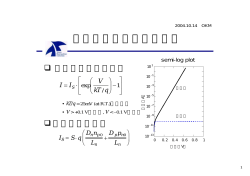
1 生体膜の電気伝導現象と膜電位 生体膜の機能として、(1) 物質の選択透過や能動輸送、 (2) 外界情報の受容、増幅、伝 播、 (3) エネルギー変換、 (4) 免疫応答、細胞間相互作用などがあげられる。 これらの 機能の多くは膜電位の変化と関係があり、またイオン透過と共役している。 従って、細 胞膜における電位発生の機構やイオン透過の仕組みを理解することは重要である。 1. 膜のイオン透過 1.1.電解質水溶液の導電現象 電解質溶液の電圧 V と電流 I の関係(電極面積 S ,間隔 d) I = GV = V R (ohm の法則)(G=コンダクタンス、 R=1/G) G =κ S d • (κ=導電率、抵抗率ρ=1/κ) 導電率κ:κ=zFcu (z,イオン価;F,ファラデー定数;c,イオン濃度; u,電気化学的移動度) 粘性流体(粘度η)中にある荷電粒子(イオン)(電荷 ze、半径 a) 粒子が電場によって受ける電気力:zeE 速度 v で動いている粒子が粘性流体から受ける流体抵抗力:ξv (ξ:摩擦係数、ξ=6πaη(球形イオンの場合の Stokes の式)) 速度一定 ⇒ 電気力=抵抗力、 zeE = ξv v=zeE /ξ=uE (u:電気化学的移動度, u = ze /ξ) 流束 J:単位時間、単位面積を通過する粒子の数 =粒子の濃度c×粒子の速度 v ⇒ J = cv = cuE 電流: I = zFJS = zFcuES = zFcuVS / d (電極面積 S、間隔 d) コンダクタンス G: G = I / V = zFcu S d = κ S d • ⇒ κ=zFcu 輸率 全電流=陰イオン電流+陽イオン電流(Kohlrausch のイオン独立移動の法則) I = I + + I − = (FV S d )( z + u + c + + z − u − c − ) 輸率 t+, t−:全電流に対する陰イオンや陽イオンの寄与 ( ) ( z + c + = − z − c − ⇒ t + = I + I = u + u + − u − 、 t − = I − I = − u − u + − u − ) 2 1.2. Nernst-Planck のイオン拡散式 溶質粒子 i の流束: Ji (mol/cm2s)=粒子の移動速度 vi (cm/s)×溶質濃度 ci (mol/cm3) ⇒ J i = ci vi 移動速度 vi ∝溶質粒子に作用する駆動力 fi ⇒ J i = ci ω i f i (ωi:モル移動度、molar mobility) ~ / dx 駆動力 fi ∝ 電気化学ポテンシャルの勾配 dµ ~ / dx ) ⇒ J i = − ci ω i ( dµ (Nernst-Planck) (正の流束:電気化学ポテンシャル高から低側への移動) µ~i = µ i0 + z i Fφ + RT ln ci ( φ :電位) ( µ i0 は x に依存しなとする) ⎧ ⇒ J i = −ω i ci ⎨ RT ⎩ d ln ci z F dφ ⎫ dφ ⎫ ⎧ dci + zi F + ci i ⎬ = − Di ⎨ ⎬ dx dx ⎭ RT dx ⎭ ⎩ dx (拡散定数 D=ωiRT) • Nernst-Planck 積分形 ⎧d ⎡ ⎛ z Fφ ⎞ ⎛ z Fφ ⎞⎤ ⎫ ⎛ z i Fφ ⎞ ⎟⎥ ⎬ ⎟ = − Di ⎨ ⎢ci exp⎜ i ⎟ ⇒ J i exp⎜ i dx RT ⎝ RT ⎠ ⎝ ⎠⎦ ⎭ ⎝ RT ⎠ ⎣ ⎩ 上式の両辺 × exp⎜ 定常状態 ⇒ Ji は場所によらず一定 膜相の x=o から x=d まで積分(DI は定数とする) ⇒ J i = [ − Di cid exp( z i F∆φ RT ) − cio ∫ d 0 exp( z i Fφ RT )dx ] (Nernst-Planck 積分形) ( cio , cid :x=o, d での濃度; φo、 φd :x=o, d での電位;△φ = φd − φo ) • µ i0 が x に依存する場合(膜内にエネルギ−障壁wI)の Nernst-Planck 微分形 ⎧ ⇒ J i = −ω i ci ⎨ RT ⎩ d ln ci d (wi + z i Fφ ) ⎫ c d (wi + z i Fφ ) ⎫ ⎧ dci + i + ⎬ ⎬ = − Di ⎨ dx dx dx ⎭ ⎭ ⎩ dx RT x=0, d で wio = wid = 0 ⇒ J i = [ − Di cid exp( z i F∆φ RT ) − ci0 ∫ d 0 ] exp(wi RT + z i Fφ RT )dx 3 1.3. Goldman の定電場の式 Nernst-Planck 式(微分方程式)を解くために 膜内に定電場を仮定 ∆ φ = φ i -φ o o 側 φo φo φi φi i側 ∆ φ = φ i -φ o 膜モデル(右図参照) co  ̄の付いた変数:膜内に関するパラメータ c 上付 o, i:膜の外側と内側のパラメータ o 膜 βi 膜電位は o 側基準 d ci βo ci 膜中での流束(エネルギー障壁の無い場合) 膜モデル Nernst-Planck の積分形 ⎡ ⎞ ⎛ z F∆φ ⎟⎟dx = − Di ⎢cii exp⎜⎜ i ⎠ ⎝ RT ⎣ d ⎛ z Fφ J i ∫ exp⎜⎜ i 0 ⎝ RT ⎞ o⎤ ⎟⎟ − ci ⎥ ⎠ ⎦ Goldman の仮定: 膜内の電位勾配一定 積分 ⇒ J i ∫ d 0 ⎛ RTd ⎛ z F∆φ x ⎞ ⎟⎟dx = J i ⎜⎜ exp⎜⎜ i ⎝ RTd ⎠ ⎝ z i F∆φ ⇒ J i = − • ⎞ ⎡ ⎛ z i F∆φ ⎟⎟ ⎢exp⎜⎜ ⎠ ⎣ ⎝ RT Di z i F∆φ cii exp(z i F∆φ / RT ) − cio RTd exp(z i F∆φ / RT ) − 1 ⎞ ⎤ ⎟⎟ − 1⎥ ⎠ ⎦ (Goldman の式) 膜内のパラメーター( ∆φ 、 ci0 、 cii )は観測不可 ⇒ 膜の両溶液のパラメーター(観測可能量)で表す cio = β i cio , cii = β i cii 、 βi:膜と溶液間の分配係数(濃度に依らないと仮定) 膜/溶液の界面電位を膜両側で等しいと仮定 ( φ o − φ o = φ i − φ i ) ⇒ φ i − φ o = φ i − φ o = ∆φ Pi z i F∆φ cii exp( z i F∆φ / RT ) − cio ⇒ J i = − (Goldman-Hodgin-Katz (GHK)の式) RT exp( z i F∆φ / RT ) − 1 (PI:透過係数、 Pi = Di β d ) • GHK 式を用いた計算例 Ji と△φの関係(右下図) 一価カチオン(zi=1)、Pi=0.01cm/hr、 cio =100mM(o 側濃度) cii =10, 50, 100mM (i 側の濃度) 4 • cio = cii =cI のとき Ji J i = −ci Pi z i F∆φ / RT cio=100mM (Jj−△φ:直線関係) Pi=0.01 cm/hr 100 mM 50 mM 2 zi=1 Ii = ziFSJI (S 膜面積) 3 1 電流-電圧関係(ohm の法則) ( ) -100 I i = ci Pi z i2 F 2 S / RT ∆φ = ∆φ / Ri -50 -2 (膜抵抗、 Ri = RT / ci Pi z i2 F 2 S ) • ∆φ 100 50 10 mM 0 -1 -3 GHK式による流束の計算 cio > cii のとき Ji と△φの関係:整流性 o 側から i 側への流束 (負側) > i 側から o 側の流れ (正側) ( Ji=0 の膜電位: ∆φ = ( z i RT / F ) ln cio / cii ) (Nernst の平衡電位) im 1.4. 反応速度論的取り扱い o 側 δG om i 側 モデル:二つのエネルギー障壁(山)と イオンの結合サイト(谷) (右図) m ∆Gom ∆Gim ∆Gb 粒子の拡散⇒障壁を飛び越え移動 −∆φim 速度定数 kom (om ピーク⇒m 谷) kom = (k BT / h )exp(− ∆Gom / RT ) ∆φ mi −∆φmo δφ ∆φ=φi-φο om (kB、Boltzmann 定数;h、Planck 定数) ∆φ 膜に電場印加(膜に定電場を仮定) 二つのエネルギー障壁と一つの結合部 反応の速度定数 位をもつモデル ( * * k om (o→m) : k om = k om exp − zF∆φ om / RT ) (△φom:o 側から om ピークまでの電位差) * k mo * kmi ( * (m→o): k mo = k mo exp − zF∆φ mo / RT ( * (m→i): k mi = k mi exp − zF∆φ mi / RT ) ) 5 * kim ( * (i→m): k im = k im exp − zF∆φ im / RT ) * * o 側から膜内への流束 jom : J om = k om cio