は,腎血 (表3),運動後急性腎不全(ALPE)を起こす運 管攣縮による痛みをまさに表しており,真の 動としては,100m,200m,400m走など全力疾走 renal angina であると著者は考えている13). する無酸素運動の短距離走が最も多い(221例中 140例)17).他に,無酸素運動である重量挙げで の発症はもちろん無酸素運動の繰り返しを含む 3)斑状腎虚血 Delayed CTによって楔形の造影剤残存を確認す 運動,すなわち,サッカーや野球,バスケット, るには,血清クレアチニン1.5-3.0mg/dlの腎機 競輪,スキー,競泳などでみられる2,3).また警官 能時に造影剤40mlを静脈内投与し,1日後(数 が犯人を追いかけて発症している例もある. 時間から2日後)に単純CTを撮ると良い(図1) . わが国で運動後急性腎不全(ALPE)が多い理 血清クレアチニンが高いと造影剤残存部は広く 由の一つに,日本では運動会が富国強兵策の一 なる12,13).したがって透析が必要な重症例などで 148 痛風と核酸代謝 第34巻 第 2 号 (平成22年) 図1 楔形造影剤残存(delayed CT)と図によ 4).この中でも,腎性低尿酸血症患者でベース る説明 の血清尿酸値が1mg/dlを切るような状況がハ イリスクである6). 5)運動後急性腎不全(ALPE)診断のクライ テリア 本疾患はdelayed CTによる楔形の造影剤残存に よって発見された 12)が,以下の3つの項目が揃 ったとき本疾患と診断する2,3,13). 1)短時間の激しい運動(無酸素運動)後に 発生した急性腎不全 2)背腰痛 3)血清CK値が基準値以内か軽度上昇(基準 値上限の9倍以内) の3項目である. は,回復時で血清クレアチニン1.5-3.0mg/dlの 横紋筋融解は無いかあっても軽度なので,血 腎機能時にdelayed CTを行うと,両腎に造影剤の 清CK値や血清ミオグロビン値は基準値内か,9 楔形残存を認める2,3).またこの楔形の造影剤残存 ないし6倍までの軽度の上昇に留まり,ミオグ は腎性低尿酸血症例でもdelayed CTで同様に見ら ロビン尿は認めない13). れ 12,13),感度は落ちるもののMDP骨スキャン 14), MRI2,3,15),造影超音波検査16)でも斑状の造影剤残 (6)腎性低尿酸血症 存に一致する所見がみられる.運動誘発試験で 腎性低尿酸血症患者が,運動後急性腎不全 斑状楔形造影剤残存が証明された例も,Ohら 19) (ALPE)を起こしやすいことは,1990年Ishikawa らの「運動後急性腎不全(ALPE)の自験例1) 13 によって報告されている. これらの点から著者らは運動後急性腎不全 (ALPE)で認められる楔形の造影剤残存は,腎 例を調べ,3例が腎性低尿酸血症患者であった」 という記載に始まり1),その後,腎性低尿酸血症 虚血(血管攣縮)が両腎に斑状に現れるため, 患者は運動後急性腎不全(ALPE)を50倍起こし すなわち,斑状になるのは虚血病変に強いとこ やすいこと2,3),ほとんどの症例でベースの血清尿 ろと弱いところが存在するからと考える.そし 酸値が1mg/dl以下である6)ことがわかった. てこの腎血管の攣縮が背腰痛の原因になってい また運動後急性腎不全(ALPE)患者のうち腎 性低尿酸血症患者の占める割合は,これまでの 13) るのではないかと考えている . なお血管攣縮は腎だけに起こるが,ごくまれ 自験例23例中5例で,1990年の自験例13例中3 に,脳血管にも攣縮が起こり,ALPE症例に可逆 例と変わらなかった.しかし著者らによる報告 性後頭葉白質脳症(PRES)を合併したのではな 例の集計では,運動後急性腎不全(ALPE)201 20,21) いかという症例が2-3例報告されている . 例中,腎性低尿酸血症患者は116例(58%)と多 くを占めていた17).これは2002年Enomotoら22)に 4)運動後急性腎不全(ALPE)発生のリスク よってURAT1がクローニングされ,腎性低尿酸 ファクター 血症が注目されるようになったために,腎性低 発生のリスクを調べると,かぜ気味である, かぜ薬やNSAIDを服用後に全力疾走した,腎性 低尿酸血症患者である 13,17) などがあげられる(表 尿酸血症患者でより多く報告されているのか, 真に発生数が多いのか,今後検討が必要である. 運動後急性腎不全(ALPE)の自験例を含め, 149 Gout and Nucleic Acid Metabolism Vol.34 No.2(2010) 表4 運動後急性腎不全(ALPE)221例のまとめ17) あり なし あり なし 150 痛風と核酸代謝 第34巻 第 2 号 (平成22年) 報告例221例をまとめた17)(表4).運動後急性腎 血清尿酸値の平均が0.17mg/dlと著しく低値で, 不全(ALPE)は,主に若い男性に発生し,初診 FEUAは>150%を示し,うち3例に運動後急性腎 時,背腰痛以外に,悪心・嘔吐,微熱,CRP陽 不全の既往があったと記載されている23,41). 性を呈する例が多い.血清クレアチニン値は初 診時4mg/dl(中央値),最大値5.6mg/dlを示し, 2)外国での関心 透析必要例は23%にみられた.再発例は20%であ 腎性低尿酸血症に伴う運動後急性腎不全の症 った.両側斑状の楔形造影剤残存は,血清クレ 例は,日本・韓国以外にも,米国 42 ) ・台湾 24 ), アチニン値が1.5-3.0mg/dl時に検査を行えば, 34) ほぼ全例に見られている. いる.Sebastaらによる2010年開催のEDTAでの発 ・カナダ32)・イスラエル23,41)からも報告されて 運動後急性腎不全(ALPE)患者でも腎性低尿 表43)によると,チェコ人3,500人の検体から569人 酸血症と非腎性低尿酸血症で比較すると,腎性低 の低尿酸血症患者を検出し,この中にURAT1の 尿酸血症を伴う患者では,再発は29%と多く,初 異常を8人とURATv1(GLUT9)の異常(het- 診時の血清クレアチニン4.7mg/dl(中央値),最 erozygote2人,compound heterozygote5人, 大血清クレアチニン6.6mg/dlと非腎性低尿酸血症 homozygote2人)を9人認め,これら計17人中 患者よりいずれも高値で,透析必要例も29%と, 3人に急性腎不全と腎結石の既往があると報告 . 急性腎不全が重度になりやすかった17)(表5) した.彼らは,日本や韓国以外でも腎性低尿酸 日本では腎性低尿酸血症患者の頻度が0.1-0.4% 血症はもっと多い可能性を指摘している. と他国と比べ高いことと,先に述べた運動会の また2010年Kidney Int 5月号の"make your diag- 開催頻度が高いことが,わが国における運動後 nosis"でも本疾患が取り上げられており,外国で 急性腎不全(ALPE)の発生が多いことと関連し も本疾患に興味を持つ人が増えてきているよう ているように思われる. に思われる.しかしこれによると,Yanら34)は単 に運動後に腎性低尿酸血症患者に急性腎不全が 1)運動後急性腎不全(ALPE)とURAT1 起こったとしか記載していなかった.そこで著 (SLC22A12),GLUT9(SLC2A9)変異との 者は,Kidney Intのletter to editor44)で,「本例は単 なる運動後の急性腎不全ではなく,運動後急性 関係 URAT1(SLC22A12)遺伝子変異(遺伝性腎性 腎不全(ALPE)である.したがって腎性低尿酸 低尿酸血症タイプ1:RHUC1)としてはW258X 血症患者は他の種々の原因によっても虚血性急 homozygoteが最も多いが,W258Xとのcompound 性腎不全を起こしやすいというわけではない」 heterozygote,W258X heterozygoteの他,R90Hの と反論した. homozygoteやcompound heterozygoteの例も報告さ 21-40 ) つまりURAT1やGLUT9の変異による腎性低尿 ( 表 6 ). 著 者 ら が 経 験 し た 父 が 酸血症患者は運動後に急性腎不全を起こしやす R90H homozygoteで,息子がR90HとW258Xの いという記載は,正確には運動後急性腎不全 compound heterozygoteの例は,共にdelayed CTな (ALPE),または英語でexercise-induced acute kid- どの画像診断で両腎に楔形造影剤残存が認めら ney injury(ALPE)を起こしやすいとすべきであ れている 2,3,27) れた . 最近,URAT1の変異による運動後急性腎不全 (ALPE)の症例だけでなく,GLUT9(SLC2A9) の変異(遺伝性腎性低尿酸血症タイプ2:RHUC る.言いかえれば,腎性低尿酸血症患者は,運 動性横紋筋融解による急性腎不全を起こしやす いのではなく,著者が提唱するALPEを起こしや すいのである. 2)による運動後急性腎不全(ALPE)の症例も 報 告 さ れ た ( 表 6 ). そ れ に よ る と G L U T 9 (SLC2A9)変異でL75R homozygote を示す7人は, (7)運動後急性腎不全(ALPE)の発生機序に 対する説 Gout and Nucleic Acid Metabolism Vol.34 No.2(2010) 151 表5 腎性低尿酸血症を伴う運動後急性腎不全(ALPE)と腎性低尿酸血症を伴わない運動後急性腎不 全(ALPE)の比較(n=201)17) 悪心 152 痛風と核酸代謝 第34巻 第 2 号 (平成22年) 表6 運動後急性腎不全(ALPE)におけるSLC22A12,SLC2A9変異 153 Gout and Nucleic Acid Metabolism Vol.34 No.2(2010) の攣縮 12)を起こし,その結果運動後急性腎不全 1)運動後急性腎不全(ALPE)の発生機序 どうして運動後急性腎不全(ALPE)が発生す るのかという疑問に対しては,筋原性腎虚血説 13) (ALPE)を起こすという説である.運動を負荷 して抗酸化力に対する酸化ストレス度を調べた が最も説明しやすい(図2).しかし,無酸素運 ところ,コントロールでは3.2%しか上昇しなか 動でタイプ2筋線維から遊出し,腎に作用する ったのに対し,患児では38.5%上昇し相対的抗酸 ミオグロビン以外の血管作動性物質はまだ同定 化力の不足が示された29).しかし1例だけの報告 されていない.著者らはSELDI-TOF MSでこれら であり,方法論も含め今後の検討が必要である. の物質を調べてみたが,具体的な物質は同定さ 尿酸トランスポータ変異説は,尿酸が再吸収 れなかった2,3). されないため,これと交換に排泄される乳酸な 図2 運動後急性腎不全(ALPE)の発生機序 どのアニオンが尿細管内に蓄積し,急性腎不全 を起こすという説である.ただし,運動後急性 3) (仮説) 腎不全(ALPE)は腎性低尿酸血症を伴っても伴 わなくても,症状,検査所見,delayed CT所見に 差がなく同じ疾患であるので,尿酸トランスポ ータ変異説だけでは,腎性低尿酸血症を伴わな い運動後急性腎不全(ALPE)の病因の説明が出 来ない. 現在のところ,筋原性腎虚血説と活性酸素消 去不全・腎虚血説が支持を受けつつあるが,今 後のさらなる研究が待たれる. (8)運動後急性腎不全(ALPE)の非典型例 2)腎性低尿酸血症患者に発生しやすい機序 腎性低尿酸血症患者ではそうでない人に比べ 前述した診断基準の3項目のうち,3)のCK 値が基準値以内か軽度上昇の項は必須であるが, 50倍発生しやすい.この理由として,次の3つ 運動をした覚えがない例2,3,47)や,背腰痛を訴えな の仮説が考えられている.急性尿酸腎症説 42,45), かったという例がまれにみられる2,3).著者はこの 活性酸素消去不全・腎虚血説12,46),尿酸トランス 様な例は運動後急性腎不全(ALPE)の非典型例 ポータ変異説 26,41) である. 急性尿酸腎症説 42,45) と考えている. は,FEUAが高い腎性低尿 酸血症患者では,運動によって尿酸の産生が増 加すると,さらに多くの尿酸が尿細管の内で結 (9)運動後急性腎不全(ALPE)の治療,予後, 再発,再発予防法 晶化し尿細管を閉塞するため,急性尿酸腎症を 治療としては体液量を正常に保つ,すなわち 呈するというもので,最も考えやすい説である 脱水があれば補液を,溢水があれば水分制限を 42,45) 行い保存的にみる2,3).大部分で症例の予後は良好 発症早期の腎生検でも急性尿酸腎症の所見が1- である.ただし,重症例や,発症後に背腰痛で 2例を除き認められない.このためこの説は否 NSAIDが投与され急性腎不全が増悪した例では, 定的である. 透析療法が必要となる.報告例の23%の症例で透 .ところが,運動後急性腎不全(ALPE)では, 次に,魅力的な説として,活性酸素消去不全 46 ) 析療法が必要であった17). ・腎虚血 12)説がある.腎性低尿酸血症患者で また運動後急性腎不全(ALPE)で問題となる は,活性酸素をスカベンジする尿酸が少ないた のは再発率が高い点である.特に,腎性低尿酸 め,腎は活性酸素の影響を強く受けて,腎血管 血症患者に発生した場合は,再発率がさらに高 154 痛風と核酸代謝 第34巻 第 2 号 (平成22年) くなる17)(表5) . and acute kidney injury.N Engl J Med 361: 再発予防法として次のような点に注意する. 62-72,2009. 1)患者に,運動後急性腎不全(ALPE)を起こ 9)Clarkson PM:Exertional rhabdomyolysis and しやすい無酸素運動は,どのようなものかを十 acute renal failure in marathon runners.Sports 分説明し,これらの運動を避けさせる.2)脱 Med 37:361-363,2007. 水を予防させる.3)かぜ気味のときやNSAID 10)Zager RA:Rhabdomyolysis and myohemoglo- 服用後は全力疾走させない.4)発生したら背 binuric acute renal failure.Kidney Int 49:314- 腰痛があっても極力NSAIDを服用させない.ど うしても必要なら非麻薬性鎮痛薬を使用する. 326,1996. 11)Ishikawa I,Onouchi Z,Yuri T et al:Acute 5)再発予防の効果について明確なデータはま renal failure with severe loin pain and patchy だないが,allopurinol42)を勧める記載はある.6) renal vasoconstriction. Acute Renal Failure ベースの血清尿酸値が1mg/dl以下の著しい腎 (Ed Eliahou,H.E. )John Libbey & Company 性低尿酸血症患者に対して,部活でどの運動を Ltd London:224-229,1982. 選択すべきか,運動会などで全力疾走して良い 12) Ishikawa I, Saito Y, Shinoda A et al: か否かなど,医学的見地からの見解が現在,求 Evidence for patchy renal vasoconstriction in められている. man: observation by CT scan.Nephron 27: 31-34,1981. 文 献 13)Ishikawa I:Acute renal failure with severe loin 1)Ishikawa I,Sakurai Y,Masuzaki S et al: pain and patchy renal ischemia after anaerobic Exercise-induced acute renal failure in 3 patients exercise in patients with or without renal with renal hypouricemia. Nippon Jinzo Gakkai hypouricemia. Nephron 91:559-570,2002. Shi 32: 923-928,1990. 14)Ishikawa I,Ishii H,Saito T et al:Increased 2)Ishikawa I:Exercise-Induced Acute Renal patchy renal accumulation of 99mTc-methylene Failure-Acute Renal Failure with Severe Loin diphosphonate in a patient with severe loin pain Pain and Patchy Renal Ischemia after Anaerobic Exercise,Springer Japan,Tokyo 1-108, 2007. 3)石川勲:運動後急性腎不全(ALPE),金沢 医科大学出版局,内灘.1-65,2006. after exercise.Nephron 47:29-31,1987. 15)Kim SH,Han MC,Han JS et al:Exerciseinduced acute renal failure and patchy renal vasoconstriction:CT and MR findings.J Comput Assist Tomogr 15:985-988,1991. 4)荻野和秀,久留一郎:運動性の高尿酸血症 16)Goto S,Kono K,Fujii H et al:Contrast- と尿酸産生異常.