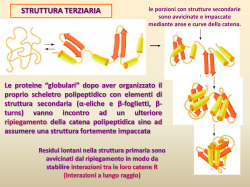
L’analisi di singoli ribosomi mentre traducono un mRNA rivelano che la traduzione avviene attraverso cicli di traslocazione e successive pause con una durata media di 2,8 sec La traslocazione di un codone si verifica in meno di 0,1 sec La lunghezza delle pause dipende dalla struttura secondaria dell’mRNA; la forza applicata dal ribosoma destabilizza la struttura secondaria e diminuisce la durata della pausa, ma non influisce sui tempi della traslocazione Following translation by single ribosomes one codon at a time - Nature 452, 598-603 (3 April 2008) doi:10.1038/nature06716 - Jin-Der Wen, Laura Lancaster, Courtney Hodges, Ana-Carolina Zeri, Shige H. Yoshimura, Harry F. Noller, Carlos Bustamante & Ignacio Tinoco I ribosomi in stallo possono essere dissociati in subunità, il peptidil-tRNA nella subunità 60S viene ubiquitinato da Ltn1 e Cdc48, il tRNA viene staccato e la proteina viene inviata ai proteasomi per la degradazione Questi meccanismi possono offrire una certa protezione contro gli effetti deleteri che derivano dall'accumulo di proteine mal ripiegate e aggregate Il sistema RQC elimina le proteine aberranti che si formano dalla traduzione di mRNA aberranti Protein degradation: Ensuring quality at the ribosome - Nature Reviews Molecular Cell Biology 14, 1 (January 2013) | doi:10.1038/nrm3499 - Katharine H. Wrighton Protein quality control systems associated with no-go and nonstop mRNA surveillance in yeast - Genes Cells. 2013 Nov 21. doi: 10.1111/gtc.12106. [Epub ahead of print] - Matsuda R, Ikeuchi K, Nomura S, Inada T. Molecular biology: RNA lost in translation - David Tollervey - Nature 440, 425-426(23 March 2006) - doi:10.1038/440425a La traduzione del ribosoma avviene con un meccanismo complesso che coinvolge l'interazione coordinata di diversi acidi nucleici e proteine L'accumulo intracellulare di proteine mal ripiegate in inclusioni amiloidi, tipico di molte malattie neurodegenerative è una conseguenza del fallimento dei meccanismi di controllo della qualità delle proteine sintetizzate Real-time tRNA transit on single translating ribosomes at codon resolution - Nature 464, 1012-1017 (15 April 2010) | doi:10.1038/nature08925 – Uemura et al. Misfolded proteins partition between two distinct quality control compartments - Nature 454, 1088-1095 (28 August 2008) - doi:10.1038/nature07195 - Daniel Kaganovich, Ron Kopito & Judith Frydman Le proteine del citosol che sono non funzionanti o danneggiate verranno inviate alla distruzione nei proteasomi Il particolare che accomuna le proteine non funzionanti è quello di avere regioni idrofobiche esposte sulla superficie Il processo di demolizione chiama in causa l’ubiquitina e gli enzimi che la legano alle proteine da demolire Le funzioni chaperon della Hsp70 (heat shock protein 70) dipendono dalle interazioni tra il dominio che lega ATP ed il dominio che lega la proteina substrato La presenza di ATP legato regola l'affinità per le proteine, le proteine che vengono legate provocano l’idrolisi dell’ATP Le Hsp70 esistono in almeno due stati conformazionali per legare le proteine mal avvolte e per assisterle nell’assumere la corretta conformazione Hsp70 chaperone dynamics and molecular mechanism - Trends Biochem Sci. 2013 Oct;38(10):507-14. doi: 10.1016/j.tibs.2013.08.001 - Mayer MP. Una proteina mal avvolta o danneggiata presenta regioni idrofobiche esterne La proteina Hsp70 lega le proteine mal avvolte ed idrolizza ATP per assisterle nel raggiungere la conformazione corretta senza regioni idrofobiche esterne Hsp70 chaperone dynamics and molecular mechanism - Trends Biochem Sci. 2013 Oct;38(10):507-14. doi: 10.1016/j.tibs.2013.08.001 - Mayer MP. La Hsp70 può intervenire per tentare di districare aggregati proteici indesiderati, fornendo alla proteina rilasciata la possibilità di ripiegarsi in modo corretto Diverse Hsp70s possono lavorare sullo stesso aggregato proteico contemporaneamente Hsp70 chaperone dynamics and molecular mechanism - Trends Biochem Sci. 2013 Oct;38(10):507-14. doi: 10.1016/j.tibs.2013.08.001 - Mayer MP. Le proteine mal avvolte e danneggiate vengono sequestrate in due compartimenti intracellulari La ripartizione delle proteine nei due compartimenti per il controllo di qualità sembra dipendere dal loro status di ubiquitinazione e dallo stato di aggregazione Le proteine mal ripiegate ed ubiquinate si accumulano in un compartimento vicino al nucleo (juxtanuclear) dove si