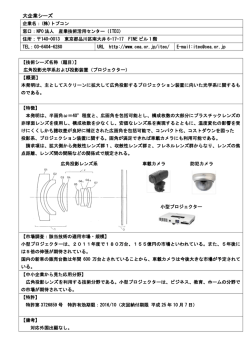
−1− (2) 特表平9−508789 【特許請求の範囲】 1.天然の樹脂を殺菌し、この樹脂から生物体を抽出しこの抽出した生物体を 培養することによって天然の樹脂から得られる生物体を含む生物体の培養。 2.天然の樹脂が、コハク及び/又はコーパルから成るグループの中から選択 される、請求の範囲第1項に記載の培養。 3.天然の樹脂が、レバノン産コハク、ワシントン州産コハク、メキシコ産コ ハク、カナダ産コハク、日本産コハク、バルト諸国産コハク、ドイツ産コハク、 コロンビア産コーパル、ビルマ産コハク、ドミニカ産コハク、中国産コハク及び /又はアフリカ産コーパルから成るグループの中から選択される、請求の範囲第 2項に記載の培養。 4.天然の樹脂が表面殺菌されている、請求の範囲第1項に記載の培養。 5.生物体が、粉砕抽出方法によって天然の樹脂から抽出される、請求の範囲 第1項に記載の培養。 6.樹脂が、粉砕に先立ち液体窒素で前処理される、請求の範囲第1項に記載 の培養。 7.樹脂を粉砕に先立ち約−20℃から約−140℃で凍結させる、請求の範囲第 1項に記載の培養。 8.樹脂を約−80℃に凍結させる、請求の範囲第7項に記載の培養。 9.生物体が、粗砕抽出方法によって、天然の樹脂から抽出される、請求の範 囲第1項に記載の培養。 10.生物体が、せん孔抽出方法によって、天然の樹脂から抽出される、請求の 範囲第1項に記載の培養。 11.生物体が、フラッシング抽出方法によって、天然の樹脂から抽出さいる、 請求の範囲第1項に記載の培養。 12.生物体が樹脂中に化石化された昆虫材料内に含有されている、請求の範囲 第1項に記載の培養。 13.昆虫材料が、昆虫組織、昆虫の糞便及び昆虫の巣を含むグループの中から 選択される、請求の範囲第12項に記載の培養。 −2− (3) 特表平9−508789 14.生物体が、樹脂の中に化石化された植物材料の中に含有される、請求の範 囲第1項に記載の培養。 15.植物材料が、根、葉、種子、小枝、樹皮、植物組織、胞子、花粉、花、イ ネ科牧草、シダ及びマメ科植物を含むグループの中から選択される、請求の範囲 第14項に記載の培養。 16.生物体が樹脂中で化石化された土壌の中に含有される、請求の範囲第1項 に記載の培養。 17.生物体が樹脂中で化石化された水の中に含有される、請求の範囲第1項に 記載の培養。 18.培養を含む生物体の年令が核酸分析によって決定される、請求の範囲第1 項に記載の培養。 19.生物体が微生物である、請求の範囲第1項に記載の培養。 20.生物体が、細菌、真菌、ウイルス、原生動物、微小藻類、節足動物及び/ 又は線虫から成るグループの中から選択される、請求の範囲第1項に記載の培養 。 21.(a)天然の樹脂を殺菌する段階、(b)生物体を天然の樹脂から抽出す る段階、及び(c)前記抽出された生物体を培養する段階、を含んで成る天然の 樹脂からの生物体の培養を得るための方法。 22.天然の樹脂が、コハク及び/又はコーパルから成るグループの中から選択 される、請求の範囲第21項に記載の方法。 23.天然の樹脂が、レバノン産コハク、ワシントン州産コハク、メキシコ産コ ハク、カナダ産コハク、日本産コハク、バルト諸国産コハク、ドイツ産コハク、 コロンビア産コーパル、ビルマ産コハク、ドミニカ産コハク、中国産コハク及び /又はアフリカ産コーパルから成るグループの中から選択される、請求の範囲第 21項に記載の方法。 24.天然の樹脂が表面殺菌されている、請求の範囲第21項に記載の方法。 25.生物体が、粉砕抽出方法によって天然の樹脂から抽出される、請求の範囲 第21項に記載の方法。 −3− (4) 特表平9−508789 26.樹脂が、粉砕に先立ち液体窒素で前処理される、請求の範囲第25項に記載 の方法。 27.樹脂を粉砕に先立ち約−20℃から約−140℃で凍結させる、請求の範囲第2 5項に記載の培養。 28.樹脂が−80℃で凍結される、請求の範囲第27項に記載の培養。 29.生物体が、粗砕抽出方法によって、天然の樹脂から抽出される、請求の範 囲第21項に記載の方法。 30.生物体が、せん孔抽出方法によって、天然の樹脂から抽出される、請求の 範囲第21項に記載の方法。 31.生物体が、フラッシング抽出方法によって天然の樹脂から抽出される、請 求の範囲第21項に記載の方法。 32.生物体が樹脂中に化石化された昆虫材料内に含有されている、請求の範囲 第21項に記載の方法。 33.昆虫材料が、昆虫組織、昆虫の糞便及び/又は昆虫の巣を含むグループの 中から選択される、請求の範囲第32項に記載の方法。 34.生物体が、樹脂の中に化石化された植物材料の中に含有され ている、請求の範囲第21項に記載の方法。 35.植物材料が、根、葉、種子、小枝、樹皮、植物組織、胞子、花粉、花、イ ネ科牧草、シダ及びマメ科植物を含むグループの中から選択される、請求の範囲 第34項に記載の方法。 36.生物体が樹脂中で化石化された土壌の中に含有されている、請求の範囲第 21項に記載の方法。 37.生物体が樹脂中で化石化された水の中に含有されている、請求の範囲第21 項に記載の方法。 38.培養を含む生物体の年令が核酸分析によって決定される、請求の範囲第21 項に記載の方法。 39.生物体が、細菌、真菌、ウイルス、原生動物、微小藻類、節足動物及び/ 又は線虫から成るグループの中から選択される、請求の範囲第21項に記載の方法 −4− (5) 特表平9−508789 。 40.生物体が微生物である、請求の範囲第21項に記載の方法。 41.天然の樹脂を殺菌し、この樹脂から植物細胞を抽出し、この抽出された植 物細胞を培養することによって、天然の樹脂から得られる植物細胞又は胞子を含 む植物細胞の培養。 42.植物細胞が、天然の樹脂の中で化石化された真菌の菌糸体から得られる、 請求の範囲第41項に記載の培養。 43.天然の樹脂内で化石化された細菌又は真菌の胞子から得られる、請求の範 囲第41項に記載の培養。 44.植物細胞が、天然の樹脂の中で化石化された植物胞子から得られる、請求 の範囲第41項に記載の培養。 45.植物細胞が、天然の樹脂の中で化石化された原生動物の嚢子又は胞子から 得られる、請求の範囲第41項に記載の培養。 46.天然の樹脂を殺菌し、この樹脂から植物細胞を抽出し、この抽出した植物 細胞を培養することによって天然の樹脂から得られる 植物細胞又は胞子を含む植物細胞の培養を得るための方法。 47.古代生物体の年令を決定するための方法において、(a)古代生物体から 核酸を抽出する段階;(b)リボソームリボ核酸をコードする核酸配列を増幅す る段階、(c)増幅した配列をクローニングする段階、(d)クローニングされ た核酸を配列決定する段階、(e)最も近い実在の微生物についてのリボソーム リボ核酸配列とクローニングされた核酸の配列を整列させる段階;(f)古代微 生物についての核酸配列と実在の生物体のリボソームリボ核酸配列の間のヌクレ オチド置換の数を決定する段階;(g)2(0.3−0.4×109 )×〔比較されるヌ クレオチドの数〕により置換数を除する段階、を含んで成る方法。 48.古代生物体が微生物である、請求の範囲第47項に記載の方法。 49.天然の樹脂から分離された生物体の培養。 50.生物体が、細菌、真菌、ウイルス、植物、原生動物、微小藻類、節足動物 及び/又は線虫から成るグループの中から選択される、請求の範囲第47項に記載 −5− (6) 特表平9−508789 の培養。 51.天然の樹脂がコハク又はコーパルである、請求の範囲第49項に記載の培養 。 52.生物体が樹脂中で化石化された昆虫材料から分離される、請求の範囲第49 項に記載の生物体の培養。 53.生物体が、樹脂の中で化石化された土壌から分離される、請求の範囲第49 項に記載の生物体の培養。 54.生物体が、樹脂の中で化石化された植物材料から分離される、請求の範囲 第49項に記載の生物体の培養。 55.生物体が、樹脂の中で化石化された水から分離される、請求の範囲第49項 に記載の生物体の培養。 56.生物体が微生物である、請求の範囲第47項、第48項、第49項、第50項、第 51項、第52項、第53項、第54項又は第55項に記載の培養。 57.請求の範囲第1項、第41項又は第49項に記載の生物体により産生される化 学療法作用物質。 58.抗菌活性を有する、請求の範囲第57項に記載の化学療法作用物質。 59.抗微生物活性を有する、請求の範囲第57項に記載の化学療法作用物質。 60.抗真菌活性を有する、請求の範囲第57項に記載の化学療法作用物質。 61.抗ガン活性を有する、請求の範囲第57項に記載の化学療法作用物質。 62.抗原生動物活性を有する、請求の範囲第57項に記載の化学療法作用物質。 63.抗炎症活性を有する、請求の範囲第57項に記載の化学療法作用物質。 64.酵素阻害物質である、請求の範囲第57項に記載の化学療法作用物質。 65.免疫調節物質である、請求の範囲第57項に記載の化学療法作用物質。 66.請求の範囲第1項、第41項又は第49項の生物体によって産生される酵素。 67.プロテアーゼ、アミラーゼ、エステラーゼ及びホスホターゼを含むグルー プの中から選択される、請求の範囲第66項に記載の酵素。 68.請求の範囲第1項、41項又は49項に記載の生物体によって産 −6− (7) 特表平9−508789 生される生物殺虫作用物質。 69.請求の範囲第1項、41項又は49項に記載の生物体によって産生される生物 除草作用物質。 70.請求の範囲第1項、41項又は49項に記載の生物体によって産生されるビタ ミン。 71.生物体を発酵プロセスにおいて使用し得る、請求の範囲第1項、第41項又 は第49項に記載の生物。 72.組換え型ベクターとして請求の範囲第1項、第41項又は第49項の生物体を 使用するための方法。 73.NRRLに寄託され、B-21177という受入れ番号が割当てられた、古代バシラ ス属菌株BCA1。 74.NRRLに寄託され、B-21178という受入れ番号が割当てられた、古代バシラ ス属菌株BCA3。 75.NRRLに寄託され、B-21179という受入れ番号が割当てられた、古代バシラ ス属菌株BCA5。 76.NRRLに寄託され、B-21180という受入れ番号が割当てられた、古代バシラ ス属菌株BCA7。 77.NRRLに寄託され、B-21181という受入れ番号が割当てられた、古代バシラ ス属菌株BCA13。 78.NRRLに寄託され、B-21182という受入れ番号が割当てられた古代バシラス 属菌株BCA15。 79.NRRLに寄託され、B-21184という受入れ番号が割当てられた古代バシラス 属菌株BCAEx2。 80.NRRLに寄託され、B-21183という受入れ番号が割当てられた古代バシラス 属菌株BCA16。 81.NRRLに寄託され、B-21357という受入れ番号が割当てられた古代シュード モナス属菌株AG-10-DA-1。 82.NRRLに寄託されY-21355という受入れ番号が割当てられた古代サッカロミ −7− (8) 特表平9−508789 セス属菌株AG-11-DM-6。 83.NRRLに寄託され21356という受入れ番号が割当てられた古代アクチノミセ ス属菌株AG-11-AC-14。 84.NRRLに寄託され21354という受入れ番号が割当てられた古代ペニシリウム 属菌株AG-11-BA-5。 85.NRRLに寄託され21353という受入れ番号が割当てられた古代クラドスポリ ウム属菌株AG-13-LA-5。 86.NRRLに寄託され21352という受入れ番号が割当てられた古代ストレプトミ セス属菌株AG-15-WA-19。 87.請求の範囲第73,74,75,76,77,78,79,80,81,82,83,84,85又は 86項に記載の生物体の副産物。 −8− (9) 特表平9−508789 【発明の詳細な説明】 古代生物体 発明の技術分野 本発明は、コハクといった供給源からの古代生物体の回収及びかかる古代生物 体を分離し培養する方法に関する。本発明は、化石化された生物体を回収し培養 して生活能力を呈するようにすることができるということを初めて明らかにする ものである。本発明の古代生物体及びその副産物に関しては、農業、工業プロセ ス、生物治療、診断及び疾病処置におけるその利用を含め、多くの用途が構想さ れている。 発明の背景 コハクは、その他の合成重合体ガラスに共通の機械特性、誘電特性及び熱特性 をもつ植物からの天然の無定形重合体ガラスの化石樹脂である、Pointer,G.O., 「コハクの中の生命」、Stanford University Press,Stanford,CA(1992)。コ ハクの源である植物樹脂は、テルペノイド化合物、酸、アルコール及び精油の複 合混合物を含んでいる。樹脂は老化するにつれて、より硬いものとなりコーパル として知られる半化石化産物を形成する。近年では、(約400万年前以後の)化 石化した木の樹脂はコーパルとみなされ、それより古い木の化石樹脂は一般にコ ハクとみなされている。コハクとコーパルは、融点、硬度、可溶性などといった さまざまな物理的特性を研究することによって、さらによく区別できる。 既知の最古のコハクは古生代の石炭紀(3億6000万∼2億8600万年前)から回 収されたものである。充分に研究されたコハクの大部 分は、中生代の白亜紀(6500万年∼1億4400万年前)及び新生代の第3紀(200万 年∼6500万年前)からのものである。コハク及びその特性についての広範な論述 に関しては、上述のPoninarを参照のこと。 コハクは、細胞オルガネラを含む生物学的材料を保存する著しい能力を有する と思われる。