高尿酸血症と痛風 17: induced ultrasonography with Levovist in a case 147-152,2009. of exercise-induced acute kidney injury without 5)Ejaz AA,Mu W,Kang DH et al:Could uric acid have a role in acute renal failure? Clin J Am Soc Nephrol 2:16-21,2007. 6)石川勲:リスクファクターとしての低尿酸 血症.高尿酸血症と痛風 18:59-66,2010. 7)Patel DR,Gyamfi R,Torres A:Exertional rhabdomyolysis and acute kidney injury.Phys Sportsmed 37:71-79,2009. 8)Bosch X,Poch E,Grau JM:Rhabdomyolysis rhabdomyolysis.Clin Nephrol 73:170-171, 2010. 17)石川勲:運動で生じる急性腎不全.日内会 誌 99:970-976,2010. 18)Goldstein SL,Chawla LS:Renal angina.Clin J Am Soc Nephrol 5:943-949,2010. 19)Oh KJ,Lee HH,Lee JS et al:Reversible renal vasoconstriction in a patient with acute renal failure after exercise.Clin Nephrol 66: 155 Gout and Nucleic Acid Metabolism Vol.34 No.2(2010) 297-301,2006. acute renal failure.Abstract for ASN,poster 20)Kimura T,Iio K,Imai E et al:Exercise- presentation [SA-PO002],2007. induced acute kidney injury with reversible pos- 29)Kaneko K,Taniguchi N,Tanabe Y et al: terior leukoencephalopathy syndrome.Clin Exp Oxidative imbalance in idiopathic renal Nephrol 14:173-175,2010. hypouricemia.Pediatr Nephrol 24:869-871, 21)島友子,中西浩一,戸川寛子 他:運動後 2009. 急 性 腎 不 全 と PRES( Posterior reversible 30)Mima A,Ichida K,Matsubara T et al:Acute encephalopathy syndrome)の合併を反復した renal failure after exercise in a Japanese sumo 腎性低尿酸血症の1例.日腎会誌 51:778, wrestler with renal hypouricemia.Am J Med 2009. Sci 336:512-514,2008. 22)Enomoto A,Kimura H,Chairoungdua A et 31)Ohtsuka Y,Zaitsu M,Ichida K et al:Human al: Molecular identification of a renal urate uric acid transporter 1 gene analysis in familial anion exchanger that regulates blood urate renal hypo-uricemia associated with exercise- levels.Nature 417:447-452,2002. induced acute renal failure.Pediatr Int 49: 23)Bahat H,Dinour D,Ganon L et al:Non-urate transporter 1-related renal hypouricemia and acute renal failure in an Israeli-Arab family. Pediatr Nephrol 24:999-1003,2009. 235-237,2007. 32)Ouellet G,Nolin L,Bonnardeaux A:Acute impaired renal function induced by the exercise. Ann Biol Clin Que 43:45-46, 2006. 24)Cheng C-J,Chu P,Lin Y-F et al:Renal 33)Tanaka M,Itoh K,Matsushita K et al:Two hypouricemia:An easily-neglected cause of male siblings with hereditary renal hypouricemia exercise-induced acute renal failure.202nd and exercise-induced ARF.Am J Kidney Dis Renal Conference in Northern Meeting in Taiwan Nephrology Association,2006. 42:1287-1292,2003. 34)Yan MT,Cheng CJ,Chen JS et al:The 25) Cheong HI, Kang JH, Lee JH et al: case:a young man with acute kidney injury Mutational analysis of idiopathic renal after exercise.The diagnosis:exercise induced hypouricemia in Korea.Pediatr Nephrol 20: acute kidney injury in hereditary renal 886-890,2005. hypouricemia. Kidney Int 77: 935-936, 26)Ichida K,Hosoyamada M,Hisatome I et al: Clinical and molecular analysis of patients with renal hypouricemia in Japan-influence of URAT1 gene on urinary urate excretion.J Am Soc Nephrol 15:164-173,2004. 27)Ishikawa I,Nakagawa M,Hayama S et al: Acute renal failure with severe loin pain and patchy renal ischaemia after anaerobic exercise (ALPE)(exercise-induced acute renal failure) in a father and child with URAT1 mutations beyond the W258X mutation.Nephrol Dial Transplant 20:1015,2005. 28)Kaito H,Kandai N,Kanda K et al: Genetic backgrounds in patients with exercise-induced 2010. 35)金本勝義,飛田尚美,安齋未知子 他:運 動後急性腎不全を呈した腎性低尿酸血症の1 例.日児誌 113:142,2009. 36)市田公美:急性腎不全を繰り返す低尿酸血 症.高尿酸血症と痛風 16:68-71,2008. 他:運動 37)石村春令,中下尚登,小河原悟 後に急性腎不全を呈した腎性低尿酸血症の 一例.日腎会誌 50:683,2008. 38)長場泰,佐野隆,竹内康雄 他:運動後急 性腎不全を反復した腎性低尿酸血症の一例. 日腎会誌 46:609,2004. 39)奈良健平,仁村隆,茂木睦仁 他:腎性低 尿酸血症に伴う運動後急性腎不全の1例. 156 痛風と核酸代謝 第34巻 第 2 号 (平成22年) 第186回日本内科学会東北地方会2008.9. 20, 2008. 40)藤木拓磨,犀川太,野津寛大:典型的な経 過をたどった運動後急性腎不全(ALPE)の 44)Ishikawa I:Comments on 'A young man with acute kidney injury after exercise'.Kidney Int78:1047-1048,2010. 45)Erley CM,Hirschberg RR,Hoefer W et al: CT所見とβ2MGの推移.日腎会誌 52:405, Acute renal failure due to uric acid nephropathy 2010. in a patient with renal hypouricemia. Klin 41)Dinour D,Gray NK,Campbell S et al: Wochenschr 67:308-312,1989. Homozygous SLC2A9 mutations cause severe 46)Murakami T,Kawakami H,Fukuda M et al: renal hypouricemia.J Am Soc Nephrol 21:64- Patients with renal hypouricemia are prone to 72, 2010. develop acute renal failure--why? Clin Nephrol 42)Yeun JY,Hasbargen JA:Renal hypouricemia: 43:207-208,1995. prevention of exercise-induced acute renal fail- 47)Sugimoto T,Ide R,Uzu T et al:Recurring ure and a review of the literature.Am J Kidney exercise-induced acute renal failure with usual Dis 25:937-946,1995. 43)Sebesta I,Stiburkova B,Bartl J:Diagnostic procedures for hereditary renal hypouricemia. Abstract for EDTA,poster presentation Su084, 2010. daily work.Nephrology(Carlton)12:110, 2007. Gout and Nucleic Acid Metabolism Vol.34 No.2(2010) 157 Exercise-induced acute renal failure (ALPE:Acute renal failure with severe Loin pain and Patchy renal ischemia after anaerobic Exercise) Isao Ishikawa There are two types of acute renal failure induced cases,the median serum creatinine at the time of by exercise. One is due to myoglobin and occurs after consultation was 4.0 mg/dl,the median maximal a marathon or other such prolonged exercise(exer- value of serum creatinine was 5.6 mg/dl and most tional rhabdomyolysis),and the other type is not patients recover conservatively,however,hemodialy- related to myoglobin and occurs after a dash or other sis was required in 23% of cases. Fifty-eight percent anaerobic exercise(exercise-induced acute renal fail- of reported cases show renal hypouricemia(23% of ure: ALPE).Typically,a young male takes 200- our series).Renal ischemia,which is demonstrated meter dash at an athletic meeting and then complains by patchy wedge-shaped contrast enhancement on of severe loin pain, nausea and vomiting after several delayed CT,is related to the etiology and loin pain. hours,and comes to an emergency clinic at night. Renal ischemia due to unknown metabolic substance This type of acute renal failure is called as ALPE. released from muscle during anaerobic exercise is Exercise-induced acute renal failure is ALPE,not proposed as the pathogenesis of ALPE without ample exertional rhabdomyolysis with acute renal failure. evidence.Hypotheses on the high risk for ALPE in Renal hypouricemia is the most important risk factor patients with renal hypouricemia,are acute uric acid for this exercise-induced acute renal failure(ALPE). nephropathy,poor scavenging activity of reactive The serum CK value is within the normal limits,or oxygen and abnormality of the urate transporter. less than 9 times normal.In a survey of 221 ALPE Key words:acute renal failure;acute kidney injury,anaerobic exercise,loin pain,renal ischemia, uric acid transporter 159 Gout and Nucleic Acid Metabolism Vol.34 No.2(2010) 総説2 痛風の病因遺伝子 松尾 洋孝 はじめに 痛風は,高尿酸血症に引き続いておこる生活 習慣病で,common disease(ありふれた疾患)の ことが多く,ヒトを対象とした解析,特に,ヒ トの疾患における臨床遺伝学的解析とそれに基 づく分子機能解析が不可欠である2). 1つである.激痛を伴う関節痛を生じるのみな 本総説では,上記の解析の実施により明らか らず,高血圧,虚血性心疾患,脳卒中などのリ になってきた尿酸トランスポーター遺伝子の生 スクとなることが知られている.生活習慣の欧 理学的および病態生理学的役割 3, 4)や,ゲノムワ 米化および高齢化に伴い患者数が増加している イド解析による新しい知見を含めて紹介する. が,食生活を含む生活習慣のほか,遺伝的要因 さらに,最近明らかになってきたcommon disease も関与していると考えられてきた. としての痛風の主要な病因となる遺伝子5)につい ヒトを含む霊長類の一部では尿酸分解酵素で ても紹介する. あるウリカーゼが欠損しているため,ウリカー ゼの機能が保たれているマウスのような哺乳類 1.痛風を伴う稀な先天性代謝異常症の遺伝子 と比較すると,ヒトの血清尿酸値は高値を示す 「痛風の病因遺伝子」としてこれまで報告さ ことが知られている1).また,その欠損のためヒ れてきたものは,痛風を伴う稀な先天性疾患か トにおいて尿酸はプリン代謝の最終代謝産物と ら同定されてきた6).痛風をきたしうる先天性疾 なり,腎臓や腸管から排泄される.したがって, 患としては,表1にあげたように,Lesch-Nyhan ヒトにおける尿酸の代謝,輸送動態やその異常 症候群やKelley-Seegmiller症候群などがある.と に起因する疾患については,ノックアウトマウ もに,ヒポキサンチン-グアニンホスホリボシル スなどのモデル動物を用いては解析困難である トランスフェラーゼ(HPRT)という酵素の遺伝 表1 痛風を伴う稀な先天性異常症の遺伝子 防衛医科大学校 分子生体制御学講座 Hirotaka Matsuo key words:尿酸トランスポーター,ABCG2/BCRP,ゲノムワイド関連解析(GWAS),痛風,高尿酸血症 160 痛風と核酸代謝 第34巻 第 2 号 (平成22年) 子が病因遺伝子となっているが,前者がHPRTの つ遺伝子として,URAT1遺伝子は2002年に発見 完全欠損症で,後者がHPRTの部分欠損症である された3).URAT1は腎臓特異的に発現し,近位尿 とされている.