concentrano i proteasomi Le proteine che invece formano aggregati caotici sono sequestrate in strutture perivacuolari. La huntingtina ed i prioni sono preferenzialmente indirizzati al compartimento perivacuolare Real-time tRNA transit on single translating ribosomes at codon resolution - Nature 464, 1012-1017 (15 April 2010) | doi:10.1038/nature08925 – Uemura et al. Misfolded proteins partition between two distinct quality control compartments - Nature 454, 1088-1095 (28 August 2008) - doi:10.1038/nature07195 - Daniel Kaganovich, Ron Kopito & Judith Frydman Il proteasoma è il principale macchinario proteolitico che si trova nel citoplasma e nel nucleo delle cellule eucariotiche Curr Opin Struct Biol. 2014 Feb;24:156-64. doi: 10.1016/j.sbi.2014.02.002. Epub 2014 Mar 14. Inobe & Matouschek - Paradigms of protein degradation by the proteasome La struttura molten globule espone gruppi idrofobici all’esterno Il pathway ubiquitina/proteasoma degrada migliaia di proteine, sia importanti come fattori di trascrizione e regolatori del ciclo cellulare, oppure proteine mal ripiegate da eliminare per impedire la loro aggregazione e tossicità. Una proteina poliubiquitinata viene comunemente indirizzata al proteasoma. Nel proteasoma sono presenti recettori per l’ubiquitina che consentono di legare le proteine da demolire. Come mostrato nella parte a destra, le ATPasi all'interno del proteasoma srotolano la proteina da demolire e la traslocano nel nucleo 20S del proteasoma dove i siti proteolitici la riducono a piccoli peptidi che vengono rilasciati mentre l’ubiquitina viene normalmente riciclata. Origin and function of ubiquitin-like proteins - Mark Hochstrasser – Nature 458, 422-429(26 March 2009) - doi:10.1038/nature07958 Progetto Proteoma Umano Il genoma umano contiene circa 21 mila geni che codificano per proteine Il trascrittoma umano contiene oltre 100 mila trascritti Il proteoma umano contiene oltre un milione di proteine Questo evidente aumento di complessità dipende sia dallo splicing alternativo che dalle modificazioni post-traduzionali delle proteine Le modificazioni post-traduzionali (PTM = glicosilazione, fosforilazione, acetilazione e metilazione, ecc.) delle proteine sono usate dalle cellule eucarioti per diversificare le funzioni delle proteine e coordinare dinamicamente le loro reti di signalling. Difettosa PTM è associata a numerosi disturbi dello sviluppo ed a patologie umane. Questo mette in evidenza l'importanza delle PTM nel mantenere in buona salute la cellula. Cell Res. 2014 Feb;24(2):143-60. doi: 10.1038/cr.2013.151. Epub 2013 Nov 12. Wang et al. - Protein posttranslational modifications and regulation of pluripotency in human stem cells Figura 1 – Nelle cellule eucariotiche le proteine possono essere modificate dopo la traduzione mediante una grande varietà di meccanismi sia reversibili che irreversibili. La struttura, la stabilità e la funzione delle proteine nelle cellule possono essere modificate dinamicamente dalla PTM. Nella figura sono rappresentati quattro tipi di PTM (glicosilazione, fosforilazione, acetilazione e metilazione). La N- e la O-glicosilazione delle proteine avviene nel RER e nell'apparato di Golgi Le vie del signalling regolate dalla fosforilazione di proteine sono coinvolte nello sviluppo embrionale e nella regolazione delle staminali pluripotenti. Molti recettori per fattori di crescita sulla membrana cellulare sono protein chinasi che, quando legano il ligando, si attivano ed avviano una cascata di fosforilazioni che regolano la traslocazione e l'attività di diversi fattori di trascrizione e quindi l'espressione dei geni per la pluripotenza o per il differenziamento. Queste vie di trasduzione del segnale sono altamente interattive tra loro e sono influenzate da altre proteine sulla membrana o nel citosol. Diverse protein fosfatasi (come PTEN e SHP2) controllano negativamente la fosforilazione proteica e giocano ruoli critici nella modulazione del signalling e nel differenziamento delle staminali pluripotenti.44 Le azioni antagoniste di HAT e HDAC sono necessari per regolare l'acetilazione di istoni e proteine non istoniche in molti tipi di cellule di mammifero e umane. Le HAT trasferiscono gruppi acetile sulle proteine target, le HDAC rimuovono i gruppi acetile. Lo stato di acetilazione degli istoni influisce sulla struttura della cromatina e determina l'accessibilità delle regioni dei promotori al macchinario trascrizionale e quindi l'attivazione