例えば、今世紀に始まって、科学者達はコハク封入体の中の組織の 保存を観察してきた。上述のPoinar 266∼271を参照。特定的には、ミトコンド リア及び染色質を含む保存状態の良好な細胞及び細胞構成要素が、バルト諸国産 −9− (10) 特表平9−508789 コハクの中に化石化された4000万年前のハエの中に観察されてきた。Poinar及び Hess,Science 215:1241∼1242(1982)。 さらに最近では、科学者達は、コハクが無傷の古代遺伝物質を保存できるか否 かを評価し始めている。1980年中期に始まって、例えば科学者達は、広範囲にわ たる古代の供給源から生活能力ある生物材料、特定的は古代DNAを回収すること を試みた。古代DNAを抽出する初期の実験は、博物館保存の外皮について行なわ れた。Higuchi et al.,Nature 312:282-284(1984),Thomas et al.,J.Mol.Ev ol.31:101-112(1990)、ミイラ化組織、Paabo,Nature 314:644-645(1985)a nd Lawlor et al.,Nature 349:785-788,(1991),bones,Hagelberg et al., Nature 342:485(1989);Hagelberg et al.,Phil.Trans.R.Soc.Lond.B.333 :399-407(1991);Hanni et al.,Acad.Sci.Paris Ser.III.310:365-370( 1990);Hagelberg and Clegg.Proc.R.Soc.Lond.B.244:45-50(1991);Horai et al.,Phil.Trans.R.Soc.Lond.B.333:409-417(1991);Hummel and Herrma n,Naturwissenschaften 78:266-267(1991)、植物化石、Golenberg,Nature 344 :656-658(1990)、凍結したマンモス、Higuchi and Wilson,Federation Proc. 43:1557(1984);Ch erfas,Science 253:1354-1356(1991),and 古代種子、Rogers and Bendich,Pl ant Mol.Biol.5:69-76(1985),Rollo et al.,Nature 335:774(1988)。しか しながら、コハクの中に埋没させられた化石化材料から生活能力あるDNAを分離 しようとするこれらの繰返しの試みは失敗に終わった。 コハク化石からの古代DNAの分離の成功が初めて報告されたのは、1992年であ り、このとき、2500万年∼4000万年前のドミニカ産コハクの中に保存された絶滅 したハチ、Proplebeia dominicanaからDNAフラグメントが分離された。Cano et al.,Med.Sci.Res.20;249-251及び619-622(1992)。きわめて最近になって、古 代レバノン産コハクの中に保存された1億2000万年∼1億3500万年前のゾウムシ からDNAがPCR増幅され、配列決定された。Cano et al.,Nature 363:536-538(1 993)。これらの報告書は、コハクが或る程度古代遺伝子情報を保存できるという ことを示している。 −10− (11) 特表平9−508789 それでも、コハク及びその他の古代材料からの古代DNAの回収の成功の報告に もかかわらず、これらの研究において配列決定された遺伝物質が真に古代のもの であるのか又は近年の汚染物質によるものであるのかに関して、多々の懐疑論が 科学界に存在している(「古代DNA」、Springer-Verlag,10,62∼64,158∼160 ,221∼222を参照のこと)。その他の科学者は、DNA分子、コハクに固有の不安 定さのため、生活能力あるDNAが何百万年もの間無傷の状態にとどまることは不 可能である、と主張している。Lindhal,Nature;362;709∼715(1993)。 さまざまな供給源からの古代DNAの抽出に関して実施されている豊富な研究作 業にもかかわらず、コハク又はそれに類似する天然に発生する樹脂材料の中に保 存された古代細菌及びその他の古代生物材料の回収に関しては、きわめてわずか な研究作業しか実施されて いない。この研究作業の意味はさらに、結果が確定的でないことによっても妨げ られている。例えば、細菌杆状体及び真菌胞子(Micrococcus electroni,Bacill us electroni,Longibacillus electroni及びSpirillum electroni)ならびに花 粉が、1929年にすでにテルペンチン(松やに)中でコハクを溶解させた後に観察 されているが(Blunk,G.,Bacterienneischlusse imm Berstein,Centraalblat t fur Mineralogie,Geologie und Palaontoligie(ABt.B,nom II 554-5)(1929) 、引用書Pointer,G.O.,「コハク中の生命」350,Stanford University Press ,Stanford,CA(1992)中)、これらの生物体は近代の実験室汚染物質のせいで あった。 同様に、古生代の塩から分離された生活能力ある細菌の報告書(Dombrosky,H. J.,Zentr.Bakteriol,ParasitenK.Abt.I.178;83-90(1960);Dombrosky.H. J.,Zentr.Bakteriol Parasitenk.Abt.1.183:173-179(1961))は、このような 土壌沈積物内に捕えられた古代微生物のプールが地下水の溶浸を介してより近年 の微生物で汚染された状態になった確率が高いことから、近代の細菌汚染物質の せいであるとされた。1983年に、メキシコ産コハクの中に無傷の細菌細胞が保存 されていることが観察された(上述のPoinar)。しかしながらこの場合も、細胞 の年令も真正性も確認されておらず、近代の細菌による汚染が疑われている。従 −11− (12) 特表平9−508789 って、コハクから生活能力ある細菌を回収するこれらの試みの各々の結果は、確 定的なものではなかった。 過去80年間、医療、工業及び農業の応用分野及び生物治療のための微生物及び 微生物副産物の集約的な商業的開発が行なわれてきた。本書に参考として内含さ れているHarvey,ed.,天然産物からの薬物:医薬及び農薬、Ellis Horwood Ltd .,England(1993)。例えば、抗生物質の大規模な科学的開発が、真菌Penicilliu m notatum によって産生されたペニシリンについての研究作業により1928年に開始された。 同様にして、工業的用途のための微生物プロセスの集約的使用は、嫌気細菌Clos tridium acetobutylicumを用いたアセトンについての研究、Domain and Solomon 、「工業的微生物学と遺伝子工学の到来」Scientific American p3∼11,W.H.Fr eeman and Co.San Francisco(1981)に伴い、1940年代に増大した。微生物の比 較的わずかな種が現在抗微生物化合物の産生のために活用されてきており、医療 又は工業プロセスでの使用のためにはその他の微生物副産物が活用されている。 例えば、抗微生物化合物及び工業的微生物副産物を大量に産生するためには、わ ずか3つの微生物グループ(糸状真菌、非糸状細菌及び糸状菌又はアクチノミセ ス)の微生物しか使用されない。 抗微生物化合物を産生することに加えて、微生物は、心臓血管及び抗炎症作用 物質、免疫調節物質、抗腫瘍化合物、中枢神経系の調節物質及び酵素阻害物質を 含む(但しこれに制限されるわけではない)その他の薬剤利用分野として使用さ れてきたさまざまな代謝産物を産生する。例えば、Harwood,C.R.,バイオテク ノロジー便覧:バシラス、294以降、Plenum Press,New York(1989)を参照の こと。微生物代謝産物は同様に、合成化学及び合理的薬物設計のためのプラット フォーム分子としても役立ち、ワクチン製造の基盤として広く用いられている。 Harvey,ed.,「天然産物からの薬物;医薬及び農薬」、Ellis Horwood Ltd,En gland(1993)参照。 微生物及び微生物副産物は、酵素及びビタミンの産生及び発酵といったさまざ まな工業プロセスの中でも使用されてきた。例えば、B.subtilisは、洗たく洗 −12− (13) 特表平9−508789 剤の中に広く用いられているさまざまな熱安定性セリンプロテアーゼ酵素を産生 する。同上。ビール、ワイン、酒及びその他の食料品ならびにさまざまなビタミ ンの製造にお いて、サッカロミセス、アスペルギルス及びカンジダを含むさまざまな真菌種が 用いられる。微生物副産物は、化粧品、バイオポリマー、界面活性剤、プリンヌ クレオシド及びフェノール系殺菌剤の製造においても使用されている。同上。 さらに最近では、環境汚染についての関心が増大しているため、生物殺虫剤及 び生物治療のために微生物及び微生物副産物が使用されつつある。例えば、さま ざまな植物及び昆虫有害生物を撲滅するための生物殺虫剤としてB.sphaericus 及びB.thuringiensisが商業的に使用されている。上述のHarwood,309、を参照 のこと。同様に、桿菌種が、有毒汚染物質からの環境浄化において日常的に使用 されている。同上、313;Debabov、「桿菌の分子生物学」第1巻(D.A.Dubnau, ed)中の「桿菌の工業的利用」、Academic Press,New York(1982),331∼370。 同書は本書中に引用により組み入れられている。 さらに、微生物及び微生物副産物は、診断検定において増々使用されてきてい る。例えば、B.subtilisは、ストレプトマイシン、ペニシリン及びカトマイシ ンを検出するための検定において使用されている。J.Bact.45:408∼409(1943) ,J.Bact.49:411(1945),Appl.Microbiol.4:307-310(1956)を引用するATCC カタログ、34(1994)を参照のこと。 動物及び人間の健康、農業、工業及び環境上のプロセスにおけるパターン変化 のため、医療、診断及び工業的利用分野のための微生物副産物の新しい供給源を 探求することが必要となっている。例えば、数多くの病原体が、現在近代の微生 物から誘導されている抗生物質に対する耐性をもつようになってきている。Cook sey,R.C.,「抗微生物作用物質に対する耐性のメカニズム」臨床微生物学マニュ アル、1099、第5版、Am.Soc.for Microbiology(1991)。同様に 、広く用いられている数多くの農薬が、植物相及び動物相に対し有害で水資源に 対する汚染をひき起こすものであることが実証されてきた。従来の抗微生物化合 −13− (14) 特表平9−508789 物に対する耐性の増大、近代の工業プロセスにより作り出された新しい需要、化 学的殺虫剤及び除草剤に対する代替物を開発する必要性そして環境汚染物質を削 除する必要性が、新しい微生物とその副産物を使用するための好機を作り上げて いる。今日までに知られている微生物副産物のアイデンティティ及び既存の用途 の広範な要約は、本書に引用により組み入れられているBiotechnology、第1巻 ∼8巻、H.J.Rehm及びG.Reed、編集、Verlag Chemie(1986)の中に記されている 。 発明の要約 本発明は古代生物体及び生物材料を何百万年も保存することのできる供給源か ら古代生物体及び生物材料を回収することに関する。より特定的に言うと、本発 明はコハク及び類似の天然に発生する樹脂から分離された古代微生物に関する。 かかる微生物には、細菌、真菌、原生動物、ウイルス及び微小藻類が含まれるが 、これらに限られるわけではない。さらに、本発明は、花粉、植物、異なる分類 群の節足動物及び線虫を含め、コハク及び類似の天然に発生する樹脂から古代生 物体及び生物物質を回収することに関する。 本発明は、古代微生物菌株を2500万年以上の間コハクの中に化石化されていた 後に回収し生活能力ある状態へと生き返らせることができるというきわめて驚く べき発見に基づいている。以下でより詳細に記述する本発明の特定の一実施態様 においては、約2500万年∼4000万年前にドミニカ産コハクの中に化石化された4 匹の古代無針バチProplebeia dominicanaの保存された腹腔組織から、古代バシ ラス菌の約12の生活能力ある菌株が分離された。本発明の古代バシ ラスを回収するのに用いられたコハク標本は、厳密に認証され、抽出及び無菌培 養条件は、近代微生物による汚染の可能性を除去するように設計され、抽出の結 果もたらされる生活能力あるあらゆる微生物の古代状態を確認するためさまざま な対照及び偽似抽出が実施された。 コハク由来の生物体の古代性を立証するため、コハクに埋没したハチの近代子 孫の腹腔から近代バシラス種を分離し、これを分類学的、生化学的及び遺伝学的 分析を介して古代バシラス分離株と比較した。古代及び近代の両方の細菌分離株 −14− (15) 特表平9−508789 の16SrDNAの一部分を、PCRを用いて増幅し、配列決定した。近代及び古代バシラ スの配列を比較し約93∼95%の相同性を有することを発見した。以下でより詳し く記述する「分子時計」分析を用いて、古代分離株のいくつかのヌクレオチド置 換率を計算した。