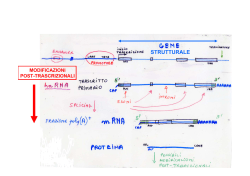
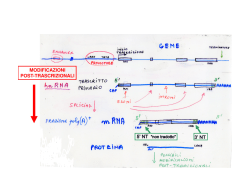
RNA polimerasi II trascrive i geni per la sintesi dei precursori degli mRNA, molti snRNA (U1, U2, U3, U4, U5, …), un gran numero di long non coding RNA mRNA di Eucarioti e Procarioti TFIIF e la RNA polimerasi … alla fine si lega la RNA polimerasi II seguita da altri fattori di trascrizione TFIIE e TFIIH Apr 2014 Il confronto tra gemelli umani identici, solo uno dei quali con sindrome di Down, rivela un generale appiattimento dei livelli di espressione genica nella persona affetta da trisomia La sindrome di Down si verifica quando un essere umano ha una copia extra del cromosoma 21. Poiché ogni cromosoma contiene molti geni regolatori dell’espressione genica, è stato presunto per decenni che la condizione patologica fosse causata principalmente da una sovrabbondanza di prodotti dei geni del cromosoma 21 in più. Recentemente è stato segnalato un caso di sindrome di Down che è associato con alterata espressione dei geni di tutti i cromosomi, non solo del cromosoma 21. Questa osservazione implica che l'espressione di un qualsiasi gene su un qualsiasi cromosoma possa contribuire alla sindrome di Down, e solleva la possibilità che una copia extra di qualsiasi cromosoma possa distruggere la regolazione generale dei geni. Appiattimento dell'espressione genica nella trisomia 21. In una persona con sindrome di Down i domini genomici che sono normalmente associati con bassi o alti livelli di espressione genica sono rispettivamente up-regolati o downregolati, rispetto a quanto avviene nel suo gemello identico che non ha la trisomia. Il risultato è un appiattimento dell’espressione genica nell’intero genoma. La stabilità del complesso di trascrizione, e quindi del numero di copie di RNA sintetizzate dalla RNA polimerasi II, dipende dai fattori di trascrizione (proteine) che compongono il complesso Una volta correttamente legata al complesso dei fattori di trascrizione sul promotore, Pol II apre la doppia elica del DNA ed inizia a legare i primi nucleotidi. Fatto questo, la subunità di inizio viene sostituita dal fattore di elongazione che stabilizza la forma attiva dell’enzima Enzima che lega il cap all’RNA in corso di sintesi cap Una volta raggiunte le sequenze di stop, il pre-RNA si stacca insieme al fattore di elongazione, ma la sintesi prosegue rallentando gradualmente … L’enzima che lega il cap si stacca alla fine della trascrizione Apr 2014 Durante il suo viaggio nel gene, l'RNA polimerasi II deve fare i conti con una serie di ostacoli alla sua progressione, tra cui i nucleosomi, le varie proteine che legano il DNA, i danni al DNA e le sequenze che sono intrinsecamente difficili da trascrivere. Non sorprende quindi che un gran numero di fattori di allungamento si siano evoluti per garantire che non si verifichi stallo o arresto della trascrizione. Quando capita che la RNA polimerasi non si riesce a riavviare, viene poliubiquitinata e degradata dal proteasoma. Questo processo è altamente regolato in modo tale da assicurare che siano degradate soltanto le molecole di RNA polimerasi II che non possono essere altrimenti recuperate. La trascrizione della RNA polimerasi II non è un processo continuo, ma frequentemente l’enzima va in stallo, talvolta anche a marcia indietro, portando talvolta all'arresto della trascrizione. Condizioni che portano a stallo possono essere in cis, ad esempio sequenze difficili da trascrivere, vincoli topologici, o la struttura stessa della cromatina, oppure in trans, ad esempio la diminuzione degli XTP necessari per la sintesi di RNA. La presenza di uno stallo temporaneo può portare l’enzima a fare marcia indietro o all’arresto trascrizionale, e questo può spesso essere risolto con l'assunzione di nuovi fattori di allungamento come TFIIS. La grande stabilità del complesso di elongazione della RNA polimerasi II permette all’enzima di rimanere attaccato al suo DNA stampo senza dissociarsi. Tale stabilità del complesso di allungamento è una benedizione in quanto assicura alta fedeltà nelle lunghe trascrizioni, ma anche una maledizione quando polimerasi ferma la sua progressione, poiché in quel caso il gene coinvolto risulta bloccato. Poliubiquitinazione e degradazione della RNA polimerasi II. L’enzima può risultare arrestato o totalmente bloccato a causa di un'ampia varietà di condizioni. Questo può innescare ubiquitinazione e degradazione da parte del proteasoma della RNA polimerasi II. Il complesso della RNA polimerasi II viene smontato e la subuntà Rbp1 viene poli-ubiquitinata e risucchiata nel proteasoma mentre le altre subunità vengono rilasciate. Appare persino sbagliato affermare “un gene codifica per un mRNA” che viene tradotto in una proteina. Nel lievito, sono stati trovati quasi 2 milioni di diversi trascritti di RNA per un genoma che contiene appena circa 6.000 geni codificanti per proteine. Un gene produce RNA condiffererenti sequenze di inizio e di terminazione. Eterogeneità intrinseca Molecular Biology: The ends justify the means - Nature. 2013 May 2;497(7447):48-9. doi: 10.1038/nature12098 - Pugh B.F. Appare persino sbagliato affermare “un gene codifica per un mRNA” che viene tradotto in una proteina. Nel lievito, sono stati trovati quasi 2 milioni di diversi trascritti di RNA per un genoma che contiene appena circa 6.000 geni codificanti per proteine. L’analisi genomica rivela che ogni gene produce una ricca complessità di RNA con differerenti sequenze di inizio e di terminazione, indicate come TIF (transcript isoforms). Questa pletora di isoforme deriva da un piccolo numero di RNA funzionalmente distinti per ciascun gene che sembrano originarsi per imprecisioni nell’inizio della trascrizione. Le popolazioni cellulari non possono mai essere omogenee, nonostante il fatto che derivano da una singola cellula di partenza. Tale eterogeneità intrinseca ha vaste implicazioni, dal fornire opportunità per l'adattamento nel corso dell'evoluzione e per spiegare in parte perché è difficile per uccidere tutte le cellule tumorali in un tumore. Molecular Biology: The ends justify the means - Nature. 