λ o − k mo cim λ m ( cio 、o 側の濃度; cim 、膜内の濃度;λ、一回のジャンプで障壁を飛び越える ことのできる粒子の存在する範囲) * m * i ci λ m − k im ci λi 膜内から i 側への流束 Jmi: J mi = k mi J i = J om = J mi (定常状態)、 cim λ m 消去、 λ = λ o = λ i [ ( ) ][ ( )( * * * * * * ⇒ J i = k om λ cio − k mo / k mi k im* / k om cii / 1 + k mo / k mi ( )] ) ⎡ zF ∆φ mo − ∆φ mi ⎤ k mo exp ⎢− ⎥ k mi RT ⎣ ⎦ mo ⎡ zF ∆φ − ∆φ mi ⎤ ⎛ ∆Gmo − ∆Gmi ⎞ = exp⎜ − ⎟ exp ⎢− ⎥ RT RT ⎝ ⎠ ⎣ ⎦ = exp(δG / RT ) exp( zFδϕ / RT ) * * k mo / k mi = ( (k * mo )( * * * / k mi k im / k om ) ) ( ) ⎡ zF ∆φ mo + ∆φ im − ∆φ mi − ∆φ om ⎤ = (k mo k im / k mi k om ) exp ⎢− ⎥ RT ⎣ ⎦ = (k mo k im / k mi k om ) exp[zF∆φ / RT ] [ ] * k om λ cio − cii (k mo k im / k mi k om ) exp( zF∆φ / RT ) ⇒ J i = 1 + exp(δG / RT ) exp( zFδφ / RT ) kmokim/kmikom=1 (o 側と i 側との間の化学的な自由エネルギー差を 0 とする) [ ] * k om λ cio − cii exp( zF∆ϕ / RT ) ⇒ J i = 1 + exp(δG / RT ) exp( zFδφ / RT ) 6 C 11.5 9.5 8.5 O H H O 9.5 A D C B O B A 0.5 0.5 C D O 0 -1.0 Na イオンチャンネルのバリアーと結合サイト 1.5.イオン透過に対するエネルギー障壁 イオン透過に対するエネルギー障壁のプロファイルを決める要素 (1)イオンを水相から油の膜相へ持ち込むのに必要なエネルギー Born エネルギー(半径 a、電気量q、媒質の誘電率ε): E B = • q2 8aπε 0 ε q=e, a=0.2 nm、ε=2(油相中)⇒ 約 70kT(41.7kcal/mol) ε=80(水相中)⇒ 約 18kT(1.05kcal/mol) 水相から油相にイオンを持ち込むにはイオン一個あたり 68kT の仕事必要 チャンネル:誘電率の高い水で満 たされた穴を膜に開ける キャリア:イオンを誘電率の高い 殻で覆う • エネルギー障壁の計算(右図) 厚さ 7 nm の油の膜 ⇒ 40.1 kcal/mole 半径 0.5 nm の穴を持つチャンネル ⇒ 6.7 kcal/mole キャリアー(殻の誘電率無限大) ⇒ 16.7 kcal/mole Energy of charging (kcal/mole) (2)イオンが透過するための工夫 41.7 40.1 26.7 ε=10 21.7 ε=5 16.7 ε=∞ 6.7 1.05 In H2O In hc Finite Pore Spherical ε=80 ε=2 slab r=5 Å Jacket r=5 Å d=7 nm エネルギー障壁の大きさ 7 イオンチャンネルのエネルギー障壁 (1) 膜と水相やチャンネル内の水相とチャンネル壁では誘電率の異なる相が接して いる。この界面にイオンが近づくとイオンの電場により、界面に分極を引き起 こす。イオンは水相にあるので、この場合界面にはイオンと同じ極性の電荷が 生ずることになり、イオンがチャンネル内部に進入するのを妨げるように働く。 (2) チャンネル壁に双極子や固定電荷がある場合:イオン−双極子、イオン−イオ ン相互作用 (3) 表面電荷による電気二重層 (4) イオンの水和、脱水和 グラミシジンチャンネルのエネルギ−障壁 グラミシジンA(ペプチド性の抗生物質)が脂質 Solvation energy 二分子膜につくるイオンチャンネル (生体イオンチャンネルのモデル) • チャンネル入り口の山は脱水和に対応 • 内部の小さい山と谷の連なりはカルボニル基 とカチオンとの相互作用(溶媒和と呼んでい Electrostatic energy る) • 近距離の相互作用+遠距離の静電相互作用(界 面分極に相当する)⇒全体のエネルギー障壁 のプロファイル グラミシジンチャンネル のエネルギ−障壁 8 2. 