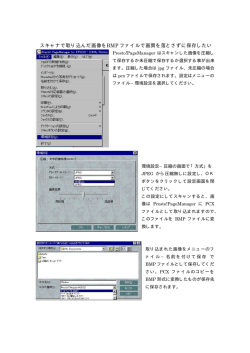
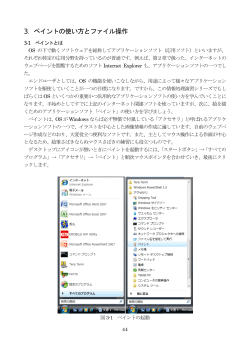
教授 上野 直人 准教授 木下 典行 助教 高橋 弘樹 鈴木 誠 技術課技術職員 高木 知世 博士研究員 田尾 嘉誉 進藤 麻子 アフリカツメガエルの形態形成と、その基盤となる細胞運動やシグナル伝達 総合研究大学院大学 大学院生 森田 仁 動物はひとつの受精卵から細胞分裂を繰り返して細胞の数を増やし、それぞれの 細胞の性質を変えながら、生物として固有の形づくり ( 形態形成 ) を行う。その 過程には細胞同士のコミュニケーション、すなわち細胞間相互作用が重要である ことが知られている。細胞間相互作用は、ある細胞集団から細胞増殖因子などが 分泌され、標的となる細胞のシグナル伝達系を活性化し、その細胞の中で転写調 節因子や細胞骨格の働きを調節することによって細胞分化、細胞運動をダイナ ミックに制御する。私たちはこの過程をプログラムとして理解し、動物種間で比 較することによって、形態形成メカニズムの共通性、多様性に迫りたいと考えて いる。 24 原 佑介 技術支援員 山本 隆正 村上 美智代 事務支援員 三宅 智子 柘植 豊子 形態形成研究部門 http://www.nibb.ac.jp/morphgen/ 生きものかたちづくりに共通する分子基盤 この脊索をもった最も原始的な動物であり、カエルやヒトに 地球上の生き物の形は実に多様である。球形の卵からこれ 比べるとその形成過程は単純であることから、脊索ができる ら動物の複雑なかたちはどのようにできるのか、その仕組み 分子メカニズムを研究するのに適した動物といえる。私たち を分子や細胞レベルで解き明かすのが私たちの目標である。 はホヤの脊索形成の基本形として、動物の脊索形成を理解し 最近の研究の進歩によって、一見多様に見える生物もそれら ようとしている ( 図 1)。 をかたちづくる基本の仕組み自体には大きな違いはなく、良 く似た遺伝子を少しだけ使い分けたり、使う時期や場所を変 細胞極性の確立 えることによって、多様な形を作りだしていることが分かっ 形ができる仕組みを理解するためには個体全体をマクロな てきた。私たちはアフリカツメガエルを用いた研究から、発 視点で見ることも大切だが、個体を構成する個々の細胞の振 生後期では骨分化を制御する骨形成タンパク質 (BMP) が、 る舞いとして理解することも重要である。個体が正しく形づ 発生初期には、背腹軸形成に決定的な役割を担っていること くられるためには個々の細胞が自己の形態や相対位置、運動 を明らかにした。後に BMP は、1920 年代にシュペーマ の向きを決めるための基準、「細胞極性」を読みとる必要が ンによって発見され、長い間その物質的基盤が不明であった ある。細胞極性が確立するためには細胞内外の分子の協調的 「オーガナイザー」の謎を解く鍵となった。オーガナイザー な働きが必要であることが分かってきた。私たちは、この細 は胚に移植することによって、「誘導」と呼ばれる細胞間相 胞極性がどのように形成されるのか、細胞はどのようにそれ 互作用を引き起こし、もうひとつの体を作ることができる能 を読みとって形や運動の変化へと結びつけるのかといった問 力を持っている。このオーガナイザーの神秘的な活性の本体 題を、生体分子や反応をリアルタイムで可視化する「バイオ は BMP に対し拮抗作用をもつ複数の分泌性タンパク質であ イメージング」をとりいれながら研究している ( 図 2)。 ることが明らかとなった。ホヤやショウジョウバエといった 脊椎動物とはかけ離れたかたちの動物たちもこの BMP を持 ち、発生制御に共通の役割、異なる役割を使い分けている。 事実、ショウジョウバエを使った研究から、シグナル伝達に 関わる多くの因子が発見され BMP の作用メカニズムの解明 に大きく貢献して いる。つまり、発 生など生命現象の 基本原理を明らか にするためにどの ような動物を使う かはあまり大きな 問題ではなく、知 りたいと思う現象 を観察するために 最も適した動物を 図 1. ホヤの胚の伸長運動 左は胚の表面、右は断面図。脊索は尾の中心部の細長い 構造。 使うことが大切だ と考えている。 脊索形成のメカニズムを探る 昆虫にはみられず、ある動物群にだけ特徴的な構造がある。 脊索と呼ばれる組織である。脊索は発生の過程では体の中心 構造としてつくられるが、将来脊椎骨に置き換わる。ホヤは 教授 上野 直人 准教授 木下 典行 助教 高橋 弘樹 図 2. EB3-GFP による微小管ダイナミクスのリアルタイム解析 左は極性化していないアニマルキャップ細胞、右は極性化した中胚葉細胞。 参考文献 1. Chung, H. A., Yamamoto, T. S., and Ueno, N. (2007). ANR5, an FGF Target Gene Product, Regulates Gastrulation in Xenopus. Curr. Biol. 17, 932-939. 2. Hotta, K., Yamada, S., Ueno, N., Satoh, N., and Takahashi, H. (2007). Brachyury-downstream notochord genes and convergent extension in Ciona intestinalis embryos. Dev. Growth. Differ. 49, 373-382. 3. Ogata, S. Morokuma, J. Hayata, T. Kolle, G. Niehrs, C. Ueno, N.* and Cho, K. W.* (*corresponding authors). (2007). TGF-beta signalingmediated morphogenesis: modulation of cell adhesion via cadherin endocytosis. Genes Dev. 21, 1817-1831. 4. Iioka, H., Iemura, S., Natsume, T. and Kinoshita, N. (2007). Wnt signalling regulates paxillin ubiquitination essential for mesodermal cell motility. Nat. Cell Biol. 9, 813-821. 5. Lee, R. H., Iioka, H., Ohashi, M., Iemura, S., Natsume, T. and Kinoshita, N. (2007). XRab40 and XCullin5 form a ubiquitin ligase complex essential for the noncanonical Wnt pathway. EMBO J. 26, 3592-3606. 助教 鈴木 誠 25
© Copyright 2026 Paperzz