2013 May 2;497(7447):48-9. doi: 10.1038/nature12098 - Pugh B.F. Il numero di geni codificanti proteine nei vertebrati non è radicalmente diverso dal numero di geni negli invertebrati (es. circa 20.000 geni nell’uomo contro 19.000 geni in Caenorhabditis elegans). La prevalenza dello splicing alternativo nei vertebrati è importante per loro superiore complessità Lo splicing alternativo è prevalente negli eucarioti multicellulari (nei mammiferi riguarda il 95% dei geni); quelli unicellulari hanno per lo più un solo introne o introni corti e presenti solo in alcuni geni Alternative splicing: a pivotal step between eukaryotic transcription and translation - Nat Rev Mol Cell Biol. 2013 Mar;14(3):153-65. doi: 10.1038/nrm3525 - Kornblihtt et al. - Erratum in Nat Rev Mol Cell Biol. 2013 Mar;14(3). doi:10.1038/nrm3560 Tipi diversi di alternative splicing Differenze nello splicing alternativo hanno un ruolo nell’evoluzione delle caratteristiche delle varie specie Anche se i geni di scimpanzé ed uomo sono identici per oltre il 99 %, il 6-8 % dei loro esoni alternativi subiscono uno splicing diverso in tessuti equivalenti La comparazione di trascrittomi di organi di specie di vertebrati che abbracciano ~ 350 milioni anni di evoluzione ha rivelato che i pattern di splicing alternativo si evolvono rapidamente e che, in un periodo evolutivo di appena 6 milioni di anni, i profili di splicing di organi equivalenti si discostano così tanto da risultare più dipendenti dall’identità della specie che dal tipo di organo Alternative splicing: a pivotal step between eukaryotic transcription and translation - Nat Rev Mol Cell Biol. 2013 Mar;14(3):153-65. doi: 10.1038/nrm3525 - Kornblihtt et al. - Erratum in Nat Rev Mol Cell Biol. 2013 Mar;14(3) - doi:10.1038/nrm3560 Lo splicing occupa una posizione centrale nel flusso delle informazioni genetiche degli eucarioti, dipende da vari fattori di regolazione e dal tipo di cromatina Lo splicing alternativo è almeno altrettanto importante per il differenziamento come la regolazione della trascrizione Nella comunicazione tra cellule, varie molecole segnale influenzano lo splicing alternativo (i) modificando l’attività di chinasi e fosfatasi che controllano trascrizione e traduzione, (ii) la localizzazione subcellulare dei regolatori dello splicing e (iii) l’acetilazione degli istoni che influenzano la struttura della cromatina Alternative splicing: a pivotal step between eukaryotic transcription and translation - Nat Rev Mol Cell Biol. 2013 Mar;14(3):153-65. doi: 10.1038/nrm3525 - Kornblihtt et al. - Erratum in Nat Rev Mol Cell Biol. 2013 Mar;14(3). doi:10.1038/nrm3560 L'organizzazione dei geni in esoni ed introni e l'evoluzione dello splicing alternativo hanno portato due vantaggi, (i) la produzione di proteine che uniscono, in una singola molecola, domini funzionali già testati dalla selezione naturale e (ii) la possibilità per un singolo gene di produrre due o più varianti di mRNA maturo, simili ma non identiche, ampliando notevolmente la capacità codificante dei genomi eucariotici Lo splicing è realizzato dallo spliceosoma assemblato nei siti di splicing di ogni introne. Sulla base della sua sequenza, un sito di splicing può essere forte o debole. La competizione tra siti forti e deboli vicini nel pre-mRNA è un fattore che porta a splicing alternativo. Alternative splicing: a pivotal step between eukaryotic transcription and translation - Nat Rev Mol Cell Biol. 2013 Mar;14(3):153-65. doi: 10.1038/nrm3525 - Kornblihtt et al. - Erratum in Nat Rev Mol Cell Biol. 2013 Mar;14(3) - doi:10.1038/nrm3560 Gen 2014 I geni eucarioti sono