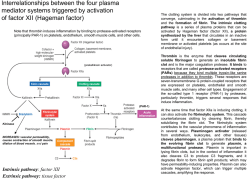
Istologia 16 – Sangue (parte 3), Emopoiesi, Gruppi sanguigni Piastrine (trombociti) Sono meno numerose dei globuli rosso (in un microlitro di sangue sono circa 200 400.000). Si tratta di frammenti cellulari, derivati dal distacco di pezzi del megacariocita, la loro cellula madre (che si trova nel midollo osseo). I trombociti hanno una vita breve: 5-9 giorni. Hanno una forma di lente biconvessa, con un diametro che può variare da 2 a 4 m. Le piastrine sono formate due parti: Cromomero. È la porzione più interna e centrale. È maggiormente colorabile ed è anche detta granomero, in quanto contiene diversi tipi di granuli (perlopiù basofili). In particolare, sono presenti: Granuli (80%). Sono piuttosto grandi. Contengono fattori della coagulazione (tra cui il fibrinogeno) e fattori di crescita (ad esempio PDGF) Granuli densi o δ. Sono più piccoli e meno abbondanti, ma più elettrondensi. Contengono calcio, ADP, fattori della coagulazione e la serotonina (o 5-idrossitriptamina), che ha una spiccata funzione vasocostrittrice (antagonista dell’istamina). Granuli lisosomiali o λ: contengono idrolasi lisosomiali. Ialomero. È la porzione di citoplasma più periferica, più chiara rispetto al cromomero Le piastrine, inoltre, producono anche le prostaglandine. Si tratta di mediatori chimici che hanno varie funzioni, tra cui quella dell’innesco della emostasi (vedi dopo) La funzione principale dei trombociti è quella di tamponare le ferite. Emostasi L’emostasi è l’arresto di un’emorraggia, cioè della fuoriuscita di sangue da un vaso a seguito di una lesione. Essa si attua con una serie di meccanismi. In condizioni normali la piastrina scorre nel torrente circolatorio liberando in continuazione prostaglandina G2; questa viene captata dall’endotelio e convertita (da un enzima endoteliale chiamato prostaciclinsintetasi) nella prostaglandina I2 detta anche prostaciclina che è un potentissimo fattore di inibizione delle piastrine. In condizioni patologiche, si attuano i seguenti meccanismi: 1. Nella zona in cui l’endotelio è venuto a mancare, non si ha la conversione della prostaglandina G2 a prostaciclina. La prostaglandina G2 in eccesso viene captata dalla stessa piastrina che la trasforma in trombossano, un’ altra prostaglandina che attiva le piastrine. 2. Contemporaneamente, nella zona in cui manca l’endotelio, emerge il connettivo della lamina propria. Le fibre collagene in esso contenute interagiscono con le piastrine e le attivano. 3. All’attivazione piastrinica fa seguito l’adesione piastrinica. Le piastrine, tramite trasformazioni della propria struttura citoscheletrica, si modificano ed assumono un aspetto estremamente irregolare che consente alle varie piastrine di incastrarsi reciprocamente nella zona priva di endotelio. Istologia 17 – Sangue (parte 3) e Gruppi sanguigni 1 4. Contemporaneamente, avviene l’esocitosi massiccia dei granuli piastrinici. In particolare, vengono secreti il fibrinogeno e la serotonina (5-idrossitriptamina), che determina una diminuzione del flusso sanguigno proprio a livello del vaso in cui si ha un’emorragia. 5. Man mano che ciò avviene, sopra il primo strato di piastrine se ne formano altri. Questo processo prende il nome di aggregazione piastrinica. Al termine del processo, si viene a creare un primo “tappo”, detto trombo bianco o trombo piastrinico, nel varco da cui fuoriusciva il sangue. 6. L’aggregazione piastrinica porta all’innesco di una seconda fase dell’emostasi, la coagulazione. La membrana della piastrina contiene un fosfolipide particolare, il fattore piastrinico 3 (PF3). Questo interagisce con la tromboplastina, una molecola rilasciata dai tessuti danneggiati. PF3, tromboplastina ed altri fattori formano l’attivatore protrombinico, vero e proprio interruttore del processo coagulativo. 