これらの酵素などの病因遺伝子 細管の管腔側に局在する尿酸再吸収トランスポ の異常により,尿酸の産生過剰などが起こるこ ーターであり,高尿酸血症治療薬であるベンズ とで,高尿酸血症及び痛風が引き起こされると ブロマロンの分子標的であることもあわせて報 されている. 告された3).このようなURAT1の生理学的な機能 このような,単一遺伝子の異常に伴う疾患は 「単一遺伝子疾患(monogenic disease)」または「メ ンデル遺伝病(Mendelian disease)」と呼ばれる. は,URAT1/SLC22A12が腎性低尿酸血症1型の病 因遺伝子として同定されたことに基づいており, 自衛隊熊本病院の症例解析により証明された3). そのほとんどが青年期までに発症し,神経症状 腎性低尿酸血症においては,合併症としての など他の随伴症状を認めることもあるが,患者 尿路結石や運動後急性腎不全 7, 8)が臨床上の問題 数は少なく一般臨床の場で診る機会は多くはな となる.URAT1/SLC22A12遺伝子における腎性低 い.すなわち,common diseaseの1つである,一 尿酸血症1型の病因変異としては,機能が全く 般的な痛風の遺伝的原因にはなりにくいと考え 消失するW258X(G774A)変異が日本人に最も られる. 多く,74.1%を占めていた9).このW258X変異は, URAT1タンパク質の258番目のアミノ酸であるト 2.血清尿酸値に関連する遺伝子 リプトファン(W)が終止コドン(X)となるナ 1)腎性低尿酸血症1型の病因遺伝子URAT1/ ンセンス変異と呼ばれる変異であり,URAT1の分 SLC22A12 子機能が完全に消失することがわかっている. 痛風の遺伝要因としては,痛風を引き起こす W258X変異は日本人において頻度の高い一塩基多 先天性代謝異常症の研究が進んでおり,上述の 型(single nucleotide polymorphism, SNP)であり, ように,プリン代謝に関わる酵素などの遺伝子 アレル頻度は2.30∼2.37%と報告されている 2,10). が明らかになっている.common diseaseとしての このことは,染色体100本(50人相当)あたり2 痛風の遺伝要因の探索のためには,それらの酵 本程度にW258X変異を認めるということを意味 素を含む代謝系の分子を対象とする研究に加え, している.日本人の腎性低尿酸血症ではそのほ 尿酸の輸送系に関わるトランスポーター分子を とんどにURAT1の変異が認められるが,一部に 対象とした研究の重要性が最近注目されている. URAT1の変異を認めない症例が存在することも この血清尿酸値を調節するトランスポーター分 報告されており 9,11),URAT1以外の腎性低尿酸血 子には,多型性の高いものもあり,また,血清 症の病因遺伝子が存在することが示唆されてい 尿酸値を高める作用のある分子と下げる作用の た.痛風症例を対象とした症例対照研究では, ある分子が存在する.それぞれ低尿酸血症や高 W258X(G774A)が認められる場合には,痛風 尿酸血症の候補病因分子となるが,これまでの になりにくいことも報告されている12). 研究で,これらの尿酸関連疾患の病因遺伝子で あり,かつ生理学的な血清尿酸値の調整で重要 2)腎性低尿酸血症2型の病因遺伝子GLUT9/ な役割を担う尿酸トランスポーター分子の実体 SLC2A9 が明らかになってきた. ヒトゲノム情報の解読後,ゲノムワイド関連 血清尿酸値を調節する遺伝子として同定され 解析(genome-wide association study, GWAS)に たのは,Urate transporter 1(URAT1/SLC22A12) よる疾患関連遺伝子の探索が盛んに行われるよ 遺伝子が初めてであった.解読されたばかりの うになった.血清尿酸値に関わるGWASも, ゲノム情報概要版を用いて有機アニオントラン 2007年以降,複数のグループにより実施され, スポーター遺伝子OAT4/SLC22A11と相同性をも 尿酸値の変動に関与する遺伝子としてGlucose 161 Gout and Nucleic Acid Metabolism Vol.34 No.2(2010) transporter 9(GLUT9/SLC2A9)が報告された13-16). ロンにより抑制されることも報告している 15) . これにより,GLUT9がヒトにおいて生理学的に GLUT9による尿酸輸送能は,その後の報告でも 重要な尿酸トランスポーターの候補であること 確認され4,17,18),従来,主要な輸送基質と考えられ が示された.最初のGWASの報告は,4,731名の ていたD-グルコース,D-フルクトースなどの糖 イタリアのサルデーニャ人を対象としたLiらの 輸送活性よりも尿酸輸送活性の方が数十倍高い 報告によるもので,初めてGLUT9遺伝子と血清 ことが示されている17). 尿酸値変動との関連が明らかにされた13).Liらの GLUT9/SLC2A9が尿酸値の変動に関与すること 報告後も,血清尿酸値変動とGLUT9のSNPに関 に加えて,過去の報告で近位尿細管における 連があるという報告が相次ぎ14-16),血清尿酸値を GLUT9の発現が示されていることから, 指標とするGWASにおいてはGLUT9が最も有意 GLUT9/SLC2A9遺伝子が腎性低尿酸血症の第2の な相関を示すことが確認された(表2). 病因遺伝子である可能性が示唆されていた. Vitartらは,GLUT9が尿酸を輸送することを 我々は過去10年間にわたる85万セットの健康診 GWASの報告の際に初めて記載し,さらに,そ 断データを有する海上自衛隊の健康診断データ の輸送動態(Km値,890 μM)についても明ら ベースを活用することにより,十分な症例数を かにした15).また,VitartらはURAT1と比べて緩 確保した上で,GLUT9遺伝子を対象とした低尿 やかではあるが,GLUT9の機能がベンズブロマ 酸血症の臨床遺伝学的解析を実施した.その詳 表2 血清尿酸値変動を対象とした主なゲノムワイド関連解析 162 痛風と核酸代謝 第34巻 第 2 号 (平成22年) 細は他の総説に記したが 2,19),この解析により腎 風の関連も複数の施設の症例対照研究において 性低尿酸血症を来す2つの機能消失型のミスセ 示されているが,その分子機構は明らかにされ ンス変異(R198CとR380W)を見いだすことが ておらず,今後の研究の進展が期待される. でき,かつGLUT9がその生理学的機能として, ヒトの近位尿細管における尿酸の再吸収という 3.痛風の主要病因遺伝子ABCG2/BCRP 役割を担っていることを示すことができた4).こ 1)なぜABCG2は有望な候補遺伝子であったか? の腎性低尿酸血症の新規病因遺伝子GLUT9の同 2004年に台湾の研究グループにより報告され 定により,既知病因遺伝子であるURAT1変異に たゲノムワイド連鎖解析により,ヒトの第4染色 よるものが「腎性低尿酸血症1型」(RHUC1, 体長腕に未知の痛風遺伝子が存在する候補領域 renal hypouricemia type 1, MIM 220150),GLUT9 があることが報告された 22).そのため,我々は, 変異によるものが「腎性低尿酸血症2型」 (RHUC2, この領域に存在する複数のトランスポーターの renal hypouricemia type 2, MIM 612076)と初めて うち,個人差が大きく,かつ尿酸と構造が類似 分類されるようになった2). したAIDS治療薬3'-azido-3'-deoxythymidine (AZT) URAT1変異以外の病因による腎性低尿酸血症2 などの物質を輸送するトランスポーターの遺伝 型の概念が確立したことにより2),腎性低尿酸血 子であるATP-binding cassette transporter G2 症の重要な合併症である運動後急性腎不全が, (ABCG2/BCRP)が最も有望な候補であると考え URAT1の機能低下が原因であるのか、あるいは た.初期のGWASでは,GLUT9遺伝子のみが尿 腎性低尿酸血症という病態そのものによるもの 酸値と関連する遺伝子として検出される報告が かを検討することが可能となった.最近, 相次いたが,表2に示すように,さらに多くの GLUT9のホモ病因変異を認める海外の複数の症 サンプル数を扱ったGWASにより,GLUT9以外 例が報告され,血清尿酸値が1.0 mg/dl以下となり, に,ABCG2を含む遺伝子領域が尿酸値の変動に 尿中尿酸排泄率(FEUA)が150%以上であった 関わることが報告され23-25),GWASの結果からも ことが報告された .すなわちGLUT9の尿酸再吸 ABCG2は極めて有望な痛風の候補遺伝子である 収における影響力はURAT1と比べても高いこと ことが示唆された. 20) が示唆された.さらに,この家族例を含む2種 類のホモ変異症例において,運動後急性腎不全 2)高容量性尿酸トランスポーターABCG2 や尿路結石の合併が報告されたことにより,病 ABCG2の分子機能の解析のために,我々は細 因遺伝子の種類にかかわらず,腎性低尿酸血症 胞 膜 小 胞 ( ベ シ ク ル ) の 解 析 系 を 用 い 5), という病態により,運動後急性腎不全をきたす Woodwardらはアフリカツメガエル卵母細胞の解 ことが明らかとなった20). 析系を用いて 26),それぞれ独立にABCG2が尿酸 これまでの解析により,URAT1及びGLUT9の を輸送することを示した.我々が採用した細胞 両遺伝子に変異を認めない腎性低尿酸血症例が 膜小胞を用いた分子機能解析は,ABCトランス 存在することも確認されており,今後,未知の ポーターの解析において一般的な方法であり, 病因遺伝子異常による「腎性低尿酸血症3型」 ABCG2をHEK293細胞に発現させたのち,細胞膜 (RHUC3, renal hypouricemia type 3)が見いだされ 小胞を調製してRIで標識した基質の輸送を評価 る可能性がある 2,19).このほか,低尿酸血症の1 することにより実施した.前述のように,尿酸 例にGLUT9のP412R変異を認めたという報告 が と構造が類似した物質であるAZTをABCG2が輸 あるが,報告された機能変化の程度が小さいこ 送することが分かっているため,まずは既知の と18),および機能解析結果が別のグループにより 輸送基質である硫酸エストロンの輸送に対する, 再現できていないこと4)から,今後の検討が必要 AZTと尿酸の阻害効果を解析した.その結果, とされている .また,GLUT9遺伝子のSNPと痛 ABCG2による硫酸エストロンの輸送は,AZTに 18) 21) 163 Gout and Nucleic Acid Metabolism Vol.34 No.2(2010) 図1 高容量性尿酸トランスポーターとしてのABCG2(文献5より引用,改変) より濃度依存性に阻害されるが,尿酸によって も同様な阻害が観察されることが分かった. ABCG2において尿酸の輸送能を解析すると,生 理的に到達しうる尿酸濃度においても輸送飽和 の生じない,高容量性の尿酸輸送能が観察され た(図1).前述のように,ABCトランスポータ ーの機能解析法としては,細胞膜小胞を用いた 解析系が適しており,Woodwardらが採用した卵 母細胞の解析系では濃度依存性やATP依存性を 調べることは困難である.そのため,我々は細 胞膜小胞を用いた解析を実施することにより ABCG2が高容量性尿酸排泄トランスポーターで あることが初めて示すことができた5). 3)ABCG2遺伝子における病因変異候補の同定 ABCG2において病態に関わる遺伝子変異を見 いだすために,高尿酸血症症例90名において ABCG2遺伝子の全コーディング領域を対象とし たリシークエンスを実施した.これにより,図 2Aに示すようなアミノ酸置換を伴う6つの変異 ( V12M, Q126X, Q141K, G268R, S441N, F506SfsX4)が見いだされた.細胞膜小胞を用い た輸送実験系でABCG2の変異体における尿酸輸 送を計測すると,V12M以外の5変異で機能の低 下が認められた.この5変異のうち,Q141Kでは 機能が半分に減少し,残りの4変異では機能が完 全に消失することが観察された(図2B).また, 図2 アミノ酸置換を伴うABCG2の変異 A,ABCG2のトポロジーモデルと特異部位 B,ABCG2変異体による尿酸輸送能 (文献5より引用,改変) 164 痛風と核酸代謝 第34巻 第 2 号 (平成22年) これらの5変異のうち,Q126X とQ141Kはそれぞ れ日本人の5.5%及び53.6%と高い頻度で認めら れることが報告されており,以降の臨床遺伝学 的解析はこの2変異に注目して実施した. 4)ヒトの生理学的な尿酸排泄を司るABCG2ト ランスポーター ABCG2の生体における機能を検討するために, 739名の日本人の健康診断受診者のサンプルを用 いて,血清尿酸値とABCG2の変異による関係に ついて量的形質座位(QTL, Quantitative trait locus) 解析を実施した.その結果,機能半減変異であ るQ141K変異を有する数が多いほど,血清尿酸 値が上昇することがわかった(図3A-C). これらの所見と,ABCG2トランスポーターの 腎臓27),肝臓および小腸28)における発現パターン から,我々は図4に示したようなABCG2によるヒ ト腎臓および肝臓,腸管からの尿酸排泄機構にお いて,生理学的なモデルと病態生理学的なABCG2 機能不全モデルを提唱することができた .すな 5) わち,生理学的なモデルにおいては,腎臓の近 図3 ABCG2の機能低下型SNP(Q141K)によ る血清尿酸値の上昇 C/Cは野生型,C/Aはヘテロ変異,A/Aはホモ変異 を示す(文献5より引用,改変) 図4 ABCG2を介したヒト尿中及び糞中への尿酸排泄機構 (文献5より引用,改変) 165 Gout and Nucleic Acid Metabolism Vol.34 No.2(2010) 表3 ABCG2の機能低下による痛風発症リスクの顕著な上昇 下線はリスク変異を示す(文献5より引用,改変) 位尿細管,肝細胞および小腸上皮細胞の管腔側 したリスクであることが分かった5).したがって, に局在するABCG2は,それぞれ尿中および糞中 これらの2つの変異を調べるだけでヒトのABCG2 への尿酸排泄を担っている.肝細胞からの胆汁 トランスポーターの機能低下の程度をほぼ予測 中への尿酸排泄は,小腸上皮細胞からの小腸の できることがわかり,簡便な検査によりリスク 管腔内への尿酸排泄とともに,糞中への尿酸排 の予測が可能であることが見いだされた. 泄(腸管排泄)に関与していると考えられる. このような解析の結果,痛風の症例の10%に ABCG2機能不全モデルにおいては,近位尿細管, ABCG2トランスポーターの機能が4分の1以下に 肝細胞および小腸上皮細胞の管腔側における尿 なる遺伝子変異パターンが認められ,痛風の発 酸排泄の障害があり,この機能不全により血清 症リスクを約26倍高めることが明らかとなった 尿酸値が高まることが示唆された.ヒトにおい (表3).また,ABCG2トランスポーターの機能 て尿酸は,3分の2が腎臓から尿中へ,残りの3分 低下が,日本人の痛風症例の約8割に見られるこ の1は主に小腸などから糞中へ排泄されることが とがわかり,3倍以上の発症リスクを認めること 教科書的にも記載されていたが,ABCG2がそれ が分かった5) (図5). らの尿酸排泄を担う分子的実体であることが示 唆された. 生活習慣病などのcommon diseaseを対象とした これまでのゲノムワイド関連解析などで同定さ れる疾患関連遺伝子は,通常,リスクの増加が 5)痛風の主要病因遺伝子としてのABCG2 2倍以下のものがほとんどであった.したがっ 痛風や高尿酸血症の発症におけるABCG2トラ て,生活習慣病の遺伝子解析において,上記の ンスポーターの役割を解析するために,日本人 知見は,ABCG2遺伝子が痛風の主要病因遺伝子 男性の痛風症例161例を含む228名の高尿酸血症 であることを示しており,疾患の病態解明にせ 症例と,血清尿酸値が正常な日本人男性865名を まる日本発の大きな成果となった. 対象として,ABCG2の主な遺伝子多型について 激しい国際競争の中,Woodwardらも,前述の 検索した.その結果,Q126XというABCG2の遺 卵母細胞の解析系を用いてQ141Kにより輸送機 伝子多型は,高尿酸血症と痛風の両方の発症リ 能が低下することを報告した 26).しかしながら, スクを増加させることがわかり,高尿酸血症の 彼らの解析では痛風のリスクは2倍以下にとど 発症と比べて,痛風の発症により強く関わるこ まり,これまでの他の生活習慣病での成果と大 とが明らかとなった.また,ハプロタイプ頻度 きく変わらないものであった.我々,日本の研 解析という方法により,ABCG2遺伝子のQ126X 究グループは,より包括的な研究アプローチを とQ141Kという2つの変異は,1つの染色体上で 独立して実施したことにより,最終的に頻度の は同時に存在しないことが明らかとなり,独立 比較的高い2つのSNPの組み合わせにも注目する 166 痛風と核酸代謝 第34巻 第 2 号 (平成22年) 図5 ABCG2の機能低下と痛風発症の関係 下線はリスク変異を示す(文献5より引用,改変) ことができた.さらに,それらの遺伝子型だけ でなくヒトの個体における分子機能に着目する もとに今後解明されていくものと期待される. 2009年にはKolzらが,これまでのGWASの成 ことにより,痛風症例の8割に3倍から26倍の発 果をもとにして,2万8千人以上を対象としたメ 症リスクを認めることを見出すことができた. タ解析を実施し,血清尿酸値の変動に関わるさ これにより,痛風の主要な病因遺伝子の解明を らに多くの遺伝子群が報告された24).この報告で 含む重要な知見を報告することができた . は,これら3つの遺伝子領域のほかに,新たに, 5) 6つの遺伝子領域が見いだされた.