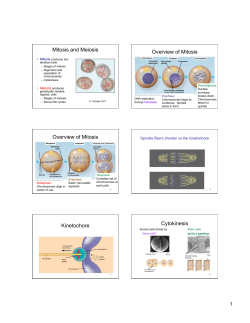
j Raptor Res. 27(4):196-202 ¸ 1993 The Raptor ResearchFoundation, Inc. CHROMOSOMES OF FIVE BUTEONINE NORTH HAWKS AMERICAN SHEILAM. SCHMUTZ,JANE S. MOKER AND TRACEYD. THUE Departmentof Animal and PoultryScience,Universityof Saskatchewan, Saskatoon,SK, Canada S7N OWO ABSTRACT.--Five species of buteoninehawkswere karyotypedfor the first time, includingthe Swainson's Hawk (Buteoswainsoni),Gray Hawk (B. nitidus),FerruginousHawk (B. regalis),Harris' Hawk (Parabuteo unicinctus),and Broad-winged Hawk (B. platypterus).All five specieshave 68 chromosomes.The karyotypesof the first four speciesappear to be very similar to eachother and to other buteosreported previouslyin the literature, differing in the shapeof only one chromosome.The Broad-wingedHawk karyotypediffers in the morphologyof severalchromosomes. Cromosomas de cincoespecies norteamericanas de halconesdel g•nero Buteo RESUMEN.--Porprimera vez han sido examinadosy caracterizadoslos cariotiposde cincohalconesdel g6neroButeo(Aguililla Migratoria Mayor Buteoswainsoni, Aguililla Gris B. nitidus,Aguililla Real B. regalis,Aguililla Rojinegra Parabuteounicinctus,y Aguililla Migratoria Menor B. platypterus).Todas las cinco especiestienen 68 cromosomas.Parece ser que los cariotipos de halconesde las cuatro primeras especies(B. swainsoni, B. nitidus,B. regalis,y Parabuteounicinctus)sonmuy similaresentre ellos,asl como lo sonconlosdeotroshalcones deesteg6nero ya referidos en literaturaanterior.l•stossediferencian s61opor la forma de s61oun cromosoma,mientrasque el Aguililla Migratoria Menor difiere por la morfologlade varioscromosomas. [Traducci6n de Eudoxio Paredes-Ruiz] Chromosome analysishasbeenusedto studyphy- pus; Bulatova1977), the RoadsideHawk (B. maglogeneticrelationshipsin severalspecies,such as nirostris;de Lucca 1983), and the White-tailed Hawk falcons (Schmutz and Oliphant 1987) and owls (B. albicaudatus;de Lucca 1985). (Schmutz and Moker 1991). In our view, cytogeneticsis well suitedto cladisticanalysisbecausethe MATERIALS AND METHODS phylogenies are basedon centricfusions(RobertThe blood sample from the Ferruginous Hawk was sonian translocations)and inversions(White 1973, Hsu 1979), bothof whichalsoimpair fertility (Therman 1980, Diedrichet al. 1983,Lippman-Hand and collectedin Alberta, Canada. Bloodsamplesfrom the Harris' and four Gray Hawks were collectedin Arizona. The bloodsamplesfrom the Swainson'sHawk and the Broadwinged Hawk were collectedfrom birds in captivityfor Vekemans1983), part of the classicaldefinitionof rehabilitationat the Western Collegeof Veterinary Medspeciesseparation. icine after injury near Saskatoon,Saskatchewan,Canada. Lymphocytecultureswere establishedfrom 0.5 ml of In an ongoingattemptto collectcytogenetic data on raptorsfor this purpose,we havekaryotypedfive whole blood and chromosomesprepared as describedby Schmutzand Oliphant (1987). Karyotypeswere basedon species of buteonine hawksnot previously reported. the bestmetaphase(s)obtainedon completecells.Several The five species of hawksare the Swainson's Hawk cellswere photographedand the chromosomesarranged (Buteoswainsoni), Gray Hawk (B. nitidus),Ferru- Comparativestudiesof karyotypesnot donein our own ginousHawk (B. regalis),Harris' Hawk (Parabuteo lab are baseduponfiguresin the originalpublishedpapers unicinctus),and Broad-wingedHawk (B. platyp- when availableor photocopiesthrough interlibrary loan. terus). Prior to our study,fiveotherspecies of buteoshad beenkaryotyped.Thesewerethe CommonBuzzard RESULTS Although most bird specieshave a large number (B. buteo;RenzoniandVegni-Talluri 