膜電位 膜に発生する起電力を膜電位と呼ぶ。 膜電位は (1)膜内のイオンの拡散による拡散 電位と、 (2)膜表面や膜内の固定電荷による表面電位やドナン電位との和となる。神経 や筋肉細胞膜では刺激により,膜電位が一過性に変化する興奮現象が見られる。この 膜電位の変化はイオンの透過性の変化により引き起こされる。 2.1. 拡散電位 (diffusion potential) (1)1 種類の一価陽イオンと 1 種類の一価陰イオン(z+=−z−)の場合 Nernst-Planck の式より (¯の付いた変数、膜内の量を表す) [ [RT (d ln c ( )] F (dφ / dx )] J + = −c +ω + RT (d ln c + / dx ) + z + F dφ / dx J − = −c −ω − − / dx ) − z − 電気的中性条件(膜内) イオン濃度: c+o = c−o = cio (膜内 o 側), c+i = c−i = cii (膜内 i 側) 流束: J + = J − = J i ⇒ ω − ω − RT d (ln ci ) dφ =− + dx ω + + ω − z + F dx ω + − ω − RT cii ln (積分)⇒ ∆φ = φ − φ = − ω + + ω − z + F cio 分配係数 β i = cii / cii = cio / cio ω − ω − RT cii ln 膜電位⇒ ∆φ = φ i − φ o = − + ω + + ω − z + F cio 2 RTω + ω − dci dc 流束 ⇒ J i = − = − D± i (=Fick の拡散の式) ω + + ω − dx dx i • o D±(電解質の拡散係数) : D± = RTω ± = 2 RT ω +ω − ω +ω − (ω ± = 2 ) ω+ +ω− ω+ + ω− 外部から電位差を加え、電流を流さない限り陽イオンと陰イオンの流束は等しい ⇒ 陰イオンと陽イオンは同じ速度で拡散。このときの移動度は ω ± となる • 発生する電位は,ω+ と ω− の差が大きいほど大 ω+≠0、 ω-=0 のときは、J=0、 Nernst の平衡電位 (2)3種類以上の一価イオンが存在する場合 Goldman-Hodgkin-Katz の式(定電場の仮定使用) 9 一価イオンの流束 J + = − o i o i P+ F∆ϕ c+ − c+ exp( F∆ϕ / RT ) P F∆ϕ c− − c− exp( F∆ϕ / RT ) 、 J− = − RT RT 1 − exp( F∆ϕ / RT ) 1 − exp( F∆ϕ / RT ) 電流ゼロの条件: I = F (∑ J − ∑ J ) = 0 + − o i RT ∑ P+ c + + ∑ P− c − 膜電位: ∆φ = φ − φ = ln F ∑ P+ c+i + ∑ P− c−o i o 主イオン種が Na+, K+, Cl−のとき(生体膜の場合) o + PK c Ko + PCl cCli RT PNa c Na ln (Goldman-Hodgkin-Katz の式) ∆φ = i F PNa c Na + PK c Ki + PCl cClo 2.2. 表面電位 (Gouy−Chapman の拡散電気二重層) 膜界面に固定電荷⇒界面近傍の溶液内のイオン分布(Poisson−Boltzmann) φ= 2k B T exp(eφ 0 / 2k B T ) + 1 + [exp(eφ 0 / 2k B T ) − 1]exp(− κx ) ln e exp(eφ 0 / 2k B T ) + 1 − [exp(eφ 0 / 2k B T ) − 1]exp(− κx ) 2n 0 e 2 φ0 at x=0; κ = εε 0 k B T φ0 が小さいとき (Debye-Huckel 近似): φ = φ 0 exp(− κx ) 1/κ:拡散二重層の厚さ (Debye length) 2.3.