dell'espressione genica. Nello stato pluripotente, le cellule hanno livelli più elevati di acetilazione degli istoni ed una maggiore accessibilità della cromatina per il macchinario di trascrizione. Nella cellula, le metilazioni di arginina e lisina sono catalizzate da metiltrasferasi differenti (PRMT e PKMT) Modificazioni post-traduzionali del core di istoni di un nucleosoma per acetilazione, metilazione e fosforilazione. Alcuni lisine (ad esempio, H3K4, H3K9 e H3K27) possono essere modificate sia per acetilazione che metilazione. La trimetilazione H3K4 e H3K27 origina una marcatura divalente della cromatina che si trova nei siti di inizio di trascrizione di molti geni coinvolti nel committement e nel differenziamento di hPSC indifferenziate. In generale, i geni indotti durante il differenziamento delle hPSC conservano la marcatura H3K4, mentre i geni silenziati durante il differenziamento conservano la marcatura H3K27. La fosforilazione di residui di amminoacidi (ad esempio, H3S10 e H3S28) in prossimità di quelle lisine potrebbe influenzare il loro switch metilazione-acetilazione. I filamenti intermedi (IF) sono strutture del citoscheletro e del nucleoscheletro che forniscono resistenza meccanica consentendo alle cellule di far fronte allo stress. I filamenti intermedi includono le lamine nucleari e varie proteine del citoplasma (cheratine, vimentina, desmina, neurofilamenti, ecc.) che sono funzionalmente regolate da modificazioni post-traduzionali (PTM). I progressi della proteomica evidenziano l'enorme complessità e le potenzialità di regolazione delle PTM delle proteine degli IF, tra cui fosforilazione, glicosilazione, sumoilazione, acetilazione e prenilazione. Nat Rev Mol Cell Biol. 2014 Mar;15(3):163-77. doi: 10.1038/nrm3753. Snider & Omary - Posttranslational modifications of intermediate filament proteins: mechanisms and functions. L’ubiquitinazione è un processo di modificazione post-traduzionale che è implicato nella regolazione della risposta immunitaria innata e adattativa. Sia l’ubiquitinazione che la deubiquitinazione giocano un ruolo cruciale non solo durante lo sviluppo del sistema immunitario, ma anche nell'orchestrazione della risposta immunitaria per garantire il corretto funzionamento dei diversi tipi di cellule che costituiscono il sistema immunitario. Conclusione - L'ubiquitinazione (e la deubiquitinazione) di proteine chiave nelle pathway di signalling dei recettori è essenziale per suscitare una risposta immunitaria adeguata ma limitata. Difetti di ubiquitinazione sono stati implicati nella patogenesi di una varietà di malattie, tra cui autoimmunità, autoinfiammazione ed immunodeficienza, sottolineando l'importanza della sua stretta regolazione e controllo. Siamo lontani dal comprendere pienamente questi processi al punto da poter intervenire terapeuticamente per il trattamento di malattie e condizioni patologiche in cui il sistema immunitario non agisce al massimo delle sue capacità, o entro i suoi limiti fisiologicamente definiti. EMBO Rep. 2014 Jan;15(1):28-45. doi: 10.1002/embr.201338025. Epub 2013 Dec 27. Zinngrebe et al. - Ubiquitin in the immune system Ubiquitina ed Ubiquitinazione L’ubiquitinazione è una modificazione post-traduzionale energiadipendente in cui la piccola proteina ubiquitina (appena 76 amminoacidi) viene legata covalentemente ad uno o più residui di lisina di una proteina substrato. Il processo di ubiquitinazione è una sequenza di tre eventi noti come attivazione, coniugazione e legatura, che coinvolge tre diversi enzimi: • E1, lega su di l’ubiquitina con un legame tioestere usando l’energia dell’idrolisi di ATP • E2 catalizza il trasferimento dell’ubiquitina da E1 alla cisteina presente nel suo sito attivo • E3 lega la glicina C-terminale dell’ubiquitina ad un residuo di lisina di una proteina bersaglio che può essere anche una ubiquitina, con formazione di un legame ubiquitinaubiquitina. L’ubiquitina ha sette residui di lisina e quindi ha la possibilità di formare sette diversi tipi di collegamenti, K6, K11, K27, K29, K33, K48 e K63. Ubiquitina, piccola proteina di 76 ammino acidi Legata con un legame isopeptidico (fra il gruppo amminico ε della lisina (K) ed il gruppo carbossilico della glicina 76 dell’ubiquitina Nelle catene di ubiquitina possono essere usate tante lisine interne: K6, K11, K27, K29, K33, K48, K63 Altra possibilità è il legame tra il gruppo amminico α della metionina N‐terminale (M1) ed il gruppo carbossilico della glicina 