Paper
zz
Explore Categories
Log in
Create new account
No category
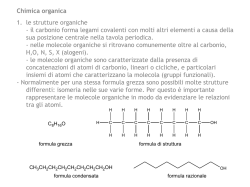
Dal carbonio al biotech Chimica organica, biochimica e
Download
Report
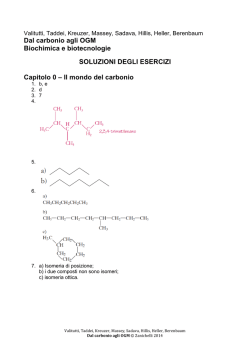
Dal carbonio agli OGM – Soluzioni degli esercizi
L`OSSERVATORE ROMANO
Reigen / Liebelei: 2 Theaterstücke
OWNER`S MANUAL
Biologia 14 - Biochimica strutturale
I grandi esploratori
CO2 IN RECIPIENTI ERMETICI (prova del CO2 nello spazio di testa)
Cap.11_par.04_Leggi di Keplero - Liceo Classico Psicopedagogico
Liceo delle Scienze Umane - Istituto Maria Ausiliatrice
Roma - Socrate srl
iWind - Italsol
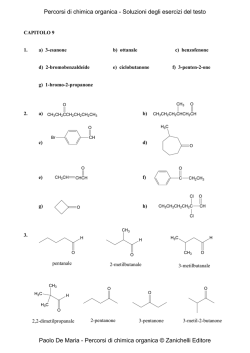
Capitolo 9 Aldeidi e chetoni - Zanichelli online per la scuola
© Copyright 2026 Paperzz
About Paperzz
DMCA / GDPR
Report