ドナン電位 膜あるいは膜表面近傍に固定電荷 ⇒ 膜と水溶液の間でイオンの分布 ⇒ 界面にドナン電位(Donnan potential) 膜に負の固定電荷(電荷密度 X−)、1−1 電解質を考える 界面でのイオン分布は平衡状態 電気化学ポテンシャルは膜と溶液界面の間で等しい RT ln c + + Fϕ = RT ln c + + Fϕ 、 RT ln c − + Fϕ = RT ln c − + Fϕ (標準電気化学ポテンシャルは膜と溶液の両相で変わらないと仮定) ドナン電位: ∆ϕ = ϕ − ϕ = − RT ⎛ c + ln⎜ F ⎜⎝ c + ⎞ RT ⎛ c − ⎟⎟ = ln⎜⎜ ⎠ F ⎝ c− ⎞ ⎟⎟ ⎠ ⇒ c + c − = c + c − 10 電気的中性条件 ⇒ c + = c − = c 、 c + = c − + X − ⇒ c + c − = (c − + X − )c − = c + (c + − X − ) = c ( ⇒ c + = X − + ) X −2 + 4c 2 / 2 、 2 ) ( c − = X − − X −2 + 4c 2 / 2 膜−溶液間のイオンの分配係数: ) ( ) ( β + = c + / c + = X − + X −2 + 4c 2 / 2c 、 β − = c − / c − = X − − X −2 + 4c 2 / 2c ⇒ ドナン電位: ∆ϕ = − RT RT ln (β + ) = ln(β − ) F F 2.4. 膜電位の測定法 電極:カロメル電極、銀塩化銀電極(Ag-AgCl) 銀塩化銀電極の電極反応: AgCl + e- = Ag + Cl- ; 標準電極電位 V0=0.2225V 起電力 Ve: Ve = V0 − (RT F ) ln a cl 、 ( V0 = − ∆G0 / F ) 銀塩化銀電極を用いた測定系 Ag | AgCl(固体) | NaCl(a1) | 膜 | NaCl(a2) | AgCl(固体) | Ag Vle Vm Vre 測定される起電力(電位差): Vob = Vle + Vm + Vre (電位は左の電極が基準) 正起電力:右の電極は左の電極に比べ正(電子不足) 電極間を導線で結ぶ⇒ 導線を電子は左から右(電流は右から左) 左の電極反応: Ag + Cl- = AgCl + e- Vle = −[V0 − (RT F ) ln a1 ] 右の電極反応: AgCl + e- = Ag + Cl- Vre = V0 − (RT F ) ln a 2 実測電位差=膜電位差+電極電位差: Vob = Vle + Vm + Vre = − RT ⎛ a 2 ln⎜ F ⎜⎝ a1 ⎞ ⎟⎟ + Vm ⎠ 膜の両側での Cl-イオンの活量既知 ⇒ 電極電位の補正可能 生物細胞では,細胞内の Cl-イオンの活量を正確に求めること困難 ⇒ 塩橋 (salt bridge)を用いた方法(電極電位差をほぼゼロ→実測電位差≈膜電位差) • 塩橋の作り方:濃厚な 3M KCl 溶液に寒天を加え、熱して溶解する。この液をポリエチ レンやガラスのチューブに詰めたあと、冷やして固める。 11 Ag|AgCl|KCl(a3) - 塩橋|NaCl(a1)|膜|NaCl(a2)|塩橋 - KCl(a3)|AgCl|Ag Vle Vlj Vm Vrj Vre 左右の銀塩化銀電極は同じ濃度の KCl 溶液中にある ⇒ Vle=−Vre 塩橋 KCl(活量 a3)と NaCl(活量 a1)と液間電位 Vlj ⇒ Vlj = RT a1 (ω Na − ω Cl ) − a3 (ω K − ω Cl ) a1 (ω Na + ω Cl ) ln F a1 (ω Na + ω Cl ) − a3 (ω K + ω Cl ) a 3 (ω K + ω Cl ) a3≫a1、 a3≫a2、 ωK≒ωCl ⇒ Vlj≈0,Vrj≈0 ⇒ Vob ≒ Vm (Henderson の式) 12 2. 膜興奮現象とイオンチャンネル 2.1.膜の興奮現象 神経や筋肉の細胞膜は刺激によって膜電位が一過性に変化する興奮現象を示す。この膜 電位の変化は膜のイオン透過性を巧みに変化させることによって引き起こされる。 • 細胞内外のイオン組成 能動輸送(Na-K pump, Na-K ATPase)⇒ K+イオン取り込み、Na+、Ca2+イオン排出 ⇒ 細胞内は K+イオン濃度が高く、Na+や Ca2+イオンの濃度は低い イカ神経繊維とカエル筋繊維のイオン組成(mM) イオン K+ Na+ Ca2+ Mg2+ Cl- イカ神経繊維 原形質 血液 400 20 50 0.4 10 40 440 10 54 560 海水 10 460 10 53 540 カエル筋繊維 筋繊維 血漿 125 2.5 10.5 4.9 14 1.5 10.9 2.1 1.25 77.5 K+イオンを選択的透過する細胞膜⇒K+イオンの拡散によって膜電位は負(細胞外を基準) 膜電位(静止膜電位)≈ EK (K+イオンの Nernst 電位 EK) E K = K RT K o ln = 58 