1966, De Boer of microchromosomesand few macrochromosomes, 1976), the Red-tailed Hawk (B. jamaicensis;Shoff- the hawksandeaglesare atypicalin that few of their ner 1974, Pape and Ogasawara 1978, Stock and chromosomes could be called microchromosomes. The Worthen 1980), the Rough-leggedHawk (B. lago- Swainson'sHawk (Fig. 1), Gray Hawk (Fig. 2), 196 DECEMBER 1993 BUTEO CHROMOSOMES 197 Figure 1. The karyotypeof a male Swainson'sHawk (Buteoswainsoni)with an arrow marking chromosome7, the variable chromosomewhich is metacentricin this species. FerruginousHawk (Fig. 3), and Harris' Hawk (Fig. 4) each had 68 chromosomesincluding one large metacentricpair (the sex chromosomes), five large submetacentric pairs, six medium-sizedmetacentric pairs, eight small metacentricpairs, and 13 small acrocentricpairs, and one pair which was variable in morphology.This variable chromosomewas either a large submetacentricor large acrocentric.It was the seventhlargest by size and therefore we refer to it as chromosome7. Traditionally chromosomesare groupedby centromerepositionand thensecondarily by size.We havedecidedto arrange the chromosomes for easeof comparisonamongthe species,in the traditional order for the $wainson's The Broad-winged hawk differed from the other four hawks we studied although the total number remained 68 (Fig. 5). The largest pair was again metacentricand we presume it to be the sex chromosomepair. However, therewere only 16 submetacentric or metacentricpairs in total as opposedto 20-21 pairs. We subdivided the buteonine hawks into two ma- jor groups basedon the different morphologiesof chromosome 7 (Fig. 6) and placedB. platypterusin a third separate group since chromosomes16-20 differed.Althoughthe karyotypeof the Gray Hawk (Fig. 2) doesnot have an obviousacrocentricchromosome7, it is the clearest karyotype overall and Hawk and Common Buzzard but with chromosome thereforewe choseto publish it. Other karyotypes 7 out of its traditional place for the other species. we studied clearly indicate that chromosome7 is Other authors, such as Stock and Worthen (1980) acrocentricin this species.Less clear, is the morwho have publishedonly a singlespecieskaryotype, phologyof chromosome20 which also appearsacplace this chromosomewith the other acrocentrics rocentricin this photograph(Fig. 2). so it would be number 21 in their karyotype. We attemptedto include the other buteo species 198 SGHMUTZ ET AL. VOL. 27, NO. 4 Figure 2. The karyotypeof a male Gray Hawk (Buteonitidus)with an arrow markingchromosome 7, the variable chromosomewhich is acrocentricin this species. Figure3. The karyotypeof a femaleFerruginousHawk (Buteoregalis)with an arrow markingchromosome 7, the variable chromosomewhich is acrocentricin this species. z w Figure 4. The karyotypeof a female Harris' Hawk (Parabuteounicinctus)with an arrow marking chromosome7, the variable chromosome which is acrocentricin this species. z z Figure 5. The karyotypeof a femaleBroad-wingedHawk (B. platypterus).The karyotypeis arrangedto emphasize that chromosomes 16-20 in the middle row are acrocentricin this speciesas opposedto metacentricin the other buteos. 