これらの計算に基づいて、古代生物は約5000万年前のものであ ることが決定され、これは、それらの古代生物が分離されたドミニカ産コハクの おおよその年代(2500万年∼4000万年)と整合性あるものである。 結果として、出願人が分離し本書で記述しているバシラスが実際に2500万年∼ 4000万年の間休眠状態にあり続けた回復した古代生物体であることには全く疑い がなかった。 本発明のもう1つの特定の実施態様においては、250年から1億2000万年まで の年代のさまざまなコーパル及びコハクの中の数多くのタイプの生物学的封入体 から、10以上のさまざまなバシラス以外の古代微生物が分離された。これらの生 物体には、アクチノミセス、ストレプトミセス、ペニシリウム、酵母及びシュー ドモナスの各種が含まれるが、これらに限られるわけではない。 これらの古代微生物は、その形態学的、生化学的及び酵素的プロフィールに関 して記述され、さまざまな商業的に有用な薬学製品、 酵素(例えばプロテアーゼ、アミラーゼなど)及び防腐剤の製造用、生物学的殺 虫剤として、及びさまざまな工業的プロセス内で、といったものを含む(ただし これらに限られるわけではない)数多くの農業、工業、環境及び医療用途をもつ ものと予想されている。さらにこれらの古代微生物のいくつかは、連鎖球菌及び ブドウ球菌といった或る種の近代動物病原体ならびにErwina carotovaraといっ た或る種の農業病原体の成長を阻害する能力を示す抗微生物性物質を分泌し、従 って抗生物質の製造のために使用することができる。古代微生物、特に古代桿菌 は、組換え型DNAベクターのための宿主としても使用できる。古代微生物及びそ れらが分泌する物質は、環境浄化、採鉱、診断及び食品製造の分野でも応用でき る。 同様に、古代微生物及びその副産物は、有用な工業的利用分野を有する可能性 がある。例えば、近代微生物は、現在工業的酵素の重要な生産者である:サッカ −15− (16) 特表平9−508789 ロミセス種といった酵母の菌株はアルコール飲料の醸造において使用されている 。このような酵母は、土壌及び堆積物試料の中にも見い出すことができる。一般 にDomain及びSolomon「工業用微生物の生物学」Benjamin/Cummings Publishing Company(1985)を参照のこと。微生物は現在生物殺虫剤としても用いられている 。たとえば、Bacillus sphaericus及びBacillus thuringiensisは往々にして昆 虫と結びついた状態で発見される。Mitscherlich及びMartin、「環境内での微生 物の生存」、Springer-Verlag(1984)。この情報は、潜在的な商業的利用分野を もつ古代微生物を誘導できるコハク又は樹脂内の生物材料の供給源を同定する上 で有用である。しかしながら、これらの古代微生物は遺伝的にその近代相対物と 異なっていることから、古代微生物が、潜在的に新規な化学構造をもつ異なる副 産物を産生できるのではないかと期待することができよう。 図面の簡単な説明 図1−2500万年∼4000万年前のコハクの中に化石化された古代無針バチの腹部 組織から抽出されている試料の写真。 発明の詳細な説明 古代微生物沈積物の多様性 コハク及びそれに関連する天然に発生する樹脂から回収することのできる生物 には、細菌、真菌、原生動物、ウイルス、微小藻類、花粉、植物(コケ及び地衣 類、シダ及び高等植物)、さまざまな分類群の節足動物及び線虫が含まれるが、 これらに制限されるわけではない。本発明の1つの態様は、細菌、特にバシラス 属の一員を含む(但しこれに限られるわけではない)内生胞子形成菌といった古 代微生物を分離することに関する。 本発明のその他の態様は、アクチノミセス属、ストレプトミセス属、サッカロ ミセス属、アスペルギルス属、ペニシリウム属、ミクロコッカス属、アルトロバ クター属、コリネ型属及びシュードモナス属の一員を含む(ただしこれに限られ るわけではない)細菌及び真菌といった古代微生物を分離することに関する。本 発明のさらなる態様は、pH及び温度の両方の極限範囲を含む(ただしこれに限ら れるわけではない)極限条件下で成長することのできる細菌及び真菌といった生 −16− (17) 特表平9−508789 物を含む(ただしこれに限られるわけではない)古代微生物を分離することに関 する。 古代微生物供給源 古代生物体及び生物材料は、約50年から1億2000万年以上前までの年代範囲で さまざまな地理的場所からのコハク及び関連する天然に発生する樹脂から回収す ることができる。生物は、樹脂マトリックス自体からだけでなく、植物胞子、微 生物胞子、土壌標本、水及 び気泡、微生物、真菌、原生動物、昆虫の巣、昆虫の糞便、昆虫、植物材料例え ば葉、枝、樹皮、根、花、牧草、シダ、マメ科植物及びさや、花粉、線虫、節足 動物及び貝がらを含む、樹脂内に埋没した状態で発見されたさまざまな封入体か らも回収され得る。 真正なコハク又は類似の天然に発生する樹脂は、ビルマ、ドミニカ共和国、バ ルト海地方、カナダ、中国、コロンビア、スマトラ、チアパス、ルソン、ルーマ ニア、シシリア、アラスカ北極沿岸、アメリカ北東部、マニトバのシーダー湖地 方、フランス北部、レバノン、イスラエル、ヨルダン及び東タイムイル(北極ソ 連)を含む世界のいくつかの地域に見い出すことができる。古代生物体又は生物 材料が回収される供給源が実際に真正なものであることを確認することが重要で ある。樹脂標本の供給源を入念に追跡し確認することに加えて、標本の年代を、 できれば標本を採取した地層内で発見される化石を調査することによって間接的 に決定することができる。化石化した樹脂沈積物をとり囲む化石の年代は、化石 化した樹脂のおおよその年代を提供してくれる。例えば、いくつかの貝がら形成 原生動物は、特定の地質学的時期に豊富であったし、このような原生動物の化石 がコハクと共に発見された場合、コハクがその地質学的時期の間に形成されたも のと仮定することができる。標本を真正性認証し、特徴づけし直接年代決定する ためのその他の方法には、x線回折、核磁気共鳴分光法、熱分解ガスクロマトグ ラフィ及び赤外線分光法(一般に上述のPoiner参照)が含まれる。 近代生物の既知の生息地に関する情報を用いて、当該出願の古代生物体の潜在 的供給源を同定することができる。 −17− (18) 特表平9−508789 例えば、さまざまな微生物と昆虫の間の今日の関係についての知識は、古代微 生物を回収することのできる供給源を同定するための指針を提供することができ る。例えば、ハチ及びさまざまな生物体 は、自らを病原体から保護しかつその食物を加工し保存するのに共生微生物を利 用するように思われる。従って、ハチのこのような微生物共生体についての現在 の知識に基づいて、ハチの祖先も古代ハチ病原体と戦いその食料を加工し保存す るために微生物共生体を使用していたと推論することができる。より特定的には 、熱帯の無針バチMelipona fasciataは、自らの巣の中に食物を保存するために 胞子形成(芽胞)菌を使用すると思われる。B.megaterium,B.circulans及びB .alveiを含むM.fasciataの巣の中に保管された食物から、さまざまなバシラス 生物が分離された(Gilliam et al.,Apidologie 21:89-97(1990))。これらの細 菌の生化学特性に基づいて、著者は、ハチがこれらの細菌を自らの食物を保存し 加工するために使用したということを示唆した。細菌とハチの間の関連性につい てのその他のケースも報告されてきた。例えば、B.brenis,B.megaterium,B .coagulans,B.cereus、及びB.subtilisは、ミツバチのさまざまな器官から 分離された(Apis mellifera,Gilliam et al.,J.Invert.Pathol.31:389-391( 1978)。B.subtilis及びB.cereusは同様に、単生バチCraw fordapsis luctuosa からも分離されている。Gilliam et al.,Apideologie 21:99-105(1990)。 細菌に加えて、酵母も、ハチが収集した花粉を加工する上で1つの役割を果た していると思われる。Gilliam et al.,Apidiologie10:53-53(1979)。ハチ又は ハチの巣から分離されたいくつかの細菌(例えば、B.subtilis)は、マイコサブ チリンといった抗生物質を産生する。J.Antibiotics 29:1043-1049(1976)。従 って、このような共生細菌が、病原体から自らを防護する上でハチを援助すると いうことが示唆されてきた。Gilliam et al.,Apidiologie 21:99-105(1990) 。古代のハチにおいても古代微生物共生体によって 類似の役割が演じられてきた可能性があるものの、古代に存在した病原体の共同 体及び温度、気体濃度といったきわめて異なる環境条件から見て、このような微 −18− (19) 特表平9−508789 生物の祖先が、正確な近代の同等物が全く存在しないような何らかの物質を産生 していた確率も高い。 同様に、アクチノミセス、ストレプトミセス、ペニシリウム及びセファロスポ リウムといった抗生物質産生微生物が、地中生息地内に一般に発見されるか、又 は植物材料と結びつけられている。同様に、Bacillus subtilisといった工業用 酵素の既知の生産者である微生物も又、地中生息地内に見い出される。アルコー ル飲料の醸造に用いられるサッカロミセス種といった酵母の菌株も、土壌及び堆 積物標本中に発見され得る。一般に、Domain and Solomonの「工業用微生物の生 物学」、Benjamin/Cummings Publishing Company 1985を参照のこと。Bacillus sphaericus及びBacillus thuringiensisといった現在生物殺虫剤として用いら れている微生物は往々にして昆虫と結びついた状態で発見される。Mitscherlich and Martin、「環境内の微生物の生存」Sprirger-Verlag(1984)。ここでも又、 古代微生物が遺伝的にその近代相対物と異なることから、古代微生物が潜在的に 新しい化学構造をもつ異なる副産物を産生する能力をもつということを期待する ことができる。 古代微生物を分離し培養するための方法 適切な封じ込め及び汚染防止手段が用いられることを条件として、本発明の1 つの態様を実施する上で、コハク又はそれに類する天然に発生する樹脂又はこの ようなコハク又は樹脂の中に化石化された生物体から直接、微生物、生物体及び その他の生物材料を分離するためのさまざまな方法を使用することができる。こ のような防止手段の1つには、化石化された生物材料又は生物体を含有する樹脂 標本の外部表面の殺菌が関与する。 本発明の1つの好ましい実施態様においては、コハク標本の外部表面は、以下 のように殺菌される:すなわち、コハク標本を(1)風袋を計った秤量ボート内 で秤量する;(2)真空下で12時間35℃で2%の緩衝グルタルアルデヒド内に浸 漬させる;(3)無菌の2度精製された水(SDDW)の中で3度洗浄し、次にトリ プチカーゼダイズ寒天(TSA)(BBL,Cockeysuille,MD)上で洗浄水を培養して殺菌 手順の有効性を評価する;(4)37℃でさらに6時間10%の漂白剤中に浸漬させ −19− (20) 特表平9−508789 る;(5)SDDW中で3回洗浄し、次に洗浄水をTSA上で培養して殺菌手順の有効 性を評価する;(6)35℃で2時間70%のエタノール中に浸漬させる;そして( 7)アルコールを蒸発させるため炎に露呈する。この表面殺菌されたコハクの標 本をトリプチカーゼダイズブロス(TSB)上で培養させて殺菌プロセスを評価する 。 コハク及び/又はその他の天然に発生する樹脂標本を殺菌し及び/又は除染す るその他の方法としては、プラズマエッチング、音波処理、定着剤、溶剤、ガス くん蒸、殺胞子剤の適用及び標本からの樹脂外部層の除去、がある。 化石材料の標本を次に、無菌計器、無菌技術を用い、無菌条件下で抽出する。 例えば、埋没した化石の特定の組織を露呈するため液体窒素内での凍結後にコハ クを粗砕(クラッキング)することができる。さらに、目に見える生物材料、鉱 物又は生物体化石を伴う又は伴わない標本を、標本の表面の除染の後無菌条件下 で粉末又は半粉末材料の形へと摩砕及び/又は粉砕することができる。無菌砂又 はモルタル及び乳棒の使用を含め任意の数の方法によって標本を粉末又は半粉末 の形に摩砕することができる。さほど破壊的でなくより標的を定めた分離方法は 、標本の保存そして古代微生物を回収するために化石又はその他の封入体のどの 部分が除去されるかについ ての制御という2重の利点を提供してくれる。このような方法としては、例えば 、標本全体を破壊することなく化石又はその他の封入体の非常に小さい試料を抽 出することのできる中空中ぐりマイクロドリルの使用が考えられる。このような 方法には、さらに、樹脂標本を凍結させる技術の使用、指向性レーザー、標本を 粉末に摩砕するための無菌砂の使用、熱、無菌粉砕プレス及び化学処理の使用が 含まれるかもしれない。乾燥した化石の水和作用が標本の抽出を容易にすること ができる。 抽出された標本は、次に、封じ込められ除染された環境内で生活能力ある古代 微生物を回収するためのさまざまな条件下で微生物を増殖させるのに現在用いら れているさまざまなタイプの増殖培地内に入れることができる。 特定のタイプの微生物に分離をターゲティングするため、古代微生物が生活能 −20− (21) 特表平9−508789 力ある培養を生成できるようにする条件の変動を用いることができる。