log10 o F Ki Ki (at 25℃) 細胞内外の K+ の濃度と静止電位 ヤリイカ巨大神経繊維 コウイカ巨大神経繊維 カエル横紋筋 ラット心筋 Ki/mM Ko/mM Ek/mV(計算値) 実測値(mV) 410 360 125 140 10 10 2.5 2.7 -93 -90 -98 -99 -61 -62 -95 -92 K+イオン、Na+イオン、Cl-イオンの透過を考慮 Goldman-Hodgkin-Katz の式 ∆φ = RT PNa c ln F PNa c o Na i Na + PK c + P c + PK c + P c o K i K i Cl Cl o Cl Cl 膜電位/mV 実測膜電位と EK との違い 2 0 1 −20 −40 −60 傾斜 −58 mV 曲線1: PK:PNa:PCl=1:0.04:0.45 曲線2: PK:PNa:PCl=1:0.0025:0.3 10 100 500 [Ko] / mM 静止電位と外液 K+濃度 13 (1)神経細胞の膜 V 電位記録法(微小電 I V V 極法) (右図参照) 電極:ガラスピペッ ト(先端直径~0.1 µm、 内 部 電 解 質 ( KCl ), 銀塩化銀電極) 膜電位/mV 40 0 活動電位 V −40 静止膜電位 −80 膜電流 外向き a I b 内向き a. 微小電極(電位測 定用)刺入→電位 d c 膜電位の記録と活動電位 -60 mV(静止電位) b. 微小電極(電流供給用)で、外向電流(膜電位 (分極)を打ち消す方向の電流)→脱分極(電 80 流×膜抵抗分の電位が変化) c. 内向き電流→過分極 40 膜電位/ mV d. 外向き電流→活動電位(膜電位は正方向にスパ イク状に変化) (2)活動電位の発生機構 活動電位 0 −40 細胞外液の Na+濃度と,活動電位のピーク電位や静 静止電位 止電位との関係 −80 100 活動電位は Na+の濃度の対数に比例 ⇒ 興奮時は Na+の拡散による電位が支配的 + ⇒ Na イオンの透過性の増加 1000 [Na]o / mM 外液の Na+濃度と活動電位 フィードバック 膜電位測定 増幅器 神経 固定電位 膜電流/ mAcm−2 電流測定 膜電位/ mV (3)電位固定法(膜電位を固定して、イオン流を電流として測定) -10 脱分極 -65 1 容量電流 0 内向き電流 −1 0 イカ巨大神経繊維を用いた膜電位固定法 遅い外向き電流 2 時間/ ms 4 14 フィードバック回路から電流供給 ⇒ 膜の電位を一定に保持 静止電位から脱分極(図では‐10 mV に固定) ⇒電流は内向き(細胞外から内へ) 、その後外向きに変化 内向き電流:Na+の濃度勾配に従った、外から内への Na+の拡散 外向き電流:K+の濃度勾配に従った,内から外への K+の拡散 ⇒ 一過性の Na+イオン透過性増加、続いて、遅い K+イオン透過性増加 • Na+、K+の透過性制御は電圧依存性の Na+チャンネルと K+チャンネルで行われる • テトロドトキシン(TTX):Na+チャンネルを特異的にブロック、内向き電流消失 電流 mA/cm 2 TEA (tetraethyl ammonium chloride):K+チャンネルをブロック, 外向き電流消失 2.5 0 −2.5 TTX 0 TEA 1 3 2 時間/ ms TTX による Na+イオン流ブロック 2.2.膜電位の光学的測定 中枢神経系(多くのニューロン(神経細胞)からなる複雑なネットワーク) ⇒ ニューロン間のユニークな連結により、複雑で多様な情報処理 ネットワークでの信号(活動電位)伝播可視化 ⇐ 活動電位の光学的計測法 電位感受性プローブ(膜電位によって吸光度や蛍光強度が変化する分子) Membran δ− δ+ δ+ δ− δ+ δ− 膜電位による光学変化の機構 膜電位の光学プローブ 4 δ+ δ− 15 膜電位変化に伴う吸光度の変化 (メロシアニン・ローダニン色 A ∆A/Ao 2×10−4 1×10−3 素染色イカの神経繊維) 吸光度の変化 ∝ 膜電位 V 神経膜興奮時の活動電位は吸光 I 50 mV 100 mV -1×10-3 5 nA/cm2 度の変化と一致 3 ms 5×10−4 ニワトリ初期胚心臓からの活動 電位の吸光度シグナル(下図) A 50 mV V 活動電位の間のわずかな遅れ ⇒信号伝達ルート推定 1 ms 膜電位(V)に依存した吸光度変化(∆A) 10−3 2×10−3 8 somites ① ④ ② ③ ⑤ 200 µm 2.0 s 0.5 s ニワトリ初期胚心臓の活動電位の光学記録 3.