7. L’attivatore protrombinico converte la protrombina, normalmente presente nel sangue, in trombina. Questa esplica azione proteolitica sul fibrinogeno, che, da essere solubile e circolare normalmente nel sangue, viene trasformato in una proteina filamentosa insolubile, la fibrina. 8. La fibrina neoformata si deposita e si stratifica formando un fitto reticolato proteico al di sopra del trombo piastrinico. In questa precipitazione tumultuosa della fibrina tutto ciò che c’è dintorno rimane intrappolato in questa rete proteica, comprese alcune cellule del sangue come i globuli rossi: in questo modo si forma, al di sopra del trombo bianco, il cosiddetto trombo rosso o coagulo che è molto più resistente del trombo piastrinico. 9. Nelle zone circostanti il coagulo, le cellule endoteliali proliferano, passano sopra il coagulo e rinsaldano la continuità della parete endoteliale. Successivamente, tramite la produzione di enzimi proteolitici, esse digeriscono tutta la zona del tappo piastrinico e del coagulo. In questo modo, viene nuovamente ricostituita la struttura fisiologica del vaso sanguigno. Un’anomala attivazione delle piastrine al di fuori di questa necessita può portare a fenomeni di trombosi, cioè alla formazione di trombi e coaguli all’interno dei vasi sanguigni senza che ce ne sia la necessità. La potenza della fibrina come meccanismo di tamponamento delle lesioni dei vasi sanguigni è stata utilizzata anche in chirurgia per le colle fibriniche. Sono formate da fibrina sintetizzata in laboratorio o isolata dal plasma degli animali da macello e sono usate al posto dei punti chirurgici. Emopoiesi E’ il processo che porta alla formazione di tutti gli elementi corpuscolati del sangue. Nell’individuo adulto, avviene nel midollo osseo. Questo, tuttavia, ha diversi gradi di attività nel corso della vita. Nell’individuo giovane è ricco di cellule progenitrici del sangue, soprattutto degli eritrociti; per questo viene chiamato midollo rosso. Nell’adulto e soprattutto nell’anziano, invece, l’attività emopoietica diminuisce e le cellule emopoietiche vengono via via rimpiazzate da tessuto adiposo; si forma così il midollo giallo. Le vie emopoietiche sono complesse e coinvolgono diversi tipi di cellule, che hanno come evoluzione finale la cellula matura. Tuttavia, possiamo riconoscere due tipi di cellule da cui partono le due vie emopoietiche. Esse sono: Progenitore comune mieloide. Dà origine a tutti gli elementi corpuscolati del sangue, tranne i linfociti. Dal progenitore comune mieloide originano tre varianti cellulari: Megacarioblasta, da cui avrà origine il megacariocita (e quindi le piastrine) Proeritroblasta, da cui avranno origine gli eritrociti. Questo processo si chiama eritrocitopoiesi. Di tutte le cellule di questa via emopoietica, rivestono un ruolo importate i reticolociti. Si tratta di eritrociti giovani, appena immessi in circolo. Pur essendo privi di nucleo, contengono ancora dei ribosomi attraverso cui ultimano la produzione di emoglobina. Nel giro di 24h diventano eritrociti maturi. Istologia 17 – Sangue (parte 3) e Gruppi sanguigni 2 Mieloblasto, da cui hanno origine tutti i granulociti e i monociti. Il processo di formazione dei granulociti si chiama granulocitopoiesi; il processo di formazione dei monociti si chiama monocitopoiesi. Il progenitore comune mieloide dà origine, infine, al mastocita. Tuttavia, la differenziazione del mastocita è separata da quella delle cellule del sangue. Progenitore comune linfoide. Mediante un processo noto come linfopoiesi dà origine sia ai