例えば、 増殖温度を極限に調整することにより、異常な条件下で増殖することのできる微 生物を培養する一方で、かかる条件で生き延びることのできないその他の微生物 を選び除くことができる。増殖培地の成分も選抜を目的として調整することがで きる。本書に記述されている方法に加えて、本書には明示的に記述されていない その他の殺菌方法及び増殖条件も当業者には直ちに明らかになることと思われ、 適正な汚染防止手段を維持しかつ古代微生物を分離し増殖させるためにこれを使 用することも可能である。 本発明の一実施態様においては、古代微生物は、樹脂標本を粉砕し(「粉砕抽 出方法」)次に抽出された微生物を培養することによって、殺菌されたコハク又 はそれに類する天然の樹脂標本から抽出される。特定的には、以下のプロトコル に従うことができる。 1.上述の通り樹脂標本を殺菌する。 2.安全キャビネット内で、その中の無菌モルタルの中に表面殺菌された標本を 入れ、液体窒素でフラッディングする。 3.無菌乳棒を用いてか又は無菌粉砕プレスを通して過冷却したコハクを置くこ とによって、過冷却コハクを破砕し粉砕する。 4.(a)アクチノミセスブロス(Difco);(b)1%のペプトン(フラスコ2 本);及び(c)トマトジュース寒天(Difco)である無菌ブロス25mlの入った4 本の250ml入りエルレンマイヤーフラスコの各々の中に同等にコハク粉末を分配 する。 5.1%のペプトンを含む1本のフラスコを55℃で、又残りの3本のフラスコを 30℃で、200RPMの軌道振とう機内にてインキュベートする。 6.第1週目は各フラスコから毎日、次に4週間の間、週に一度、500μlをサ ンプリングして次の培地上に広げる:(a)TSA;(b)ポテトデキストロース 寒天(PDA)、(c)0.4%の酵母エキスを伴うツザペク(Czapek)寒天(Difco); (d)2%の無菌スキムミルク(SMA)を伴う生菌数寒天(plate count agar,D ifco);(e)SMA pH 8.1;及び(f)でんぷん寒天(Difco)。各皿(plate)を適 −21− (22) 特表平9−508789 当な温度でインキュベートする。(55℃でインキュベートすべき皿は、加湿した 無菌ペーパータオル又はスポンジと共に無菌の密封できる袋の内部に入れなくて はならない)。 7.コロニーの存在について毎日皿を検査する。 8.分離したコロニーを採収し、各々の微生物分離株(isolate)を適切な培地上 へ継代培養する。皿上の各コロニーを(−80℃で)凍結させる。各分離株の形態 及びグラム染色特性を評価するためグラム染色を行なう。ラクトフェノールコッ トンブルー封入剤の湿潤封入内で真菌及び酵母を検査しなければならない。 9.各皿から少なくとも10の分離株を凍結し、凍結乾燥させる。こ の時点で、各個体群の内部の遺伝的多様性の度合いを立証するため、各分離株に ついて、FAME(脂肪酸メチルエステル)分析を実施しなければならない。 この方法において、標本は、液体窒素を用いずに粉砕できる。 第2の方法では、古代微生物を抽出するべくコハク又はそれに類する天然の樹 脂は、殺菌後「粗砕」され(「粗砕抽出方法」)、次に、このような微生物が培 養される。特定的には、次のようなプロトコルを使用することができる: 1.上述のようにコハク又はそれに類する天然の樹脂を殺菌する。 2.安全キャビネット内で、無菌ペトリ皿底面上に望ましい封入体を伴う無菌標 本を置き、液体窒素でこれをフラッディングする。液体窒素を完全に蒸発させる 。過冷却された標本の外側に高温無菌食塩水を数滴落とす。 3.無菌の27ゲージの針を用いて、標本片を粗砕して望ましい組織を露呈させる 。 4.無菌針で組織を収穫し、無菌の1%のトリプトンブロスが入った2本の1.5m l入り無菌マイクロ遠心分離管の各々の中に移す。 5.15分間80℃にセットされた水浴中で1本の管に熱ショックを加え、次に両方 の管を適当なインキュベータ又は乾式浴の中で30℃でインキュベートする。 6.第1週目は毎日各々の管から、そしてその後は4週間週に一度、以下の培地 上へ1ループ分を画線する:(a)TSA;(b)PDA;(c)0.4%の酵母エキス を伴うツザペク寒天(Difco);(d)SMA;(e)pH8.1のSMA;(f)でんぷん寒 −22− (23) 特表平9−508789 天(Difco)。30℃で各皿をインキュベートし、コロニーの存在について毎日皿を 検査する。 7.分離したコロニーを採収し、各々の微生物分離株を適切な培地 上に継代培養させる。皿上の各コロニーを凍結させる(−80℃で)。各分離株の 形態及びグラム染色特性を評価するためグラム染色を実施する。真菌及び酵母を ラクトフェノールコットンブルー封入剤の湿潤封入内で検査しなければならない 。 8.各皿から少なくとも10の分離株を凍結及び凍結乾燥させる。この時点で、各 個体群内の遺伝的多様性の度合いを立証するため各分離株に対してFAME(脂肪酸 メチルエステル)分析を実施しなければならない。 第3の実施態様においては、古代微生物又は生物材料は、殺菌されたコハク又 はその他の天然の樹脂標本からせん孔によって抽出され(「せん孔抽出方法」) 、次に以下のプロトコルに従って培養される。 1.上述のとおりコハクを殺菌する。 2.安全キャビネット内で、無菌ペトリ皿上に急速硬化性膠を一滴落として、望 ましい気体、水又は液体封入体を伴うコハク片を置く。膠が硬化するまで待って から先に進む(約5分)。 3.無菌ドリルプレスに取り付けた無菌の0.5mm∼1.0mmのチタンドリルビットを 用いて、コハクの中に穴をあけ、このとき気体、水又は液体の泡に刺し通すよう 注意する。 4.ドラマンドマイクロピペットにとりつけた無菌のマイクロピペットチップを 用いてコハク封入体内から気体、水又は流体を収集する。無菌の1%のトリトン ブロスが入った1.5mlの無菌マイクロ遠心分離管の中へ、気体、水又は流体を移 す。 5.適切なインキュベータ又は乾式浴内で25∼30℃で管をインキュベートする。 6.第1週目は毎日各々の管から、そしてその後は4週間週に一度、以下の培地 上へ1ループ分を画線する:(a)TSA;(b)PDA −23− (24) 特表平9−508789 、(c)0.4%の酵母エキスを伴うツザペク寒天(Difco);(d)SMA;(e)pH 8.1のSMA;(f)でんぷん寒天(Difco)。25℃∼30℃で各皿をインキュベートし 、コロニーの存在について毎日皿を検査する。 7.分離したコロニーを採収し、各々の微生物分離株を適切な培地上に継代培養 させる。皿上の各コロニーを凍結させる(−80℃で)。各分離株の形態及びグラ ム染色特性を評価するためグラム染色を実施する。真菌及び酵母をラクトフェノ ールコットンブルー封入剤の湿潤封入内で検査しなければならない。 8.各皿から少なくとも10の分離株を凍結及び凍結乾燥させる。この時点で、各 個体群内の遺伝的多様性の度合いを立証するため各分離株に対してFAME(脂肪酸 メチルエステル)分析を実施しなければならない。 第4の実施態様においては、古代微生物又は生物材料は、生物体又は昆虫の化 石の組織に対しせん孔しペプトンブロス内で乾燥した組織を水和させることによ って、殺菌された化石樹脂から抽出され(「せん孔抽出方法」)、その後以下の プロトコルに従って培養される。 1.上述のとおりにコハクを殺菌する。 2.安全キャビネット内で、無菌ペトリ皿上に急速硬化性膠を一滴落として望ま しい封入体を伴うコハク片を置く。膠が硬化するまで待ってから先に進む(約5 分)。 3.無菌のドリルプレスに取りつけられた無菌の0.5mm∼1.0mmのチタンドリルビ ットを用いて、コハクの中に穴をあけるが、このとき封入体を通して刺すように 注意する。 4.ドラマンドマイクロピペットにとりつけた無菌のマイクロピペットチップを 用いて50μlの無菌の1%ペプトンブロスで封入体内 の組織を洗い流す。無菌の1%トリプトンブロスが入った1.5mlの無菌マイクロ 遠心分離管の中に流体を移す。 5.適当なインキュベータ又は乾式浴の中で25∼30℃で管をインキュベートする 。 6.第1週目は毎日各々の管から、そしてその後は4週間週に一度、以下の培地 −24− (25) 特表平9−508789 上へ1ループ分を画線する:(a)TSA;(b)PDA、(c) 0.4%の酵母エキ スを伴うツザペク寒天(Difco);(d)SMA;(e)pH 8.1のSMA;(f)でんぷ ん寒天(Difco)。25℃∼30℃で各皿をインキュベートする。 7.分離したコロニーを採収し、各々の微生物分離株を適切な培地上に継代培養 させる。平板上の各コロニーを凍結させる(−80℃で)。各分離株の形態及びグ ラム染色特性を評価するためグラム染色を実施する。真菌及び酵母をラクトフェ ノールコットンブルー封入剤の湿潤封入内で検査しなければならない。 8.各平板から少なくとも10の分離株を凍結及び凍結乾燥させる。この時点で、 各個体群内の遺伝的多様性の度合いを立証するため各分離株に対してFAME(脂肪 酸メチルエステル)分析を実施しなければならない。 これらのプロトコルの各々について、環境汚染物質についてテストするためフ ード内にTSAの皿を3枚置くことができる。皿は、抽出手順全体にわたり開放し た状態に置かれる。手順の終りで、ペトリ皿のフタを戻し、皿を2週間35℃でイ ンキュベートする。 古代微生物の同定及び特徴づけ 本発明の古代微生物を同定し特徴づけするため、近代微生物を同定し特徴づけ するさまざまな一般的及び特定的方法を使用することができる。この点で、古代 微生物は、形態学的、生化学的、遺伝学的及び生物学的に評価することができる 。例えば、さまざまな形態 学的特性を評価するため拡大して及び拡大せずに、古代細菌を目で検査すること ができる。菌株親和力、内生胞子特性及び成長条件も同様に、一般的に評価され る。古代細菌を特徴づけするためには、近代微生物を分類するのに標準的に用い られるもののような生化学特性を使用することもできる。付加的には、近代生物 体と古代生物体の間の関係又は区別をひき出すための手段としてさまざまな近代 生物体の特徴である広範囲の酵素の産生について、古代微生物をテストすること ができる。さらに、生物体を同定し特徴づけするための手段としてのみならず生 物体により産生される有用な生物活性をもつ分子を同定するために、当業者にと っては周知の技術を用いて、抗生物質及び/又はその他の生物活性をもつ物質の −25− (26) 特表平9−508789 産生について、古代生物をテストすることができる。 さらに、古代生物体を、その遺伝情報を評価することによって特徴づけするこ とができる。遺伝分析の1つのアプローチには、近代生物体の遺伝子配列から特 許されたプライマを用いて、ポリメラーゼ連鎖反応又はリガーゼ連鎖反応方法と いった適切な任意のDNA又はRNA増幅技術を用いて古代核酸を増幅することを含む 。近代生物体の遺伝子に関係づけすることのできる古代生物体の遺伝子をクロー ニングし配列決定して、遺伝的関係性及び進化上の分岐を評価し、かつ中でコー ド化されたタンパク質の構造を決定しなければならない。代替的には、古代微生 物の遺伝情報を、当該技術分野において既知のものである無作為プライマー増幅 又は配列非依存型プライマー増幅技術を用いて、分離することが可能である。さ らに、配列分析から大量のゲノムDNAを分離するために、古代微生物の培養を使 用することができる。 本発明の1つの実施態様においては、推定上の古代微生物分離株からDNAが抽 出され、適切なプライマーを用いて16S rRNA遺伝子の セグメントが増幅される。PCR増幅は、50μlの全体量で、1単位の低DNA Taqポ リメラーゼ(AmpliTaq-LD DNAポリメラーゼ、PerKin Elmer,Norwalk,CT)、2μ g/mlのウシ血清アルブミン分画V(Sigma,St Louis,MD)、各々0.5μMのプラ イマー、2.0mMのMgCl2 及び0.2mMのデオキシヌクレオチド三リン酸(dNTP)を用 いて行なわれる。全ての試薬混合および標本希釈は氷水浴中で行なわれ、熱ブロ ックが80℃に達した後管を熱サイクラーの中に置く。適切なプロトコル(適切な 鋳型DNA及びプライマーを用いて最適化されたもの)で熱サイクラー乾式浴を用 いて、ポリメラーゼ連鎖反応を行なう。PCR産物を適当なクローニングベクター 内にクローニングし、次に標準プロトコルを用いて配列決定する。 古代微生物の年代決定 上述のPCR増幅技術を用いて、古代微生物の年代を決定することができる。配 列決定に続いて、手作業か、又はアラインメントソフトウェアパッケージを用い て、配列を整列させ、配列を評価することができる。16S rRNA遺伝子のヌクレオ チド置換率rは、以前の報告書に基づき一年につき1つの位置あたり0.3∼0.4× −26− (27) 特表平9−508789 10- 9 置換であると仮定される(Ochman,H.& Wilson,A.C.J.Mol.Evol.26:74 -86(1987);Moran,N.A.,Munson,M.A.,Baumann,P.,& Ishikawa,H.Proc .R.Soc.Lond.B.253:167-171(1993))。2つの分類群の間の分岐時点(推定上 の古代微生物が分離されたコハクの年代)は、推定上の古代生物体とその最も近 い現存の血族の間の置換数を、分析されたヌクレオチドの数の2r倍で除するこ とによって決定される。2つの分類群の間の分岐時点は、分離株が得られたコハ クの年代とほぼ同じであるはずである。 