イオンチャンネル 神経や筋肉細胞の興奮現象についての電気生理学的研究から、イオンチャンネルの概念 が生まれた。チャンネルの存在は、特定イオンの透過を特異的に阻害するブロッカーが見 つかったことからより確かになった。 (1)チャンネル蛋白質の構造解析 • ブロッカーを利用したアフィニティ・カラムクロマト⇒チャンネル蛋白質の分離精製 • チャンネル蛋白質の一次配列の決定:DNA の配列⇒蛋白質配列 • 一次配列⇒二次構造、膜貫通部分の推定 16 • X 線回折による三次元結晶解析 porin (Kresch and Schulz, 1994) potassium channel (Doyle et al., Science 280:69-77, 1998), iron transporting channel (Ferguson et al., 1998), mechanosensitive channel (Chang et al., 1998), chloride channel (2002)(全てバクテリア由来) • 電子顕微鏡電子線回折による二次元結晶解析 aquaporin(Murata et al., Nature 407:599-605 (2000), acetylcholine receptor (2)機能解析 • パッチクランプ法による単一イオンチャンネル解析 • パッチクランプ法と遺伝子工学的手法を組み合わせた機能構造の相関研究 カエル卵や培養細胞にチャンネル蛋白質の変異 DNA 導入⇒細胞膜にチャンネル発現 ⇒パッチクランプ法による解析 3.1.イオンチャンネルの性質 イオンチャンネルの3つの素機能 (1)フィルター機能:イオンの 選択的透過 Voltage sensor (2)センサー機能:電圧変化や 刺激分子(神経伝達物質、 Ca++イオンなど)の結合を 検出 Narrow selectivity filter (3)ゲート機能:チャンネルの 開閉 電圧依存性イオンチャンネルのイ オン選択性(下表) nm イオンチャンネルの機能 gate 17 Current 2 0.08 mA/cm イオンチャンネルのイオン透過性 Time / ms ゲーティング電流 • ゲーティング電流: チャンネルの開閉に伴って流れる電流(≠イオン透過によるチャンネル電流) ⇐ チャンネルタンパク質のコンフォ−メーションの変化に伴う電荷の移動 ゲーティング電流の測定(ゲーティング電流<<イオン透過電流): 外部溶液の Na+を除き、tetrodotoxin で Na+イオン流をブロック tetramethylammonium fluoride で K+イオン流をブロック 3.2.パッチクランプ法 単一のイオンチャンネルを流れる電流はどのくらいの量になるのか? • 単位面積あたりのチャンネル数(数密度)の見積 :50-500 チャンネル/µm2 Na+チャンネルの数密度(イカ巨大神経繊維) (アイソトープ・ラベルしたテトロドトキシンを用いて) 単一チャンネル電流=興奮時の Na+チャンネル電流/チャンネルの数密度 =数 pA(1pA=0.62×107 ions/s) pipette • 単一チャンネルの測定に必要な条 cell 件 ノイズレベル<0.1pA muscle fiber membrane suction 測定する膜面積を微小領域に限定 (チャンネルが数個含まれ程度に) ガラスピペット パッチクランプ法 先端径∼1µm,滑らかなガラス先端部分をもつ ピペット内部に電解質をつめ Ag-AgCl 電極を装着 ピペット先端を細胞に押付⇒ピペット内部を陰圧⇒細胞膜とピペット先端を密着 18 ⇒ 電気的に絶縁された膜微小領域(パッチ)⇒単一チャンネル電流測定 3.3.チャンネルの構造 ペプチドのアミノ酸側鎖の疎水性度 ⇒ 膜内領域と膜外領域の推定 • Na+チャンネル:4つのドメイン、S1−S6 の膜貫通領域/ドメイン 膜貫通領域:二十数残基のアミノ酸からなるヘリックス構造 H5(S5-S6 間)領域:β構造のループで、穴の壁を構成 S4:正電荷を持つアミノ酸残基を三つおきに持つ(電圧のセンサー領域) • Ca++チャンネルも Na+チャンネルと似た構造、同じファミリー • K+チャンネル蛋白質:Na+チャンネルの一つのドメインに相当する配列をもち、四量 Hydropathyl Index 体でイオンチャンネルを構成 Amino acid no. Na+チャンネルの hydropathy (疎水性度)plot K-channel Na チャンネルの膜貫通セグメント 3.4. チャンネルの構造と機能の相関 Na- and Ca-channels 電圧依存性チャンネルのドメイン構造 19 遺伝子工学と電気生理学を組合せた方法 アミノ酸配列の一部を変異させた mRNA を細胞に導入 ⇒変異イオンチャンネルの発現 ⇒パッチクランプ法適用 ⇒電圧センサー、フィルター、ブロッ カーの結合サイトなどの推定 変異遺伝子導入と発現した変異 イオンチャンネルの解析 3.5.