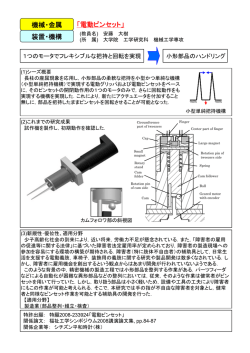
記憶をリン酸化で語る 東北大学大学院薬学研究科薬理学分野 福永 浩司 学習あるいは経験は神経のネットワークに記憶される。記憶の回路形成には、ネットワークを構 成するニューロン間のシナプス伝達効率が促進されることが重要である。 記憶の素過程には このシナプス伝達効率の変化(シナプス可塑性)が関わっている。 1982 年に私達が脳で発 見した カルシウム/カルモデュリン依存性プロテインキナーゼ II (CaM キナーゼ II) はシ ナプスにおいて記憶を作るときに必須のリン酸化酵素である。記憶のプロセスは記銘 (encording)、保持 (maintenance)、固定 (consolidation)、想起 (recall) に分けられるが,これ らに関与する神経機構は異なっている。図 1 に示したように、視覚、聴覚、触覚、知覚から得 られた情報(入力)は海馬を通過する間に統合・選別されて、情報に対応した大脳皮質の記憶 の回路に送られる。情動を伴った重要な情報が海馬に入ってくると神経終末から多くのグルタ ミン酸が放出され、シナプス後部で NMDA 受容体が活性化される。海馬の NMDA 受容体の 活性化反応は記憶するための最初のステップであるが、大脳皮質の記憶回路形成の分子機 構も基本的に同じである。NMDA 受容体からのカルシウム流入に続いてシナプスの後部では CaM キナーゼ II が活性化される。 固定 記銘 目, 耳, 感覚 から 入っ て きた 情報 は脳 内の 海馬 と いう 部位 を通 って 神経 細 胞の ネッ トワ ーク に入 っ てい く。 海馬 一度 作ら れた 神経 のネ ット ワー クで はシ ナプ スの 結合 数や 形態 変化 が長 期に 持続 する こと によ って 記憶 は固 定さ れる (長期 記憶 )。 想起 維持 シナ プス 神経 のネ ットワ ーク 記憶 はシ ナプ スで 作ら れ る。 外界 から の刺 激 によ って 同じ 神経 ネッ トワ ーク が興 奮し て, 記憶 が再 生さ れる (想起 )。 図 1 . 記憶のプロセス 遺伝子改変マウスの学習行動実験から、CaM キナーゼ II は記憶の記銘と維持機構に関与 していることが明らかとなった。一度、形成された記憶の回路は時折、活性化されることによっ て維持される。この繰り返される記憶回路の活性化反応が長期記憶形成に重要である。記憶 の維持には CaM キナーゼ II 以外にも CaM キナーゼ IV と MAP キナーゼが関わってい る。 シナプス入力線維を高頻度刺激(数十から数百 Hz,数秒間:HFS) すると、刺激をやめた 後、数時間から数日間、シナプス伝達効率が増強される現象が海馬で見出された (1)。この 現象は長期増強 (long-term potentiation; LTP) と名付けられた。最初に貫通線維から歯状 回顆粒細胞への入力線維に見出された現象である。この LTP 現象は海馬 CA1、CA3 領域、 大脳皮質、扁桃核など様々な部位で観察される。 CaM キナーゼ II は記憶分子として働くた めの面白い性質を持っている。一つは CaM キナーゼ II は分子内の 286 番目のスレオニ ン残基 (Thr-286)が自己リン酸化されることである。次に、本酵素は同じサブユニットが 10 分 子会合して、オリゴマーを形成することである(図 4)。自己リン酸化されるとカルシウム非依存 性となり、オリゴマーを形成することで恒常的に活性を持つようになる。すなわち、 高頻度刺 激により NMDA 受容体イオンチャネルが活性化される。後シナプスのカルシウムイオンの上 昇は一過性であるが、この一過性のカルシウム上昇によって活性化された CaM キナーゼ II は恒常的に活性を持つと考えられる (2)。 C A 30 CaM キナーゼ II 記録電極 ** Ca 2+-非依存性活性 (nmol/mg/min) 刺激電極 シャッファー側枝 CA1 交連線維 歯状回 CA3 貫通繊維 苔状線維 刺激前 EPSP 10 msec 200 150 100 50 20 20 220 刺激後 EPSP 250 0 ** 0 10 20 30 40 50 60 D 1 mV EPSP 振幅 (%) 300 ** 25 15 高頻度刺激 40 時間 (min) 60 80 CaM キナーゼ IV 活性(%) B 高頻度刺激 コントロール CaM キナーゼ IV 高頻度刺激 ** 200 180 160 140 * 120 100 80 0 10 20 30 40 50 60 時間 (min) 図 2. ラット海馬 CA1 領域 LTP における CaM キナーゼ II と CaM キナーゼ IV の活性変化 実際に、高頻度刺激して LTP を誘発した海馬 CA1 領域で CaM キナーゼ II の活性を測 定した。対照として CaM キナーゼ IV の活性も同時に測定した(図 2)。 CaM キナーゼ II のカルシウム非依存性活性は高頻度刺激直後に上昇して,その後 1 時間以上持続した (3)。 一方、核内に存在する CaM キナーゼ IV の活性化反応は一過性であり、持続しない。活性 化された CaM キナーゼ II の存在する部位を自己リン酸化された Thr-286 を認識する抗 体を用いて調べると,CA1 錐体細胞の樹状突起とスパインで自己リン酸型 CaM キナーゼ II が増えていることが解る(図 3)。 コン トロ ール LTP 図 3 . 