discontinui costituiti da introni ed esoni. Il processo di splicing dipende da un insieme di piccoli RNA nucleari (snRNA), che si assemblano con proteine in un complesso noto come spliceosoma. Studio del ciclo di vita degli snRNA spliceosomali dalla loro trascrizione, esportazione dal nucleo e reimportazione nel nucleo per il loro assemblaggio nello spliceosoma. Il processo di assemblaggio può anche influenzare la regolazione dello splicing alternativo ed ha implicazioni per le malattie umane. Le RiboNucleoProteine sono complessi formati da RNA e proteine Struttura della RNP U1 attggaaaccgaaacccgttggtcacctctgcaatagccctccctccctcacttctacaattttgtgaca gtggtcttgttttctgcattctctgcttcacgtgcttgttttgttggagcgcgtttgcatgctgctttaa attctgaaatattaaaaaaatttcgaagtttttcagcacatgggatgggagttttgaatttcaatttttt aaaaacatttttctgtgattagtgccgtcgtggcacggctgttagccgcctatccggtttattcgatact ttGTGAGTTTTTTGTAACTTTATGGTCGTCGAAATGGGAAAACTTGGCCACCAATATAAGTTTGGAAAAC AATTTCCTAAAAATAAAATAATTGAACTTTTCCGATGAATAAAAAAATCGATCAGATATTCTGGAAAAAA AATCGATAAATTAATCGATTTTCTTGGAAAATACATCGAAAAATTGAGAAAAATAGAAAAATGAATGTTT TTCGATTACCGATTTATTGATTTTTCGTGAAAACTGAGTTCAGATAATTTTAAAAGCAATGTTTTTCATT TTTCAAATCAGAATCACTATAGTTTTGAAAAATCAATAATTAATTTATTGATTTTTCAATATAATTTTTT GGAAAAAATAGAAAAATCCCTTTCTAAAAGTTTTAAATTTCCAAGAAAAATTCATTTTCAAAATCACCAA CGCGCTCTATAGAGTAGTCGATGAAAATCTCCGTTAAGGGTGCATGGGCAAAACGCGCTCGAACGACAAT TGTTATTGTATGTTTGGTCTTGCAACGAAAAGTTTGAAAAATTGAAAAAAAGTTGTGTCTGATACATTTT TTTTTGGCATTTTCTGCTATTTTACACCAGAAAAAATTTAATAAACATAAAAAATCGAAATTTTTCAAGT TGGACAATTTTCAGtgagcatcttatccatcctagttctcagttcaggacttgtgcacattcgtttagag ccagatattcgcaaagccttttcaccggatgattcagatgctggataGTAAGTGACTACTGACCTTGAAG CCTCCTTCCTCCACCAGTCAGAAATAACACGTTTTTTCGCAATGTTTTTCTTTTTCTAATTCGATTTCCC TTTCTCCCTTTCTTATTGTGATTTGGTCAATGTTTGGTTGACTGGGAAGAAAATTGAATTTTTTTGGAAT TCCACTTGAAGTTAAAAAACCCAAAATAAATATTTGATCAAAAATAAATAAGAAAAAAAAGAAAACTTTA AAGCAAATGAAAATTTCGTTCGTAACTATTTTGTTAATTTTTTTAAAACTCCTATTTTAAATATATGCTT TTTGCGGAAATTTCTATAAATTTTTTTACATTTTTCAGtgaaacccgtgtctggctggaatactacggac tcgacatctatccggaacgagcattctgtatttttaccgccaagcgcgaaaattccagtattctccagga aggcgcactggcagacGTAAGTTGATTCTCCGTCACGCCCACTTTTCTGGCGGGAATTTAAAAAATTTCA Gatttatactgtggacaatcgactatcggcggcagttggctaccaagatggggatggacgaaaaaattgc gatccactctgcgacttgaacagcccctttcacttgttagcgGTAGGTGGTGGTCTAGGGTGTCATTTTT CGATTTTTTCAATTATTCGATGTTTTTAGTGAAAATCGAAAAATCTAAAAATTGAAAATCGAAAAATGAA AGAAACATTGTTTTTTGGGGACCAAACATCTTAATGAATTTAACAACAGGGAAAACTGAACAGAAACCTG GACGGTCTTATCCCATTTATCTATATTCTTAAAATGAATGATGGAGAAAAAAGTTAAAATAAAAACATTA TCAGCTTTTTGTAAGTTTTTCTCAAAAATTGTTCGATTTTTCGATTTTCTAAAAAGTCGAAAAACCGAAA CCCTTGGTGGTGGTGGTGGTGGACTAGAAAACTCTTCAACGACCACATGGCAATTTTCAGaatttgacgc ggagaaacaatggtaccacaagtgtattcacctatccggatatgccatatagcggactggatattttcct gggacttcacttgagtaatgcggattttggtaagattttttttgaaatgttaaatgaaaagttgaaaaat agtttttatgatttagccactttccagttaaaatttcatttttttaactataaaaagttctggaaaaatg aatttctAGgccgccgatcctaaaAGTgcaccatttcgcAGaAGTacGTacAGTttcccatctatccctA GTgGTcttGTtttctgcattctctgcttcacGTgcttGTtttGTtggAGcgcGTttgcatgctgctttaa attctgaaatattaaaaaaatttcgaAGTttttcAGcacatgggatgggAGTtttgaatttcaatttttt aaaaacatttttctGTgattAGTgccGTcGTggcacggctGTtAGccgcctatccgGTttattcgatact ttGTGAGTTTTTTGTAACTTTATGGTCGTCGAAATGGGAAAACTTGGCCACCAATATAAGTTTGGAAAAC