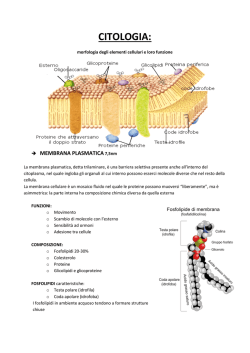
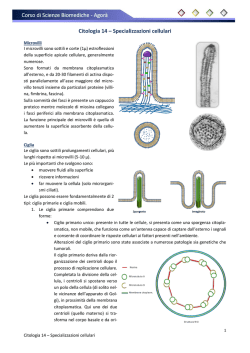
La cellula (seconda parte) LA LEZIONE Parete cellulare In alcuni organismi (protisti e piante) la parte più esterna della membrana cellulare è ricoperta da una parete cellulare (fig.1), con spiccate funzioni di protezione dovute alla sua rigidità. Nei batteri tale parete è formata da peptidoglicani. La variabilità nella sua composizione determina differenze nella reazione alla colorazione di Gram (contenente il colorante cristalvioletto), permettendo di distinguere i batteri in Grampositivi (che diventano blu) o Gram-negativi (che si decolorano). Nei vegetali la parete presenta una composizione stratificata più complessa e, procedendo dall'esterno verso l'interno, è possibile evidenziare: una lamella mediana, a base soprattutto di pectine; una parete primaria, dotata di elasticità e formata anch'essa soprattutto da pectine; una parete secondaria, più rigida della primaria e contenente in modo particolare cellulosa, nonché altre sostanze come la lignina. Il principale componente della parete cellulare è comunque la cellulosa, che nella parete interna e in quella esterna è presente in forma di fibrille. Figura 1 - Schema di parete cellulare eucariotica Citoplasma L'interno della cellula è in buona parte occupato dal citosol, una soluzione salina acquosa con consistenza gelatinosa (colloidale), costituita per circa il 70% da acqua. Il valore del pH è vicino alla neutralità. Oltre ad acqua e sali, contiene anche carboidrati, aminoacidi, enzimi, nucleotidi, metaboliti, prodotti intermedi delle reazioni, ATP (adenosine triphosphate), proteine varie, per esempio quelle di trasporto, ecc. Il citosol, insieme agli organelli cellulari e al citoscheletro in esso contenuti, forma il citoplasma. In tutte le cellule il citoplasma rappresenta il compartimento operativo della vita cellulare, ossia quello in cui la maggior parte dei segnali, di varia natura e origine, viene elaborata per produrre le opportune risposte. Le differenze osservabili nella presenza degli organelli citoplasmatici permettono di distinguere le cellule in procariotiche o eucariotiche. Parleremo prima di quelle più complesse, ossia delle cellule eucariotiche, che sono tipiche sia delle piante sia degli animali. Hanno molte caratteristiche comuni, per le quali possono essere distinte con facilità dalle cellule procariotiche, ma presentano anche qualche differenza tra di loro, legata alla grande diversità funzionale degli organismi ai quali appartengono. Citoscheletro L’organizzazione interna e il movimento delle cellule eucariotiche dipendono da una complicata rete di filamenti proteici (filamenti di actina, microtubuli, filamenti intermedi) presenti nel citoplasma e che costituiscono, nel loro complesso, il citoscheletro (fig.2). Tale struttura ha funzioni al tempo stesso statiche e dinamiche. Infatti, è prima di tutto responsabile dell'organizzazione interna della cellula e del mantenimento della sua forma, ma garantisce anche i cambiamenti di conformazione, dando alla cellula la necessaria plasticità per affrontare le sollecitazioni dell'ambiente esterno. Inoltre, il citoscheletro contribuisce al movimento della cellula, al trasporto interno di molecole, vescicole e organelli, indirizzandoli verso la specifica destinazione, alla formazione del fuso mitotico, fondamentale per la divisione cellulare, e all'ancoraggio degli organelli. I microtubuli hanno origine dal centrosoma, che è preposto anche alla loro organizzazione. Figura 2 - Citoscheletro in cellule endoteliali fluorescenti viste al microscopio confocale: la tubulina è marcata in verde, l'actina in rosso Estroflessioni cellulari Alcune cellule sono dotate di estroflessioni collegate al citoscheletro, dotate di funzioni diverse. Tali estroflessioni sono microvilli, ciglia e flagelli. I primi sono estensioni della membrana plasmatica dell'apice cellulare e tappezzano gli epiteli di rivestimento delle cavità interne dell'organismo. La loro funzione è quella di aumentare la superficie di scambio, garantendo così alle cellule il necessario assorbimento di acqua e sostanze nutritive. I microvilli sono costituiti da circa trenta filamenti di actina, disposti in parallelo e uniti da ponti proteici. Le ciglia sono solitamente numerose e brevi (ca. 10 μm), capaci di muoversi all'unisono creando correnti intorno alla cellula, utili per esempio per allontanare sostanze indesiderate, come accade nelle mucose delle vie respiratorie, o per permettere lo spostamento della cellula uovo nelle tube di Falloppio. Le ciglia, così come i flagelli, richiedono energia (ATP) per svolgere il loro compito. I flagelli sono invece molto lunghi (fino a 500 μm) e generalmente sulla singola cellula ne è presente uno solo. Producono un'elevata velocità di propulsione, come nel caso degli spermatozoi che si muovono verso l'ovulo. La parte interna di un ciglio o di un flagello è detta assonema o centriolo e presenta una struttura costante in quasi tutti gli eucarioti: una membrana che racchiude nove coppie di microtubuli, che a loro volta circondano due microtubuli centrali non accoppiati. L'assonema è attaccato al cosiddetto corpuscolo basale, anch'esso formato da microtubuli, ma con una struttura leggermente diversa da quella dell'assonema: nove triplette esterne che circondano due microtubuli singoli centrali. Per maggiori informazioni sul movimento della cellula si può consultare questo video. Organelli cellulari Gli organelli o organuli cellulari (mitocondri, reticolo endoplasmatico, plastidi, ribosomi, lisosomi, cloroplasti ecc.) sono strutture delimitate da membrane fosfolipidiche. Ognuno di loro è dotato di specifiche funzioni determinate dal proprio corredo enzimatico. Inoltre, gli organuli sono in grado di interagire tra di loro, scambiandosi materiali grazie ai processi di esocitosi ed endocitosi. Durante la mitosi (processo di divisione cellulare di cui si parlerà più diffusamente nella voce Cellula, terza parte) gli organelli sono ripartiti fra le due cellule figlie, tranne i mitocondri e i cloroplasti che sono in grado di duplicarsi da soli. Reticolo endoplasmatico Questo organello è costituito da un esteso sistema di membrane che formano cisterne (strutture la cui forma ricorda quella di un sacco appiattito) e tubuli collegati tra loro fino a costruire una rete. Le cisterne sono coperte da complessi di ribosomi (poliribosomi o polisomi), mentre i tubuli sono lisci: per questo motivo si parla, rispettivamente, di reticolo granulare (o rugoso) e di reticolo liscio (o agranulare). La presenza di polisomi indica come il reticolo sia sede di sintesi proteica, analogo in questo al citosol, che contiene polisomi non legati a membrana e quindi definiti liberi. Questa duplice localizzazione di un processo fondamentale è dovuta al fatto che le proteine sono differenti tra loro, sia per struttura molecolare sia per funzioni, e quindi le proteine destinate al citosol, alla faccia citosolica delle membrane, al nucleo e agli organuli semiautonomi, sono sintetizzate nei polisomi liberi; invece, quando le proteine sono destinate alla secrezione (nel caso di proteine solubili) o all'inserimento nel lume o nelle membrane degli organuli connessi funzionalmente con i processi di esocitosi ed endocitosi, la sintesi ha luogo quasi sempre nei ribosomi legati alle membrane del reticolo. Nella sua attività di esocitosi il reticolo funziona in stretta correlazione con l'apparato del Golgi. Il reticolo endoplasmatico svolge anche altre funzioni importanti: per esempio, la sintesi di fosfolipidi, destinati a essere poi trasferiti in larga misura ad altre membrane attraverso il citosol o tramite l'esocitosi; l'accumulo di Ca 2+ a rapido scambio, fondamentale per il funzionamento cellulare di tutte le attività calcio-dipendenti, in regioni specializzate del reticolo endoplasmatico (come il reticolo sarcoplasmatico del muscolo) ove viene trattenuto e quindi liberato quando stimolazioni elettriche (nel muscolo scheletrico) o da parte di secondi messaggeri (nelle altre cellule) inducono l'apertura di canali nella membrana di tali regioni. Apparato del Golgi E' un organello composto da 3-5 cisterne appiattite distinte l'una dall'altra e disposte parallelamente, denominate cis (la più interna, che guarda verso il nucleo e la cui superficie è nota come faccia cis, oppure faccia concava o immatura) e trans (la più periferica, che guarda verso la membrana e la cui superficie è chiamata faccia trans, o anche faccia matura o convessa) e da una rete composta da tubuli e vescicole (trans-Golgi network, TGN) ampiamente