200 SCHMUTZET AL. I 2 3 4 5 VOL. 27, NO. 4 6 Buteo Buteo 7 8 9 10 11 12 X X X x x x 13 14 15 16 17 18 n Buteo jamaicensis Buteo iagopus 19 Buteo n nn 21 22 23 24 25 26 28 29 30 31 32 33 I 2 3 4 5 6 7 8 • •0 n X X X x • • x 13 14 15 16 17 18 19 21 22 23 24 25 26 27 29 30 31 32 33 2 $ 4 Parabuteo 27 Buteo 1 X X Xx 7 8 9 10 & x x 11 12 13; 14 n n n n 16 17 18 19 20 22 25 24 25 26, 29 30 31 32 33 21 28 swainsoni • • z x tl unicinctus Buteo buteo •2 5 aibicaudatus Buteo magnirostris n • regalis nitidus Buteo platypterus 27 Figure 6. A cladogram illustrating therelationships among thebuteonine hawks studied todate andcomputer drawn idiograms ofthethreekaryotypes observed, uponwhichthephylogeny isbased. studiedpreviously,althoughthe copiesof the kar- tailed Hawk (B. albicaudatus; de Lucca 1985) match yotypesthat we usedmakeour interpretation ten- the buteoswe did very closely.However,a minute tative.The karyotypes of theCommon Buzzard( B. metacentricwas described(De Boer 1976, Stockand buteo;De Boer 1976), the Red-tailedHawk (B. maicensis; Stockand Worthen 1980), and the White- Worthen 1980) which we cannotverify or dispute sincethe small chromosomesin our karyotypes are DECEMBER 1993 BUTEO CHROMOSOMES not elongatedenoughto determinethis.We worked with poorphotocopies of karyotypesof the RoughleggedHawk (B. lagopus;Bulatova1977) and the RoadsideHawk (B. magnirostris; de Lucca 1983) andcanonlysaythat thetoptwo rows(Fig. 6) agree in general with the karyotypesof the buteoswe studied. DISCUSSION Four speciesof buteoninehawks studiedhere for the first time have one of the sametwo karyotypes. This similarity supportsthe closetaxonomicrelationshipBrownandAmadon(1968) suggested among thesespecies. Further differentiationamongsubfamilies would needto be basedon other typesof data, suchasDNA sequence datawhichis currentlyunder investigationby Bob Sheehy(pers.comm.). The differencein morphologyof chromosome number7 is likely due to an inversion,in this case a pericentricinversion,one of the two cytogenetic changesthat typicallyoccurduring evolution(Hsu 1979). We are unableto saywhichis the "ancestral" shapeof chromosome 7 and thereforeour cladogram is "unrooted" and shows the two main branches as 201 mosomesto demonstratespecifictranslocationsand inversionsin mammals (Ward et al. 1991). Avian karyotypesare much more difficultto obtainthan mammaliankaryotypes,primarily dueto the lower numbersof dividing cells stimulatedby mitogens (Prus and Schmutz1987). Low numbersof dividing cells therefore make chromosomebanding difficult sincemost techniqueswork well on only a small proportion of these cells. Although chromosome banding is necessaryto determinewith precision which chromosomes are involved in translocations, bandingis not necessary to identifythe presenceor absenceof translocations,which is the key to phylogeneticcomparisons. In addition to a poor responseto mitogens,cytogeneticstudiesof birds have been further hamperedbecausefreshbloodsamplesor growingfeathers, capableof further cell divisionin culture, are required for analysis.Many rare and endangered birds exist in remote areas. Field collection of sam- plesfrom wild caughtbirds is occasionally possible but the logisticsof air transportto an appropriate lab within 24 hr is frequentlyimpossible.Despite suchdifficulties,chromosomeanalysiscan providea derivingat an equal point in time (Fig. 6). useful tool in taxonomic studiesand we encourage The Harris' Hawk is placedin the genus?ara- raptorbiologists to collaborate in suchstudies. buteoas opposedto Buteo;however, its karyotype ACKNOWLEDGMENTS would not distinguishit from mostof the other Buteo We thank JosefK. Schmutz,Bob Sheehy,and Collette species studiedto date.This suggests that the Harris' Wheler for collectingthe bloodsampleswhich madethis Hawk is indeed a close relative of the buteos. study possible.We acknowledgefunding from the Sas- In contrastthe karyotypeof the Broad-winged katchewanAgriculturalDevelopmentFund and the NaHawk showsfive chromosomes, numbers 16-20, that differ in morphologyfromthe restof thebuteos(Fig. 5). This would suggestthat it is the most divergent in the subfamily.It is more closelyrelated to the Swainsoh's Hawk and Common Buzzard than the tional Scienceand EngineeringResearchCouncil which supportsour laboratory. LITERATURE CITED BROWN,n. AND D. AMADON. 