トランスポ 6)ABCG2以外の重要な遺伝子の存在 ーター遺伝子の領域としては,URAT1/SLC22A12, 2008年末にDehghanらにより報告されたGWAS OAT4/SLC22A11, MCT9/SLC16A9が挙げられ,そ では,GLUT9以外に,ABCG2,SLC17A3を含む遺伝 の他に,PDZK1,GCKR,LRRC16A-SCGNといっ 子領域が血清尿酸値の変動に関わることが報告され た様々な遺伝子領域が報告された(表2).上記 .連鎖不平衡の問題があり,特に後者の た (表2) のうち,LRRC16A-SCGN以外は,その後のrepli- 遺伝子領域は,SLC17A3_SLC17A1_SLC17A4 と複数 cation studyにおいても血清尿酸値への影響の再現 のトランスポーター遺伝子を含む領域にまたが 性が認められている 32).Kolzらの報告において, っているため,どのトランスポーター遺伝子が URAT1のSNPと血清尿酸値変動との関わりが, 血清尿酸値の変動において,より生理学的に重 GWASにおいても初めて報告された24). 23) 要であるのか,GWAS後のさらなる詳細な解析 PDZドメインタンパク質PDZK1は,URAT1を が必要である.そのうち,NPT1/SLC17A1につい はじめとするトランスポーターと結合してその ては,その遺伝子のSNPが痛風の発症に関連して 機能を高めることが報告されており33),尿酸トラ い る こ と に つ い て Uranoら 29) が 報 告 し て い る . ンスポートソーム(尿酸輸送分子複合体)にお NPT1/SLC17A130),NPT4/SLC17A331)ともにそれぞ ける尿酸輸送調節機構の解明につながることが れ尿酸を輸送することが最近報告されており, 期待される.OAT4についても尿酸輸送活性があ SLC17A3_SLC17A1_SLC17A4の遺伝子領域におい ることが既に示されており34, 35),高尿酸血症や低 てどの分子が重要であるのか,これらの知見を 尿酸血症などの疾患との関連が解明されていく 167 Gout and Nucleic Acid Metabolism Vol.34 No.2(2010) ものと考えられている.その他の遺伝子につい も記載されていた腎外排泄(腸管排泄)の生理 ては,尿酸動態との関連が不明である.GCKR 学的分子機構がABCG2により担われていること (glucokinase regulatory protein)はグルコースセン も示唆された37).これらの知見は,尿酸動態の生 サーとして作用する解糖系酵素の調節因子であ 理学的な分子機構や,痛風や高尿酸血症の病態 り,2型糖尿病を対象としたGWASにおいて 解明をさらに進めるものであり,さらに今後の GCKR遺伝子の同じSNPが中性脂肪値の変動に関 ABCG2以外の遺伝子における研究の発展も期待 連することが報告された36). されることから,新たな視点からの痛風の予防 最近,Kamataniらにより,日本人における GWASの 結 果 も 報 告 さ れ , URAT1, GLUT9, 法や治療薬の開発につながることが大いに期待 される. ABCG2が血清尿酸値と関連することが示される と と も に , 新 た な 遺 伝 子 と し て low density 文 献 lipoprotein receptor-related protein 2(LRP2)と尿 1)Wu XW, Lee CC, Muzny DM et al : Urate oxi- 酸値との関連も指摘される25)など,これらの遺伝 dase: primary structure and evolutionary implica- 子と尿酸関連疾患との関係についても,今後の tions. Proc Natl Acad Sci U S A 86: 9412-9416, 研究の進歩が期待される. 1989. 2)松尾洋孝: 尿酸の再吸収機構と輸送体病−ゲ おわりに ヒトゲノムの解読後のポストゲノムシークエ ノ ム ワ イ ド 関 連 解 析 後 の 新 展 開 . Annual Review 腎臓 2010. 御手洗哲也, 東原英二, 秋 ンス研究として重要な位置づけにあるGWASな 澤忠男ら(編). 中外医学社 pp9-20, 2010. どのゲノムワイド解析により,疾患に関連する 3)Enomoto A, Kimura H, Chairoungdua A et al : 遺伝子が次々と同定されているが,その後の病 Molecular identification of a renal urate anion 態解明は困難な場合もある.GWASの成果を効 exchanger that regulates blood urate levels. 率的に引き出す重要なポイントの1つは,対象 となる遺伝子の機能を評価し,そのSNPによる影 Nature 417: 447-452, 2002. 4)Matsuo H, Chiba T, Nagamori S et al : Mutations 響を適切に判定できることであると考えられる. in glucose transporter 9 gene SLC2A9 cause そのため,分子機能の評価法が確立しており, renal hypouricemia. Am J Hum Genet 83: 744- かつ,様々な疾患の病態に密接に関わるトラン 751, 2008. スポーター分子は格好のターゲットとなる.今 5)Matsuo H, Takada T, Ichida K et al : Common 回紹介した,痛風のリスクを著明に高める尿酸 defects of ABCG2, a high-capacity urate 排泄トランスポーターABCG2遺伝子のSNPsの同 exporter, cause gout: a function-based genetic 定は,痛風を含めたcommon diseaseを対象とした analysis in a Japanese population. Sci Transl 「個人差に応じた早期予防や早期治療法」(テー ラーメイド医療)の確立のためにも,極めて重 要な知見になると考えられる. ABCG2遺伝子が,痛風の主要病因遺伝子であ るとともに,高容量性の尿酸排泄トランスポー Med 1: 5ra11, 2009. 6)Zaka R, Williams CJ:New developments in the epidemiology and genetics of gout. Curr Rheumatol Rep 8: 215-223, 2006. 7)Ishikawa I: Acute renal failure with severe loin ターをコードしていることもあわせて解明され, pain and patchy renal ischemia after anaerobic ABCG2がヒトの生理学的な尿酸排泄に関わるこ exercise in patients with or without renal とが示唆された.さらに,ABCG2が腎臓のみな hypouricemia. Nephron 91: 559-570, 2002. らず,肝臓や小腸にも発現していることから, 8)Ishikawa I, Nakagawa M, Hayama S et al : Acute 尿酸の腎臓からの排泄のみならず,教科書的に renal failure with severe loin pain and patchy 168 痛風と核酸代謝 第34巻 第 2 号 (平成22年) renal ischaemia after anaerobic exercise (ALPE) 17)Caulfield MJ, Munroe PB, O'Neill D et al : (exercise-induced acute renal failure) in a father SLC2A9 is a high-capacity urate transporter in and child with URAT1 mutations beyond the W258X mutation. Nephrol Dial Transplant 20: 1015, 2005. 9)Ichida K, Hosoyamada M, Hisatome I et al : Clinical and molecular analysis of patients with renal hypouricemia in Japan-influence of URAT1 gene on urinary urate excretion. J Am Soc Nephrol 15: 164-173, 2004. humans. PLoS Med 5: e197, 2008. 18)Anzai N, Ichida K, Jutabha P et al : Plasma urate level is directly regulated by a voltage-driven urate efflux transporter URATv1 (SLC2A9) in humans. J Biol Chem 283: 26834-26838, 2008. 19)松尾洋孝, 市田公美: GLUT9の異常症. 高尿酸 血症と痛風18: 84-89, 2010. 20) Dinour D, Gray NK, Campbell S et al : 10)Iwai N, Mino Y, Hosoyamada M et al : A high Homozygous SLC2A9 mutations cause severe prevalence of renal hypouricemia caused by renal hypouricemia. J Am Soc Nephrol 21: 64- inactive SLC22A12 in Japanese. Kidney Int 66: 72, 2010. 935-944, 2004. 11) Wakida N, Tuyen DG, Adachi M et al : 21)金井好克: 尿酸排泄異常の成因 尿酸トランス ポーター. 高尿酸血症と痛風 17: 21-27, 2009. Mutations in human urate transporter 1 gene in 22) Cheng LS, Chiang SL, Tu HP et al : presecretory reabsorption defect type of familial Genomewide scan for gout in Taiwanese aborig- renal hypouricemia. J Clin Endocrinol Metab 90: ines reveals linkage to chromosome 4q25. Am J 2169-2174, 2005. Hum Genet 75: 498-503, 2004. 12)Taniguchi A,Urano W,Yamanaka M et al:A 23) Dehghan A, K ttgen A, Yang Q et al : common mutation in an organic anion trans- Association of three genetic loci with uric acid porter gene,SLC22A12,is a suppressing fac- concentration and risk of gout: a genome-wide tor for the development of gout.Arthritis association study. Lancet 372: 1953-1961, 2008. Rheum 52:2576-2577,2005. 24)Kolz M, Johnson T, Sanna S et al : Meta-analysis 13)Li S, Sanna S, Maschio A et al : The GLUT9 of 28,141 individuals identifies common variants gene is associated with serum uric acid levels in within five new loci that influence uric acid con- Sardinia and Chianti cohorts. PLoS Genet 3: e194, 2007. centrations. PLoS Genet 5: e1000504, 2009. 25)Kamatani Y, Matsuda K, Okada Y et al : 14)D ring A, Gieger C, Mehta D et al : SLC2A9 Genome-wide association study of hematological influences uric acid concentrations with pro- and biochemical traits in a Japanese population. nounced sex-specific effects. Nat Genet 40: 430436, 2008. Nat Genet 42: 210-215, 2010. 26)Woodward OM, K ttgen A, Coresh J et al : 15)Vitart V, Rudan I, Hayward C et al : SLC2A9 is Identification of a urate transporter, ABCG2, a newly identified urate transporter influencing with a common functional polymorphism caus- serum urate concentration, urate excretion and ing gout. Proc Natl Acad Sci U S A 106: 10338- gout. Nat Genet 40: 437-442, 2008. 10342, 2009. 16) McArdle PF, Parsa A, Chang YP et al : 27)Huls M, Brown CD, Windass AS et al : The Association of a common nonsynonymous vari- breast cancer resistance protein transporter ant in GLUT9 with serum uric acid levels in old ABCG2 is expressed in the human kidney proxi- order amish. Arthritis Rheum 58: 2874-2881, mal tubule apical membrane. Kidney Int 73: 220- 2008. 225, 2008. 169 Gout and Nucleic Acid Metabolism Vol.34 No.2(2010) 28)Maliepaard M, Scheffer GL, Faneyte IF et al : 33)Anzai N, Miyazaki H, Noshiro R et al : The mul- Subcellular localization and distribution of the tivalent PDZ domain-containing protein PDZK1 breast cancer resistance protein transporter in regulates transport activity of renal urate-anion normal human tissues. Cancer Res 61: 3458- exchanger URAT1 via its C terminus. J Biol 3464, 2001. Chem 279: 45942-45950, 2004. 29)Urano W, Taniguchi A, Anzai N et al : Sodium- 34)木村弘章, 市田公美, 細山田真 他: 近位尿細管 dependent phosphate cotransporter type 1 管腔膜側に存在するヒト有機陰イオントラ sequence polymorphisms in male patients with ンスポーターhOAT4(human Organic Anion gout. Ann Rheum Dis 69: 1232-1234, 2010. Transporter 4)における尿酸輸送の解析. 痛 30)Iharada M, Miyaji T, Fujimoto T et al : Type 1 風と核酸代謝 25: 113-120, 2001. sodium-dependent phosphate transporter 35)Hagos Y, Stein D, Ugele B et al : Human renal (SLC17A1 Protein) is a Cl--dependent urate organic anion transporter 4 operates as an asym- exporter. J Biol Chem 285: 26107-26113, 2010. metric urate transporter. J Am Soc Nephrol 18: 31)Jutabha P, Anzai N, Kitamura K et al : Human sodi- 430-439, 2007. um phosphate transporter 4 (hNPT4/SLC17A3) as a 36)Saxena R, Voight BF, Lyssenko V et al : common renal secretory pathway for drugs and Genome-wide association analysis identifies loci urate. J Biol Chem 285: 35123-35132, 2010. for type 2 diabetes and triglyceride levels. 