continui tra loro (fig.3). Figura 3 - Apparato del Golgi osservato al microscopio elettronico Il ruolo dell'apparato del Golgi è quello di modificare le proteine provenienti dal reticolo tramite coniugazione con zuccheri (glicosilazione), oppure con residui fosfato (fosforilazione) o solfato (solfatazione). Grazie alla molteplicità delle cisterne è possibile assicurare l'ordinato svolgimento dei numerosi eventi connessi con questi processi; infatti, vescicole contenenti le proteine prodotte dal reticolo endoplasmatico rugoso sono trasportate alla faccia cis dell'apparato del Golgi, si fondono con la sua membrana e rilasciano il proprio contenuto all'interno dell'apparato. Qui le molecole sono modificate, ed è bene sottolineare che gli apparati del Golgi generalmente sono più grandi e numerosi nelle cellule dotate di intensa attività di sintesi e secrezione (per esempio, i linfociti che producono anticorpi). Successivamente, il trasporto prosegue attraverso la faccia trans e poi la TGN, dove le proteine sono raggruppate, inglobate in altre vescicole e spedite verso la propria specifica destinazione. In molte cellule, inoltre, nella rete di tubuli dell'apparato del Golgi si può verificare un mutamento dello stato di condensazione di alcune proteine che, non trovandosi più in soluzione, formano aggregati specifici che vanno a costituire i granuli di secrezione. Questo meccanismo permette alla cellula di accumulare i prodotti della propria secrezione finché un segnale (per esempio, l'aumento del Ca 2+ nel citosol) non induca la fusione (esocitosi) sincrona della membrana di molti granuli con il plasmalemma, con conseguente secrezione extracellulare del loro contenuto. Mitocondri I mitocondri sono organuli presenti nel citoplasma di tutte le cellule eucariotiche, sia animali sia vegetali, mentre mancano nelle cellule procariotiche. Tra le funzioni svolte, la più importante è la demolizione di molecole organiche per ricavare energia, che viene poi conservata sintetizzando ATP, la molecola che fornisce il “carburante” necessario ai processi vitali della cellula (v. Glicolisi, Respirazione cellulare). Nei batteri tale funzione è invece espletata da enzimi contenuti nella membrana cellulare e in sue introflessioni dette mesosomi. Il mitocondrio è comunque coinvolto anche in altri processi, tra i quali, a titolo di esempio, si possono ricordare: la regolazione dell'apoptosi (morte programmata della cellula), la sintesi del gruppo eme, la sintesi del colesterolo, la produzione di calore nel tessuto adiposo bruno (negli animali in letargo e in quelli adattati al clima freddo). I mitocondri hanno forma solitamente allungata e sono costituiti da due membrane, una interna e l'altra esterna; lo spazio presente tra le due è chiamato spazio intermembrana, mentre lo spazio delimitato dalla membrana interna è detto matrice mitocondriale (fig. 4). Figura 4 - Mitocondrio visto al microscopio elettronico La membrana interna ha un aspetto molto caratteristico, poiché si estende nella matrice formando delle pieghe dette creste mitocondriali; in esse è contenuto l'apparato di proteine necessario alla sintesi di ATP a partire da molecole precursori, ossia gli enzimi coinvolti nella fosforilazione ossidativa e quelli dell'ATP sintetasi. In breve, attraverso il citosol entrano nel mitocondrio sostanze da metabolizzare, alle quali gli enzimi della matrice sottraggono elettroni, ossidandole. Gli elettroni passano ad altri enzimi, allineati nella membrana interna e responsabili della catena di trasporto degli elettroni. Il passaggio degli elettroni lungo tale catena garantisce una caduta progressiva, a salti, della loro energia libera, fino a raggiungere l'ossigeno con il quale gli elettroni si legano formando acqua. La graduale perdita di energia è utilizzata per espellere H+ dal mitocondrio, creando così un gradiente elettrochimico tra la membrana interna, che diviene elettronegativa, e la matrice intermembrana, che diventa alcalina. Queste condizioni favoriscono il rientro degli H+, rendendo così disponibile energia per il funzionamento di una ATP-sintetasi che genera ATP. Grazie a questo perfetto sistema il mitocondrio è una delle macchine più efficienti esistenti in natura: pochissima dell'energia liberata dall'ossidazione dei substrati viene infatti