piccoli che ai grandi linfociti Le cellule emopoietiche, quindi, possono essere separate a seconda del tipo di cellula a cui daranno origine; per questo, si parla di colony-forming units (CFU), veri e propri gruppi di cellule che danno origine a uno stesso tipo di cellule. Esistono, quindi, CFU-E (colony-forming units – erythrocytes), CFU-GM (colonyforming units – granulo-monocytes),… . Istologia 17 – Sangue (parte 3) e Gruppi sanguigni 3 Gruppi sanguigni Il gruppo sanguigno è una delle numerose caratteristiche di un individuo (come il colore degli occhi e dei capelli, etc) ed è determinato dai geni. I gruppi sanguigni vengono definiti da: Presenza o assenza di alcuni antigeni presenti sulla membrana cellulare dei globuli rossi: Alcuni anticorpi nel plasma sanguigno. Esistono diversi tipi di classificazione dei gruppi sanguigni (in base agli antigeni considerati). I più comuni (e più utilizzati) sono il sistema AB0 ed il sistema Rh. Sistema AB0 In base al sistema AB0, i soggetti possono essere: Gruppo A. Sui globuli rossi è presente l’antigene A, mentre nel plasma si riscontra la presenza degli anticorpi anti-B. Gruppo B. Sui globuli rossi è presente l'antigene B, mentre nel plasma si riscontra la presenza degli anticorpi anti-A. Gruppo AB. Sui globuli rossi sono presenti entrambi gli antigeni (sia A che B), mentre nel plasma non si riscontra la presenza né di anticorpi anti-A, né di anticorpi anti-B. Gruppo 0. Sui globuli rossi sono assenti antigeni, mentre il plasma sanguigno possiede sia gli anticorpi anti-A che quelli anti-B. Sistema Rh In base al sistema Rh, i soggetti possono essere: Rh positivi (Rh+). Sul globulo rosso è presente una glicoproteina, il fattore Rh. Rh negativi (Rh-). Sono Rh- (Rh-negativi) i soggetti che non hanno il fattore Rh. L’85% della popolazione è Rh+ ed il 15% è Rh-. Diversamente dal sistema AB0, nel caso del sistema Rh il plasma sanguigno dei soggetti Rhnon contiene anticorpi anti-Rh. Questo perché il fattore Rh è assente nei soggetti Rh- (che quindi non possono sviluppare gli anticorpi). Istologia 17 – Sangue (parte 3) e Gruppi sanguigni 4 Compatibilità dei gruppi sanguigni Gli anticorpi e gli antigeni del sangue pongono delle barriere alle trasfusioni di sangue tra persone di gruppi diversi. Infatti, se un soggetto ad esempio possiede degli anticorpi anti-A (ed è quindi B), non potrà ricevere sangue da un soggetto di gruppo A (con antigene A e anticorpi antiB). Infatti, da un lato gli anticorpi anti-A riconoscerebbero gli eritrociti con l’antigene A e dell’altro gli anticorpi anti-B riconoscerebbero gli eritrociti con l’antigene B, causandone l’eliminazione. La situazione si complica ulteriormente se introduciamo il sistema Rh. Vediamo cosa accade caso per caso. Soggetto 0 RhPuò donare a tutti. Infatti, non c’è nessuno dei due antigeni sui globuli rossi ed il fattore Rh è assente. Può ricevere solo da soggetti 0 Rh-. Essendo 0, infatti, contiene sia anti-A che anti-B; inoltre, se venisse in contatto con il fattore Rh, svilupperebbe degli anticorpi contro di esso. Soggetto 0 Rh+ Può donare a tutti i gruppi, purchè i soggetti siano Rh+. Il sangue di questo soggetto, infatti, svilupperebbe anti-Rh in soggetti Rh-. Può ricevere solo da soggetti 0, sia Rh- che Rh+. Infatti, non possiede anti-Rh; quindi l’ingresso del sangue Rh- sarebbe del tutto innocuo. Soggetto A RhPuò donare solo ad A e AB (sia Rh+ che Rh-). Può ricevere da soggetti 0 Rh- e A Rh-. Soggetto A Rh+ Può donare solo ad A e AB (solo Rh+). Può ricevere da soggetti 0 (Rh+ e Rh-) e A (Rh+ e Rh-). Soggetto B RhPuò donare solo a B e AB (sia Rh+ che Rh-). Può ricevere da soggetti 0 Rh- e B Rh-. Soggetto B Rh+ Può donare solo a B Rh + e