AATTTCCTAAAAATAAAATAATTGAACTTTTCCGATGAATAAAAAAATCGATCAGATATTCTGGAAAAAA AATCGATAAATTAATCGATTTTCTTGGAAAATACATCGAAAAATTGAGAAAAATAGAAAAATGAATGTTT TTCGATTACCGATTTATTGATTTTTCGTGAAAACTGAGTTCAGATAATTTTAAAAGCAATGTTTTTCATT TTTCAAATCAGAATCACTATAGTTTTGAAAAATCAATAATTAATTTATTGATTTTTCAATATAATTTTTT GGAAAAAATAGAAAAATCCCTTTCTAAAAGTTTTAAATTTCCAAGAAAAATTCATTTTCAAAATCACCAA CGCGCTCTATAGAGTAGTCGATGAAAATCTCCGTTAAGGGTGCATGGGCAAAACGCGCTCGAACGACAAT TGTTATTGTATGTTTGGTCTTGCAACGAAAAGTTTGAAAAATTGAAAAAAAGTTGTGTCTGATACATTTT TTTTTGGCATTTTCTGCTATTTTACACCAGAAAAAATTTAATAAACATAAAAAATCGAAATTTTTCAAGT TGGACAATTTTCAGtgAGcatcttatccatcctAGTtctcAGTtcAGgacttGTgcacattcGTttAGAG ccAGatattcgcaaAGccttttcaccggatgattcAGatgctggatAGTAAGTGACTACTGACCTTGAAG CCTCCTTCCTCCACCAGTCAGAAATAACACGTTTTTTCGCAATGTTTTTCTTTTTCTAATTCGATTTCCC TTTCTCCCTTTCTTATTGTGATTTGGTCAATGTTTGGTTGACTGGGAAGAAAATTGAATTTTTTTGGAAT TCCACTTGAAGTTAAAAAACCCAAAATAAATATTTGATCAAAAATAAATAAGAAAAAAAAGAAAACTTTA AAGCAAATGAAAATTTCGTTCGTAACTATTTTGTTAATTTTTTTAAAACTCCTATTTTAAATATATGCTT TTTGCGGAAATTTCTATAAATTTTTTTACATTTTTCAGTgaaacccGTGTctggctggaatactacggac tcgacatctatccggaacgAGcattctGTatttttaccgccaAGcgcgaaaattccAGTattctccAGga AGgcgcactggcAGacGTAAGTTGATTCTCCGTCACGCCCACTTTTCTGGCGGGAATTTAAAAAATTTCA GatttatactGTggacaatcgactatcggcggcAGTtggctaccaAGatggggatggacgaaaaaattgc gatccactctgcgacttgaacAGcccctttcacttGTtAGcgGTAGGTGGTGGTCTAGGGTGTCATTTTT CGATTTTTTCAATTATTCGATGTTTTTAGTGAAAATCGAAAAATCTAAAAATTGAAAATCGAAAAATGAA AGAAACATTGTTTTTTGGGGACCAAACATCTTAATGAATTTAACAACAGGGAAAACTGAACAGAAACCTG GACGGTCTTATCCCATTTATCTATATTCTTAAAATGAATGATGGAGAAAAAAGTTAAAATAAAAACATTA TCAGCTTTTTGTAAGTTTTTCTCAAAAATTGTTCGATTTTTCGATTTTCTAAAAAGTCGAAAAACCGAAA CCCTTGGTGGTGGTGGTGGTGGACTAGAAAACTCTTCAACGACCACATGGCAATTTTCAGaatttgacgc ggAGaaacaatgGTaccacaAGTGTattcacctatccggatatgccatatAGcggactggatattttcct gggacttcacttgAGTaatgcggattttgGTaAGattttttttgaaatGTtaaatgaaaAGTtgaaaaat AGTttttatgatttAGccactttccAGTtaaaatttcatttttttaactataaaaAGTtctggaaaaatG Duplice effetto dell’allungamento trascrizionale sullo splicing alternativo: (a) Durante una fase di allungamento veloce, la RNA pol II favorisce il reclutamento dello spliceosoma al sito forte di splicing al 3’ di un introne a valle di un sito debole di plicing al 3’ dell'introne a monte, il risultato è lo skipping dell’esone. Invece, durante un allungamento lento (destra) viene favorito il reclutamento dello spliceosoma sull’introne a monte, il risultato è lo splicing con inclusione dell'esone; Alternative splicing: a pivotal step between eukaryotic transcription and translation - Nat Rev Mol Cell Biol. 2013 Mar;14(3):153-65. doi: 10.1038/nrm3525 - Kornblihtt et al. - Erratum in Nat Rev Mol Cell Biol. 2013 Mar;14(3) - doi:10.1038/nrm3560 Duplice effetto dell’allungamento trascrizionale sullo splicing alternativo: (b) quando entrambi i siti di splicing al 3’ sono ugualmente forti e l'introne a monte ha un sito di legame (verde) per un fattore di splicing che inibisce l’inclusione dell'esone (fattore negativo NF), l’allungamento veloce della Pol II (a sinistra) favorisce il reclutamento dello spliceosoma su ambedue gli introni, garantendo l’inclusione dell'esone. Al contrario, l’allungamento lento (destra) dà il tempo al fattore di splicing negativo di legarsi prima dello spliceosoma, che quindi si legherà al sito di splicing al 5‘ dell'introne a valle con risultante skipping dell’esone. Alternative splicing: a pivotal step between eukaryotic transcription and translation - Nat Rev Mol Cell Biol. 2013 Mar;14(3):153-65. doi: 10.1038/nrm3525 - Kornblihtt et al. - Erratum in Nat Rev Mol Cell Biol. 2013 Mar;14(3) - doi:10.1038/nrm3560 Lo splicing alternativo è governato da sequenze cis regolatorie nel pre-mRNA, ESE (exonic splicing enhancers) ESS (exonic splicing silencers) ISE (intronic splicing enhancers) ISS (intronic splicing silencers) e da due principali famiglie di proteine regolatrici dello splicing alternativo, •SRSF (Ser/Arg-rich splicing factors) •hnRNP (heterogeneous nuclear ribonucleoproteins) Alternative splicing: a pivotal step between eukaryotic transcription and translation - Nat Rev Mol Cell Biol. 2013 Mar;14(3):153-65. doi: 10.1038/nrm3525 - Kornblihtt et al. - Erratum in Nat Rev Mol Cell Biol. 2013 Mar;14(3). doi:10.1038/nrm3560 L’efficienza del processo di splicing non è mai del 100% per tutti gli esoni Anche se le sostituzioni nella terza base di codoni sinonimi non cambia l’amminoacido codificato, talvolta si produce inaspettatamente un aumento dell’efficienza dello splicing e WT AAA GAT GCT GAT TTG TAT TTA TTA GAC TCT CCT TTT GGA A1 A2 A3 A4 A5 A6 A7 A8 A9 A10 A11 A12 AAG AAG AAA AAG AAG AAG AAA AAA AAA AAG AAA AAA GAC GAC GAC GAC GAC GAC GAC GAC GAT GAC GAC GAC GCA GCG GCT GCG GCC GCC GCG GCA GCC GCT GCA GCA GAT GAC GAT GAC GAC GAC GAT GAT GAC GAC GAC GAC TTG TTG TTG TTG TTG TTA TTG TTA TTA TTG TTG TTA TAC TAT TAC TAC TAT TAC TAT TAT TAT TAT TAT TAC TTA TTA TTA TTA TTG TTG TTA TTG TTG TTA TTG TTA TTA TTA TTG TTG TTG TTG TTG TTG TTG TTG TTG TTG GAT GAT GAT GAC GAC GAC GAT GAC GAT GAC GAC GAC TCA TCG TCA TCC TCT TCG TCA TCC TCA TCC TCA TCA CCC CCG CCG CCC CCG CCT CCT CCG CCC CCA CCG CCG TTC TTC TTC TTC TTC TTT TTC TTT TTC TTT TTC TTT GGA GGC GGA GGT GGT GGC GGC GGA GGC GGG GGT GGT A13 A14 A15 A16 A17 AAA AAA AAG AAA AAA GAC GAT GAT GAT GAC GCA GCG GCT GCG GCA GAT GAC GAT GAT GAT TTG TTG TTA TTG TTA TAT TAT TAT TAT TAC TTA TTA TTA TTG TTG TTG TTG TTA TTA TTG GAT GAT GAC GAC GAT TCT TCG TCT TCA TCC CCG CCA CCG CCG CCC TTT TTT TTC TTT TTC GGG GGT GGT GGC GGC A18 AAA GAT GCA GAT TTG TAC TTG TTA GAC TCG CCC TTT GGC A19 AAG GAC GCA GAT TTG TAT TTG TTA GAC TCC CCA TTC GGG A20 AAG GAC GCT GAC TTA TAC TTG TTA GAT TCC CCT TTC GGT A21 AAG GAT GCA GAT TTA TAT TTA TTA GAC TCC CCT TTT GGT non una diminuzione come ci si sarebbe invece aspettato L’esempio sopra riportato riguarda l’esone 12 del gene CFTR che codifica per una proteina di membrana (canale dello ione Cl-) responsabile della fibrosi cistica Assemblaggio co-trascrizionale delle snRNP e loro rimodellamento dalla trascrizione alla traduzione e alla degradazione Lo splicing che forma mRNA avviene in modo co-trascrizionale con l’assemblaggio delle particelle snRNP La presenza del cap facilita lo splicing, la formazione della estremità 3’ e quindi l’esportazione di mRNA attraverso i pori nucleari Nel citoplasma, il CBC (cap-binding complex) viene scambiato con il fattore di inizio traduzione 4E (EIF4E) e con il ribosoma CPSF6 (cleavage and polyadenylation specificity factor 6), EJC (exon junction complex); hnRNPC (heterogeneous nuclear ribonucleoprotein C), NXF1 (nuclear export factor 1), PABPC1 (cytoplasmic poly(A) binding protein 1), PABPN1 (nuclear poly(A)-binding protein 1), TREX1(transcription export complex 1), XRN1 (5′–3′ exoribonuclease 1). How cells get the message: dynamic assembly and function of mRNA–protein complexes - Nat Rev Genet. 