76 Esistono anche ulteriori possibilità di legame che consentono un complesso signalling in vari sistemi, come quello immunitario Il fatto che il genoma umano codifichi per solo due E1, ma almeno 38 E2 e più di 600 E3, rende il processo di ubiquitinazione molto vario e complesso, ma anche molto specifico. I diversi punti di legame dell’ubiquitina svolgono differenti funzioni fisiologiche: REAZIONI DI UBIQUITINAZIONE Ub E1 attiva l’Ubiquitina E3 lega una o più Ubiquitine alla Proteina da distruggere Ub E2 coniuga una o più Ubiquitine La malattia di Huntington è una malattia neurodegenerativa ereditaria dominante causata dall’espansione della tripletta CAG nel gene della huntingtina, caratterizzata dall’accumulo di aggregati di proteine con la coda di poliglutammine nei neuroni L’arricchimento anomalo di inclusioni con proteine ubiquitinate, caratteristica diagnostica comune ad altre malattie neurodegenerative come Alzheimer e Parkinson, suggerisce che la disfunzione nel metabolismo dell’ubiquitina può contribuire alla patogenesi di queste malattie Poiché la modificazione delle proteine per aggiunta di catene di poliubiquitina regola molti processi cellulari essenziali, tra cui la degradazione delle proteine, il ciclo cellulare, la trascrizione, la riparazione del DNA ed il traffico di vescicole, la perturbazione di questo meccanismo di segnalazione rischia di avere gravi conseguenze per la funzione neuronale e la sopravvivenza Global changes to the ubiquitin system in Huntington's disease - Nature 448, 704-708 (9 August 2007) doi:10.1038/nature06022 - Bennett et al. La propagazione cellula-cellula di proteine mal ripiegate si verifica in tutte le malattie neurodegenerative? Una recente ricerca dimostra la propagazione della huntingtina mutante in colture di sezioni Lug 2014 sottili di cervello in vivo, estendendo in tal modo il processo di propagazione cellulacellula di proteine mal ripiegate anche alla malattia di Huntington. Nel complesso, i nostri risultati suggeriscono che la propagazione transneuronale della huntingtina mutante potrebbe essere costituire un importante e sottovalutato contributo alla fisiopatologia della malattia di Huntington Nat Neurosci. 2014 Aug;17(8):1064-72. doi: 10.1038/nn.3761. Epub 2014 Jul 13. Pecho-Vrieseling et al. Transneuronal propagation of mutant huntingtin contributes to non-cell autonomous pathology in neurons •Legare ad una lisina di una proteina una singola ubiquitina oppure una catena di ubiquitine (monoubiquitazione e poliubiquitinazione) permette l’interazione con un sottoinsieme di domini (UBD) in proteine bersaglio e con enzimi deubiquitinanti. •Segnali che provengono dal contesto, come la localizzazione cellulare in cui si verifica la modificazione, possono contribuire a determinare le conseguenze fisiologiche della mono- e poli-ubiquitinazione. Origin and function of ubiquitin-like proteins - Mark Hochstrasser – Nature 458, 422-429(26 March 2009) doi:10.1038/nature07958 •Catene di ubiquitina legate alla lisina 48 sono comunemente associate all’interazione con il proteasoma ed alla degradazione della proteina, ma possono anche regolare la trascrizione nel nucleo. •Catene di ubiquitina legate alla lisina 63 intervengono nella trasduzione del segnale, nel traffico di vescicole, nell’endocitosi e nella riparazione del DNA. •La monoubiquitiazione specifica regola quasi tutti questi processi e la trascrizione genica modulata dalla cromatina. Origin and function of ubiquitin-like proteins - Mark Hochstrasser – Nature 458, 422-429(26 March 2009) doi:10.1038/nature07958 L’ubiquitinazione è una modificazione post-traduzionale cruciale che regola molti processi cellulari negli eucarioti, incluse le risposte infiammatorie, il traffico endocitico ed il ciclo cellulare. L’ubiquitinazione ha anche un ruolo centrale nella modulazione dei sistemi di difesa degli eucarioti. Ci sono crescenti evidenze sul fatto che molti batteri patogeni hanno imparato a sfruttare a loro beneficio i sistemi che usano ubiquitina. In questa review sono descritte le modalità con le quali batteri patogeni umani colpiscono l’ubiquitinazione per sovvertire e manipolare I sistemi di difesa dell’ospite, speso utilizzando il mimetismo molecolare di proteine batteriche effettrici secrete. Queste strategie consentono ai batteri patogeni di massimizzare la funzione effettrice e di ottenere nutrienti, promuovendo così la proliferazione dei batteri. Mag 2014
© Copyright 2026 Paperzz