AB Rh +. Può ricevere da soggetti 0 (Rh + e Rh-) e B (Rh + e Rh-). Soggetto AB RhPuò donare solo a AB (Rh + e Rh -). Può ricevere da tutti i soggetti (dato che non ha né anti-A né anti-B), ma purchè siano Rh-. Soggetto AB Rh+ Può donare solo a AB Rh. Può ricevere da tutti i soggetti. Istologia 17 – Sangue (parte 3) e Gruppi sanguigni 5 Ereditarietà del gruppo sanguigno Il gruppo sanguigno, come tutti i caratteri dell’essere umano, è determinato da due alleli presenti su due cromosomi omologhi. Gli alleli sono forme diverse di uno stesso gene e, insieme, codificano per un’informazione; l’informazione finale, quindi, è sempre il risultato di due “messaggi” (i due alleli, appunto). I due alleli assieme formano il genotipo di quel carattere; ciò che invece si manifesta è il fenotipo. Nel sistema AB0 esistono tre geni (A,B e 0) che sono responsabili della trasmissione del gruppo sanguigno. I geni A e B sono geni codominanti, cioè si esprimono entrambi se sono presenti assieme. Il gene 0 è un gene recessivo, cioè la sue espressione è “coperta” da quella dell’altro gene. Nel sistema Rh i geni sono due: D (dominante e responsabile dell'Rh+) e d (recessivo: Rh-). Fenotipo Bombay Si tratta di un fenotipo raro, il cui nome deriva dal fatto che venne riscontrato per la prima volta in una famiglia indiana di Bombay. Sia l’antigene A che l’antigene B si impiantano sulla membrana dei globuli rossi grazie ad un supporto, detto supporto H. Questo viene prodotto se gli alleli dell’individuo sono del tipo HH o Hh. Se, invece, il soggetto è hh, allora non si ha la produzione del supporto H e si manifesta il fenotipo Bombay. Le persone con il fenotipo Bombay non possono produrre il supporto H; per cui, pur avendo i geni per produrre gli antigeni A e B, non li produce (o meglio … questi non si formano sulla superficie dei globuli rossi, in quanto mancano del supporto). In conseguenza di ciò, egli svilupperà gli anticorpi anti-A e anti-B (dovuti all’assenza di questi antigeni) e produrrà anche gli anticorpi anti-H (cioè diretti verso il supporto di sostegno degli antigeni A e B). Fenotipicamente, quindi, saranno soggetti di tipo 0 (assenza di A e B e presenza di anti-A e anti-B). Genotipicamente, però, potranno possedere combinazioni varie (AA, A0, AB, BB, B0). Quando un soggetto con fenotipo Bombay si incrocia con un soggetto non Bombay, produrrà tutte le combinazioni possibili derivanti dai suoi alleli e da quelli del partner. Istologia 17 – Sangue (parte 3) e Gruppi sanguigni 6 Malattia emolitica del neonato Se una madre Rh- concepisce un figlio Rh+, le emazie del figlio passano nella madre (al momento del parto) e la madre si immunizza, formando anticorpi anti-Rh. Alla seconda gravidanza, se il figlio è di nuovo Rh+, gli anticorpi anti-Rh passano la placenta e distruggono i globuli rossi del figlio, con conseguente anemia e possibile morte del feto (malattia emolitica del neonato). Tutto questo può essere prevenuto iniettando, entro 72 ore dal primo parto, anticorpi anti-Rh alla madre. Questi anticorpi bloccano e distruggono eventuali globuli rossi fetali passati nel sangue materno, per cui il sangue materno non svilupperà anticorpi anti-Rh che ad una successiva gravidanza potrebbero attaccare i globuli rossi del figlio. Per lo stesso motivo, soggetti Rh+ non possono donare sangue a soggetti Rh- perché se questi ultimi sono stati precedentemente sensibilizzati con sangue Rh+ (precedenti trasfusioni o somministrazione di derivati del sangue, etc) potrebbe scatenarsi una reazione da incompatibilità dovuta agli anticorpi sviluppati con la precedente sensibilizzazione. Istologia 17 – Sangue (parte 3) e Gruppi sanguigni 7
© Copyright 2026 Paperzz