32)van der Harst P, Bakker SJ, de Boer RA et al : Replication of the five novel loci for uric acid Science 316: 1331-1336, 2007. 37)松尾洋孝, 高田龍平, 市田公美 他: 痛風の主要 concentrations and potential mediating mecha- な病因遺伝子ABCG2の同定. 実験医学28: nisms. Hum Mol Genet 19: 387-395, 2010. 1285-1289, 2010. 171 Gout and Nucleic Acid Metabolism Vol.34 No.2(2010) 原著1 尿酸トランスポーターURAT1トランスジェニックマウスにおける 尿酸の体内動態 塚田 愛1) 安西 尚彦2) 木村 徹2) 市田 公美1) Jutabha Promsuk2) 櫻井 裕之2) 尿酸はヒトにおけるプリン体の最終代謝産物 子はTgマウスにおいて発現が有意に増加してい であり,主に腎臓から排泄される.腎臓での尿 たが,他の尿酸トランスポーターや尿酸代謝酵 酸の排泄亢進や排泄低下によって腎性低尿酸血 素の遺伝子の発現量に変化は見られなかった. 症や高尿酸血症が引き起こされるため,腎臓で 以上の結果より,URAT1過剰発現は,マウス生 の尿酸輸送を理解することは臨床的に重要であ 体内での尿酸動態および他の尿酸トランスポー る.尿酸トランスポーターURAT1は,腎臓近位 ターや尿酸代謝酵素の遺伝子発現に大きな影響 尿細管管腔側において尿酸再吸収を行う分子で を及ぼさないことが示された. ある.このURAT1は尿酸降下薬の作用点であり, また遺伝子変異により腎性低尿酸血症をきたす. 緒 言 よってURAT1は血中尿酸値に大きく影響を及ぼ 尿酸はヒトにおけるプリン体の最終代謝産物 す因子であると考えられるが,その発現が尿酸 である.体内で合成された内因性プリン体や食 値や他の尿酸トランスポーターにどのような影 事から得られる外来性プリン体は,ヒポキサン 響を及ぼすのか明らかになっていない.そこで, チン,キサンチンを経て尿酸に代謝される.多 URAT1トランスジェニック(Tg)マウスを用い くの哺乳類では,尿酸はウリカーゼにより水溶 て,尿酸値測定およびマイクロアレイと定量 性が高いアラントインへ代謝されるが,ヒトを PCRによる遺伝子発現変化の解析を行った. 含む霊長類ではウリカーゼを遺伝的に欠損して RT-PCRの結果から,Urat1 mRNAが腎臓特異 いるため,尿酸がプリン体の最終代謝産物とな 的に発現しており,Tgマウスでは野生型と比べ る1).従って,ヒトの血中尿酸値は他の哺乳類に て発現量の上昇が見られた.ウエスタンブロッ 比べて高値を示すことが知られている. ト お よ び 免 疫 組 織 染 色 に よ り , 導 入 し た HA- 尿酸は主に肝臓で合成され,その約70%が腎 mURAT1タンパク質が腎臓の近位尿細管管腔側 臓,残りの約30%が小腸から排泄される.腎臓に に発現していることを確認した.以上の結果か おいて糸球体濾過された尿酸は,尿細管におい ら,導入したHA-mURAT1がすでに報告されてい て再吸収と分泌が行われ,最終的に糸球体ろ過 るmURAT1と同様の発現分布を示し,URAT1が 量の約10%が尿中に排泄される.血中尿酸値は 過剰発現していることを確認できた.そこで, 生体内における尿酸の産生と排泄のバランスに このマウスをモデルとして解析を行った.血中 よって決定されるが,痛風患者の半数以上が尿 および尿中尿酸値を測定した結果,どちらの尿 酸排泄の低下が原因であり,尿酸産生亢進との 酸値にも変化が見られなかった.マイクロアレ 混合型を合わせると85%以上の患者に尿酸排泄 イおよび定量PCRにて検討した結果,Urat1遺伝 の低下が見られる 2).血液中の尿酸値は通常2.8- 受付:2010年6月7日,受理:2010年10月6日 1)東京薬科大学病態生理学 Ai Tsukada,Kimiyoshi Ichida 2)杏林大学医学部薬理学 Toru Kimura,Jutabha Promsuk,Naohiko Anzai,Hiroyuki Sakurai key words:尿酸,尿酸トランスポーター 172 痛風と核酸代謝 第34巻 第 2 号 (平成22年) 4.1 mg/dlの範囲に保たれており,主に腎臓におけ グを挿入することにした.HA-mURAT1 Tgマウ る尿酸輸送が血中尿酸値を調節していると考え スは,フェニックスバイオ(株)に作出を依頼 られている3).従って,主要排泄器官である腎臓 した.マウスUrat1遺伝子をコードするゲノムク での尿酸輸送を理解することは重要である. ローンのコード配列の開始コドン直後にHAエピ URAT1は,SLC22A12遺伝子によりコードされ トープタグ配列を挿入した組換えBACクローン るOATファミリーに属するトランスポーターで を構築した.この組換えゲノムクローンを あり,主に腎近位尿細管の管腔側膜に発現する. Jcl:BDF1マウスの受精卵に導入してTgマウスを URAT1は細胞内の乳酸やニコチン酸,pyrazine 作出することにより,マウス個体本来の発現制 carboxylic acid(PZA)などの有機酸との交換に 御下でタグ付きmURAT1を発現する動物モデル 4) より尿酸を尿細管から再吸収する .尿酸排泄促 を得た10).本研究の動物実験は,杏林大学におけ 進薬(痛風治療薬)であるベンズブロマロンや る動物実験規定に則って行った. プロベネシドは,URAT1の阻害が主な作用機序 と考えられている5).また,抗結核薬であるピラ ジナミドは,URAT1がピラジナミド活性代謝物 2. RT(reverse-transcription)- PCR 約12週令のマウスからエーテル麻酔下で腎臓 であるPZAと尿酸とを交換輸送することにより, を摘出し,腎臓の皮質部位を採取した.フェノ 尿酸の再吸収を促進する5,6).さらに,URAT1遺伝 ール・クロロホルム抽出により,total RNAを得 子変異により尿酸再吸収が阻害されると,腎性 た . 逆 転 写 反 応 お よ び PCR反 応 は , そ れ ぞ れ 低尿酸血症が引き起こされることが知られてい Super Script III First-Strand Synthesis System,Go る4).従って,URAT1は血中尿酸値に大きく影響 Taq Polymerase(Invitrogen)を用い,そのプロト を及ぼすトランスポーターとして重要であり, コールに従って行った. 腎臓における尿酸輸送を理解する上で不可欠な 因子である. 3.腎皮質タンパク質の調製 これまでin vitroで尿酸輸送能を持つトランス 7) TS buffer(5mM Tris-HCl,pH7.4,250mM ポーターは数々報告されている が,in vivoでそ sucrose)にプロテアーゼインヒビター(1μg/ml の輸送が証明された,つまり遺伝子変異によっ leupeptin,1μ pepstatinA,1μg/ml aprotinin, 10 て病態を引き起こすことが示された尿酸トラン μ/ml PMSF)を加え,採取した腎臓皮質をホモ スポーターは,URAT1 4),URATv1 8),ABCG2 9) ジナイズした.4℃,800×gで10分間遠心を行い, の3つである.URAT1は尿酸再吸収における管 上清をさらに4℃,100,000×gで90分間,遠心し 腔側の尿酸の取り込みにおいて重要であるが, た.沈殿物にTS bufferを加え,ホモジナイザーに 実際にURAT1が血中尿酸値にどれほど影響を及 て懸濁した. ぼすのかは未だ明らかになっていない.そこで 本研究では,生体内での尿酸輸送機構を明らか 4.血漿・尿中の生化学的検査 にすることを目的として,URAT1の過剰発現マ エーテル麻酔下でマウスを開胸し,ヘパリン ウスを作出し,尿酸輸送におけるURAT1の役割 処理を施したシリンジを用いて心臓から血液を 解明を行った. 採取した.800×gで10分間遠心を行い,上清の 血漿を得た.また膀胱穿刺により,尿を採取し 方 法 1. HA-mURAT1 トランスジェニック(Tg) た.生化学測定はオリエンタル酵母株式会社, 長浜LSLに委託した. マウスの作製 Tgマウス作製にあたり,導入したURAT1を検 出するため,hemagglutinin(HA)エピトープタ 5.DNAマイクロアレイ解析 CodeLink Gene Expression Sytem(Applied 173 Gout and Nucleic Acid Metabolism Vol.34 No.2(2010) Microarrays社)の製品説明書に従って行った. mUrat1 cDNAを,ネガティブコントロールとし bioarray chamber はarray WoRxe(GE Healthcare製) て,逆転写反応を行わなかったものをそれぞれ を用いて走査し,CodeLink Expression Analysis 用いた.その結果,WTマウス,Tgマウスともに, Version 4.2ソフトウエアを用いてmRNA発現量解 腎臓においてmUrat1 mRNAの発現が確認できた. 析を行った.Student’ s t-testを用い,p<0.05を統 肝臓においてはmUrat1のバンドは確認できなか 計的に有意と判定した.Fold changeは,野生型 った.脳,小腸においても同様にRT-PCRを行っ (WT)マウスでのある遺伝子のmRNA発現量を たが,mUrat1 mRNAの発現は見られなかった 1とした時,Tgマウスの同じ遺伝子のmRNA発 ( デ ー タ 示 さ ず ). 以 上 の 結 果 か ら , m U r a t 1 現の変化量で示した(n=3). mRNAは腎臓特異的に発現しており,また過剰 発現させても腎臓以外に発現することはないこ 6.定量PCR解析 とが示された.腎臓において,mUrat1 mRNAの マウス腎臓皮質からtotal RNAを調製し,逆転 発現が WTマウスに比べてTgマウスで増加して 写反応によってcDNAを調製した.定量PCR反応 いた.また,雄性マウスの方が雌性マウスに比 は,THUNDERBIRD SYBR qPCR Mix (TOY- べてmUrat1 mRNAの発現増加が顕著であった. OBO) を 用 い た . 測 定 は ABI PRISM 7700 Sequence Detector( ABI社 , ソ フ ト ウ エ ア : 2.導入したHA-URAT1タンパク質の腎臓に Sequence Detector Software 1.91allias)で行った. おける発現 11) 発現量解析はΔΔ Ct法により行った . 抗HA抗体によるウエスタンブロットを用い て,導入したHA-mURAT1のタンパク質発現を確 結 果 1.RT-PCR法によるmUrat1 mRNA発現の確認 認した(Fig. 1B).mURAT1は糖タンパク質であ るため,コアとなるタンパク質が合成された後, マウスUrat1(mUrat1)のmRNA発現をRT- 小胞体でハイマンノース型,ゴルジ体でコンプ PCR法にて検討を行った(Fig.1A).ポジティブ レックス型の糖鎖修飾を受け原形質膜に到達す コントロールとして,RT反応液の代わりに る.Tgマウスではハイマンノース型およびコン プレックス型糖鎖修飾を受けたHA-mURAT1のタ ンパク質発現が確認できた.また,抗HA抗体を 用いた免疫組織染色の結果から,導入したHAmURAT1が腎近位尿細管の管腔側に発現してい ることを確認した(データ示さず). Fig1. Expression of URAT1 mRNA and protein A. mURAT1 mRNA expression in the kidney and the liver was examined by RT-PCR. 1mg total RNA was reverse-transcribed and amplified with PCR. B. HA-URAT1 protein expression was detected by Western blotting. 50 mg protein was loaded on SDS-PAGE and HA-URAT1 was detected with anti-HA antibody. HA-URAT1 modified with high-mannose and complex type carbohydrate chains was expressed in Tg mice. 174 痛風と核酸代謝 第34巻 第 2 号 (平成22年) Fig2. Urate concentration in plasma and urine Plasma (A)and Urine(B)urate concentration was measured by enzyme method. 3.血漿中および尿中の尿酸値測定 mURAT1を過剰発現させたことによる尿酸値 の変動を検討するために,血漿中および尿中の 尿酸値の測定を行った(Fig. 2).尿酸値の測定 は,URAT1の発現が顕著である雄性マウスを用 Tgマウスの血中尿酸値および尿中尿酸排泄量に いた.WTマウスおよびTgマウスの血漿尿酸値を 変化は見られなかった. 測定した結果,WTマウス(3.33±1.34mg/dl, n=19)とTgマウス(3.05±1.36mg/dl,n=18) 4.腎臓における尿酸トランスポーターの発現変 の間には有意な差は認められなかった.また, 動 それぞれの尿中尿酸値を測定した結果,WTマウ URAT1を過剰発現させても,血中・尿中尿酸 ス(0.35±0.08g/g cre,n=10)とTgマウス 値に変化が見られなかった.1つの理由として, (0.42±0.09g/g cre,n=13)の間には有意な差 他の尿酸トランスポーターが代償的にその発現 は認められなかった.以上の結果から,URAT1 変化を起こしていると考え,定量PCRを用いて を過剰発現させたにもかかわらず,WTマウスと 尿酸を運ぶとされているトランスポーターの 175 Gout and Nucleic Acid Metabolism Vol.34 No.2(2010) mRNAの定量を行った(Table1).それぞれの尿 ことからも,URAT1が血中尿酸値の調節に重要 酸トランスポーターのマウス腎臓皮質における な役割を果たしていると考えられる 5).そこで mRNA発現量は,ΔΔCt法を用いて相対発現量変 我々は,URAT1過剰発現マウスを作出し,その 化として解析した 11).mUrat1遺伝子は,Tgマウ 血中・尿中尿酸値に対する寄与に関して検討を スにおいて有意に発現増加が認められ,導入し 行った.URAT1が尿酸再吸収を行うトランスポ たHA-mUrat1が腎臓に過剰発現していることが ーターのため,再吸収が亢進し血中尿酸値が上 確認された.しかしながら,他の尿酸トランス 昇すると予測したが,血中尿酸値に変化が見ら ポーター遺伝子に関しては,その発現量に有意 れなかった.マウスにはヒトと異なり,尿酸を な変化は見られなかった. アラントインに代謝する酵素ウリカーゼが発現 しているため,血中尿酸値の変化が見られない 5.尿酸代謝酵素遺伝子の発現変動(Table2) 可能性がある12).そこで,尿中尿酸値の測定も行 血中・尿中尿酸値に変化が見られなかった他 った.実際,URAT1,OAT1,OAT3のノックア の理由として,尿酸代謝酵素の発現変化を考え, ウトマウスでは,血中尿酸値に変化が見られな DNAマイクロアレイを用いて尿酸代謝酵素 いものの,尿中尿酸排泄量に変化が見られてい mRNAの発現量を比較した.総遺伝子数36227に る13).しかしながら,URAT1 Tgマウスでは,尿 関して腎臓皮質におけるmRNAの発現解析を行 中尿酸値にも変化が見られなかった. った.その結果,Tgマウスの方がWTマウスに比 そこで,URAT1を過剰発現させたことによっ べて2倍以上発現量が増加していたものが55遺 て代償的に他の尿酸トランスポーターや尿酸代 伝子,1/2以下に減少していたものが38遺伝子存 謝酵素が発現変化を起こし,尿酸値に変化が見 在した.DNAマイクロアレイにおいても, られなかった可能性を考え,それらの遺伝子発 mUrat1 mRNAの発現量増加が確認できた.しか 現をDNAマイクロアレイおよび定量PCRで解析 しながら,尿酸代謝酵素や他の尿酸トランスポ した.しかしながら,既知の尿酸トランスポー ーターに関しては,有意な発現量の変化は見ら ターや尿酸代謝酵素のmRNAの発現に変化は見 れなかった.同様に,尿酸の主要代謝器官であ られなかった.このように,今回の研究では る肝臓についてもDNAマイクロアレイ解析を行 mRNAレベルでの発現の定量・比較検討のみを ったが,尿酸代謝酵素の発現に有意な変化は見 行ったが,mRNAの発現レベルはタンパク質の られなかった. それと比例しないこともある.また我々の研究 グループは,URAT1がスカフォールドタンパク 考 案 PDZK1と結合することによって細胞膜上で安定 本研究では,腎臓における尿酸輸送機構を解 化され,尿酸輸送活性が上昇することを示した14). 