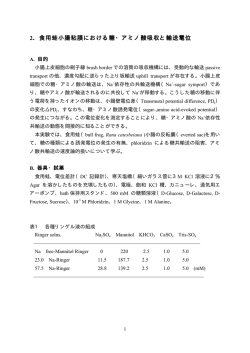
V V∼const. [S] [S] 図 2.3: 酵素反応速度の基質濃度依存性. 全酵素濃度は一定であるとする. 反応速度は基質濃度が低いとき基質濃度にほぼ比例するが、基質濃度が増 加するに従い、反応速度は飽和し基質濃度がどんなに増加しても一定と なる. 2.1.4 酵素反応速度論 (Enzyme Kinetics) 基質 (Substrate)S が酵素 (Enzyme)E を触媒 (catalyst) として生成物 (Product)P となる反応を考えよう。酵素反応には、図 2.3 に示したよう に、基質濃度 [S] がどんなに高くても (酵素濃度 [E] は一定という条件で)、 反応速度 V(生成物 P の生成速度) は頭打ちになるという特徴がある。つ まり、基質濃度に対して常に線形で増加するわけではない。酵素反応の特 徴を纏めると次のようになる。 (1) 基質濃度 [S] が一定のとき、[E] に比例する。 (2) 基質濃度 [S] が低濃度のとき、一定の酵素濃度では [S] に比例する。 (3) 基質濃度 [S] が高濃度のとき、一定になる。 従って、単純に質量作用の法則を適用することはできない。その理由を 考えてみよう。いま、酵素反応について、反応物を S, E、生成物を P と して、 k S + E!P + E とのように反応すると考えると、 V = d[P] = k[S][E] dt 11 のように書けると思うかもしれない。しかし、これでは基質濃度 [S] に 対して常に [E] に比例して増加してしまうことになってしまう。そこで Michaelis と Menten(1913) は、反応中間体 (複合体 (Complex))C の存在 を仮定し次の反応式を提案した。 k+1 k S + E ! C ⇀2 P + E k−1 (2.8) 彼らの功績から酵素反応速度に関する関係式をミカエリス-メンテンの 式 (Michaelis-Menten equation)(2.18) という。この式の導出法に関して 後に Briggs と Haldane (1925) によって修正が加えられ一般化された。こ こでは彼らの方法に従って解説する。彼らは、[S] ≫ [E] を仮定し、複合 体 C が分解して酵素 E が解放されるとすぐさま別の基質 S と結合するこ とに着目した。また、反応 (2.8) の後半では生成物 P はすぐさま除去さ れ (実際に細胞内では生成物は直ぐに別の反応に用いられることになるの で、この仮定は妥当である。) その逆反応は無視できるものとしている。 これらのことから、C の生成速度と崩壊速度が均しい状態、準定常状態 (quasi-steady state) 6 を仮定した。これについては次のセクションで解説 する。 簡単のためにそれぞれの化学種の濃度を s = [S], e = [E], c = [C], p = [P] とおくことにする。質量作用の法則から、それぞれの化学物質の生成速度 に関する方程式を次のように書くことができる。 ds dt de dt dc dt dp dt = k−1 c − k+1 se (2.9) = k−1 c + k2 c − k+1 se (2.10) = −k−1 c − k2 c + k+1 se (2.11) = k2 c (2.12) いま、ここで行いたいのは、ミカエリス–メンテンの式の導出である。つ まり、生成物の反応速度 V と基質の濃度 s の関係式の導出を行う。V = dp dt なので、式 (2.12) を最終的に用いることがわかる。従って、残りの式から 6 準定常状態では、ある物理量が、長い時間スケールで見たら変化しているが、短い時 間スケールで見たらほとんど変化していないので、変化がないものと近似できるような場 合である。どのような時間スケールでも変化しない定常状態とは異なる。 12 c を求めればよい。ところで、式 (2.10) と式 (2.11) の右辺は符号が異なる だけなので、次の関係式が成立する。 de dc + =0 dt dt e + c = const ≡ e0 (2.13) (2.14) ここで、e0 は遊離酵素、結合酵素を含めた全酵素の濃度である。この式 から変数 e は消去されて、本質的には c と s だけの 2 変数だけ考えれば良 いことがわかる。 