古代微生物を用いた有用なタンパク質の産生 古代微生物は、近代微生物により産生されるタンパク質に関係す るものの、より高い温度、異なる大気条件でかつ/又は異なる基質に対して生物 活性を有するものである酵素及びその他のタンパク質を産生することができる。 一例としては、古代のハチがその食物を加工し保存するために用いた古代細菌は 、異なる環境条件下で生長していたし、工業的な用途をもち得る新しい代謝産物 の産生を導く生化学経路を示していたかもしれない。従って、古代微生物によっ て産生される祖先の酵素は、例えばより高い温度又は極限のpH条件での最適な性 能といったような独特の工業的に有用な生化学特性をもつのものと予想されてい る。 従って、本発明の1つの実施態様は、工業的、農業的、医療上及び診断上の用 途をもつタンパク質及び酵素を含む(ただしこれらに限られるわけではない)さ まざまな物質の産生のための古代微生物の培養に関する。このような物質は、近 代の微生物からの類似の物質の分離及び精製のために現在用いられているさまざ まな技術を用いて、培養から直接分離でき、好ましくは精製することができる。 代替的には、このようなタンパク質及び酵素は、生化学に合成されてもよいし、 或いは又組換え型DNA技術を用いて産生することもできる。 本発明の特定の実施態様においては、コハク中の2500万年∼4000万年前の昆虫 化石から分離された古代バシラス菌を、アミラーゼ、リパーゼ、ホスファターゼ 及びプロテアーゼを含む(ただしこれに限られるわけではない)さまざまなタン パク質及び酵素の産生のために使用することができる。さらに、本発明の古代バ −27− (28) 特表平9−508789 シラスのいくつかは、その近代の子孫に比べて、生活能力をもち増強された又は 著しく広い宿主範囲特性を備えた抗生物質を産生するのに使用することができる 。古代桿菌又はその他の微生物がこのようなタンパク質を産生する能力は、以下 で記述するような生化学的及び酵素によ る検定、又はその他の周知で広く用いられている検定、例えばGordon et al.,「 バシラス属USDA便覧427(1973);以下で記述するAPI ZYM酵素プロフィルテストを 用いて、測定することができる。バシラス属の細菌によって産生されるさまざま な酵素及びその他の産物、その同定のための検定、産生方法、及びさまざまな工 業的用途及びその他の用途についての完全な論述に関しては、その全体が本書に 引用により組み入れられているHarwood,C.R.,バイオテクノロジー便覧:バシ ラス、Plenum Press,New York(1989)を参照のこと。 本発明の一実施態様においては、「BCA5」と呼ばれる古代バシラスの1つが、 他の化合物の中でも、酸性ホスファターゼを産生した。表IIIを参照のこと。本 発明の別の実施態様では、古代Bacillussubtilis様の生物体は、アルカリ性ホス ファターゼ、ブチレートエステラーゼ、カプリレートエステラーゼ及び/又はβ −ガラクトシダーゼを産生し(例3参照)、古代Bacillus sphaericus様生物は 、カプリレートエステラーゼ及びキモトリプシン(例3参照)を産生し、古代Ba cillus cereus様生物体はアルカリ性ホスファターゼ、ブチレートエステラーゼ 、カプリレートエステラーゼ、ロイシンアミノペプチダーゼ、キモトリプシン、 酸性ホスファターゼ及びβ−グルコロニダーゼを産生し、古代Bacillus pumillu s様生物体は、カプリレートエステラーゼを産生した。 これらの古代酵素に対する現代の相対物は、プロテアーゼ及びアミラーゼ活性 を決定するための検定における使用、皮革もどし、ビール内でのタンパク質の曇 りの形成の防止、小麦粉のグルテン含有量の軽減、そして家庭用洗剤への内含、 を含めた広い商業的利用分野を有する。例えば、本書に引用により組み入れられ ているHarwood,C.R.,バイオテクノロジー便覧:バシラス、294以降、Plenum Press,New York(1989)を参照のこと。同様に、以上の論述も参照のこと。酵 −28− (29) 特表平9−508789 素を産生する古代微生物は、その現代の相対物と同一ではないが、近代微生物が 産生した酵素の商業的利用は、商業的環境における広範な一連の用途全体におけ る古代微生物によって産生された酵素の応用を示唆している。例えば、以下で論 述するBacillus subtillis様の微生物の1つの形態は、活性でかつ安定したアル カリ性プロテアーゼを産生することが示された。例3参照。 古代微生物からの抗生物質及び抗微生物物質 古代微生物は、今日の微生物界ではもはや存在しないか又はさほど優勢又は有 効でない、さまざまな古代生物体に対して有効な抗生物質を産生していたかもし れない。近代微生物の進化には、その祖先がその古代微生物近隣者の抗微生物作 用に耐えることができるようにしたいくつかの特性の喪失が付随していた確率が 高い。究極的には、微生物の中には、おそらくは単に長い年月にわたる古代病原 体の優勢度の低下をその理由として、その祖先が容易に耐えることのできたもの と同じ病原体に対して自らを防御できないほどにまで進化してしまったものもあ るかもしれない。この関係で、かつて古代では生存のために不可欠であった抗微 生物物質が、場合によって完全に不必要又は余分なものとなってしまった可能性 があり、従って、時間と進化が、もはやこれらの活性物質を産生できない近代微 生物を導き出したかもしれない。かくして、古代微生物からの抗生物質は、近代 微生物の感染、特に新しくかつ急速に進化する抗生物質耐性の病原菌株によって ひき起こされるものの治療のためにきわめて有効である可能性がある。従って、 本発明は、時代をさかのぼり、長い間に絶滅したさまざま生命形態の中で、今日 のものとは徹底的にかつ根本的に異なるものであった環境の中で何百万年も前に 生きていた微生物の抗微生物特性を検査する好機を与える医療科学 を提供している。 従って、本発明のもう1つの実施態様は、古代微生物を使用する抗生物質の産 生に関する。より特定的に言うと、出願人が分離し以下でより完全に記述するい くつかの古代バシラス及びアクチノミセス菌株は、ヒトの微生物病原体に対し強 力な抗微生物活性を示す。古代バシラス及びアクチノミセスの多くはさまざまな 古代菌株に対して抗微生物活性を示すが、一般にB.subtilis様及びB.lichenfo −29− (30) 特表平9−508789 rmis様として同定されている古代桿菌は、最強の抗微生物活性を産生すると思わ れた。「BCA1」と呼称されB.subtllis様菌として同定される古代バシラスの1 つは、Staphylococcus aureus及びStreptococcus pyogenesの増殖を完全に阻害 することができる。さらに、BCA1は、Salmonella pullorum,E.coli,Agrobacte r tumefaciens,Acinetobacter calcoacetlcus,Erwinia caratovora,Verticil lum dahliae及びPenicillum notatumの増殖を阻害することもできる。かくして 、BCA1は、さまざまな病原性微生物によりひき起こされる感染を処理する上で有 用であると期待されている非常に強力な抗生物質を産生する。全て、B.lichenf ormis様菌株として同定される古代バシラスBCA12,BCA13及びBCA15は、B.liche nformisから誘導された近代バシトラシンの分子祖先の産生のために有用であり 得る。 微生物の抗微生物活性をテストするための方法は、当該技術分野において長い 間知られてきており、本発明の古代微生物の場合にも同様に充分利用できるもの である。このような方法としては、例えば、日常的に用いられている交差画線方 法(Colome et al.,「微生物学における実験室演習」、West Educational Publ. ,St.Paul.MN(1986))、ならびにJ.Antibiotics 45(6):832-838内のChatterje e et al.により記述されている方法が含まれる。Colome et al及 びChatterJee et alの文献は両者共、本書に引用により組み入れられている。 抗微生物、抗炎症、CNS、心臓血管及び化学療法用の作用物質を含むその他の 有用な作用物質も同様に古代微生物を用いて産生することができる。Harvey ed. ,「天然産物からの薬物」;「医薬及び農薬」、Ellis Horwood Ltd.,England. (1993)。 古代微生物のためのその他の用途 本発明のさらにもう1つの実施態様では、昆虫の生物学的制御において古代微 生物を使用することができる。例えば、今日使用されている全ての微生物防除剤 のうちで最も成功したものの中に含まれるBacillus thuringiensis及びBacillus popillaeといったバシラスのいくつかの種が、現在、昆虫個体群を制御するの に用いられている。さらに、Bacillus sphaericusのいくつかの菌株が、蚊の幼 −30− (31) 特表平9−508789 虫に対して病原性をもつものと思われ、最近の現場試験では、B.sphaericusが化 学的殺虫剤の貴重な代替物となりうることが示唆されている。出願人が分離し本 書に記述している「BCA Ex2」及び「BCA16」と呼称された古代バシラス生物体の うちの2つは、さまざまな形態学的及び生化学的特性に基づいてBacillus sphae ricus様菌株として固定されてきた。これらの古代菌株は近代昆虫に対し病原性 をもち、従って生物学的殺虫剤として有用である可能性がある。 古代微生物、特に古代バシラス菌は、さまざまなタンパク質の産生におけるク ローニング及び発現宿主としても用いることができる。例えば、Bacillus subti lisは現在、組換え型DNA技術において広く用いられ、高レベルの組換え型産物を 容易に分泌することができるという利点をもつ。同様にして、古代微生物特に古 代バシラス菌は、診断検定における試薬としても使用することができる。例え ば、Bacillus subtilisは、ストレプトマイシンの検出のための診断検定におい て広く用いられる。 本発明の古代微生物は同様に工業的な利用分野も有している。例えば、ドミニ カ産コハクから分離されたサッカロミセス種は、CO2 (気体)及びエタノール( アルコール)の産生を伴ってグルコース、スクロース、ラクトース及びガラクト ースを発酵させ、任意の数の発酵プロセス中で使用することができる。Demain a nd Solomon,「工業用微生物の生物学」、Benjamin/Cummings Publishing Compa ny,1985)。古代微生物は、生物殺虫剤としても使用できる。Baclllus sphaeri cus及びBacillus thuringiensisは、現在生物殺虫剤として使用されており、こ のことはすなわち、その古代相対物が類似の有用性を有しうることを表わしてい る。Mitscherlich及びMartin,「環境内での微生物の生存」、Springer-Verlag(1 984)。 近代微生物代謝産物は、抗真菌化合物としても使用される。例えば、さまざま な真菌性疾患の治療において使用されるグリセオフルビンは、Penicillium種に より産生される。局所的抗真菌薬であるピロールニトリンは、異なるシュードモ ナス種によって産生され、同様に、幾分かの抗原生動物活性も示す。同じく局所 的抗真菌治療薬であるバリオチンは、真菌Paecilomyces variottiiの産物である −31− (32) 特表平9−508789 。皮ふの真菌性疾患の治療に用いられるシカニンは、ライ麦の草の寄生カビであ るHelminthosporium siccansから誘導される。上述の「バイオテクノロジー」第 4巻、248∼486を参照のこと。 同様にして、微生物によって産生される近代抗微生物化合物は、動物の飼料に 対する添加剤として、及び或る種の動物の疾病を治療するため、栽培植物を細菌 、真菌及びウイルスの感染、寄生虫病及び競合的草本から保護するため、糖尿病 、子宮アトニー、片頭痛、起立性循環器系障害、老化、高血圧、過プロラクチン 血症、先端巨 大症及びパーキンソン病、ガン、心臓血管疾患、高コレステロール血症、動脈硬 化症及び冠状動脈疾患といったような或る種の代謝障害を治療するためにも使用 される。上述のバイオテクノロジー、第4巻、510∼620を参照。微生物は、さま ざまなビタミン及び助酵素を産生するものとしても知られ(上述のバイオテクノ ロジー、第4巻、115∼159参照)、かつその外部環境から重金属及び放射性核種 を蓄積させる能力をもつ(上述のバイオテクノロジー、第6b巻、402∼403参照 )。同様にして、金属を浸出させる微生物の能力は、採鉱及び冶金業界において 用いられる。近代生物体の副産物のこれらの既知の用途は、その対応する古代先 祖の副産物も同様にして使用できるということの証拠を提供している。 以上で本発明について記述してきたが、制限的意味のない例として本発明を例 示するために、以下の例を提供する。 実施例 例1:古代バシラス生物の分離と特徴づけ 試料供給源。 化石組織を抽出するために、化石化された無針バチProblebeia dominicanaを含む2500万年∼4000万年前の構造的に無傷のドミニカ産コハク標 本を使用した。これらの試料を厳密に真正性認証し、コハク試料が開かれ化石組 織が抽出される全ての条件を殺菌して、近代微生物での汚染の可能性を排除した 。抽出プロセス内の全ての段階は、クラスIIaの層流フード内で行なった。