明することを目的として,URAT1過剰発現マウ こういった,PDZK1や未知の結合タンパク質と スを作出し,その解析を行った. の相互作用によって過剰発現させたURAT1また 食物から摂取した尿酸や生体内で合成された は他の尿酸トランスポーターが機能調節されて 尿酸は糸球体濾過を受けた後,腎近位尿細管に いる可能性もある.さらに,他のトランスポー おいて再吸収,分泌,分泌後再吸収を受け,最 ターの様にリン酸化やニトロソ化などの翻訳後 終的に約10%のみが排泄される.この過程の再吸 修飾によって機能調節されている可能性もある. 収を担っている分子の1つがURAT1であり,そ 従って,今後はタンパク質レベルでの発現・機 の遺伝的機能欠損によって尿酸再吸収が阻害さ 能調節について検討することが必要であろう. 4) れ,腎性低尿酸血症を引き起こす .また,尿酸 2008年に我々は,グルコーストランスポータ 降下薬として用いられているベンズブロマロン ーGLUT9(SLC2A9)が腎臓近位尿細管の血管側 やプロベネシドがこのURAT1を標的としている に存在する電位依存性尿酸トランスポーター 176 痛風と核酸代謝 第34巻 第 2 号 (平成22年) URATv1であり,管腔側のURAT1とタンデムに 4)E nomoto A,Kimura H,Chairoungdua A: 働くことによって尿酸の経細胞輸送を担うとい Molecular identification of a renal urate anion 8) う モ デ ル を 提 唱 し た . 近 年 , Genome Wide exchanger that regulates blood urate levels. Association Study(GWAS)によって,血中尿酸 Nature 23 : 447-452,2002. 値や痛風と関連する遺伝子の報告が多数なされ 5)Ichida K,Hosoyamada M,Hisatome I et al: ているが,尿酸値は管腔側のURAT1よりも血管 Clinical and molecular analysis of patients with 側の URATv1の方が遺伝子多型との関連を示さ renal hypouricemia in Japan-influence of れている場合が圧倒的に多い 15-18) .このことと今 回の結果を考え合わせると,高尿酸血症発症に は管腔側での尿酸の細胞内への取り込みよりも, URAT1 gene on urinary urate excretion.J Am Soc Nephrol 15:164-173,2004. 6)Roch-Ramel F,Guisan B,Schild L:Indirect 血管側での尿酸の排出の方がより重要である可 coupling of urate and p-aminohippurate transport 能性が示唆される. to sodium in human brush-border membrane 以上より,腎近位尿細管管腔側の尿酸再吸収 トランスポーターURAT1を過剰発現させても, 他の尿酸トランスポーターや尿酸代謝酵素の mRNA発現に変化を及ぼさず,血中・尿中尿酸 値も変化しないことが示された. vesicles. Am J Physiol 270:F61-68,1996. 7)安西尚彦,Jutabha Promsuk,木村徹 他:腎 臓の尿酸トランスポーター:最近の進歩 痛風と核酸代謝 33:7-15,2009. 8)Anzai N,Ichida K,Jutabha P et al:Plasma urate level is directly regulated by a voltage-driv- 謝 辞 Tgマウスの作出,ならびに研究遂行に際して 御助言をいただいた大阪大学大学院医学系研究 科生体システム薬理学,金井好克教授に感謝い たします.Tgマウスの維持,スクリーニング, en urate efflux transporter URATv1(SLC2A9) in humans.J Biol Chem 283:26834-26838, 2008. 9 ) Matsuo H, Takada T, Ichida K et al: Common defects of ABCG2,a high-capacity サンプル調製に協力を頂いた,杏林大学医学部 urate exporter,cause gout:a function-based 薬理学教室,高橋美知氏に感謝いたします.本 genetic analysis in a Japanese population.Sci 研究は科学研究費補助金(特定領域研究,若手 Transl Med 1:p 5ra11,2009. 研究),財団法人痛風研究会の支援によって一部 行われました. 10)Takahashi R,Ueda M:Generation of transgenic rats using YAC and BAC DNA constructs. Methods Mol Biol 597:93-108,2010. 文 献 1)Wu XW,Lee CC,Muzny DM et al:Urate oxidase:primary structure and evolutionary implications.Proc Natl Acad Sci U S A.86: 9412-9416,1989. 11)Livak KJ,Schmittgen TD:Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T))Method. Methods 25:402-408,2001. 12)Wu X,Wakamiya M,Vaishnav S et al: 2)中村 徹:高尿酸血症の原因(尿酸排泄低 Hyperuricemia and urate nephropathy in urate 下),高尿酸血症・痛風.新しい診断と治療 oxidase-deficient mice.Proc Natl Acad Sci U S のABC37,鎌谷直之編,pp34-47,最新医学 社,東京,2006. 3)Hediger MA,Johnson RJ,Miyazaki H et al: A 91:742-746,1994. 13)Eraly SA,Vallon V,Rieg T et al:Multiple organic anion transporters contribute to net renal Molecular physiology of urate transport. excretion of uric acid.Physiol Genomics 33: Physiology (Bethesda) 20:125-133,2005. 180-192,2008. 177 Gout and Nucleic Acid Metabolism Vol.34 No.2(2010) 14)Anzai N,Miyazaki H,Noshiro R et al:The 17)Stark K,Reinhard W,Neureuther K et al: multivalent PDZ domain-containing protein Association of common polymorphisms in PDZK1 regulates transport activity of renal GLUT9 gene with gout but not with coronary urate-anion exchanger URAT1 via its C artery disease in a large case-control study. terminus.J Biol Chem 279:45942-45950, 2004. PLoS One 3:e1948,2008. 18)Dehghan A,K tgen A,Yang Q et al: 15)D ing A,Gieger C,Mehta D et al:SLC2A9 Association of three genetic loci with uric acid influences uric acid concentrations with pro- concentration and risk of gout:a genome-wide nounced sex-specific effects.Nat Genet 40: association study.Lancet 372:1953-1961, 430-436,2008. 2008. 16)Vitart V,Rudan I,Hayward C et al:SLC2A9 is a newly identified urate transporter influencing serum urate concentration,urate excretion and gout.Nat Genet 40:437-442,2008. 178 痛風と核酸代謝 第34巻 第 2 号 (平成22年) Urate metabolism in urate transporter URAT1 transgenic mice Ai Tsukada1) Toru Kimura2) 2) Jutabha Promsuk Naohiko Anzai2) 1) Kimiyoshi Ichida Hiroyuki Sakurai2) Urate is a final metabolite of purines and the major the urate concentration and DNA microarray and excretory organ is the kidney. Renal urate handling is quantitative PCR analysis. RT-PCR showed that clinically important because over-excretion and expression of Urat1 mRNA is specific in the kidney under-excretion of urate cause hypouricemia and and the expression level in Tg mice was increased hyperuricemia, respectively. We identified a urate- compared with that in wild type(WT)mice. anion exchanger, URAT1, localized at the apical side Western blot and immunohistochemistry showed that in the renal proximal tubules. This transporter is likely exogenous URAT1 was expressed only in the renal to act on urate reabsorption, because URAT1 is a tar- proximal tubule cells in the kidney. Serum urate con- get for anti-gout drugs and mutation in URAT1 causes centration in URAT1 Tg mice was not significantly renal hypouricemia. Thus, URAT1 is one of the different from that in WT mice. The urate level in important molecules for maintaining the serum urate urine from URAT1 Tg mice was the same as that level. However, little is known about how URAT1 from WT mice. DNA microarray and quantitative controls the serum urate level and its expression influ- PCR analysis showed that in URAT1 Tg mice,the ences other urate transporters and urate metabolic expression levels of urate metabolic enzymes and enzymes in vivo. We constructed URAT1 transgenic urate transporters other than Urat1 were the same as mice(Tg)to investigate the urate transport mecha- those in WT mice. nism in vivo. We used Tg mice for measurement of 1)Department of Pathophysiology, Tokyo University of Pharmacy and Life Science 2)Department of Pharmacology and Toxicology, Kyorin University School of Medicine key words:Urate,Urate transporters 179 Gout and Nucleic Acid Metabolism Vol.34 No.2(2010) 原著2 炎症性サイトカイン過剰産生ヒト胸腺癌細胞株(ThyL-6)における Flavopiridolによる転写阻害を介したlL-6分泌抑制 高木 和貴1) 岩崎 博道1) 背 稲井 邦博2) 上田 孝典1) 内木延宏2) 景 結 Interleukin-6 (IL-6)はde novo急性炎症反応に 果 おける重要なメディエーターとして産生され, ThyL-6細胞が分泌しているサイトカインとし 細胞受容体に結合すると核内に伝達し転写因子 てIL-6に加え,IL-8,VEGF,TNFα,IL-1α, を活性化する.近年IL-6受容体抗体等生物学的製 RANTESが検出された.IL-6については,細胞に 剤が登場し,関節リウマチ患者にも約3割程度 1nMから100nM Flavopiridolを4時間添加すると, に寛解を得ることが可能になった.Flavopiridol 濃度依存性に培養上清中IL-6濃度が減少した. はDysoxylum binectariferumより抽出されるフラボ Western blot法による検討ではFlavopiridol投与に ノイド成分由来の半合成のフラボン誘導体であ より濃度依存性にIL-6の発現が抑制され,時間・ る.Flavopiridolは殆どのcyclin-dependent kinases 濃度依存性にRNA polymerase IIのdegradationが見 (CDK)に対する阻害作用により,転写レベルに られた. 於いてRNA polymerase IIのリン酸化を阻害し, 結 論 細胞の転写活性を抑制する事が近年明らかにさ Flavopiridolは転写阻害によりIL-6産生細胞での れつつある. IL-6産生を抑制した.FlavopiridolがCDKを標的と 目 的 IL-6を含む炎症性サイトカイン産生細胞株を用 いて,FlavopiridolがIL-6産生を転写レベルで阻害 したIL-6産生抑制による新規分子標的療法とし て,関節リウマチなど炎症性疾患に対治療戦略 になりうる可能性が示唆された. することにより,関節リウマチなど炎症性疾患 はじめに に対する新規分子標的薬となりうるか検討する. 関節リウマチなどの炎症性疾患,重症細菌感 方 法 染症などによる発熱時では産生・惹起された炎 高熱・白血球増加・血清IL-6値の高値・尿酸排 症性サイトカインが病因病態形成上,重要な役 泄亢進による低尿酸血症が見られる57歳胸腺癌 割を占めている1).特に敗血症など重症感染症に 患者から我々が樹立したThyL-6細胞株について, 罹患した患者では,炎症性サイトカイン血症2)か サイトカイン抗体アレイにて細胞が分泌するサ ら播種性血管内凝固,多臓器障害など不可逆的 イトカインをスクリーニングした.In vitroにお な病態を来しうるため,感染症など原因治療に いて細胞にFlavopiridolを添加し,western blot法に 並行して高サイトカイン血症に対するステロイ てRNA polymerase IIの発現を観察した. ドパルス療法や吸着療法3)などが必要になる場合 がある. 受付:2010年9月8日,受理:2010年11月25日 1)福井大学医学部 血液腫瘍内科 Kazutaka Takagi,Hiromichi Iwasaki,Takanori Ueda 2)福井大学医学部 分子病理学 Kunihiro Inai,Nobuhiro Naiki Key words:Flavopiridol,炎症性サイトカイン,IL-6,RNA polymerase II 180 痛風と核酸代謝 第34巻 第 2 号 (平成22年) Interleukin-6 (IL-6)はde novo急性炎症反応に し転写因子を活性化することが知られている4, 5). おける重要なメディエーターとして産生され, また多発性骨髄腫など一部の悪性腫瘍において, 肝臓のIL-6受容体に結合しgp130とオリゴマー複 自己分泌も含めたIL-6依存性の細胞増殖が知られ 合 体 を 形 成 す る ( 図 1 ). さ ら に シ グ ナ ル が ている 6-8).近年関節リウマチの臨床に於いて抗 JAK/Tyk キナーゼ,STAT3を介して核内に伝達 IL-6受容体抗体やTNF-α阻害剤などいわゆる生 物学的製剤が登場し,関節リウマチ患者にも約 3割程度に寛解を得ることが可能になってきた 9) .