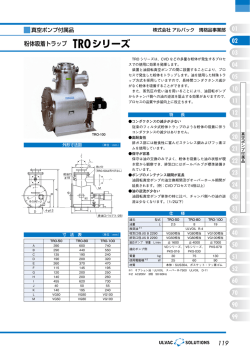
精子の発生機序とその形態 ( 資料提供 :佐藤和文先生) J ohnsen の分類( 1970) J3お) J J S I QY M ーH N I ー セ M[イ p I RYfャ [ 10 : 精細胞の層が正し い配列、 多数の精子を伴う j@ I セ セ W QI I 完全な精子形成能。 9: 多数の精子を認めるが、細胞配列に乱れ有り。 8: 精細管腔内に 精子が 5 から 10個程度。 7 : 精子を認めず。精子細胞が多数有り。 Cρ ュヂ忌治l d弘三仇か J 6: 精子を認めず。精子細胞が 5 から 10個程度。 aYQ セcI II 5: 精子及び精子細胞を認めず、精母細胞は多数有り 。 4: 精子及び精子細胞を認めず、精母細胞は数個程度 3 : 精細胞は精祖細胞のみ。 められる Ap. B I: 1It1l l 鋸 飽 . PL. L. Z. P I! 綱!i j 飯島弘 S a かも sd は綿子細胞である. Rb はi量検体 で Ad. 1 I j ' " 娘級殺 . 2 : 精細胞は欠如、 Sertol i cell のみ。 1 : 精細管内に細胞成分を認めず。 精子発生: Spermat ogenesi s 精祖細胞が精子に変わるまでの全過程 1 期 : Spermat ocyt egenesi s : 有糸分裂で幹細胞を残して増殖する数世代の精祖細胞。4 倍体 Ad"' " B 26 日 A d : dar k : 濃染する 卵円形の核に空胞あり 、核小体は核膜近くにある A p : pal e B: 淡染する楕円 円形の核、核小体は核膜に接する 円形 の核、核膜に沿って濃染する染色質 2 期 : PL" " ' I I : 精母細胞が 2 回の減数( 成熟) 分裂を経て染色体数は半数 32 日 II : 第 2 精 母 細 胞 ( 精 娘 細 胞 ) 第 2 減数分裂 Prel ept ot ene,L eptotene,Zygot ene,Pachyt ene : 第 1 精母細胞 : 第 l 減数分裂 16 日 3 期: Spermi ogenesi s : 精子形成 ・精子細胞が精子に変化す る。1 倍体 先体形成、核濃縮 ・形態変化 、 尾部( 鞭毛) 発生 Sa""' Sd→精子、 R b は遺残体 発育合計日数 74 日 nHU ハhU $ert oli Cel l s Spermat ogoni a I一 一 ー 一 切 一 一 '一 ー Pri mary $ p e m祖 tocyl es - 一一一一ー-- -1 Prel ept ot ene Zygot ene Pachyt ene M刷 。t時 Fi gures $ e∞ndary $ p e m祖 t ocy悼 s Mei oti c Fi gures 精細管内の細胞( 1 ) セルトリ細胞と生殖細胞の精子形成と減数分裂を示した 。 ( a ---i) はHEPES- TC199中 5 C 3 日 間分散培養した生きた細胞をノマルスキー光学で観察し、 (j ---q) はエポキシ樹脂切片 20 μ m標 本の精細管上皮。 生きたセル卜リ細胞 ( a) は有意に大型で、輪郭が号 │き伸ばされ辺縁がはっきりし 、核膜はぎざ ぎざして特有な核小体を吊るしている( 矢印) 。 細胞質はさまざまな大きさの頼粒を含んでいる 。 ( j ) の精細管固定セル卜リ細胞でも類似した核がある 。 細胞質と核の比、核の大きさと形は in vi tro (b- i ) 、固定された (k-q) の生殖細胞と類似している。 精祖細胞 in vi tro(b) 、固定された (k) は輪郭のはっきりした核膜と頼粒状クロマチンと 1 個ま たはより多くの核小体を持つ 。 第一精母細胞 in vi tro(c-e) 、固定された( 1 - n) はクロマチン粒 の増加と特徴的な核の膨化が見 られる 。 減数分裂直前の発育の間中、球状の核が残る。 前レプトテン期・レプトテン期精祖細胞 (c と1 ) は核膜が明瞭でなく、クロマチン粒は細かく核 の大きさは精組細胞より少し大きし '0 しばしばクロマチンが移動して少ない一部分( 矢印) が前レプトテン期 ・レプトテン期の核 に見られる 。 ジゴテン期精祖細胞 ( d , m) はも っと大きな核を持ち 、前より 大きくなったク ロマチ ン塊が平等 に散らされレプトテン期精祖細胞に比較して核膜は十分に明瞭である 。 