チャンネルの三次元構造とイオン透過 K-チャンネル三次元結晶解析の結果 (1)フィルター部分のモデル(右図) out 細胞質側から細胞外の方向に従って 長いトンネル(1.8 nm)から広い cavity(直 径 1 nm)(穴の壁は疎水性、水和イオンとし て透過) ⇒フィルター部分の入り口で脱水和 ⇒フィルター(主鎖のカルボニル酸素がイ オンを安定化) in K−チャンネルのイオン透過孔モデル • イオンはフィルター部分に2個、cavity に1個存在 • cavity の正電荷は,cavity から遠ざかる 方向に双極子をもつヘリックスで囲まれ ることにより,安定化 • 疎水性の壁はイオンとの相互作用が少な い⇒イオン透過性を上げる • チャンネルの両側:負電荷アミノ酸残基 ⇒カチオンを引き寄せる S4 セグメントの電圧 センシング・モデル (2)電圧センシング機構のモデル(右図) 20 4.モデルチャンネルと平面脂質二分子膜法 イオンチャンネルのイオン透過機構のを検討する上で,単純なモデル系を用いた研究は重 要なヒントを与える。 モデルチャンネル系:構造の解析の容易な低分子量のチャンネル分子+脂質二分子膜 チャンネル形成分子:合成ペプチド、ペプチド性抗生物質など 脂質二分子膜:平面脂質二分子膜(直接チャンネル電流を測定可能) リポソ−ム(脂質二分子膜をもつ小胞、分光学的な方法適用容易) 4.1. 平面脂質二分子膜の作成法 平面脂質二分子膜の作り方 (A) 刷毛塗り法(painting method): A 脂質の溶液をテフロンのポ C ットあるいはシートに開け た穴に塗り付け⇒膜が自然 に薄化(あるいは電圧をか けて強制的に)⇒ 二分子 B 膜 欠点:溶媒(デカンなど) が膜中に残る(溶媒を選ぶ ことにより,残存溶媒がほ とんど無い膜を作ることも 平面脂質二分子膜の作り方 可能) (B) 張り合わせ法(folding method): 液面をテフロンシートに開けた穴の下におき、気液界面に脂質単分子膜作成⇒液面を上 下げして、穴を通過⇒二分子膜形成 特徴:溶媒を含まない膜 (C) ティップ・ディップ法(tip-dip method) 先端を細くしたガラスのキャピラリー(内部に電解質)使用 先端を気液界面に作った単分子膜を通過させる⇒先端に二分子膜形成 特徴:小さい膜面積(電気的なノイズを抑えることができる) 4.2. 脂質二分子膜の性質とイオノフォアの作用 脂質二分子膜と生体膜の比較:膜の厚さ、電気容量、破壊電圧,水透過性はほぼ同じ 最も大きな違いは膜抵抗とイオン透過性 21 この脂質二分子膜にイオノフォアとよばれる一群の物質をあたえると、膜抵抗の減少やイ オン透過性の増加が見られる。イオノフォアにはキャリアー型とチャンネル型がある。 キャリアー型:イオンと抱接化合物をつくり膜を透過 バリノマイシン、クラウンエーテル、X-537A(下図) キャリア型イオノフォア チャンネル型:脂質二分子膜にイオンチャンネルをつくる グラミシジン,アラメチシン チャンネル型では離散的なチャンネル電 流が観測される グラミシジン・チャンネル電流(右図) グラミシジンのチャンネル電流揺らぎ グラミシジンの構造と チャンネルモデル 22 チャンネル型イオノフォア 2.3 人工チャンネル形成分子 人工チャンネル形成分子のデザインについては、大きく分けて3つのアプローチがある。 (1)天然のチャンネル形成ペプチド(抗生物質や毒素)を修飾して、機能改変をする。 アラメチシン、グラミシジンがよく使われる。(2)生体のチャンネルタンパク質の穴形 成部分と推定されるペプチドを合成して、チャンネル形成の可否や基となる生体のチャン ネルと比較する。(3)基本骨格にペプチドを用いず、チャンネルの機能的特徴だけを模 倣するようにデザインした人工チャンネル分子。 アセチルコリンレセプ ター M2 セグメント 人工イオンチャンネル
© Copyright 2026 Paperzz



![名前:解答例 問題6 (1) 純アルミニウムにおいては、 eV /atom]、ASV /R](http://s3.paperzz.com/store/data/005656799_1-6de218d9e340e146495f2f9cbf2e9e02-250x500.png)