2013 Apr;14(4):275-87. doi: 10.1038/nrg3434 - Müller-McNicoll M, Neugebauer KM. Figura – Meccanismi di esportazione di mRNP dal nucleo al citosol. Esempi dei diversi tipi di esport di mRNP (particelle ribonucleoproteiche messaggero) riguardanti quattro diverse classi di trascritti della RNA polimerasi II (Pol II) le loro diverse modalità di uscita dal nucleo: - mRNP mature dopo aver completato lo splicing, sono normalmente esportate dal nucleo tramite NXF1 (nuclear RNA export factor 1) che viene reclutato da complessi di proteine adattatrici, quali TREX1 (transcription export complex 1) ed altre (UAP56, CPSF6, UIF, SR, …) con lunghi RNA pieni di introni che stanno subendo splicing; - mRNP sono troppo grandi per passare attraverso i pori nucleari, possono uscire attraverso la gemmazione di vescicole dall’involucro nucleare. - mRNA privi di introni che codificano per istoni vengono esportati da SLBP (stem-loop-binding protein) che viene reclutata dal CBC (cap-binding complex) e da NELF (negative elongation factor); - TREX2 può esportare mRNP maturi attraverso i pori nucleari interagendo con NXF1; - l’esportazione di lunghi trascritti privi di introni avviene attraverso una pathway alternativa di esportazione (ALREX = alternative RNA export) ed altre proteine che reclutano NXF1 anche in assenza di splicing; - piccoli trascritti senza introni vengono legati da PHAX e sono trasportati fuori dal nucleo da altre proteine (CRM1 e la GTPasi della famiglia RAN). How cells get the message: dynamic assembly and function of mRNA–protein complexes -Nat Rev Genet. 2013 Apr;14(4):275-87. doi: 10.1038/nrg3434 - Müller-McNicoll Neugebauer Packaging co-trascrizionale dell’RNA, maturazione ed esportazione di mRNA: (a) l’assemblaggio della snRNP sul nascente pre-mRNA impedisce la formazione di loop deleteri; (b) splicing costitutivo: l’assemblaggio delle snRNP in corso di splicing mantiene gli esoni consecutivi in stretta vicinanza e contribuisce a garantire il corretto ordine degli esoni durante lo splicing; (c) splicing alternativo e poliadenilazione alternativa (APA): l’assemblaggio sul pre-mRNA potrebbe nascondere forti siti di splicing o esoni alternativi per promuovere l’exon skipping, oppure potrebbe esporre deboli siti di splicing per facilitare l'inclusione dell'esone durante lo splicing alternativo. Ne potrebbe derivare l’occultamento di segnali deboli di poliadenilazione (PAS) presenti negli introni oppure l’esposizione di PAS per una poliadenilazione (APA) alternativa. How cells get the message: dynamic assembly and function of mRNA–protein complexes -Nat Rev Genet. 