さら に、抽出の結果もたらされる生活能力のある全ての微生物の古代状態を確認する ため、さまざまな対照及び偽似抽出を実施した。 −32− (33) 特表平9−508789 コハク殺菌及び粗砕手順。 2%のグルタルアルデヒド溶液内に一晩浸し無菌 脱イオン水(SDW)で3回洗い流すことによって、コハク供試体を表面殺菌した。 次に、洗浄済み試験片を5分間2%の漂白剤溶液中に浸し、SDW中で2回洗い流 した。コハクを70%のエタ ノール中に30分間浸し、火にあてて殺菌プロセスを完了した。次に無菌供試体を 以下のとおり液体窒素で粗砕した。無菌ガラスペトリ皿の中にコハク供試体を置 き、液体窒素で覆い、その後液体窒素を蒸発させた。コハク片全体の上に高温、 無菌の生理食塩水を滴下し、記述されたとおり化石化されたハチの組織を露呈す る平面に沿ってコハクを粗砕させた、Cano et al.,Med.Sci.Res.20:249-251( 1992)。 化石抽出。 無菌の27ゲージの針のついた無菌のツベルクリン注射器を用いて 、化石化されたハチの組織(胸郭及び腹部)の少量の標本をコハクから取り出し た。個々の標本を、TSBの入った個々の容器の中で懸濁させ、2週間35℃でイン キュベートして、存在するあらゆる細菌の増殖を可能にした。この期間の後、微 生物増殖を示す容器からのアリコートを固体培地トリプチカーゼダイズ寒天(TSA )上で継代培養させ、全く異なる細菌コロニーの視覚化及びさらなる分離を可能 にした。 分離され古代細菌の同定及び特徴づけ。 古代細菌分離株の同定は、基本的に Gordon et al.,「バシラス属」、農業省便覧427、ワシントンD.C.(1973)に記 述されている通りに形態学的及び生化学的特性を評価することによって行なわれ た。形態学的及び生化学的評価は全て層流フード内で実施した。19の酵素の検出 を可能にする半定量的システムを用いて古代バシラス培養の酵素活性を評価した (API ZYM Methodology,API Analytab Products)。BCA分離株の酵素活性プロ フィールを、米国標準培養収集機関(American Type Culture Collection)から 得た現存のB.subtilisのものと比較した。 全ての古代細菌保存培養は、安全で施錠された場所に保管され、全ての分離株 の試料は−70℃で安全な冷凍装置内で凍結され保管さ −33− (34) 特表平9−508789 れた。 抗微生物活性検定。 交差画線方法(Colome et al.,1986)を用いて、ヒト及 び植物に対する病原性をもつさまざまな細菌及び真菌に対する抗微生物活性につ いて、古代細菌をテストした。古代細菌をミュラー−ヒルトン寒天上に画線し、 24∼48時間35℃でインキュベートした。90度の角度で病原体を交差画線し、皿を 、細菌病原体について35℃で、又真菌病原体について室温でインキュベートした 。交差画線した皿を次に、交差画線から48時間後に増殖阻害についてそしてそれ 以降最高2週間にわたって監視した。 対照及び偽似抽出。分離された古代細菌が、分離プロセス中に拾われた近代の 汚染物質でなかったことをさらに確認するため、いくつかの対照実験及び偽似抽 出が行なわれた。全ての抽出手順において、フードの中に存在する細菌汚染物質 を検出するため歩哨(sentinel)ペトリ皿が存在していたが、どれも現われなか った。 殺菌、粗砕及び化石抽出手順で用いられた溶液の試料及びコハク供試体の内部 及び外部の試験片を、TSBを接種し微生物増殖についてチェックすることによっ て微生物汚染物質について検定した。いずれの溶液にもコハク試験片にも、細菌 又は真菌は全く検出されなかった。溶液の100マイクルリットルの試料及びコハ ク供試体内部の小片をケラックス溶液内に接種し、現存のバシラスに特異なプラ イマを用いてバシラスDNAを抽出及びPCR増幅することによって、バシラスDNAの 存在について検定を行なった。溶液中及び殺菌されたコハク供試体の表面上には 幾分かバシラスDNAが検出されたものの、ハコク内部から取られた試料内には全 く検出されなかった。 生きたBacillus subtilisで前処理されたコハク供試体を除染する上で殺菌プ ロセスが有効か否かを決定するためにもう1つの実験が行なわれた。簡単に言う と、化石封入体無しのコハク供試体を一 晩内生胞子含有Bacillus subtilis培養の中に沈めた。次に、上述の通り、そし て以下で例示されている通りに、コハク供試体を殺菌し、寛容な条件下でTSB中 でインキュベートさせて、汚染性のあらゆる細菌成長を可能にした。この対照実 −34− (35) 特表平9−508789 験を11回実施し、各々のケースにおいて、汚染物質は全く検出されなかった。か くして殺菌手順は、コハク供試体の表面上の近代の細菌汚染物質についての潜在 性を排除する上で有効である。 抽出手順中の試料の潜在的環境汚染を評価するために、擬似抽出を行なった。 化石封入体の無いコハク供試体を上述の通りに表面殺菌し、以下に記す抽出手順 をシミュレートした。さらに、溶液及びコハク外部及び内部の試料を、バシラス 内生胞子の存在についてテストした。2つの擬似抽出を行なったが、いずれも細 菌の増殖を全くもたらさなかった。 結果。 以下で記述する通り、殺菌した2500万年∼4000万年前のコハク供試体 から抽出された化石化したProblebeiのdominicanaの胸部及び腹部の組織の試料 の細菌成長培地に接種することによって、いくつかの古代細菌培養が生成された 。これらの分離株の形態学的及び生化学的特性が評価され、こうして古代細菌の 全てがバシラス又はバシラス様の生物体であることが示された。さらに、酵素活 性プロフィールが決定された。いくつかの近代バシラスについての形態、生化学 特性及び酵素活性プロフィールも同じく、並行して評価された。次に、これらの 回収された古代バシラスの体系的分類をより特定的に規定するため、近代及び古 代バシラスの特性が比較された。 形態学的及び生化学的評価の結果は、表I及びIIにそれぞれ示されている。古 代細菌の酵素プロフィールは表IIIに示されており、古代及び近代/子孫細菌の 比較が表IVに示されている。 −35− (36) −36− 特表平9−508789 (37) −37− 特表平9−508789 (38) −38− 特表平9−508789 (39) −39− 特表平9−508789 (40) −40− 特表平9−508789 (41) 特表平9−508789 古代細菌の酵素プロフィールは、近代バシラス種のものといくつかの点で相関 関係を有していたが、差異もいくつか観察された。例えば、古代細菌は、近代バ シラスに比べ、より多いアルカリ性ホスファターゼ及びロイシンアミノペプチタ ーゼ活性を生成すると思われた。同様に、近代B.subtilis及びB.cereusはα− −41− (42) 特表平9−508789 グルコシダーゼを産生するが、一方この活性は古代細菌には検出されなかった。 これとは対照的に、古代B.lichenformis様細菌はα−グルコシダーゼを産生し たが、近代B.lichenformisはこれを産生しなかった。さらに、古代B.lichenfo rmis様細菌は、近代B.lichenformis菌株に比べてより多くのβ−ガラクトシダ ーゼを産生するように思われる。 これらの観察事実に基づいて、出願人は、その見かけ上最も近い現存のバシラ ス子孫を基準にしてさまざまな古代細菌分離株を同定した。従って、以下の古代 細菌が以下に作表されたように同定される: 興味深いことに、B.subtilis,B.megaterium,B.cereus,B.lichenformis ,及びB.sphaericusが全て、或る種の現存のHymenopteraバチの消化管を阻害す ることが知られている。 古代バシラス菌株は同様に、上述の交差画線法を用いて抗微生物化合物の産生 に至るまで評価された。有意な量の阻害(すなわち、古代細菌画線から少なくと も2mm以内に増殖無し)は全て「部分阻害」として評点された。増殖が全く観察 されなかった場合にのみ、「完全阻害」の評点が割合てられた。検定は2度行な われ、テスト病原体の増殖を再現可能な形で阻害した古代細菌のみが、完全又は 部分阻害のいずれかの能力をもつものとして評点された。結果は表VIに示されて −42− (43) 特表平9−508789 いる。 例2:コハク種の除染 11のコハク片(封入体無し)を表面殺菌し、上述の手順に従って Bacillus su btilisのトリプチカーゼダイズ寒天(BBL,CocKeysville,MD)内生胞子形成培養 内で粗砕する。特定的に言うと、コハクと、(1)風袋測定された秤量ボート内 で秤量し、(2)真空内で少なくとも12時間35℃で2%の緩衝グルタルアルデヒ ド内に浸漬させ、(3)約250mlの無菌で2度精製された水(SDDW)とTSB上の培 養洗浄水の中で3回洗い流して殺菌手順の有効性を評価し、(4)37℃でさらに 6時間10%の漂白剤中に浸漬させ、(5)SDDW及びTSB上の培養洗浄水中で3回 洗い流して殺菌手順の有効性を評価し、(6)35℃で2時間70%のエタノール中 に浸漬させ、(7)アルコールを蒸発させるため炎に露呈した。 内生胞子の数は(直接計数により)、TSB1mlあたり約1.6×107 個であると見 積られた。殺菌手順を実施する前に培養中に一晩コハクを浸した。10mlのTSBが 入った管に表面殺菌したコハクを接種し、14日間35℃でインキュベートした。2 −43− (44) 特表平9−508789 週間のインキュベーション期間の後、11のTSB管のいずれの中にも、細菌成長は 全く検出されなかった。 例3:さまざまなコハク供給源からの古代微生物の抽出(粉砕方法) ドイツ産コハクからのBacillus subtillus様の微生物の分離。 土壌及び昆虫 デブリ(debris)を含むドイツ産コハク(bitterfelder)供試体(1500万年∼25 00万年前のもの)を、上述の殺菌及び粉砕抽出方法を用いて、表面殺菌及び組織 収獲した。30℃で4日間のインキュベーションの後、TSB寒天の表面上に、クリ ーム色の粘液状コロニーが見られた。分離株はAG-14-BF-2とコード化された。顕 微鏡検査により、胞子のうを膨潤させない、卵形の胞子を伴う直径1.0μm未満 のグラム陽性杆状体が明らかになった。バシラス種は 厳密に好気性で、カタラーゼ陽性で、VP反応陽性であり(VPブロスのpHは6未満 )、カゼイン、ゼラチン及びでんぷんを加水分解した。分離株は、推定的にBaci llus subtillis様として同定された。 API-ZYMプロフィールは以下のとおりであった:アルカリ性ホスファターゼ( +)、ブチレートエステラーゼ(+/−)、カプリレートエステラーゼ(+)、 ミリステートリパーゼ(−)、ロイシンアミノペプチダーゼ(−)、バリンアミ ノペプチダーゼ(−)、シスチンアミノペプチダーゼ(−)、トリプシン(−) 、キモトリプシン(−)、酸性ホスファターゼ(−)、ホスフォアミダーゼ(− )、α−ガラクトシダーゼ(−)、β−ガラクトシダーゼ(−)、β−グルコロ ニダーゼ(−)、α−グルコシダーゼ(−)、β−グルコシダーゼ(−)、N− アセチル−β−グルコースアミダーゼ(−)、α−マンノシダーゼ(−)、β− フコシダーゼ(−)。 ワシントン州Calmount産コハクからのアクチノミノス種様の微生物の分離。 土壌粒子を含むワシントン州Calmount産コハクの供試体(3800万年∼5400万年前 のもの)を、上述の滅菌及び粉砕抽出方法を用いて、表面殺菌し組織を収獲した 。30℃で8日間のインキュベーションの後、4%の酵母エキスを伴うツザペク寒 天平板の表面上に白っぽい粉末状のコロニーが現われた。分離株はAG-16-WA-3と コード化された。顕微鏡検査により、グラム陽性の分枝フィラメント及び短い杆 −44− (45) 特表平9−508789 状体が明らかにされた。分離株は任意に嫌気性であり、でんぷん−カゼイン寒天 中でよく成長し、7日間の増殖の後、直径1.0∼2.5mmの乾燥した粉末状のコロニ ーを産生した。分離株は、pHが8.1に調整された状態でSMA寒天平板内のカゼイン の消化により決定される通り、アルカリ性プロテアーゼを産生することが立証さ れた。分離株は、推定的に、アクチノミノス種として同定された。 中国産コハクからのBacillus sphaericus様微生物の分離。 土壌粒子を含む 中国産コハク供試体(4000万年∼5300万年前のもの)を、上述の殺菌及び粉砕抽 出方法を用いて、表面殺菌し組織を収獲した。30℃で5日間のインキュベーショ ンの後、TSA寒天の表面上には、白っぽいペースト状のコロニーが見られた。分 離株はAG-17-CH-1とコード化された。顕微鏡検査により、胞子のうを膨潤させな い丸い胞子を伴う直径1.0μm未満のグラム陽性杆状体が明らかになった。バシ ラス種は任意に嫌気性で、カタラーゼ陽性で、VP反応陰性(VPブロスのpH>7) であり、カゼイン、ゼラチン及びでんぷんを加水分解した。分離株は、推定的に 、Bacillus sphaericus様として同定された。 API-ZYMプロフィールは以下のとおりであった:アルカリ性ホスファターゼ( −)、ブチレートエステラーゼ(−)、カプリレートエステラーゼ(+)、ミリ ステートリパーゼ(−)、ロイシンアミノペプチダーゼ(−)、バリンアミノペ プチダーゼ(−)、シスチンアミノペプチダーゼ(−)、トリプシン(−)、キ モトリプシン(+)、酸性ホスファターゼ(−)、ホスフォアミダーゼ(−)、 α−ガラクトシダーゼ(−)、β−ガラクトシダーゼ(−)、β−グルコロニダ ーゼ(−)、α−グルコシダーゼ(−)、β−グルコシダーゼ(−)、N−アセ チル−β−グルコースアミダーゼ(−)、α−マンノシダーゼ(−)、β−フコ シダーゼ(−)。 