パキテン期精祖細胞 (e , n) は全ての細胞中、最大の核を持ち 、十分に明瞭な核膜と 1 個または より多い大きな核小体がある 。 大中期板( 有糸分裂で染色体が赤道面に添って縦に分割する時期) の細胞中で減数分裂が進 行中 in vi tro( f ) で、固定標本 ( 0) も第一減数分裂の大中期板と似ている O 第二精母細胞細胞の in vi tro(g) と固定標本 (p) の両方とも前レプ卜テン期精祖細胞に見られた より細かいクロマチン粒と少し小さいサイズ、の核を持つ。 第二精母細胞細のクロマチン粒は核膜に接して不均等な分布である。 精娘細胞 in vi tro( h,i ) と固定標本 (q) に第二精母細胞細胞が分離し第二減数分裂を停止した結 果、クロマチン濃縮( 矢印) が見られる。 これらの細胞( 技術を加えられた精子細胞) は in vi troで観察された。 しかし本来の分化は知 られていない。 これらの細胞は定型的な円形精子細胞( 精細管内の細胞のに類似しているか、または小型 0 だろう 。 - 69 QM セN セN⦅M Sa 精細管内の細胞 Sa Msー・イ Sa S b1 ュ ウ S b2 _._.-.._.ー勾 叩.-. .-..- 日 ・ ... ._ - - 向日刊日" “ " “ “ ‘ 白川日目白“ . _ ---- 自白一一 i Sper mat ozoa S b2 Sc sd 1 Sd 1 S d2 (2) in vi tro(a-o) は生きた細胞をノマルスキー光学で見た精子細胞と精巣精子、 (p-z) はエポキシ 樹脂 20μm切片中の精細管上皮である 。 第二精母細胞( 精細管内の細胞 1 ) より明瞭な核膜を持つ Sa精子細胞の核が見られるがクロマ b) と精細管固定標本 ( p) の両方とも類似している。 チン粒の大きさは in vitroの生きた細胞の (a, 新しい精子細胞中にアクロゾーム小胞体や先体様構造は見られない。 アクロゾーム様小胞体( 矢印) は ( d, g) にはっきり見える 。 Sa精子細胞の核の上に平らにされたアクロゾーム様小胞体と in vi tro(e) 、固定 (8) にゴルジ体 輪郭( 矢印) が見られる 。 in vi tro(c) 、固定( r ) は細胞直径の拡大を終了し十分に発達した鞭毛が見える。 (f とt) のSbl 期の発育段階では核の球形喪失が始まりアクロゾームキャップ( 矢印) が生じる。 in vi tro( g ,i ) と固定 ( u, v) の両方とも Sb2精子細胞はマンチェッ卜( 精子頚の周囲にある一時 的な帯 ・矢印) が存在し続ける特徴がある。 アクロゾ ーム様キャップ( 開いた矢印) は鞭毛が付いている核の反対側に付く特徴がある。 精子細胞のマンチェッ卜は in vi tro(g ,i ) と精細管上皮 ( u, y) も同様に明瞭でない。 in vitro(j) と固定 ( w) の両方とも Sc精子細胞はアクロゾーム様の目立った隆起とマンチェット は失われる 。 Sc精子細胞は大変太い鞭毛を持ち、核付近の尾部に環( 矢印・中心体) と鞭毛中心部を被う ミ卜コンドリ アは、未だ持っていない。 in vi tro( k ,i ) と固定 (x) の両方とも Sdl 精 子 細 胞 は 短 い 核 の 輪 が 中 央 部 か ら 末 尾 へ 移 動 す る ( 矢印) 。 ( m, y) Sd2精子細胞は発育が続いてきた精子細胞の中で最も小さい頭部を持ち 、大きくな った 中心体( 矢印) と伸び、た尾部を持つ 。時々中心体の末尾に細胞残存物を持つ。 in vi tro( m) と固定 (y) の両方とも Sd2精子細胞は頭部も尾部も精巣精子の大きさに似ている。 in vi tro(n, o) と固定 (z) の両方とも時には尾部が頭部周囲を囲ったり、頭部も尾部も細胞膜に 包まれる事がある 。 Sc-Sd2精子細胞の発育中は、時々核の中に空胞を持つ (j-o, w-z) in vi tro(a-o) と精細管 (p-z) のヒト精子形成中、最初に目立つ事は核が小さくなる濃縮、そして アクロゾーム機小胞からアクロゾームが作られ、余分な細胞質を脱ぎ捨てる事が観察された。 70
© Copyright 2026 Paperzz