dispersa in calore. Un aspetto molto interessante dei mitocondri, ma anche dei cloroplasti, è la presenza in essi di DNA e ribosomi, ossia un intero patrimonio genetico (anche se molto ridotto), completo di sistema di espressione, simile a quello delle cellule procariotiche. Questa proprietà ha fatto ipotizzare che tali organelli siano il prodotto di un rapporto di simbiosi nato nel passato: infatti, si pensa che circa un miliardo e mezzo di anni fa qualche batterio sia entrato in alcune cellule eucariotiche, trovando un riparo e a propria volta fornendo l'apparato per svolgere un metabolismo ossidativo di tipo aerobio, di cui le cellule erano prive. Successivamente, le cellule hanno inglobato nel proprio patrimonio genetico la maggior parte di quello batterico, lasciandone però una porzione sufficiente ad assicurare una certa autonomia a quelli che ormai erano diventati organuli cellulari. Il mitocondrio presenta un'altra caratteristica molto studiata: questo organello si trova nelle cellule uovo mentre è generalmente assente negli spermatozoi, ovvero è ereditato dalla madre (via matrilineare). Si tratta di un'eredità non mendeliana. Il DNA mitocondriale dei figli è quindi quasi uguale a quello materno, a meno che non siano avvenute mutazioni. Questa proprietà risulta molto utile per esaminare determinate patologie a trasmissione mitocondriale. Inoltre, è alla base di importanti ricerche sull'evoluzione del genere umano: infatti, grazie a studi di genetica delle popolazioni condotti analizzando il genoma del mitocondrio, è stato possibile determinare gradi di parentela, migrazioni e discendenze, arrivando a teorizzare l'esistenza di una progenitrice che gli scienziati hanno soprannominato Eva mitocondriale. Cloroplasti I cloroplasti, presenti solo nelle piante e nelle alghe, sono molto simili ai mitocondri. Infatti, la loro funzione consiste nello svolgere i processi del metabolismo aerobico. Inoltre, dal punto di vista morfologico, sono circondati da una doppia membrana, una esterna che delimita un primo compartimento, e una interna che circonda un compartimento più ampio (stroma). All'interno dello stroma è presente un altro complesso sistema formato da membrane, i tilacoidi (fig.5). Figura 5 - Cloroplasto visto al microscopio elettronico Questi sono membrane appiattite in forma di dischi impilati (grana), oppure semplici lamine (lamellae stromatiche), e in essi si trova la clorofilla, un pigmento che è capace di captare la luce in un processo che porta alla sintesi di ATP, utilizzato per fissare l'anidride carbonica nelle sintesi degli zuccheri (v. Fotosintesi). Molto brevemente, il processo inizia con la captazione dei fotoni, che fornisce l'energia necessaria a trasportare protoni (H+) attraverso la membrana nel lume dei tilacoidi; in questo modo si crea un potenziale elettrochimico (con l'interno dei tilacoidi acido e caricato positivamente), che rappresenta una fonte di energia. Tale energia è rilasciata nel processo di ritorno degli H+ nello stroma ed è utilizzata dall'ATP-sintetasi per sintetizzare ATP attraverso la fosforilazione dell'ADP (adenosine diphosphate). L'ATP è poi fonte di energia per sintetizzare i carboidrati a partire da CO 2 (ciclo di fissazione della CO2). Questo insieme di reazioni garantisce la base dell'equilibrio ecologico sulla Terra, perché permette la vita delle piante e degli animali che di esse si nutrono; senza queste reazioni non sarebbe possibile la vita come noi la conosciamo. Anche per i cloroplasti, come già detto, si ipotizza un'origine simbiotica, poiché sono anch'essi dotati di DNA e ribosomi e quindi capaci di replicarsi. Lisosomi e perossisomi I lisosomi sono organelli cellulari impegnati nei processi di degradazione e digestione di molecole provenienti dall'ambiente esterno (eterofagia) ma anche da quello interno (autofagia). Questa capacità è dovuta al fatto che i lisosomi contengono in abbondanza enzimi idrolitici (idrolasi acide), capaci di demolire proteine, carboidrati e lipidi fino a ottenere i costituenti di base; questi ultimi, grazie al basso peso molecolare, possono diffondere attraverso le membrane per essere poi utilizzati nuovamente dalla cellula nei suoi processi di sintesi. Il fatto che gli enzimi lisosomiali funzionino a un pH basso rappresenta una protezione per la