ドミニカ産コハクからのArthrobacter aurescens様微生物の分離。 絶滅した マメ科植物Hymenaea proteraの葉及び花を含むドミニカ産コハク供試体(2500万 年∼4000万年前のもの)を、上述の殺菌及び粉砕抽出方法を用いて、表面殺菌し 組織を収獲した。30℃で7日間のインキュベーションの後、TSA寒天の表面上に 黄色のペースト状コロニーが見られた。分離株はAMG7とコード化された。顕微鏡 −45− (46) 特表平9−508789 検査により、杆状体−球菌サイクルに向かう明確な傾向をもつグラム陽性で運動 性の多形の杆状体が明らかになった。いかなる菌糸体も産生されず、厳密に好気 性であった。分離株は、黄色色素の存在及びでんぷんを加水分解する能力に基づ き、推定的にArthrobacter aurescens様であるものとして同定された。この生物 は、交差画線検定においてStaphyrococcus aureus及びEscherichia coliの成長 を阻害することが立証された。 レバノン産コハクからのBacillus subtilis様微生物の分離。 土壌粒子を含 むレバノン産コハク供試体(1億2000万年∼1億3500万年前のもの)を、上述の 殺菌及び粉砕抽出方法を用いて、表面殺菌し、組織を収獲した。30℃で5日間の インキュベーションの後、TSA寒天の表面上に、クリーム色の粘液状のコロニー が見られた。分離株はAG-14-LA-1とコード化された。顕微鏡検査は、胞子のうを 膨潤させない卵形の胞子を伴う直径1.0μm未満のグラム陽性杆状体を明らかに した。バシラス種は、厳密に好気性で、カタラーゼ陽性で、V−P(Voges-Pros kaner)反応陽性であり(V−PブロスのpH<6)であり、カゼイン、ゼラチン 及びでんぷんを加水分解した。分離株は、推定的にBacillus subtilis様である ものとして同定された。さらに、この生物体は、65℃で安定であると同時にpH 8 .1であるアルカリ性プロテアーゼを産生することが立証された。 API-ZYMプロフィールは以下の通りであった:アルカリ性ホスファターゼ(+ )、ブチレートエステラーゼ(+)、カプリレートエステラーゼ(+)、ミリス テートリパーゼ(−)、ロイシンアミノペプチダーゼ(−)、バリンアミノペプ チダーゼ(−)、シスチンアミノペプチダーゼ(−)、トリプシン(−)、キモ トリプシン(−)、酸性ホスファターゼ(−)、ホスフォアミダーゼ(−)、α −ガラクトシダーゼ(−)、β−ガラクトシダーゼ(+)、β−グ ルコロニダーゼ(−)、α−グルコシダーゼ(−)、β−グルコシダーゼ(−) 、N−アセチル−β−グルコースアミダーゼ(−)、α−マンノシダーゼ(−) 、β−フコシダーゼ(−)。 バルト諸国産コハクからのストレプトミセス種様の微生物の分離。 土壌粒子 及び植物デブリを含むバルト諸国産コハク供試体(4000万年前のもの)を、上述 −46− (47) 特表平9−508789 の殺菌及び粉砕抽出方法を用いて、表面殺菌し、組織を収獲した。30℃で7日間 のインキュベーションの後、4%の酵母エキスを伴うツザペク寒天平板の表面上 に、気菌糸体を伴う灰色がかった粉末状のコロニーが現われた。分離株はAG-11BA-11とコード化された。顕微鏡検査により、鎖状の胞子のう産生胞子及び短か い杆状体を伴う、グラム陽性の分枝フィラメントの存在が明らかになった。分離 株は、SMA寒天平板内でカゼインの消化により決定されるようにプロテアーゼを 産生することが立証された。分離株は、推定的にストレプトミセス種として同定 された。 ドミニカ産コハクからのサッカロミセス種様の分離。 土壌粒子及び昆虫の排 泄物を含むドミニカ産コハク供試体(4000万年前)を、上述の殺菌及び粉砕方法 を用いて表面殺菌し組織を収獲した。30℃で7日間のインキュベーションの後、 PDA寒天の表面上に白色のクリーム状コロニーが現われた。分離株はAG-11-DM-6 としてコード化された。顕微鏡検査により、直径5.0∼7.5μmの丸乃至は卵形の 出芽する酵母細胞が明らかになった。この生物体は、被包されておらず、Levine のEMB寒天(Difco)上で仮性菌糸を形成せず、グルコース、スクロース、ラクトー ス及びガラクトースを発酵させてCO2 (気体)及びエタノール(アルコール)を産 生した。分離株は、推定的にサッカロミセス種として同定された。 コロンビア産コーパルからのMicrococcus roseus様の微生物。 土壌粒子及び 昆虫排泄物を含むコロンビア産コーパル供試体(約2 00万年前のもの)を、上述の殺菌及び粉砕抽出方法を用いて表面殺菌し、組織を 収穫した。30℃で5日間のインキュベーションの後、TSA寒天の表面上に赤くペ ースト状のコロニーが現われた。分離株は、AG-11-CC-5とコード化された。顕微 鏡検査は、対で、4つ組及び不規則な束の形に配置されたグラム陽性球菌を明ら かにした。分離株は酸性グルコースを産生したか、グリセロールからではなく、 でんぷんを加水分解した。分離株は、推定的にMicrococcus roseus様であるもの として同定された。 バルト諸国産のコハクからのペニシリウム様生物体の分離。 土壌粒子を含む バルト諸国産(リトアニア産)コハクの供試体(4000万年前のもの)を上述の殺 −47− (48) 特表平9−508789 菌及び粉砕抽出方法を用いて表面殺菌し組織を収穫した。30℃で7日間のインキ ュベーションの後、PDA及びツザペク寒天の表面上には、黄色がかった膜様で逆 の拡散不能な色素を伴う粉末状で灰緑色のコロニーが現われた。分離株はAG-11BA-5とコード化された。ラクトフェノールコットンブルー(LPB)湿潤封入の顕微 鏡検査により、ペニシリア内の鉢状体の分生子産生細胞を伴うhylineの有隔性菌 糸が明らかになった。フィアロコニジアは、平滑な又は棘状の外壁を伴う鎖状を していた。分離株は推定的に、ペニシリウム株として同定された、交差画線検定 により、Staphylococcus aureus,Streptococcus pyogenes及びEscherichia col iに対する増殖阻害物質の存在が明らかになった。コロニーは、SMA上で増殖させ られたとき、カゼインを消化した。 アフリカ産コーパルからのアクチノミセス種様の微生物の分離。 土壌粒子及 び昆虫デブリを含むアフリカ産コーパルの供試体(約5000年前のもの)を、上述 の殺菌及び粉砕抽出方法を用いて表面殺菌し組織を収穫した。30℃で7日間のイ ンキュベーションの後、4%の酵母エキスを伴うツザペク寒天平板の表面上には 、白っぽい粉 末状のコロニーが現われた。分離株はAG-11-AC-14とコードされた。顕微鏡検査 により、グラム陽性の分枝フィラメント及び短かい杆状体の存在が明らかになっ た。分離株は、任意に嫌気性であり、でんぷん−カゼイン寒天内で充分に増殖し 、7日間の増殖後、直径1.5∼2.5mmの白色の乾燥した粉末状コロニーを産生し た。分離株は、SMA寒天平板内でカゼイン消化により決定されるようにプロテア ーゼを産生することが立証された。分離株は、推定的にアクチノミセス株として 同定された。 ビルマ産コハクからのCoryneform Thermophile様微生物の分離。 土壌粒子を 含むビルマコハク標本(3800万年∼5400万年前)を上述の通りに表面殺菌し、同 じく上述の粉砕抽出方法を用いて組織を収穫した。55℃で7日間のインキュベー ションの後、TSA皿の表面上に黄色がかったペースト状コロニーが現われた。分 離株はAG-12-BM-2とコード化された。顕微鏡検査は、鎖、柵及び杭垣パターンに 配置されたグラム陽性杆状体の存在を明らかにした。分離株は任意に嫌気性であ −48− (49) 特表平9−508789 り、でんぷん−カゼイン寒天内でよく成長した。分離株は、SMA寒天皿中でカゼ インの消化により決定されるようにタンパク質を産生することが立証された。分 離株は、試験的にcoryneform thermophileとして同定された。 コロンビア産コーパルからのバシラス種様の微生物の分離。 土壌粒子を含む コロンビア産コーパルの試料(約200万年前のもの)を上述の殺菌及び粉砕抽 出方法を用いて表面殺菌し組織を収穫した。55℃で5日間のインキュベーション の後TSA寒天の表面上にクリーム色の薄片状のコロニーが見られた。分離株はAC13-CC-33とコード化された。顕微鏡検査は、胞子のうを膨張させる卵形胞子を伴 う直径1.0μm以上のグラム陽性杆状体を明らかにした。バシラス株は、厳密に 好気性で、カタラーゼ陽性、V−P反応陽性(V−P ブロスのpH<6)であり、カゼイン、ゼチラン及びでんぷんを加水分解した。分 離株は、推定的にバシラス種として同定された。さらにこの生物は、65℃で安定 であると同時にpH 8.1で活性であるアルカリ性プロテアーゼを産生することが立 証された。 API-ZYMプロフィールは、以下の通りであった:アルカリ性ホスファターゼ( +)、ブチレートエステラーゼ(+)、カプリレートエステラーゼ(+)、ミリ ステートリパーゼ(−)、ロイシンアミノペプチダーゼ(−)、バリンアミノペ プチダーゼ(−)、シスチンアミノペプチダーゼ(−)、トリプシン(−)、キ モトリプシン(+)、酸性ホスファターゼ(−)、ホスフォアミダーゼ(−)、 α−ガラクトシダーゼ(−)、β−ガラクトシダーゼ(−)、β−グルコロニダ ーゼ(−)、α−グルコシダーゼ(−)、β−グルコシダーゼ(−)、N−アセ チル−β−グルコースアミダーゼ(−)、α−マンノシダーゼ(+)、β−フコ シダーゼ(−)。 日本産コハクからのタンパク質分解性ココバシラス(球杆菌)様微生物の分離 。 土壌粒子を含む日本産コハク試料(約8000万年前のもの)を上述の殺菌及び 粉砕抽出方法を用いて表面殺菌し組織を収穫した。30℃で5日間のインキュベー ションの後TSA寒天の表面上に白っぽい粘液状のコロニーが見られた。分離株は 、AG-19-JA-2とコード化された。顕微鏡検査により、胞子のうを膨潤させる卵形 −49− (50) 特表平9−508789 胞子を伴う直径1.0μm以上のグラム陽性球杆菌が明らかになった。バシラス種 は任意に好気性で、カタラーゼ陽性、オキシダーゼーであり、カゼイン及びゼラ チンを加水分解したがでんぷんは加水分解しなかった。分離株は、試験的にグラ ム陽性タンパク質分解性球杆菌として同定された。 ワシントン州産コハクからのストレプトミセス種様の微生物の分離。 土壌粒 子及び植物デブリを含むワシントン州産コハク試料( 3800∼5400万年前のもの)を上述のとおり表面殺菌し粉砕抽出方法を用いて組織 を収穫した。30℃で7日間のインキュベーションの後、5%の酵母エキスを伴う ツザペク寒天皿の表面上に気菌糸体を伴う灰色がかった粉末状のコロニーが現わ れた。培養は、ストレプトミセスに特徴的なカビ臭さを有していた。分離株はAG -15-WA-18とコード化された。顕微鏡検査により、鎖状の胞子を産生する胞子の うを伴うグラム陽性分枝フィラメント及び短かい杆状体の存在が明らかになった 。分離株は、Staphylococcus aureus,Bacillus subtilis及びEscherichia coli を含む、グラム陽性菌とグラム陰性菌の両方を阻害した抗微生物剤を産生するこ とが立証された。分離株は、推定的に、ストレプトミセス種様のものとして同定 された。 レバノン産コハクからのクラドスポリウム種様の微生物の分離。 土壌粒子を 含むレバノン産コハク供試体(1億2000万年∼1億3500万年前のもの)を上述の とおりに表面殺菌し、同じく上述の粉砕抽出方法を用いて組織を収穫した。25℃ で10日間のインキュベーションの後、ツザペク寒天の表面上に、暗く膜様の逆の 及び拡散不能の色素を伴う粉末状で濃黄緑色のコロニーが現われた。分離株はAG -13-LA-5とコード化された。ラクトフェノールコットンブルー(LPB)湿潤封入の 顕微鏡検査は、染色体分離体(dysjunctor)を伴う卵形で暗色の胚分生子を伴う 暗色の有隔性菌糸を明らかにした。遮へい細胞から分枝は明白であった。分離株 は、推定的にクラドスポリウム種様のものと同定された。コロニーは、SMA上で 成長させられた時点でキチンを消化し、グルコース及びスクロースを含めさまざ まな炭水化物を同化した。 例4:さまざまなコハク供給源からの古代微生物の抽出(粗砕方法) −50− (51) 特表平9−508789 ビルマ産コハクからのバシラス種様の微生物の分離。 昆虫及び 花粉粒を含むビルマ産コハク試料(3800万年∼5400万年前のもの)を、上述の殺 菌及び粗砕抽出方法を用いて表面殺菌し、組織を収穫した。30℃で6日間のイン キュベーションの後、TSA寒天の表面上には、クリーム色の粘液状のコロニーが 見られた。分離株はAG-15-BM-1とコード化された。顕微鏡検査により、胞子のう を膨潤させない卵形胞子を伴う直径1.0μm以上のグラム陽性杆状体が明らかに なった。バシラス種は任意に嫌気性で酸耐性をもち、カタラーゼ陽性、V−P反 応陽性(V−PブロスのpH<6)で、カゼイン、ゼラチン及びでんぷんを加水 分解し、唯一の炭素供給源としてクエン酸塩を利用した。分離株は、推定的にBa cillus cereus様のものとして同定された。 