1968. Eagles, hawks and falcons of the world. McGraw Hill, New York, NY other buteos,basedon the morphologyof chromoU.S.A. some7 (Fig. 6). BULATOVA,N.S. 1977. Chromosomestructure and evoIn contrastto the similarityfoundamongthe bulution in birds. NAUKA Publishing House, Novositeos,the nine species of falconsstudiedto dateexhibit birsk, Russia. four differentkaryotypesin the genus(Schmutzand DE BOER,L.E.M. 1976. The somaticchromosomecomplementsof 16 speciesof Falconiformes(Aves)and the Oliphant 1987, Longmireet al. 1988). The primary karyoloõicalrelationships of the order.Genetica 46:77cytogeneticdifferencesamong the falconsare cen113. tric fusionsor translocationsas opposedto inversions, with the Merlin (Falco columbarius)having BE LUCCA, E.J. 1983. Somatic chromosomesof Falco sparverius andButeornagnirostris (Falconiformes: Aves). the greatestnumber of chromosomes (Longmire et Nucleus. 26:48-56. al. 1988) and therefore being the most diverged, 1985. Kar¾otypeand nucleolusorganizing followedbythe PeregrineFalcon(F. peregrinus) and regionsin somaticchromosomes of the White-tailed Prairie Falcon(F. mexicanus; SchmutzandOliphant Hawk Buteoalbicaudatus (Falconiformes:Aves). 1987). Cytogeneticists frequently use banding of chro- roblos 42:7-13. DIEDRICH, U., I. HANSMANN,D. JANKE,O. OPITZ AND 202 SGHMUTZ ET AL. H.-D. PROBECK. 1983. Chromosome anomalies in 136 coupleswith a historyof recurrentabortions.Hum. VOL. 27, NO. 4 SCHMUTZ,S.M. ANDJ.S. MOKER. 1991. A cytogenetic comparisonof someNorth American owl species.Genome 34:714-717. Genet. 65:48-52. Hsu, T.C. 1979. Human and mammalian cytogenetics --AND L.W. OLIPHANT.1987. A chromosome study of the Peregrine,Prairie, and Gyrfalconswith implian historicalperspective. Springer-Verlag,New York, NY U.S.A. cationsfor hybrids.J. Hered. 78:388-390. LIPPMAN-HAND,A. ANDM. VEKEMANS. 1983. Balanced SHOFFNER, R.N. 1974. Chromosomesof birds. Pages 223-287 in H. Busch (ED.), The cell nucleus. Acatranslocationsamongcoupleswith two or more spondemic Press, New York, NY U.S.A. taneousabortions:are malesand femalesequallylikely to be carriers? Hum. Genet. 63:252-257. LONGMIRE, J.L., A.K. LEWIS, N.C. BROWN, J.M. BUCKINGHAM, L.M. CLARK, M.D. JONES, L.J. MEINCKE,J. MEYNE, R.L. RATLIFF,F.A. RAY, R.P. WAGNER AND R.K. MOYZIS. 1988. Isolation and mo- lecular characterizationof a highly polymorphiccentromerictandemrepeatin the family Falconidae.Genomics 2:14-24. PAPE,K.L. AND F.X. OGASAWARA.1978. Studies on the karyotypeof the Red-tailedHawk. RaptorRes.12:2834. STOCK,A.D. ^ND G.L. WORTHEN. 1980. Identification of the sexchromosomes of the Red-tailed Hawk (Buteo jamaicensis)by C- and G-banding.RaptorRes. 14:6568. THERMAN, E. 1980. Human chromosomesstructure, behavior, effects. Springer-Verlag, New York, NY U.S.A. WARD,O.J., A.S. GRAPHODATSKY, D.H. WURSTER-HILL, V.R. EREMINA,J.P. PARKAND (•. YU. 1991. Cytogeneticsof beavers:a case study of speciationby monobrachial centtic fusion. Genome 34:324-328. WHITE, M.J.D. 1973. Animal cytologyand evolution. CambridgeUniv. Press,Cambridge,U.K. ficiencyandaccuracy of surgicalandcytogenetic sexing in psittacines.Avian Dis. 31:420-424. PRUS,S.E. ANDS.M. SCHMUTZ. 1987. Comparative ef- RENZONI,A. AND M. VEGNI-TALLURI. 1966. The kar- yogramsof someFalconiformes and Strigiformes. Chromosoma 20:133-150. Received25 April 1993; accepted30 August 1993
© Copyright 2026 Paperzz