cellula, che avendo un pH neutro costituisce un ambiente sfavorevole per l'attività di tali enzimi, nell'eventualità che questi fuoriescano dai lisosomi. In generale i lisosomi svolgono il loro compito fondendosi con le vescicole che racchiudono le molecole da digerire, creando così un'unica vescicola. Essi garantiscono l'eliminazione di sostanze di rifiuto e tossiche o di componenti malfunzionanti; inoltre la loro attività è alla base dell'apoptosi. I lisosomi hanno origine per gemmazione dall'apparato del Golgi, mentre gli enzimi idrolitici provengono dal reticolo endoplasmatico e sono successivamente modificati nell'apparato del Golgi. Gli enzimi sono quindi rilasciati entro vescicole endocitosiche che si fondono con i lisosomi versando in essi il proprio contenuto. Anche i perossisomi sono organelli cellulari dotati di enzimi, però di tipo ossidativo. Questi enzimi permettono svariate funzioni, per esempio, di ossidare gli acidi grassi a lunga catena (β-ossidazione), di sintetizzare colesterolo e acidi biliari, di partecipare al metabolismo di aminoacidi e purine, di contribuire allo smaltimento dei metaboliti tossici. La caratteristica distintiva dei perossisomi è quella di produrre acqua ossigenata (perossido d'idrogeno) come scarto finale di reazione. Ribosomi Questi organelli sono legati al reticolo endoplasmatico rugoso e alla membrana nucleare (nel caso di cellule che secernono le proteine prodotte) ma si ritrovano anche liberi nel citoplasma, oppure nei mitocondri e cloroplasti (quando le proteine prodotte dalla cellula sono immagazzinate). I ribosomi sono formati da RNA (ribonucleic acid) e da proteine e hanno un coefficiente di sedimentazione di 80S (espresso in unità Svedberg). Sono costituiti da due subunità di dimensioni differenti (fig.6), la più grande denominata 60S (coefficiente di sedimentazione, espresso in unità Svedberg) e la più piccola 40S. Figura 6 - Schema di ribosoma eucariotico Le due subunità sono a loro volta scomponibili: la maggiore è formata da tre molecole di rRNA (5S, 5,8S, 28S); la minore da una singola molecola di rRNA (18S). Tali molecole (tranne 5S) sono sintetizzate nei nucleoli, dove l'enzima RNA polimerasi agisce su più punti della catena di DNA, trascrivendoli; in questo modo si formano strutture (cistroni) che emergono dalla catena di DNA, così come i rami di un albero si dipartono dal tronco. Questo RNA ribosomale (RNA 45S) è detto pre-rRNA, perché sono necessari tagli per formare le subunità 18S e 32S, quest'ultima ulteriormente tagliata a dare le subunità 28S e 5,8S. I ribosomi intervengono nella sintesi proteica. Per svolgere la loro attività, le due subunità del ribosoma devono essere unite, per poter così leggere una molecola di mRNA (RNA messaggero) tripletta per tripletta, associando a ognuna di queste il rispettivo aminoacido portato dal tRNA (RNA di trasporto) e formando una specifica proteina (v. anche Cellula, terza parte; Le molecole informazionali e il DNA). Centrosoma e centrioli Il centrosoma è una struttura priva di membrana, posta vicino al nucleo e circondata da una massa amorfa di materiale. La sua funzione è quella formare, organizzare e distruggere i microtubuli. Nel centrosoma sono visibili i centrioli, coppia di piccole strutture cilindriche, cave e con nove triplette di microtubuli disposte in cerchio. Il centrosoma è fondamentale nella mitosi, che è regolata grazie ai movimenti dei centrioli e dei microtubuli (per una più ampia trattazione di questo processo v. Cellula, terza parte), ma ha un ruolo di grande importanza anche nella formazione delle estroflessioni cellulari che contengono microtubuli, come microvilli, ciglia e flagelli, e inoltre contribuisce a regolare il trasporto interno alla cellula che fa capo al reticolo endoplasmatico e all'apparato del Golgi. Vacuoli. Nelle cellule vegetali sono presenti organuli chiamati vacuoli, soprattutto deputati ad accumulare e conservare acqua e materiali in essa disciolti o comunque dispersi (come ioni minerali, metaboliti secondari ecc.). I vacuoli originano da una regione specifica del reticolo endoplasmatico e nella cellula giovane sono piccoli e numerosi, mentre nella cellula matura è solitamente presente un solo grande vacuolo, formatosi a seguito della fusione dei vacuoli più piccoli.
© Copyright 2026 Paperzz