現在使用可能な生物学的製剤である抗TNF-α 単クローン性抗体・可溶性TNF受容体や抗IL-6受 容体抗体は,滑膜マクロファージや線維芽細胞の 活性化に重要なTNF-αやIL-6の作用を直接抑制す ることで滑膜や線維芽細胞の増殖を抑制する. Flavopiridol(図2)は熱帯・亜熱帯に生息す るMahogany科に属する植物の木皮である Dysoxylum binectariferum(中国では「葱臭木」) より抽出されるフラボノイド成分から人工的に 合成された半合成のフラボン誘導体である. Flavopiridolはcyclin-dependent kinase (CDK)の 図1 IL-6シグナル伝達 我々の樹立したThyL-6細胞は炎症性サイトカインを分泌し, 自己分泌されたIL-6により増殖する.IL-6は細胞膜IL-6受容体 ATP結合部位に結合し,低濃度で殆どのCDKに 対して高い阻害効果を示す10, 11).単剤での臨床試 に結合しgp130とオリゴマー複合体を形成する.さらにシグナ 験(phase II)ではCDK inhibitorとして転移性悪 ルがJAK/Tyk kinases,STAT3を介して核内に伝達し転写因子 性黒色腫12),子宮体癌13),多発性骨髄腫14)に対す を活性化する.FlavopiridolはRNA pol II阻害により転写を阻害 る臨床試験が行われたが効果は不充分で14),強い する. 骨髄毒性以外に非血液毒性として下痢が見られ た.Flavopiridolは現在慢性リンパ性白血病15)や, ハイリスク急性骨髄性白血病に対して,araC等 との併用療法として臨床治験が行われている16). Flavopiridolは 細 胞 内 転 写 レ ベ ル に 於 い て , CDK阻害を介してRNA polymerase IIのリン酸化 を阻害し転写活性を抑制する事が近年明らかに されている 16).Flavopiridolのターゲットである cyclin H-CDK7は,CDK-activating kinase (CAK) 活性を持っている他にRNA polymerase II (pol II) のCOOH-terminal domain(CTD)アミノ酸配列 Y1S2P3T4S5P6S7の52回繰り返し構造のうち,5番目 図2 Flavopiridolの分子構造 のセリン(S 5)をリン酸化することによりRNA 構造式名, (-)-2-(2-Chlorophenyl)-5,7-dihydroxy-8-[(3r,4s)-3- pol IIの転写開始を促進する活性を持つ 17).また hydroxy-1-methyl-4-piperidinyl]-4h-1-benzopyran-4-one 同様にFlavopiridolのターゲットであるcyclin T- hydrochloride.マホガニー科Dysoxylum binectariferumとい う木皮から発見され,分子構造をヒントに合成されたフラボノ CDK9は,RNA pol II CTDの2番目セリン(S2)を イド. リン酸化することによりRNA pol IIの転写伸長を 促進する.Houらは肝癌細胞(HepG2)を用いて 181 Gout and Nucleic Acid Metabolism Vol.34 No.2(2010) Flavopiridolが細胞のIL-6産生を転写レベルで産生 患者胸水から採取した胸腺癌細胞を10%FBS添加 阻害することにより抗炎症作用を発揮し得るこ RPMI1640培地 (100 U/ml ペニシリン,100 ug/ml 18 ) と を 示 し た . さ ら に Flavopiridolが vascular ストレプトマイシン含有)を用いて37℃, endothelial growth factor (VEGF)蛋白の発現を 5%CO 2条件下で継代培養し,細胞株(ThyL-6細 抑制することも示されている19).我々はIL-6等多 胞)として樹立した20).実験には対数増殖期の細 様な炎症性サイトカインを過剰産生するヒト胸 胞を用いた.細胞培養上清中のサイトカイン分 腺癌細胞株(ThyL-6)を樹立した20).今回我々は 泌の定量は細胞増殖がconfluentとなった時期の培 ThyL-6細胞株を用いてFlavopiridolがIL-6産生を抑 養上清を採取し,測定まで-80℃で保存した. 制することを示し,関節リウマチなど炎症性疾 患に対する免疫抑制薬としての分子標的療法薬 に成りうる可能性が示唆されたので報告する. Western blot法:細胞から抽出した蛋白と等 量の2x SDS-PAGE sample buffer (100mM TrisHCl pH 6.8, 4% SDS, 20% Glycerol, 20% β-mercap- 方 法 toethanol and 0.02% Bromophenol blue)を準備し, 試 薬 : Flavopiridol hydrochloride, 2-( 2- SDS-PAGEゲル(7%, 10% or 15%)に添加し泳動 Chlorophenyl)-5,7-dihydroxy-8-[(3r,4s)-3- した.ゲル内に分画された蛋白は Immobilon-P hydroxy-1-methyl-4-piperidinyl]-4h-1-benzopyran-4- 0.45 μm PVDF membrane (Millipore, Billerica, one hydrochloride ( Sigma, St. Louis, MO), MA)に転写し,抗体とTBS-T (25 mM Tris, R P M I 1 6 4 0 培 地 , 牛 胎 児 血 清 ( F B S ), 1 0 % 130mM NaCl, pH 8.4,5 mM リン酸2水素カリウ trypsin-EDTA,MTT assay kit (以上Sigma), ム, 5%ドライミルク,0.1% Tween 20)を用いて TranSignal Human Cytokine Antibody Array 3.0 kit 培養した. (Panomics, Redwood City, CA),ヒト遺伝子組み 換 え IL-6( 分 子 量 約 24kD), IL-8, RANTES サイトカイン抗体アレイ解析:サイトカイン ( Regulated upon Activation, Normal T-cell 抗体アレイ(TranSignal Human Cytokine Antibody Expressed, and Secreted)(以上PeproTech EC, Array 3.0 kit:Panomics, Fremont, CA) を用いて, London, UK) ,ウサギ多クローン抗ヒトIL-6抗体, 細胞が分泌するサイトカインプロフィールをス マウスモノクローナル抗ヒトIL-8抗体,ヤギ多ク クリーニングした.2mLの対数増殖期細胞培養 ローン抗ヒトRANTES抗体(以上Sigma).RNA 上清中にメンブレンを室温で2時間培養した. polymerase II (pol II) (N-20),phospho-Ser5 Pol メンブレンに二次抗体処理してサイトカインを II (clone 8A7) (以上Santa Cruz Biotechnology Chemiluminescence Image Analyzer ( Alpha Inc., CA), phospho-Ser 2 Pol II ( ab5095) Innotech, San Leandro, CA)にて可視化した. (Abcam, Cambridge, MA),HRP結合二次抗体 (Santa Cruz, CA) . 統計処理:図4,図5におけるデータ群間の 検定は,ANOVA分散分析と多重比較のpost hoc ThyL-6細胞の樹立:57歳男性.持続する高熱 を主訴として福井大学医学部附属病院血液腫瘍 検定をStatView 5.0 (SAS Institute Inc.)を用いて 行った. 内科に紹介初診となった.CT等にて縦隔腫瘍と 鎖骨上リンパ節腫脹を認め,生検にて胸腺癌と 診断した.38℃以上の高熱の持続と白血球増加, 結 果 ThyL-6細胞のcharacterization. 患者胸水から採 血清CRP高値,血清IL-6値の高値,血清尿酸値 取した胸腺癌細胞を継代培養しThyL-6細胞を樹 1.7 mg/dl,尿酸クリアランス18.2 mL/minと著し 立した20).免疫染色による検討ではThyL-6細胞で い尿酸排泄亢進による低尿酸血症を呈していた. は上皮系のマーカーが発現し,リンパ球系のマ 182 痛風と核酸代謝 第34巻 第 2 号 (平成22年) UK)より若干大きな分子量であった(図3A). 自己分泌IL-6依存性増殖:ThyL-6細胞培養上 清に抗モノクローナルIL-6抗体を投与すると,濃 度依存性にThyL-6細胞増殖が抑制され(図3B ),ThyL-6細胞増殖は自己分泌IL-6に依存するこ とが示唆された.ThyL-6細胞が分泌するIL-6の生 物学的活性の有無を検討するために,培養上清 をIL-6依存性に増殖する多発性骨髄腫細胞 (ILKM-3細胞) 21) に添加した.添加用量依存性 にILKM-3細胞増殖を促進した(データ省略). ThyL-6が分泌するIL-6が生物学的活性を持つこと が示唆された. FlavopiridolによるThyL-6細胞のIL-6分泌抑 制:Flavopiridolは細胞CDKを阻害し,転写レベ ルに於いてRNA polymerase IIのリン酸化を阻害 表1 ThyL-6細胞の表面抗原(免疫染色) ーカーは陰性であった(表1).G-band分染法で は染色体領域7p11, 16q24, 17p11等の異常が見ら れた.サイトカインアレイを用いて測定した ThyL-6細胞が分泌するサイトカインプロフィー ルを示す(表2).ThyL-6細胞が分泌しているサ イトカインとしてIL-6,IL-8,RANTES,IL-1α, VEGF,soluble TNFα receptor I,eotoxin,cytotoxic T lymphocyte antigen (CTLA)が検出され, GM-CSF, EGF, MIP1α , MIP1β , MIP-4, MIP-5,TGFβ,IFNγ,TNFR II,VCAMP-1, IL-1β,IL-1Rα,IL-2,IL-3,IL-5,IL-6R,IL10,IL-15,IL-17は検出されなかった(表2). Western blot法ではThyL-6細胞が分泌するIL-6は ヒト遺伝子組み換えIL-6 (PeproTech EC, London, 表2 ThyL-6細胞が分泌するサイトカイン 183 Gout and Nucleic Acid Metabolism Vol.34 No.2(2010) とし,RNA polymerase II (pol II) のCOOH-terminal domain(CTD)リン酸化を阻害する可能性 が想定されている 17 ).ThyL-6細胞に見られた FlavopiridolのRNA pol II 転写阻害活性を検討する ために100nM Flavopiridol存在下に4時間培養し, ThyL-6細胞のIL-6もしくはRNA polymerase II (RNA pol II)の発現の変化をwestern blot法で検 討した.Flavopiridol投与により濃度依存性にIL-6 の発現が抑制され,時間依存性,濃度依存性に RNA pol IIのdegradationが見られた(図5). RNA pol II の2つのサブユニット(IIoとIIa)の うち,RNA pol IIのDNAへの結合期(initiation) サブユニット(IIa)において100nMの 図3 A)ThyL-6細胞から分泌されたIL-6(western blot法) . ThyL-6細胞培養上清を直接15% sodium dodecylsulfatepolyacrylamide gel electrophoresis (SDS-PAGE)で泳動し,膜 を抗ヒトモノクローナルIL-6抗体で培養した.細胞が分泌する 蛋白はヒト遺伝子組み換えIL-6より若干大きい分子量と考えら れた. B)抗IL-6抗体投与によるThyL-6細胞増殖抑制 96穴プレート培地に1x105のThyL-6細胞を一晩培養し洗浄し た後,10%, 1.0%, もしくは0.1%抗IL-6抗体含有RPMI培地で ThyL-6細胞を24時間培養した。培養上清に抗モノクローナル IL-6抗体を投与によりThyL-6細胞の増殖がコントロールと比較 して抑制され(MTT assay, p<0.05),ThyL-6細胞増殖は自己 分泌IL-6に依存することが示唆された. し,細胞蛋白の転写活性を抑制する事が近年明 らかにされている22). FlavopiridolのThyL-6細胞 に対する増殖抑制効果は72時間 in vitro MTT assayによるIC50値では400±10 nMであったが,今 回我々は~100 nMのFlavopiridolにおけるThyL-6細 胞に対する薬理作用を検討した.これまでの臨 床治験に於いて,患者でのFlavopiridolの血中濃 度は投与法にもよるが,1μM程度まで到達可能 であるため10),今回1nM, 10nM, 100nMで4時間 培養した.この濃度条件のFlavopiridol短時間パ ルス投与により,非投与時と比べて投与量依存 性に細胞培養上清中IL-6濃度が抑制された(図 4) . FlavopiridolによるThyL-6細胞転写抑制: Flavopiridolはcyclin H-CDK7/cyclinT-CDK9を標的 Flavopiridol投与により阻害が見られ,伸長期 (elongation)サブユニット(IIo)ではFlavopiridol 投与によりdegradationが見られた(図5B) . 考 察 我々は発熱・白血球増加・著しい高CRP血症, そして排泄促進による血漿尿酸値低下を呈した 胸腺癌患者からThyL-6細胞を樹立した20).今回行 ったサイトカインアレイによるスクリーニング ではThyL-6細胞はIL-6等いくつかの炎症性サイト カインを分泌していた.本細胞の増殖は自己が 分泌するIL-6にup regulateされていたが(図3), IL-6以外にもTNF-αやIL-8等いくつかの炎症性サ イトカインも高濃度に産生されており,これら 他のサイトカインの影響も否定はできない.臨 床的にも一部の多発性骨髄腫や悪性リンパ腫細 胞に於いて,IL-6依存性に増殖する腫瘍が知られ ている6-8).Frassanitoらはflowcytometerを用いて多 発性骨髄腫患者47人と,本態性M蛋白血症 (MGUS)患者15人の骨髄腫細胞から分泌される IL-6量が臨床病期と相関することや,再発難治性 細胞が自己分泌IL-6に関連することを示した 6). これらデータからFlavopiridolによる癌細胞IL-6転 写抑制を標的とした治療戦略は,難治性多発性 骨髄腫や悪性リンパ腫8)などIL-6/STAT3経路を耐 性機序とした治療抵抗性がん細胞治療に応用で きうる可能性も示唆された.しかしIL-6/STAT3 経路以外に,低悪性度リンパ腫におけるBcl-2な 184 痛風と核酸代謝 第34巻 第 2 号 (平成22年) 図5 FlavopiridolによるIL-6転写抑制 図4 Flavopiridolによる細胞IL-6分泌抑制 ThyL-6細胞(1x105)を一晩培養し洗浄した後,図に示した濃度 のFlavopiridol含有RPMI培地で4時間培養し,培養液中のIL-6 濃度を測定した.100nM Flavopiridol投与により培養上清中へ のIL-6分泌がコントロールと比較して抑制された(p<0.05) ど抗アポトーシス蛋白転写抑制の可能性も考え A) ThyL-6細胞培養液中に100nM Flavopiridolを投与し4時間 培養した後,細胞を採取し細胞RNA polymerase II (RNA pol II)の発現をwestern blotで観察した.上のバンド(IIo) はリン酸化 RNA pol IIを,下のバンド(IIa)は非リン酸化 RNA pol IIを示す.4時間の薬剤投与によりRNA pol II の degradationが見られた. B) RNA pol II degradationはFlavopiridol濃度依存性が観察さ れた. 害している可能性が示唆された.1-10 nMという 極めて低い濃度の4時間パルス投与で 8) られる . Flavopiridolはpan CDK inhibitorであり 16, 23 ) , FlavopiridolがThyL-6細胞に於いてIL-6のdegrada- cyclin T-CDK9/cyclin H-CDK7を阻害し,RNA tionを誘導可能であったことは,Flavopiridolが低 polymerase II (pol II)のCOOH-terminal domain 用量で転写阻害を介したIL-6産生抑制作用を有 (CTD)アミノ酸配列Y S P T S P S の52回繰り返 し,新規抗炎症剤として関節リウマチなど炎症 し構造のうち,S もしくはS をリン酸化し,RNA 性疾患に有用となりうる可能性が導き出される. pol IIのDNAへの結合(initiation)と伸長(elon- 関節リウマチなど炎症性疾患に対する抗サイト 1 2 2 3 4 5 6 7 5 17) gation)により,転写活性を正に制御する .今 回血中濃度として到達可能な 10) ,100nMの カイン治療として既にヒト化抗IL-6受容体抗体 (tocilizumab)による優れた関節リウマチ症状改 Flavopiridol存在下に4時間まで細胞を培養しRNA 善効果・骨破壊進行抑制効果が得られている24). pol IIa サブユニット優位のdegradationが見られた 滑膜マクロファージから分泌されたIL-6が破骨細 (図5).