API-ZYMプロフィールは、以下の通りであった:アルカリ性ホスファターゼ( +)、ブチレートエステラーゼ(+)、カプリレートエステラーゼ(+)、ミリ ステートリパーゼ(−)、ロイシンアミノペプチダーゼ(+)、バリンアミノペ プチダーゼ(−)、シスチンアミノペプチダーゼ(−)、トリプシン(−)、キ モトリプシン(+)、酸性ホスファターゼ(+)、ホスフォアミダーゼ(−)、 α−ガラクトシダーゼ(−)、β−ガラクトシダーゼ(−)、β−グルコロニダ ーゼ(+)、α−グルコシダーゼ(−)、β−グルコシダーゼ(−)、N−アセ チル−β−グルコースアミダーゼ(−)、α−マンノシダーゼ(−)、β−フコ シダーゼ(−)。 カナダ産コハクからのバシラス種様の微生物の分離。 昆虫部分を含むカナダ 産コハク供試体(6800万年∼7500万年前のもの)を、上述の殺菌及び粗砕抽出方 法を用いて表面殺菌し組織を収集した。30℃で5日間のインキュベーションの後 、TSA寒天の表面上にクリーム色のペースト状コロニーが見られた。分離株は、A G-18-CA-2とコード化された。顕微鏡検査により、胞子のうを膨潤させる卵形胞 子を伴う直径1.0μm以上のグラム陽性杆状体が明らかになった。バシラス種は 任意に好気性で、カタラーゼ陽性、V−P反応陰性(V−PブロスのpH>7)で あり、カゼイン及びゼラチンを加水分解したがでんぷんは加水分解しなかった。 −51− (52) 特表平9−508789 分離株は、推定的に、バシラス種として同定された。 例5:さまざまなコハク供給源からの古代微生物の抽出(フラッシング方法) 。 ドミニカ産コハクからのシュードモナス種様の生物の分離。 水滴(土壌が中 に含まれているものもある)を含有するドミニカ産コハク供試体(2500万年∼40 00万年前のもの)を、前述の殺菌及びフラッシング抽出方法を用いて表面殺菌し 組織を収穫した。30℃で5日間のインキュベーション後、TSA寒天の表面上に黄 色がかった湿っぽいきらめくコロニーが見られた。分離株は、AG-10-DA-1とコー ド化された。顕微鏡検査により、グラム陽性の運動型杆状体が明らかになった。 分離株は好気性で、硝酸塩を還元し、カタラーゼ陽性で、オキシダーゼ陽性であ り、カゼイン及びゼラチンを加水分解したが、でんぷんは加水分解しなかった。 分離株は、推定的に、シュードモナス種として同定された。 例6:さまざまなコハク供給源からの古代微生物の抽出(せん孔方法) メキシコ産コハクからのBacillus pumilus種の生物の分離。 Pleheia silaceas の封入体を内含するメキシコ産コハク供試体(2000万年∼3500万年前のもの)を 、上述の殺菌及びせん孔抽出方法を用いて、表面殺菌し組織を収穫した。25℃で 4日間のインキュベーションの後、TSA寒天の表面上には、淡黄褐色でクリーム 状のコロニーが見られた。分離株はAG-12-MX-2とコード化された。顕微鏡検査は 、胞子のうを膨潤させない卵形胞子を伴う直径1.0μm未満のグラ ム陽性杆状体を明らかにした。バシラス種は、厳密に好気性で、カタラーゼ陽性 、V−P反応陽性(V−PブロスのpH<6)であり、カゼイン及びゼラチンを 加水分解したがでんぷんは加水分解しなかった。分離株は、推定的にBacillus p umilus様のものとして同定された。 API-ZYMプロフィールは、以下の通りであった:アルカリ性ホスファターゼ( −)、ブチレートエステラーゼ(−)、カプリレートエステラーゼ(+)、ミリ ステートリパーゼ(−)、ロイシンアミノペプチダーゼ(−)、バリンアミノペ プチダーゼ(−)、シスチンアミノペプチダーゼ(−)、トリプシン(−)、キ モトリプシン(−)、酸性ホスファターゼ(−)、ホスフォアミダーゼ(−)、 −52− (53) 特表平9−508789 α−ガラクトシダーゼ(−)、β−ガラクトシダーゼ(−)、β−グルコロニダ ーゼ(−)、α−グルコシダーゼ(−)、β−グルコシダーゼ(−)、N−アセ チル−β−グルコースアミダーゼ(−)、α−マンノシダーゼ(−)、β−フコ シダーゼ(−)。 例7:核酸配列分析による有効化 プライマー対BCF1(CGGGAGGCAGCAGTAGGGAAT)(配列番号1)及びBCR2(CTCCCC AGGCGGAGTGCTTAAT)(配列番号2)を用いて、鋳型として、推定上の古代分離株B CA16(Baclilus spharicusとして同定)から抽出したDNAを用いた。これらのプ ライマーは、B.circulans(GenBank受入れ番号X60613)の16S rRNA配列から得 られたものであり、この遺伝子の530塩基対領域を増幅する。Cano及びPoinar,C ano,R.J.& Poinar,H.N.,Biotechniques 15:8-11(1993)により記述されて いる通り、シリカゲル懸濁液を用いてDNAを抽出した。PCR増幅のためには、PCR による酵素的増幅のためのDNA供給源として、抽出されたDNAを5μl用いた。2 単位の低DNA Ampli-Taq DNAポリメラーゼ(Perkin-Elmer,Norwalk,CT)、2μg /ml のウシ血清アルブミン、各々0.5μMのBCA341F及びBCA871Rプライマー、2.0mMの MgCl2 及び0.2mMのデオキシヌクレオチド三リン酸、を合計体積50μlで用いて、 対称的PCR増幅を実施した。全ての試薬の混合及び標本希釈は、氷水浴中で実施 し、ヒートブロックが80℃に達した後熱サイクラー内に管を入れた。Temp-Troni c(商標)熱サイクラー乾式浴(Thermolyne,Dubuque,IA)を用いて、ポリメラーゼ 連鎖反応を行なった。最初のサイクル中、95℃で2分間DNA鋳型を変性させ、そ れに続いて1分間58℃でプライマーアニーリング段階を、そして72℃で1分間拡 張段階を行なった。その後の30サイクルは、1分間の94℃での変性段階、1分間 の58℃でのプライマーアニーリング段階及び1分間の72℃でのプライマー拡張段 階から成っていた。最後のサイクルは、1分間の変性段階、1分間のプライマー アニーリング段階及び10分間のプライマー拡張段階で構成されていた。各々の増 幅の産物を、トリス−酢酸塩−EDTA緩衝液中の1.2%のアガロースによって評価 した。メーカーの説明書に従って、TAクローニング(商標)キット(Invitrogen −53− (54) 特表平9−508789 ,San Diego,CA)を用いて、増幅産物をクローニングした。α−説明書を伴うS equenase II DNA配列決定キット(USB,Cleueland,OH)を用いて、クローニン グされたプラスミドDNAを配列決定した。SP6及びT7配列決定プライマーを両 方共用いて、各試料について3つのクローンを配列決定した。6%の配列決定ゲ ル(GEL-MIX6 Gibco BRL,Gaithersburg,MD)の中で、配列決定産物の電気泳動 を実施した。空気乾燥した配列決定ゲルのオートラジオグラフを、XARX線フィル ム(Kodak,Rochester,MN)を用いて作成した。配列をSparc 10ステーション( Sun Microystems,Mountain View,CA)内へ走査し、Bio Image DNA分析ソフト ウェア(Millipore,Bedford,MA)を用いて分析した。コンピュータ分析した配 列を確認するためオートラジオグ ラフを手作業で評価した。Genetic Data Environment(GDE2.1)テキスト編集プ ログラムを用いて、手作業で配列を整列させた。 BCA16の16S rDNAヌクレオチド配列、 ヌクレオチド置換率rは、推定上の古代B.sphaericusと現存のB.sphaericus の間の置換数を分析されたヌクレオチド部位の数で除することにより、計算され る。この数Kは、次に、2つの配列の間の分岐(拡散)時期をTとして2Tによ って除される。Li,Wen-Hsiung,& Dan Grauer、「分子進化の基礎」、Sinauer Associates,Inc.,Sunderland,Massachusetts(1991)。分岐時期(T)と同 様、コハク片の年数も、nを分析されたヌクレオチドの数(487)としrを一年、 一位置あたりの0.3−0.4×10- 9 としてK/n2Rを除することによって計算でき る。BCA16と現存のB.sphaericusの間のヌクレオチド置換数は、13であった。こ のKの値に基づくと、Tは33.3×106 年∼44.4×106 年である。ドミニカ産コハク 鉱山の年代は2500万年∼4000万年の間であることから、推定上の古代B.sphaeric usと現存のものの間のヌクレオチド置換数はコハクの年代と一貫しており、従っ て、結果はBCA16がドミニカ産コハク内のB.spahericusの古代分離株であると いう主張を有効化している。 例8:ヌクレオチド配列分析による分離株の古代性の有効化 推定上の古代分離株からDNAを抽出し、適切なプライマーを用いて16S rRNA遺 −54− (55) 特表平9−508789 伝子又はrbcL遺伝子(光合成生物について)の1セグメントを増幅させた。合計 体積50lで、1単位の低DNA Taqポリメラーゼ(Ampli Taq-LD DNAポリメラーゼ、 Perkin Elmer,Norwalk, CT)、2.5g/mlのウシ血清アルブミン分画V(Sigma ,St.Louis,MO)、各々0.55Mのプライマー、2.0mMのMgCl2 、及び0.2mMのデオ キシヌクレオチド三リン酸(dNTPs )を用いて、PCR増幅を行なう。全ての試薬混合 及び試料希釈は、氷水浴内で実施し、ヒート ブロックが80℃に達した後、熱サイクラ内に管を入れる。(適切な鋳型DNA及び プライマを用いて最適化された)適切なプロトコルで熱サイクラー乾式浴を用い て、ポリメラーゼ連鎖反応を行なう。PCR産物を適当なクローニングベクター内 へクローニングし、次に標準プロトコルを用いて配列決定する。配列を手作業で か又はアラインメントソフトウェアパッケージを用いて整列させ、配列を評価す る。16S rRNA遺伝子のためのヌクレオチド置換率rは、以前の報告書(Ochman an d Wilson J.Mol.Evol.26:74-86(1987);Moran et al.,Proc.R.Soc.Lo nd.B.253:167-171(1993))に基づいて一年一位置あたり0.3∼0.4×10- 9 の置換 ;そしてさまざまな植物分類群のrbcL遺伝子については1.9×10- 9 (Zurawski an d Clegg,Zurawski an Clegg,Annual Review Plant Physiology(植物生理学年 次評論)38:391-418(1987))であるものと仮定されている。2つの分類群の間 の分岐時期(推定上の古代微生物が分離されたコハクの年代)は、推定上の古代 生物体とその最も近い現存の血族の間の置換数を分析されたヌクレオチド数の2 r倍で除することによって決定される。2つの分類群の間の分岐時期は、分離株 を得たコハクの年代とほぼ同じであるはずである。 微生物の寄託 以下の古代細菌の培養が1994年1月28日(「BCA培養」)及び1994年11月1日 (「AG」培養)付けでNRRL,Peoria,Illinoisに寄託され、以下の受入れ番号が 割当てられた: −55− (56) 特表平9−508789 本発明は、本発明の一態様の単なる例示として意図されている本書で開示した 実施態様又は寄託された細菌培養に範囲が限定されるべきものではなく、機能的 に等価である全てのものが本発明の範囲内に入る。実際、本書に示され記述され ているものに加えて、上述の説明から当業者には本発明のさまざまな修正が明ら かとなるだろう。このような修正は添付のクレームの範囲内に入るものとされて いる。本書中に言及されている全ての特許、特許出願はここに引用により組み入 れられている。 −56− (57) −57− 特表平9−508789 (58) 【図1】 −58− 特表平9−508789 (59) 【国際調査報告】 −59− 特表平9−508789 (60) −60− 特表平9−508789 (61) −61− 特表平9−508789 (62) −62− 特表平9−508789 (63) −63− 特表平9−508789 (64) 特表平9−508789 ───────────────────────────────────────────────────── フロントページの続き (51)Int.Cl.6 C12N 識別記号 庁内整理番号 5/04 9282−4B 7/00 8827−4B 9/18 9/18 8827−4B 9/28 9/28 9152−4B 9/50 9/50 7823−4B 15/09 ZNA 1/02 9282−4B 1/68 9282−4B //(C12N 1/20 C12R C12R (C12N C12R C12N 1:24) 1/14 1:80) 1/16 1:85) (31)優先権主張番号 330,894 (32)優先日 1994年10月28日 (33)優先権主張国 米国(US) (81)指定国 EP(AT,BE,CH,DE, DK,ES,FR,GB,GR,IE,IT,LU,M C,NL,PT,SE),OA(BF,BJ,CF,CG ,CI,CM,GA,GN,ML,MR,NE,SN, TD,TG),AP(KE,MW,SD,SZ),AM, AU,BB,BG,BR,BY,CA,CN,CZ,E E,FI,GE,HU,JP,KE,KG,KR,KZ ,LK,LR,LT,LV,MD,MG,MN,MW, MX,NO,NZ,PL,RO,RU,SD,SI,S K,TJ,TT,UA,UZ,VN −64− 7/00 C12Q 1/02 1/68 Z C12N 5/00 F 15/00 ZNAA 7823−4B C12Q (C12N FI
© Copyright 2026 Paperzz