2.1.5 準定常状態近似 (quasi-steady state approximation) 前のセクションで触れたように Briggs と Haldane は C の生成速度が崩 壊速度と均しいとし、C の準定常状態を仮定した。つまり式 (2.11) で、 dc =0 dt (2.15) とおける。この条件と、式 (2.14) より、e を消去すると、 c= e0 s s + Km (2.16) が求まる。ここで、 Km ≡ k−1 + k2 k+1 (2.17) とおいている。Km をミカエリス定数 (Michaelis constant) という。生成 物の反応速度 V は式 (2.12) より次式で与えられるので、式 (2.16) より、 dp = k2 c dt Vmax s = s + Km V = (2.18) が導出できる。ここで、 Vmax = k2 e0 とおいた。式 (2.18) をミカエリス-メンテンの式 (Michaelis-Menten equation) という。これを図示したものが、図 2.4(a) であり、節 2.1.4 の冒頭で 挙げた酵素反応の特徴をよく表している。反応速度がちょうど反応速度の 最大値 Vmax の半分となったときの基質濃度 s が Km と等しくなる。(各 自確かめておくこと。) つまり、Km は反応がその最大速度に達するのに 必要な基質濃度の目安となる。この値が小さいほど、基質と親和性の高い 酵素ということになる。これは次の Lineweaver-Burk プロットという形 13 (a) (b) V 1/V Vmax Vmax /2 1/Vmax s Km -1/K m slope = Km /Vmax 1/s 図 2.4: ミカエリス-メンテンの式 (2.18) による s−V プロット (a), 1/s−1/V プロット (b). 式に変形でき、実験ではこのプロット (図 2.4(b)) から、Vmax , Km を求め る 7。 1/V = s + Km Vmax s = 1/Vmax + (2.19) Km 1/s Vmax (2.20) (練習問題 2-4) ミカエリス-メンテンの式 (2.18) で、本節の最初に整理し たミカエリス-メンテン則の性質が表現できているか以下の方法で各自確 認してみよう。各条件を s = const., s が非常に小さい場合, s が非常に大 きい場合として、式 (2.18) がどう近似されるか示して議論しなさい。 (練習問題 2-5) ミカエリス-メンテンの式 (2.18) を線形にプロットする手 法として、Lineweaver-Burk プロット (1/s–1/V ) の他に、Eadie-Hofstee プロット (V /s–V ) および Hanes-woolf プロット (s–s/V ) と呼ばれるもの がある。これらのプロットを作成するために式 (2.20) を参考に、それぞ れ対応する関係式を導出なさい。図 2.4(b) に習ってプロットを作成せよ。 さらに、3 つの手法の利点、欠点について議論せよ。 2.1.6 平衡近似 (equilibrium approxmation) 前セクションでは複合体の準定常状態を仮定して、ミカエリス-メンテ ン式を導出したが、遊離酵素 E と基質 S から 複合体 C 形成する反応 (反 7 コンピュータによる解析が発達する以前は、このような手法でデータを線形の関係に 変換してから線形回帰によって各係数を求めていた。しかし線形の関係に変換する際に、 値によって誤差が異なり係数の見積もりを誤ることがある。現在では加工前のデータから 非線形回帰によって各係数を求める。 14 応式 (2.8) の1番目の反応) が平衡状態にあると仮定して導出する方法も あり、この方法を平衡近似という。歴史的にはこちらの方が先であり、複 雑な系を概観するには適しているので紹介する。 式 (2.7) より平衡定数は、その反応の反応物、生成物の濃度比で決まる ことを思い出しておこう。このとき、Ks = k−1 /k+1 とおけば 8 、 se − Ks c = 0 (2.21) となることがわかり、これより c が求まる。従って、式 (2.14) と併せて、 dp = k2 c dt Vmax s = s + Ks V = (2.22) が求まる。式 (2.18) と式の形は同じであるが、Ks は Km とは定義が異な ることに注意しよう。 8 解離定数という。節 2.1.3 で定義した平衡定数の逆数となっていることに注意。 15
© Copyright 2026 Paperzz