このことはFlavopiridolがRNA pol IIの 胞を活性化して骨破壊を増悪させることからも, DNAへの結合(initiation)阻害を優位に転写を阻 関節リウマチにおいてIL-6産生を制御する必要が 185 Gout and Nucleic Acid Metabolism Vol.34 No.2(2010) あり,Flavopiridolは低濃度(∼100nM)でIL-6を Orientia tsutsugamushi infection. J Clin 抑制し関節破壊を抑制する可能性が示唆された. Microbiol 35:3320-3322,1997. Sekineらはマウス関節リウマチモデルを用いて 3)Takagi K,Shimizu H,Iwasaki H et al:Serum FlavopiridolがCDK inhibitionを介する滑膜細胞増 cytokine level during continuous venovenous 殖阻害により組織学的,臨床的に関節炎症状を hemofiltration in toxic shock-like syndrome due 改善するする事を示した25).Houらはin vitro炎症 to group G beta Streptococcus bacteremia in a モデルとしてHepG2肝細胞癌細胞においてIL-6に patient with idiopathic thrombocytopenic purpu- より誘導されるCDK9-STAT3複合体を介したγ- ra. Scand J Infect Dis 34:403-406,2002. フィブリノーゲン遺伝子プロモーター領域への 4)Murakami M,Hibi M,Nakagawa N et al: RNA plo II活性を,Flavopiridol(500nM)が阻害 IL-6-induced homodimerization of gp130 and することでFlavopiridolなどCDK9阻害剤が抗炎症 associated activation of a tyrosine kinase. 26) 作用を持ちうることを示した .上記報告のいず Science 260:1808-1810,1993. れも数百nMの濃度でのFlavopiridolで有意な滑膜 5)Moran DM,Mattocks MA,Cahill PA et al: 細胞増殖抑制効果など抗炎症効果が見られてお Interleukin-6 mediates G(0)/G(1) growth り,今回我々の細胞を用いた検討では100nMの arrest in hepatocellular carcinoma through a 低濃度でもIL-6蛋白のdegradationが観察された STAT 3-dependent pathway.J Surg Res 147: (図5) . 23-33,2008. 図1にIL-6刺激による細胞シグナル伝達の概念 6)Frassanito MA,Cusmai A,Iodice G et al: 図を示す.IL-6はde novo急性炎症反応における Autocrine interleukin-6 production and highly 重要なメディエーターとして産生され,肝臓細 malignant multiple myeloma: relation with resist- 胞のIL-6受容体に結合しgp130とオリゴマー複合 ance to drug-induced apoptosis.Blood 97: 体を形成する.さらにシグナルがJAK/Tyk キナ 483-489,2001. ーゼ,STAT3を介して核内に伝達し転写因子を 7)Brocke-Heidrich K,Kretzschmar AK,Pfeifer 活性化することが知られている4).ThyL-6細胞の G et al:Interleukin-6-dependent gene expres- 染色体異常として7p11,8p11,9q11,16q24, sion profiles in multiple myeloma INA-6 cells 17p11,21p11での異常が検出されたが,そのう reveal a Bcl-2 family-independent survival path- ち17p11にはgp130をコードする遺伝子が存在し way closely associated with Stat3 activation. ており,ThyL-6細胞もgp130を介したIL-6シグナ Blood 103:242-251,2004. ル伝達異常メカニズムの存在が示唆され(デー 8)Alas S,Bonavida B:Inhibition of constitutive タ省略) ,今後の検討を要する. STAT3 activity sensitizes resistant non- *本研究の一部は財団法人痛風研究会の研究助成 Hodgkin's lymphoma and multiple myeloma to 金により行った. chemotherapeutic drug-mediated apoptosis. Clin Cancer Res 9:316-326,2003. 文 献 9)Bathon JM,Cohen SB:The 2008 American 1)Ronnblom L,Elkon KB:Cytokines as thera- College of Rheumatology recommendations for peutic targets in SLE.Nat Rev Rheumatol 6: the use of nonbiologic and biologic disease-mod- 339-347,2010. ifying antirheumatic drugs in rheumatoid arthri- 2)Iwasaki H,Takada N,Nakamura T et al: Increased levels of macrophage colony-stimulat- tis: where the rubber meets the road.Arthritis Rheum 59:757-759,2008. ing factor,gamma interferon,and tumor 10)Senderowicz AM:The cell cycle as a target for necrosis factor alpha in sera of patients with cancer therapy:basic and clinical findings with 186 痛風と核酸代謝 第34巻 第 2 号 (平成22年) the small molecule inhibitors flavopiridol and UCN-01. Oncologist 7 Suppl 3:12-19,2002. ic acid in human neuroblastoma. Int J Cancer 99:658-664,2002. 11)Carlson BA,Dubay MM,Sausville EA et al: 20)Inai K, Takagi K, Takimoto N et al:Multiple Flavopiridol induces G1 arrest with inhibition of inflammatory cytokine-productive ThyL-6 cell cyclin-dependent kinase (CDK) 2 and CDK4 line established from a patient with thymic carci- in human breast carcinoma cells. Cancer Res 56:2973-2978,1996. noma. Cancer Sci 99:1778-1784,2008. 21)Shimizu S, Yoshioka R, Hirose Y et al: 12)Burdette-Radoux S,Tozer RG,Lohmann RC Establishment of two interleukin 6 (B cell stim- et al:Phase II trial of flavopiridol,a cyclin ulatory factor 2/interferon beta 2)-dependent dependent kinase inhibitor, in untreated human bone marrow-derived myeloma cell lines. metastatic malignant melanoma. Invest New J Exp Med 169:339-344,1989. Drugs 22:315-322,2004. 22) Sedlacek HH: Mechanisms of action of 13)Grendys EC,Jr.,Blessing JA,Burger R et al:A phase II evaluation of flavopiridol as second-line chemotherapy of flavopiridol. Crit Rev Oncol Hematol 38:139170,2001. endometrial 23)Shah MA, Schwartz GK:Cyclin-dependent carcinoma:a Gynecologic Oncology Group kinases as targets for cancer therapy. Cancer study. Gynecol Oncol 98:249-253,2005. Chemother Biol Response Modif 21:145-170, 14)Dispenzieri A,Gertz MA,Lacy MQ et al: 2003. Flavopiridol in patients with relapsed or refracto- 24)Nishimoto N, Kishimoto T:Interleukin 6: from ry multiple myeloma: a phase 2 trial with clinical bench to bedside. Nat Clin Pract Rheumatol 2: and pharmacodynamic end-points. Haematologica 91:390-393,2006. 619-626,2006. 25) Sekine C, Sugihara T, Miyake S et al: 15)Byrd JC, Lin TS, Dalton JT et al:Flavopiridol Successful treatment of animal models of administered using a pharmacologically derived rheumatoid arthritis with small-molecule cyclin- schedule is associated with marked clinical effi- dependent kinase inhibitors. J Immunol 180: cacy in refractory, genetically high-risk chronic 1954-1961,2008. lymphocytic leukemia. Blood 109:399-404, 2007. 16)Dickson MA, Schwartz GK:Development of cell-cycle inhibitors for cancer therapy. Curr Oncol 16:36-43,2009. 17)Svejstrup JQ:The RNA polymerase II transcription cycle:cycling through chromatin. Biochim Biophys Acta 1677:64-73,2004. 18)Hou T, Ray S, Brasier AR:The functional role of an IL-6 inducible CDK9-STAT3 complex in human gamma -fibrinogen gene expression. J Biol Chem 2007. 19)Rapella A, Negrioli A, Melillo G et al: Flavopiridol inhibits vascular endothelial growth factor production induced by hypoxia or picolin- 26)Hou T, Ray S, Brasier AR:The functional role of an interleukin 6-inducible CDK9.STAT3 complex in human gamma-fibrinogen gene expression. J Biol Chem 282:37091-37102,2007. 187 Gout and Nucleic Acid Metabolism Vol.34 No.2(2010) Flavopiridol inhibits interleukin-6 secretion via degradation of RNA polymerase Ⅱ in multi-cytokine-producing ThyL-6 cells originally established from patient with thymus cancer Kazutaka Takagi 1) Nobuhiro Naiki 2) Kunihiro Inai2) Hiromichi Iwasaki1) Interleukin-6 (IL-6), an inflammatory cytokine Takanori Ueda known to be an inhibitor of pan-cyclin-dependent produced by various cells, regulates production of kinases (CDKs). In this paper, we demonstrate that acute-phase proteins in liver, and induces proliferation Flavopiridol inhibits IL-6 production at concentrations in IL-6-dependent cells. After IL-6 binds to an IL-6 below 100 nM in inflammatory cytokine-producing receptor (IL-6R) on the target cell, the signal acti- cells (ThyL-6)originally established from a 57- vates gp-130-associated Janus tyrosine kinase (Jak), year-old patient with thymus cancer. Moreover, which then stimulates transcription, cell proliferation Flavopiridol was able to inhibit RNA polymerase II and cytokine production via signal transducers and phosphorylation, which is necessary for CDK-7 or activators of transcription 3 (STAT3). Monoclonal CDK-9 activation, thus suggesting that this flavonoid anti-IL-6R antibody therapy (tocilizumab 1) , human- drug can inhibit transcription activity at clinically ized monoclonal antibody) has recently been found achievable incubation times and concentrations.In to be effective for rheumatoid arthritis (RA)and conclusion, Flavopiridol is a novel therapeutic tool for some IL-6-dependent cancers, and about 30% of RA IL-6-dependent cancer, and is an anti-inflammation patients have achieved remission by combination use agent for conventional therapy-resistant rheumatologi- with methotrexate. Flavopiridol, a semi-synthetic cal diseases. flavonoid isolated from Dysoxylum binectariferum, is 1) Hematology and Oncology Division, Faculty of Medical Science, University of Fukui. 2) Division of Molecular Pathology, Faculty of Medical Science, University of Fukui.
© Copyright 2026 Paperzz