2013 Apr;14(4):275-87. doi: 10.1038/nrg3434 - Müller-McNicoll Neugebauer Negli eucarioti lo splicing del pre-mRNA è co-trascrizionale, spazialmente e temporalmente separato dalla traduzione dell'mRNA che avviene fuori dal nucleo L’assemblaggio del pre-mRNA nel complesso di splicing è importante per l’esportazione di mRNA. Ci sono due classi distinte di RNP competenti per l’export: lunghi mRNA, che vengono esportati da NXF1 (nuclear export factor 1) e piccoli RNA che vengono esportati da CRM1 (chromosomal region maintenance protein 1) attraverso una pathway dipendente da RAN·GTP. Il complesso che lega il cap (CBC) è il primo evento di elaborazione del mRNA nascente che emerge dalla Pol II. CBC interagisce con diversi TREX1 (transcription export complex 1). How cells get the message: dynamic assembly and function of mRNA–protein complexes -Nat Rev Genet. 2013 Apr;14(4):275-87. doi: 10.1038/nrg3434 - Müller-McNicoll Neugebauer Forte legame tra splicing alternativo e malattia, mutazioni nelle sequenze regolatrici che influenzano lo splicing alternativo sono cause diffuse di malattie ereditarie umane e di cancro Mutazioni nelle sequenze codificanti possono essere male interpretate considerando solo gli effetti putativi che avrebbero nella sequenza della proteina codificata Ad esempio, la variazione di un singolo nucleotide che non cambia la sequenza amminoacidica di una proteina (mutazione silente) può facilmente essere scambiata per un polimorfismo neutrale quando in realtà potrebbe causare l’interruzione di un ESE cruciale con conseguente origine di una patologia Le sequenze degli esoni che codificano proteine sono sottoposte a duplice pressione selettiva, (1) conservazione della sequenza amminoacidica codificata e (2) degli elementi esonici che regolano lo splicing alternativo del pre-mRNA Alternative splicing: a pivotal step between eukaryotic transcription and translation - Nat Rev Mol Cell Biol. 2013 Mar;14(3):153-65. doi: 10.1038/nrm3525 - Kornblihtt et al. - Erratum in Nat Rev Mol Cell Biol. 2013 Mar;14(3) - doi:10.1038/nrm3560 Le più note patologie causate da mutazioni cis -acting sono le taupatie come la demenza frontotemporale con parkinsonismo, la distrofia miotonica e l'atrofia muscolare spinale (SMA) Anche mutazioni in geni che codificano per fattori trans-acting che regolano lo splicing alternativo possono causare patologie. A differenza delle mutazioni cis-acting che interessano solo il gene compromesso, questo secondo tipo di mutazione può influenzare grandi insiemi di geni I disturbi più studiati che derivano da questo tipo di mutazione sono la distrofia muscolare facioscapuloomerale e vari tipi di tumori legati alla sovraespressione di SRSF1 La ricerca nel campo dello splicing alternativo è vitale per prevedere i pattern di splicing alternativo e per progettare protocolli di terapia genica in grado di correggere i difetti dello splicing alternativo Alternative splicing: a pivotal step between eukaryotic transcription and translation - Nat Rev Mol Cell Biol. 2013 Mar;14(3):153-65. doi: 10.1038/nrm3525 - Kornblihtt et al. - Erratum in Nat Rev Mol Cell Biol. 2013 Mar;14(3) - doi:10.1038/nrm3560 Nel nucleo, il processo di editing altera la sequenza degli mRNA serve per il trasporto di colesterolo nel sangue serve per l’assorbimento di lipidi nell’intestino Il processo di editing dell’RNA altera la sequenza di mRNA, ad esempio mediante inserzione di uracile (U) mRNA maturo (dopo splicing ed eventuale editing) che può essere esportato nel citoplasma per essere tradotto in proteina
© Copyright 2026 Paperzz