海馬 L T P 誘導後の C A 1 錐体細胞における活性型 C a M キナーゼ II の局在の免疫組織化学的変化 NMDA 受容体は可塑性に関与する受容体であるが、通常のシナプス伝達は AMPA 受容 体が担っている。CaM キナーゼ II は活性化されたスパインにおいて AMPA 受容体をリン 酸化することによって(4)、細胞質からシナプス後部膜への AMPA 受容体の輸送(delivery) を促進することによって (5)、 シナプス後部での AMPA 受容体イオンチャネルのコンダクタン スを上昇させる。海馬の LTP 現象はこの AMPA 受容体の持続的な活性化反応を反映して いるのである。さらに、 CaM キナーゼ II は細胞骨格蛋白質のリン酸化反応によってスパイ ンの形態変化にも関与している (6)。この記憶の分子機構は大脳新皮質における記憶形成で も同じである。したがって CaM キナーゼ II は大脳皮質においては長期記憶の形成にも関 わっている (7)。 一方,錐体細胞の核内に存在する CaM キナーゼ IV は CaM キナーゼ・キナーゼで Thr-196 がリン酸化され,活性化される。 LTP においては核内で一過性に活性化され, cAMP 反応エレメント結合蛋白質 (CREB) をリン酸化する (8)。 長期記憶形成には遺伝子 発現と蛋白質合成を介した新しいシナプスや受容体の発現が必要である。記憶の固定・維持 に CaM キナーゼ IV による CREB のリン酸化反応、続いて脳由来神経栄養因子 (BDNF) の発現が関わっている。 CREB によって誘導されるその他の記憶関連遺伝子の同定が進 んでいる。 MAP キナーゼも基本的にはこの遺伝子発現を介する長期記憶形成に関わって いる。 記憶の形成と同様に、記憶の消去のメカニズムの研究も進んでいる。記憶装置にも容量が あり、重要ではない情報、忘れたい記憶は消去する必要がある。 グ ルタ ミン 酸神 経 終末 神 経細 胞 体 AMPA 受容 体 P ス パイ ン 電 位依 存性 Ca2+ チ ャ ネル NMDA 受 容体 受 容体 の 輸送 P Ca2 + P CaM kinase kinase Ca2 + CaM kinase II CaM kinase IV MAP kinase MAP kin ase P 樹 状突 起 P CREB P 転写 LTP の発現 BDNF 自 己リ ン 酸化 型 C aM キ ナー ゼ II 図 4. 海馬における LTP の発現と維持機構 LTP の維持 面白いことに記憶の消去にも NMDA 受容体が使われる。海馬においては 10-100 Hz の刺 激で誘発した LTP を 1-5 Hz の低頻度刺激で消去することができる。あるいは 1-5 Hz の低 頻度刺激では長期抑圧(Long-term depression; LTD)という現象も観察される。図 5 に示す ように、高頻度刺激 (HFS) では後シナプスでのカルシウム上昇は大きく細胞内カルシウム濃 度は 1〜2 µM 以上に達する。この濃度では CaM キナーゼ II とカルシニューリン(カルシウ ム依存性脱リン酸化酵素)が活性化される。実際にはリン酸化される基質に対する親和性の違 いから CaM キナーゼ II によるリン酸化反応が優位に起こる。 前 シナ プ ス 細 胞内 C a 2 + レ ベ ル 後 シナ プ ス HFS [Ca 2+ ] > 2 µM CaM kinase II NMDA 受 容 体 Ca2+ 蛋 白質 リ ン 酸化 反応 蛋 白質 脱 リ ン酸 化 反応 LFS [Ca 2+ ] < 1 µM Calcineurin 図 5. 記憶の消去の メカニズム 一方、記憶を消去する低頻度(1 〜 5 Hz) の刺激(LFS)では後シナプスでのカルシウム上 昇は小さく 1 µM 以下である。この濃度では CaM キナーゼ II は活性化されない。カルシウ ムに高親和性のカルシニューリンのみが活性化される。したがって、記憶に関わるシナプスで のリン酸化蛋白質は脱リン酸化されることになる。このカルシニューリンによる脱リン酸化反応 がシナプス伝達効率を低下させる。本研究では海馬におけるシナプス可塑性にリン酸化反応 が関わっていること。 CaM キナーゼ II やカルシニューリンなどのカルシウム依存性酵素が 記憶形成のシグナル伝達の一部を担っていることを明らかにした。アルツハイマー病や脳血 管障害においては神経細胞が脱落することによって記憶が部分的に障害されていく。私達は 最近、神経細胞死が起こらない程度の脳虚血においても CaM キナーゼ II が神経細胞にお いて変性する現象を見出した。 CaM キナーゼ II の神経細胞における変性と記憶障害の関 連を今後、明らかにしていきたい。 参考文献 1) Bliss TVP sand Lømo T. J Physiol 232:331-356, 1973 2) Fukunaga K et al. Neurosci Res 38: 3-17, 2000 3) Fukunaga K et al. J Biol Chem 268: 7863-7867, 1993 4) Barria A et al. Science 276: 2042-2045, 1997 5) Hayashi Y et al. Science 287: 2262-2267, 2000 6) Toni N et al. Nature 402: 421-425, 1999 7) Frankland PW et al. Nature 411: 309-313, 2001 8) Kasahara J et al. J Biol Chem 276: 24044-24050, 2001
© Copyright 2026 Paperzz